418 Accepted by B. Rossaro: 24 Dec. 2013; published: 12 Feb. 2014 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2014 Magnolia Press Zootaxa 3764 (4): 418–426 www.mapress.com/zootaxa/ Article http://dx.doi.org/10.11646/zootaxa.3764.4.2 http://zoobank.org/urn:lsid:zoobank.org:pub:72B9303F-EAAA-4074-AF98-01C7B4F49453 A new species of sponge-dwelling Oukuriella (Chironomidae) from Brazil LÍVIA MARIA FUSARI 1 , CAROLINA FERRAZ BELLODI 2 , & CARLOS JOSÉ EINICKER LAMAS 1 1 Universidade de São Paulo, Museu de Zoologia - MZUSP, Laboratório de Diptera, São Paulo, SP, Brazil. C.P.: 42494. E-mail: [email protected]; [email protected] 2 Programa de Pós-Graduação em Entomologia e Conservação da Biodiversidade, Universidade Federal da Grande Dourados - UFGD, Dourados, MS, Brazil. E-mail: [email protected] Abstract Oukuriella Epler, 1986 is an exclusively Neotropical genus that comprises two well supported clades: inhabitants of freshwater sponges and inhabitants of immersed wood (saproxylophilic). In this work, males of a new chironomid species, Oukuriella froehlichi sp. n., are described and illustrated. Specimens were collected near stream in the Brazilian Atlantic Forest. Phylogenetic analyses performed in this study show that the newly discovered species belongs in the clade of freshwater sponge inhabitants. Key words: Neotropics, Brazil, Chironomidae, freshwater sponges Introduction The genus Oukuriella, exclusively found in the Neotropical region, was originally erected by Epler (1986) based on the morphology of O. albistyla, O. fasciata and O. simulatrix. This group is characterized by the absence of the anal point and bifid superior volsella. Several new species of Oukuriella were subsequently described by Epler (1996), Messias & Fittkau (1997), Messias (1998, 2000), Messias & Oliveira (1998), Messias et al. (2000), Trivinho-Strixino & Messias (2005) and Fusari et al. (2008; 2009). Oukuriella was recently reviewed by Fusari et al. (2013) and six new species were added to genus. So, Oukuriella comprises 21 valid species, but the immature stages are known from only five species: O. epleri Messias et Fittkau, 1997, O. intermedia Messias, Fittkau et Oliveira 2000, O. pesae Fusari, Roque et Hamada, 2009, O. pinhoi Fusari, Roque et Hamada, 2013 and O. jatai Trivinho-Strixino et Messias, 2005. The phylogeny of the genus was recently inferred based on the morphology of adult males (Fusari et al. 2013). Topologies obtained in this study indicated that the chironomids found on freshwater sponges and immersed wood constitute two major sister clades within Oukuriella, in addition there are other basal branches with unpredicted habitat. The evidence of a correlation between evolutionary relationships and life histories allows to infer the habitat of a larva basing on the placement of a species in the phylogenetic tree. Material and methods Examined material. Chironomids were collected in ethanol 70%, and deposited in the Museu de Zoologia da Universidade de São Paulo, São Paulo, Brazil (MZUSP). Specimens were slide-mounted using Euparal, following the procedures outlined by Sæther (1969). The terminology follows Sæther (1980). Measurements were made according to the method suggested by Sæther (1968) and Schlee (1966). The measurements are given with the observed range, followed by the mean when more than five specimens are measured. Phylogenetic analysis. Phylogenetic analysis was carried out include Oukuriella froehlichi sp. n. in the phylogenetic tree. The data matrix was evaluated under the maximum parsimony criterion using the TNT software

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2014 Magnolia Press
Zootaxa 3764 (4): 418–426
www.mapress.com/zootaxa/Article
http://dx.doi.org/10.11646/zootaxa.3764.4.2
http://zoobank.org/urn:lsid:zoobank.org:pub:72B9303F-EAAA-4074-AF98-01C7B4F49453
A new species of sponge-dwelling Oukuriella (Chironomidae) from Brazil
LÍVIA MARIA FUSARI1, CAROLINA FERRAZ BELLODI2, & CARLOS JOSÉ EINICKER LAMAS1 1Universidade de São Paulo, Museu de Zoologia - MZUSP, Laboratório de Diptera, São Paulo, SP, Brazil. C.P.: 42494.
E-mail: [email protected]; [email protected] 2Programa de Pós-Graduação em Entomologia e Conservação da Biodiversidade, Universidade Federal da Grande Dourados -
UFGD, Dourados, MS, Brazil. E-mail: [email protected]
Abstract
Oukuriella Epler, 1986 is an exclusively Neotropical genus that comprises two well supported clades: inhabitants of
freshwater sponges and inhabitants of immersed wood (saproxylophilic). In this work, males of a new chironomid species,
Oukuriella froehlichi sp. n., are described and illustrated. Specimens were collected near stream in the Brazilian Atlantic
Forest. Phylogenetic analyses performed in this study show that the newly discovered species belongs in the clade of
freshwater sponge inhabitants.
Key words: Neotropics, Brazil, Chironomidae, freshwater sponges
Introduction
The genus Oukuriella, exclusively found in the Neotropical region, was originally erected by Epler (1986) based on the morphology of O. albistyla, O. fasciata and O. simulatrix. This group is characterized by the absence of the anal point and bifid superior volsella. Several new species of Oukuriella were subsequently described by Epler (1996), Messias & Fittkau (1997), Messias (1998, 2000), Messias & Oliveira (1998), Messias et al. (2000), Trivinho-Strixino & Messias (2005) and Fusari et al. (2008; 2009). Oukuriella was recently reviewed by Fusari et
al. (2013) and six new species were added to genus. So, Oukuriella comprises 21 valid species, but the immature stages are known from only five species: O. epleri Messias et Fittkau, 1997, O. intermedia Messias, Fittkau et
Oliveira 2000, O. pesae Fusari, Roque et Hamada, 2009, O. pinhoi Fusari, Roque et Hamada, 2013 and O. jatai
Trivinho-Strixino et Messias, 2005.The phylogeny of the genus was recently inferred based on the morphology of adult males (Fusari et al. 2013).
Topologies obtained in this study indicated that the chironomids found on freshwater sponges and immersed woodconstitute two major sister clades within Oukuriella, in addition there are other basal branches with unpredicted habitat. The evidence of a correlation between evolutionary relationships and life histories allows to infer the habitat of a larva basing on the placement of a species in the phylogenetic tree.
Material and methods
Examined material. Chironomids were collected in ethanol 70%, and deposited in the Museu de Zoologia da Universidade de São Paulo, São Paulo, Brazil (MZUSP). Specimens were slide-mounted using Euparal, following the procedures outlined by Sæther (1969). The terminology follows Sæther (1980). Measurements were made according to the method suggested by Sæther (1968) and Schlee (1966).
The measurements are given with the observed range, followed by the mean when more than five specimens are measured.
Phylogenetic analysis. Phylogenetic analysis was carried out include Oukuriella froehlichi sp. n. in the phylogenetic tree. The data matrix was evaluated under the maximum parsimony criterion using the TNT software
418 Accepted by B. Rossaro: 24 Dec. 2013; published: 12 Feb. 2014

(Goloboff et al. 2003). Characters were coded for O. froehlichi sp. n. and added as a new terminal to the Oukuriella
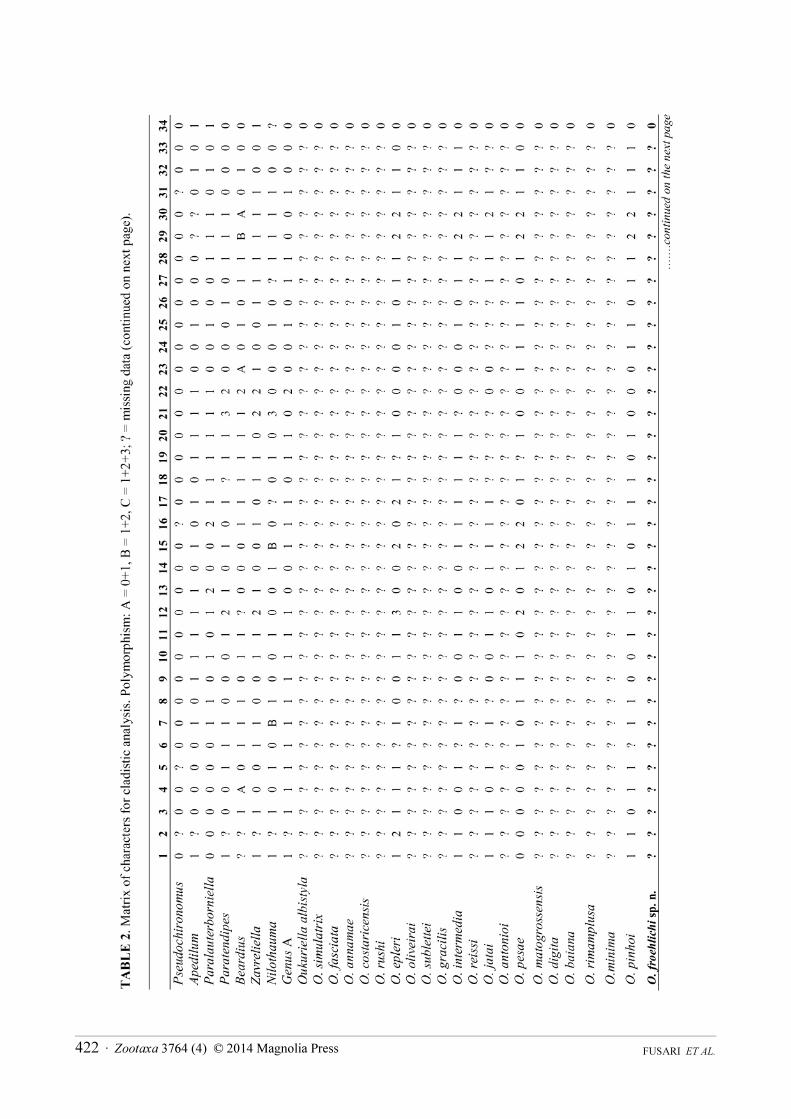
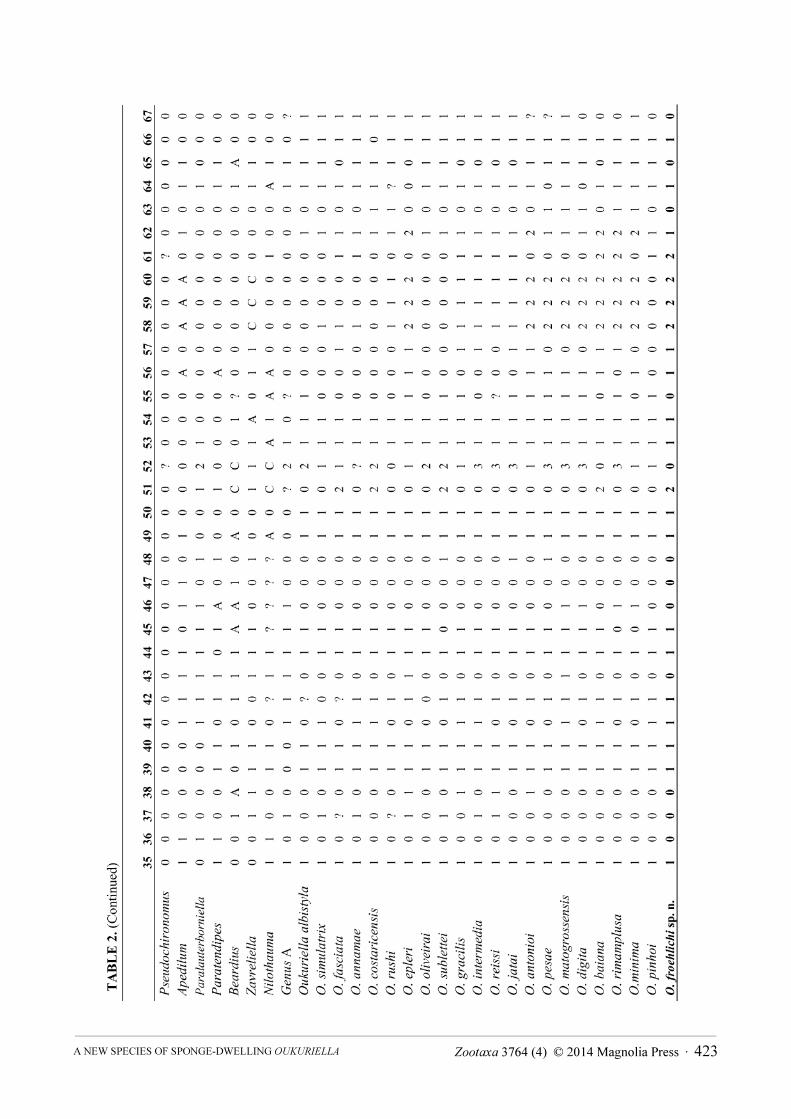
matrix, originally used in Fusari et al. (2013), into a Nexus Data Editor (NDE version 0.5.0; Page 2001) file. The original analyses of Fusari et al. (2013) was implemented based on 67 morphological characters and 21 ingroup taxa (Table 2). The outgroup includes a single undetermined species of the following genera: Apedilum Townes, 1945, Beardius Reiss et Sublette, 1985, Genus A (a new Neotropical Chironomini genus that is currently being described by Pinho, Mendes et Andersen), Nilothauma Kieffer, 1921, Paralauterborniella Lenz, 1941, Paratendipes Kieffer, 1911, Pseudochironomus Malloch, 1915 and Zavreliella Kieffer, 1920 (see Fusari et al. 2013).
Heuristic tree searches were performed using 1000 replications with the tree bisection and reconnection (TBR) branch swapping algorithm (Swofford & Olsen 1990), retaining 100 trees, random seed=0. All characters were treated as unordered under the implied weighting (Goloboff 1993). We used a TNT script (propk.run) written by Salvador Arias to calculate the appropriate value for constant k (for details see Goloboff et al. 2008). The script returned a value of k=4.062500 for our data set, which was selected for data analysis.
Results
Oukuriella froehlichi sp. n.
Type material: Holotype male, BRAZIL, São Paulo, Salesópolis, Estação Biológica de Boracéia, 23°31’55”S 45°50’45”W, 10.x.2007, adult male, light trap, leg. Froehlich, C. G. (MZUSP). Paratypes: 2 males, with the same data as holotype (MZUSP).
Etymology. The species is in honour of Dr. Claudio Gilberto Froehlich (FFCLRP-USP), who dedicated his life to Science and also collected all type-series.
Diagnosis. Oukuriella froehlichi sp. n. differs from other congeneric species by: male, wings with dark macula, thorax lacking scutal tubercle, legs light brown with dark brown spot, tergites II–VII with tufts of setae, lateral projections on the posterior margin of tergite IX absent, concavity width equivalent to its depth, and posterior margin of tergite IX with tufts of setae, seen in ventral view. O. froehlichi sp. n. closely resembles O.
baiana Fusari, Roque et Hamada, 2013, from which it can be distinguished by the shape of the superior volsella, and tufts of setae in the ventral margin of tergite IX.
Description. Male (n= 2–3)Color: Thorax light brown with dark brown areas (Fig. 1B). Wings with dark markings, veins light brown (Fig.
1C). Legs light brown with dark brown rings (Fig. 1D). Abdomen light brown, except for third tergite, dark brown; anterior transversal band of fourth tergite dark brown (Fig. 1E).
Total length 2.94–3.52 mm. Wing length 2.22–2.59 mm, width 0.69–0.81 mm. Total length/wing length 1.30–1.40. Wing length/profemur length ratio 1.5.
Head. (Fig. 1A) AR 0.97–1.00. Apical flagellomere 625–735 µm long. Temporals 6. Clypeus with 34–56 setae. Lengths of palpomeres 1–5 (in µm): 35–38; 37–40; 188–195; 154–162; 185–189.
Thorax. (Fig. 1 B) Without scutal tubercle. Ac 5–10; Dc 7–14; Scts 9–13.Wing. (Fig. 1 C) VR 1.25–1.30. Brachiolum with 2–3 setae. R with 8–10, R
1 with 16–24, R
4+5 with 25–32 setae,
remaining veins and cell membranes bare.Legs. (Fig. 1 D) Lengths (in µm) and proportions of leg segments as in Table 1. Tibial spur lengths (in µm):
fore 57–60, middle 79–85, hind 92–96. Abdomen (Fig. 1E). Tergites II–VII with large tuft of setae in anterior median region, and pair of small tufts of
setae on latero-median region.Hypopygium. (Figs. 1 F, 2 A–C) Laterosternite IX with 6–7 setae. Posterior margin of tergite IX without
projections. Concavity of posterior margin of tergite IX as wide as deep. Margin of tergite in ventral view with tufts of setae (Fig. 2B). Phallapodeme 53–65 µm long, transverse sternapodeme 36–57 µm. Superior volsella falciform 101–110 µm long, with 11 setae. Digitus long, lacking microtrichae, with same shape of superior volsella, 104–114 µm long. Inferior volsella 150–159 µm long, with 2 basal setae, 2–4 dorsal setae and 13–15 apical setae. Gonocoxite 175–192 µm long. Gonostylus with 211–233 µm long. HR 0.82–0.83. HV 1.40–1.51.
Female and immatures. Unknown.
Zootaxa 3764 (4) © 2014 Magnolia Press · 419A NEW SPECIES OF SPONGE-DWELLING OUKURIELLA

FIGURE 1. Oukuriella froehlichi sp. n., male. A–Head, B–Thorax, C–Wing, D–Legs, E–Abdomen, F–Hypopygium.
FUSARI ET AL.420 · Zootaxa 3764 (4) © 2014 Magnolia Press
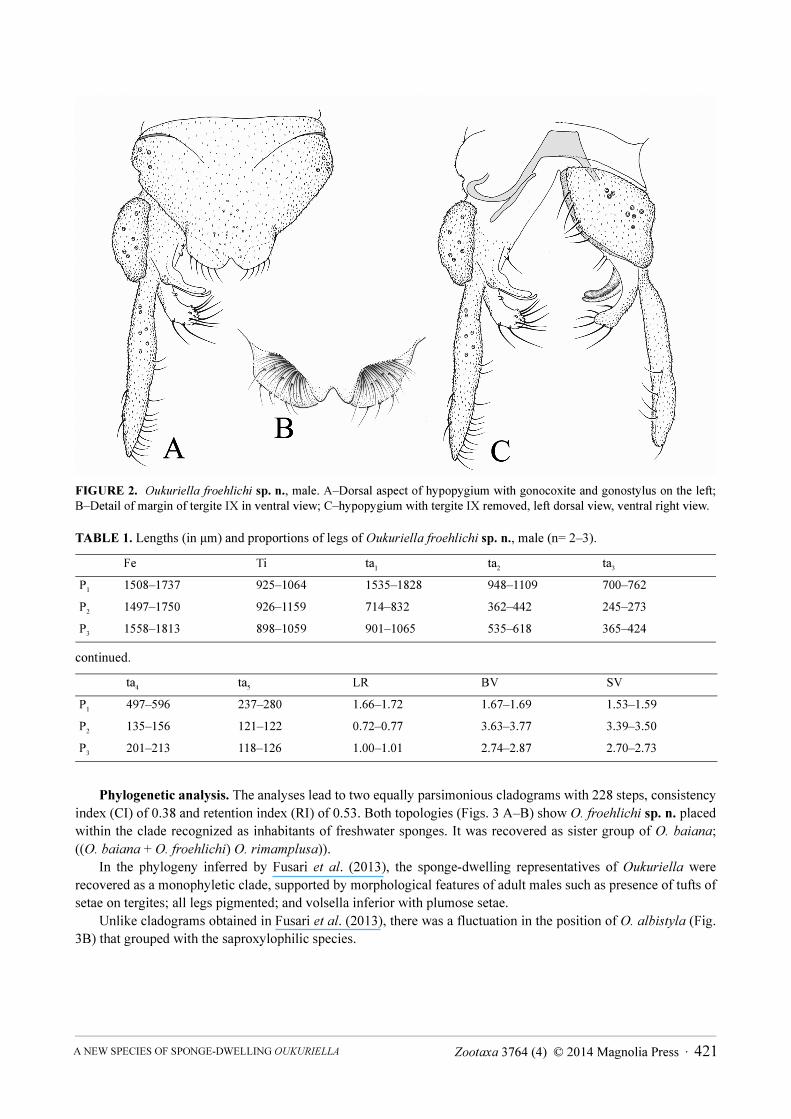
FIGURE 2. Oukuriella froehlichi sp. n., male. A–Dorsal aspect of hypopygium with gonocoxite and gonostylus on the left;
B–Detail of margin of tergite IX in ventral view; C–hypopygium with tergite IX removed, left dorsal view, ventral right view.
TABLE 1. Lengths (in µm) and proportions of legs of Oukuriella froehlichi sp. n., male (n= 2–3).
continued.
Phylogenetic analysis. The analyses lead to two equally parsimonious cladograms with 228 steps, consistency index (CI) of 0.38 and retention index (RI) of 0.53. Both topologies (Figs. 3 A–B) show O. froehlichi sp. n. placed within the clade recognized as inhabitants of freshwater sponges. It was recovered as sister group of O. baiana; ((O. baiana + O. froehlichi) O. rimamplusa)).
In the phylogeny inferred by Fusari et al. (2013), the sponge-dwelling representatives of Oukuriella were recovered as a monophyletic clade, supported by morphological features of adult males such as presence of tufts of setae on tergites; all legs pigmented; and volsella inferior with plumose setae.
Unlike cladograms obtained in Fusari et al. (2013), there was a fluctuation in the position of O. albistyla (Fig. 3B) that grouped with the saproxylophilic species.
Fe Ti ta1
ta2
ta3
P1
1508–1737 925–1064 1535–1828 948–1109 700–762
P2
1497–1750 926–1159 714–832 362–442 245–273
P3
1558–1813 898–1059 901–1065 535–618 365–424
ta4
ta5
LR BV SV
P1
497–596 237–280 1.66–1.72 1.67–1.69 1.53–1.59
P2
135–156 121–122 0.72–0.77 3.63–3.77 3.39–3.50
P3
201–213 118–126 1.00–1.01 2.74–2.87 2.70–2.73
Zootaxa 3764 (4) © 2014 Magnolia Press · 421A NEW SPECIES OF SPONGE-DWELLING OUKURIELLA
FUSARI ET AL.422 · Zootaxa 3764 (4) © 2014 Magnolia Press
Zootaxa 3764 (4) © 2014 Magnolia Press · 423A NEW SPECIES OF SPONGE-DWELLING OUKURIELLA
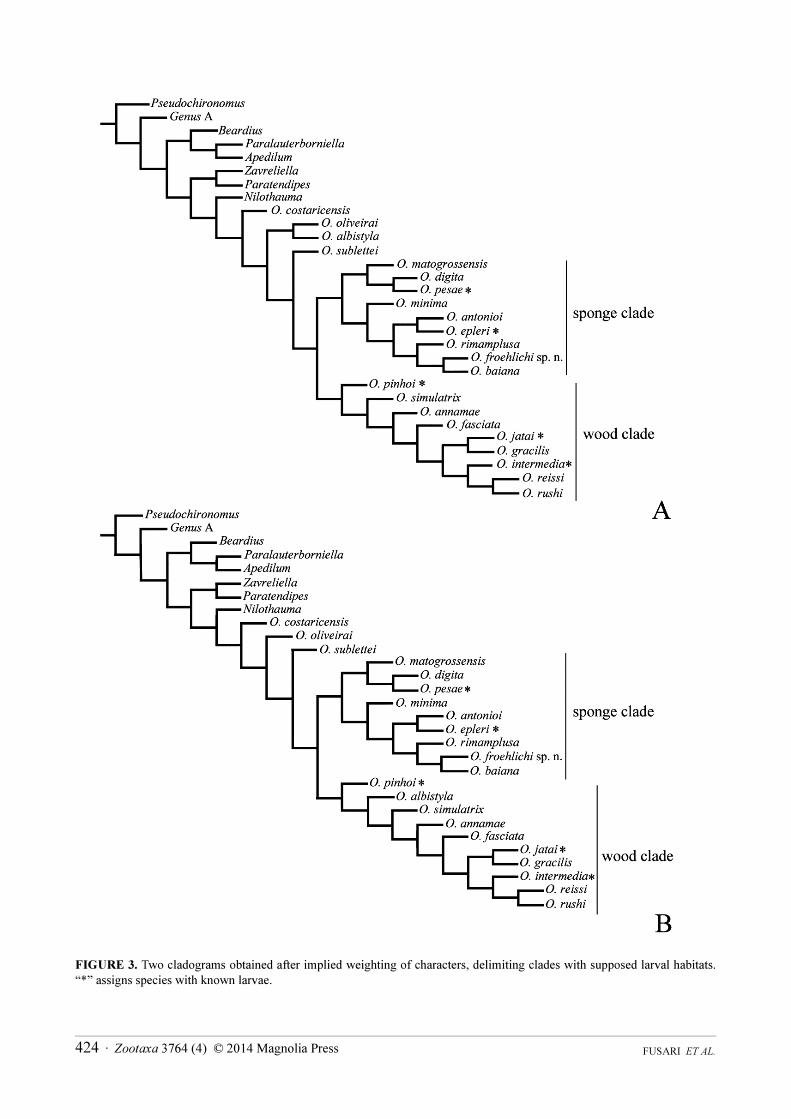
FIGURE 3. Two cladograms obtained after implied weighting of characters, delimiting clades with supposed larval habitats.
“*” assigns species with known larvae.
FUSARI ET AL.424 · Zootaxa 3764 (4) © 2014 Magnolia Press

Discussion
Relationships between chironomids and their specific microhabitats (e.g. submerged wood and animals) are thought to represent major driving forces in the evolution of the family (Cranston & McKie 2006). Furthermore, interspecific interactions such as commensalism, parasitism and mutualism play an important role in the co-evolutionary history of chironomids with several aquatic organisms (fish, insects, and mollusks) (Tokeshi 1993; Roque et. al. 2004).
In Fusari et al. (2013), the topologies yielded strong support to the sponge-dwelling group based on adult male morphology. Our results also show two well supported clades that correspond to the distribution of the species habitats as immatures. So in Oukuriella, the phylogenetic placement of a given species is likely to be predictive of its immature life history, therefore it is suggested that O. froehlichi sp. n. is a sponge-dwelling species.
It is emphasized that studies on the biology of immature stages is encouraged together with phylogenetic analysis. That as demonstrated by these results, the topologies can provide information about the habitat of the larvae, thus directing field studies.
A fluctuation in the position of O. albistyla Epler, 1986 suggests that the species belongs to the group of the saproxylophilic species. It is suggested that saproxylophilic condition is plesiomorphic as it is observed also in related basal genera (Nilothauma and Beardius).
The larval habitats of O. albistyla, O. costaricensis Epler, 1996, O. oliveirai Messias et Fittkau, 1997 and O.
sublettei Messias et Oliveira, 1998 are still unknown, so both taxonomy and ecology of these groups need to be addressed in future studies.
Acknowledgments
Dr. Olivia Evangelista and Dr. Fabio de Oliveira Roque provided valuable comments on an earlier draft of this manuscript draft. Dr. Claudio G. Froehlich for collecting the specimens. Salvador Arias for providing the propk.run script for TNT. L. M. Fusari is currently supported by CNPq through the SISBIOTA Program (Grant number 150078/2012-9; 150434/2013-8). C. F. Bellodi thanks CAPES/DS. C. J. E. Lamas is CNPq research fellow.
References
Cranston, P.S. & McKie, B. (2006) Aquatic Wood—an insect perspective. In: Simon, J.G. & James, L.H. (Eds.), Insect
Biodiversity and Dead Wood: Proceedings of a Symposium for the 22nd International Congress of Entomology. General
Technical Report SRS-93. US Department of Agriculture Service, Southern Research Station Asheville, NC, pp. 9–14.
Epler, J.H. (1986) Oukuriella, a new genus of Chironomidae (Diptera) from South America. Entomologica Scandinavica, 17,
157–163.
http://dx.doi.org/10.1163/187631286x00341
Epler, J.H. (1996) New species of the Oukuriella (Diptera: Chironomidae) from Costa Rica. Hydrobiologia, 318, 3–11.
http://dx.doi.org/10.1007/bf00014126
Fusari, L.M., Roque, F.O. & Hamada, N. (2008) Sponge-dwelling Chironomids in the upper Paraná River (Brazil): Little
known but potentially threatened species. Neotropical Entomology, 37 (5), 522–527.
http://dx.doi.org/10.1590/s1519-566x2008000500004
Fusari, L.M., Roque, F.O. & Hamada, N. (2009) Oukuriella pesae, a new species of sponge-dwelling chironomid (Insecta:
Diptera) from Amazonia, Brazil. Zootaxa, 2146, 61–68.
Fusari, L.M., Roque, F.O. & Hamada, N. (2013) Systematics of Oukuriella Epler, 1986, including a revision of the species
associated with freshwater sponges. Insect Systematics & Evolution, 44, 1–41.
Goloboff, P.A. (1993) Estimating character weights during tree search. Cladistics, 9, 83–91.
http://dx.doi.org/10.1006/clad.1993.1003
Goloboff, P.A., Carpenter, J.M., Arias, J.S., Esquivel, D.R.M., (2008) Weighting against homoplasy improves phylogenetic
analysis of morphological data sets. Cladistics, 24, 758–773.
http://dx.doi.org/10.1111/j.1096-0031.2008.00209.x
Goloboff, P., Farris, S. & Nixon, K. (2003) TNT: Tree analysis using New Technology. Version 1.0, Beta test v. 0.2. Published
by the authors. Tucumán, Argentina. Program and documentation. Available from: http://www.zmuc.dk/public/phylogeny/
TNT/ (accessed 7 January 2014)
Messias, M.C. (1998) Contribuição ao estudo do gênero Oukuriella Epler, 1986. (Insecta, Diptera, Chironomidae). Doctoral
Zootaxa 3764 (4) © 2014 Magnolia Press · 425A NEW SPECIES OF SPONGE-DWELLING OUKURIELLA

Thesis, Instituto Oswaldo Cruz, Rio de Janeiro, Brazil, 157 pp.
Messias, M.C. (2000) Oukuriella reissi, a new species of the genus Oukuriella Epler, 1986 (Insecta, Diptera, Chironomidae).
In: Baehr, M. & Spies, M. (Eds.), Contributions to chironomid research in memory of Dr. Friedrich Reiss. Spixiana, 23 (2),
pp. 159–161.
Messias, M.C, Fittkau, E.J. & Oliveira, S.J. (2000) A new species of Oukuriella Epler, 1986 (Insecta, Diptera, Chironomidae)
with first descriptions of the immature stages for the genus. In: Hoffrichter, O. (Ed.), Late 20th Research on Chironomidae:
an Anthology from the Proceedings of the 13th International Symposium on Chironomidae. Freibeurg, Aachen Shaker, pp.
183–188.
Messias, M.C. & Fittkau, E.J. (1997) Two new species of the Neotropical genus Oukuriella Epler, 1986. (Insecta, Diptera,
Chironomidae). Spixiana, 20 (3), 255–260.
Messias, M.C. & Oliveira, S.J. (1998) On a new species of the genus Oukuriella Epler, 1986. (Insecta, Diptera, Chironomidae).
Journal of Kansas Entomological Society, 71 (3), 260–262.
Page, R.D.M. (2001) Nexus Data Editor (for Windows) Version 0.5.0. University of Glasgow, Glasgow. Available from: http://
taxonomy.zoology.gla.ac.uk/rod/NDE/nde.html (accessed 7 January 2014)
Roque, F.O., Trivinho-Strixino, S., Jancso, M. & Fragoso, E.N. (2004) Records of Chironomidae larvae living on other aquatic
animals in Brazil. Biota Neotropica, 4, 1–9.
http://dx.doi.org/10.1590/s1676-06032004000200018
Schlee, D. (1966) Präparation und Ermittlung von Meßwerten an Chironomidae (Diptera). Gewässer und Abwässer, 41/42,
169–193.
Sæther, O.A. (1968) Chironomids of the Finse Area, Norway, with special reference to their distribution in a glacier brook.
Archiv für Hydrobiologie, 64, 426–483.
Sæther, O.A. (1980) Glossary of chironomid morphology terminology (Diptera: Chironomidae). Entomologica Scandinavica,
Supplement, 14, 1–51.
Swofford, D.L. & Olsen, G. (1990) Phylogeny reconstruction. In: Hillis, D.M. & Moritz, C. (Eds.), Molecular Systematics.
Sinauer, Sunderland, MA, pp. 411–501.
Tokeshi, M. (1993) On the evolution of commensalism in the Chironomidae. Freshwater Biology, 29, 481–489.
http://dx.doi.org/10.1111/j.1365-2427.1993.tb00782.x
Trivinho-Strixino, S. & Messias, M.C. (2005) A new species of Oukuriella Epler, 1986 (Insecta, Diptera, Chironomidae,
Chironominae) from São Paulo State, Brazil. Entomologia y Vectores, 12, 283–291.
http://dx.doi.org/10.1590/s0328-03812005000200012
FUSARI ET AL.426 · Zootaxa 3764 (4) © 2014 Magnolia Press
Related Documents











