Structural and functional genomics and evolutionary relationships in the cluster of genes encoding murine 2,5-oligoadenylate synthetases Tomoji Mashimo, a Philippe Glaser, b Marianne Lucas, c Dominique Simon-Chazottes, a Pierre Emmanuel Ceccaldi, d Xavier Montagutelli, a Philippe Despre `s, c and Jean-Louis Gue ´net a, * a Unite ´ de Ge ´ne ´tique des Mammife `res, Institut Pasteur, 75724 Paris Cedex 15, France, b Laboratoire de Ge ´nomique des Microorganismes Pathoge `nes, Institut Pasteur, 75724 Paris Cedex 15, France, c Unite ´ des Interactions Mole ´culaires Flavivirus-Ho ˆtes, Institut Pasteur, 75724 Paris Cedex 15, France, d De ´partement de Virologie, Institut Pasteur, 75724 Paris Cedex 15, France Received 3 January 2003; accepted 30 May 2003 Abstract 2,5-Oligoadenylate synthetases (2,5-OASs) are interferon-inducible enzymes. Some of these proteins play an important role in cellular physiology, in particular, in the innate defense mechanisms against RNA virus infections. In the present publication we report the complete genomic structure of the cluster of genes encoding mouse 2,5-OAS, with all its transcription units, their predicted functions, and their evolutionary relationships. We found that mouse Oas2/Oas3 genes have a genomic structure similar to that of human OAS2/OAS3, while the mouse equivalent of human OAS1 is composed of eight (Oas1a to Oas1h) tandemly arranged transcription units. For all these eight genes a specific inducible promoter controls transcription. The possible functions of this family of proteins are discussed. © 2003 Elsevier Inc. All rights reserved. Keywords: 2,5-Oligoadenylate synthetases; Evolution; Functional genomics; mouse In mammals, 2,5-oligoadenylate synthetases (2,5- OASs) represent a relatively homogeneous family of enzy- matic proteins with a remote evolutionary origin, since molecules with a similar structure have been identified in a wide range of species including birds [1], reptiles [2], and even the marine sponge Geodia cydonium [3]. At least some of these proteins are interferon-inducible enzymes and play an important role in the host’s innate defense mechanisms against viral infections, in particular where positive- stranded RNA viruses are concerned [4 – 8]. Recently, we and others reported that a nonsense mutation in the gene encoding the Oas1/L1 isoform of the mouse is constantly associated with, and presumably responsible for, the sus- ceptibility of several inbred strains to experimental infec- tions with flaviviruses [9,10]. This antiviral activity of 2,5- OASs might also be direct and interferon-independent, since plants transgenic for genes of this family and for the gene encoding RNase L were found to be resistant to ex- perimental infections with a number of viruses such as tobacco mosaic virus, cucumber mosaic virus, and potato virus Y [11,12]. Finally, in addition to the role they play in the host’s defense mechanisms, proteins of this family have been demonstrated to play a role in some other important cellular processes such as growth control, differentiation, and apoptosis [13–17]. Considering the multiplicity of the OAS-encoding genes [4 – 8], their evolutionary conservation, and the fundamental functions they appear to be involved in, it is reasonable to assume that this family of genes is important for the main- tenance of homeostasis at the cellular level and, accord- ingly, it would be interesting to investigate in detail the regulation and function(s) of each of its individual mem- bers, using some of the modern strategies of genetic engi- neering, both in vitro and in vivo, in the laboratory mouse. * Corresponding author. Fax: 33-1-45-68-86-34. E-mail address: [email protected] (J.-L. Gue ´net). R Available online at www.sciencedirect.com Genomics 82 (2003) 537–552 www.elsevier.com/locate/ygeno 0888-7543/03/$ – see front matter © 2003 Elsevier Inc. All rights reserved. doi:10.1016/S0888-7543(03)00176-9

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Structural and functional genomics and evolutionary relationships in thecluster of genes encoding murine 2�,5�-oligoadenylate synthetases
Tomoji Mashimo,a Philippe Glaser,b Marianne Lucas,c Dominique Simon-Chazottes,a
Pierre Emmanuel Ceccaldi,d Xavier Montagutelli,a Philippe Despre`s,c
and Jean-Louis Gue´neta,*a Unite de Genetique des Mammiferes, Institut Pasteur, 75724 Paris Cedex 15, France,
b Laboratoire de Genomique des Microorganismes Pathogenes, Institut Pasteur, 75724 Paris Cedex 15, France,c Unite des Interactions Moleculaires Flavivirus-Hotes, Institut Pasteur, 75724 Paris Cedex 15, France,
d Departement de Virologie, Institut Pasteur, 75724 Paris Cedex 15, France
Received 3 January 2003; accepted 30 May 2003
Abstract
2�,5�-Oligoadenylate synthetases (2�,5�-OASs) are interferon-inducible enzymes. Some of these proteins play an important role incellular physiology, in particular, in the innate defense mechanisms against RNA virus infections. In the present publication we report thecomplete genomic structure of the cluster of genes encoding mouse 2�,5�-OAS, with all its transcription units, their predicted functions, andtheir evolutionary relationships. We found that mouseOas2/Oas3 genes have a genomic structure similar to that of humanOAS2/OAS3,while the mouse equivalent of humanOAS1 is composed of eight (Oas1a to Oas1h) tandemly arranged transcription units. For all these eightgenes a specific inducible promoter controls transcription. The possible functions of this family of proteins are discussed.© 2003 Elsevier Inc. All rights reserved.
Keywords: 2�,5�-Oligoadenylate synthetases; Evolution; Functional genomics; mouse
In mammals, 2�,5�-oligoadenylate synthetases (2�,5�-OASs) represent a relatively homogeneous family of enzy-matic proteins with a remote evolutionary origin, sincemolecules with a similar structure have been identified in awide range of species including birds [1], reptiles [2], andeven the marine spongeGeodia cydonium [3]. At least someof these proteins are interferon-inducible enzymes and playan important role in the host’s innate defense mechanismsagainst viral infections, in particular where positive-stranded RNA viruses are concerned [4–8]. Recently, weand others reported that a nonsense mutation in the geneencoding the Oas1/L1 isoform of the mouse is constantlyassociated with, and presumably responsible for, the sus-ceptibility of several inbred strains to experimental infec-tions with flaviviruses [9,10]. This antiviral activity of 2�,5�-
OASs might also be direct and interferon-independent,since plants transgenic for genes of this family and for thegene encoding RNase L were found to be resistant to ex-perimental infections with a number of viruses such astobacco mosaic virus, cucumber mosaic virus, and potatovirus Y [11,12]. Finally, in addition to the role they play inthe host’s defense mechanisms, proteins of this family havebeen demonstrated to play a role in some other importantcellular processes such as growth control, differentiation,and apoptosis [13–17].
Considering the multiplicity of the OAS-encoding genes[4–8], their evolutionary conservation, and the fundamentalfunctions they appear to be involved in, it is reasonable toassume that this family of genes is important for the main-tenance of homeostasis at the cellular level and, accord-ingly, it would be interesting to investigate in detail theregulation and function(s) of each of its individual mem-bers, using some of the modern strategies of genetic engi-neering, both in vitro and in vivo, in the laboratory mouse.
* Corresponding author. Fax:�33-1-45-68-86-34.E-mail address: [email protected] (J.-L. Gue´net).
R
Available online at www.sciencedirect.com
Genomics 82 (2003) 537–552 www.elsevier.com/locate/ygeno
0888-7543/03/$ – see front matter © 2003 Elsevier Inc. All rights reserved.doi:10.1016/S0888-7543(03)00176-9

However, using these strategies requires that the structure ofthe OAS-encoding cluster of genes be precisely known atthe genomic level. For this reason, we undertook to analyzein detail the structure of the whole region in the mouse andfound that it displays a genomic organization different fromthat of human [5,6]. In addition to the fine genomic structureof the cluster, we established the evolutionary relationshipsamong these genes, investigated their tissue-specific expres-sion, and discuss their possible functions.
Results
Fine mapping of the Oas cluster of genes on mousechromosome 5
The genetic localization of the 2�,5�-OAS cluster wasachieved while we were mapping the major dominant allelethat determines resistance to an experimental infection witha highly neurovirulent strain of the West Nile flavivirus(IS-98-ST1 strain) [9]. Linkage analysis of the resistance/susceptibility phenotype was performed on the progeny offour intersubspecific backcrosses set between the suscepti-
ble laboratory strains (BALB/c or C57BL/6) and the resis-tant, wild derived, inbred strains (MBT/Pas or MAI/Pas)[18]. This allowed us to establish that the locus governingresistance/susceptibility maps to chromosome 5 (Chr 5)between markers D5Mit431 and D5Mit242 (Fig. 1).
Matching our linkage results to the genetic databasefor radiation hybrid (RH) mapping (http://www.jax.org/resources/documents/cmdata/rhmap) indicated that the clus-ter of genes encoding 2�,5�-OAS (Oas) mapped to the sameregion, making this cluster of genes a front-line candidatebased on its known functions (Fig. 1). Using an EST fromthe 3� UTR region of the Oasl cDNA (the mouse orthologof human OASL (OAS-like protein—AF068835), wemapped this gene between marker D5Mit158 and the locusof Rnf10 (Ring finger protein 10), �150 cR centromeric tothe Oas locus (Fig. 1).
We also found that both gene order and relative distanceswere highly conserved between the region of mouse Chr 5and the orthologous region of human chromosome 12, withthe exception of a small fragment spanning from the CDV1locus (carnitine deficiency-associated gene expressed inventricle 1) to the OASL locus, which appears to be invertedin the mouse compared to human (Fig. 1).
Fig. 1. Genetic localization of the 2�,5�-OAS locus (Oas) on mouse chromosome 5. Distances between loci are indicated to the left of the chromosome, incM on the genetic linkage map and in cR on the RH map. The 95% confidence intervals for the genetic distances are indicated in brackets. The Wnv locus(now identified as Flv or Oas1/L1) determines the resistant/susceptible phenotype to an experimental infection with West Nile flavivirus in mice [9]. Bothlinkage and RH maps demonstrate that the mouse Oas region is highly conserved in the orthologous segment of human chromosome 12q24 (http://genome.ucsc.edu), with the exception of a chromosomal inversion spanning from the OASL to the CDV1 locus.
538 T. Mashimo et al. / Genomics 82 (2003) 537–552
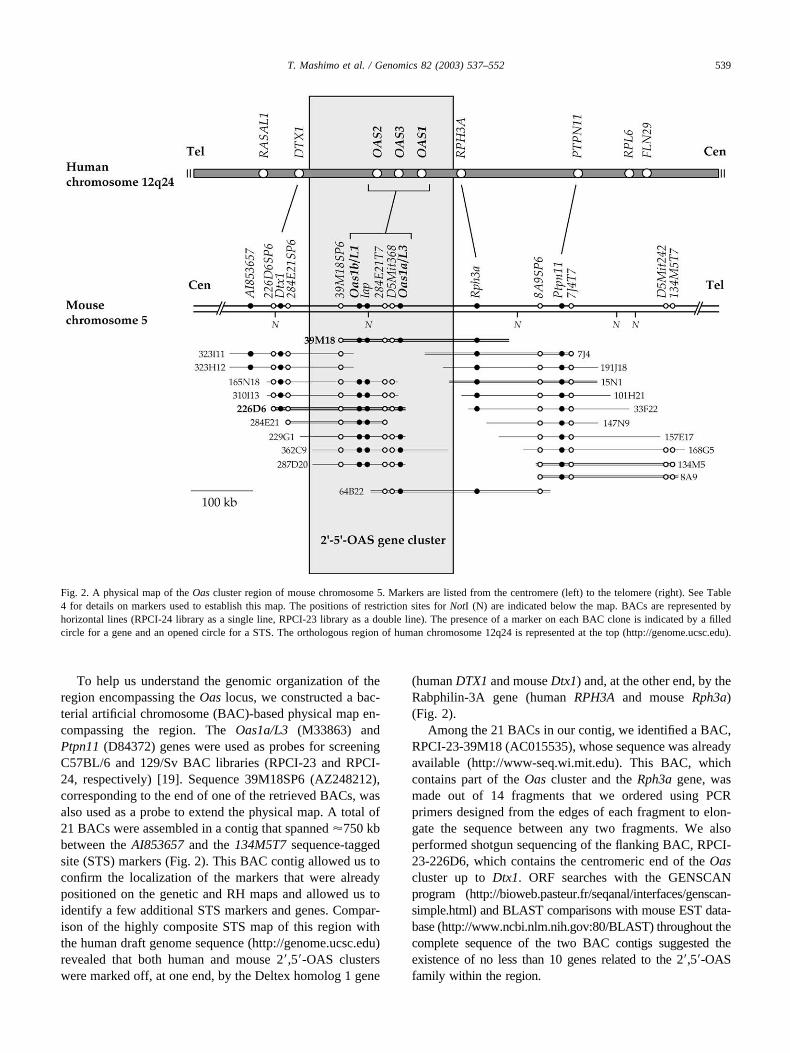
To help us understand the genomic organization of theregion encompassing the Oas locus, we constructed a bac-terial artificial chromosome (BAC)-based physical map en-compassing the region. The Oas1a/L3 (M33863) andPtpn11 (D84372) genes were used as probes for screeningC57BL/6 and 129/Sv BAC libraries (RPCI-23 and RPCI-24, respectively) [19]. Sequence 39M18SP6 (AZ248212),corresponding to the end of one of the retrieved BACs, wasalso used as a probe to extend the physical map. A total of21 BACs were assembled in a contig that spanned �750 kbbetween the AI853657 and the 134M5T7 sequence-taggedsite (STS) markers (Fig. 2). This BAC contig allowed us toconfirm the localization of the markers that were alreadypositioned on the genetic and RH maps and allowed us toidentify a few additional STS markers and genes. Compar-ison of the highly composite STS map of this region withthe human draft genome sequence (http://genome.ucsc.edu)revealed that both human and mouse 2�,5�-OAS clusterswere marked off, at one end, by the Deltex homolog 1 gene
(human DTX1 and mouse Dtx1) and, at the other end, by theRabphilin-3A gene (human RPH3A and mouse Rph3a)(Fig. 2).
Among the 21 BACs in our contig, we identified a BAC,RPCI-23-39M18 (AC015535), whose sequence was alreadyavailable (http://www-seq.wi.mit.edu). This BAC, whichcontains part of the Oas cluster and the Rph3a gene, wasmade out of 14 fragments that we ordered using PCRprimers designed from the edges of each fragment to elon-gate the sequence between any two fragments. We alsoperformed shotgun sequencing of the flanking BAC, RPCI-23-226D6, which contains the centromeric end of the Oascluster up to Dtx1. ORF searches with the GENSCANprogram (http://bioweb.pasteur.fr/seqanal/interfaces/genscan-simple.html) and BLAST comparisons with mouse EST data-base (http://www.ncbi.nlm.nih.gov:80/BLAST) throughout thecomplete sequence of the two BAC contigs suggested theexistence of no less than 10 genes related to the 2�,5�-OASfamily within the region.
Fig. 2. A physical map of the Oas cluster region of mouse chromosome 5. Markers are listed from the centromere (left) to the telomere (right). See Table4 for details on markers used to establish this map. The positions of restriction sites for NotI (N) are indicated below the map. BACs are represented byhorizontal lines (RPCI-24 library as a single line, RPCI-23 library as a double line). The presence of a marker on each BAC clone is indicated by a filledcircle for a gene and an opened circle for a STS. The orthologous region of human chromosome 12q24 is represented at the top (http://genome.ucsc.edu).
539T. Mashimo et al. / Genomics 82 (2003) 537–552

Cloning the genes within the 2�,5�-OAS cluster region
When we started our project, only a partial cDNA se-quence of the Oas1/L1 isoform was reported (X55982—now Oas1b) [20]. Using this sequence as well as the cDNAsequences deduced from mRNAs extracted from primarycultures of neural cells induced by interferon (IFN), wecould retrieve from the mouse EST database a highly ho-mologous EST clone (BC012877) containing the full-lengthORF for Oas1b. We previously described the Oas1b/L1gene of most laboratory inbred strains, indicating that it ismutated with a stop codon in exon 4, resulting in thetruncation of the encoded protein [9]. This stop codon wasalso identified in the BAC sequence (from strain C57BL/6),but not in the sequence of BC012877, suggesting that thisEST derives from a strain recently established from wildspecimens or from a laboratory strain showing resistance toflavivirus infection such as the PL/J inbred strain [9].
The genomic sequences corresponding to the other 2�,5�-OAS genes, Oas1g, Oas1a, and Oas1c, which were alreadyknown as Oas1/L2 and L3 isoforms [20,21] and Oasl5 [22],respectively, were identified in BAC 39M18 sequence. Inaddition, we identified four novel 2�,5�-OAS genes, threematching the cDNA clones deposited in the GenBank da-tabase, Oas1d (AB067532), Oas1e (AB067531), and Oas1h(AB067530), and one showing strong homology to severalESTs (BG630095 and AW743276) in the BAC sequence.Although this last gene lacked the 5� region with a startcodon, comparison with the sequences of the other 2�,5�-OAS genes allowed us to predict its complete cDNA se-quence. It was designated Oas1f. In total, at least eight
mouse Oas1 genes, whose sequences appear to be highlyhomologous to human OAS1 when translated to amino acids[4–6], were found within the cluster.
In addition to the eight Oas1 genes listed above, we alsoidentified genomic sequences previously deposited in Gen-Bank and matching two murine cDNA clones: Oasl11(AB067535) and Oasl10 (AB0675324). Considering thetranslated amino acid sequence of these sequences, theyclearly appeared orthologous to human OAS2 [23] andOAS3 [24], respectively (see Fig. 3).
Our findings are consistent with recent publications[7,8,10] on this subject reporting 10 murine 2�,5�-Oasgenes: Oas1a, Oas1b, Oas1c, Oas1d, Oas1e, Oas1f, Oas1g,Oas1h, Oas2, and Oas3. These 10 genes are structurallyarranged as shown in Fig. 3.
Fine genomic structure of the genes encoding 2�,5�-OAS
Matching the complete cDNA sequence for each of the10 2�,5�-OAS genes to the genomic sequence of the BACs(BLAST analysis) allowed us to order, size, and orient eachtranscription unit (Fig. 3). Beginning at the proximal edgeof the 2�,5�-OAS gene array, Oas2 and Oas3 appear withthe same order and orientation as OAS2 and OAS3 in human[4–6]. The eight Oas1 genes are ordered differently: Oas1a,Oas1c, Oas1e, and Oas1g are transcribed in the same ori-entation as Oas2 and Oas3, consistent with human OAS1transcriptional orientation. Oas1b, Oas1d, Oas1f, andOas1h are transcribed in the opposite direction. Finally, amouse intracisternal A particle [25] was identified 3� of theOas1b gene.
Fig. 3. Schematic organization of the cluster of genes encoding the 2�,5�-OAS. Exons of the 2�,5�-OAS genes are represented by black boxes and the directionof transcription is indicated by arrows. The BACs used in the sequence analysis are shown above. The organization of the human 2�,5�-OAS cluster, asestablished by Rebouillat et al. [5] and Justesen et al. [6], is indicated at the top.
540 T. Mashimo et al. / Genomics 82 (2003) 537–552
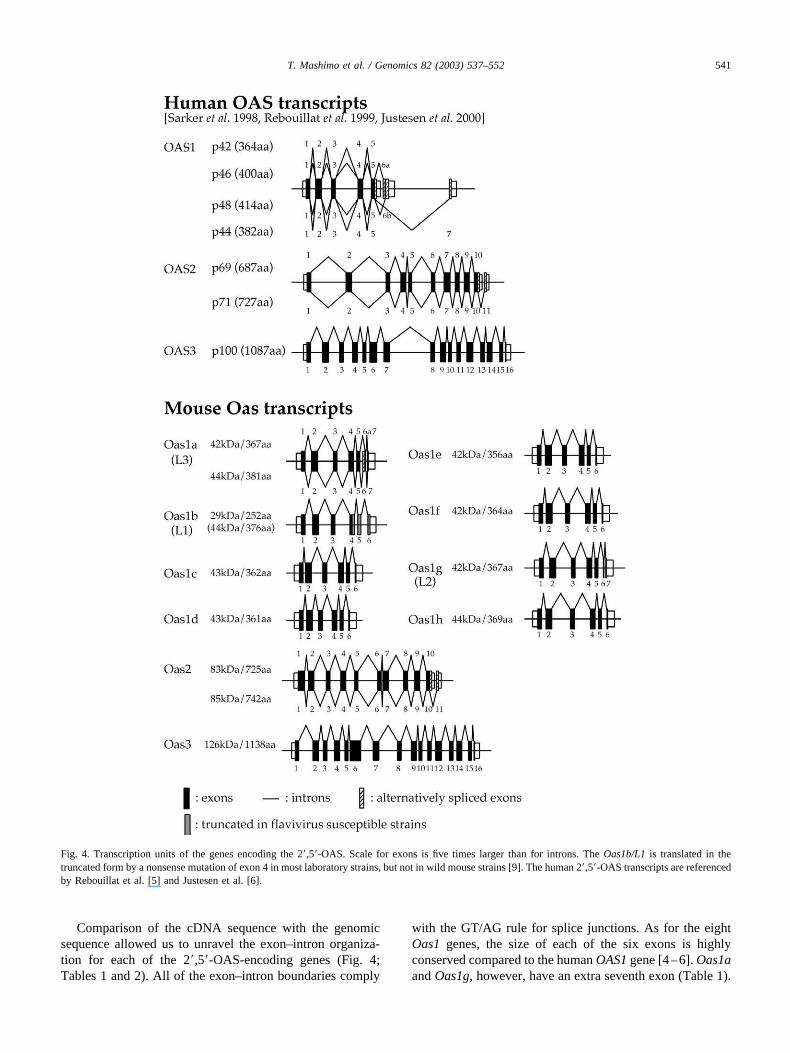
Comparison of the cDNA sequence with the genomicsequence allowed us to unravel the exon–intron organiza-tion for each of the 2�,5�-OAS-encoding genes (Fig. 4;Tables 1 and 2). All of the exon–intron boundaries comply
with the GT/AG rule for splice junctions. As for the eightOas1 genes, the size of each of the six exons is highlyconserved compared to the human OAS1 gene [4–6]. Oas1aand Oas1g, however, have an extra seventh exon (Table 1).
Fig. 4. Transcription units of the genes encoding the 2�,5�-OAS. Scale for exons is five times larger than for introns. The Oas1b/L1 is translated in thetruncated form by a nonsense mutation of exon 4 in most laboratory strains, but not in wild mouse strains [9]. The human 2�,5�-OAS transcripts are referencedby Rebouillat et al. [5] and Justesen et al. [6].
541T. Mashimo et al. / Genomics 82 (2003) 537–552

Table 1Exon–intron organization in mouse 2�–5�-OAS genes
Exon Gene Exon (bp) 5� Donor sequence 3� Acceptor sequence Intron (bp)
1 OAS1 �180 tctcctgtcaATGATGGATC GGTGGTAAAGgtgagtccag 1316Oas1b/L1 �180 tggacctaggATGGAGCAGG AGGGGTCAAGgtgagccttc 1545Oas1g/L2 �183 cagacttagcATGGAGCACG GGTGGTGAAGgtgagccctc 1318Oas1a/L3 �183 cagacttagcATGGAGCACG GGTGGTGAAGgtgagccctc 1409Oas1f �183 tggccataaaATGGTGAAGG GGTTGTGATGgtgagtccca 592Oas1c �183 tggcctcaggATGGAGAATG GGTTGTGATGgtgagtccca 332Oas1h �204 tgccctcaggATGGCGAAAA GGTTGTGATGgtgagtccca 875Oas1e �183 tggcctcaggATGGCGAGGG GGTTGTGATGgtgagtccca 922Oas1d �183 tggcctcaggATGGCGAGGG GGTTGTGATGgtgagtccca 595
2 OAS1 289 tctttttcagGGTGGCTCCT GATGCCCTGGgtgagagctc 2226Oas1b/L1 277 tgtctttcagGGCAAAGGCA GATTTACTGGgtaaggcagc 2910Oas1g/L2 289 tgtctttcagGGTGGCTCCT GATGTCCTGGgtaagccagc 3703Oas1a/L3 289 tgtctttcagGGTGGCTCCT GATGTCCTGGgtaagcctgc 3541Oas1f 289 tgtcttttagGGCGGCTCCT GATGTCCTGTgtaagtcaac 3014Oas1c 289 tgtcttttagGGCGGCTCCT GATGTCCTATgtaagtcagc 2359Oas1h 289 tgttttttagGGTGGTTCCT GATGCCCTGTgtaagtcagc 4162Oas1e 289 tgtcttttagGGAGGCTCCT GATGCTCTGTgtaagtcaga 2125Oas1d 289 tgtcttttagGGTGGCTGCT GATGCCCTGTgtaagtcaac 888
3 OAS1 185 gcccgaacagGTCAGTTGAC GTACCAAAATgtatggccct 5273Oas1b/L1 185 ttgtttacagATCATCTTAA GTACCAACTGgtaaggcatt 3396Oas1g/L2 185 tcctttacagGTCATGGTAG GTACCAACTGgtaaggcatc 2552Oas1a/L3 185 tcctttatagGTCATGTTAA GTACCAACTGgtaaggcatc 2564Oas1f 197 ttctttatagATGAACTGAG GTATCAACTGgtaaggcatc 3727Oas1c 197 ttctttatagATGAACTGAG GTATCAACTGgtaaggcatc 2346Oas1h 197 ttctttacagATGAAGTGAG GTATCAACTGgtaaggcatc 3738Oas1e 197 tccttcatagATGAAGGGGG GTATCAACTGgtaaggcatc 2811Oas1d 197 tcctttatagATGAAGTAAG GTATCAACTGgtaaggcatc 1966
4 OAS1 230 ctcttctcagtGTAAGAAGA CGAAACCCAGgtatgctatc 808Oas1b/L1 233 ctcttctcagTGTAAGGAGA AAAAAGACAGgtagtctgtc 680Oas1g/L2 233 ctcttctcagTGTAAGGAGA GAAAAGCCAGgtagtgtcca 625Oas1a/L3 233 ctcttctcagTGTAAGGAGA GAAAAGCCAGgtagtgtcca 600Oas1f 233 ctcttctcagTGTAAGGAGA AAAAAGTCAGgtagtctgtc 691Oas1c 236 ctcttctcagTGTAAGGAGA AAAAAGCCAGgtagcttgtc 627Oas1h 236 ctcttctcagTGTAAGGAGA AAAGAGTCAGgtagcttgtc 635Oas1e 233 ctcttctcagTGTAAGGAGA AAAAAGCCAGgtagcttgtc 639Oas1d 233 ctcttctcagTGTAAGGAGA AAAAAGCCAGgtagcttgtc 662
5 OAS1 154 tgctctgcagGCCTGTGATC GATTCTGCTGgtgagacctc 1688Oas1b/L1 154 tgctctgcagGCCTGTGATC GGAGGTGCCGgtgagaacat 1158Oas1g/L2 154 tcctctgcagGCCTGTGATC GGATGTGCCGgtgagacccc 1111Oas1a/L3 154 tcctctgcagGCCTGTGATC GGATGTGCCGgtgagacccc 1112Oas1f 154 tgctctgcagGCCTGTGATC GGGTGTGCCGgtgagagcct 1114Oas1c 154 tgctctgcagGCCTCTGATC GGATTTGTCGgtgagaccct 977Oas1h 154 tgctctgcagTCCTCTGATC GGATGTGCCGgtgagaccct 973Oas1e 154 tgctctgcagGCCTCTGATC GGAAGTGTCGgtaagaccct 1007Oas1d 154 tgctctgcagGCCTCTGATC GGAAGTGATGgtgagaccct 985
6 OAS1 �165 accctttcagGCTGAAAGCA CATCCTCTGAatgccagtgc —Oas1b/L1 �102 tctattacagACGGAGGTTG ATCTGGATAGcccagagtgt —Oas1g/L2 27 ttcattgcagACGGTGGTTC TTTTGAGCAGgttagcattt 155Oas1a/L3 27 ttcattgcagACGGTGGTTC TTTTGAGCAGgttagcattt 155Oas1f �39 tccgttacagATGAAGGTCG CCTTCTATGAgcaccaaagc —Oas1c �30 ttcattgcagCCAGAGAGAC CTTCCTGTGAgcacccccag —Oas1h �30 ttcattgcagCCAGACAAAA CTTCCTGTGAgcacccaagc —Oas1e �15 ttcattgcagCCAGAGAAAC GAAACAATAAtgtgtcttcc —Oas1d �30 ttcattgcagCCAGAGAGAC CTTCCAGTGAgcagtgcagc —
7 OAS1 — — —Oas1b/L1 — — —Oas1g/L2 �33 ccatcctcagGTGGAGGAGA CCTGCTGTGAgcacagcagcOas1a/L3 �33 ccatcctcagGTAGAAGAGA CCTGCTGTGAgcacagcagcOas1f — — —Oas1c — — —Oas1h — — —Oas1e — — —Oas1d — — —
Note. Intron–exon organization in human OAS1 was established by comparing the cDNA sequence (NM-016816) with the genomic sequence (NT-009763).The START and STOP codons are in bold face.
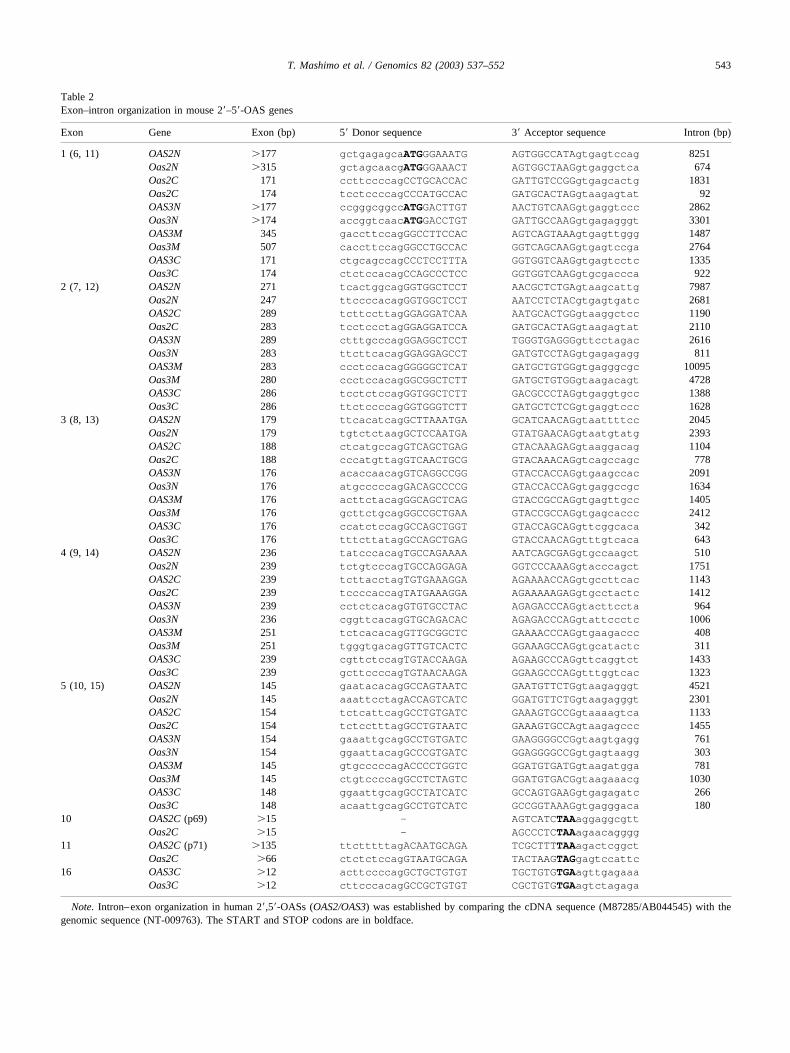
Table 2Exon–intron organization in mouse 2�–5�-OAS genes
Exon Gene Exon (bp) 5� Donor sequence 3� Acceptor sequence Intron (bp)
1 (6, 11) OAS2N �177 gctgagagcaATGGGAAATG AGTGGCCATAgtgagtccag 8251Oas2N �315 gctagcaacgATGGGAAACT AGTGGCTAAGgtgaggctca 674Oas2C 171 ccttccccagCCTGCACCAC GATTGTCCGGgtgagcactg 1831Oas2C 174 tcctccccagCCCATGCCAC GATGCACTAGgtaagagtat 92OAS3N �177 ccgggcggccATGGACTTGT AACTGTCAAGgtgaggtccc 2862Oas3N �174 accggtcaacATGGACCTGT GATTGCCAAGgtgagagggt 3301OAS3M 345 gaccttccagGGCCTTCCAC AGTCAGTAAAgtgagttggg 1487Oas3M 507 caccttccagGGCCTGCCAC GGTCAGCAAGgtgagtccga 2764OAS3C 171 ctgcagccagCCCTCCTTTA GGTGGTCAAGgtgagtcctc 1335Oas3C 174 ctctccacagCCAGCCCTCC GGTGGTCAAGgtgcgaccca 922
2 (7, 12) OAS2N 271 tcactggcagGGTGGCTCCT AACGCTCTGAgtaagcattg 7987Oas2N 247 ttccccacagGGTGGCTCCT AATCCTCTACgtgagtgatc 2681OAS2C 289 tcttccttagGGAGGATCAA AATGCACTGGgtaaggctcc 1190Oas2C 283 tcctccctagGGAGGATCCA GATGCACTAGgtaagagtat 2110OAS3N 289 ctttgcccagGGAGGCTCCT TGGGTGAGGGgttcctagac 2616Oas3N 283 ttcttcacagGGAGGAGCCT GATGTCCTAGgtgagagagg 811OAS3M 283 ccctccacagGGGGGCTCAT GATGCTGTGGgtgagggcgc 10095Oas3M 280 ccctccacagGGCGGCTCTT GATGCTGTGGgtaagacagt 4728OAS3C 286 tcctctccagGGTGGCTCTT GACGCCCTAGgtgaggtgcc 1388Oas3C 286 ttctccccagGGTGGGTCTT GATGCTCTCGgtgaggtccc 1628
3 (8, 13) OAS2N 179 ttcacatcagGCTTAAATGA GCATCAACAGgtaattttcc 2045Oas2N 179 tgtctctaagGCTCCAATGA GTATGAACAGgtaatgtatg 2393OAS2C 188 ctcatgccagGTCAGCTGAG GTACAAAGAGgtaaggacag 1104Oas2C 188 cccatgttagGTCAACTGCG GTACAAACAGgtcagccagc 778OAS3N 176 acaccaacagGTCAGGCCGG GTACCACCAGgtgaagccac 2091Oas3N 176 atgcccccagGACAGCCCCG GTACCACCAGgtgaggccgc 1634OAS3M 176 acttctacagGGCAGCTCAG GTACCGCCAGgtgagttgcc 1405Oas3M 176 gcttctgcagGGCCGCTGAA GTACCGCCAGgtgagcaccc 2412OAS3C 176 ccatctccagGCCAGCTGGT GTACCAGCAGgttcggcaca 342Oas3C 176 tttcttatagGCCAGCTGAG GTACCAACAGgtttgtcaca 643
4 (9, 14) OAS2N 236 tatcccacagTGCCAGAAAA AATCAGCGAGgtgccaagct 510Oas2N 239 tctgtcccagTGCCAGGAGA GGTCCCAAAGgtacccagct 1751OAS2C 239 tcttacctagTGTGAAAGGA AGAAAACCAGgtgccttcac 1143Oas2C 239 tccccaccagTATGAAAGGA AGAAAAAGAGgtgcctactc 1412OAS3N 239 cctctcacagGTGTGCCTAC AGAGACCCAGgtacttccta 964Oas3N 236 cggttcacagGTGCAGACAC AGAGACCCAGgtattccctc 1006OAS3M 251 tctcacacagGTTGCGGCTC GAAAACCCAGgtgaagaccc 408Oas3M 251 tgggtgacagGTTGTCACTC GGAAAGCCAGgtgcatactc 311OAS3C 239 cgttctccagTGTACCAAGA AGAAGCCCAGgttcaggtct 1433Oas3C 239 gcttccccagTGTAACAAGA GGAAGCCCAGgtttggtcac 1323
5 (10, 15) OAS2N 145 gaatacacagGCCAGTAATC GAATGTTCTGgtaagagggt 4521Oas2N 145 aaattcctagACCAGTCATC GGATGTTCTGgtaagagggt 2301OAS2C 154 tctcattcagGCCTGTGATC GAAAGTGCCGgtaaaagtca 1133Oas2C 154 tctcctttagGCCTGTAATC GAAAGTGCCAgtaagagccc 1455OAS3N 154 gaaattgcagGCCTGTGATC GAAGGGGCCGgtaagtgagg 761Oas3N 154 ggaattacagGCCCGTGATC GGAGGGGCCGgtgagtaagg 303OAS3M 145 gtgcccccagACCCCTGGTC GGATGTGATGgtaagatgga 781Oas3M 145 ctgtccccagGCCTCTAGTC GGATGTGACGgtaagaaacg 1030OAS3C 148 ggaattgcagGCCTATCATC GCCAGTGAAGgtgagagatc 266Oas3C 148 acaattgcagGCCTGTCATC GCCGGTAAAGgtgagggaca 180
10 OAS2C (p69) �15 — AGTCATCTAAaggaggcgttOas2C �15 — AGCCCTCTAAagaacagggg
11 OAS2C (p71) �135 ttctttttagACAATGCAGA TCGCTTTTAAagactcggctOas2C �66 ctctctccagGTAATGCAGA TACTAAGTAGgagtccattc
16 OAS3C �12 acttccccagGCTGCTGTGT TGCTGTGTGAagttgagaaaOas3C �12 cttcccacagGCCGCTGTGT CGCTGTGTGAagtctagaga
Note. Intron–exon organization in human 2�,5�-OASs (OAS2/OAS3) was established by comparing the cDNA sequence (M87285/AB044545) with thegenomic sequence (NT-009763). The START and STOP codons are in boldface.
543T. Mashimo et al. / Genomics 82 (2003) 537–552

Fig. 5. Comparison of the predicted amino acid sequences for the Oas1 genes. The alignment of the deduced amino acid sequences of the mouse Oas1 genesand the human OAS1 (NM-016816) was obtained using CLUSTALW with BOXSHADE (http://bioweb.pasteur.fr), in which gaps are marked as dashes andconserved residues are highlighted by gray boxes (significantly) and by black boxes (completely). Potential domains of enzymatic activity are shown above[1,4,25,40,41]. The three conserved aspartic acid residues that catalyze the nucleotidyl transferase reaction are shown by arrows. The stop codon in exon 4of mouse Oas1b/L1 is shown in a shaded box. The downstream sequence (italicized) is not translated in most laboratory strains.

We found two alternatively spliced forms of Oas1a amongthe mRNA molecules transcribed in IFN-induced mouseneural cells (Fig. 4).
The Oas2 and Oas3 genes also showed high conserva-tion of their genomic structure between human and mouse(Table 2 and Fig. 4). Like for human OAS2 [23], the mouseOas2 gene can be transcribed into one of two possibleRNAs by alternative splicing; the first consists of 10 exonsand the other of 11. The Oas3 gene consists of 16 exons,like human OAS3 [24], and no alternatively spliced forms ofOas3 were found.
Sequence comparison of the various Oas1 isoforms
To help in understanding the function(s) of each of theeight Oas1 isoforms, we compared their predicted aminoacid sequences with the sequence of the human orthologousOAS1 gene (Fig. 5), considering that the sequences of thefunctional domains would be the most conserved. Thesedomains are: (i) two � helices and five � strands, which arealso conserved as catalytic domains in the other members ofthe nucleotidyl transferase superfamily [26]; (ii) a P-loopmotif responsible for double-stranded RNA (dsRNA) bind-ing [27]; (iii) three aspartic acid residues that are essentialfor the Mg2�-dependent catalytic activity of the nucleotidyltransferase reaction [4]; (iv) two substrate ATP-bindingsites, in which one serves as acceptor and the other as donor[28]; (v) the highly conserved domain in the 2�,5�-OASgene family [6]; and finally (vi) a CFK motif that is requiredfor tetramerization of OAS1 into its active form [26,27].Comparison of the predicted amino acid sequences indi-cated, on average, a relatively high level of conservation inthe functional domains although a few alterations werenoted with implications for some of the protein functions(see Discussion for details). In addition, as a consequence ofthe presence of a nonsense mutation in exon 4, the last twodomains are both absent in the product of the Oas1b/L1gene in most strains of laboratory mice [9].
Evolutionary relationships among the 2�,5�-OAS genefamily
The sequence-based phylogenetic analysis of each iso-form allowed us to establish the evolutionary relationshipsamong the family of the 2�,5�-OAS genes (Fig. 6). Asexpected, all mouse Oas1 genes and human OAS1 fall intothe same group. The two known rat 2�,5�-OAS genes, ratOas1a (AAC19135) and Oas1b (CAA79317), were shownto be orthologous to mouse Oas1a/Oas1g and Oas1b, re-spectively. It is possible that additional Oas1 genes exist inthe rat, but they have not been identified yet. Since allmouse Oas1 genes fall into the same cluster they can beconsidered as being the consequence of duplications of anancestral copy, after divergence of the primates and rodentsphyla.
The genes encoding mouse OAS-like proteins, Oasl1
(AF1204) and Oasl2 (AB067533), which have been re-cently cloned [7,8,29] and mapped �150 cR centromeric tothe Oas locus in this study (Fig. 1), fall into the same groupas the human OASL gene and chicken 2�,5�-OAS genes(AB002586). The function of these proteins is not yetknown.
Promoter sequences and expression profiles of the genesencoding 2�,5�-OAS
To understand the functions of the genes encoding thevarious isoforms of 2�,5�-OAS, we analyzed the sequencesof their 5� regulatory regions using the transcription factordatabase TRANSFAC. This computational analysis de-tected several regions capable of binding transcriptionalfactors as shown in Fig. 7A. A canonical interferon-stimu-lated responsive element (ISRE) was predicted in the pro-moter regions of Oas1b/L1, Oas1g/L2, Oas1a/L3, andOas2, and binding motifs for basal levels of transcription,such as AP1 and SP1, were found upstream of Oas1c,Oas1h, Oas1e, and Oas1d. From this analysis it is clear thateach of the 10 mouse 2�,5�-OAS genes exhibits a uniquebattery of transcriptional regulatory elements, suggestingthat each of these transcription units has the potential to bedifferentially regulated.
Considering the particular importance of the Oas1b iso-
Fig. 6. Phylogenetic tree based on the comparison of the amino acidsequences of the different proteins encoded in the 2�,5�-OAS cluster ofgenes. GenBank accession numbers of human OAS genes are as follows:OAS1 (NM-016816), OAS2 (M87285), OAS3 (AB044545), OASL (NM-003733). The two domains of human OAS2 (N, amino-terminal; C, car-boxyl-terminal) and the three domains of human OAS3 (N, amino-terminal;M, middle; C, carboxyl-terminal) are delimited as described in Rebouillatet al. [5]. The scale bar indicates an evolutionary distance of 0.1 amino acidsubstitutions per position in the sequence.
545T. Mashimo et al. / Genomics 82 (2003) 537–552

form in the innate defense mechanisms after RNA virusinfection, we sequenced a 520-bp stretch of the promoterregion, upstream of the transcription start site, in sevendifferent inbred strains of mouse, including susceptible andresistant ones. We found several changes at this level (Fig.8B), but none of these SNPs is consistent with the pattern ofresistance or susceptibility for the selected set of strains.
RT-PCR was performed on RNA templates extractedfrom several adult and fetal tissues, either before or afterinduction with dsRNA. Expression of all 10 isoforms in the2�,5�-OAS cluster is represented in Fig. 7B. In this pictureit is clear that the induction of transcription is effective 24 hafter an injection of poly(I,C), although the level of induc-tion for each gene appears variable. Expression was alsodetected in fetal tissues for all genes. Finally, it is interestingto note that some genes showed a slightly different patternof expression according to the tissues analyzed. Oas1b/L1,for example, is readily detectable in the liver and lungs,while Oas1a/L3 and Oas1g/L2 are abundant in the liver,
suggesting that these different isoforms might have differentfunctions.
Discussion
In the present publication we report the completegenomic structure of the cluster of genes encoding 2�,5�-oligoadenylate synthetases in the mouse, with all its tran-scription units, their predicted function, and their evolution-ary relationships. We found that mouse Oas2/Oas3 geneshave a genomic structure very similar to that of humanOAS2/OAS3, while the mouse equivalent of human OAS1exhibits a different organization with no less than eight(Oas1a to Oas1h) tandemly arranged transcription units. Allthese genes are transcribed and we could identify a specifictranscript for each of them. We also found that, for most ofthese isoforms, transcription is regulated by a specific in-ducible promoter since, in the normal cells of the adultorganism, the gene products are in general much less abun-
Fig. 7. (A) Computational analysis of the promoter regions of the different genes in the 2�,5�-OAS cluster. This analysis was performed using the TFSEARCHprogram. (B) Expression profile of the genes encoding 2�,5�-OASs in different tissues. RT-PCR was performed on RNA extracted from several tissues (brain,liver, kidney, lung, and testis) after induction with an injection of poly(I,C) or not, from fetal brain and liver, and from mouse neuroblastoma cells (Neuro2a)grown in vitro. Genomic DNA of BALB/c mice was used as a negative control. The hypoxanthine phosphoribosyltransferase gene (Hprt) was used a controlfor expression.
546 T. Mashimo et al. / Genomics 82 (2003) 537–552

Fig. 8. (A) Comparison of the promoter regions of mouse and human OAS1 genes. The promoter of the Oas1b isoform exhibits an NF-�B binding site thatis closely associated to an ISRE binding site. This association, which has been reported to be capable of triggering gene transcription when bound by somespecific viral RNAs/DNAs [32], might explain the importance of this isoform in the 2�,5�-OAS/RNase L cascade. Oas1b and OAS1 also have in commona GAS-like element at �149 bp that can bind with strong affinity to STAT1 homodimers. (B) Sequence alignments in the promoter region of Oas1b in sevendifferent inbred strains of mouse: MAI/Pas, MBT/Pas (resistant—derived from Mus musculus musculus); STF/Pas, SEG/Pas (resistant—derived from Musspretus); and C57BL/6 (B6), BALB/c (C), and 129/Sv (129), susceptible. SNPs, indicated by asterisks, are either outside the binding sites or not consistentwith the pattern of susceptibility/resistance.
547T. Mashimo et al. / Genomics 82 (2003) 537–552

dant than in cells stimulated by an injection of poly(I,C).Interferon also appears as a strong inducer for these genes.
If we consider that proteins with a structure similar tothat of the 2�,5�-OAS family are found in remotely relatedspecies, it makes sense to believe that these proteins prob-ably have important functions at the cellular level. So far,unfortunately, little is known about these functions and twoquestions can be addressed: (i) why do so many genes existin the mouse, where only one is (apparently) sufficient inhuman? and (ii) do the different isoforms at the Oas1 locushave essentially the same function, or does each isoformhave a specific function? Situations of that kind are notuncommon, in which mouse and human genomes harbororthologous genes in variable copy number, and the olfac-tory receptors that are at least three times more numerous inthe mouse than in the human offer another example [30]. Inthis case it was suggested that processes were operating toshape the mammalian genomes under selective pressures ofthe environment with the mouse having a number (and/orvariety) of receptors greater than that of human, becauseolfaction is more important for survival in this species.
Considering the great similarities at the DNA or proteinsequence levels, it is tempting to consider that most of thetranscription units in the cluster are duplicates, in otherwords extra copies of one, maybe two, essential/ancestralgene(s). However, in a previous study we reported that anonsense mutation in exon 4 of the gene encoding the2�,5�-OAS/L1 isoform was constantly associated with thesusceptibility of laboratory mice to an experimental infec-tion with a pathogenic strain of West Nile virus (flavivirus),suggesting that the inactivation of this isoform is the directcause of the susceptibility [9,10]. If this observation isconfirmed this would indicate that at least one of the iso-forms in the OAS cluster—Oas1b—has a specific functionthat cannot be performed by the others. The susceptibility oflaboratory mice to West Nile virus, and presumably to allother members of the family Flaviviridae, was discoveredbecause mice were experimentally infected with viruses ofthis kind. One may then hypothesize that each isoform in theOAS cluster has its own specificity, making the animalresistant to several more or less related RNA viruses.
According to Perelygin and colleagues [10], the flavivi-rus-specific activity of the Oas1b isoform on viral replica-tion might be correlated to a 4-amino-acid deletion in theP-loop motif that is found only in the sequence of thisisoform and does not appear to exist in either rat or human.This 4-aa deletion would allow the Oas1b protein to recog-nize and bind a specific conserved RNA structure unique toflavivirus RNAs. Even if this hypothesis is supported byother experiments in vitro [31], an alternative explanation toOas1b specific activity on flavivirus replication may also befound in its promoter sequence, in which several bindingsites (NF-�B, GAS, and ISRE) exhibit a unique organiza-tion (Fig. 8). In particular, it is noteworthy that Oas1b is theonly gene in which the two binding sites NF-�B and ISREare closely associated in tandem, producing a genomic
structure that has previously been reported as capable oftriggering gene expression upon viral induction [32].
Sequencing the promoter regions of the Oas1b isoformin remotely related mouse species did not provide evidencethat a particular structural change in this promoter might beassociated with the phenotype of resistance or susceptibilityafter flavivirus infection. This substantiates the hypothesisthat the stop codon found in the Oas1b coding sequence ofmost laboratory strains is related directly to this phenotype.
The human OAS1 gene encodes four different OAS pro-teins (p42, p44, p46, and p48) resulting from alternativesplicing of the first five exons with the three exons of theC-terminal region (see Fig. 4 for details) [6]. So far, we haveidentified alternatively spliced transcripts only for themouse Oas1a/L3 isoform. As in human these two tran-scripts are generated through alternative splicing of a part ofthe sixth exon and we cannot rule out the existence of otherforms. To elucidate this point other exhaustive screenings ofcDNA libraries prepared from cells induced by interferonwould be necessary. However, it must be noted that one ofthe alternatively spliced isoforms of OAS1, the 9-2 isozyme,which contains a BH3 domain at its C-terminal end and canbind to antiapoptotic proteins, thus promoting cellular apo-ptosis [33], has not been found in the mouse. Finally, wefound that two alternatively spliced isoforms were encodedin mouse Oas2, a finding that is consistent with the exis-tence of human isoforms (p69 and p71) encoded in humanOAS2 [23], and like in human, we did not identify anyalternatively spliced forms from mouse Oas3 [24].
Alignment of the predicted amino acid sequences for theproteins encoded by the eight Oas1 genes indicates thatOas1c, Oas1d, Oas1e, Oas1f, and Oas1h are structurallysimilar and lack functional domains, such as the LXXXPAmotif [27], the highly conserved aspartic acid residues inexon 2 [4], and the CFK motif [27]. These observationssuggest that if these isoforms have retained their bindingactivity to dsRNA, they have lost their Mg2�-dependentcatalytic activity [4,22] and accordingly they may actuallybe inactive pseudogenes rather than real isoforms. In con-trast, Oas1g/L2 and Oas1a/L3 encode proteins that could befunctional in the 2�,5�-OAS/RNase L cascade.
Finally, and strangely, we noted that despite the remoteorigin of this family of proteins no sequences related tothose of 2�,5�-OAS genes could be identified either in Cae-norhabditis elegans or in Drosophila melanogaster basedon a BLAST search.
Material and methods
Mouse strains and crosses
In our experiments, we used the 219 progenies of fourintersubspecific backcrosses set from a combination of fourdifferent mouse inbred strains: the two classical laboratorystrains BALB/c and C57BL/6 on the one hand and two
548 T. Mashimo et al. / Genomics 82 (2003) 537–552

inbred strains derived from recently trapped wild progeni-tors of the Mus musculus musculus subspecies on the other.All these four strains are highly inbred. BALB/c andC57BL/6 are susceptible to experimental infections with thehighly virulent strain of West Nile flavivirus IS-98-ST1,while MAI/Pas and MBT/Pas are resistant [9].
Drawing up the genetic and radiation hybrid map of themouse chromosome 5 region, containing theOAS-encoding cluster of genes
For the establishment of the genetic map encompassingthe Oas locus, we analyzed the genetic constitution of a totalof 203 mice: 81 from the (MBT/Pas � BALB/c)F1 �BALB/c backcross, 55 from the (MBT/Pas � C57BL/6)F1� C57BL/6 backcross, 25 from the (MAI/Pas � BALB/c)F1 � BALB/c backcross, and 42 from the (MAI/Pas �C57BL/6)F1 � C57BL/6 backcross. Genomic DNA wasextracted from the tail tip of each of these mice using aDNA extraction kit (Qiagen DNeasy Tissue Kit) and then,each DNA sample was genotyped using a set of geneticmarkers known to be polymorphic in the critical region.Polymorphism was revealed either by analysis of single-strand length polymorphism (SSLP) or single-strand con-formation polymorphism (SSCP) (GenGel Excel 12.5/24Kit and Automated Silver Staining, Americium PharmacyBiotech).
The sources of markers were MGI-Chromosome Com-mittee Report, chromosome 5; TJL RH Database, chromo-some 5; and mouse ESTs/genes positive after a BLASTanalysis (BLAST server at the National Center for BiologyInformation [34]) with genes localized in the region ofconserved synteny in human (12q24.1–q24.2). Microsatel-lite markers that were polymorphic (i) by SSLP, D5Mit17,D5Mit239, D5Mit158, D5Mit406, D5Mit319, D5Mit407,D5Mit68, D5Mit431, D5Mit368, D5Mit242, D5Mit369,D5Mit95, D5Mit96, D5Mit426, D5Mit321, D5Mit247, and(ii) by SSCP, D5Mit159, were ordered first in the Oasregion. Markers listed in Table 3 were later ordered in thesame region.
PCR amplification was carried out with the followingconditions: 94°C for 5 min for 1 cycle; 94°C for 1 min,55°C for 1 min, 72°C for 1 min for 30 cycles. PCR productswere electrophoresed on 4% agarose gels and visualizedwith ethidium bromide staining. Genotype data were ana-lyzed using the MapManager QTb28 program [35] to makea genetic linkage map.
RH panel clones were purchased from Research Genet-ics. Typing of each clone for Ung, Oasrp, Nos1, Oasl,Ptpn11, Aldh2, Eif2b, and Ubc markers (Table 3) was de-termined by PCR analysis in duplicate. Typing data of othermarkers was obtained from The Jackson Laboratory (http://www.jax.org) and combined with our data using the Map-Manager QTb28 program [35].
Table 3Primers used for genetic and physical mapping
Marker GenBank Size (bp) Forward primer Reverse primer
Markers used to construct genetic and RH maps
Golge3 AB029526 SSLP 253 CCTCCCAGTTCAGTCTCTAA ATTCCCTTGTCTCAGTTTTCNos1 AF135825 SSLP 271 TACCCAGCATCTCTCACCAG AAGGACCTAAGTTCAAAAGCUng Y08975 STS 110 TTTTGTCTGGAAAGGAACCC ACTGTGAACCCAGGGAGAACOas1 (Oas12) AF068835 STS 114 TCCCCGACCCCCAACTCTAAAC GCTGAGGGAAGGATGTTGAGGAOas (Oas1a) AI449562 STS 94 GTGGACAGATGGACACCAAG CAGCCATGTTTGACTCCTGTPtpn11 D84372 STS 158 AGACTAGACGAGCGTTCCCA TGTCAAGAACGCAGAGCAGTA1dh2 U07235 STS 123 AAGAACTCGTAAAGCGGCAT AAGACACTTTGGTGAAGGGGEif2b AA266894 STS 187 CAGGAAGGACAACTGAGATTACATC CTCCTGTCTGGCTTGGGCTGCTGAGUbc AI194771 STS 135 CCATCACACCCAAGAACAAG CTGACAGGCAAGACCATCAC
STS markers used to construct the BAC contig
AI853657 AI853657 STS 143 ATCTCAAGGACCCCAAACAC ATAGAGACAACGGTCCCCAGDtx1 AW556360 STS 128 GCTCAGCTTGATGCGTGTAT AAGACATCGTCCTTGCCTCTOas1b/L1 AC015535 STS 418 ACACAGTGTCCATCTCAACCA TGTTGGTGCAGGTATTCAGAGACIap M10062 STS 1053 GGGGAGAGAGAGTAGGAACAAAGAG GCTCTGACTCTCCAAAAATACGCTCOas1a/L3 AI449562 STS 94 GTGGACAGATGGACACCAAG CAGCCATGTTTGACTCCTGTRph3a D29965 STS 214 CTTTCATGTCTCCCCCACTTCA TATAATTTCTGCCCTCCTCCTG226D6SP6 AZ702819 STS 130 TCTTTATTCCCAACATTCAGG TTCCCCGTCTTTTCATCTTCC284E21SP6 AZ035255 STS 357 GGGGGATGAGGAGCAGACAGC CAAGCAAAGGAGGCAAAATGG39M18SP6 AZ248212 STS 119 TCCCTGGGGACTGTATTTGTGA TATTTGAGATGTAGGGGGTTCC284E21T7 AQ978022 STS 150 TGGATGGCAGAGCCTGAGAAG CCATAAAAGTGAAAACACAAG8A9SP6 AZ093599 STS 218 GCAGCTTCTCCTTGTTCTTGG GTTTGTTCCCTTTTGTGTTTG7J4T7 AZ091372 STS 146 CCTTCCAAGTAGCCAGTTTCT GACAACGAGGTCAGCATAGGC134M5T7 AZ277309 STS 115 CAGCCAGGGTTGATTTTATTA CCTACCAGAGCCAGTGTTGTG
BAC end sequences are available from the TIGR database at http://www.tigr.org/tdb/bac_ends/mouse/bac_end_intro.html. SSLP, simple sequence lengthpolymorphism; STS, sequence tagged site.
549T. Mashimo et al. / Genomics 82 (2003) 537–552

Identification of BAC clones from libraries
RPCI-23 (C57BL/6) and RPCI-24 (129/Sv) mouse BAClibraries from Research Genetics were screened using PCRprobes of Oas3 (M33863), Ptpn11 (D84372), and39M18SP6 (AZ248212) (see Fig. 2 and Results).
BAC DNA isolation was performed with a Plasmid MidiPrep procedure with modifications suggested by the manu-facturer (Qiagen, Inc.). DNA from BACs with long inserts(180–230 kb), from RPCI-23 and RPCI-24 mouse BAClibrary, was prepared using the Nucleobond AX500 (Mach-erey–Nagel) protocol. For shotgun sequencing of BACclone RPCI-23-226D6, bacterial DNA-free BAC DNA wasisolated using the Qiagen Large-Construct Kit.
BAC DNA was digested with NotI restriction enzymesand resolved by pulsed-field gel electrophoresis (Gene Nav-igator electrophoresis unit; Pharmacia) under the followingconditions: 1% Gigaphor agarose (Eurobio) gel in 0.5�TBE buffer, 190 V, for 3 � 6 h at 14°C using successively5-, 10-, and 15-s switch times. DNA in gels was transferredto Hybond-N� membranes (Amersham) by conventionalSouthern blotting and hybridized with radiolabeled probes.
Generation of novel sequence-tagged sites
Sequences for STS-content mapping were derived frommouse ESTs/genes positive after a BLAST analysis(BLAST server at the National Center for Biology Informa-tion (NCBI) [34]) with genes localized in the region ofconserved synteny in human (12q24.1–q24.2) or BAC-endsequences that are available online at the TIGR site, forsome RPCI-23 library BAC clones (http://www.tigr.org/tdb/bac_ends/mouse/bac_end_intro.html).
We performed shotgun sequencing of the BAC (RPCI-22-226D6), which contains the Oas cluster and the Dtx1gene. Briefly, 30 �g of BAC DNA isolated as describedabove was sheared by nebulization, end-repaired using T4DNA polymerase, and ligated to BstXI adaptors (Invitro-gen). The ligation mixture was fractionated by agarose gelelectrophoresis and fragments from 1 to 3 kb were ligatedinto BstXI-digested pcDNA2.1 (Invitrogen). The plasmids
were used as templates for cycle sequencing reactions con-sisting in 35 cycles (96°C for 30 s, 50°C for 15 s, 60°C for4 min) in a thermocycler. Samples were precipitated andloaded onto an ABI 3700 automatic capillary DNA se-quencer (Applied Biosystems). Shotgun clone sequencesfrom both ends were assembled into eight contigs and editedusing the PHRED [36,37], PHRAP (P. Green, unpublished),and CONSED [38] softwares.
Cloning of the murine 2�-5�-OAS genes
To prepare mRNA induced by IFN, we used primarycultured neurons derived from the brain of a 14-day mouseembryo as previously described [9]. Briefly, the brain wasdissected, minced, and subsequently plated onto tissue cul-ture dishes. The cells were induced by interferon-� in dosesof 20 IU/ml (GIBCO BRL). Total RNA was extracted fromthe induced cells using the RNAxel Kit (Eurobio). First-strand cDNA was synthesized with oligo(dT) primers usingreverse transcriptase (GIBCO BRL) and the followingcDNA amplification was performed using the AdvantageHigh-Fidelity PCR Kit (Clontech) with PCR primers aslisted in Table 4. Amplified products were gel purified andcloned into a dT-tailed pCR4-TOPO vector (Invitrogen). Atleast three independent clones for each cDNA fragmentswere sequenced.
Sequence alignment and phylogenetic analysis
The deduction of open reading frames in the sequence ofBAC clones was done using the GENSCAN program(http://bioweb.pasteur.fr/) or by BLAST search with amouse EST database through the NCBI Web servers (http://www.ncbi.nlm.nih.gov/). Sequence alignment of each Oasgene was performed with the CLUSTALW software (http://bioweb.pasteur.fr/). The phylogenetic tree was constructedusing the PHYLIP program with the parsimony method forprotein sequences (http://bioweb.pasteur.fr/). Graphical out-put of the bootstrap figure was produced by DRAWTREE(http://bioweb.pasteur.fr/).
Table 4Primers used to amplify specifically the cDNA of murine Oas genes
Gene Forward Reverse Size (bp)
Oas1b/L1 AGGTAAAAGCTGGACCTAGG CTGTCTGTGCAGGGGTTGAGG 1447Oas1g/L2 GGAGACCCAGGAAGCTCCAGA TTGAGGAAGGCTGGCTGTGATTG 1294Oas1a/L3 GGAGACCCAGGAAGCTCCAGA TTGAGGAAGGCTGGCTGTGATTG 1294Oas1f TGGCCATAAAATGGTGAAGGAT TCATATCCCTGACTCTTGAGCA 1148Oas1c AACACTCCTGGCCTCAGGATG AGGTCTTGCAGCAGAGGAAGGT 1171Oas1h TGCAGTCAGCAAACATTCCTG TCTCCTTGGCAGATGCTTTGG 1288Oas1e AACACTCCTGGCCTCAGGATG CCTCTCTAGGTCAAAGTCTTAC 1181Oas1d AACACTCCTGGCCTCAGGATG AGGTCTTACAGAAGAGGAGGAC 1169Oas2 AACGATGGGAAACTGGCTGAC GGTAGATCTGACCTCCACAGC 2326Oas3 TACTGAGAGTACCGGTCAACA ACACAGACATACCACAAATAC 3570
550 T. Mashimo et al. / Genomics 82 (2003) 537–552

Identification of regulatory elements and expressionanalysis
Identification of transcriptional binding factors in the 5�regulatory region was performed using the TFSEARCHprogram (http://www.tfsearch.com) with the TRANSFACdatabase. A thousand base pairs from the predicted tran-scription initiation site were analyzed. Comparison of thepromoter sequence intra- and interspecies also provided theprediction of regulatory elements.
Expression profiles of several different tissues were in-vestigated through brain, liver, kidney, lung, and testis,either before or after induction of interferon with an injec-tion of poly(I,C) (Amersham). Male BALB/c mice wereinjected intravenously with 200 �g of poly(I,C) prepared asdescribed by Flenniken et al. [39]. Control animals wereinjected with phosphate-buffered saline. Animals were sac-rificed at 24 h after the injection. RT-PCR amplification wascarried out under the following conditions: 94°C for 1 min,60°C for 1 min, 72°C for 1 min for 35 cycles. Primers usedfor RT-PCR are listed in Table 5.
Acknowledgments
We thank our colleagues in the Transverse ResearchProgram 21 for their interest in this project. We also thankSoline Vigneau for technical help in analyzing the promoterregion and Gordon Langsley for critically reading the manu-script. This work was funded by the Transverse ResearchProgram network (TRP21, Pasteur Institute) and Programmede Recherche Fondamentale en Microbiologie et MaladiesInfectieuses et Parasitaires, Ministere de l’Education Natio-nale, de la Recherche et de la Technologie, France.
References
[1] A Yamamoto, et al., Two types of chicken 2�,5�-oligoadenylate syn-thetase mRNA derived from alleles at a single locus, Biochim. Bio-phys. Acta 1395 (1998) 181–191.
[2] P.J. Cayley, R.F. White, J.F. Antoniw, N.J. Walesby, I.M. Kerr,Distribution of the ppp(A2�p)nA-binding protein and interferon-re-
lated enzymes in animals, plants, and lower organisms, Biochem.Biophys. Res. Commun. 108 (1982) 1243–1250.
[3] M. Wiens, A. Kuusksalu, M. Kelve, W.E. Muller, origin of theinterferon-inducible (2�-5�)oligoadenylate synthetases: cloning of the(2�-5�)oligoadenylate synthetase from the marine sponge Geodia cy-donium, FEBS Lett. 462 (1999) 12–18.
[4] S.N. Sarkar, A. Ghosh, H.W. Wang, S.S. Sung, G.C. Sen, The natureof the catalytic domain of 2�-5�-oligoadenylate synthetases, J. Biol.Chem. 274 (1999) 25535–25542.
[5] D. Rebouillat, A.G. Hovanessian, The human 2�,5�-oligoadenylatesynthetase family: interferon-induced proteins with unique enzymaticproperties, J. Interferon Cytokine Res. 19 (1999) 295–308.
[6] J. Justesen, R. Hartmann, N.O. Kjeldgaard, Gene structure and func-tion of the 2�-5�-oligoadenylate synthetase family, Cell Mol. Life Sci.57 (2000) 1593–1612.
[7] S. Eskildsen, R. Hartmann, N.O. Kjeldgaard, J. Justesen, Gene struc-ture of the murine 2�-5�-oligoadenylate synthetase family, Cell Mol.Life Sci. 59 (2002) 1212–1222.
[8] S. Kakuta, S. Shibata, Y. Iwakura, Genomic structure of the mouse2�,5�-oligoadenylate synthetase gene family, J. Interferon CytokineRes. 22 (2002) 981–993.
[9] T. Mashimo, et al., A nonsense mutation in the gene encoding2�-5�-oligoadenylate synthetase/L1 isoform is associated with WestNile virus susceptibility in laboratory mice, Proc. Natl. Acad. Sci.USA 99 (2002) 11311–11316.
[10] A.A. Perelygin, et al., Positional cloning of the murine flavivirusresistance gene, Proc. Natl. Acad. Sci. USA 99 (2002) 9322–9327.
[11] A. Mitra, et al., A mammalian 2-5A system functions as an antiviralpathway in transgenic plants, Proc. Natl. Acad. Sci. USA 93 (1996)6780–6785.
[12] T. Ogawa, T. Hori, I. Ishida, Virus-induced cell death in plantsexpressing the mammalian 2�,5� oligoadenylate system, Nat. Biotech-nol. 14 (1996) 1566–1569.
[13] G. Rysiecki, D.R. Gewert, B.R. Williams, Constitutive expression ofa 2�,5�-oligoadenylate synthetase cDNA results in increased antiviralactivity and growth suppression, J. Interferon Res. 9 (1989) 649–657.
[14] B.A. Hassel, P.O. Ts’o, A sensitive assay for the IFN-regulated 2-5Asynthetase enzyme, J. Virol. Methods 50 (1994) 323–334.
[15] J.C. Castelli, et al., A study of the interferon antiviral mechanism:Apoptosis activation by the 2-5A system, J. Exp. Med. 186 (1997)967–972.
[16] S. Salzberg, et al., Ectopic expression of 2-5A synthetase in myeloidcells induces growth arrest and facilitates the appearance of a myeloiddifferentiation marker, Cancer Res. 57 (1997) 2732–2740.
[17] A. Zhou, et al., Interferon action and apoptosis are defective in micedevoid of 2�,5�-oligoadenylate-dependent RNase L, EMBO J. 16(1997) 6355–6363.
[18] F. Bonhomme, J.L. Guenet, The laboratory mouse and its wild rela-tives, in: M.F. Lyon, S. Rastan, S.D.M. Brown (Eds.), In Genetic
Table 5Primers used for expression analysis (RT-PCR) of individual Oas genes
Gene Forward Reverse Size (bp)
Oas1b/L1 AGGCTGCCGCCTGGCTGCAAT TAAGGCAGGAGGATGGCAATA 213Oas1g/L2 ATGGTAGTATCAATAAGAAGC GCATAGACAGTGAGTAGCTCC 247Oas1a/L3 ATGTTAATACTTCCAGCAAGC GCAAAGACAGTGAGCAACTCT 247Oas1f CTGGGGTGTGCCGATGAAGGT GACCGGATGCAGGGGTGAGAA 226Oas1c AACATTCGAATAAAAATCAAG TCTTGTCTCTCTGGCGACAAA 334Oas1h GGATGTGCCGCCAGACAAAAA GAAGTCAAATCTCTGTCCATC 293Oas1e TGATATGTCCCTTGTGAATGG CCTTCTAGGCAAGTGCTACAC 88Oas1d TCGACTTTCTTTACGAACCAT CCTAGGCAGATGCTGGGGGTGC 105Oas2 GTGGCACAGATGACTTTGACA TCAGCAATCCCAACCTACTAC 392Oas3 GAGGGCTTCCGCACTGTTCTG TGTCACTTTCTATCAATGGTG 329
551T. Mashimo et al. / Genomics 82 (2003) 537–552

Variants and Strains of the Laboratory Mouse, Oxford Univ. Press,Oxford, 1996, pp. 1577–1596.
[19] K. Osoegawa, et al., Bacterial artificial chromosome libraries formouse sequencing and functional analysis, Genome Res. 10 (2000)116–128.
[20] M.N. Rutherford, A. Kumar, A. Nissim, J. Chebath, B.R. Williams,The murine 2-5A synthetase locus: Three distinct transcripts fromtwo linked genes, Nucleic Acids Res. 19 (1991) 1917–1924.
[21] Y. Ichii, R. Fukunaga, S. Shiojiri, Y. Sokawa, Mouse 2-5A synthetasecDNA: Nucleotide sequence and comparison to human 2-5A syn-thetase, Nucleic Acids Res. 14 (1986) 10117.
[22] S. Shibata, S. Kakuta, K. Hamada, Y. Sokawa, Y. Iwakura, Cloningof a novel 2�,5�-oligoadenylate synthetase-like molecule, Oasl5 inmice, Gene 271 (2001) 261–271.
[23] I. Marie, A.G. Hovanessian, The 69-kDa 2-5A synthetase is com-posed of two homologous and adjacent functional domains, J. Biol.Chem. 267 (1992) 9933–9939.
[24] D. Rebouillat, A. Hovnanian, G. David, A.G. Hovanessian, B.R. Wil-liams, Characterization of the gene encoding the 100-kDa form of human2�,5� oligoadenylate synthetase, Genomics 70 (2000) 232–240.
[25] A. Chang-Yeh, D.E. Mold, M.H. Brilliant, R.C. Huang, The mouseintracisternal A particle-promoted placental gene retrotransposition ismouse-strain-specific, Proc. Natl. Acad. Sci. USA 90 (1993) 292–296.
[26] S.N. Sarkar, S. Bandyopadhyay, A. Ghosh, G.C. Sen, Enzymaticcharacteristics of recombinant medium isozyme of 2�-5� oligoadeny-late synthetase, J. Biol. Chem. 274 (1999) 1848–1855.
[27] A. Ghosh, S.N. Sarkar, W. Guo, S. Bandyopadhyay, G.C. Sen, En-zymatic activity of 2�-5�-oligoadenylate synthetase is impaired byspecific mutations that affect oligomerization of the protein, J. Biol.Chem. 272 (1997) 33220–33226.
[28] S.N. Sarkar, M. Miyagi, J.W. Crabb, G.C. Sen, Identification of thesubstrate-binding sites of 2�-5�-oligoadenylate synthetase, J. Biol.Chem. 277 (2002) 24321–24330.
[29] M. Tiefenthaler, et al., M1204, a novel 2�,5� oligoadenylate syn-thetase with a ubiquitin-like extension, is induced during maturationof murine dendritic cells, J. Immunol. 163 (1999) 760–765.
[30] J.M. Young, B.J. Trask, The sense of smell: genomics of vertebrateodorant receptors, Hum. Mol. Genet. 11 (2002) 1153–1160.
[31] N. Urosevic, et al., Development and characterization of new flavi-virus-resistant mouse strains bearing Flv(r)-like and Flv(mr) allelesfrom wild or wild-derived mice, J. Gen. Virol. 80 (1999) 897–906.
[32] G. Cheng, A.S. Nazar, H.S. Shin, P. Vanguri, M.L. Shin, IP-10 genetranscription by virus in astrocytes requires cooperation of ISRE withadjacent kappaB site but not IRF-1 or viral transcription, J. InterferonCytokine Res. 18 (1998) 987–997.
[33] A. Ghosh, S.N. Sarkar, T.M. Rowe, G.C. Sen, A specific isozyme of2�-5� oligoadenylate synthetase is a dual function proapoptotic pro-tein of the Bcl-2 family, J. Biol. Chem. 276 (2001) 25447–25455.
[34] S.F. Altschul, et al., Gapped BLAST and PSI-BLAST: A new gen-eration of protein database search programs, Nucleic Acids Res. 25(1997) 3389–3402.
[35] K.F. Manly, J.M. Olson, Overview of QTL mapping software andintroduction to Map Manager QT, Mamm. Genome 10 (1999) 327–334.
[36] B. Ewing, L. Hillier, M.C. Wendl, P. Green, Base-calling of auto-mated sequencer traces using PHRED. I. Accuracy assessment, Ge-nome Res. 8 (1998) 175–185.
[37] B. Ewing, P. Green, Base-calling of automated sequencer traces usingPHRED, II. Error probabilities. Genome Res. 8 (1998) 186–194.
[38] D. Gordon, C. Abajian, P. Green, CONSED: A graphical tool forsequence finishing, Genome Res. 8 (1998) 195–202.
[39] A.M. Flenniken, J. Galabru, M.N. Rutherford, A.G. Hovanessian,B.R. Williams, Expression of interferon-induced genes in differenttissues of mice, J. Virol. 62 (1988) 3077–3083.
[40] N. Kon, R.J. Suhadolnik, Identification of the ATP binding domain ofrecombinant human 40-kDa 2�,5�-oligoadenylate synthetase by pho-toaffinity labeling with 8-azido-[alpha-32P]ATP, J. Biol. Chem. 271(1996) 19983–19990.
[41] I. Marie, D. Rebouillat, A.G. Hovanessian, The expression of bothdomains of the 69/71 kDa 2�,5� oligoadenylate synthetase generates acatalytically active enzyme and mediates an anti-viral response, Eur.J. Biochem. 262 (1999) 155–165.
552 T. Mashimo et al. / Genomics 82 (2003) 537–552
Related Documents











