The FASEB Journal • Research Communication Stress-inducible and constitutive phosphoinositide pools have distinctive fatty acid patterns in Arabidopsis thaliana Sabine Ko ¨nig, Alina Mosblech, and Ingo Heilmann 1 Department of Plant Biochemistry, Albrecht-von-Haller-Institute for Plant Sciences, Georg-August- University Go ¨ttingen, Go ¨ttingen, Germany ABSTRACT Function and development of eukaryotic cells require tight control of diverse physiological pro- cesses. Numerous cellular processes are regulated by polyphosphoinositides, which interact with protein partners or mediate release of the second messenger, inositol 1,4,5-trisphosphate (InsP 3 ). Emerging evidence suggests that different regulatory or signaling functions of polyphosphoinositides may be orchestrated by the establishment of distinct subcellular pools; the princi- ples underlying pool-formation are, however, not un- derstood. Arabidopsis plants exhibit transient increases in polyphosphoinositides with hyperosmotic stress, providing a model for comparing constitutive and stress-inducible polyphosphoinositide pools. Using a combination of thin-layer-chromatography and gas- chromatography, phospholipids from stressed and nonstressed Arabidopsis plants were analyzed for their associated fatty acids. Under nonstress conditions structural phospholipids and phosphatidylinositol con- tained 50 –70 mol% polyunsaturated fatty acids (PUFA), whereas polyphosphoinositides were more sat- urated (10 –20 mol% PUFA). With hyperosmotic stress polyphosphoinositides with up to 70 mol% PUFA were formed that differed from constitutive species and coincided with a transient loss in unsaturated phospha- tidylinositol. The patterns indicate inducible turnover of an unsaturated phosphatidylinositol pool, which accumulates under standard conditions and is primed for phosphorylation on stimulation. Metabolic analysis of wild-type and transgenic plants disturbed in phos- phoinositide metabolism suggests that, in contrast to saturated species, unsaturated polyphosphoinositides are channeled toward InsP 3 -production.—Ko ¨nig, S., Mosblech, A., Heilmann, I. Stress-inducible and consti- tutive phosphoinositide pools have distinctive fatty acid patterns in Arabidopsis thaliana. FASEB J. 21, 1958 –1967 (2007) Key Words: hyperosmotic stress plant PtdIns (4,5) P 2 pools signal transduction Phosphoinositides are centrally involved in the regulation of a multitude of cellular processes in ani- mal, fungal, and plant systems, and these regulatory functions have been extensively reviewed (1–5). Phos- phoinositide regulation involves the interaction of the lipids with protein partners that may be recruited to target membranes or are modified in their biochemical activity by their phosphoinositide ligands. Examples from both the plant and animal research fields include the regulation of ion-channel or ATPase activity (6, 7), cytoskeletal dynamics (8 –11), and hormonal and stress signaling (1–5). As yet, the involvement in vesicle trafficking has only been demonstrated with animal cells (8, 12). There is an obvious need for tight spatial and temporal regulation of phosphoinositide-protein interactions in all eukaryotes, because a phosphoinosi- tide may bind to various alternative partners, with potentially conflicting effects on physiology. Emerging evidence suggests that phosphoinositide-protein in- teractions are restricted according to physiological requirements by compartmentation into autono- mous phosphoinositide pools. Phosphatidylinositol 4,5-bisphosphate [PtdIns(4,5)P 2 ] is the best-studied regulatory phosphoinositide and can, in both plants and animals, perform cellular functions either as an intact lipid ligand for protein partners (13–15) or as a substrate for phospholipase C (PLC), which hydro- lyzes the lipid into inositol 1,4,5-trisphosphate (InsP 3 ) and diacylglycerol (DAG) (3, 16). The list of PtdIns(4,5)P 2 functions reported from various eu- karyotic systems (1–5) suggests the presence of vari- ous physiological PtdIns(4,5)P 2 pools compartmen- talized along organellar borders or in microdomains within one membrane. Whereas the presence of such independent pools of PtdIns(4,5)P 2 has been dem- onstrated before in plants and animals (17–20), previous studies from the animal field have capitalized on the recognition of polyphosphoinositide head- groups by various target proteins (14, 21), and so far the lipids constituting distinct phosphoinositide pools have not been characterized regarding differences in 1 Correspondence: Department of Plant Biochemistry, Al- brecht-von-Haller-Institute for Plant Sciences, Georg-August- University Go ¨ ttingen, Justus-von-Liebig-Weg 11, 37077 Go ¨ttin- gen, Germany. E-mail: [email protected] doi: 10.1096/fj.06-7887com 1958 0892-6638/07/0021-1958 © FASEB

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The FASEB Journal • Research Communication
Stress-inducible and constitutive phosphoinositide poolshave distinctive fatty acid patterns in Arabidopsisthaliana
Sabine Konig, Alina Mosblech, and Ingo Heilmann1
Department of Plant Biochemistry, Albrecht-von-Haller-Institute for Plant Sciences, Georg-August-University Gottingen, Gottingen, Germany
ABSTRACT Function and development of eukaryoticcells require tight control of diverse physiological pro-cesses. Numerous cellular processes are regulated bypolyphosphoinositides, which interact with proteinpartners or mediate release of the second messenger,inositol 1,4,5-trisphosphate (InsP3). Emerging evidencesuggests that different regulatory or signaling functionsof polyphosphoinositides may be orchestrated by theestablishment of distinct subcellular pools; the princi-ples underlying pool-formation are, however, not un-derstood. Arabidopsis plants exhibit transient increasesin polyphosphoinositides with hyperosmotic stress,providing a model for comparing constitutive andstress-inducible polyphosphoinositide pools. Using acombination of thin-layer-chromatography and gas-chromatography, phospholipids from stressed andnonstressed Arabidopsis plants were analyzed for theirassociated fatty acids. Under nonstress conditionsstructural phospholipids and phosphatidylinositol con-tained 50–70 mol% polyunsaturated fatty acids(PUFA), whereas polyphosphoinositides were more sat-urated (10–20 mol% PUFA). With hyperosmotic stresspolyphosphoinositides with up to 70 mol% PUFA wereformed that differed from constitutive species andcoincided with a transient loss in unsaturated phospha-tidylinositol. The patterns indicate inducible turnoverof an unsaturated phosphatidylinositol pool, whichaccumulates under standard conditions and is primedfor phosphorylation on stimulation. Metabolic analysisof wild-type and transgenic plants disturbed in phos-phoinositide metabolism suggests that, in contrast tosaturated species, unsaturated polyphosphoinositidesare channeled toward InsP3-production.—Konig, S.,Mosblech, A., Heilmann, I. Stress-inducible and consti-tutive phosphoinositide pools have distinctive fatty acidpatterns in Arabidopsis thaliana. FASEB J. 21, 1958–1967(2007)
Key Words: hyperosmotic stress � plant � PtdIns (4,5) P2 pools� signal transduction
Phosphoinositides are centrally involved in theregulation of a multitude of cellular processes in ani-mal, fungal, and plant systems, and these regulatory
functions have been extensively reviewed (1–5). Phos-phoinositide regulation involves the interaction of thelipids with protein partners that may be recruited totarget membranes or are modified in their biochemicalactivity by their phosphoinositide ligands. Examplesfrom both the plant and animal research fields includethe regulation of ion-channel or ATPase activity (6, 7),cytoskeletal dynamics (8–11), and hormonal and stresssignaling (1–5). As yet, the involvement in vesicletrafficking has only been demonstrated with animalcells (8, 12). There is an obvious need for tight spatialand temporal regulation of phosphoinositide-proteininteractions in all eukaryotes, because a phosphoinosi-tide may bind to various alternative partners, withpotentially conflicting effects on physiology. Emergingevidence suggests that phosphoinositide-protein in-teractions are restricted according to physiologicalrequirements by compartmentation into autono-mous phosphoinositide pools. Phosphatidylinositol4,5-bisphosphate [PtdIns(4,5)P2] is the best-studiedregulatory phosphoinositide and can, in both plantsand animals, perform cellular functions either as anintact lipid ligand for protein partners (13–15) or asa substrate for phospholipase C (PLC), which hydro-lyzes the lipid into inositol 1,4,5-trisphosphate(InsP3) and diacylglycerol (DAG) (3, 16). The list ofPtdIns(4,5)P2 functions reported from various eu-karyotic systems (1–5) suggests the presence of vari-ous physiological PtdIns(4,5)P2 pools compartmen-talized along organellar borders or in microdomainswithin one membrane. Whereas the presence of suchindependent pools of PtdIns(4,5)P2 has been dem-onstrated before in plants and animals (17–20),previous studies from the animal field have capitalizedon the recognition of polyphosphoinositide head-groups by various target proteins (14, 21), and so farthe lipids constituting distinct phosphoinositide poolshave not been characterized regarding differences in
1 Correspondence: Department of Plant Biochemistry, Al-brecht-von-Haller-Institute for Plant Sciences, Georg-August-University Gottingen, Justus-von-Liebig-Weg 11, 37077 Gottin-gen, Germany. E-mail: [email protected]
doi: 10.1096/fj.06-7887com
1958 0892-6638/07/0021-1958 © FASEB

fatty acid compositions, which may define distinct mo-lecular lipid species.
The key hypothesis of this study is that phosphoi-nositides constituting distinct physiological pools maydiffer in their fatty acid make-up. In this context notethat preferences for phosphoinositide substrates withcertain fatty acid compositions have been demon-strated in vitro for recombinant enzymes, includingphosphatidylinositolphosphate kinase (PIP kinase) andPLC from insect cells (22) and different phosphoinosi-tide phosphatases deficient in patients suffering fromLowe’s oculocerebrorenal syndrome (23). So far, it isnot clear whether such preferences have physiologicalrelevance and how association of phosphoinositideswith functional pools correlates with fatty acid patternsof the lipids.
The generation of PtdIns(4,5)P2 species with distinctfatty acid compositions would in part be defined by thebiosynthetic enzymes. The parent lipid, PtdIns, is syn-thesized in the endoplasmic reticulum [ER, (24, 25)]from cytidinediphospho-diacylglycerol (CDP-DAG)and myo-inositol by phosphatidylinositol synthase (PIsynthase) and is distributed to various cellular locationsof plant and animal cells by an unknown mechanismpossibly involving targeted vesicle flow or the action oflipid transfer proteins (26, 27). PtdIns can be sequen-tially phosphorylated by phosphatidylinositol kinases(PI kinases) and PIP kinases, which occur in numerousisoforms in Arabidopsis, encoded by multigene families(4). PI kinase and PIP kinase activities have been foundassociated with various plant subcellular fractions, in-cluding the plasma membrane, the actin cytoskeleton,and endomembrane compartments (4, 5, 28), and itcan reasonably be concluded that PtdIns(4,5)P2 can begenerated from PtdIns in compartmentalized pools.
PIP kinase activity has been shown to increase tran-siently after application of various stresses in plants andalgae (17, 29, 30), and in animal systems differentialactivation of PIP kinase isoforms associated with differ-ent phosphoinositide pools is being discussed as ameans of harmonizing diverse physiological roles ofPtdIns(4,5)P2 (31, 32). Transient increases in the levelsof regulatory phospholipids have previously been re-ported for Arabidopsis plants challenged by osmoticstimulation, including those for PtdIns(4,5)P2 (33, 34)and phosphatidic acid (PtdOH) (35). Based on strongcorrelative evidence, PtdIns(4,5)P2 transiently increas-ing in Arabidopsis plants with osmotic stress has beensuggested to be a substrate for PLC, generating InsP3and acting as part of the osmotic signaling cascadeleading to Ca2� release from internal stores (33).Although downstream effects of phosphoinositide sig-naling with osmotic stress are not well defined in plants,phosphoinositides transiently increasing with hyperos-motic stress clearly represent a physiological pool func-tionally different from the majority of constitutivephosphoinositide pools of nonchallenged plants.
In this study, PtdIns(4,5)P2, its precursors, and im-mediate derivatives were chosen for the targeted bio-chemical characterization of associated fatty acid com-
positions, because PtdIns(4,5)P2 is a multifunctionalsignaling component and is well-characterized regard-ing its stress-inducible and constitutive dynamics (33,34). Whereas a number of studies have characterizedpolyphosphoinositide molecular species from nonchal-lenged animal systems (36–39), no such information isavailable for plants. Here we show that PtdIns specieswith characteristic fatty acyl moieties can supply differ-ent physiological pools of polyphosphoinositides thatare defined by their constitutive nature or by theirstress-inducible dynamics. The data presented indicatea discrete function of highly unsaturated molecularphosphoinositide species in InsP3 production duringosmotic stress.
MATERIALS AND METHODS
Plant growth and stress treatment
Arabidopsis ecotype Columbia (col-0) plants were grown onsoil under exposure to 140 �mol photons m–2 sec–1 of light inan 8 h light and 16 h dark regime. Rosette leaves wereharvested at 6 wk and frozen in liquid nitrogen. Plant materialfrom 6–8 plants was combined. Arabidopsis ecotype Columbia(col-0) plants and col-0 plants expressing the human type Iinositolpolyphosphate 5-phosphatase [(40), expression line2–8 (41)] destined for stress treatments were grown understerile conditions in sealed jars on 0.5% Murashige and Skoogmedium including modified vitamins (Duchefa) containing1% (w/w) sucrose and 0.25% (w/w) Gelrite (Roth). After14 d plants were transferred to hydroponic cultures in liquidmedia as described (42). Hydroponic cultures were exposedto 140 �mol photons m–2 sec–1 of light in an 8 h light and16 h dark regime and continuously aerated. Eight to ten-week-old plants were treated by adding NaCl or sorbitol infinal concentrations of 0.4 M and 0.8 M, respectively, to thehydroponic media. Rosette leaves were harvested beforetreatment and after various periods of stimulation, as indi-cated in the results section, and immediately frozen in liquidnitrogen. Care was taken to perform experiments over thesame day-time period within the light-dark-regime, and not tocross the light-dark transition.
Lipid extraction and biochemical analyses
Plant material was ground under liquid nitrogen to a finepowder. Polyphosphoinositides were extracted from pow-dered plant material by using an acidic extraction protocol(43). Lipids were separated by thin-layer-chromatography(TLC) on silica gel plates (Merck) using developing solventsfor optimal resolution: for phosphoinositides and PtdOH,CHCl3:CH3OH:NH4OH:H2O [57:50:4:11(v/v/v/v)] (44); forPtdCho and PtdEtn, acetone:toluol:water [91:30:7 (v/v/v)](45); for isolating phosphatidylinositol, CHCl3:methyl ace-tate:isopropanol:CH3OH:0.25% aqueous potassium chloride[25:25:25:10:9 (v/v/v/v/v)] (46). Lanes with lipid standards(5 �g) run in parallel to biological samples were cut andlipids were visualized in aqueous 10% (w/w) CuSO4 (Sigma,St. Louis, MO, USA) containing 8% H3PO4 (Sigma) andsubsequent heating to 180°C. Unstained lipids were locatedon the remaining parts of the TLC plates, according tostandard migration; scraped; redissolved in their respectivedeveloping solvents; and dried under N2 flow. Lipids weretransmethylated (47), fatty acid methyl esters dissolved inacetonitril and analyzed using a GC6890 gas chromatograph
1959FATTY ACID PATTERNS OF POLYPHOSPHOINOSITIDE POOLS
with flame-ionization detection (Agilent, Boblingen, Ger-many) fitted with a 30 m � 250 �m DB-23 capillary column(Agilent). Helium flowed as a carrier gas at 1 ml min–1.Samples were injected at 220°C. After 1 min at 150°C, theoven temperature was raised to 200°C at a rate of 8°C min–1,then to 250°C at 25°C min–1, and then kept at 250°C for 6min. Fatty acids were identified according to authentic stan-dards and by their characteristic mass spectrometric fragmen-tation patterns (data not shown), and quantified according tointernal tripentadecanoic acid standards of known concentra-tion. Variation in fatty acid patterns obtained with materialsampled on different days did not exceed that denoted by sd.Due to limiting material in samples representing isolatedminor lipids, fatty acids of low abundance may be absent fromfatty acid patterns. InsP3 levels were determined using the[3H]InsP3 receptor binding assay system (GE Healthcare) asdescribed previously (29).
RESULTS
Arabidopsis polyphosphoinositides are moresaturated than PtdIns
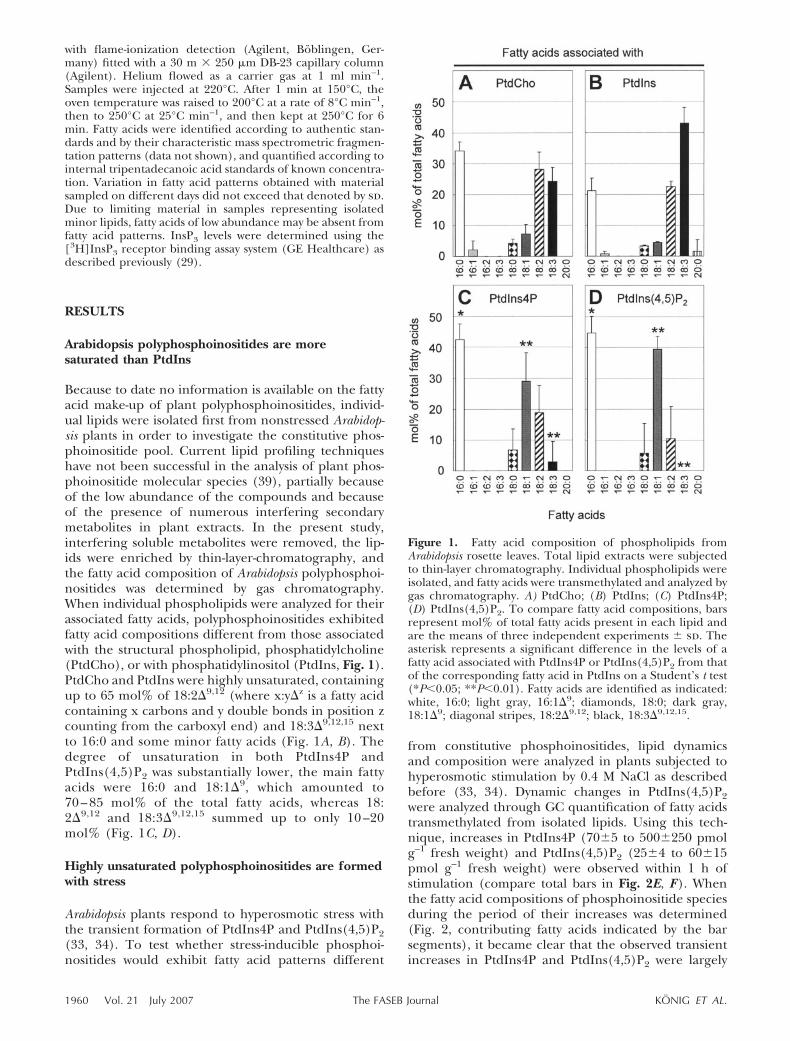
Because to date no information is available on the fattyacid make-up of plant polyphosphoinositides, individ-ual lipids were isolated first from nonstressed Arabidop-sis plants in order to investigate the constitutive phos-phoinositide pool. Current lipid profiling techniqueshave not been successful in the analysis of plant phos-phoinositide molecular species (39), partially becauseof the low abundance of the compounds and becauseof the presence of numerous interfering secondarymetabolites in plant extracts. In the present study,interfering soluble metabolites were removed, the lip-ids were enriched by thin-layer-chromatography, andthe fatty acid composition of Arabidopsis polyphosphoi-nositides was determined by gas chromatography.When individual phospholipids were analyzed for theirassociated fatty acids, polyphosphoinositides exhibitedfatty acid compositions different from those associatedwith the structural phospholipid, phosphatidylcholine(PtdCho), or with phosphatidylinositol (PtdIns, Fig. 1).PtdCho and PtdIns were highly unsaturated, containingup to 65 mol% of 18:2�9,12 (where x:y�z is a fatty acidcontaining x carbons and y double bonds in position zcounting from the carboxyl end) and 18:3�9,12,15 nextto 16:0 and some minor fatty acids (Fig. 1A, B). Thedegree of unsaturation in both PtdIns4P andPtdIns(4,5)P2 was substantially lower, the main fattyacids were 16:0 and 18:1�9, which amounted to70 – 85 mol% of the total fatty acids, whereas 18:2�9,12 and 18:3�9,12,15 summed up to only 10 –20mol% (Fig. 1C, D).
Highly unsaturated polyphosphoinositides are formedwith stress
Arabidopsis plants respond to hyperosmotic stress withthe transient formation of PtdIns4P and PtdIns(4,5)P2(33, 34). To test whether stress-inducible phosphoi-nositides would exhibit fatty acid patterns different
from constitutive phosphoinositides, lipid dynamicsand composition were analyzed in plants subjected tohyperosmotic stimulation by 0.4 M NaCl as describedbefore (33, 34). Dynamic changes in PtdIns(4,5)P2were analyzed through GC quantification of fatty acidstransmethylated from isolated lipids. Using this tech-nique, increases in PtdIns4P (70�5 to 500�250 pmolg–1 fresh weight) and PtdIns(4,5)P2 (25�4 to 60�15pmol g–1 fresh weight) were observed within 1 h ofstimulation (compare total bars in Fig. 2E, F). Whenthe fatty acid compositions of phosphoinositide speciesduring the period of their increases was determined(Fig. 2, contributing fatty acids indicated by the barsegments), it became clear that the observed transientincreases in PtdIns4P and PtdIns(4,5)P2 were largely
Figure 1. Fatty acid composition of phospholipids fromArabidopsis rosette leaves. Total lipid extracts were subjectedto thin-layer chromatography. Individual phospholipids wereisolated, and fatty acids were transmethylated and analyzed bygas chromatography. A) PtdCho; (B) PtdIns; (C) PtdIns4P;(D) PtdIns(4,5)P2. To compare fatty acid compositions, barsrepresent mol% of total fatty acids present in each lipid andare the means of three independent experiments � sd. Theasterisk represents a significant difference in the levels of afatty acid associated with PtdIns4P or PtdIns(4,5)P2 from thatof the corresponding fatty acid in PtdIns on a Student’s t test(*P�0.05; **P�0.01). Fatty acids are identified as indicated:white, 16:0; light gray, 16:1�9; diamonds, 18:0; dark gray,18:1�9; diagonal stripes, 18:2�9,12; black, 18:3�9,12,15.
1960 Vol. 21 July 2007 KONIG ET AL.The FASEB Journal
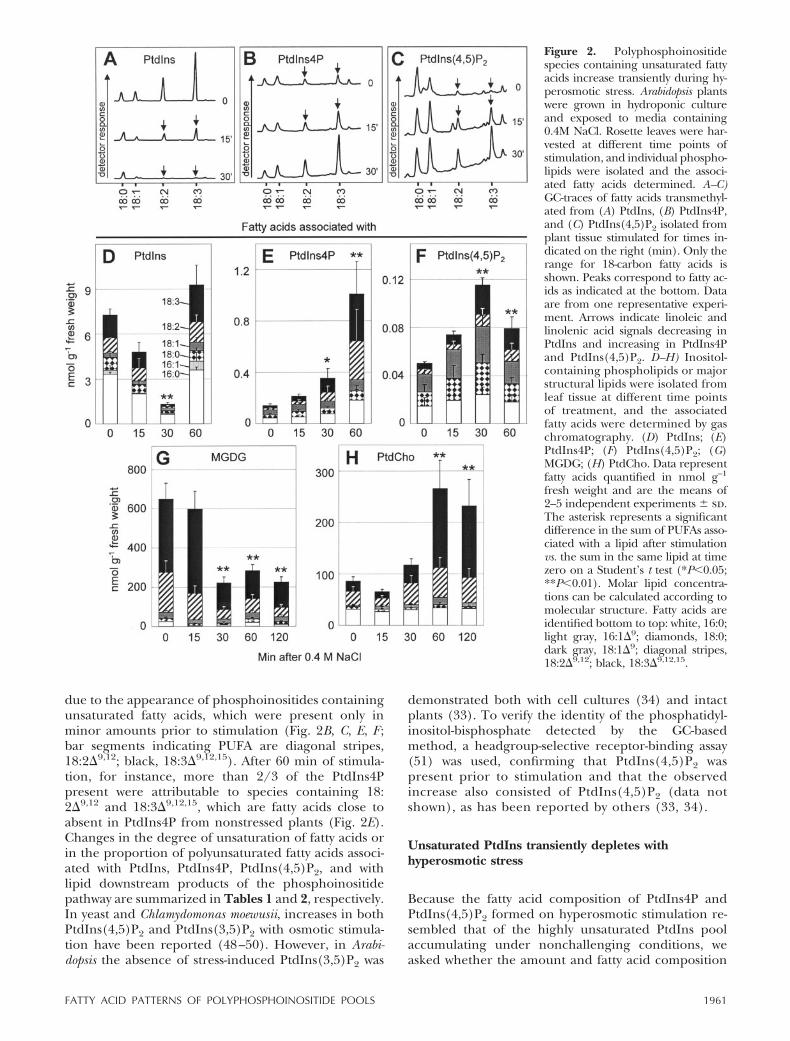
due to the appearance of phosphoinositides containingunsaturated fatty acids, which were present only inminor amounts prior to stimulation (Fig. 2B, C, E, F ;bar segments indicating PUFA are diagonal stripes,18:2�9,12; black, 18:3�9,12,15). After 60 min of stimula-tion, for instance, more than 2/3 of the PtdIns4Ppresent were attributable to species containing 18:2�9,12 and 18:3�9,12,15, which are fatty acids close toabsent in PtdIns4P from nonstressed plants (Fig. 2E).Changes in the degree of unsaturation of fatty acids orin the proportion of polyunsaturated fatty acids associ-ated with PtdIns, PtdIns4P, PtdIns(4,5)P2, and withlipid downstream products of the phosphoinositidepathway are summarized in Tables 1 and 2, respectively.In yeast and Chlamydomonas moewusii, increases in bothPtdIns(4,5)P2 and PtdIns(3,5)P2 with osmotic stimula-tion have been reported (48–50). However, in Arabi-dopsis the absence of stress-induced PtdIns(3,5)P2 was
demonstrated both with cell cultures (34) and intactplants (33). To verify the identity of the phosphatidyl-inositol-bisphosphate detected by the GC-basedmethod, a headgroup-selective receptor-binding assay(51) was used, confirming that PtdIns(4,5)P2 waspresent prior to stimulation and that the observedincrease also consisted of PtdIns(4,5)P2 (data notshown), as has been reported by others (33, 34).
Unsaturated PtdIns transiently depletes withhyperosmotic stress
Because the fatty acid composition of PtdIns4P andPtdIns(4,5)P2 formed on hyperosmotic stimulation re-sembled that of the highly unsaturated PtdIns poolaccumulating under nonchallenging conditions, weasked whether the amount and fatty acid composition
Figure 2. Polyphosphoinositidespecies containing unsaturated fattyacids increase transiently during hy-perosmotic stress. Arabidopsis plantswere grown in hydroponic cultureand exposed to media containing0.4M NaCl. Rosette leaves were har-vested at different time points ofstimulation, and individual phospho-lipids were isolated and the associ-ated fatty acids determined. A–C)GC-traces of fatty acids transmethyl-ated from (A) PtdIns, (B) PtdIns4P,and (C) PtdIns(4,5)P2 isolated fromplant tissue stimulated for times in-dicated on the right (min). Only therange for 18-carbon fatty acids isshown. Peaks correspond to fatty ac-ids as indicated at the bottom. Dataare from one representative experi-ment. Arrows indicate linoleic andlinolenic acid signals decreasing inPtdIns and increasing in PtdIns4Pand PtdIns(4,5)P2. D–H) Inositol-containing phospholipids or majorstructural lipids were isolated fromleaf tissue at different time pointsof treatment, and the associatedfatty acids were determined by gaschromatography. (D) PtdIns; (E)PtdIns4P; (F) PtdIns(4,5)P2; (G)MGDG; (H) PtdCho. Data representfatty acids quantified in nmol g–1
fresh weight and are the means of2–5 independent experiments � sd.The asterisk represents a significantdifference in the sum of PUFAs asso-ciated with a lipid after stimulationvs. the sum in the same lipid at timezero on a Student’s t test (*P�0.05;**P�0.01). Molar lipid concentra-tions can be calculated according tomolecular structure. Fatty acids areidentified bottom to top: white, 16:0;light gray, 16:1�9; diamonds, 18:0;dark gray, 18:1�9; diagonal stripes,18:2�9,12; black, 18:3�9,12,15.
1961FATTY ACID PATTERNS OF POLYPHOSPHOINOSITIDE POOLS
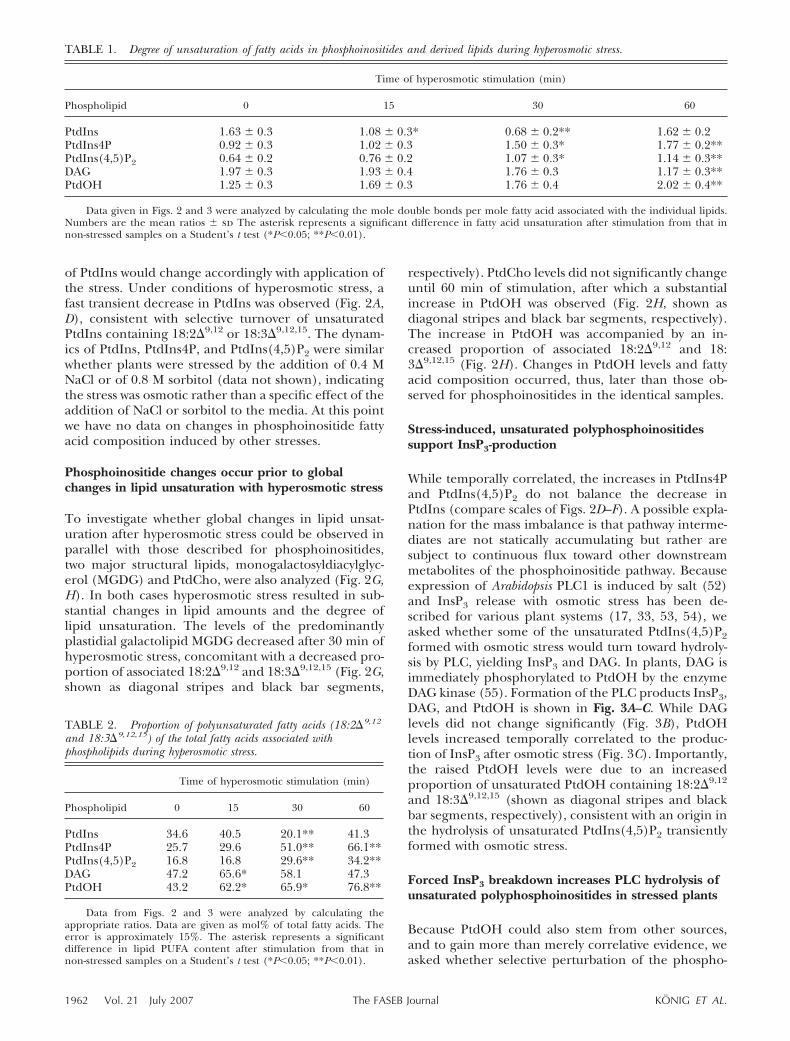
of PtdIns would change accordingly with application ofthe stress. Under conditions of hyperosmotic stress, afast transient decrease in PtdIns was observed (Fig. 2A,D), consistent with selective turnover of unsaturatedPtdIns containing 18:2�9,12 or 18:3�9,12,15. The dynam-ics of PtdIns, PtdIns4P, and PtdIns(4,5)P2 were similarwhether plants were stressed by the addition of 0.4 MNaCl or of 0.8 M sorbitol (data not shown), indicatingthe stress was osmotic rather than a specific effect of theaddition of NaCl or sorbitol to the media. At this pointwe have no data on changes in phosphoinositide fattyacid composition induced by other stresses.
Phosphoinositide changes occur prior to globalchanges in lipid unsaturation with hyperosmotic stress
To investigate whether global changes in lipid unsat-uration after hyperosmotic stress could be observed inparallel with those described for phosphoinositides,two major structural lipids, monogalactosyldiacylglyc-erol (MGDG) and PtdCho, were also analyzed (Fig. 2G,H). In both cases hyperosmotic stress resulted in sub-stantial changes in lipid amounts and the degree oflipid unsaturation. The levels of the predominantlyplastidial galactolipid MGDG decreased after 30 min ofhyperosmotic stress, concomitant with a decreased pro-portion of associated 18:2�9,12 and 18:3�9,12,15 (Fig. 2G,shown as diagonal stripes and black bar segments,
respectively). PtdCho levels did not significantly changeuntil 60 min of stimulation, after which a substantialincrease in PtdOH was observed (Fig. 2H, shown asdiagonal stripes and black bar segments, respectively).The increase in PtdOH was accompanied by an in-creased proportion of associated 18:2�9,12 and 18:3�9,12,15 (Fig. 2H). Changes in PtdOH levels and fattyacid composition occurred, thus, later than those ob-served for phosphoinositides in the identical samples.
Stress-induced, unsaturated polyphosphoinositidessupport InsP3-production
While temporally correlated, the increases in PtdIns4Pand PtdIns(4,5)P2 do not balance the decrease inPtdIns (compare scales of Figs. 2D–F). A possible expla-nation for the mass imbalance is that pathway interme-diates are not statically accumulating but rather aresubject to continuous flux toward other downstreammetabolites of the phosphoinositide pathway. Becauseexpression of Arabidopsis PLC1 is induced by salt (52)and InsP3 release with osmotic stress has been de-scribed for various plant systems (17, 33, 53, 54), weasked whether some of the unsaturated PtdIns(4,5)P2formed with osmotic stress would turn toward hydroly-sis by PLC, yielding InsP3 and DAG. In plants, DAG isimmediately phosphorylated to PtdOH by the enzymeDAG kinase (55). Formation of the PLC products InsP3,DAG, and PtdOH is shown in Fig. 3A–C. While DAGlevels did not change significantly (Fig. 3B), PtdOHlevels increased temporally correlated to the produc-tion of InsP3 after osmotic stress (Fig. 3C). Importantly,the raised PtdOH levels were due to an increasedproportion of unsaturated PtdOH containing 18:2�9,12
and 18:3�9,12,15 (shown as diagonal stripes and blackbar segments, respectively), consistent with an origin inthe hydrolysis of unsaturated PtdIns(4,5)P2 transientlyformed with osmotic stress.
Forced InsP3 breakdown increases PLC hydrolysis ofunsaturated polyphosphoinositides in stressed plants
Because PtdOH could also stem from other sources,and to gain more than merely correlative evidence, weasked whether selective perturbation of the phospho-
TABLE 1. Degree of unsaturation of fatty acids in phosphoinositides and derived lipids during hyperosmotic stress.
Time of hyperosmotic stimulation (min)
Phospholipid 0 15 30 60
PtdIns 1.63 � 0.3 1.08 � 0.3* 0.68 � 0.2** 1.62 � 0.2PtdIns4P 0.92 � 0.3 1.02 � 0.3 1.50 � 0.3* 1.77 � 0.2**PtdIns(4,5)P2 0.64 � 0.2 0.76 � 0.2 1.07 � 0.3* 1.14 � 0.3**DAG 1.97 � 0.3 1.93 � 0.4 1.76 � 0.3 1.17 � 0.3**PtdOH 1.25 � 0.3 1.69 � 0.3 1.76 � 0.4 2.02 � 0.4**
Data given in Figs. 2 and 3 were analyzed by calculating the mole double bonds per mole fatty acid associated with the individual lipids.Numbers are the mean ratios � sd The asterisk represents a significant difference in fatty acid unsaturation after stimulation from that innon-stressed samples on a Student’s t test (*P�0.05; **P�0.01).
TABLE 2. Proportion of polyunsaturated fatty acids (18:2�9,12
and 18:3�9,12,15) of the total fatty acids associated withphospholipids during hyperosmotic stress.
Time of hyperosmotic stimulation (min)
Phospholipid 0 15 30 60
PtdIns 34.6 40.5 20.1** 41.3PtdIns4P 25.7 29.6 51.0** 66.1**PtdIns(4,5)P2 16.8 16.8 29.6** 34.2**DAG 47.2 65.6* 58.1 47.3PtdOH 43.2 62.2* 65.9* 76.8**
Data from Figs. 2 and 3 were analyzed by calculating theappropriate ratios. Data are given as mol% of total fatty acids. Theerror is approximately 15%. The asterisk represents a significantdifference in lipid PUFA content after stimulation from that innon-stressed samples on a Student’s t test (*P�0.05; **P�0.01).
1962 Vol. 21 July 2007 KONIG ET AL.The FASEB Journal
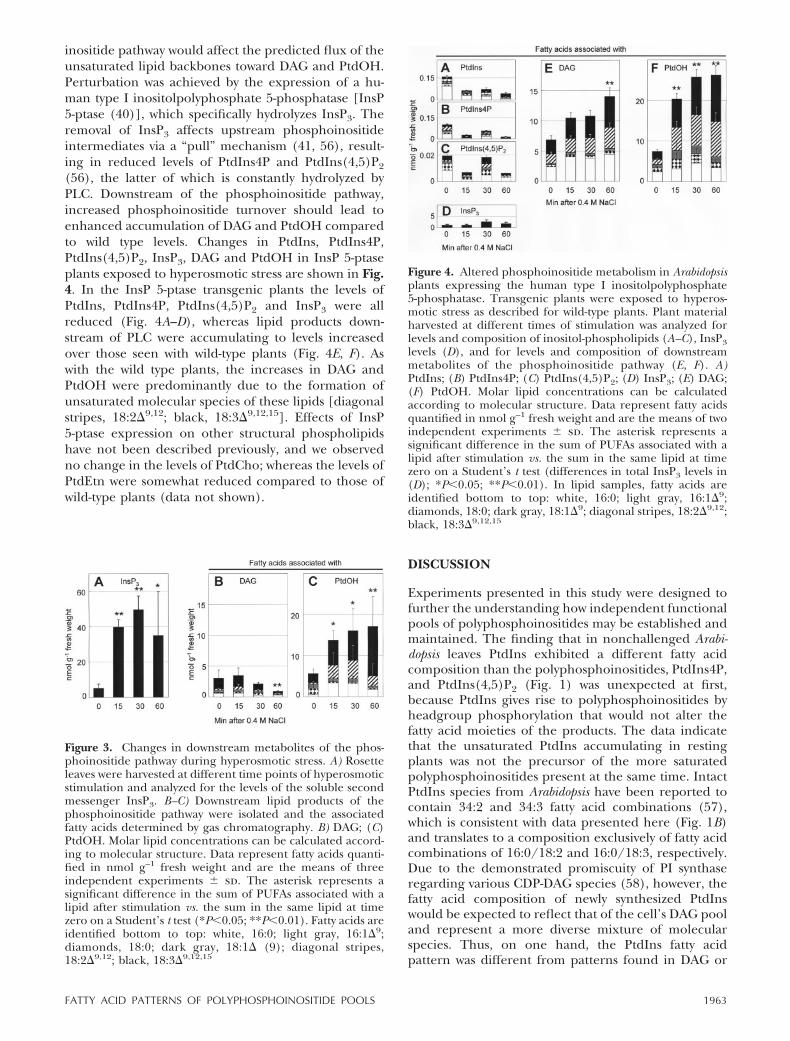
inositide pathway would affect the predicted flux of theunsaturated lipid backbones toward DAG and PtdOH.Perturbation was achieved by the expression of a hu-man type I inositolpolyphosphate 5-phosphatase [InsP5-ptase (40)], which specifically hydrolyzes InsP3. Theremoval of InsP3 affects upstream phosphoinositideintermediates via a “pull” mechanism (41, 56), result-ing in reduced levels of PtdIns4P and PtdIns(4,5)P2
(56), the latter of which is constantly hydrolyzed byPLC. Downstream of the phosphoinositide pathway,increased phosphoinositide turnover should lead toenhanced accumulation of DAG and PtdOH comparedto wild type levels. Changes in PtdIns, PtdIns4P,PtdIns(4,5)P2, InsP3, DAG and PtdOH in InsP 5-ptaseplants exposed to hyperosmotic stress are shown in Fig.4. In the InsP 5-ptase transgenic plants the levels ofPtdIns, PtdIns4P, PtdIns(4,5)P2 and InsP3 were allreduced (Fig. 4A–D), whereas lipid products down-stream of PLC were accumulating to levels increasedover those seen with wild-type plants (Fig. 4E, F). Aswith the wild type plants, the increases in DAG andPtdOH were predominantly due to the formation ofunsaturated molecular species of these lipids [diagonalstripes, 18:2�9,12; black, 18:3�9,12,15]. Effects of InsP5-ptase expression on other structural phospholipidshave not been described previously, and we observedno change in the levels of PtdCho; whereas the levels ofPtdEtn were somewhat reduced compared to those ofwild-type plants (data not shown).
DISCUSSION
Experiments presented in this study were designed tofurther the understanding how independent functionalpools of polyphosphoinositides may be established andmaintained. The finding that in nonchallenged Arabi-dopsis leaves PtdIns exhibited a different fatty acidcomposition than the polyphosphoinositides, PtdIns4P,and PtdIns(4,5)P2 (Fig. 1) was unexpected at first,because PtdIns gives rise to polyphosphoinositides byheadgroup phosphorylation that would not alter thefatty acid moieties of the products. The data indicatethat the unsaturated PtdIns accumulating in restingplants was not the precursor of the more saturatedpolyphosphoinositides present at the same time. IntactPtdIns species from Arabidopsis have been reported tocontain 34:2 and 34:3 fatty acid combinations (57),which is consistent with data presented here (Fig. 1B)and translates to a composition exclusively of fatty acidcombinations of 16:0/18:2 and 16:0/18:3, respectively.Due to the demonstrated promiscuity of PI synthaseregarding various CDP-DAG species (58), however, thefatty acid composition of newly synthesized PtdInswould be expected to reflect that of the cell’s DAG pooland represent a more diverse mixture of molecularspecies. Thus, on one hand, the PtdIns fatty acidpattern was different from patterns found in DAG or
Figure 3. Changes in downstream metabolites of the phos-phoinositide pathway during hyperosmotic stress. A) Rosetteleaves were harvested at different time points of hyperosmoticstimulation and analyzed for the levels of the soluble secondmessenger InsP3. B–C) Downstream lipid products of thephosphoinositide pathway were isolated and the associatedfatty acids determined by gas chromatography. B) DAG; (C)PtdOH. Molar lipid concentrations can be calculated accord-ing to molecular structure. Data represent fatty acids quanti-fied in nmol g–1 fresh weight and are the means of threeindependent experiments � sd. The asterisk represents asignificant difference in the sum of PUFAs associated with alipid after stimulation vs. the sum in the same lipid at timezero on a Student’s t test (*P�0.05; **P�0.01). Fatty acids areidentified bottom to top: white, 16:0; light gray, 16:1�9;diamonds, 18:0; dark gray, 18:1� (9); diagonal stripes,18:2�9,12; black, 18:3�9,12,15
Figure 4. Altered phosphoinositide metabolism in Arabidopsisplants expressing the human type I inositolpolyphosphate5-phosphatase. Transgenic plants were exposed to hyperos-motic stress as described for wild-type plants. Plant materialharvested at different times of stimulation was analyzed forlevels and composition of inositol-phospholipids (A–C), InsP3levels (D), and for levels and composition of downstreammetabolites of the phosphoinositide pathway (E, F). A)PtdIns; (B) PtdIns4P; (C) PtdIns(4,5)P2; (D) InsP3; (E) DAG;(F) PtdOH. Molar lipid concentrations can be calculatedaccording to molecular structure. Data represent fatty acidsquantified in nmol g–1 fresh weight and are the means of twoindependent experiments � sd. The asterisk represents asignificant difference in the sum of PUFAs associated with alipid after stimulation vs. the sum in the same lipid at timezero on a Student’s t test (differences in total InsP3 levels in(D); *P�0.05; **P�0.01). In lipid samples, fatty acids areidentified bottom to top: white, 16:0; light gray, 16:1�9;diamonds, 18:0; dark gray, 18:1�9; diagonal stripes, 18:2�9,12;black, 18:3�9,12,15
1963FATTY ACID PATTERNS OF POLYPHOSPHOINOSITIDE POOLS

other structural phospholipids, such as PtdCho, where18:2/18:3 and 18:3/18:3 combinations occur (57). Onthe other hand, the PtdIns pattern also differed sub-stantially from the patterns found in PtdIns4P andPtdIns(4,5)P2 (Fig. 1C, D), which carried more satu-rated fatty acids. An equivalent pattern has been ob-served in nonchallenged mouse RAW 264.7 cells, wherevery long chain polyunsaturated fatty acids were foundin PtdIns, but not in derived polyphosphoinositides(38). A possible explanation for the observed patternsis that a diverse mixture of PtdIns molecular species isdrawn on to generate two different pools of polyphos-phoinositides, according to fatty acid composition: Un-der nonchallenging conditions saturated and monoun-saturated PtdIns species are used to produce thecorresponding species of polyphosphoinositides withconstitutive regulatory functions. In this scenario, apool of highly unsaturated PtdIns may accumulate,which is primed to respond to stress and represents thePtdIns molecular species observed in nonchallengedcells. Between 15 and 30 min of hyperosmotic stress,the primed, highly unsaturated PtdIns is turned over(Fig. 2), leading to a transient decrease in PtdIns levelsand to concomitant formation of highly unsaturatedpolyphosphoinositides, as is shown in this study (Fig. 2;Tables 1, 2). The complex, and at first unintuitive, fattyacid patterns observed may, thus, be explained by theoutlined concept of PtdIns partitioning into constitu-tive and stress-inducible lipid pools. Considering thatPtdIns is formed in the ER (24, 25), it is possible thatPtdIns species with certain fatty acid compositions areretained in the ER, whereas others are delivered tovarious subcellular locations, including the plasmamembrane, the Golgi, or the nucleus, where stress-inducible or developmentally controlled PI kinases andPIP kinases may produce polyphosphoinositides withdifferent regulatory effects. After 60 min of hyperos-motic stress, PtdIns has recovered to starting levels (Fig.2D), suggesting increased resynthesis of this lipid oraction of polyphosphoinositide phosphatases. As con-comitant with the PtdIns increase, at 60 min PtdIns4Paccumulates (Fig. 2E) and only PtdIns(4,5)P2 decreases(Fig. 2F), phosphatase action appears unlikely. Thepattern suggests inactivation of a stress-induced PIPkinase, perhaps via a feedback loop involving termina-tion of the InsP3 signal, and consequential accumula-tion of the precursor lipids, PtdIns and PtdIns4P.
Hyperosmotic stress results in global changes in lipidunsaturation in eukaryotic cells, and increases in lipidunsaturation are required to maintain membrane in-tegrity (59). The elevated degree of unsaturation ob-served for PtdCho after 60 min of hyperosmotic stress(Fig. 2H) may be part of such a survival mechanism.Note that prior to the changes in PtdCho, MGDGunsaturation in the plastid decreases (Fig. 2G), whichsuggests recycling of PUFAs from plastidial MGDG tosupport unsaturation of PtdCho in the plasma mem-brane. Changes in phosphoinositide unsaturation withhyperosmotic stress occur earlier than those of other,
structural lipids, and, therefore, may not be a result ofchanges in global lipid unsaturation.
The transient decrease in PtdIns with stress treat-ment observed in this study is contrasted by a previousreport of no PtdIns decrease with stress (57). Possibleexplanations include the transient nature of thechanges or differences in the stresses applied or theexperimental setups. Slight differences in fatty acidcomposition of PtdIns from nonchallenged plantsgrown under different conditions (compare Fig. 1Bwith Fig. 2D) are likely the result of growth on soil vs.hydroponic culture, which poses a stress to the plants byitself and may cause some turnover of unsaturated PtdIns.
To test whether increased polyphosphoinositide un-saturation (Fig. 2, Tables 1, 2) during the initial phaseof adaptation to hyperosmotic stress was a consequenceof fatty acid desaturase action, plant material washarvested over the critical period of osmotic stimula-tion, total lipid extracts were prepared, and the propor-tions of the associated fatty acids determined. Theamounts of 18:2�9,12 and 18:3�9,12,15 transmethylatedfrom the total lipid extracts did not change substan-tially over 60 min of application of the stress (data notshown), indicating that no net increase in unsaturatedfatty acids had occurred. Because the contribution ofphosphoinositides to the total phospholipids is onlysmall, specific phosphoinositide desaturation wouldnot be apparent and cannot be ruled out. However, todate no fatty acid desaturases have been reported tohave exclusive preference for fatty acids esterified topolyphosphoinositides. The fatty acid composition oftotal lipid extracts did not substantially change, suggest-ing fatty acid desaturases were not responsible for theincreased degree of polyphosphoinositide unsaturationafter osmotic stimulation.
Previous reports of increased phosphorylation ofPtdIns and of PtdIns4P with hyperosmotic stress (33,34), as well as the data presented here, suggest that theincrease in polyphosphoinositides likely originated instress-induced activation of PI kinases and PIP kinases,which generate PtdIns4P and PtdIns(4,5)P2, respec-tively, from primed, unsaturated PtdIns. Preferences ofrecombinant enzymes toward phosphoinositide sub-strates with defined fatty acid compositions have beendemonstrated in vitro for insect phosphoinositide ki-nases (22) and for human phosphoinositide phospha-tases (23), suggesting that the fatty acid composition ofthe phosphoinositide substrate is a modulator of en-zymes regulating cellular phosphoinositide production(22) and/or distribution. With respect to physiology,PtdIns pools with different fatty acid compositions maybe phosphorylated in response to different endogenousor exogenous cues by different sets of lipid kinasesacting on their respective preferred saturated or unsat-urated phosphoinositide substrates.
The transient nature of the increases in unsaturatedpolyphosphoinositides raises the question as to whatphysiological role the transient phosphoinositide in-creases may have in stress adaptation. Here we aimed todecide whether transiently formed PtdIns(4,5)P2 would
1964 Vol. 21 July 2007 KONIG ET AL.The FASEB Journal

remain intact, serving as a ligand to potential proteinpartners, or whether the lipid would turn toward hy-drolysis by PLC and InsP3 production. Our results withwild-type Arabidopsis and the transgenic plants express-ing the human InsP 5-ptase indicate the latter scenario(Figs. 3, 4). Arguments in support of this interpretationinclude: (A) the increase in PtdIns(4,5)P2 was tempo-rally correlated with a concomitant increase in InsP3;(B) the downstream lipid products of PLC action, DAGand PtdOH, accumulated in parallel with InsP3 produc-tion; (C) the increases in DAG and PtdOH were largelydue to increased formation of highly unsaturated mo-lecular species of these lipids, consistent with theirorigin in the unsaturated PtdIns(4,5)P2 increasing withosmotic stress; and (D) the effects of osmotic stimula-tion on DAG and PtdOH formation and compositionwere increased in the transgenic Arabidopsis plantsexpressing the human InsP 5-ptase, for which increasedphosphoinositide turnover by PLC has been previouslyreported (41, 56). While PtdOH can be formed from allglycerophospholipids by action of phospholipase D(60), the patterns observed here indicate that PtdOHformed with hyperosmotic stress in Arabidopsis is likely aconsequence of combined action of PLC and DAGkinase.
The apparent channeling of unsaturated phosphoi-nositides through a pathway for InsP3 production leadsus to speculate how the unsaturated lipids could becompartmentalized. Despite a generally smaller pro-portion in glycerophospholipids in sphingolipid- andsterol-enriched lipid microdomains (“lipid rafts”) (61),phosphoinositides are thought to be present in raft-likemicrodomains in mammalian membranes (62, 63).Unsaturated acyl groups associated with a phosphoino-sitide will compensate the shape of a large, chargedheadgroup and result in a more cylindrical, bilayer-forming lipid, whereas saturated acyl chains will createan inversely cone-shaped lipid (64, 65) that generatesmembrane curvature strain (64–66). Such differencesin hydrophobic tails alone have been shown in animalcells to be sufficient for differential lateral distributionand accumulation of lipids in membrane microdo-mains without action of accessory proteins (67). Thedata presented here indicate concerted, stress-induc-ible turnover of unsaturated phosphoinositidesthrough a pathway defined by PI kinase, PIP kinase,PLC, and DAG kinase. Tobacco homologs of all en-zymes of this postulated pathway have been found to beenriched in isolated detergent-insoluble membranepreparations (“lipid-rafts”) from tobacco cell plasmamembranes resembling microdomains (F. Furt and S.Mongrand, personal communication). Differences inphosphoinositide fatty acid composition, such as thosedescribed in this study, may restrict certain phospho-inositide species to raft-like microdomains or precludeothers from forming microdomains in Arabidopsis. Itremains to be seen whether on hyperosmotic stress therespective enzymes may also be found in the detergent-soluble fraction of the plasma membrane, or whetherthe pathway is restricted to rafts.
In summary, an attempt was made to disentangle thecomplex regulatory network of polyphosphoinositidesby comparing distinct phosphoinositide pools that areconstitutively present in Arabidopsis leaves with thosetransiently formed during conditions of stress. Consti-tutive and inducible phosphoinositide pools consist oflipids with distinct fatty acid compositions, and likelydiffer in their biophysical properties. Tracing of theunsaturated lipid backbones of intermediates derivedfrom stress-induced phosphoinositides indicatesPLC-mediated hydrolysis of this signaling pool, lead-ing to production of the second messenger, InsP3.Future refinements in mass-spectrometric analysismay provide tools to monitor some phosphoinositidepools in complex metabolite mixtures based ondifferent molecular composition in order to eluci-date physiological functions of individual pools notonly for PtdIns(4,5)P2, but also for the multitude ofother known polyphosphoinositides.
The transgenic Arabidopsis plants expressing the humantype I inositolpolyphosphate 5-phosphatase were kindly pro-vided by Dr. Imara Perera (North Carolina State University,Raleigh, NC, USA). We thank Dr. Ivo Feussner (Georg-August-University Gottingen, Germany) and Dr. Imara Pereraand Dr. Wendy Boss (North Carolina State University, Ra-leigh, NC, USA) for their helpful discussion. We are gratefulfor generous funding through an Emmy Noether Fellowshipfrom the German Research Foundation (D. F. G., to I.H.).
REFERENCES
1. Santarius, M., Lee, C. H., and Anderson, R. A. (2006) Super-vised membrane swimming: small G-protein lifeguards regulatePIPK signalling and monitor intracellular PtdIns(4,5)P2 pools.Biochem. J. 398, 1–13
2. Balla, T. (2006) Phosphoinositide-derived messengers in endo-crine signaling. J. Endocrinol. 188, 135–153
3. Meijer, H. J., and Munnik, T. (2003) Phospholipid-based signal-ing in plants. Annu. Rev. Plant Biol. 54, 265–306
4. Mueller-Roeber, B., and Pical, C. (2002) Inositol phospholipidmetabolism in Arabidopsis. Characterized and putative isoformsof inositol phospholipid kinase and phosphoinositide-specificphospholipase C. Plant Physiol. 130, 22–46
5. Stevenson, J. M., Perera, I. Y., Heilmann, I., Persson, S., andBoss, W. F. (2000) Inositol signaling and plant growth. TrendsPlant Sci. 5, 252–258
6. Suh, B. C., and Hille, B. (2005) Regulation of ion channels byphosphatidylinositol 4,5-bisphosphate. Curr. Opin. Neurobiol. 15,370–378
7. Cote, G. G., Yueh, Y. G., and Crain, R. C. (1996) Phosphoino-sitide turnover and its role in plant signal transduction. Subcell.Biochem. 26, 317–343
8. Wenk, M. R., and De Camilli, P. (2004) Protein-lipid interac-tions and phosphoinositide metabolism in membrane traffic:insights from vesicle recycling in nerve terminals. Proc. Natl.Acad. Sci. U. S. A. 101, 8262–8269
9. Wasteneys, G. O., and Galway, M. E. (2003) Remodeling thecytoskeleton for growth and form: an overview with some newviews. Annu. Rev. Plant Biol. 54, 691–722
10. Doughman, R. L., Firestone, A. J., and Anderson, R. A. (2003)Phosphatidylinositol phosphate kinases put PI4,5P(2) in itsplace. J. Membr. Biol. 194, 77–89
11. Lemmon, M. A., Ferguson, K. M., and Abrams, C. S. (2002)Pleckstrin homology domains and the cytoskeleton. FEBS Lett.513, 71–76
12. Roth, M. G. (2004) Phosphoinositides in constitutive membranetraffic. Physiol Rev. 84, 699–730
1965FATTY ACID PATTERNS OF POLYPHOSPHOINOSITIDE POOLS

13. Balla, T. (2005) Inositol-lipid binding motifs: signal integratorsthrough protein-lipid and protein-protein interactions. J. CellSci. 118, 2093–2104
14. Lemmon, M. A. (2003) Phosphoinositide recognition domains.Traffic 4, 201–213
15. Drobak, B. K., Watkins, P. A. C., Valenta, R., Dove, S. K., Lloyd,C. W., and Staiger, C. J. (1994) Inhibition of plant plasmamembrane phosphoinositide phospholipase C by the actin-binding protein, profilin. Plant J. 6, 389–400
16. Berridge, M. J. (1983) Rapid accumulation of inositol trisphos-phate reveals that agonists hydrolyse polyphosphoinositidesinstead of phosphatidylinositol. Biochem. J. 212, 849–858
17. Heilmann, I., Perera, I. Y., Gross, W., and Boss, W. F. (2001)Plasma membrane phosphatidylinositol 4,5-bisphosphate levelsdecrease with time in culture. Plant Physiol. 126, 1507–1518
18. Kost, B., Lemichez, E., Spielhofer, P., Hong, Y., Tolias, K.,Carpenter, C., and Chua, N. H. (1999) Rac homologues andcompartmentalized phosphatidylinositol 4, 5-bisphosphate actin a common pathway to regulate polar pollen tube growth.J. Cell Biol. 145, 317–330
19. Liu, Y., Casey, L., and Pike, L. J. (1998) Compartmentalizationof phosphatidylinositol 4,5-bisphosphate in low-density mem-brane domains in the absence of caveolin. Biochem. Biophys. Res.Commun. 245, 684–690
20. King, C. E., Stephens, L. R., Hawkins, P. T., Guy, G. R., andMichell, R. H. (1987) Multiple metabolic pools of phosphoi-nositides and phosphatidate in human erythrocytes incubatedin a medium that permits rapid transmembrane exchange ofphosphate. Biochem. J. 244, 209–217
21. Lemmon, M. A., and Ferguson, K. M. (2000) Signal-dependentmembrane targeting by pleckstrin homology (PH) domains.Biochem. J. 350 Pt. 1, 1–18
22. Carricaburu, V., and Fournier, B. (2001) Phosphoinositide fattyacids regulate phosphatidylinositol 5-kinase, phospholipase Cand protein kinase C activities. Eur. J. Biochem. 268, 1238–1249
23. Schmid, A. C., Wise, H. M., Mitchell, C. A., Nussbaum, R., andWoscholski, R. (2004) Type II phosphoinositide 5-phosphataseshave unique sensitivities towards fatty acid composition andhead group phosphorylation. FEBS Lett. 576, 9–13
24. Collin, S., Justin, A. M., Cantrel, C., Arondel, V., and Kader, J. C.(1999) Identification of AtPIS, a phosphatidylinositol synthasefrom Arabidopsis. Eur. J. Biochem. 262, 652–658
25. Justin, A. M., Kader, J. C., and Collin, S. (2002) Phosphatidyl-inositol synthesis and exchange of the inositol head are catal-ysed by the single phosphatidylinositol synthase 1 from Arabi-dopsis. Eur. J. Biochem. 269, 2347–2352
26. Phillips, S. E., Vincent, P., Rizzieri, K. E., Schaaf, G., Bankaitis,V. A., and Gaucher, E. A. (2006) The diverse biological func-tions of phosphatidylinositol transfer proteins in eukaryotes.Crit. Rev. Biochem. Mol. Biol. 41, 21–49
27. Vincent, P., Chua, M., Nogue, F., Fairbrother, A., Mekeel, H.,Xu, Y., Allen, N., Bibikova, T. N., Gilroy, S., and Bankaitis, V. A.(2005) A Sec14p-nodulin domain phosphatidylinositol transferprotein polarizes membrane growth of Arabidopsis thaliana roothairs. J. Cell Biol. 168, 801–812
28. Bunney, T. D., Watkins, P. A., Beven, A. F., Shaw, P. J.,Hernandez, L. E., Lomonossoff, G. P., Shanks, M., Peart, J., andDrobak, B. K. (2000) Association of phosphatidylinositol 3-ki-nase with nuclear transcription sites in higher plants. Plant Cell12, 1679–1688
29. Perera, I. Y., Heilmann, I., and Boss, W. F. (1999) Transient andsustained increases in inositol 1,4,5-trisphosphate precede thedifferential growth response in gravistimulated maize pulvini.Proc. Natl. Acad. Sci. U. S. A. 96, 5838–5843
30. Cho, M. H., Shears, S. B., and Boss, W. F. (1993) Changes inphosphatidylinositol metabolism in response to hyperosmoticstress in Daucus carota L. cells grown in suspension culture. PlantPhysiol. 103, 637–647
31. Roy, A., and Levine, T. P. (2004) Multiple pools of phosphati-dylinositol 4-phosphate detected using the pleckstrin homologydomain of Osh2p. J. Biol. Chem. 279, 44683–44689
32. Balla, T., and Varnai, P. (2002) Visualizing cellular phosphoino-sitide pools with GFP-fused protein-modules. Sci. STKE 2002,PL3
33. DeWald, D. B., Torabinejad, J., Jones, C. A., Shope, J. C.,Cangelosi, A. R., Thompson, J. E., Prestwich, G. D., and Hama,H. (2001) Rapid accumulation of phosphatidylinositol 4,5-
bisphosphate and inositol 1,4,5-trisphosphate correlates withcalcium mobilization in salt-stressed Arabidopsis. Plant Physiol.126, 759–769
34. Pical, C., Westergren, T., Dove, S. K., Larsson, C., and Somma-rin, M. (1999) Salinity and hyperosmotic stress induce rapidincreases in phosphatidylinositol 4,5-bisphosphate, diacylglyc-erol pyrophosphate, and phosphatidylcholine in Arabidopsisthaliana cells. J. Biol. Chem. 274, 38232–38240
35. Munnik, T., Meijer, H. J., Ter Riet, B., Hirt, H., Frank, W.,Bartels, D., and Musgrave, A. (2000) Hyperosmotic stress stim-ulates phospholipase D activity and elevates the levels of phos-phatidic acid and diacylglycerol pyrophosphate. Plant J. 22,147–154
36. Pettitt, T. R., Dove, S. K., Lubben, A., Calaminus, S. D., andWakelam, M. J. (2006) Analysis of intact phosphoinositides inbiological samples. J. Lipid Res. 47, 1588–1596
37. Guan, X. L., and Wenk, M. R. (2006) Mass spectrometry-basedprofiling of phospholipids and sphingolipids in extracts fromSaccharomyces cerevisiae. Yeast 23, 465–477
38. Milne, S. B., Ivanova, P. T., DeCamp, D., Hsueh, R. C., andBrown, H. A. (2005) A targeted mass spectrometric analysis ofphosphatidylinositol phosphate species. J. Lipid Res. 46, 1796–1802
39. Welti, R., and Wang, X. (2004) Lipid species profiling: ahigh-throughput approach to identify lipid compositionalchanges and determine the function of genes involved in lipidmetabolism and signaling. Curr. Opin. Plant Biol. 7, 337–344
40. Laxminarayan, K. M., Chan, B. K., Tetaz, T., Bird, P. I., andMitchell, C. A. (1994) Characterization of a cDNA encoding the43-kDa membrane-associated inositol-polyphosphate 5-phos-phatase. J. Biol. Chem. 269, 17305–17310
41. Perera, I. Y., Hung, C. Y., Brady, S., Muday, G. K., and Boss, W. F.(2006) A universal role for inositol 1,4,5-trisphosphate-mediatedsignaling in plant gravitropism. Plant Physiol. 140, 746–760
42. Randall, P. J., and Bouma, D. (1973) Zinc deficiency, carbonicanhydrase, and photosynthesis in leaves of spinach. Plant Physiol.52, 229–232
43. Cho, M. H., Chen, Q., Okpodu, C. M., and Boss, W. F. (1992)Separation and quantification of [3H]inositol phospholipidsusing thin-layer-chromatography and a computerized 3H imag-ing scanner. LC-GC. 10, 464–468
44. Perera, I. Y., Davis, A. J., Galanopoulou, D., Im, Y. J., and Boss,W. F. (2005) Characterization and comparative analysis ofArabidopsis phosphatidylinositol phosphate 5-kinase 10 revealsdifferences in Arabidopsis and human phosphatidylinositolphosphate kinases. FEBS Lett. 579, 3427–3432
45. Hartel, H., Dormann, P., and Benning, C. (2000) DGD1-independent biosynthesis of extraplastidic galactolipids afterphosphate deprivation in Arabidopsis. Proc. Natl. Acad. SciU. S. A. 97, 10649–10654
46. Christie, W. W. (2003) Thin-layer chromatography. In LipidAnalysis, Vol. 15 pp. 142–152, The Oily Press, Bridgwater, UK
47. Hornung, E., Pernstich, C., and Feussner, I. (2002) Formationof conjugated Delta11Delta13-double bonds by Delta12-linoleicacid (1,4)-acyl-lipid-desaturase in pomegranate seeds. Eur. J. Bio-chem. 269, 4852–4859
48. Dove, S. K., Cooke, F. T., Douglas, M. R., Sayers, L. G., Parker,P. J., and Michell, R. H. (1997) Osmotic stress activates phos-phatidylinositol-3,5-bisphosphate synthesis. Nature 390, 187–192
49. Bonangelino, C. J., Nau, J. J., Duex, J. E., Brinkman, M.,Wurmser, A. E., Gary, J. D., Emr, S. D., and Weisman, L. S.(2002) Osmotic stress-induced increase of phosphatidylinositol3,5-bisphosphate requires Vac14p, an activator of the lipidkinase Fab1p. J. Cell Biol. 156, 1015–1028
50. Meijer, H. J., Divecha, N., van den Ende, H., Musgrave, A., andMunnik, T. (1999) Hyperosmotic stress induces rapid synthesisof phosphatidyl-D-inositol 3,5-bisphosphate in plant cells. Planta208, 294–298
51. Chilvers, E. R., Batty, I. H., Challiss, R. A., Barnes, P. J., andNahorski, S. R. (1991) Determination of mass changes inphosphatidylinositol 4,5-bisphosphate and evidence for agonist-stimulated metabolism of inositol 1,4,5-trisphosphate in airwaysmooth muscle. Biochem. J. 275 (Pt 2), 373–379
52. Hirayama, T., Ohto, C., Mizoguchi, T., and Shinozaki, K. (1995)A gene encoding a phosphatidylinositol-specific phospholipaseC is induced by dehydration and salt stress in Arabidopsisthaliana. Proc. Natl. Acad. Sci. U. S. A. 92, 3903–3907
1966 Vol. 21 July 2007 KONIG ET AL.The FASEB Journal

53. Drobak, B. K., and Watkins, P. A. (2000) Inositol(1,4,5)-trisphosphate production in plant cells: an early response tosalinity and hyperosmotic stress. FEBS Lett. 481, 240–244
54. Heilmann, I., Perera, I. Y., Gross, W., and Boss, W. F. (1999)Changes in phosphoinositide metabolism with days in cultureaffect signal transduction pathways in Galdieria sulphuraria. PlantPhysiol. 119, 1331–1339
55. Munnik, T. (2001) Phosphatidic acid: an emerging plant lipidsecond messenger. Trends Plant Sci. 6, 227–233
56. Perera, I. Y., Love, J., Heilmann, I., Thompson, W. F., and Boss,W. F. (2002) Up-regulation of phosphoinositide metabolism intobacco cells constitutively expressing the human type I inositolpolyphosphate 5-phosphatase. Plant Physiol. 129, 1795–1806
57. Welti, R., Li, W., Li, M., Sang, Y., Biesiada, H., Zhou, H. E.,Rajashekar, C. B., Williams, T. D., and Wang, X. (2002) Profilingmembrane lipids in plant stress responses. Role of phospho-lipase D alpha in freezing-induced lipid changes in Arabidopsis.J. Biol. Chem. 277, 31994–32002
58. Justin, A. M., Kader, J. C., and Collin, S. (2003) Syntheticcapacity of Arabidopsis phosphatidylinositol synthase 1 ex-pressed in Escherichia coli. Biochim. Biophys. Acta 1634, 52–60
59. Kinnunen, P. K. (2000) Lipid bilayers as osmotic responseelements. Cell Physiol. Biochem. 10, 243–250
60. Wang, X. (2002) Phospholipase D in hormonal and stresssignaling. Curr. Opin. Plant Biol. 5, 408–414
61. Laloi, M., Perret, A. M., Chatre, L., Melser, S., Cantrel, C.,Vaultier, M. N., Zachowski, A., Bathany, K., Schmitter, J. M.,Vallet, M., Lessire, R., Hartmann, M. A., and Moreau, P. (2006)Insights into the role of specific lipids in the formation anddelivery of lipid microdomains to the plasma membrane ofplant cells. Plant Physiol. 143, 461–472
62. Shibasaki, Y., Ishihara, H., Kizuki, N., Asano, T., Oka, Y., andYazaki, Y. (1997) Massive actin polymerization induced byphosphatidylinositol-4-phosphate 5-kinase in vivo. J. Biol. Chem.272, 7578–7581
63. Raucher, D., Stauffer, T., Chen, W., Shen, K., Guo, S., York,J. D., Sheetz, M. P., and Meyer, T. (2000) Phosphatidylinositol4,5-bisphosphate functions as a second messenger that regulatescytoskeleton-plasma membrane adhesion. Cell 100, 221–228
64. Cooke, I. R., and Deserno, M. (2006) Coupling between lipidshape and membrane curvature. Biophys. J. 91, 487–495
65. van Meer, G. (2005) Cellular lipidomics. EMBO J. 24, 3159–316566. Janmey, P. A., and Kinnunen, P. K. (2006) Biophysical properties
of lipids and dynamic membranes. Trends Cell Biol. 16, 538–54667. Mukherjee, S., Soe, T. T., and Maxfield, F. R. (1999) Endocytic
sorting of lipid analogues differing solely in the chemistry oftheir hydrophobic tails. J. Cell Biol. 144, 1271–1284
Received for publication December 15, 2006.Accepted for publication January 25, 2007.
1967FATTY ACID PATTERNS OF POLYPHOSPHOINOSITIDE POOLS
Related Documents











