REPRODUCTIVE ENDOCRINOLOGY AND INFERTILITY Stromal cell–specific apoptotic and antiestrogenic mechanisms may explain uterine defects in humans after clomiphene citrate therapy Magdalena Nutu, PhD; Yi Feng, MD, PhD; Emil Egecioglu, PhD; Birgitta Weijdegård, MSc; Elisabet Stener-Victorin, RPT, PhD; Ruijin Shao, MD, PhD OBJECTIVE: The purpose of this study was to investigate clomiphene citrate (CC)–induced modulation of uterine cell function in vivo. STUDY DESIGN: Prepubertal female Sprague-Dawley rats were treated intraperitoneally with CC for 6 or 24 hours or with a combination of CC and/or 17--estradiol (E2) for 4 days. RESULTS: Chronic CC treatment induced apoptosis in a fraction of uter- ine stromal cells by activating the caspase-3-mediated apoptotic path- way. The damage was prevented by successive E2 treatment; however, pretreatment or concomitant treatment with E2 did not protect against CC-induced uterine apoptosis. CC decreased the protein expression of estrogen receptor and increased its phosphorylation but did not affect estrogen receptor expression or phosphorylation. Furthermore, changes in Hoxa11, p27, and progesterone receptor protein levels and localization were associated with CC treatment. CONCLUSION: We provide novel mechanistic insights into cellular and molecular events by which CC regulates uterine stromal cell function and hence the implantation process and pregnancy outcome. Key words: apoptosis, clomiphene citrate, implantation, uterus Cite this article as: Nutu M, Feng Y, Egecioglu E, et al. Stromal cell–specific apoptotic and antiestrogenic mechanisms may explain uterine defects in humans after clomiphene citrate therapy. Am J Obstet Gynecol 2010;203:65.e1-10. C lomiphene citrate (CC), a non- steroid tissue-selective estrogen re- ceptor (ER) modulator, 1 is used com- monly to treat various infertilities in women, most notably polycystic ovary syndrome (PCOS) and unexplained in- fertility. 2,3 Several clinical and epidemi- ologic studies have indicated that suc- cessful ovulation occurs in 70 –99% of women who are treated with CC, 2,4 whereas ultimate pregnancy rates are only 27– 40%. 2,5 The total CC-treated pregnancy rates are approximately 10 times lower than “natural” rates. 6 Further- more, spontaneous abortion and miscar- riage occur frequently in women who re- ceive CC therapy. 2,4,7 Although it has been proposed that the antiestrogenic effects of CC may lead to early pregnancy failure, 4 the precise molecular events underlying CC-induced uterine defects remain in- completely understood. The mammalian uterus is a dynamic re- productive organ that undergoes cyclic changes in response to ovarian steroid hor- mones. 8 The biologic effects of 17--estra- diol (E2) and CC are mediated primarily by 2 nuclear receptors, ER- and/or ER-, which are encoded by 2 different genes and function as transcription factors. 9 Al- though the distribution and relative levels of ER- and ER- expression are tissue specific and diverse, 10 both ER- and ER- are expressed in the rodent uterus and human endometrium. 11 It has been reported that inappropriate activation or inhibition of ER subtypes may cause or contribute to a variety of uterine diseases, such as endometriosis and endometrial cancer. 11 The protective effects of E2 in uterine homeostasis are evident both from in vivo and in vitro studies: the presence of E2 inhibits uterine apopto- sis in vivo; 12 however, human endome- trial cells undergo apoptosis when E2 is withdrawn. 13 Previous studies from our laboratory and others have shown that CC enhances apoptotic processes in the ova- ries, fallopian tubes, villi, and decidual tis- sues. 14-16 Less is known, however, about From the Department of Physiology/Endocrinology, Institute of Neuroscience and Physiology, The Sahlgrenska Academy at University of Gothenburg, Gothenburg, Sweden (all authors); and the Department of Neurobiology and Integrative Medicine, Shanghai Medical College of Fudan University, Shanghai (Dr Feng), and the Department of Obstetrics and Gynecology, First Affiliated Hospital, Heilongjiang University of Chinese Medicine, Harbin (Dr Stener-Victorin), China. Received Nov. 16, 2009; revised Jan. 10, 2010; accepted March 17, 2010. Reprints: Ruijin Shao, MD, PhD, Department of Physiology/Endocrinology, Institute of Neuroscience and Physiology, The Sahlgrenska Academy at Gothenburg University, Medicinareg 11, PO Box 434, 413 90 Gothenburg, Sweden. [email protected]. This study was supported by the Swedish Medical Research Council (Grant no. 5859), the Svenska Sällskapet för Medicinsk Forskning, Sahlgrenska Academy Research Council, Göteborgs Läkaresällskap, Fred G. and Emma E. Kanolds Stiftelse, Anna Cederbergs Stiftelse, Fredrik and Ingrid Thurings Stiftelse, and the Hjalmar Svenssons Research Foundation. 0002-9378/free • © 2010 Mosby, Inc. All rights reserved. • doi: 10.1016/j.ajog.2010.03.039 For Editors’ Commentary, see Table of Contents Research www. AJOG.org JULY 2010 American Journal of Obstetrics & Gynecology 65.e1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

R
SmiME
Oc
Sia
Riwp
Cc
Ccmwsfocww
FPaCG(
R
RN1
TSGF
0
Research www.AJOG.org
EPRODUCTIVE ENDOCRINOLOGY AND INFERTILITY
tromal cell–specific apoptotic and antiestrogenicechanisms may explain uterine defects
n humans after clomiphene citrate therapyagdalena Nutu, PhD; Yi Feng, MD, PhD; Emil Egecioglu, PhD; Birgitta Weijdegård, MSc;
lisabet Stener-Victorin, RPT, PhD; Ruijin Shao, MD, PhD
Ceecl
Cma
BJECTIVE: The purpose of this study was to investigate clomipheneitrate (CC)–induced modulation of uterine cell function in vivo.
TUDY DESIGN: Prepubertal female Sprague-Dawley rats were treatedntraperitoneally with CC for 6 or 24 hours or with a combination of CCnd/or 17-�-estradiol (E2) for 4 days.
ESULTS: Chronic CC treatment induced apoptosis in a fraction of uter-ne stromal cells by activating the caspase-3-mediated apoptotic path-ay. The damage was prevented by successive E2 treatment; however,
lomiphene citrate therapy. Am J Obstet Gynecol 2010;203:65.e1-10.
optmrcpCt
C-induced uterine apoptosis. CC decreased the protein expression ofstrogen receptor � and increased its phosphorylation but did not affectstrogen receptor � expression or phosphorylation. Furthermore,hanges in Hoxa11, p27, and progesterone receptor protein levels andocalization were associated with CC treatment.
ONCLUSION: We provide novel mechanistic insights into cellular andolecular events by which CC regulates uterine stromal cell function
nd hence the implantation process and pregnancy outcome.
retreatment or concomitant treatment with E2 did not protect against Key words: apoptosis, clomiphene citrate, implantation, uterus
ite this article as: Nutu M, Feng Y, Egecioglu E, et al. Stromal cell–specific apoptotic and antiestrogenic mechanisms may explain uterine defects in humans after
Cc
pcmdbwftosEaricscufpstwler
lomiphene citrate (CC), a non-steroid tissue-selective estrogen re-
eptor (ER) modulator,1 is used com-only to treat various infertilities inomen, most notably polycystic ovary
yndrome (PCOS) and unexplained in-ertility.2,3 Several clinical and epidemi-logic studies have indicated that suc-essful ovulation occurs in 70 –99% ofomen who are treated with CC,2,4
hereas ultimate pregnancy rates are
rom the Department of Physiology/Endocrinhysiology, The Sahlgrenska Academy at Uniuthors); and the Department of Neurobiologollege of Fudan University, Shanghai (Dr Feynecology, First Affiliated Hospital, Heilong
Dr Stener-Victorin), China.
eceived Nov. 16, 2009; revised Jan. 10, 2010;
eprints: Ruijin Shao, MD, PhD, Department ofeuroscience and Physiology, The Sahlgrenska1, PO Box 434, 413 90 Gothenburg, Sweden.
his study was supported by the Swedish Medicvenska Sällskapet för Medicinsk Forskning, Saöteborgs Läkaresällskap, Fred G. and Emma Eredrik and Ingrid Thurings Stiftelse, and the Hja
002-9378/free • © 2010 Mosby, Inc. All rights
For Editors’ Commentary, see Table
nly 27– 40%.2,5 The total CC-treatedregnancy rates are approximately 10
imes lower than “natural” rates.6 Further-ore, spontaneous abortion and miscar-
iage occur frequently in women who re-eive CC therapy.2,4,7 Although it has beenroposed that the antiestrogenic effects ofC may lead to early pregnancy failure,4
he precise molecular events underlying
gy, Institute of Neuroscience andity of Gothenburg, Gothenburg, Sweden (allnd Integrative Medicine, Shanghai Medical, and the Department of Obstetrics andg University of Chinese Medicine, Harbin
epted March 17, 2010.
siology/Endocrinology, Institute ofademy at Gothenburg University, [email protected].
esearch Council (Grant no. 5859), theenska Academy Research Council,anolds Stiftelse, Anna Cederbergs Stiftelse,r Svenssons Research Foundation.
erved. • doi: 10.1016/j.ajog.2010.03.039
ontents
sJULY 2010 Ameri
C-induced uterine defects remain in-ompletely understood.
The mammalian uterus is a dynamic re-roductive organ that undergoes cyclichanges in response to ovarian steroid hor-ones.8 The biologic effects of 17-�-estra-
iol (E2) and CC are mediated primarilyy 2 nuclear receptors, ER-� and/or ER-�,hich are encoded by 2 different genes and
unction as transcription factors.9 Al-hough the distribution and relative levelsf ER-� and ER-� expression are tissuepecific and diverse,10 both ER-� andR-� are expressed in the rodent uterusnd human endometrium.11 It has beeneported that inappropriate activation ornhibition of ER subtypes may cause orontribute to a variety of uterine diseases,uch as endometriosis and endometrialancer.11 The protective effects of E2 interine homeostasis are evident both
rom in vivo and in vitro studies: theresence of E2 inhibits uterine apopto-is in vivo;12 however, human endome-rial cells undergo apoptosis when E2 isithdrawn.13 Previous studies from our
aboratory and others have shown that CCnhances apoptotic processes in the ova-ies, fallopian tubes, villi, and decidual tis-
oloversy ang)jian
acc
PhyAcruiji
al Rhlgr. Klma
res
of C
ues.14-16 Less is known, however, aboutcan Journal of Obstetrics & Gynecology 65.e1

te
pcptpbosespeeptcfis
drmttmcbttwtttpstaupi
MAAcmbdMmw(immbAal
ET(wnLbooutqtswosi
ERpvlit8tf
ERifhm
C
N
Research Reproductive Endocrinology and Infertility www.AJOG.org
6
he effects of CC on the apoptotic machin-ry in the uterus.
The maternal endometrium showsrominent steroid-dependent cyclichanges in structure and function inreparation for the process of implanta-ion.8 Successful implantation requiresrecise coordination between the em-ryo and uterus under the influence ofvarian steroids.17 After fertilization,pecific uterine cell types undergo differ-ntiation and proliferation to provide auitable environment for embryo im-lantation and development.17 Genomicndometrial responses to estrogen aressential for the regulation of the “im-lantation window.”18 The transforma-ion of endometrial stromal cells into de-idual cells has been recognized as aundamental step during the process ofmplantation.18,19 Previously, it has been
FIGURE 1Schematic of the experimental des
C, clomiphene citrate; E2, 17-�-estradiol; Exp, experiment.
utu. Clomiphene citrate treatment and rat uterus. Am J Ob
hown that in vivo treatment with CC C
5.e2 American Journal of Obstetrics & Gynecolog
elays and/or inhibits implantation inodents,18,20 probably because of abnor-alities that are seen in the reproductive
ract (including the uterus) after CCreatment in both rats16,21 and hu-
ans.22 Furthermore, a significant de-rease in the implantation rate has alsoeen observed in rabbits that werereated with CC before and after ovula-ion.23 These observations, combinedith clinical studies, have given rise to
he hypothesis that CC may contributeo implantation-related complicationshrough an unidentified regulatoryrocess in the uterus. The goals of thistudy were (1) to determine whether CCreatment induces uterine cell apoptosisnd (2) to investigate the molecular reg-lation of ERs and other potential im-lantation and cell cycle regulators17,24
n the uteri of rats that were treated with
ynecol 2010.
C. w
y JULY 2010
ATERIALS AND METHODSnimalsll experimental procedures and proto-ols were approved by the ethics com-ittee at Gothenburg University. Prepu-
ertal female Sprague-Dawley rats (20ays old) were obtained from Taconic&B (Copenhagen, Denmark) andaintained in cages that containedood chips under defined conditions
temperature, 21 � 2°C; relative humid-ty, 45–55%; and 12-hour light/dark illu-
ination schedule). Animals were accli-ated to the animal facilities for 5 days
efore the initiation of the experiments.ll animals had free access to tap waternd were fed ad libitum with standardaboratory diet.
xperimental designhree experiments were carried out
Figure 1) in 25-day-old rats. The ratsere randomized to receive intraperito-eal injections of CC (Sigma-Aldrich, St.ouis, MO), E2 (Sigma-Aldrich), oroth. Controls were treated with vehiclenly. Bodyweight was recorded through-ut the experiment. Rats were killednder light anesthesia with sodium pen-
obarbital (0.5 mL/kg bodyweight). Ali-uots of serum were prepared fromrunk blood after heart puncture andtored at – 80°C until analysis. The uteriere dissected grossly with the removalf contaminating tissues (eg, adipose tis-ues), weighed, and immediately frozenn liquid nitrogen or fixed in formalin.
xperiment 1ats received CC (1 or 10 mg/kg intra-eritoneally) or an equivalent volume ofehicle (0.9% NaCl). Uteri were col-ected 6 and 24 hours (acute effect) afternjection or on day 4 (chronic effect) af-er daily intraperitoneal injections (n �/group). For the selected doses andreatment schedule, CC is effective in ratallopian tube and uterus in vivo.16,25
xperiment 2ats received injections of CC (10 mg/kg
ntraperitoneally) or vehicle once dailyor 4 consecutive days. Twenty-fourours after the last injection, E2 (0.3g/kg in 100 �L sesame oil) or vehicle
ign
stet G
as injected for 4 consecutive days.
ERoetmwsr25
ATWflmaTaa[CENmdNTtmaTgtwjJ(
HiUfe�m(Um
UcwBc
SbdNO
www.AJOG.org Reproductive Endocrinology and Infertility Research
JULY 2010 Ameri
xperiment 3ats were injected with E2 or vehiclence daily for 4 consecutive days, as inxperiment 2. Twenty-four hours afterhe last injection, CC (10 mg/kg), CC (10
g/kg) and E2 (0.3 mg/kg), or vehicleas injected intraperitoneally for 4 con-
ecutive days. In experiments 2 and 3, theats were killed, and uteri were collected4 hours after the final injection (n �/group).
ntibodieshe primary antibodies that were used forestern blot (WB) analysis and immuno-
uorescence were obtained from com-ercial sources (anticleaved caspase 3 and
nticleaved caspase 9 [Cell Signalingechnology, Beverly, MA]; anti-ERá,ntiphospho-ERâ [Ser87], anti-Hoxa10,nti-Hoxa11, anti-p27, and anti-p53Santa Cruz Biotechnology, Santa Cruz,A]; antiphospho-ERá [Ser118] and anti-Râ [Upstate Biotechnology, Lake Placid,Y]; anti-pan-cytokeratin, anti-á-smoothuscle actin and anti-â-actin [Sigma-Al-
rich]; and antiprogesterone receptor [PR;ovocastra Laboratories, Newcastle Uponyne, UK] antibodies). The secondary an-
ibodies for WB were goat-antimouse im-unoglobulin G (Sigma-Aldrich), goat-
ntirabbit immunoglobulin G (AC31RL;ropix, Bedford, MA), and donkey anti-oat immunoglobulin G (Santa Cruz Bio-echnology), which were all conjugatedith alkaline phosphatase. The Cy3-con-
ugated antimouse antibody was fromackson ImmunoResearch LaboratoriesWest Grove, PA).
istologic evaluation andmmunofluorescence staining
teri were fixed in neutral buffered 10%ormalin, decalcified, dehydrated, andmbedded in paraffin. Series sections (5m) were prepared and stained with he-atoxylin/eosin to visualize cell nuclei
Histocenter, Västra Frolunda, Sweden).terine sections were subjected to im-unofluorescence studies to test the lo-
EM. The single asterisk denotes P � .05; the dou-le asterisks denote P � .01; the triple asterisksenote P � .001 vs vehicle-treated controls.utu. Clomiphene citrate treatment and rat uterus. Am J
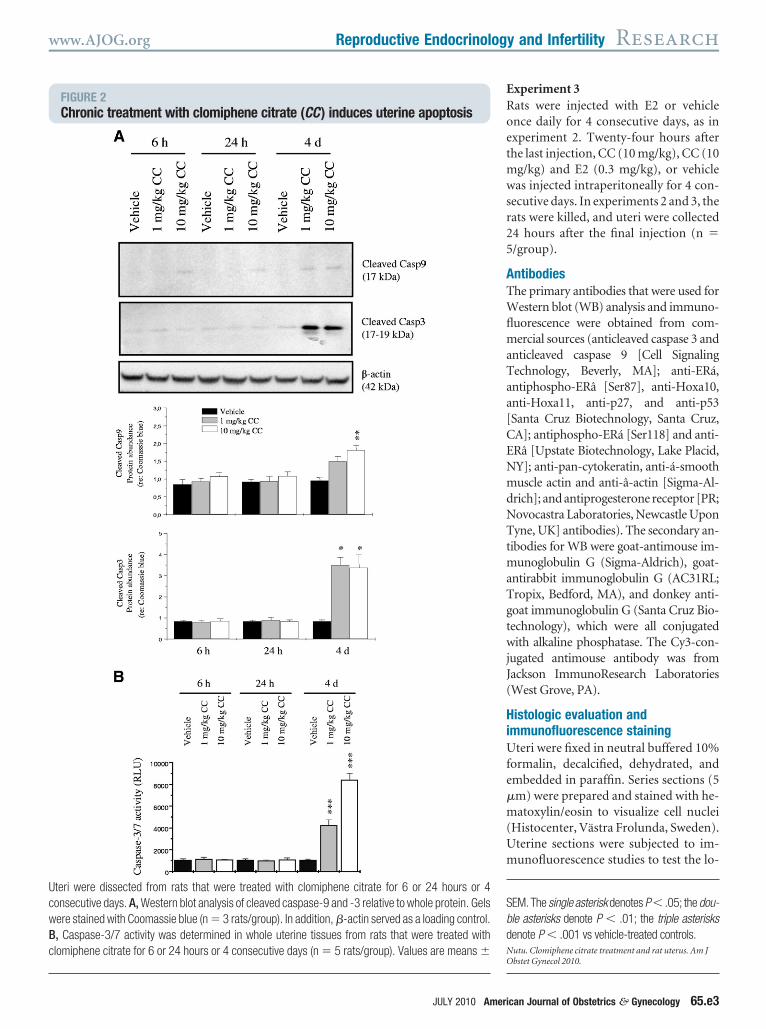
FIGURE 2Chronic treatment with clomiphene citrate (CC) induces uterine apoptosis
teri were dissected from rats that were treated with clomiphene citrate for 6 or 24 hours or 4onsecutive days. A, Western blot analysis of cleaved caspase-9 and -3 relative to whole protein. Gelsere stained with Coomassie blue (n � 3 rats/group). In addition, �-actin served as a loading control., Caspase-3/7 activity was determined in whole uterine tissues from rats that were treated withlomiphene citrate for 6 or 24 hours or 4 consecutive days (n � 5 rats/group). Values are means �
bstet Gynecol 2010.
can Journal of Obstetrics & Gynecology 65.e3
ci
IfTd
tsu(Gb
0icolw(aii3ptmotu
PWwp
CFiwmNc1cuSd
SD(TattbwwliSp
RCiiR
AmcdwdeL
N
Research Reproductive Endocrinology and Infertility www.AJOG.org
6
alization of progesterone receptor Asoform, as described previously.26
n situ detection of DNAragmentationissue sections were dewaxed, protease
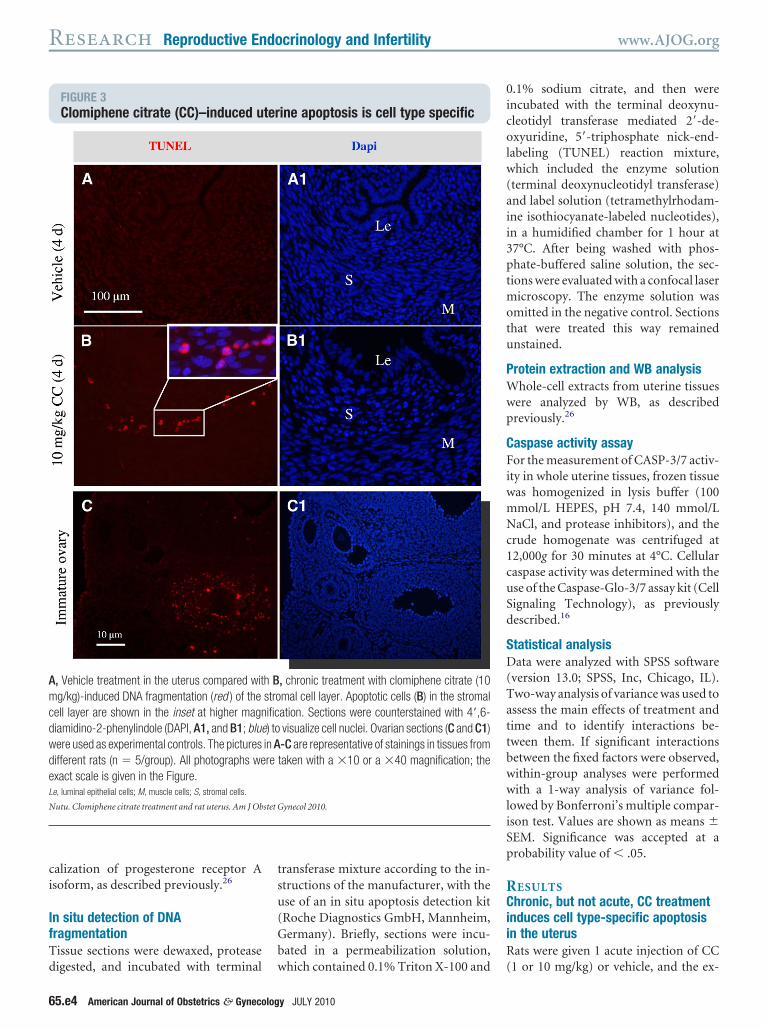
FIGURE 3Clomiphene citrate (CC)–induced u
, Vehicle treatment in the uterus compared witg/kg)-induced DNA fragmentation (red ) of the
ell layer are shown in the inset at higher magniamidino-2-phenylindole (DAPI, A1, and B1; blueere used as experimental controls. The picturesifferent rats (n � 5/group). All photographs wexact scale is given in the Figure.e, luminal epithelial cells; M, muscle cells; S, stromal cells.
utu. Clomiphene citrate treatment and rat uterus. Am J Ob
igested, and incubated with terminal w
5.e4 American Journal of Obstetrics & Gynecolog
ransferase mixture according to the in-tructions of the manufacturer, with these of an in situ apoptosis detection kitRoche Diagnostics GmbH, Mannheim,ermany). Briefly, sections were incu-ated in a permeabilization solution,
ine apoptosis is cell type specific
, chronic treatment with clomiphene citrate (10mal cell layer. Apoptotic cells (B) in the stromalation. Sections were counterstained with 4=,6-visualize cell nuclei. Ovarian sections (C and C1)-C are representative of stainings in tissues fromtaken with a �10 or a �40 magnification; the
ynecol 2010.
hich contained 0.1% Triton X-100 and (
y JULY 2010
.1% sodium citrate, and then werencubated with the terminal deoxynu-leotidyl transferase mediated 2=-de-xyuridine, 5=-triphosphate nick-end-
abeling (TUNEL) reaction mixture,hich included the enzyme solution
terminal deoxynucleotidyl transferase)nd label solution (tetramethylrhodam-ne isothiocyanate-labeled nucleotides),n a humidified chamber for 1 hour at7°C. After being washed with phos-hate-buffered saline solution, the sec-ions were evaluated with a confocal laser
icroscopy. The enzyme solution wasmitted in the negative control. Sectionshat were treated this way remainednstained.
rotein extraction and WB analysishole-cell extracts from uterine tissues
ere analyzed by WB, as describedreviously.26
aspase activity assayor the measurement of CASP-3/7 activ-
ty in whole uterine tissues, frozen tissueas homogenized in lysis buffer (100mol/L HEPES, pH 7.4, 140 mmol/LaCl, and protease inhibitors), and the
rude homogenate was centrifuged at2,000g for 30 minutes at 4°C. Cellularaspase activity was determined with these of the Caspase-Glo-3/7 assay kit (Cellignaling Technology), as previouslyescribed.16
tatistical analysisata were analyzed with SPSS software
version 13.0; SPSS, Inc, Chicago, IL).wo-way analysis of variance was used tossess the main effects of treatment andime and to identify interactions be-ween them. If significant interactionsetween the fixed factors were observed,ithin-group analyses were performedith a 1-way analysis of variance fol-
owed by Bonferroni’s multiple compar-son test. Values are shown as means �EM. Significance was accepted at arobability value of � .05.
ESULTShronic, but not acute, CC treatment
nduces cell type-specific apoptosisn the uterusats were given 1 acute injection of CC
ter
h Bstroific) toin Are
stet G
1 or 10 mg/kg) or vehicle, and the ex-
p36mtsivCb6ceuCs(wisrd(aeccsac
EuTcsvwAutwcC(
CimAwchabe
ortita(ictdtmttB
acs“lTs5Blth
CEW
Rtra(.N
www.AJOG.org Reproductive Endocrinology and Infertility Research
ression of cleaved caspase 9 and caspase(Casp3) was measured in the uterus atand 24 hours. Alternatively, CC was ad-inistered daily for 4 days (chronic
reatment), and the expression of theame apoptotic molecules was measuredn the uterus. WB analysis revealed ele-ated levels of cleaved caspase 9 andasp3 in the uteri from rats that hadeen treated with CC for 4 days but not atand 24 hours (Figure 2, A). Using the
aspase-Glo 3/7 assay, we confirmed thatlevated Casp3 activity was found only interi from rats that had been treated withC for 4 days (Figure 2, B), which is con-
istent with the levels of cleaved Casp3Figure 2, A). Next, DNA fragmentationas detected in situ with TUNEL stain-
ng. TUNEL-positive cells were presentpecifically in the stromal cell layer inats that had been treated with CC for 4ays (Figure 3, B) but not in control ratsFigure 3, A). The luminal (Figure 3, And B) and glandular (data not shown)pithelial cells and the smooth muscleell layers (Figure 3, A and B) in bothontrol and CC-treated rats showed notaining. Ovarian sections (Figure 3, Cnd C1) were used as experimentalontrols.
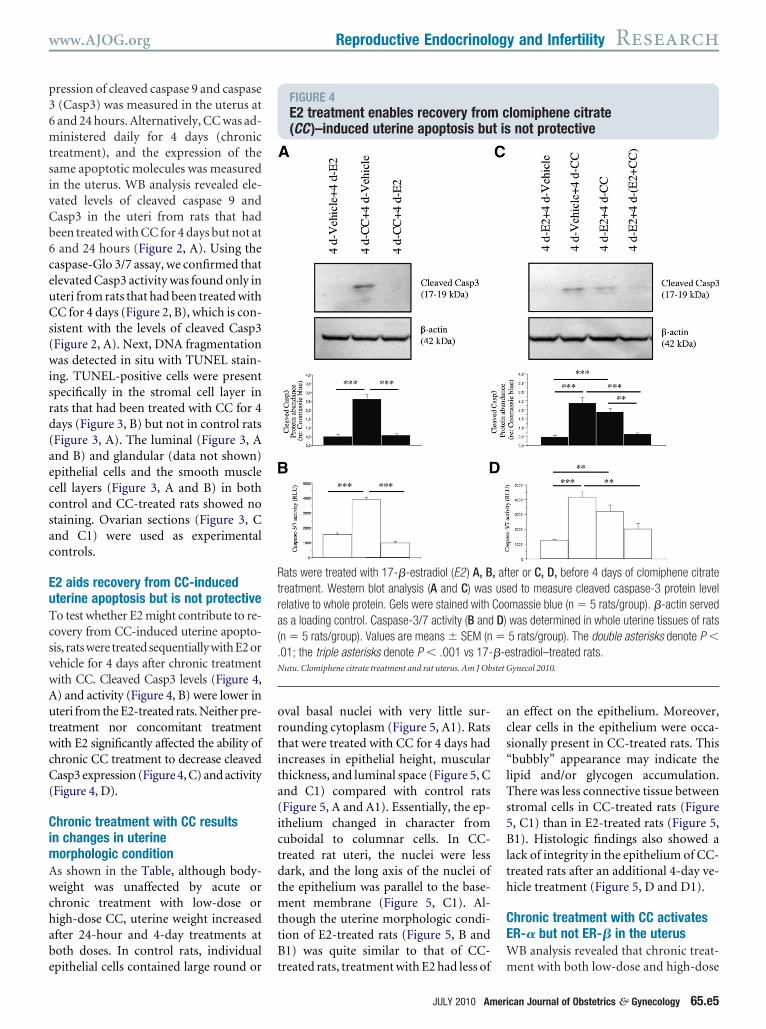
2 aids recovery from CC-inducedterine apoptosis but is not protectiveo test whether E2 might contribute to re-overy from CC-induced uterine apopto-is, ratswere treatedsequentiallywithE2orehicle for 4 days after chronic treatmentith CC. Cleaved Casp3 levels (Figure 4,) and activity (Figure 4, B) were lower interi from the E2-treated rats. Neither pre-
reatment nor concomitant treatmentith E2 significantly affected the ability of
hronic CC treatment to decrease cleavedasp3 expression (Figure 4, C) and activity
Figure 4, D).
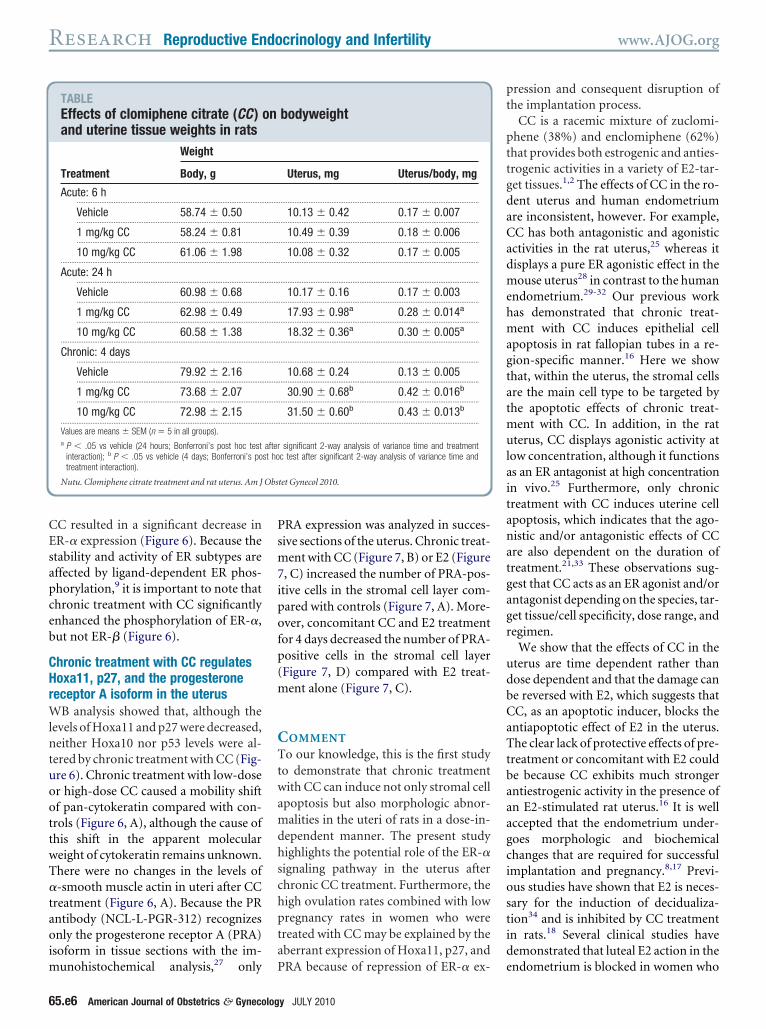
hronic treatment with CC resultsn changes in uterine
orphologic conditions shown in the Table, although body-eight was unaffected by acute or
hronic treatment with low-dose origh-dose CC, uterine weight increasedfter 24-hour and 4-day treatments atoth doses. In control rats, individual
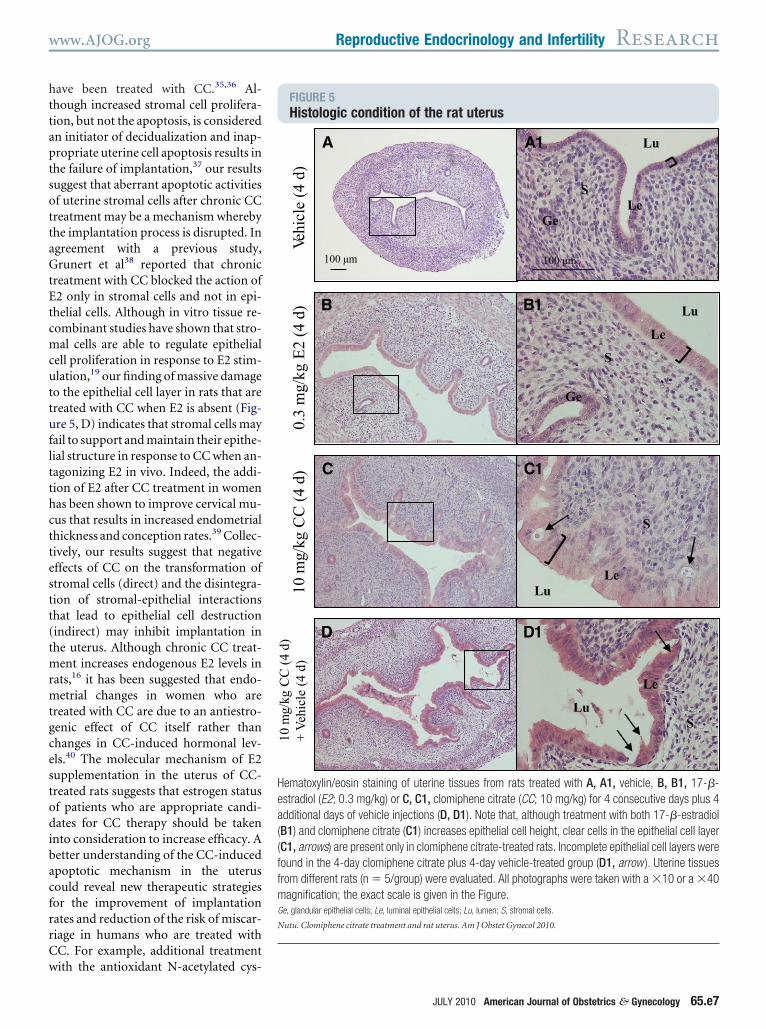
pithelial cells contained large round or tval basal nuclei with very little sur-ounding cytoplasm (Figure 5, A1). Ratshat were treated with CC for 4 days hadncreases in epithelial height, muscularhickness, and luminal space (Figure 5, Cnd C1) compared with control ratsFigure 5, A and A1). Essentially, the ep-thelium changed in character fromuboidal to columnar cells. In CC-reated rat uteri, the nuclei were lessark, and the long axis of the nuclei ofhe epithelium was parallel to the base-
ent membrane (Figure 5, C1). Al-hough the uterine morphologic condi-ion of E2-treated rats (Figure 5, B and1) was quite similar to that of CC-
FIGURE 4E2 treatment enables recovery from(CC )–induced uterine apoptosis bu
ats were treated with 17-�-estradiol (E2 ) A, Breatment. Western blot analysis (A and C) waselative to whole protein. Gels were stained withs a loading control. Caspase-3/7 activity (B andn � 5 rats/group). Values are means � SEM (n01; the triple asterisks denote P � .001 vs 17-utu. Clomiphene citrate treatment and rat uterus. Am J Ob
reated rats, treatment with E2 had less of m
JULY 2010 Ameri
n effect on the epithelium. Moreover,lear cells in the epithelium were occa-ionally present in CC-treated rats. Thisbubbly” appearance may indicate theipid and/or glycogen accumulation.here was less connective tissue between
tromal cells in CC-treated rats (Figure, C1) than in E2-treated rats (Figure 5,1). Histologic findings also showed a
ack of integrity in the epithelium of CC-reated rats after an additional 4-day ve-icle treatment (Figure 5, D and D1).
hronic treatment with CC activatesR-� but not ER-� in the uterusB analysis revealed that chronic treat-
lomiphene citratenot protective
ter or C, D, before 4 days of clomiphene citrated to measure cleaved caspase-3 protein levelmassie blue (n � 5 rats/group). �-actin servedwas determined in whole uterine tissues of rats5 rats/group). The double asterisks denote P �stradiol–treated rats.ynecol 2010.
ct is
, afuse
CooD)��-estet G
ent with both low-dose and high-dose
can Journal of Obstetrics & Gynecology 65.e5
CEsapceb
CHrWlntuoottwT�taoim
Psm7ipofp(m
CTtwamdhschpta
pt
pttgdaCadmehmagtatmulaitanatgagr
udbCaTtbaaagciostid
Obs
Research Reproductive Endocrinology and Infertility www.AJOG.org
6
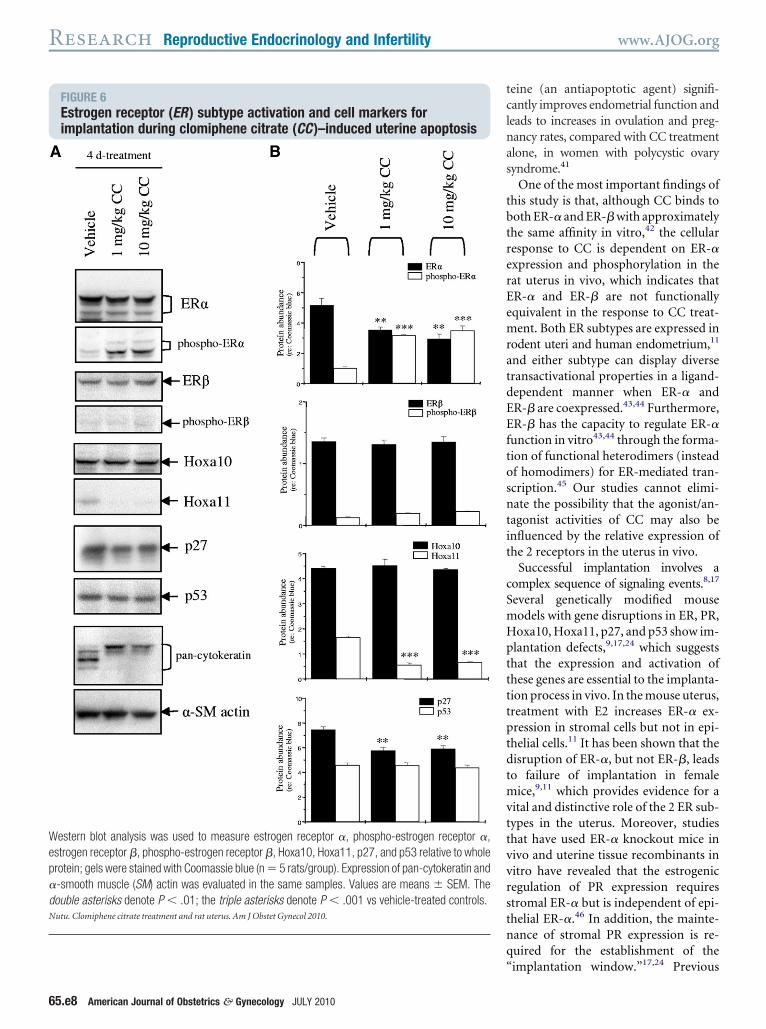
C resulted in a significant decrease inR-� expression (Figure 6). Because thetability and activity of ER subtypes areffected by ligand-dependent ER phos-horylation,9 it is important to note thathronic treatment with CC significantlynhanced the phosphorylation of ER-�,ut not ER-� (Figure 6).
hronic treatment with CC regulatesoxa11, p27, and the progesteroneeceptor A isoform in the uterus
B analysis showed that, although theevels of Hoxa11 and p27 were decreased,either Hoxa10 nor p53 levels were al-
ered by chronic treatment with CC (Fig-re 6). Chronic treatment with low-doser high-dose CC caused a mobility shiftf pan-cytokeratin compared with con-rols (Figure 6, A), although the cause ofhis shift in the apparent moleculareight of cytokeratin remains unknown.here were no changes in the levels of-smooth muscle actin in uteri after CC
reatment (Figure 6, A). Because the PRntibody (NCL-L-PGR-312) recognizesnly the progesterone receptor A (PRA)
soform in tissue sections with the im-
TABLEEffects of clomiphene citrate (CC )and uterine tissue weights in rats
Treatment
Weight
Body, g
Acute: 6 h..........................................................................................................
Vehicle 58.74 � 0.50..........................................................................................................
1 mg/kg CC 58.24 � 0.81..........................................................................................................
10 mg/kg CC 61.06 � 1.98...................................................................................................................
Acute: 24 h..........................................................................................................
Vehicle 60.98 � 0.68..........................................................................................................
1 mg/kg CC 62.98 � 0.49..........................................................................................................
10 mg/kg CC 60.58 � 1.38...................................................................................................................
Chronic: 4 days..........................................................................................................
Vehicle 79.92 � 2.16..........................................................................................................
1 mg/kg CC 73.68 � 2.07..........................................................................................................
10 mg/kg CC 72.98 � 2.15...................................................................................................................
Values are means � SEM (n � 5 in all groups).a P � .05 vs vehicle (24 hours; Bonferroni’s post hoc test
interaction); b P � .05 vs vehicle (4 days; Bonferroni’s potreatment interaction).
Nutu. Clomiphene citrate treatment and rat uterus. Am J
unohistochemical analysis,27 only P
5.e6 American Journal of Obstetrics & Gynecolog
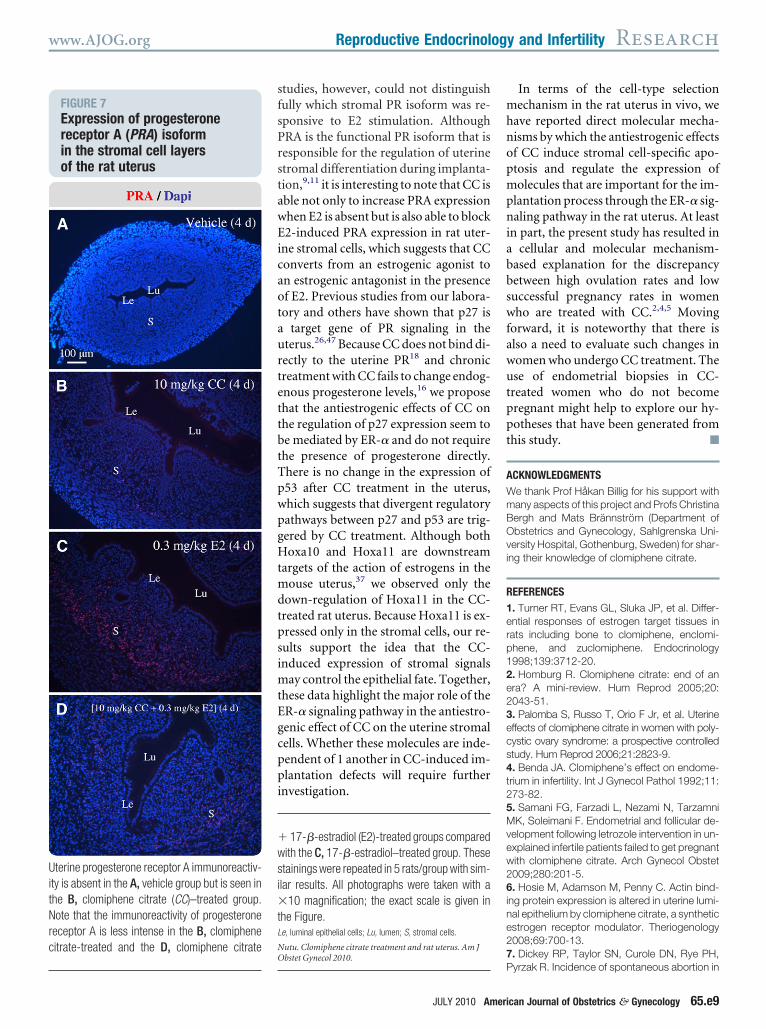
RA expression was analyzed in succes-ive sections of the uterus. Chronic treat-
ent with CC (Figure 7, B) or E2 (Figure, C) increased the number of PRA-pos-tive cells in the stromal cell layer com-ared with controls (Figure 7, A). More-ver, concomitant CC and E2 treatmentor 4 days decreased the number of PRA-ositive cells in the stromal cell layerFigure 7, D) compared with E2 treat-ent alone (Figure 7, C).
OMMENT
o our knowledge, this is the first studyo demonstrate that chronic treatmentith CC can induce not only stromal cell
poptosis but also morphologic abnor-alities in the uteri of rats in a dose-in-
ependent manner. The present studyighlights the potential role of the ER-�ignaling pathway in the uterus afterhronic CC treatment. Furthermore, theigh ovulation rates combined with lowregnancy rates in women who werereated with CC may be explained by theberrant expression of Hoxa11, p27, and
bodyweight
Uterus, mg Uterus/body, mg
..................................................................................................................
10.13 � 0.42 0.17 � 0.007..................................................................................................................
10.49 � 0.39 0.18 � 0.006..................................................................................................................
10.08 � 0.32 0.17 � 0.005..................................................................................................................
..................................................................................................................
10.17 � 0.16 0.17 � 0.003..................................................................................................................
17.93 � 0.98a 0.28 � 0.014a
..................................................................................................................
18.32 � 0.36a 0.30 � 0.005a
..................................................................................................................
..................................................................................................................
10.68 � 0.24 0.13 � 0.005..................................................................................................................
30.90 � 0.68b 0.42 � 0.016b
..................................................................................................................
31.50 � 0.60b 0.43 � 0.013b
..................................................................................................................
significant 2-way analysis of variance time and treatmentc test after significant 2-way analysis of variance time and
tet Gynecol 2010.
RA because of repression of ER-� ex- e
y JULY 2010
ression and consequent disruption ofhe implantation process.
CC is a racemic mixture of zuclomi-hene (38%) and enclomiphene (62%)hat provides both estrogenic and anties-rogenic activities in a variety of E2-tar-et tissues.1,2 The effects of CC in the ro-ent uterus and human endometriumre inconsistent, however. For example,C has both antagonistic and agonisticctivities in the rat uterus,25 whereas itisplays a pure ER agonistic effect in theouse uterus28 in contrast to the human
ndometrium.29-32 Our previous workas demonstrated that chronic treat-ent with CC induces epithelial cell
poptosis in rat fallopian tubes in a re-ion-specific manner.16 Here we showhat, within the uterus, the stromal cellsre the main cell type to be targeted byhe apoptotic effects of chronic treat-
ent with CC. In addition, in the ratterus, CC displays agonistic activity at
ow concentration, although it functionss an ER antagonist at high concentrationn vivo.25 Furthermore, only chronicreatment with CC induces uterine cellpoptosis, which indicates that the ago-istic and/or antagonistic effects of CCre also dependent on the duration ofreatment.21,33 These observations sug-est that CC acts as an ER agonist and/orntagonist depending on the species, tar-et tissue/cell specificity, dose range, andegimen.
We show that the effects of CC in theterus are time dependent rather thanose dependent and that the damage cane reversed with E2, which suggests thatC, as an apoptotic inducer, blocks thentiapoptotic effect of E2 in the uterus.he clear lack of protective effects of pre-
reatment or concomitant with E2 coulde because CC exhibits much strongerntiestrogenic activity in the presence ofn E2-stimulated rat uterus.16 It is wellccepted that the endometrium under-oes morphologic and biochemicalhanges that are required for successfulmplantation and pregnancy.8,17 Previ-us studies have shown that E2 is neces-ary for the induction of decidualiza-ion34 and is inhibited by CC treatmentn rats.18 Several clinical studies haveemonstrated that luteal E2 action in the
on
.........
.........
.........
.........
.........
.........
.........
.........
.........
.........
.........
.........
afterst ho
ndometrium is blocked in women who
httaptsottaGtEtcmcuttufltthcttestt(tmrmtgcestodibacfrrCw
Hea((ffmG
N
www.AJOG.org Reproductive Endocrinology and Infertility Research
ave been treated with CC.35,36 Al-hough increased stromal cell prolifera-ion, but not the apoptosis, is consideredn initiator of decidualization and inap-ropriate uterine cell apoptosis results inhe failure of implantation,37 our resultsuggest that aberrant apoptotic activitiesf uterine stromal cells after chronic CCreatment may be a mechanism wherebyhe implantation process is disrupted. Ingreement with a previous study,runert et al38 reported that chronic
reatment with CC blocked the action of2 only in stromal cells and not in epi-
helial cells. Although in vitro tissue re-ombinant studies have shown that stro-al cells are able to regulate epithelial
ell proliferation in response to E2 stim-lation,19 our finding of massive damage
o the epithelial cell layer in rats that arereated with CC when E2 is absent (Fig-re 5, D) indicates that stromal cells may
ail to support and maintain their epithe-ial structure in response to CC when an-agonizing E2 in vivo. Indeed, the addi-ion of E2 after CC treatment in womenas been shown to improve cervical mu-us that results in increased endometrialhickness and conception rates.39 Collec-ively, our results suggest that negativeffects of CC on the transformation oftromal cells (direct) and the disintegra-ion of stromal-epithelial interactionshat lead to epithelial cell destructionindirect) may inhibit implantation inhe uterus. Although chronic CC treat-
ent increases endogenous E2 levels inats,16 it has been suggested that endo-etrial changes in women who are
reated with CC are due to an antiestro-enic effect of CC itself rather thanhanges in CC-induced hormonal lev-ls.40 The molecular mechanism of E2upplementation in the uterus of CC-reated rats suggests that estrogen statusf patients who are appropriate candi-ates for CC therapy should be taken
nto consideration to increase efficacy. Aetter understanding of the CC-inducedpoptotic mechanism in the uterusould reveal new therapeutic strategiesor the improvement of implantationates and reduction of the risk of miscar-iage in humans who are treated withC. For example, additional treatment
ith the antioxidant N-acetylated cys-FIGURE 5Histologic condition of the rat uterus
Lu
LeGe
S
Lu
Le
Ge
S
LuLe
S
Lu
Le
S
10 m
g/kg
CC
(4
d)
0.3
mg/
kg E
2 (4
d)
Veh
icle
(4
d)10
mg/
kg C
C (
4 d)
+
Veh
icle
(4
d)
100 µm
A
100 µm
A1
B B1
C C1
D D1
ematoxylin/eosin staining of uterine tissues from rats treated with A, A1, vehicle, B, B1, 17-�-stradiol (E2; 0.3 mg/kg) or C, C1, clomiphene citrate (CC; 10 mg/kg) for 4 consecutive days plus 4dditional days of vehicle injections (D, D1). Note that, although treatment with both 17-�-estradiolB1) and clomiphene citrate (C1) increases epithelial cell height, clear cells in the epithelial cell layerC1, arrows) are present only in clomiphene citrate-treated rats. Incomplete epithelial cell layers wereound in the 4-day clomiphene citrate plus 4-day vehicle-treated group (D1, arrow ). Uterine tissuesrom different rats (n � 5/group) were evaluated. All photographs were taken with a �10 or a �40agnification; the exact scale is given in the Figure.
e, glandular epithelial cells; Le, luminal epithelial cells; Lu, lumen; S, stromal cells.
utu. Clomiphene citrate treatment and rat uterus. Am J Obstet Gynecol 2010.
JULY 2010 American Journal of Obstetrics & Gynecology 65.e7
tclnas
tbtrerEemratdEEftosntit
cSmHpttttptdtmvttvvrstnq“
Wep�
dN
Research Reproductive Endocrinology and Infertility www.AJOG.org
65.e8 American Journal of Obstetrics & Gynecology JULY 2010
eine (an antiapoptotic agent) signifi-antly improves endometrial function andeads to increases in ovulation and preg-ancy rates, compared with CC treatmentlone, in women with polycystic ovaryyndrome.41
One of the most important findings ofhis study is that, although CC binds tooth ER-� and ER-� with approximatelyhe same affinity in vitro,42 the cellularesponse to CC is dependent on ER-�xpression and phosphorylation in theat uterus in vivo, which indicates thatR-� and ER-� are not functionallyquivalent in the response to CC treat-ent. Both ER subtypes are expressed in
odent uteri and human endometrium,11
nd either subtype can display diverseransactivational properties in a ligand-ependent manner when ER-� andR-� are coexpressed.43,44 Furthermore,R-� has the capacity to regulate ER-�
unction in vitro43,44 through the forma-ion of functional heterodimers (insteadf homodimers) for ER-mediated tran-cription.45 Our studies cannot elimi-ate the possibility that the agonist/an-
agonist activities of CC may also benfluenced by the relative expression ofhe 2 receptors in the uterus in vivo.
Successful implantation involves aomplex sequence of signaling events.8,17
everal genetically modified mouseodels with gene disruptions in ER, PR,oxa10, Hoxa11, p27, and p53 show im-
lantation defects,9,17,24 which suggestshat the expression and activation ofhese genes are essential to the implanta-ion process in vivo. In the mouse uterus,reatment with E2 increases ER-� ex-ression in stromal cells but not in epi-helial cells.11 It has been shown that theisruption of ER-�, but not ER-�, leadso failure of implantation in female
ice,9,11 which provides evidence for aital and distinctive role of the 2 ER sub-ypes in the uterus. Moreover, studieshat have used ER-� knockout mice inivo and uterine tissue recombinants initro have revealed that the estrogenicegulation of PR expression requirestromal ER-� but is independent of epi-helial ER-�.46 In addition, the mainte-ance of stromal PR expression is re-uired for the establishment of the
FIGURE 6Estrogen receptor (ER ) subtype activation and cell markers forimplantation during clomiphene citrate (CC )–induced uterine apoptosis
estern blot analysis was used to measure estrogen receptor �, phospho-estrogen receptor �,strogen receptor �, phospho-estrogen receptor �, Hoxa10, Hoxa11, p27, and p53 relative to wholerotein; gels were stained with Coomassie blue (n � 5 rats/group). Expression of pan-cytokeratin and-smooth muscle (SM) actin was evaluated in the same samples. Values are means � SEM. Theouble asterisks denote P � .01; the triple asterisks denote P � .001 vs vehicle-treated controls.utu. Clomiphene citrate treatment and rat uterus. Am J Obstet Gynecol 2010.
implantation window.”17,24 Previous
sfsPrstawEicaotaurtettbtTpwpgHtmdtpsimtEgcppi
mhnopmpniabbswfawutppt
AWmBOvi
R1erp12e23ecs4t25Mvew26ine27
UitNrc
�wsi�tL
NO
www.AJOG.org Reproductive Endocrinology and Infertility Research
tudies, however, could not distinguishully which stromal PR isoform was re-ponsive to E2 stimulation. AlthoughRA is the functional PR isoform that isesponsible for the regulation of uterinetromal differentiation during implanta-ion,9,11 it is interesting to note that CC isble not only to increase PRA expressionhen E2 is absent but is also able to block2-induced PRA expression in rat uter-
ne stromal cells, which suggests that CConverts from an estrogenic agonist ton estrogenic antagonist in the presencef E2. Previous studies from our labora-ory and others have shown that p27 is
target gene of PR signaling in theterus.26,47 Because CC does not bind di-ectly to the uterine PR18 and chronicreatment with CC fails to change endog-nous progesterone levels,16 we proposehat the antiestrogenic effects of CC onhe regulation of p27 expression seem toe mediated by ER-� and do not requirehe presence of progesterone directly.here is no change in the expression of53 after CC treatment in the uterus,hich suggests that divergent regulatoryathways between p27 and p53 are trig-ered by CC treatment. Although bothoxa10 and Hoxa11 are downstream
argets of the action of estrogens in theouse uterus,37 we observed only the
own-regulation of Hoxa11 in the CC-reated rat uterus. Because Hoxa11 is ex-ressed only in the stromal cells, our re-ults support the idea that the CC-nduced expression of stromal signals
ay control the epithelial fate. Together,hese data highlight the major role of theR-� signaling pathway in the antiestro-enic effect of CC on the uterine stromalells. Whether these molecules are inde-endent of 1 another in CC-induced im-lantation defects will require further
nvestigation.
17-�-estradiol (E2)-treated groups comparedith the C, 17-�-estradiol–treated group. Thesetainings were repeated in 5 rats/group with sim-lar results. All photographs were taken with a
10 magnification; the exact scale is given inhe Figure.e, luminal epithelial cells; Lu, lumen; S, stromal cells.
utu. Clomiphene citrate treatment and rat uterus. Am J
P
JULY 2010 Ameri
In terms of the cell-type selectionechanism in the rat uterus in vivo, we
ave reported direct molecular mecha-isms by which the antiestrogenic effectsf CC induce stromal cell-specific apo-tosis and regulate the expression ofolecules that are important for the im-
lantation process through the ER-� sig-aling pathway in the rat uterus. At least
n part, the present study has resulted incellular and molecular mechanism-
ased explanation for the discrepancyetween high ovulation rates and lowuccessful pregnancy rates in womenho are treated with CC.2,4,5 Moving
orward, it is noteworthy that there islso a need to evaluate such changes inomen who undergo CC treatment. These of endometrial biopsies in CC-
reated women who do not becomeregnant might help to explore our hy-otheses that have been generated fromhis study. f
CKNOWLEDGMENTSe thank Prof Håkan Billig for his support withany aspects of this project and Profs Christinaergh and Mats Brännström (Department ofbstetrics and Gynecology, Sahlgrenska Uni-ersity Hospital, Gothenburg, Sweden) for shar-ng their knowledge of clomiphene citrate.
EFERENCES. Turner RT, Evans GL, Sluka JP, et al. Differ-ntial responses of estrogen target tissues inats including bone to clomiphene, enclomi-hene, and zuclomiphene. Endocrinology998;139:3712-20.. Homburg R. Clomiphene citrate: end of anra? A mini-review. Hum Reprod 2005;20:043-51.. Palomba S, Russo T, Orio F Jr, et al. Uterineffects of clomiphene citrate in women with poly-ystic ovary syndrome: a prospective controlledtudy. Hum Reprod 2006;21:2823-9.. Benda JA. Clomiphene’s effect on endome-rium in infertility. Int J Gynecol Pathol 1992;11:73-82.. Samani FG, Farzadi L, Nezami N, TarzamniK, Soleimani F. Endometrial and follicular de-
elopment following letrozole intervention in un-xplained infertile patients failed to get pregnantith clomiphene citrate. Arch Gynecol Obstet009;280:201-5.. Hosie M, Adamson M, Penny C. Actin bind-
ng protein expression is altered in uterine lumi-al epithelium by clomiphene citrate, a syntheticstrogen receptor modulator. Theriogenology008;69:700-13.. Dickey RP, Taylor SN, Curole DN, Rye PH,
FIGURE 7Expression of progesteronereceptor A (PRA) isoformin the stromal cell layersof the rat uterus
terine progesterone receptor A immunoreactiv-ty is absent in the A, vehicle group but is seen inhe B, clomiphene citrate (CC)–treated group.ote that the immunoreactivity of progesterone
eceptor A is less intense in the B, clomipheneitrate-treated and the D, clomiphene citrate
bstet Gynecol 2010.
yrzak R. Incidence of spontaneous abortion incan Journal of Obstetrics & Gynecology 65.e9

c18Gh29Mtc1Ctt81tA1Kes1Mtm1Td3E41sm11mspp1c31dt1nli22nd12odM2zm
tP2omR2Drp12Pip32Nfcf22pnpt12Domr72mpp3HdcE3MeoO3Hnip3Stprc3m
3cei13fdw13o3npm33pm44Bpmcm14NddS4Eda14cttrd4Eu14GfeE4Letp4p
Research Reproductive Endocrinology and Infertility www.AJOG.org
6
lomiphene pregnancies. Hum Reprod 1996;1:2623-8.. Diedrich K, Fauser BC, Devroey P, Griesinger. The role of the endometrium and embryo inuman implantation. Hum Reprod Update007;13:365-77.. DeMayo FJ, Zhao B, Takamoto N, Tsai SY.echanisms of action of estrogen and proges-
erone. Ann N Y Acad Sci 2002;955:48-59; dis-ussion 86-8, 396-406.0. Kuiper GG, Carlsson B, Grandien K, et al.omparison of the ligand binding specificity and
ranscript tissue distribution of estrogen recep-ors alpha and beta. Endocrinology 1997;138:63-70.1. Moutsatsou P, Sekeris CE. Steroid recep-ors in the uterus: implications in endometriosis.nn N Y Acad Sci 2003;997:209-22.2. Yin Y, Huang WW, Lin C, Chen H, Mac-enzie A, Ma L. Estrogen suppresses uterinepithelial apoptosis by inducing birc1 expres-ion. Mol Endocrinol 2008;22:113-25.3. Song J, Rutherford T, Naftolin F, Brown S,or G. Hormonal regulation of apoptosis and
he Fas and Fas ligand system in human endo-etrial cells. Mol Hum Reprod 2002;8:447-55.4. Chaube SK, Prasad PV, Tripathi V, ShrivastavG. Clomiphene citrate inhibits gonadotropin-in-uced ovulation by reducing cyclic adenosine’,5’-cyclic monophosphate and prostaglandin2 levels in rat ovary. Fertil Steril 2006;86(suppl):1106-11.5. Kokawa K, Shikone T, Nakano R. Apopto-is in human chorionic villi and decidua in nor-al and ectopic pregnancy. Mol Hum Reprod998;4:87-91.6. Shao R, Nutu M, Weijdegard BE, et al. Clo-iphene citrate causes aberrant tubal apopto-
is and estrogen receptor activation in rat fallo-ian tube: implications for tubal ectopicregnancy. Biol Reprod 2009;80:1262-71.7. Dey SK, Lim H, Das SK, et al. Molecularues to implantation. Endocr Rev 2004;25:41-73.8. Barkai U, Kidron T, Kraicer PF. Inhibition ofecidual induction in rats by clomiphene andamoxifen. Biol Reprod 1992;46:733-9.9. Groothuis PG, Dassen HH, Romano A, Pu-yadeera C. Estrogen and the endometrium:
essons learned from gene expression profilingn rodents and human. Hum Reprod Update007;13:405-17.0. Gupta JS, Roy SK. Effect of clomiphene onuclear estrogen receptor of the fallopian tubeuring ovum transport in rabbits. Endocr Res989;15:339-53.1. Nagao T, Yoshimura S. Oral administrationf clomiphene to neonatal rats causes repro-uctive tract abnormalities. Teratog Carcinogutagen 2001;21:213-21.2. Cunha GR, Taguchi O, Namikawa R, Nishi-uka Y, Robboy SJ. Teratogenic effects of clo-
iphene, tamoxifen, and diethylstilbestrol on (5.e10 American Journal of Obstetrics & Gynecolo
he developing human female genital tract. Humathol 1987;18:1132-43.3. Birkenfeld A, Mootz U, Beier HM. The effectf clomiphene citrate on blastocyst develop-ent and implantation in the rabbit. Cell Tissuees 1985;241:495-503.4. Franco HL, Jeong JW, Tsai SY, Lydon JP,eMayo FJ. In vivo analysis of progesterone
eceptor action in the uterus during embryo im-lantation. Semin Cell Dev Biol 2008;19:78-86.5. Young RL, Goldzieher JW, Chakraborty PK,anko WB, Bridges CN. Qualitative differences
n estrogenic/antiestrogenic effects of clomi-hene and zuclomiphene. Int J Fertil 1991;6:291-5.6. Shao R, Weijdegard B, Ljungstrom K, et al.uclear progesterone receptor A and B iso-
orms in mouse fallopian tube and uterus: impli-ations for expression, regulation, and cellularunction. Am J Physiol Endocrinol Metab006;291:E59-72.7. Samalecos A, Gellersen B. Systematic ex-ression analysis and antibody screening doot support the existence of naturally occurringrogesterone receptor (PR)-C, PR-M, or otherruncated PR isoforms. Endocrinology 2008;49:5872-87.8. Jordan VC, Rowsby L, Dix CJ, Prestwich G.ose-related effects of non-steroidal anti-estrogens and oestrogens on the measure-ent of cytoplasmic oestrogen receptors in the
at and mouse uterus. J Endocrinol 1978;8:71-81.9. Gonen Y, Casper RF. Sonographic deter-ination of a possible adverse effect of clomi-hene citrate on endometrial growth. Hum Re-rod 1990;5:670-4.0. Homburg R, Pap H, Brandes M, Huirne J,ompes P, Lambalk CB. Endometrial biopsyuring induction of ovulation with clomipheneitrate in polycystic ovary syndrome. Gynecolndocrinol 2006;22:506-10.1. Unfer V, Costabile L, Gerli S, Papaleo E,arelli G, Di Renzo GC. Low dose of ethinyl
stradiol can reverse the antiestrogenic effectsf clomiphene citrate on endometrium. Gynecolbstet Invest 2001;51:120-3.2. Yagel S, Ben-Chetrit A, Anteby E, Zacut D,ochner-Celnikier D, Ron M. The effect of ethi-yl estradiol on endometrial thickness and uter-
ne volume during ovulation induction by clomi-hene citrate. Fertil Steril 1992;57:33-6.3. Markaverich BM, Upchurch S, McCormackA, Glasser SR, Clark JH. Differential stimula-
ion of uterine cells by nafoxidine and clomi-hene: relationship between nuclear estrogeneceptors and type II estrogen binding sites andellular growth. Biol Reprod 1981;24:171-81.4. Shelesnyak MC, Tic L. Studies on theechanism of decidualization. Acta Endocrinol
Copenh) 1963;42:465-72. A
gy JULY 2010
5. Fritz MA, Holmes RT, Keenan EJ. Effect oflomiphene citrate treatment on endometrialstrogen and progesterone receptor induction
n women. Am J Obstet Gynecol 1991;165:77-85.6. Fritz MA, Westfahl PK, Graham RL. The ef-
ect of luteal phase estrogen antagonism on en-ometrial development and luteal function inomen. J Clin Endocrinol Metab 1987;65:006-13.7. Daftary GS, Taylor HS. Endocrine regulationf HOX genes. Endocr Rev 2006;27:331-55.8. Grunert G, Neumann G, Porcia M, Tcher-itchin AN. The estrogenic responses to clomi-hene in the different cell types of the rat uterus:orphometrical evaluation. Biol Reprod 1987;7:527-38.9. Dickey RP, Holtkamp DE. Development,harmacology and clinical experience with clo-iphene citrate. Hum Reprod Update 1996;2:83-506.0. Massai MR, de Ziegler D, Lesobre V,ergeron C, Frydman R, Bouchard P. Clomi-hene citrate affects cervical mucus and endo-etrial morphology independently of the
hanges in plasma hormonal levels induced byultiple follicular recruitment. Fertil Steril993;59:1179-86.1. Badawy A, State O, Abdelgawad S.-Acetyl cysteine and clomiphene citrate for in-uction of ovulation in polycystic ovary syn-rome: a cross-over trial. Acta Obstet Gynecolcand 2007;86:218-22.2. Paige LA, Christensen DJ, Gron H, et al.strogen receptor (ER) modulators each induceistinct conformational changes in ER alphand ER beta. Proc Natl Acad Sci USA999;96:3999-4004.3. Hall JM, McDonnell DP. The estrogen re-eptor beta-isoform (ERbeta) of the human es-rogen receptor modulates ERalpha transcrip-ional activity and is a key regulator of the cellularesponse to estrogens and antiestrogens. En-ocrinology 1999;140:5566-78.4. Pettersson K, Delaunay F, Gustafsson JA.strogen receptor beta acts as a dominant reg-lator of estrogen signaling. Oncogene 2000;9:4970-8.5. Pettersson K, Grandien K, Kuiper GG,ustafsson JA. Mouse estrogen receptor beta
orms estrogen response element-binding het-rodimers with estrogen receptor alpha. Molndocrinol 1997;11:1486-96.6. Kurita T, Lee KJ, Cooke PS, Taylor JA,ubahn DB, Cunha GR. Paracrine regulation ofpithelial progesterone receptor by estradiol inhe mouse female reproductive tract. Biol Re-rod 2000;62:821-30.7. Moutsatsou P, Sekeris CE. Estrogen androgesterone receptors in the endometrium.
nn N Y Acad Sci 1997;816:99-115.
Related Documents











