Brain Research 963 (2003) 203–213 www.elsevier.com / locate / brainres Research report Stereotaxic localization of corticosterone to the amygdala enhances hypothalamo-pituitary–adrenal responses to behavioral stress 1 * Jack D. Shepard , Kirk W. Barron, Dean A. Myers Department of Physiology, College of Medicine, University of Oklahoma Health Sciences Center, 800 N. Research Parkway, Oklahoma City, OK 73104, USA Accepted 29 October 2002 Abstract The amygdala is involved in behavioral, autonomic, and neuroendocrine responses to stressful stimuli. The goal of the current study was to determine the effect of directly elevating glucocorticoids in the amygdala on hypothalamo-pituitary–adrenocortical (HPA) responses to the elevated plus maze, a behavioral stressor known to activate the amygdala. Micropellets (30 mg) of crystalline corticosterone or cholesterol (control) were implanted bilaterally at the dorsal margin of the CeA in male Wistar rats; vascular catheters were also placed at this time. Five days post-surgery, blood samples were drawn at 07:00 and 19:00 h to assess diurnal rhythm of plasma corticosterone. At 7 days post-implantation, rats were subjected to behavioral stress using an elevated plus maze and blood was collected 15 min prior to stress, and at 15, 45, and 90 min after the initiation of the stressor. Corticotropin releasing factor (CRF) and arginine vasopressin (AVP) mRNA levels were analyzed by in situ hybridization in the medial parvocellular division of the hypothalamic paraventricular nucleus (mpPVN) in corticosterone- and cholesterol-implanted rats either not exposed to the elevated plus maze (control) or 4 h post-behavioral stress. Localization of corticosterone to the amygdala had no effect on diurnal rhythm of corticosterone secretion. Behavioral stress significantly increased peak plasma corticosterone levels in both groups to a similar level. However, in the corticosterone implanted rats, plasma corticosterone concentrations at 45 and 90 min post-stress were significantly greater compared to control rats indicating a prolonged corticosterone response to behavioral stress. In non-stressed rats, corticosterone delivery to the amygdala elevated basal CRF mRNA in the mpPVN to levels similar to those observed post-stress in control animals; no further increase was observed in CRF mRNA following stress. Behavioral stress resulted in a significant elevation in CRF mRNA in cholesterol controls. Basal AVP mRNA levels were unaffected by corticosterone implants. AVP mRNA did not increase in cholesterol implanted rats in response to behavioral stress. However, AVP mRNA levels were higher in corticosterone implanted rats post stress compared to cholesterol treated controls. In conclusion, direct administration of corticosterone to the amygdala increases plasma corticosterone in response to a behavioral stressor without altering the diurnal rhythm in plasma corticosterone. Elevated basal levels of mpPVN CRF mRNA, and the induction of a mpPVN AVP mRNA response to the behavioral stressor implicate enhanced ACTH secretagogue expression in the increased HPA response to corticosterone modulation of amygdala function. 2002 Elsevier Science B.V. All rights reserved. Theme: Endocrine and autonomic regulation Topic: Hypothalamic–pituitary–adrenal regulation Keywords: Paraventricular nucleus; Amygdala; Corticosterone; Corticotropin-releasing factor; Vasopressin; Stress *Corresponding author. Department of Obstetrics and Gynecology, University of Oklahoma Health Sciences Center, Suite 468, RP1, 800 N. Research Parkway, Oklahoma City, OK 73104, USA. Tel.: 11-405-271-2286; fax: 11-405-271-5440. E-mail address: [email protected] (D.A. Myers). 1 Present address: Section on Endocrine Physiology, Developmental Endocrinology Branch, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, MD 20891-1862, USA. 0006-8993 / 02 / $ – see front matter 2002 Elsevier Science B.V. All rights reserved. PII: S0006-8993(02)03978-1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Brain Research 963 (2003) 203–213www.elsevier.com/ locate/brainres
Research report
S tereotaxic localization of corticosterone to the amygdala enhanceshypothalamo-pituitary–adrenal responses to behavioral stress
1 *Jack D. Shepard , Kirk W. Barron, Dean A. MyersDepartment of Physiology, College of Medicine, University of Oklahoma Health Sciences Center, 800 N. Research Parkway, Oklahoma City,
OK 73104,USA
Accepted 29 October 2002
Abstract
The amygdala is involved in behavioral, autonomic, and neuroendocrine responses to stressful stimuli. The goal of the current studywas to determine the effect of directly elevating glucocorticoids in the amygdala on hypothalamo-pituitary–adrenocortical (HPA)responses to the elevated plus maze, a behavioral stressor known to activate the amygdala. Micropellets (30mg) of crystallinecorticosterone or cholesterol (control) were implanted bilaterally at the dorsal margin of the CeA in male Wistar rats; vascular catheterswere also placed at this time. Five days post-surgery, blood samples were drawn at 07:00 and 19:00 h to assess diurnal rhythm of plasmacorticosterone. At 7 days post-implantation, rats were subjected to behavioral stress using an elevated plus maze and blood was collected15 min prior to stress, and at 15, 45, and 90 min after the initiation of the stressor. Corticotropin releasing factor (CRF) and argininevasopressin (AVP) mRNA levels were analyzed by in situ hybridization in the medial parvocellular division of the hypothalamicparaventricular nucleus (mpPVN) in corticosterone- and cholesterol-implanted rats either not exposed to the elevated plus maze (control)or 4 h post-behavioral stress. Localization of corticosterone to the amygdala had no effect on diurnal rhythm of corticosterone secretion.Behavioral stress significantly increased peak plasma corticosterone levels in both groups to a similar level. However, in thecorticosterone implanted rats, plasma corticosterone concentrations at 45 and 90 min post-stress were significantly greater compared tocontrol rats indicating a prolonged corticosterone response to behavioral stress. In non-stressed rats, corticosterone delivery to theamygdala elevated basal CRF mRNA in the mpPVN to levels similar to those observed post-stress in control animals; no further increasewas observed in CRF mRNA following stress. Behavioral stress resulted in a significant elevation in CRF mRNA in cholesterol controls.Basal AVP mRNA levels were unaffected by corticosterone implants. AVP mRNA did not increase in cholesterol implanted rats inresponse to behavioral stress. However, AVP mRNA levels were higher in corticosterone implanted rats post stress compared tocholesterol treated controls. In conclusion, direct administration of corticosterone to the amygdala increases plasma corticosterone inresponse to a behavioral stressor without altering the diurnal rhythm in plasma corticosterone. Elevated basal levels of mpPVN CRFmRNA, and the induction of a mpPVN AVP mRNA response to the behavioral stressor implicate enhanced ACTH secretagogueexpression in the increased HPA response to corticosterone modulation of amygdala function. 2002 Elsevier Science B.V. All rights reserved.
Theme: Endocrine and autonomic regulation
Topic: Hypothalamic–pituitary–adrenal regulation
Keywords: Paraventricular nucleus; Amygdala; Corticosterone; Corticotropin-releasing factor; Vasopressin; Stress
*Corresponding author. Department of Obstetrics and Gynecology, University of Oklahoma Health Sciences Center, Suite 468, RP1, 800 N. ResearchParkway, Oklahoma City, OK 73104, USA. Tel.:11-405-271-2286; fax:11-405-271-5440.
E-mail address: [email protected](D.A. Myers).1Present address: Section on Endocrine Physiology, Developmental Endocrinology Branch, National Institute of Child Health and Human Development,
National Institutes of Health, Bethesda, MD 20891-1862, USA.
0006-8993/02/$ – see front matter 2002 Elsevier Science B.V. All rights reserved.PI I : S0006-8993( 02 )03978-1

204 J.D. Shepard et al. / Brain Research 963 (2003) 203–213
1 . Introduction would increase corticosterone secretion in response tobehavioral stress. We also hypothesized that CRF and AVP
Adequate regulation of glucocorticoid production by the mRNA levels would be elevated in the mpPVN in re-adrenal cortex is critical for preserving homeostasis during sponse to localized delivery of corticosterone to theboth non-stressed and stressed conditions [51]. Adreno- amygdala.cortical glucocorticoid biosynthesis is regulated by anteriorpituitary adrenocorticotrophic hormone (ACTH), which inturn is regulated by corticotropin releasing factor (CRF) 2 . Materials and methodsand arginine vasopressin (AVP) synthesized in neurons ofthe medial parvocellular division of the paraventricular 2 .1. Animalsnucleus of the hypothalamus (mpPVN). In response toeither homeostatic perturbances (physiologic stress) or All protocols were approved by the University ofperceived threats to homeostasis (behavioral stress) Oklahoma Health Sciences Center Institutional Animalglucocorticoid production by the adrenal cortex increases Care and Use Committee. Male Wistar rats (Charles Riverin proportion to the magnitude of the stressor [13,32,33]. Laboratories, Wilmington, MA) weighing 350–450 g wereGlucocorticoids suppress ACTH biosynthesis and release maintained on a 12-h light–dark cycle (lights off 18:00–acting at the level of the anterior pituitary, mpPVN, and 06:00 h) with food and water available ad libitum. Ratslimbic structures such as dorsal hippocampus, ventral were housed in groups of three prior to surgery andsubiculum and cingulate gyrus, assuring that corticosteroid single-housed thereafter.levels are maintained at normal physiological levels innon-stressed states and returned to basal levels following a2 .2. Experimental designstressor [14,38].
Both basal and stress-induced production of glucocor- Upon arrival, rats were habituated to the animal facilityticoids have been noted to increase following chronic for at least 1 week then acclimated to the testing room (2 hstress. Excessive glucocorticoid production following re- per day) for one additional week. The rats were thenpeated or chronic stress has been attributed in part to a stereotaxically implanted with cholesterol (n511) or cor-diminution of negative feedback mechanisms within limbic ticosterone (n512) as previously described [67] andstructures such as the dorsal hippocampus, ventral vascular catheters placed to allow blood collection. Ansubiculum and cingulate gyrus [1,12,18,38]. While the additional three rats were stereotaxically implanted withlimbic structures described above are inhibitory to HPA corticosterone for the determination of corticosteroneactivity, substantial evidence indicates a facilitatory role concentrations in neural tissue surrounding the implanta-for the amygdala in HPA regulation [9]. For instance, tion site. Acclimation to the testing room was continuedstimulation of the amygdala increases c-fos staining in the for one additional week post-operatively (2 h per day). OnmpPVN indicative of neural activation of the mpPVN [57]. day 5 post-surgery, blood (400ml) was collected fromIn addition, CRF, ACTH and corticosterone are released in indwelling jugular cannulas at 07:00 and 19:00 h toresponse to amygdala stimulation [24,27] while lesions of examine the effect of corticosterone implants on diurnalthe central (CeA) or medial (MeA) amygdaloid nuclei, rhythm of corticosterone secretion. On post-operative daysources of major efferent pathways from the amygdala, 7, between 09:00 and 10:00 h (after the 2-h acclimationblock or greatly attenuate HPA responses to stress period), rats were subjected to a behavioral stressor[4,7,8,22,43,60]. CRF and AVP expressing neurons in the (placement on an elevated plus maze for 15 min). BloodmpPVN receive multisynaptic innervation from the samples were collected at 15 min prior to and 15, 45 andamygdala providing a neuroanatomical basis for amygdala 90 min after the initiation of the stressor. Brains wereregulation of the HPA axis [30,62]. collected 4 h post stress for in situ hybridization analysis.
Similar to the limbic structures which inhibit HPA To examine the effect of corticosterone implants on basalactivity, neurons within the amygdala express both type I CRF and AVP mRNA levels, brains were collected fromand type II corticosteroid receptors with the highest density eight rats (cholesterol implantedn54, corticosterone im-located in neurons within the CeA [37,50,63]. Considering plantedn54) receiving the same treatment describedevidence supporting the amygdala as a facilitator of HPA above but were not subjected to the behavioral stressor.activation and localization of glucocorticoid receptorswithin CeA and MeA neurons, the amygdala may provide 2 .3. Surgerya unique site for glucocorticoid facilitation of HPA activi-ty. The purpose of the current study was to determine the 2 .3.1. Vascular catheterizationeffect of stereotaxic localization of corticosterone to the Rats were anesthetized with ketamine (100 mg/kg) andamygdala on HPA function under basal conditions and in acepromazine (1.5 mg/kg) and an incision made at theresponse to a behavioral stressor. We hypothesized that midpoint of the right collar bone. The external jugular veinstereotaxic delivery of corticosterone to the amygdala was exposed by blunt dissection and silicone tubing (0.64

J.D. Shepard et al. / Brain Research 963 (2003) 203–213 205
mm I.D., 1.2 mm O.D., Dow Corning, Midland, MI, USA) 2 .4. Behavioral stressfilled with sterile saline was inserted approximately 30 mminto the jugular vein, ligated to the vessel, sutured to the An elevated plus-maze was used as the behavioral stresspectoral muscle, and tunneled subcutaneously to emerge at paradigm. Animals were placed on the maze for 15 min tothe back of the neck. ensure activation of the HPA axis. The elevated plus maze
was chosen as a behavioral stressor since (1) the elevatedplus-maze has been shown to produce a robust HPA
2 .3.2. Stereotaxic surgery response by 5 min and the amount of corticosteroneRats were stereotaxically implanted (bilaterally) at the produced is proportional to time on the maze [10,26], (2)
dorsal margin of the CeA with crystalline corticosterone or numerous studies indicate that the CeA modulates thecholesterol micropellets as previously described [67]. expression of fear /anxiety, including that exhibited on theImplantation of glucocorticoids directly into discrete brain elevated plus maze [15,42,66], and (3) the elevated plusregions has been used extensively to directly examine the maze is a validated method for measurement of anxiety inactions of glucocorticoids or other compounds on discrete rats [56]. Neural responses to the elevated plus maze areCNS structures [18,25,41,49,64]. Implantation of corticos- likely to include activation of the CeA.terone at the dorsal margin of the CeA allows direct The elevated plus-maze was constructed of black plexi-evaluation of glucocorticoid responsiveness of the glass runways with two open arms (50310 cm) and twoamygdala. Stereotaxic coordinates were: 2.5 mm caudal to arms enclosed with clear plexiglass (50310340 cm) at anbregma,64.2 mm lateral to midline, 7.0 mm ventral to elevation of 50 cm above the floor. Arms of the maze formbrain surface [55]. Micropellets were constructed accord- a cross with two open arms being opposite each other.ing to a procedure previously described by our laboratory Light intensity on the maze was 140 lux. The maze wasand others [25,53,67]. Cholesterol or corticosterone (30 cleaned with a 20% ethanol solution after use and allowedmg) was tamped into 25 Gauge stainless steel cannulas and to dry completely between sessions.micropellets were extruded by inserting a stylet into thecannula after stereotaxically positioning the cannula at the 2 .5. Blood sampling and radioimmunoassaydorsal margin of the CeA. Placement of micropellets wasconfirmed histologically in each animal at the end of the Blood (400ml) was collected in chilled, heparinizedstudy (Fig. 1). tubes via jugular cannula at 15 min before and at 15, 45,
90 min after being placed on the elevated plus-maze.Blood samples were centrifuged at 4003g for 5 min at10 8C the plasma pipetted into cryotubes and stored at280 8C until assayed. The volume of plasma in eachsample was replaced with sterile heparinized saline (30IU/ml) and the red blood cells returned to the animal viathe jugular cannula following the next sample.
Corticosterone concentration was determined for plasmausing a commercially available radioimmunoassay kit(DPC, Los Angeles, CA, USA). The sensitivity of thisassay is 5.7 ng/ml and the intra-assay and inter-assaycoefficients of variation were 3.1 and 8.5%, respectively.
2 .6. Tissue collection
Rats were anesthetized with pentobarbital (75 mg/kg)and immediately decapitated. Brains were then rapidlyremoved and frozen by immersion in isopentane chilledwith pulverized dry ice. Brains were stored at280 8C untilcryosectioning. Serial coronal sections (25mm) werecryosectioned (215 8C), thaw mounted onto sialated slidesand stored at280 8C. One in five series of sectionsthrough the PVN were collected from each animal.
Fig. 1. Sites of steroid implant localization in corticosterone implanted Thymus and adrenal glands were collected, weighed, andrats. Distances from bregma are indicated. Circles with same numbers indexed to body weight as organ weight in mg per 100 g ofindicate right and left implant sites for the same animal. The schematic
body weight. Chronic activation of the HPA axis has beendiagrams are from Paxinos and Watson [55] (Abbreviations: CeM: centralshown to produce adrenal hypertrophy and reduction inamygdaloid nucleus, medial division; CeL: central amygdaloid nucleus,
lateral division; CeC: central amygdaloid nucleus, central division). thymus weight [13].

206 J.D. Shepard et al. / Brain Research 963 (2003) 203–213
2 .7. In situ hybridization Following the final wash, sections were dried overnightat RT then opposed to Hyperfilm-bmax film (Amersham,
35Antisense S-labeled CRF cRNA probes were gener- Arlington Heights, IL, USA) at 220 8C for 7 days,ated from a 700-bp cDNA fragment consisting of the rat developed (GBX developer, Eastman Kodak, Rochester,CRF coding region subcloned into pBluescript. A 2.2-kb NY, USA) and fixed (GBX fixer, Eastman Kodak) to verifyprepro AVP-neurophysin II genomic clone in pBluescript specific labeling. Slides were then dipped in nuclearwas used as a template for PCR generation of a 226-bpemulsion (NBT2, Eastman Kodak), stored at 48C for 30DNA used to generate either sense (control) or antisensedays, developed (D19, Eastman Kodak) for 5 min at 168C,cRNAs. The PCR generated probe consisted of exon cfixed (Kodak fixer), and counterstained with 0.1%(neurophysin /glycoprotein region of the carboxyl terminus toluidine blue.of preprovasopressin-neurophysin II) with either a 39-T3RNA polymerase sequence (antisense generation) or a2 .8. Image analysis59-T7 RNA polymerase sequence incorporated (plasmidsgenerously provided by J.A. Majzoub, Harvard University, The procedure for quantification of in situ hybridizationBoston, MA, USA). One hundred ng of cDNA template has been described in detail for our laboratory [52,67].was transcribed with T7 (CRF) or T3 (AVP) RNA Lightfield images were collected using an Olympus B340polymerase (Ambion, Austin, TX, USA) in 13 transcrip- microscope equipped with a COHU high-performancetion buffer containing 1 mM dithiothreitol (DTT), 40 U CCD camera (RS170; COHU, San Diego, CA, USA).
35ribonuclease inhibitor (Ambion), 125mCi S-UTP (spe- Image analysis was performed using a 7100/66 Powercific activity 1250 Ci /mM; New England Nuclear, Boston, Macintosh with NIH Image software (version 1.62, W.MA, USA), 0.5 mmol each of ATP, CTP, and GTP Rasband, National Institutes of Health,http: / / rsb.in-(Boehringer Mannheim, Indianapolis, IN, USA), in a fo.nih.gov/nih-image). Two sections from each animalvolume of 40ml. Transcription reactions were carried out within the mpPVN were analyzed. Background counters-at 378C for 60 min. At this time, deoxyribonuclease I (20 taining (toluidine blue) was reduced in each image usingU) was added with 10 U ribonuclease inhibitor and the Wratten 4A (Kodak) filtered light during image capturecDNA template digested for 10 min at 378C. Complemen- [52,67]. Once captured, images were subjected to high passtary RNA probes were purified by gel filtration column filtration to increase contrast between silver grains andchromatography (Pharmacia & Upjohn, Piscataway, NJ, underlying stained neurons. Images were then duplicatedUSA). and the duplicated images superimposed on the originals.
The procedure for in situ hybridization as performed in Maximum gray level of the histological staining wasour laboratory has been previously described in detail determined and subtracted from the duplicated images.[52,67]. Mounted tissue was fixed (4% paraformaldehyde), Thus, on the duplicated images, only gray level for silveracetylated (0.25% acetic anhydride in 0.1 M tri- grains remained (background grayscale50; silver grainethanolamine), dehydrated with progressively higher con- grayscale.0). Stained neurons or small groups of neuronscentrations of ethanol (70, 80, 95, 100%), delipidated were outlined in the original image and the neuronal(chloroform), dried, covered with 200ml prehybridization outlines transferred to the duplicate images allowing
2solution (50% formamide, 43 SSC (13 SSC50.15 M determination of silver grain number permm . The grayNaCl, 0.015 M Na citrate, pH 7.2), 2.53 Denhardt’s level for pixels over each outlined area was determinedsolution (13 Denhardt’s is 1% solution of BSA, Ficoll and and the number of pixels greater than zero (representingpolyvinylpyrrolidone), 10% dextran sulfate, 4 mM EDTA, the area of silver grains) was divided by the number of0.5 mg/ml denatured sonicated salmon sperm DNA, 0.25 pixels per silver grain providing an accurate determinationmg/ml yeast tRNA, 10 mM NaHPO , 10 mM DTT), of silver grains per area [52,67]. In preliminary experi-4
coverslipped with parafilm, and prehybridized for 2 h in a ments, there was a close linear relationship betweenmoist chamber at 558C. Hybridization was carried out in manual grain counts and this method (r50.93; n525
7prehybridization solution containing 1310 cpm/ml neurons; range of grains per neuron 5–69). BackgroundcRNA probe at 558C for 18 h. Post-hybridization, sections hybridization was calculated from 25 neurons in thewere washed twice in 43 SSC at room temperature (RT) thalamic region of each slide. Specific hybridization wasfollowed by incubation with 20mg/ml ribonuclease A and determined as an area for which the number of overlying0.5 U/ml ribonuclease T1 (Boehringer Mannheim) for 30 silver grains was greater than two standard deviationsmin at 378C. Sections were then washed in 50 mM NaCl above the background mean number of silver grains.for 30 min at 378C, 13 SSC for 15 min at RT, twice in Background number of silver grains was subtracted from0.13 SSC for 30 min each at 658C, and 0.13 SSC for 15 hybridization signal of specifically labeled neurons.min at RT. Control in situ hybridization was performed Magnocellular neurons within the mpPVN region
35using sense-strand S-labeled cRNA probes. Slides for labeled for AVP were identified and excluded from analy-each probe were processed in a single in situ hybridization sis. Magnocellular neurons were identified by plotting thereaction. number of silver grains per unit area revealing a bimodal

J.D. Shepard et al. / Brain Research 963 (2003) 203–213 207
distribution of labeled neurons. Since magnocellular neu- cycles followed by addition of 200ml of 50 mM Tris–HCl,rons express high levels of AVP the distribution represent- 10 sodium chloride, 10 mM EDTA (pH 8.0) containinging the most heavily labeled neurons were considered to be proteinase K (50mg/ml) and 0.01% Triton X-100. Themagnocellular and excluded from the data set. As predicted tissue was then incubated for 1 h at 658C with occasionalfrom previous descriptions [68], the majority of the heavily vortexing. At the end of the incubation period the sampleslabeled AVP neurons resided within the magnocellular were boiled for 5 min, chilled on ice and centrifuged atsubdivision of the PVN with a few scattered magnocellular 20003g. Corticosterone concentrations in the extract wereneurons located within the parvocellular subdivisions. then measured directly in the lysate using a corticosterone
ELISA (IDS, Bolton, UK) which was found to not be2 .9. Determination of corticosterone diffusion from the affected by the presence of Triton X-100 at the con-implant centration used. The range of the ELISA was 0.5–191
ng/ml.At 5 days post-stereotaxic surgery, rats (n53) were
anesthetized with pentobarbital and immediately decapi- 2 .10. Statistical analysistated. The brains were then rapidly removed, chilled brieflyin ice-cold saline and placed into a Stoelting rat brain Data are presented as mean6standard error of the meanslicer (No. 51384; Stoelting, Wood Dale, IL, USA). One (S.E.M.). A 2 Implant (cholesterol, corticosterone)34mm coronal sections were obtained rostral to caudal until Period (215, 15, 45, 90 min) repeated measures analysis ofthe steroid implant site was located. Sections were placed variance (ANOVA) was performed on plasma corticos-onto glass slides and a micro punch (0.74 mm diameter) terone values. A 2 Implant (cholesterol, corticosterone)32was used to obtain tissue punches (0.74 mm diameter31.0 Period (07:00, 19:00 h) ANOVA was used to determine themm depth) with the aid of a dissecting microscope (see effect of corticosterone implants on diurnal rhythm ofFig. 2 for punch sites). Tissue punches were placed into corticosterone secretion. A 2 Implant (cholesterol,1.5 ml polypropylene micocentrifuge tubes and stored at corticosterone)32 Stress (basal, stress) analysis of variance280 8C. To ascertain corticosterone concentrations in the was performed on in situ hybridization data. Main effectstissue, the punches were subjected to two freeze–thaw were analyzed with Bonferroni adjusted pairwise com-
parisons where appropriate.
3 . Results
3 .1. Plasma corticosterone
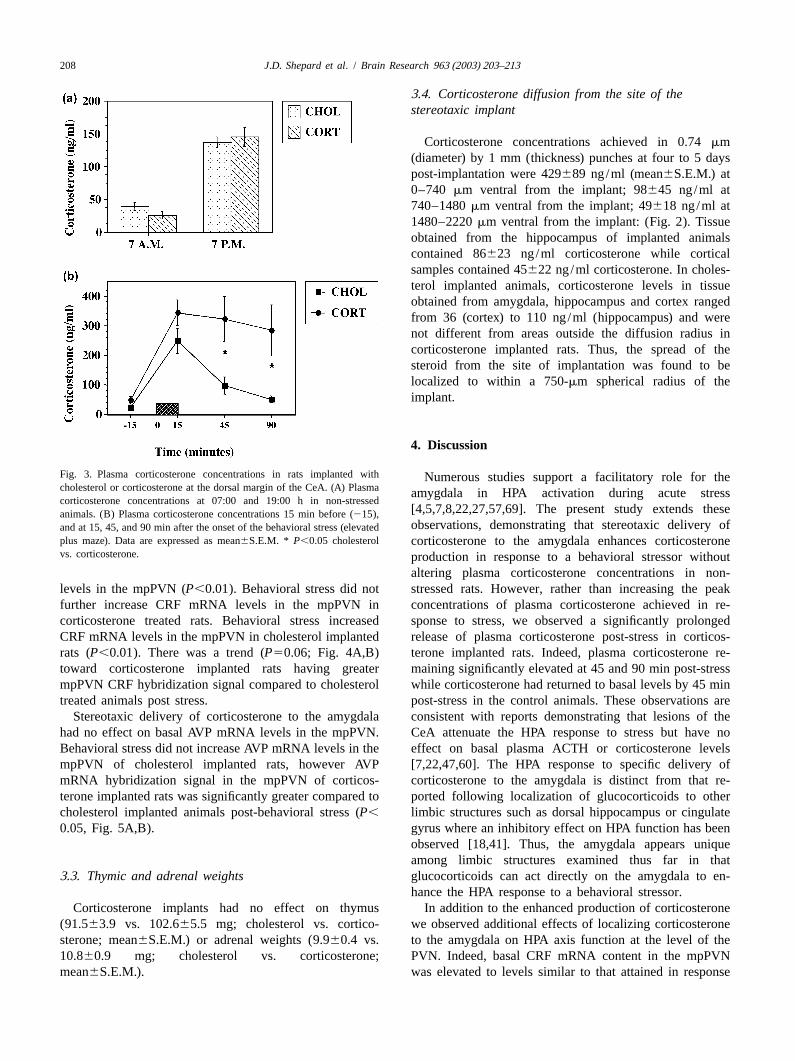
Both cholesterol and corticosterone implanted rats ex-hibited a normal diurnal pattern of corticosterone withsignificantly greater plasma corticosterone concentrationsobserved at 19:00 h compared to 07:00 h (P,0.05, Fig.3A). No differences were observed in plasma corticos-terone concentrations between cholesterol and corticos-terone-implanted animals at either time point. While peakcorticosterone concentrations attained in response to theelevated plus-maze (behavioral stressor) were not differentbetween groups, corticosterone remaining significantlyelevated at 45 and 90 min post stress in the corticosterone-implanted animals compared to the cholesterol-implantedrats implanted (P,0.05, Fig. 3B). Area under the curvemeasurement of corticosterone release post-stressor alsodemonstrated significantly greater release of corticosteronein the corticosterone implanted rats compared to control(P,0.025).
Fig. 2. Corticosterone diffusion from the steroid implant. The 0.74-mm 3 .2. CRF and AVP mRNA levels in the medialdiameter (by 1 mm thick) punches were taken where indicated (numbered parvocellular paraventricular nucleuscircles) relative to the implantation site (hatched bar). Corticosteroneconcentrations in the tissue punches were: (1) 459679 (mean6S.E.M.);
Stereotaxic implantation of corticosterone at the level of(2) 98639; (3) 39618; (4) 66623; (5) 45622 ng/ml. The schematicdiagrams are from Paxinos and Watson [55]. the amygdala resulted in increased basal CRF mRNA
208 J.D. Shepard et al. / Brain Research 963 (2003) 203–213
3 .4. Corticosterone diffusion from the site of thestereotaxic implant
Corticosterone concentrations achieved in 0.74mm(diameter) by 1 mm (thickness) punches at four to 5 dayspost-implantation were 429689 ng/ml (mean6S.E.M.) at0–740 mm ventral from the implant; 98645 ng/ml at740–1480mm ventral from the implant; 49618 ng/ml at1480–2220mm ventral from the implant: (Fig. 2). Tissueobtained from the hippocampus of implanted animalscontained 86623 ng/ml corticosterone while corticalsamples contained 45622 ng/ml corticosterone. In choles-terol implanted animals, corticosterone levels in tissueobtained from amygdala, hippocampus and cortex rangedfrom 36 (cortex) to 110 ng/ml (hippocampus) and werenot different from areas outside the diffusion radius incorticosterone implanted rats. Thus, the spread of thesteroid from the site of implantation was found to belocalized to within a 750-mm spherical radius of theimplant.
4 . Discussion
Fig. 3. Plasma corticosterone concentrations in rats implanted with Numerous studies support a facilitatory role for thecholesterol or corticosterone at the dorsal margin of the CeA. (A) Plasma amygdala in HPA activation during acute stresscorticosterone concentrations at 07:00 and 19:00 h in non-stressed
[4,5,7,8,22,27,57,69]. The present study extends theseanimals. (B) Plasma corticosterone concentrations 15 min before (215),observations, demonstrating that stereotaxic delivery ofand at 15, 45, and 90 min after the onset of the behavioral stress (elevated
plus maze). Data are expressed as mean6S.E.M. * P,0.05 cholesterol corticosterone to the amygdala enhances corticosteronevs. corticosterone. production in response to a behavioral stressor without
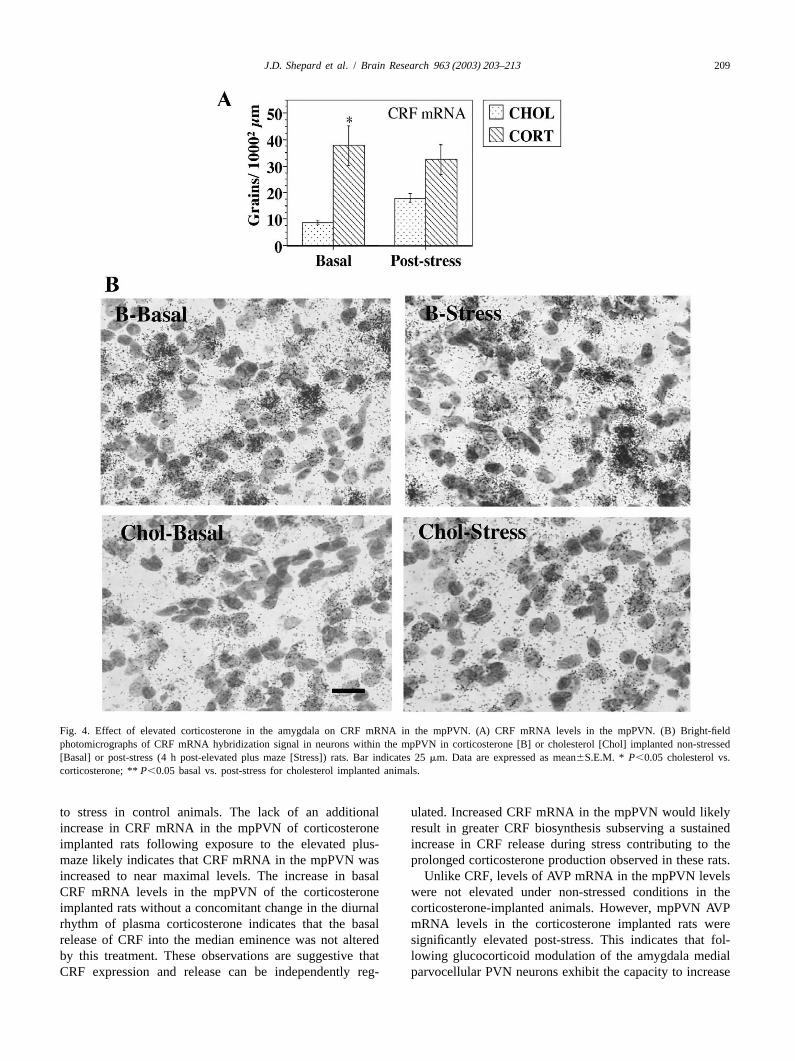
altering plasma corticosterone concentrations in non-levels in the mpPVN (P,0.01). Behavioral stress did not stressed rats. However, rather than increasing the peakfurther increase CRF mRNA levels in the mpPVN in concentrations of plasma corticosterone achieved in re-corticosterone treated rats. Behavioral stress increased sponse to stress, we observed a significantly prolongedCRF mRNA levels in the mpPVN in cholesterol implanted release of plasma corticosterone post-stress in corticos-rats (P,0.01). There was a trend (P50.06; Fig. 4A,B) terone implanted rats. Indeed, plasma corticosterone re-toward corticosterone implanted rats having greater maining significantly elevated at 45 and 90 min post-stressmpPVN CRF hybridization signal compared to cholesterol while corticosterone had returned to basal levels by 45 mintreated animals post stress. post-stress in the control animals. These observations are
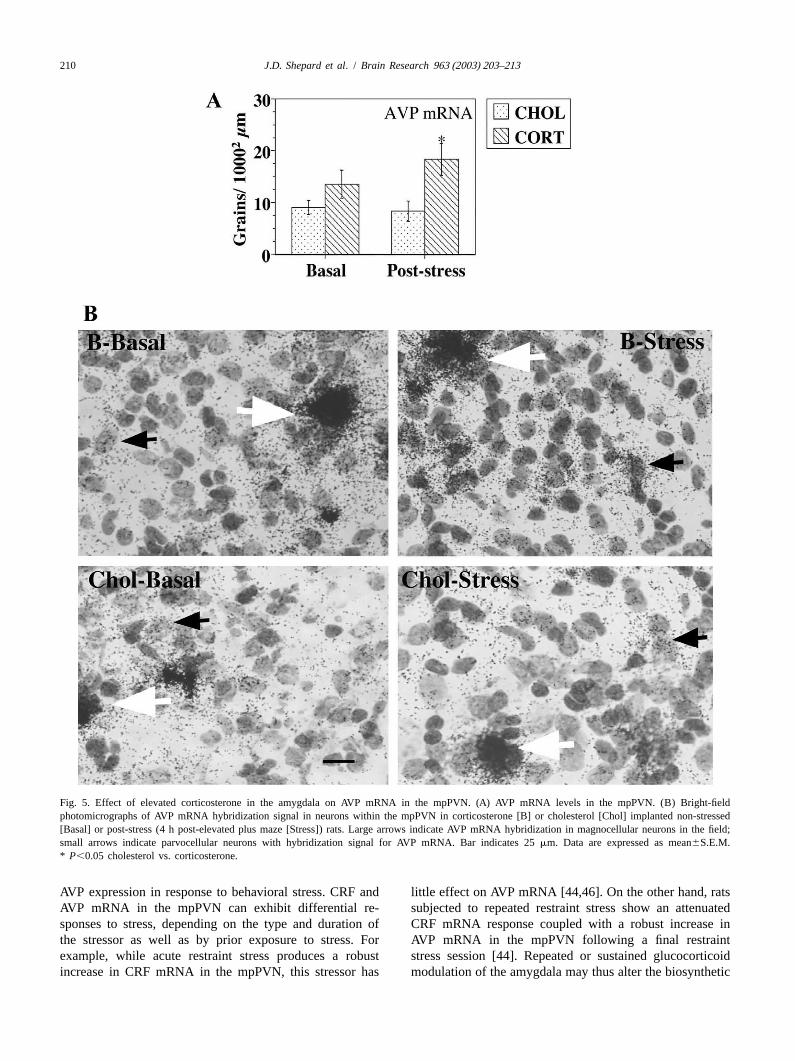
Stereotaxic delivery of corticosterone to the amygdala consistent with reports demonstrating that lesions of thehad no effect on basal AVP mRNA levels in the mpPVN. CeA attenuate the HPA response to stress but have noBehavioral stress did not increase AVP mRNA levels in the effect on basal plasma ACTH or corticosterone levelsmpPVN of cholesterol implanted rats, however AVP [7,22,47,60]. The HPA response to specific delivery ofmRNA hybridization signal in the mpPVN of corticos- corticosterone to the amygdala is distinct from that re-terone implanted rats was significantly greater compared to ported following localization of glucocorticoids to othercholesterol implanted animals post-behavioral stress (P, limbic structures such as dorsal hippocampus or cingulate0.05, Fig. 5A,B). gyrus where an inhibitory effect on HPA function has been
observed [18,41]. Thus, the amygdala appears uniqueamong limbic structures examined thus far in that
3 .3. Thymic and adrenal weights glucocorticoids can act directly on the amygdala to en-hance the HPA response to a behavioral stressor.
Corticosterone implants had no effect on thymus In addition to the enhanced production of corticosterone(91.563.9 vs. 102.665.5 mg; cholesterol vs. cortico- we observed additional effects of localizing corticosteronesterone; mean6S.E.M.) or adrenal weights (9.960.4 vs. to the amygdala on HPA axis function at the level of the10.860.9 mg; cholesterol vs. corticosterone; PVN. Indeed, basal CRF mRNA content in the mpPVNmean6S.E.M.). was elevated to levels similar to that attained in response
J.D. Shepard et al. / Brain Research 963 (2003) 203–213 209
Fig. 4. Effect of elevated corticosterone in the amygdala on CRF mRNA in the mpPVN. (A) CRF mRNA levels in the mpPVN. (B) Bright-fieldphotomicrographs of CRF mRNA hybridization signal in neurons within the mpPVN in corticosterone [B] or cholesterol [Chol] implanted non-stressed[Basal] or post-stress (4 h post-elevated plus maze [Stress]) rats. Bar indicates 25mm. Data are expressed as mean6S.E.M. * P,0.05 cholesterol vs.corticosterone; **P,0.05 basal vs. post-stress for cholesterol implanted animals.
to stress in control animals. The lack of an additional ulated. Increased CRF mRNA in the mpPVN would likelyincrease in CRF mRNA in the mpPVN of corticosterone result in greater CRF biosynthesis subserving a sustainedimplanted rats following exposure to the elevated plus- increase in CRF release during stress contributing to themaze likely indicates that CRF mRNA in the mpPVN was prolonged corticosterone production observed in these rats.increased to near maximal levels. The increase in basal Unlike CRF, levels of AVP mRNA in the mpPVN levelsCRF mRNA levels in the mpPVN of the corticosterone were not elevated under non-stressed conditions in theimplanted rats without a concomitant change in the diurnal corticosterone-implanted animals. However, mpPVN AVPrhythm of plasma corticosterone indicates that the basal mRNA levels in the corticosterone implanted rats wererelease of CRF into the median eminence was not altered significantly elevated post-stress. This indicates that fol-by this treatment. These observations are suggestive that lowing glucocorticoid modulation of the amygdala medialCRF expression and release can be independently reg- parvocellular PVN neurons exhibit the capacity to increase
210 J.D. Shepard et al. / Brain Research 963 (2003) 203–213
Fig. 5. Effect of elevated corticosterone in the amygdala on AVP mRNA in the mpPVN. (A) AVP mRNA levels in the mpPVN. (B) Bright-fieldphotomicrographs of AVP mRNA hybridization signal in neurons within the mpPVN in corticosterone [B] or cholesterol [Chol] implanted non-stressed[Basal] or post-stress (4 h post-elevated plus maze [Stress]) rats. Large arrows indicate AVP mRNA hybridization in magnocellular neurons in the field;small arrows indicate parvocellular neurons with hybridization signal for AVP mRNA. Bar indicates 25mm. Data are expressed as mean6S.E.M.* P,0.05 cholesterol vs. corticosterone.
AVP expression in response to behavioral stress. CRF and little effect on AVP mRNA [44,46]. On the other hand, ratsAVP mRNA in the mpPVN can exhibit differential re- subjected to repeated restraint stress show an attenuatedsponses to stress, depending on the type and duration of CRF mRNA response coupled with a robust increase inthe stressor as well as by prior exposure to stress. For AVP mRNA in the mpPVN following a final restraintexample, while acute restraint stress produces a robust stress session [44]. Repeated or sustained glucocorticoidincrease in CRF mRNA in the mpPVN, this stressor has modulation of the amygdala may thus alter the biosynthetic

J.D. Shepard et al. / Brain Research 963 (2003) 203–213 211
capacity of mpPVN neurons. Since AVP potentiates the in the relative roles of the CeA and MeA reported in thestimulatory effect of CRF on ACTH release, increased literature. Although the rostral CeA was targeted in theAVP expression in mpPVN neurons and co-secretion with present study, the rostral CeA and MeA were both withinCRF are important adaptations to chronic stress insuring the diffusion radius of the corticosterone implants, thussufficient HPA drive when glucocorticoids are elevated both these nuclei could play a role in modulating PVN[1,2]. The increase AVP mRNA in mpPVN in response to function [31,50,63].stress in the corticosterone-implanted rats likely contri- While stimulation of the amygdala results in activationbutes in the sustained corticosterone production in re- of mpPVN neurons, the exact pathways mediating thissponse to stress in corticosterone-implanted rats. Our effect remain to be elucidated [57,59]. CRF containingfindings provide evidence that the amygdala can modulate neurons project directly from the CeA to the mpPVNACTH secretagogue synthesis in the mpPVN and may play providing a possible mechanism for CRF synthesized ina role maintaining HPA responsiveness by influencing CRF the CeA to be released into the mpPVN [30,48]. Althoughand AVP expression in the mpPVN as well as the CRF administration into the mpPVN increases c-fos andAVP:CRF ratio. CRF expression in this nucleus the direct CeA projections
In contrast to our findings that stereotaxic delivery of to the mpPVN are relatively sparse, thus CRF release fromcorticosterone to the amygdala facilitated HPA function, CeA terminals may be insufficient for mpPVN activationAkana et al. [3] observed that localizing corticosterone to [54]. An attractive mechanism via which the amygdalathe amygdala did not effect ACTH release in response to may facilitate HPA activity is through a multisynapticrestraint stress in adrenalectomized, corticosterone re- pathway to the mpPVN via the bed nuclei of striaplaced rats. Similar to our findings these authors observed terminalis (BNST) and preoptic area. The CeA and MeAno effect on basal ACTH levels. However, in that study project heavily to the lateral and medial BNST, respective-plasma ACTH was quantified at 15 and 30 min after the ly [17]. Stimulation of the of the lateral BNST increasesonset of restraint stress during the ascending limb of the CRF mRNA in the mpPVN and plasma corticosteroneHPA response. As indicated above, rather than observing while stimulation of the medial BNST decreases CRFan increase in peak plasma corticosterone, we found a mRNA in the mpPVN and plasma corticosterone con-prolongation of corticosterone production in the corticos- centration [21,35]. Alternatively, the amygdala could en-terone-implanted rats at 45 and 90 min post-initiation of hance HPA responses to stress by projections to the medialstress. Alternatively, HPA-enhancing effects of corticos- preoptic area of the hypothalamus via the MeA. In supportterone on the amygdala may show dependence on the type of a BNST/preoptic pathway mediating amygdala stimula-of stressor and/or magnitude of stress. Furthermore, the tion of the HPA axis, bilateral lesions of the BNST orfacilitated corticosterone production seen in the glucocor- preoptic area inhibit MeA-induced activation of the HPAticoid implanted rats in the present study may be via axis. The greatest inhibition occurs when both BNST andmodulation of glucocorticoid termination (feedback) of the preoptic area are lesioned [23]. The amygdala may alsoHPA response to stress and as such would require adrenal- counter the inhibitory influence of the hippocampus onintact animals eliciting a stress-induced increase in plasma PVN function via the BNST and/or preoptic area [11,34].corticosterone. The hippocampus plays a prominent role in not only basal
How elevated glucocorticoids or prior exposure to stress HPA function but also in limiting the magnitude andmodulates information processing within the amygdala is duration of the stress response [38,39]. Recent evidencepresently unknown. It is clear that the amygdala is indicates that the ventral subiculum plays a prominent roleinvolved in processing stressful stimuli and amygdaloid in mediating hippocampal inhibition of the PVN [36]. Thenuclei may be differentially involved in modulating HPA ventral subiculum in turn mediates mpPVN function via aresponses to various forms of stress. While some inves- multisynaptic pathway including the BNST and preoptictigators report CeA activation in response to acute be- area; thus the BNST and preoptic area are likely sites ofhavioral stressors [45] others have found the MeA to have integration for hippocampus and amygdala regulation ofa more prominent role [16]. Considering that extensive the mpPVN. Our observation of the prolonged productioninterconnections exist between amygdaloid nuclei as well of corticosterone post-stress may indeed reflect a reducedas within subdivisions of nuclei (including the CeA and ability to effectively limit the duration of the stressMeA) it is likely that glucocorticoid modulation of response. In the present study, elevated corticosterone inamygdala function is complex [58]. As discussed previous- the amygdala may not act as a direct positive feedbackly many investigators have reported attenuation of the HPA signal per se but may reduce the inhibitory influence of theresponse to stress following CeA lesions. While Prewitt hippocampus on the HPA axis when circulating glucocor-and Herman [61] reproduced these effects, identical CeA ticoids are elevated.lesions in a follow up experiment failed to reduce ACTH The role of the amygdala in behavioral and neuroen-and corticosterone levels in response to restraint stress. docrine responses to stress led clinical investigators toPlasticity of the amygdala in response to prior stress or examine amygdala function in mood disorders. Functionalexposure to glucocorticoids may account for the variability imaging studies have consistently demonstrated increased

212 J.D. Shepard et al. / Brain Research 963 (2003) 203–213
[12] M.F. Dallman, Stress Update: Adaptation of the hypothalamic–activation of the amygdala in patients suffering from majorpituitary–adrenal axis to chronic stress, Trends Endocrinol. Metab. 4depression [19,20]. Furthermore, many of these patients(1993) 62–69.
present with elevated glucocorticoid levels coupled with [13] M.F. Dallman, S.F. Akana, C.S. Cascio, D.N. Darlington, L.increased cerebrospinal fluid levels of CRF [6,28,29,40]. Jacobson, N. Levin, Regulation of ACTH secretion: variations on aOne model of melancholic depression postulates that theme of B, Recent Prog. Horm. Res. 43 (1987) 113–167.
[14] M.F. Dallman, S.F. Akana, N. Levin, C.-D. Walker, M.J. Bradbury,chronically elevated glucocorticoids may result in over-S. Suemaru, K.S. Scribner, Corticosteroids and the control ofexpression of CRF in the amygdala and contribute tofunction in the hypothalamo-pituitary–adrenal (HPA) axis, Ann. NYdepressive symptoms [65,66]. The current data provideAcad. Sci. 746 (1994) 22–28.
evidence that chronically elevated glucocorticoids at the [15] M. Davis, Neurobiology of fear responses: the role of the amygdala,level of the amygdala may also be involved in HPA J. Neuropsychiatry Clin. Neurosci. 9 (1997) 382–402.
[16] C.V. Dayas, K.M. Buller, T.A. Day, Neuroendocrine responses to anactivation as well.emotional stressor: evidence for involvement of the medial but notIn conclusion, chronic corticosterone administration tothe central amygdala, Eur. J. Neurosci. 11 (1999) 2312–2322.the amygdala potentiates the plasma corticosterone re-
[17] J.S. de Olmos, L. Heimer, The concepts of the ventral striatopallidalsponse to behavioral stress. CRF mRNA was increased to system and extended amygdala, Ann. NY Acad. Sci. 877 (1999)stress levels in the mpPVN in response to elevated 1–32.
[18] D. Diorio, V. Viau, M.J. Meaney, The role of the medial prefrontalcorticosterone in the amygdala and no further increase wascortex (cingulate gyrus) in the regulation of hypothalamic–pituitary–produced in response to stress. AVP mRNA in the mpPVNadrenal responses to stress, J. Neurosci. 13 (1993) 3839–3847.was not increased in response to corticosterone treatment
[19] W.C. Drevets, Functional anatomical abnormalities in limbic andalone but was elevated in corticosterone implanted animals prefrontal cortical structures in major depression, Prog. Brain Res.following behavioral stress. Thus, the enhanced HPA 126 (2000) 413–431.responses to behavioral stress may have been mediated by[20] W.C. Drevets, Neuroimaging and neuropathological studies of
depression: implications for the cognitive–emotional features ofaugmentation of CRF expression coupled with recruitmentmood disorders, Curr. Opin. Neurobiol. 11 (2001) 240–249.of a medial parvocellular AVP response.
[21] J.D. Dunn, Plasma corticosterone responses to electrical stimulationof the bed nucleus of the stria terminalis, Brain Res. 407 (1987)327–331.
[22] S. Feldman, N. Conforti, A. Itzik, J. Weidenfield, Differential effectR eferencesof amygdaloid lesions on CRF-41 ACTH and corticosterone re-sponses following neural stimuli, Brain Res. 658 (1994) 21–26.
[1] G. Aguilera, Regulation of pituitary ACTH secretion during chronic [23] S. Feldman, N. Conforti, D. Saphier, The preoptic area and bedstress, Front. Neuroendocrinol. 15 (1994) 321–350. nucleus of the stria terminalis are involved in the effects of the
[2] G. Aguilera, Corticotropin releasing hormone, receptor regulation amygdala on adrenocortical secretion, Neuroscience 37 (1990) 775–and the stress response, Trends Endocrinol. Metab. 9 (1998) 329– 779.336. [24] S. Feldman, J. Weidenfeld, The excitatory effects of the amygdala
[3] S.F. Akana, A. Chu, L. Soriano, M.F. Dallman, Corticosterone on the hypothalamo-pituitary–adrenocortical responses are mediatedexerts site-specific and state dependent effects in prefrontal cortex by hypothalamic norepinephrine, serotonin, and CRF-41, Brain Res.and amygdala on regulation of adrenocorticotropic hormone, insulin Bull. 45 (1998) 389–393.and fat deposits, J. Neuroendocrinol. 13 (2001) 625–637. [25] S. Feldman, J. Weidenfeld, Glucocorticoid receptor antagonists in
[4] J.P. Allen, C.F. Allen, Role of the amygdaloid complexes in the the hippocampus modify the negative feedback following neuralstress-induced release of ACTH in the rat, Neuroendocrinology 15 stimuli, Brain Res. 821 (1999) 33–37.(1974) 220–230. [26] S.E. File, H. Zangrossi, F.L. Sanders, P.S. Mabbutt, Raised corticos-
[5] J.P. Allen, C.F. Allen, Amygdalar participation in tonic ACTH terone in the rat after exposure to the elevated plus-maze, Psycho-secretion in the rat, Neuroendocrinology 19 (1975) 115–125. pharmacology 113 (1994) 543–546.
[6] L. Arborelius, M.J. Owens, P.M. Plotsky, C.B. Nemeroff, The role [27] R.W. Gabr, D.L. Birkle, A.J. Azzaro, Stimulation of the amygdala byof corticotropin-releasing factor in depression and anxiety disorders, glutamate facilitates corticotropin-releasing factor release from theJ. Endocrinol. 160 (1999) 1–12. median eminence and activation of the hypothalamic–pituitary–
[7] S. Beaulieu, T. Di Paolo, N. Barden, Control of ACTH secretion by adrenal axis in stressed rats, Neuroendocrinology 62 (1995) 333–the central nucleus of the amygdala: implication of the serotoniner- 339.gic systems and its relevance to the glucocorticoid delayed negative [28] P.W. Gold, G.P. Chrousos, The endocrinology of melancholic andfeedback mechanism, Neuroendocrinology 44 (1986) 247–254. atypical depression: relation to neurocircuitry and somatic conse-
[8] S. Beaulieu, G. Pelletier, H. Vaudry, N. Barden, Influence of the quences, Proc. Assoc. Am. Physicians 111 (1999) 22–34.central nucleus of the amygdala on the content of corticotropin- [29] P.W. Gold, J. Licinio, M. Wong, G.P. Chrousos, Corticotropinreleasing factor in the median eminence, Neuroendocrinology 49 releasing hormone in the pathophysiology of melancholic and(1989) 255–261. atypical depression and in the mechanism of action of antidepressant
[9] S. Bhatnagar, M.F. Dallman, Neuroanatomical basis for facilitation drugs, Ann. NY Acad. Sci. 771 (1995) 716–729.of hypothalamic–pituitary–adrenal responses to a novel stressor [30] T.S. Gray, M.E. Carney, D.J. Magnuson, Direct projections from theafter chronic stress, Neuroscience 84 (1998) 1025–1039. central amygdaloid nucleus to the hypothalamic paraventricular
[10] N. Calvo, I.D. Martijena, V.A. Molina, M. Volosin, Metyrapone nucleus: possible role in stress-induced adrenocorticotropin release,pretreatment prevents the behavioral and neurochemical sequelae Neuroendocrinology 50 (1989) 433–446.induced by stress, Brain Res. 800 (1998) 227–235. [31] M.B. Greenwood-Van, M. Gibson, W. Gunter, J. Shepard, R.
[11] N.S. Canteras, R.B. Simberly, L.W. Swanson, Organization of Foreman, D. Myers, Stereotaxic delivery of corticosterone to theprojections from the medial nucleus of the amygdala: A PHAL amygdala modulates colonic sensitivity in rats, Brain Res. 893study in the rat, J. Comp. Neurol. 360 (1995) 213–245. (2001) 135–142.

J.D. Shepard et al. / Brain Research 963 (2003) 203–213 213
[32] M.B. Hennessy, J.P. Heybach, J. Vernikos, S. Levine, Plasma [51] A. Munck, P.M. Guyre, N.J. Holbrook, Physiological functions ofcorticosterone concentrations sensitively reflect levels of stimulus glucocorticoids in stress and their relation to pharmacologicalintensity in the rat, Physiol. Behav. 22 (1979) 821–825. actions, Endocrinol. Rev. 5 (1984) 25–44.
[33] M.B. Hennessy, S. Levine, Sensitive pituitary–adrenal responsive- [52] D.A. Myers, M.E. Bell, T.J. McDonald, T.R. Myers, Corticotropin-ness to varying intensities of psychological stimulation, Physiol. releasing factor receptor expression in the pituitary of fetal sheepBehav. 21 (1978) 295–297. after lesions of the hypothalamic paraventricular nucleus, Endo-
[34] J.P. Herman, W.E. Cullinan, Neurocircuitry of stress: central control crinology 140 (1999) 4292–4299.of the hypothalamo-pituitary–adrenocortical axis, Trends Neurosci. [53] D.A. Myers, T.J. McDonald, T.G. Dunn, G.E. Moss, P.W.20 (1997) 78–84. Nathanielsz, Effect of implantation of dexamethasone adjacent to the
[35] J.P. Herman, W.E. Cullinan, S.J. Watson, Involvement of the bed paraventricular nucleus on messenger ribonucleic acid for corticot-nucleus of the stria terminalis in tonic regulation of paraventricular ropin-releasing hormone and proopiomelanocortin during late gesta-hypothalamic CRH and AVP mRNA expression, J. Neuroendocrinol. tion in fetal sheep, Endocrinology 130 (1992) 2167–2172.6 (1994) 433–442. [54] D. Parkes, S. Rivest, S. Lee, C. Rivier, W. Vale, Corticotropin-
[36] J.P. Herman, W.E. Cullinan, M.I. Morano, H. Akil, S.J. Watson, releasing factor activates c-fos, NGFI-B, and corticotropin-releasingContribution of the ventral subiculum to inhibitory regulation of the factor gene expression within the paraventricular nucleus of the rathypothalamo-pituitary–adrenocortical axis, J. Neuroendocrinol. 7 hypothalamus, Mol. Endocrinol. 7 (1993) 1357–1367.(1995) 475–482. [55] G.T. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates,
[37] J. Honkaniemi, M. Pelto-Huikko, L. Rechardt, J. Isola, A. Lammi, Academic Press, New York, 1997.K. Fuxe, J. Gustafsson, A. Wikstrom, T. Hokfelt, Colocalization of [56] S. Pellow, P. Chopin, S.E. File, M. Briley,Validation of open: closedpeptide and glucocorticoid receptor immunoreactivities in rat central arm entries in an elevated plus-maze as a measure of anxiety in theamygdaloid nucleus, Neuroendocrinology 55 (1992) 451–459. rat, J. Neurosci. Methods 14 (1985) 149–167.
[38] L. Jacobson, R. Sapolsky, The role of the hippocampus in feedback [57] T. Petrov, J.H. Jhamandas, T.L. Krukoff, Electrical stimulation ofregulation of the hypothalamic–pituitary–adrenocortical axis, Endo- the central nucleus of the amygdala induces fos-like immuno-cr. Rev. 12 (1991) 118–134. reactivity in the hypothalamus of the rat: a quantitative study, Mol.
[39] M. Joels, E. Vreugdenhil, Corticosteroids in the brain. Cellular and Brain Res. 22 (1994) 333–340.molecular actions, Mol. Neurobiol. 17 (1998) 87–108. [58] A. Pitkanen, V. Savander, J.E. Ledoux, Organization of intra-
[40] J.W. Kasckow, D. Baker, T.D.J. Geracioti, Corticotropin-releasing amygdaloid circuitries in the rat: an emerging framework forhormone in depression and post-traumatic stress disorder, Peptides understanding functions of the amygdala, Trends Neurosci. 2022 (2001) 845–851. (1997) 517–523.
[41] K.J. Kovacs, G.B. Makara, Corticosterone and dexamethasone act at [59] Q.J. Pittman, H.W. Blume, L.P. Renaud, Connections of the hypo-different brain sites to inhibit adrenalectomy-induced adrenocor- thalamic paraventricular nucleus with the neurohypophysis, medianticotropin hypersecretion, Brain Res. 474 (1988) 205–210. eminence, amygdala, lateral septum and midbrain periaqueductal
[42] J.E. Ledoux, Fear and the brain: where have we been, and where are gray: an electrophysiological study in the rat, Brain Res. 215 (1981)we going?, Biol. Psychiatry 44 (1998) 1229–1238. 15–28.
[43] M.P. Lilly, D.J. Putney, D.E. Carlson, Potentiated response of [60] C.M. Prewitt, J.P. Herman, Lesion of the central nucleus of thecorticotropin (ACTH) to repeated moderate hemorrhage requires amygdala decreases basal CRH mRNA expression and stress-in-amygdalar neuronal processing, Neuroendocrinology 71 (2000) 88– duced ACTH release, Ann. NY Acad. Sci. 746 (1994) 438–440.98. [61] C.M. Prewitt, J.P. Herman, Hypothalamo-pituitary–adrenocortical
[44] X.M. Ma, A. Levy, S.L. Lightman, Emergence of an isolated regulation following lesions of the central nucleus of the amygdala,arginine vasopressin (AVP) response to stress after repeated re- Stress 1 (1997) 263–280.straint: a study of both AVP and corticotropin-releasing hormone [62] C.M. Prewitt, J.P. Herman, Anatomical interactions between themessenger ribonucleic acid (RNA) and heteronuclear RNA, Endo- central amygdaloid nucleus and the hypothalamic paraventricularcrinology 138 (1997) 4351–4357. nucleus of the rat: a dual tract-tracing analysis, J. Chem. Neuroanat.
[45] S. Makino, T. Shibasaki, N. Yamauchi, T. Nishioka, T. Mimoto, I. 15 (1998) 173–185.Wakabayashi, P.W. Gold, K. Hashimoto, Psychological stress in- [63] J.M.H.M. Reul, E.R. de Kloet, Two receptor systems for corticos-creased corticotropin-releasing hormone mRNA and content in the terone in rat brain: Microdistribution and differential occupation,central nucleus of the amygdala but not in the hypothalamic Endocrinology 117 (1985) 2505–2511.paraventricular nucleus in the rat, Brain Res. 850 (1999) 136–143. [64] P.E. Sawchenko, Evidence for a local site of action for glucocor-
[46] S. Makino, M.A. Smith, P.W. Gold, Increased expression of corticot- ticoids in inhibiting CRF and vasopressin expression in the paraven-ropin-releasing hormone and vasopressin messenger ribonucleic acid tricular nucleus, Brain Res. 403 (1987) 213–223.(mRNA) in the hypothalamic paraventricular nucleus during re- [65] J. Schulkin, P.W. Gold, B.S. McEwen, Induction of corticotropin-peated stress: association with reduction in glucocorticoid receptor releasing hormone gene expression by glucocorticoids: implicationmRNA levels, Endocrinology 136 (8) (1995) 3299–3309. for understanding the states of fear and anxiety and allostatic load,
[47] A. Marcilhac, P. Siaud, Regulation of the adrenocorticotrophin Psychoneuroendocrinology 23 (1998) 219–243.response to stress by the central nucleus of the amygdala in rats [66] J. Schulkin, B.S. McEwen, P.W. Gold, Allostasis, amygdala, anddepends upon the nature of the stressor, Exp. Physiol. 81 (1996) anticipatory angst, Neurosci. Biobehav. Rev. 18 (1994) 385–396.1035–1038. [67] J.D. Shepard, K.W. Barron, D.A. Myers, Corticosterone delivery to
[48] A. Marcilhac, P. Siaud, Identification of projections from the central the amygdala increases corticotropin-releasing factor mRNA in thenucleus of the amygdala to the paraventricular nucleus of the central amygdaloid nucleus and anxiety-like behavior, Brain Res.hypothalamus which are immunoreactive for corticotrophin-releas- 861 (2000) 288–295.ing hormone in the rat, Exp. Physiol. 82 (1997) 273–281. [68] L.W. Swanson, Biochemical switching in hypothalamic circuits
[49] M.J. Meaney, B.S. McEwen, Testosterone implants into the mediating responses to stress, Prog. Brain Res. 87 (1991) 181–200.amygdala during the neonatal period masculinize the social play of [69] K.V. Thrivikraman, Y. Su, P.M. Plotsky, Patterns of fos-immuno-juvenile female rats, Brain Res. 398 (1986) 324–328. reactivity in the CNS induced by repeated hemorrhage in conscious
[50] M. Morimoto, N. Morita, H. Ozawa, K. Yokoyama, M. Kawata, rats: Correlations with pituitary–adrenal axis activity, Stress 2Distribution of glucocorticoid receptor immunoreactivity and mRNA (1997) 145–158.in the rat brain: an immunohistochemical and in situ hybridizationstudy, Neurosci. Res. 26 (1996) 235–269.
Related Documents











