Starch granule initiation and growth are altered in barley mutants that lack isoamylase activity Rachel A. Burton 1 , Helen Jenner 2 , Luke Carrangis 1 Brendan Fahy 2 , Geoffrey B. Fincher 1 , Chris Hylton 2 , David A. Laurie 2 , Mary Parker 2 , Darren Waite 2 , Sonja van Wegen 1 , Tamara Verhoeven 2 and Kay Denyer 2, * 1 Department of Plant Science, University of Adelaide, Waite Campus, Glen Osmond, SA 5064, Australia 2 John Innes Centre, Norwich Research Park, Colney, Norfolk NR4 7UH, UK 3 Institute of Food Research, Norwich Research Park, Colney, Norfolk NR4 7UA, UK Received 31 October 2001; revised 18 March 2002; accepted 27 March 2002. *For correspondence (fax +44 1603 450045; e-mail [email protected]). Summary Two mutant lines of barley, Risø 17 and Notch-2, were found to accumulate phytoglycogen in the grain. Like the sugary mutants of maize and rice, these phytoglycogen-accumulating mutants of barley lack isoamylase activity in the developing endosperm. The mutants were shown to be allelic, and to have lesions in the isoamylase gene, isa1 that account for the absence of this enzyme. As well as causing a reduction in endosperm starch content, the mutations have a profound effect on the structure, number and timing of initiation of starch granules. There are no normal A-type or B-type granules in the mutants. The mutants have a greater number of starch granules per plastid than the wild-type and, particularly in Risø 17, this leads to the appearance of compound starch granules. These results suggest that, as well as suppressing phytoglycogen synthesis, isoamylase in the wild-type endosperm plays a role in determining the number, and hence the form, of starch granules. Keywords: starch, barley, granules, initiation, isoamylase, phytoglycogen. Introduction Normal starch synthesis in plants requires, in addition to starch synthases and starch-branching enzymes, a deb- ranching enzyme (DBE) that cleaves (1,6) a-linkages within amylopectin and related polysaccharides or oligosacchar- ides. Evidence that DBE is necessary for normal starch synthesis comes from the study of mutant plants and algae lacking enzymes belonging to the isoamylase class of DBE (maize, James et al., 1995; rice, Kubo et al., 1999; Arabidopsis, Zeeman et al., 1998; Chlamydomonas, Mouille et al., 1996). In these mutants, the starch content is lower than normal and there is an accumulation of a soluble (1fi4:1fi6) a-glucan, phytoglycogen. Phytogly- cogen does not accumulate in wild-type plants and algae, or in other low-starch mutants. In the isoamylase (sugary1) mutants of maize and rice, there is also a decrease in the activity of limit dextrinase (LD), another type of DBE (Nakamura et al., 1996; Pan and Nelson, 1984). It has been argued that this decrease, rather than the loss of isoamylase, may be the direct cause of phytoglycogen accumulation. There is an inverse correl- ation between LD activity and phytoglycogen accumula- tion in the endosperm of rice mutants carrying sugary1 alleles of different severity (Nakamura et al., 1997). However, LD remains at wild-type levels in the phytogly- cogen-accumulating isoamylase mutant of Arabidopsis (dbe1; Zeeman et al., 1998). Thus, it is likely that the isoamylase-type of DBE plays a specific role in the synthesis of starch that cannot be assumed by the LD-type. Two models have been put forward to explain the role of isoamylase in starch synthesis. The first proposes that isoamylase plays a direct role in the synthesis of amylopectin, the major component of starch granules (Ball et al., 1996; Myers et al., 2000). It is suggested that DBE is required for the synthesis of an amylopectin molecule capable of crystallization from a soluble pre- amylopectin precursor. In the absence of isoamylase, pre- amylopectin is further elaborated by starch synthase and starch-branching enzymes in the stroma to form phytogly- cogen. The second model (Zeeman et al., 1998) proposes that isoamylase does not play a direct role in the synthesis of amylopectin. Instead, together with other degradative enzymes, isoamylase degrades soluble a-glucans. In the The Plant Journal (2002) 31(1), 97–112 ª 2002 Blackwell Science Ltd 97

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Starch granule initiation and growth are altered in barleymutants that lack isoamylase activity
Rachel A. Burton1, Helen Jenner2, Luke Carrangis1 Brendan Fahy2, Geoffrey B. Fincher1, Chris Hylton2, David A. Laurie2,
Mary Parker2, Darren Waite2, Sonja van Wegen1, Tamara Verhoeven2 and Kay Denyer2,*1Department of Plant Science, University of Adelaide, Waite Campus, Glen Osmond, SA 5064, Australia2John Innes Centre, Norwich Research Park, Colney, Norfolk NR4 7UH, UK3Institute of Food Research, Norwich Research Park, Colney, Norfolk NR4 7UA, UK
Received 31 October 2001; revised 18 March 2002; accepted 27 March 2002.*For correspondence (fax +44 1603 450045; e-mail [email protected]).
Summary
Two mutant lines of barley, Risù 17 and Notch-2, were found to accumulate phytoglycogen in the grain.
Like the sugary mutants of maize and rice, these phytoglycogen-accumulating mutants of barley lack
isoamylase activity in the developing endosperm. The mutants were shown to be allelic, and to have
lesions in the isoamylase gene, isa1 that account for the absence of this enzyme. As well as causing a
reduction in endosperm starch content, the mutations have a profound effect on the structure, number
and timing of initiation of starch granules. There are no normal A-type or B-type granules in the
mutants. The mutants have a greater number of starch granules per plastid than the wild-type and,
particularly in Risù 17, this leads to the appearance of compound starch granules. These results suggest
that, as well as suppressing phytoglycogen synthesis, isoamylase in the wild-type endosperm plays a
role in determining the number, and hence the form, of starch granules.
Keywords: starch, barley, granules, initiation, isoamylase, phytoglycogen.
Introduction
Normal starch synthesis in plants requires, in addition to
starch synthases and starch-branching enzymes, a deb-
ranching enzyme (DBE) that cleaves (1,6) a-linkages within
amylopectin and related polysaccharides or oligosacchar-
ides. Evidence that DBE is necessary for normal starch
synthesis comes from the study of mutant plants and
algae lacking enzymes belonging to the isoamylase class
of DBE (maize, James et al., 1995; rice, Kubo et al., 1999;
Arabidopsis, Zeeman et al., 1998; Chlamydomonas,
Mouille et al., 1996). In these mutants, the starch content
is lower than normal and there is an accumulation of a
soluble (1®4 : 1®6) a-glucan, phytoglycogen. Phytogly-
cogen does not accumulate in wild-type plants and algae,
or in other low-starch mutants.
In the isoamylase (sugary1) mutants of maize and rice,
there is also a decrease in the activity of limit dextrinase
(LD), another type of DBE (Nakamura et al., 1996; Pan and
Nelson, 1984). It has been argued that this decrease, rather
than the loss of isoamylase, may be the direct cause of
phytoglycogen accumulation. There is an inverse correl-
ation between LD activity and phytoglycogen accumula-
tion in the endosperm of rice mutants carrying sugary1
alleles of different severity (Nakamura et al., 1997).
However, LD remains at wild-type levels in the phytogly-
cogen-accumulating isoamylase mutant of Arabidopsis
(dbe1; Zeeman et al., 1998). Thus, it is likely that the
isoamylase-type of DBE plays a speci®c role in the
synthesis of starch that cannot be assumed by the LD-type.
Two models have been put forward to explain the role of
isoamylase in starch synthesis. The ®rst proposes that
isoamylase plays a direct role in the synthesis of
amylopectin, the major component of starch granules
(Ball et al., 1996; Myers et al., 2000). It is suggested that
DBE is required for the synthesis of an amylopectin
molecule capable of crystallization from a soluble pre-
amylopectin precursor. In the absence of isoamylase, pre-
amylopectin is further elaborated by starch synthase and
starch-branching enzymes in the stroma to form phytogly-
cogen. The second model (Zeeman et al., 1998) proposes
that isoamylase does not play a direct role in the synthesis
of amylopectin. Instead, together with other degradative
enzymes, isoamylase degrades soluble a-glucans. In the
The Plant Journal (2002) 31(1), 97±112
ã 2002 Blackwell Science Ltd 97

absence of isoamylase, these accumulate in the form of
phytoglycogen along with reduced amounts of normal
amylopectin.
The phenotypes of the isoamylase mutants so far
described do not provide suf®cient information to allow
these models to be further evaluated. It is not clear which,
if either is correct. To investigate further the role of
isoamylase in starch synthesis, we identi®ed two allelic
isoamylase mutants of barley, Risù 17 and Notch-2, during
a screen for altered starch synthesis in the endosperm of
high-lysine mutants of barley. Detailed characterization of
starch granule structure and the sequence of the isoamy-
lase genes of these mutants have shed light on the
possible role of isoamylase during starch synthesis.
Results
Two low-starch barley mutants accumulate large
amounts of soluble a-glucan
The starch and soluble a-glucan contents of mature grains
of two wild-type barley cultivars (Bomi and Carlsberg II)
and several previously identi®ed low-starch mutants
(Balaravi et al., 1976; Bansal, 1970; Doll, 1983) were meas-
ured (Figure 1). Soluble a-glucan is material that is soluble
in aqueous extraction medium but insoluble in > 60%
aqueous methanol. Two of the low-starch mutants, Risù 17
and Notch-2, had much higher soluble a-glucan contents
than the wild-type lines and the other mutants.
The degree of branching of the a-glucans in the mature
grains of wild-type (Bomi) and mutants (Risù 17 and
Notch-2) was compared by measuring the wavelengths of
maximum absorbance (lmax) of the a-glucan-iodine
complexes. All of the starches had lmax of 550±600 nm,
which is typical of starches generally. The lmax of the
soluble a-glucan from Bomi was also 550±600 nm sug-
gesting that it had a degree of branching similar to starch.
The soluble a-glucans from the mutants had lmax values
of 420±430 nm, which indicated that they were more
highly branched than starch. These values are in the
same range as those of phytoglycogen from maize and
Arabidopsis (Zeeman et al., 1998), suggesting strongly that
the barley mutants accumulate phytoglycogen.
The soluble a-glucan in the mutants is phytoglycogen
To examine the structure of the soluble a-glucan in more
detail and to compare the starches from wild-type and
mutants, we determined the relative abundance of chains
of different lengths in these a-glucans using ¯uorophore-
assisted gel electrophoresis (O'Shea and Morell, 1996).
Starch and soluble a-glucan were extracted from endo-
sperms at ®ve different stages of development. For starch,
the pro®les obtained re¯ect the distributions of the short-
to medium-length chains of the amylopectin component of
starch.
There was little alteration in the chain-length pro®les of
the starches with developmental age, for either the wild-
type or the mutants (Figure 2a, i-iii). The mutant starches
had chain-length pro®les similar to those of the wild-type
starches except that mutant starches from older and
mature endosperms had relatively more chains of dp 10±
13 (Figure 2b, i-ii).
The chain-length pro®les of the soluble a-glucan from
the barley mutants were similar to those of phytoglycogen
from other species (e.g. maize, Dinges et al., 2001; rice,
Nakamura et al., 1997; Arabidopsis, Zeeman et al., 1998).
There was an increase in the proportion of short chains (dp
6±9) with age (Figure 2a, v-vi and Figure 2b, iii-iv). At all
stages of development, the soluble a-glucans from the
mutants had a greater proportion of chains of dp 6±8
(Figure 2b, v-vi) and, particularly in Notch-2, fewer chains
of dp 10±15 than their respective starches (Figure 2b, v-vi).
Risù 17 and Notch-2 are allelic variants
To determine whether the recessive mutations in Risù 17
and Notch-2 were in the same gene, we crossed them and
examined the F1 progeny. All of the F1 grains were
shrivelled, indicating that they had a lower than normal
starch content, and the starch granules were irregularly
shaped and small. Risù 17 and Notch-2 were crossed with
other low-starch cultivars (Risù 13, Risù 16, Risù 527, Risù
1508, Notch-1; Doll, 1983). These crosses gave F1 grains
that were all, or mostly all, normal in shape. The starch
Figure 1. Starch and soluble a-glucan contents of mature grains.a-Glucans were extracted from mature grains by homogenization inwater. Starch was puri®ed from the water-insoluble fraction and solublea-glucan by alcohol precipitation of the water-soluble fraction. Soluble a-glucan is material which is soluble in aqueous extraction medium butinsoluble in > 60% aqueous methanol. Data are means 6 SD of valuesfrom 3 to 4 separate extracts. Bomi and Carlsberg II are wild-type withrespect to their starch contents. The other cultivars were previouslyidenti®ed as low-starch mutants.
98 Rachel A. Burton et al.
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

granules in grains resulting from the cross between Risù
17 and Notch-1 were normal in shape. These results
indicate ®rstly, that the mutations in Risù 17 and Notch-2
lie in the same gene and secondly, that none of the other
low-starch cultivars carry mutations that are allelic to those
in Risù 17 or Notch-2.
Risù 17 and Notch-2 lack isoamylase activity
Isoamylase activity cannot be quanti®ed in crude extracts
due to the absence of a unique substrate for this enzyme.
However, isoamylase activity in extracts of several plant
species is revealed on native, non-denaturing polyacryla-
mide gels containing amylopectin. The enzyme appears as
a blue-staining band with low mobility when the gels are
stained with Lugol solution (Dinges et al., 2001; Kubo et al.,
1999; Zeeman et al., 1998). Such a band was observed with
extracts of developing wild-type barley endosperms of
different developmental ages (Figure 3 and data not
shown). A second, faint blue-staining band was some-
times observed in wild-type extracts (not visible in
Figure 3). Neither blue band was observed with extracts
of endosperms of any age for either Risù 17 or Notch-2. To
estimate the minimum isoamylase activity that could be
detected using this native gel method, we compared a
series of dilutions of the wild-type extract on native gels
(not shown). From this, we estimated that the isoamylase
activity in the mutants was < 4% of that in the wild-type.
The activities of several other enzymes are affected in the
mutants
The activities of many of the enzymes involved in the
conversion of sucrose to starch were determined in
endosperms from Risù 17 and its parent variety, Bomi
and in Notch-2 and the variety from which it was derived,
NP113. When the activities in grains of 45±55 mg FW in the
mutants and their corresponding wild-types were com-
pared, the activities of many enzymes were not statistically
signi®cantly different (Table 1). Some enzymes (Table 1)
did show differences but these were not consistently
different in both mutants. For example, the activity of
soluble starch synthase was higher in Risù 17 than in Bomi
but there was no signi®cant difference in the activity of this
enzyme between NP113 and Notch-2. Most of the enzymes
that differed in activity showed a higher activity in the
mutant than in the wild-type. These increases in activities
are unlikely to cause or contribute to the decrease in total
a-glucan synthesis observed in the mutants. Two
enzymes, SBE and alkaline pyrophosphatase, showed
lower activity in Risù 17 than in Bomi. These data suggest
that the mutations cause pleiotropic effects on the activ-
ities of other enzymes in the pathway of starch synthesis.
Such pleiotropic effects are common in starch mutants of
cereals (maize, Singletary et al., 1997; barley, Schulman
and Ahokas, 1990).
Attempts to assay limit dextrinase (LD) activity in crude
extracts of developing barley endosperm were unsuccess-
ful due to the presence of protein inhibitors (Macri et al.,
1993). The activity of LD measured in mixed extracts of
developing pea embryo and developing barley endo-
sperms was much less than that expected from measure-
ments of the LD activity in these tissues extracted
separately (data not shown). Inclusion of a chemical
modi®er of the LD inhibitor, phenyl glyoxal (MacGregor
et al., 2000) did not increase the measurable LD activity in
extracts of barley. As an alternative approach to estimate
the LD activity, we used native gels similar to those
described above to identify isoamylase but containing red-
pullulan rather than amylopectin. Red-pullulan is a unique
substrate for LD, and is not degraded by isoamylase or any
other enzymes. Two bands of LD activity were observed
(data not shown). Comparison of the LD activity in extracts
of mutant and wild-type developing endosperms did not
reveal any consistent differences. This suggested that in
the barley isoamylase mutants, unlike those of rice and
maize, the LD activity is not decreased relative to that in the
wild-type. However, this conclusion must be treated with
some caution as we do not know to what extent the LD
inhibitors affect the LD activity revealed in these gels.
Cloning and sequencing the isoamylase cDNA
To discover whether the lack of isoamylase activity was
due to mutations within the isoamylase gene, PCR was
used to amplify the cDNAs encoding isoamylase in the
wild-types and mutants. A nested PCR strategy, using the
high-®delity Taq polymerase Elongase, was used to obtain
full-length isoamylase cDNAs. The cDNA sequences have
been submitted to GenBank under accession numbers
AF490375 (Bomi), AF490376 (Notch-2) and AF490377 (Risù
17), and the predicted amino acid sequences are shown in
Figure 4a. This is the ®rst published report of the full-
length sequence of barley isoamylase cDNA; the sequence
previously reported by Sun et al. (1999) is truncated at both
the 5¢ and 3¢ ends. We will refer to this cDNA as isa1.
The isa1 cDNAs from the wild-types, Bomi and NP113,
are almost identical and very similar to isoamylase cDNAs
from other cereals (data not shown). The isa1 cDNAs from
both mutants differ from the wild-type isa1 cDNAs. That
from Risù 17 contains an 872-bp deletion, starting at amino
acid 338 (Figure 4a), whilst that from Notch-2 contains a
72-bp insertion. The insertion in the Notch-2 cDNA
includes two in-frame stop codons (Figure 4a). A
BLASTN search with the 72 bp sequence reveals that it is
in the same position and has 90% sequence identity with
intron 9 of a DBE gene from Aegilops tauchii (Figure 4b).
This suggests that the insertion in Notch-2 is an intron that
Isoamylase mutants of barley 99
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

is either not removed or is incorrectly spliced out of the
mRNA, possibly due to a single base change (G to A) at the
5¢ intron splice junction (Figure 4b). A thorough investiga-
tion of the Notch-2 cDNA population revealed a number of
other isoamylase cDNAs, for example, Notch-2A and
Notch-2B (Figure 4a) in which the intron appears to have
been mis-spliced. The mis-splicing in both Notch-2A and B
results in the removal of two amino acids and causes
frameshifts that result in the introduction of downstream
stop codons. No full-length, wild-type cDNAs were found
in Notch-2 or Risù 17.
The relative amounts of isa1 transcript in developing
endosperms of the wild-types and mutants were com-
pared by Northern analysis (Figure 4c). Using this tech-
nique, the transcripts in the mutants were apparently
absent. However, as transcripts were detected in the
mutants using the more sensitive PCR technique, we
assume that a very low level of isa1 transcript is present in
Figure 2a.
100 Rachel A. Burton et al.
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

the mutants but that it is below the level of detection in the
Northern analysis.
The isoamylase gene is located on chromosome 7H
The mutation responsible for the low-starch phenotype of
Risù 17 was previously assigned to barley chromosome 7H
using a set of translocation lines (Jensen, 1979).
We mapped the isoamylase gene, isa1 by RFLP in a
population of 86 doubled haploid (DH) lines from a
Chebec 3 Harrington cross. A single polymorphism
between restriction enzyme digests of the parental lines
was identi®ed. Scoring this polymorphism in the mapping
population con®rmed the chromosome 7H location
Figure 2b.
Figure 2. Analysis by gel electrophoresis of the short chains of a-glucans.a-Glucans were extracted from grains of different developmental ages. The stages of development were de®ned by the FWs of the grains as follows: s,> 20 mg; h, 20±40 mg; n, 40±70 mg; ,, 70±100 mg; e, mature grains. a-Glucan samples were debranched with isoamylase, derivatized with the¯uorophore APTS and subjected to electrophoresis in an Applied Biosystems DNA sequencer. Data were analysed using Genescan software.(a) Analysis of chain-length pro®les. The sum of the areas of peaks corresponding to individual chain lengths between 6 and 28 glucose units wascalculated and each peak area was expressed as a percentage of the total peak area. Each value is the mean of two replicate measurements. (i-iii) starch;(iv-vi) soluble a-glucan.(b) Comparison of chain-length pro®les. To compare different a-glucans, for each chain length, the difference between values for the percentage total peakarea was calculated.
Isoamylase mutants of barley 101
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

(Figure 4d) and reference to markers listed in Langridge
et al. (1995) placed the gene genetically close to the
centromere on the short arm. The gene was also mapped
in a population of 90 DH lines from a Galleon 3 Haruna
Nijo cross (Langridge et al., 1995) which gave an equiva-
lent map location (data not shown). The limit dextrinase
gene is also located on chromosome 7H (Figure 4d; R.A.
Burton and G.B. Fincher, unpublished data).
The mutants have radically altered starch-granule
architecture
Scanning electron microscopy of mature grains (Figure 5a)
showed that the endosperm of the wild-type barley, Bomi,
contained the A- and B-type starch granules typical of the
Triticeae family (Jane et al., 1994). As observed previously
(Burgess et al., 1982; Gautam et al., 1994; Sood et al., 1992;
Tester et al., 1993), the granules in the mutants were
completely different in appearance to those in the wild-
type. They were irregular in shape and intermediate in size
compared with the A- and B-type granules in Bomi.
Some of the granules in Risù 17 resembled the com-
pound granules of rice and oat endosperm. In these Risù
17 granules, a number of irregular granulae were closely
oppressed to form a smooth compound granule
(Figure 5a). Compound granules were previously
observed in the developing endosperms of Notch-2
(Sood et al., 1992). These observations and our own of
the young developing endosperm of Risù 17 (Figure 5b),
suggested that the individual components of the com-
pound granules in these mutants were initiated separately
within the plastid. Later, these component granules grew
together to ®ll the available space and therefore became
irregular in shape. The granules in these mutants do not
appear to arise as single granules which later fracture into
Figure 3. Isoforms of debranching enzymes in developing endosperms.Crude, soluble extracts of developing endosperms were loaded onto gelscontaining amylopectin. Each track contained extract (from 1 mg FW oftissue) and loading medium in a ratio of 5 : 1. After electrophoresis, gelswere incubated at 37°C for 16 h at pH 6.0 and stained with iodinesolution. The position of the blue-staining band that is due to isoamylaseactivity is indicated with an arrow. Tracks 1 and 2: Bomi, tracks 3 and 4:Risù 17, tracks 5 and 6: NP113, tracks 7 and 8: Notch-2.
Table 1. Comparison of the maximum catalytic activities of enzymes in crude extracts of developing endosperms
Enzyme
Activity (mmol min±1 g±1FW)
Bomi Risù 17 NP113 Notch-2
Sucrose synthase 3.39 6 0.16 [8] 3.79 6 0.53 [5] 2.81 6 0.72 [4] 3.80 6 0.59 [5]UDPG pyrophosphorylase 54.13 6 4.68 [7] 68.87 6 8.51 [5] 41.04 6 5.10 [5] 55.53 6 11.91 [5]Fructokinase 0.245 6 0.022 [8] 0.311 6 0.039 [5] 0.182 6 0.021 [5] 0.291 6 0.026 [5]Glucokinase 0.435 6 0.050 [8] 0.523 6 0.056 [5] 0.386 6 0.034 [5] 0.612 6 0.054 [5]Phosphoglucomutase 19.02 6 3.92 [8] 38.35 6 10.40 [6] 27.51 6 7.87 [6] 28.37 6 6.94 [6]Phosphoglucose isomerase 19.61 6 2.26 [8] 21.40 6 1.71 [4] 18.68 6 0.79 [5] 22.83 6 2.88 [5]ADPG pyrophosphorylase 4.44 6 0.37 [8] 6.88 6 0.97 [6] 3.72 6 0.28 [5] 5.25 6 0.78 [6]Soluble starch synthase 0.216 6 0.020 [8] 0.399 6 0.026 [4] 0.131 6 0.025 [5] 0.118 6 0.037 [4]Granule-bound starch synthase 0.387 6 0.048 [8] 0.373 6 0.069 [4] 0.252 6 0.051 [4] 0.222 6 0.074 [3]Alkaline pyrophosphatase 7.34 6 0.94 [8] 4.52 6 0.64 [5] 3.68 6 0.57 [5] 3.58 6 0.64 [5]Starch-branching enzyme 42.93 6 2.62 [6] 31.54 6 0.78 [5] ND ND
Developing endosperms from grains of 45±55 mg FW were extracted as described in Experimental procedures and assayed for enzymeactivities. Values are means 6 SE of measurements made on a number of independent extracts (shown in parentheses). Comparison of theactivities of each enzyme in Bomi and Risù 17 and in NP113 and Notch-2 was done using Microsoft Excel software (t-test, 2-taileddistribution, 2-sample equal variance). This statistical analysis showed that the activities of soluble starch synthase and ADPGpyrophosphorylase were higher in Risù 17 than in Bomi (P < 0.05), the activities of fructokinase and glucokinase were higher in Notch-2than in NP113 (P < 0.05) and the activities of SBE and alkaline pyrophosphatase were lower in Risù 17 than in Bomi (P = 0.05). All other pair-wise comparisons showed no statistically signi®cant difference (P > 0.05). ND = not determined.
102 Rachel A. Burton et al.
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

multiple, irregular pieces as in developing embryos of the
starch-branching enzyme mutant of pea (rr, Lloyd, 1995).
Endosperm cells contain different amounts of
phytoglycogen
To investigate the distribution of starch and phytoglyco-
gen within endosperms of barley, slices of endosperm
were cut from developing grains, ®xed and embedded in
resin. Thin sections were cut, stained and examined with
either a light microscope (Figure 6, left panels) or a
transmission electron microscope (TEM; Figure 6, right
panels).
As well as the small, irregular granules in the mutants
(Figure 6b-f), some plastids in cells of the mutant endo-
sperms contained a diffuse material that stained lightly
with toluidine blue (Figure 6b-d). The TEM pictures
showed that this material was particulate (Figure 6e,g,h).
This is likely to be phytoglycogen as it resembled similar
material seen in the leaves of isoamylase mutants of
Arabidopsis (Zeeman et al., 1998) and in sections of
developing maize endosperm from sugary1 mutant plants
prepared using the same procedures used for the barley
endosperm sections shown in Figure 6 (M. James and M.
Parker, Iowa State University and IFR Norwich, respect-
ively, personal communication).
In the endosperm of older Risù 17 grains and in both
young and older Notch-2 endosperm, most cells contained
phytoglycogen (not shown). In many plastids, the starch
granules had sharp, well-de®ned edges (e.g. Figure 6f).
However, plastids containing large amounts of phytogly-
cogen as well as starch often had granules with irregular,
diffuse edges (Figure 6c,g). At high magni®cation
(Figure 6h), it was evident that the boundary between the
granules and phytoglycogen was not well de®ned.
The phytoglycogen content of adjacent cells was vari-
able, particularly in young endosperm of Risù 17
(Figure 6b,c). Many cells contained little or no phytoglyco-
gen. The phytoglycogen-containing cells were randomly
dispersed throughout the endosperm. Within a cell, there
was variation between plastids in phytoglycogen content.
Some plastids contained large amounts of phytoglycogen
and small amounts of starch whilst other plastids con-
tained more starch and relatively little phytoglycogen
(Figure 6e). To investigate whether this could have been
due to loss of phytoglycogen during ®xation or artefacts
due to the staining procedure, we compared endosperm
sections of the maize mutant sugary1 (kindly provided by
M. James and M. Parker) with barley sections that were
prepared and stained in the same way. In the maize
sections, there was more phytoglycogen and less starch
than in the barley sections. This is consistent with meas-
urements of the phytoglycogen and starch content of
sugary1 maize endosperm (55% and 14% (w/w), respect-
ively; Dinges et al., 2001). There was also less heterogen-
eity between cells and between plastids within cells in the
amounts of these materials in maize compared to barley.
This suggested that the lack of phytoglycogen in some of
the cells and plastids of the barley endosperm was not
likely to be due to loss during preparation but to real cell-
to-cell variation in phytoglycogen and starch content. The
underlying cause of this variation is not understood.
There is a single wave of granule initiation in the
mutants
To discover whether the total number of granules in the
endosperm as a whole was altered in the isoamylase
mutants, starch was extracted from endosperms of differ-
ent ages and the numbers of granules in samples of these
were estimated using a haemocytometer (Shannon et al.,
1996). In wild-type barley endosperms, A-type granules
initiate at approximately 5±10 days after anthesis (DAA)
followed approximately 10 days later by a second wave of
granule initiation that gives rise to the B-type granules.
Our measurements of granule number in Bomi show these
two waves of initiation (Figure 7). Before 20 DAA, almost
all of the granules were large and disc-shaped, which is
typical of A-type granules. After 20 DAA, there was a
sudden increase in granule number and we observed
small, spherical B-type granules as well as the larger, A-
type granules. In the mutant Risù 17, the number of
granules per endosperm was higher than in wild-type
endosperms before 20 DAA, showing that many more
granules initiate in the young endosperm of the mutant
than in the wild-type. The mean of the values shown in
Figure 7 for endosperms less than 20 DAA was
109.6 6 15.3 million (mean 6 SE, n = 11) for Risù 17 and
19.4 6 3.8 million (mean 6 SE, n = 11) for Bomi. However,
in the mutant, there was no second wave of initiation.
Thus, the total number of granules per endosperm in the
mutant in the later half of development (after 20 DAA) was
similar to that in the endosperm of the wild-type. A
second, independent experiment on a separately grown
batch of plants gave results that showed the same trends
as those shown in Figure 7.
Discussion
In many respects, the Risù 17 and Notch-2 mutants are
similar to other cereal isoamylase mutants. They lack
isoamylase activity and have a reduced starch content, an
increased sugar content, an altered storage-protein com-
position and shrivelled grains at maturity, and they
accumulate phytoglycogen (our data and those of
Bansal, 1970; Doll, 1983; Sood et al., 1992).
The cDNA sequences con®rmed that Risù 17 and Notch-
2 have mutations in the isoamylase gene, isa1 that abolish
Isoamylase mutants of barley 103
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

function and are very likely to be responsible for the
phenotype. The gene from Risù 17 has a 872-bp deletion in
the coding region. In Notch-2, there is a single-base
substitution in a 5¢-intron splice consensus sequence
(converting GT to AT) that appears to interfere with the
normal splicing of an intron from the primary mRNA
transcript. As a result, the cDNA from Notch-2 has either a
72-bp insertion containing two in-frame stop codons or a
downstream stop codon introduced by a mis-splicing
event. In addition, the abundance of isa1 transcripts is
severely reduced in the mutants compared to the wild-
types. This could be due to the fact that the mutant
transcripts are unstable or are recognized by the plant as
aberrant and are therefore rapidly turned over.
As well as causing a decrease in starch content and an
accumulation of phytoglycogen, the mutations have a
profound effect on the structure, number and timing of
initiation of starch granules. As observed previously, in
both mutants, granules are smaller than A-type granules
from the wild-type and are irregularly shaped (Burgess
et al., 1982; Gautam et al., 1994; Sood et al., 1992; Tester
et al., 1993). In Risù 17, a single wave of granule initiation
occurs at the time in endosperm development when A-
type granules initiate in the wild-type. More than one
granule initiates per plastid in the mutant and these pack
together to form compound granules resembling those
found in normal rice and oat grains. In the early stages of
endosperm development (< 20 DAA), the total number of
Figure 4a.
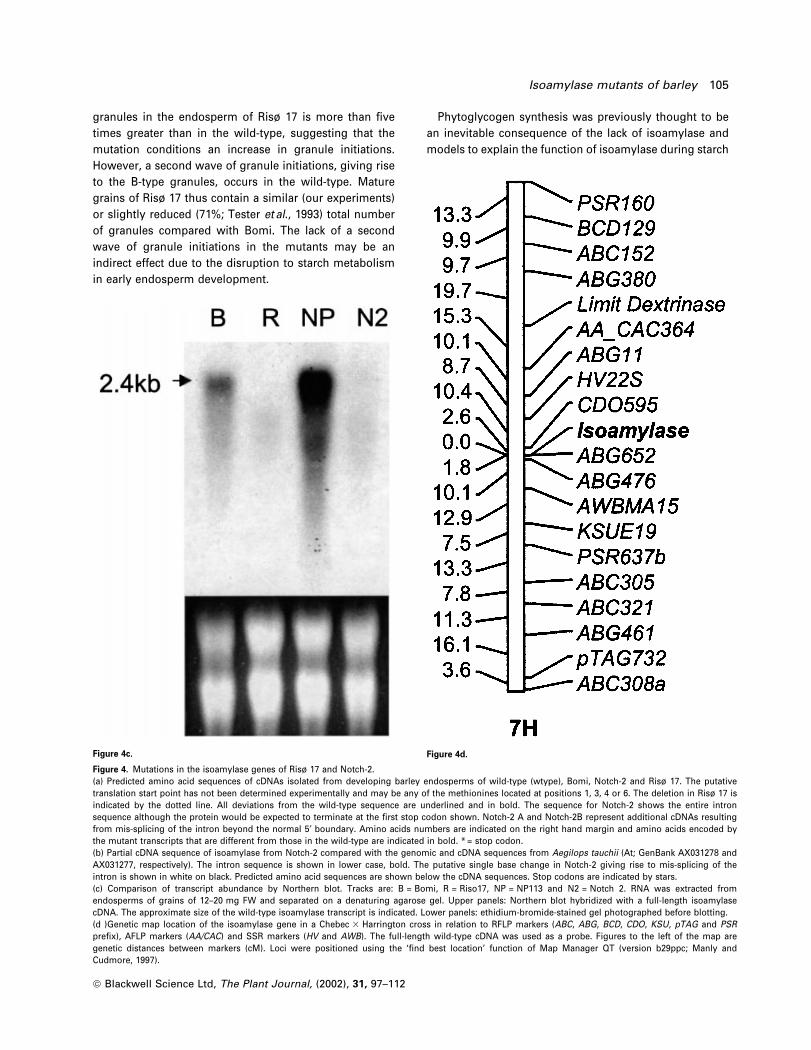
Figure 4b.
104 Rachel A. Burton et al.
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112
granules in the endosperm of Risù 17 is more than ®ve
times greater than in the wild-type, suggesting that the
mutation conditions an increase in granule initiations.
However, a second wave of granule initiations, giving rise
to the B-type granules, occurs in the wild-type. Mature
grains of Risù 17 thus contain a similar (our experiments)
or slightly reduced (71%; Tester et al., 1993) total number
of granules compared with Bomi. The lack of a second
wave of granule initiations in the mutants may be an
indirect effect due to the disruption to starch metabolism
in early endosperm development.
Phytoglycogen synthesis was previously thought to be
an inevitable consequence of the lack of isoamylase and
models to explain the function of isoamylase during starch
Figure 4c. Figure 4d.
Figure 4. Mutations in the isoamylase genes of Risù 17 and Notch-2.(a) Predicted amino acid sequences of cDNAs isolated from developing barley endosperms of wild-type (wtype), Bomi, Notch-2 and Risù 17. The putativetranslation start point has not been determined experimentally and may be any of the methionines located at positions 1, 3, 4 or 6. The deletion in Risù 17 isindicated by the dotted line. All deviations from the wild-type sequence are underlined and in bold. The sequence for Notch-2 shows the entire intronsequence although the protein would be expected to terminate at the ®rst stop codon shown. Notch-2 A and Notch-2B represent additional cDNAs resultingfrom mis-splicing of the intron beyond the normal 5¢ boundary. Amino acids numbers are indicated on the right hand margin and amino acids encoded bythe mutant transcripts that are different from those in the wild-type are indicated in bold. * = stop codon.(b) Partial cDNA sequence of isoamylase from Notch-2 compared with the genomic and cDNA sequences from Aegilops tauchii (At; GenBank AX031278 andAX031277, respectively). The intron sequence is shown in lower case, bold. The putative single base change in Notch-2 giving rise to mis-splicing of theintron is shown in white on black. Predicted amino acid sequences are shown below the cDNA sequences. Stop codons are indicated by stars.(c) Comparison of transcript abundance by Northern blot. Tracks are: B = Bomi, R = Riso17, NP = NP113 and N2 = Notch 2. RNA was extracted fromendosperms of grains of 12±20 mg FW and separated on a denaturing agarose gel. Upper panels: Northern blot hybridized with a full-length isoamylasecDNA. The approximate size of the wild-type isoamylase transcript is indicated. Lower panels: ethidium-bromide-stained gel photographed before blotting.(d )Genetic map location of the isoamylase gene in a Chebec 3 Harrington cross in relation to RFLP markers (ABC, ABG, BCD, CDO, KSU, pTAG and PSRpre®x), AFLP markers (AA/CAC) and SSR markers (HV and AWB). The full-length wild-type cDNA was used as a probe. Figures to the left of the map aregenetic distances between markers (cM). Loci were positioned using the `®nd best location' function of Map Manager QT (version b29ppc; Manly andCudmore, 1997).
Isoamylase mutants of barley 105
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

synthesis have, at their core, ideas to explain the produc-
tion of this polymer. However, the present study of the
barley isoamylase mutants reveals a different picture.
Firstly, the fact that phytoglycogen does not accumulate in
all endosperm cells shows that loss of isoamylase activity
does not necessarily result in phytoglycogen synthesis.
Secondly, the initiation of abnormally large numbers of
granules early in endosperm development in all cells of
the mutants suggests that isoamylase plays a fundamental
role in granule initiation.
There is other evidence to suggest that the lack of
isoamylase affects starch synthesis without necessarily
leading to the production of phytoglycogen. Starch and
phytoglycogen accumulation was studied in developing
endosperms of maize containing 0±3 doses of the sugary1
mutant allele (Singletary et al., 1997). A statistically
signi®cant decrease in starch content was measured in
endosperms with 2 and 3 doses of the mutant allele. At
maturity, these endosperms had 71% and 57%, respect-
ively, of the starch content per endosperm of the wild-type.
However, phytoglycogen accumulation was only observed
in endosperms containing three doses of the mutant allele.
An effect of the lack of isoamylase activity on starch
granule number and/or shape has been reported for
sugary1 mutants of maize and rice. Unlike normal maize
endosperm which has simple granules, the granules in the
sugary1 mutant of maize are compound (Boyer et al.,
1977). In the sugary1 mutant of rice, numerous small
granules not observed in the wild-type are present in
addition to the normal compound granules (Kubo et al.,
1999). This implies that in maize and rice, as in barley,
there is an increase in the number of granule initiations per
plastid in mutants that lack isoamylase.
We have two suggestions for possible mechanisms
through which isoamylase activity might suppress granule
initiation. Firstly, the number of granules that are initiated
in a plastid might be determined by the concentration and/
or structure of soluble a-glucan in the stroma at the onset
of starch synthesis. High concentrations of soluble a-
glucans with a chain-length pro®le conducive to crystal-
lization may favour more nucleation events leading to a
greater number of granules. Zeeman et al. (1998) sug-
gested that isoamylase, together with other starch-degrad-
ing enzymes, might act to reduce the synthesis of soluble
a-glucans during the growth of the granule. Similarly,
isoamylase might contribute to the destruction of soluble
a-glucans at the time of granule initiation, thus limiting the
number of nucleation events leading to granules as well as
later, inhibiting the synthesis of phytoglycogen. In the
Figure 5. Scanning electron microscopy of barley grains.Mature (a) and frozen immature (b) grains were fractured to reveal thestarch granules within endosperm cells and viewed in a SEM. Themagni®cation is indicated by the scale bar. Note that panels are atdifferent magni®cations. Labels are A-type starch granules (A), B-typestarch granules (B), compound starch granule (cs), mutant starchgranules (s), plastid stroma (ps).
106 Rachel A. Burton et al.
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112
absence of isoamylase, more soluble a-glucans capable of
crystallization may accumulate in the stroma and this may
favour more nucleation events and hence, initiation of
more than the usual number of granules.
Secondly, isoamylase may destroy a speci®c primer
required for granule initiation. In animals and yeast, a self-
glucosylated protein primer, glycogenin, is required for the
synthesis of glycogen particles. Glycogen synthase cannot
elongate small oligosaccharides unless these are attached
to glycogenin (Alonso et al., 1995). Isoamylase might act
on some glycogenin-like protein required for granule or
polymer initiation in plants, preventing the initiation of
large numbers of granules and phytoglycogen particles by
cleaving off the associated a-glucan chains. Isoamylase
from Pseudomonas is able, in vitro, to cleave the a-glucan
chain from a primed glycogenin molecule (Lomarko et al.,
1992). Limit dextrinase, the other form of starch debranch-
ing enzyme found in plants, cannot cleave this protein-
glucan complex (Lomarko et al., 1992).
The role of glycogenin-like proteins in starch synthesis is
not established. Starch synthases can elongate small
malto-oligosaccharides and so it is not necessary to
postulate the presence of a priming protein for polymer
synthesis. However, plant genes encoding glycogenin-like
proteins have been reported (e.g. Roach and Skurat, 1997)
and various self-glycosylating proteins have been identi-
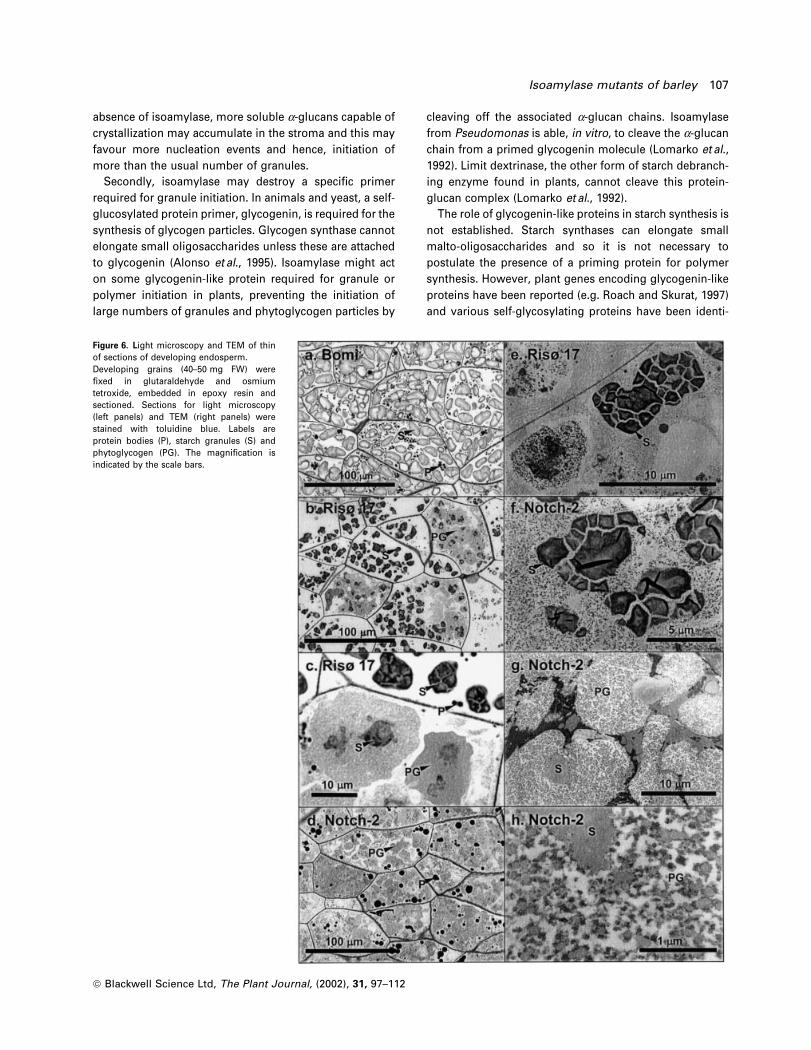
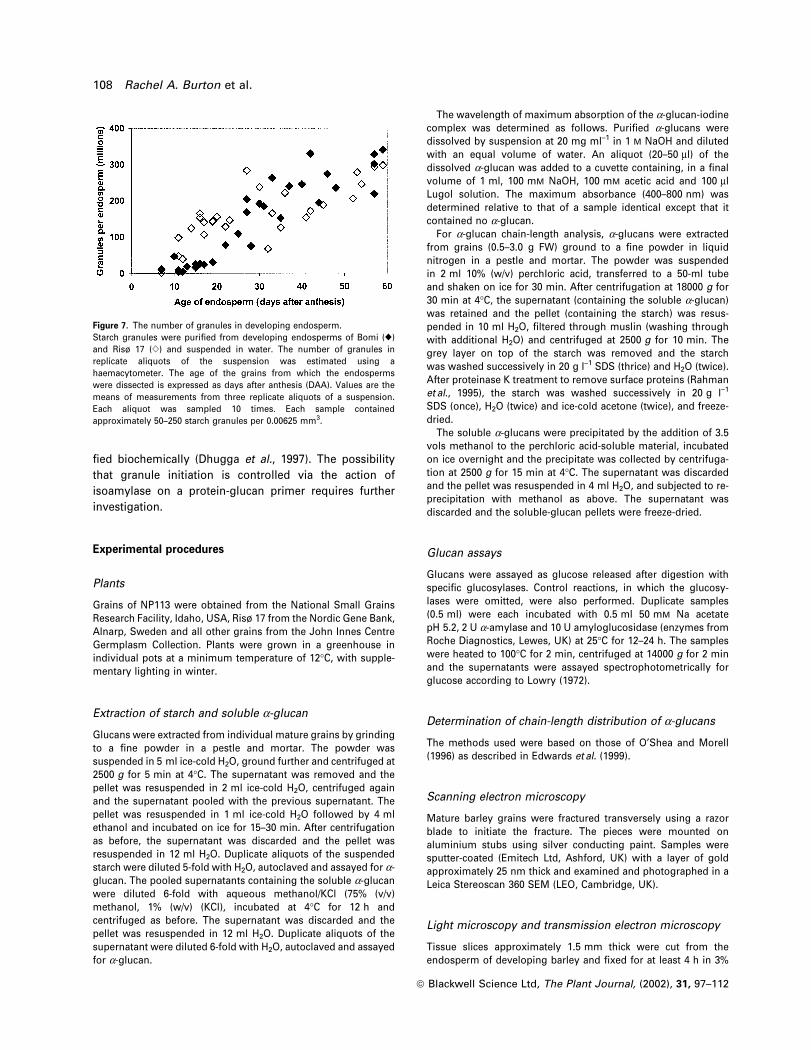
Figure 6. Light microscopy and TEM of thinof sections of developing endosperm.Developing grains (40±50 mg FW) were®xed in glutaraldehyde and osmiumtetroxide, embedded in epoxy resin andsectioned. Sections for light microscopy(left panels) and TEM (right panels) werestained with toluidine blue. Labels areprotein bodies (P), starch granules (S) andphytoglycogen (PG). The magni®cation isindicated by the scale bars.
Isoamylase mutants of barley 107
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112
®ed biochemically (Dhugga et al., 1997). The possibility
that granule initiation is controlled via the action of
isoamylase on a protein-glucan primer requires further
investigation.
Experimental procedures
Plants
Grains of NP113 were obtained from the National Small GrainsResearch Facility, Idaho, USA, Risù 17 from the Nordic Gene Bank,Alnarp, Sweden and all other grains from the John Innes CentreGermplasm Collection. Plants were grown in a greenhouse inindividual pots at a minimum temperature of 12°C, with supple-mentary lighting in winter.
Extraction of starch and soluble a-glucan
Glucans were extracted from individual mature grains by grindingto a ®ne powder in a pestle and mortar. The powder wassuspended in 5 ml ice-cold H2O, ground further and centrifuged at2500 g for 5 min at 4°C. The supernatant was removed and thepellet was resuspended in 2 ml ice-cold H2O, centrifuged againand the supernatant pooled with the previous supernatant. Thepellet was resuspended in 1 ml ice-cold H2O followed by 4 mlethanol and incubated on ice for 15±30 min. After centrifugationas before, the supernatant was discarded and the pellet wasresuspended in 12 ml H2O. Duplicate aliquots of the suspendedstarch were diluted 5-fold with H2O, autoclaved and assayed for a-glucan. The pooled supernatants containing the soluble a-glucanwere diluted 6-fold with aqueous methanol/KCl (75% (v/v)methanol, 1% (w/v) (KCl), incubated at 4°C for 12 h andcentrifuged as before. The supernatant was discarded and thepellet was resuspended in 12 ml H2O. Duplicate aliquots of thesupernatant were diluted 6-fold with H2O, autoclaved and assayedfor a-glucan.
The wavelength of maximum absorption of the a-glucan-iodinecomplex was determined as follows. Puri®ed a-glucans weredissolved by suspension at 20 mg ml±1 in 1 M NaOH and dilutedwith an equal volume of water. An aliquot (20±50 ml) of thedissolved a-glucan was added to a cuvette containing, in a ®nalvolume of 1 ml, 100 mM NaOH, 100 mM acetic acid and 100 mlLugol solution. The maximum absorbance (400±800 nm) wasdetermined relative to that of a sample identical except that itcontained no a-glucan.
For a-glucan chain-length analysis, a-glucans were extractedfrom grains (0.5±3.0 g FW) ground to a ®ne powder in liquidnitrogen in a pestle and mortar. The powder was suspendedin 2 ml 10% (w/v) perchloric acid, transferred to a 50-ml tubeand shaken on ice for 30 min. After centrifugation at 18000 g for30 min at 4°C, the supernatant (containing the soluble a-glucan)was retained and the pellet (containing the starch) was resus-pended in 10 ml H2O, ®ltered through muslin (washing throughwith additional H2O) and centrifuged at 2500 g for 10 min. Thegrey layer on top of the starch was removed and the starchwas washed successively in 20 g l±1 SDS (thrice) and H2O (twice).After proteinase K treatment to remove surface proteins (Rahmanet al., 1995), the starch was washed successively in 20 g l±1
SDS (once), H2O (twice) and ice-cold acetone (twice), and freeze-dried.
The soluble a-glucans were precipitated by the addition of 3.5vols methanol to the perchloric acid-soluble material, incubatedon ice overnight and the precipitate was collected by centrifuga-tion at 2500 g for 15 min at 4°C. The supernatant was discardedand the pellet was resuspended in 4 ml H2O, and subjected to re-precipitation with methanol as above. The supernatant wasdiscarded and the soluble-glucan pellets were freeze-dried.
Glucan assays
Glucans were assayed as glucose released after digestion withspeci®c glucosylases. Control reactions, in which the glucosy-lases were omitted, were also performed. Duplicate samples(0.5 ml) were each incubated with 0.5 ml 50 mM Na acetatepH 5.2, 2 U a-amylase and 10 U amyloglucosidase (enzymes fromRoche Diagnostics, Lewes, UK) at 25°C for 12±24 h. The sampleswere heated to 100°C for 2 min, centrifuged at 14000 g for 2 minand the supernatants were assayed spectrophotometrically forglucose according to Lowry (1972).
Determination of chain-length distribution of a-glucans
The methods used were based on those of O'Shea and Morell(1996) as described in Edwards et al. (1999).
Scanning electron microscopy
Mature barley grains were fractured transversely using a razorblade to initiate the fracture. The pieces were mounted onaluminium stubs using silver conducting paint. Samples weresputter-coated (Emitech Ltd, Ashford, UK) with a layer of goldapproximately 25 nm thick and examined and photographed in aLeica Stereoscan 360 SEM (LEO, Cambridge, UK).
Light microscopy and transmission electron microscopy
Tissue slices approximately 1.5 mm thick were cut from theendosperm of developing barley and ®xed for at least 4 h in 3%
Figure 7. The number of granules in developing endosperm.Starch granules were puri®ed from developing endosperms of Bomi (r)and Risù 17 (e) and suspended in water. The number of granules inreplicate aliquots of the suspension was estimated using ahaemacytometer. The age of the grains from which the endospermswere dissected is expressed as days after anthesis (DAA). Values are themeans of measurements from three replicate aliquots of a suspension.Each aliquot was sampled 10 times. Each sample containedapproximately 50±250 starch granules per 0.00625 mm3.
108 Rachel A. Burton et al.
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

glutaraldehyde in 0.1 M cacodylate buffer, pH 7.4. The slices werewashed 3 times in buffer, cut into smaller pieces and ®xedovernight in 1% aqueous osmium tetroxide. The pieces weredehydrated in an ethanol series with 3 changes in 100% ethanoland transferred to acetone. Tissue was in®ltrated and embeddedin epoxy resin (Spurr's, Agar Scienti®c, Stansted, UK).
Sections 1 or 1.5 mm thick were stained with 1% (w/v) toluidineblue in 1% (w/v) borax, pH 11 and examined with an OlympusBX60 microscope (Olympus Optical, Japan) and recorded digitallywith AcQuis Bio software (Synoptics, Cambridge, UK).
Sections showing silver-gold interference colours were cutfrom embedded material with a diamond knife and collected oncopper grids. They were stained sequentially in uranyl acetate andReynold's lead citrate and examined and photographed in anelectron microscope (JEOL 1200EX/B).
Native gels
All procedures were carried out at 4±6°C. Developing endospermswere homogenized in approximately 10 vols extraction medium(100 mM MOPS pH 7.2, 10 mM EDTA, 50 ml l±1 ethanediol, 1 mM
DTT) in a pestle and mortar. The extract was centrifuged at28000 g for 5 min at 4°C. The resulting supernatant was added tosample loading medium (600 ml l±1 glycerol, 2 mg ml±1 bromo-phenol blue, 20 mM DTT) in a volume ratio of 1 : 5 (loading buffer:to extract).
Native gel electrophoresis was carried out according toLaemmli (1970), except that SDS was omitted from all solutionsand the separating gel contained 0.1% (w/v) acarbose (Glucobay100, Bayer plc, Berkshire, UK) to inhibit a-amylase activity. Theseparating gels consisted of 7.5% acrylamide, were 1 mm thickand contained potato amylopectin (1 mg ml±1, Sigma, Poole, UK)or red pullulan (10 mg ml±1, Megazyme International, CountyWicklow, Ireland).
After electrophoresis at 15 mA per gel and 4°C, native gelscontaining amylopectin were rinsed in medium A (100 mM MESpH 6.0, 5 mM DTT, 50 ml l±1 ethanediol, 0.1% (w/v) acarbose),incubated in this medium for 16 h at 37°C, rinsed brie¯y in waterand then stained with Lugol's solution. Native gels containing redpullulan were rinsed in medium A and then incubated in thismedium for 6±16 h at 37°C until bands were visible. Afterincubation, gels were soaked in 50 ml l±1 aqueous ethanol toenhance the contrast between the bands and the background.
Enzyme activities
All procedures were carried out at 4±6°C. Developing endospermswere dissected from grains of 45±55 mg FW and homogenized inapproximately 5 vols extraction medium (100 mM MOPS pH 7.2,5 mM DTT, 5 mM MgCl2, 5% (v/v) glycerol, 1% (w/v) BSA, 1% (w/v)(PVP) in a pestle and mortar followed by an all-glass homo-genizer. The extract was centrifuged at 28000 g for 10 min at 4°Cand the resulting supernatant was assayed. For total starchsynthase assays, the extract was not centrifuged.
For each enzyme, the activity reported was dependent upon thepresence in the assay of all of the appropriate substrates andcofactors and also upon extract concentration within the rangeused to make the measurements. The concentrations ofcomponents of each of the assays and their pH values wereoptimized to give the maximum rate using extracts of Bomi. Therate of the reaction was linear with respect to time for at least4 min in spectrophotometric assays and for at least 10 and 30 minin assays of sucrose synthase and starch synthase, respectively.
For starch-branching enzyme, activity was calculated from therate of reaction during the phase of the assay in which it waslinear with respect to time. Reaction mixtures were as follows:
Sucrose synthase. As in Craig et al. (1999) except that the bufferwas 82 mM AMPSO (3-[(1,1-Dimethyl-2-hydroxyethyl)amino]-2-hydroxy-propanesulphonic acid) pH 9.0.
UDPG pyrophosphorylase. The assay contained, in 1 ml,100 mM HEPES pH 8.1, 2 mM MgCl2, 0.8 mM NAD, 0.8 mM UDP-glucose, 1 mM NaPPi, 2 U phosphoglucomutase (PGM), 5 UG6PDH and 10±50 ml of a 10-fold dilution of extract in extractionmedium. The reaction was initiated with sodium pyrophosphate(NaPPi) and monitored spectrophotometrically at 340 nm.
Fructokinase. As in Craig et al. (1999) except that 2.5 mM NAD,3 mM MgCl2 and 100 ml extract were used.
Glucokinase. As in Craig et al. (1999) except that 100 mM BicinepH 8.5, 2 mM NAD, 2.5 U G6PDH, 100 ml extract were used andPGM was omitted.
Phosphoglucomutase. The assay contained, in 1 ml, 50 mM
Bicine pH 8.0, 0.6 mM NAD, 6 mM glucose-1-phosphate, 2 UG6PDH, 5±10 ml extract.
Phosphoglucose isomerase. The assay contained, in 1 ml,100 mM glycyl glycine pH 8.4, 1 mM NAD, 10 mM fructose-6-phosphate, 4 U G6PDH, 5±10 ml extract.
ADPG pyrophosphorylase. As in Smith et al. (1989; assay 2b),except that 100 mM HEPES pH 7.9, 0.4 mM NAD, 1 mM ADP-glucose, 1.5 mM NaPPi and 5 U PGM were used.
Soluble starch synthase. As in Jenner et al. (1994), the resinmethod except that 0.5 mg potato amylopectin, 2 mM ADP[U-14C]glucose at 2.3 GBq mol±1 and 10 ml of extract were used.
Granule-bound starch synthase. As above for soluble starchsynthase except that extracts which had not been centrifuged toremove insoluble material (total extracts) as well as solubleextracts were assayed. The granule-bound activity was calculatedas the difference between the activities in the total and solubleextracts.
Alkaline inorganic pyrophosphatase. As in Gross and ap Rees(1986) except that 50 mM Bicine pH 8.9, 20 mM MgCl2, 1.25 mM
NaPPi and 50 ml extract were used.Starch-branching enzyme. As in Smith (1990), the phosphor-
ylase-stimulation assay using MES buffer except that assays wereprocessed using DOWEX rather than methanol/KCl essentiallyaccording to Jenner et al. (1994), the resin method. 0±10 ml of a 10-fold dilution of extract was used and activity was expressed ineach case relative to that in assays with no extract.
RNA extraction and cDNA synthesis
Total RNA was extracted from barley tissues using a commer-cially available phenol-guanidine isothiocyanate preparation(Trizol; Gibco BRL, Gaithersburg, MD, USA). Single strandedcDNA was prepared from 1 mg of total RNA with Thermoscriptreverse transcriptase (Gibco BRL) and either an oligo(dT)20 or theTRACE (Frohman et al., 1988) primer according to the manufac-turers instructions. The cDNA was treated with RNase H for20 min at 37°C prior to the PCR reaction.
Isolation of the 5¢ and 3¢ ends of the barley isoamylase
cDNA
A cDNA sequence for barley isoamylase was published by Sunet al. (1999) but comparison with published sequences from othercereals (Zea mays, Beatty et al., 1997; AF030882 and Triticumaestivum, Luetticke et al., 2000; AX010486) indicated that it was
Isoamylase mutants of barley 109
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

not full length. The missing 5¢ end of the isoamylase transcriptwas obtained using nested PCR on cDNA prepared from total RNAfrom developing grain (Bomi) 10±13 days after fertilization. Theprimer sequences were 5¢-CCGATAAATAATCCCACCTCGC-3¢ and5¢-ATCACTGCCTTAGCATAAGGATCC-3¢ for the ®rst round of PCRand 5¢-GGCTGCAGGGCATGAAGATGATGGCCAT-3¢ with 5¢-GAACCTCCTCGCTCACCCTAT-3¢ for the second, nested round,with PCR conditions of 4 min at 94°C, followed by 35 cycles of1 min at 94°C, 1 min at 53°C, 1 min at 72°C and a ®nal extensiontime of 10 min at 72°C. The 3¢ end of the cDNA sequence wasobtained using a 3¢ RACE protocol where the ®rst round of PCRwas carried out with a gene-speci®c isoamylase primer 5¢-CCACTTATTGACATGATCAGC-3¢ and the RACE 3¢ primer 5¢-GACTC GAGTCGACATCG-3¢ (Frohman et al., 1988). A secondPCR reaction was carried out with a nested gene-speci®c primer5¢-CGTCAAGCTCATTGCTGAAGC-3¢ and RACE 3¢ with PCR condi-tions of 4 min at 94°C, followed by 35 cycles of 30 sec at 94°C,30 sec at 50°C, 2 min at 72°C and a ®nal extension time of 10 minat 72°C. PCR products to be analysed were cloned into the pGEM-T Easy vector (Promega, Madison WI, USA) and sequenced usingan ABI 3700 capillary sequencer.
Isolation of full-length isoamylase cDNAs
Primers for the isolation of full-length isa1 cDNAs from wild typeand mutant barley lines were designed to the 5¢ untranslatedregion upstream from the predicted translation start site and tothe 3¢ untranslated region at the 3¢ end of the transcript. A ®rst-round PCR reaction was carried out with the primers 5¢-CCGATAAATATCCCACCTCGC-3¢ and 5¢-CCGCCGAACGACTACA-TATAC-3¢ using the high-®delity Taq polymerase Elongase(Gibco, BRL) and PCR conditions of 45 sec at 94°C, followed by35 cycles of 45 sec at 94°C, 45 sec at 54°C, 3 min 30 sec at 72°Cand a ®nal extension time of 7 min at 72°C. A fully nested, secondround of PCR was carried out with 5¢-GGCTGCAGGGCATGAA-GATGATGGCCATGG-3¢ and 5¢-TCAAACATCAGGGCGTGATAC-AA-3¢ under the same PCR conditions. Putative full-length tran-scripts were cloned into the pGEM-T Easy vector (Promega). ThecDNA transcripts were fully sequenced in both directions using aset of overlapping primers on an ABI 3700 capillary sequencer.The resultant chromatograms were edited using Chromas(Technelysium, Helensvale, Queensland, Australia) software andanalysed with Genetic Computer Group (Madison, WI, USA)software in the ANGIS suite of programs at the University ofSydney.
Northern analysis
Approximately 10 mg of total RNA was separated on a 1% agarosedenaturing gel with size standards (Promega, Madison, USA).RNA was visualized with ethidium bromide under ultraviolet lightto ensure equal loading. RNA was transferred to Duralon nylonmembrane (Stratagene, La Jolla, USA) by capillary transfer andcrosslinked using UV light. A [32P]-labelled full-length isoamylasecDNA was synthesized by random priming, essentially asdescribed by Feinberg and Vogelstein (1983) and the membranewas probed as described by Banik et al., 1996). Autoradiographywas performed for 5 days at ± 80°C with X-ray ®lm and anintensifying screen.
Mapping the isoamylase gene, isa1
The probe DNA for Southern hybridization was radioactivelylabelled using standard methods. Hybridization methods were asdescribed in Rogowsky et al., 1991), except that both pre-hybridization and hybridization were carried out in the samesolution (0.9 M NaCl, 30 mM Pipes pH 6.8, 0.75 mM EDTA, 7.5%(w/v) dextran sulphate, 0.6% (w/v) BSA, 0.6% (w/v) Ficoll 400, 0.6%(w/v) polyvinyl-pyrollidone, 250 mg ml±1 denatured salmonsperm).
Counting starch granules
All procedures were carried out at 4±6°C. Developing endospermswere dissected from immature grains and homogenized inapproximately 50 vols extraction medium (50 mM HEPESpH 7.8, 10 mM EDTA, 10 mM DTT, 0.1 mg ml±1 and 0.1 mg ml±1
Proteinase K) in a pestle and mortar. The homogenate wasincubated at 37°C for 1 h, centrifuged at 28000 g for 5 min and thesupernatant discarded. The pellet was washed successively in 1 mlaliquots of 20 g l±1 SDS (twice), water (twice), 0.5 M NaCl, water,and the resulting starch preparation was resuspended in 1±10 mlof water. Three 200-ml or 1-ml aliquots of the suspension wereremoved for analysis. For each aliquot, the number of granulesper ml was estimated using a haemocytometer slide with a unitvolume of 0.00625 mm3. Starch granules stained with Lugolsolution, were diluted to approximately 50 granules per unitvolume and viewed under a light microscope. The precisenumber of granules per unit volume was determined 5 timeseach for two replicate dilutions of each sample of resuspendedstarch. These results were used to calculate the number ofgranules per endosperm.
Acknowledgements
The John Innes Centre is supported by a competitive strategicgrant from the Biotechnology and Biological Sciences ResearchCouncil (BBSRC), UK. The authors are extremely grateful to AlisonM. Smith for support, encouragement and useful discussionsthroughout the course of this work and for constructive criticismof the manuscript. Nicola Patron and Margaret Pallota are thankedfor DNA and RNA preparation, and chromosome mapping,respectively. Tamara Verhoeven thanks the BBSRC for a researchstudentship and Syngenta for additional ®nancial support. Workat the University of Adelaide was supported by grants (to GeoffFincher) from the Grains Research and Development Corporationof Australia and (to Kay Denyer) from the JIC/CSIRO/Waite fund.
References
Alonso, M.D., Lomako, J., Lomako, W.M. and Whelan, W.J. (1995)A new look at the biogenesis of glycogen. FASEB J. 9, 1126±1136.
Balaravi, S.P., Bansal, H.C., Eggum, B.O. and Bhaskaran, S. (1976)Characterisation of induced high protein and high lysinemutants in barley. J. Sci. Fd. Agric. 27, 545±552.
Ball, S., Guan, H.P., James, M., Myers, A., Keeling, P., Mouille, G.,Buleon, A., Colonna, P. and Preiss, J. (1996) From glycogen toamylopectin: a model for the biogenesis of the plant starchgranule. Cell 86, 349±352.
Banik, M., Garrett, T.P.J. and Fincher, G.B. (1996) Molecularcloning of cDNAs encoding (1®4)-b-xylan endohydrolyases
110 Rachel A. Burton et al.
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

from the aleurone layer of germinated barley (Hordeumvulgare). Plant Mol. Biol. 31, 1163±1172.
Bansal, H.C. (1970) A new mutant induced in barley. Curr. Sci. 39,494.
Beatty, M.K., Myers, A.M. and James, M.G. (1997) Genomicnucleotide sequence of a full-length wild-type allele of themaize sugary1 (su1) gene (accession, AF030882) (PGR 97±187).Plant Physiol. 115, 1731.
Boyer, C., Daniels, R.R. and Shannon, J.C. (1977) Starch granule(amyloplast) development in endosperm of several Zea mays L.genotypes affecting kernel polysaccharides. Am. J. Bot. 64, 50±56.
Burgess, S.R., Turner, R.H., Shewry, P.R. and Mi¯in, B.J. (1982)The structure of normal and high-lysine barley grains. J. Exp.Bot. 33, 1±11.
Craig, J., Barratt, P., Tatge, H., De jardin, A., Handley, L., Gardner,C., Barber, L., Wang, T.L., Hedley, C., Martin, C. and Smith, A.M.(1999) Mutations at the rug4 locus alter the carbon and nitrogenmetabolism of pea plants through an effect on sucrosesynthase. Plant J. 17, 101±110.
Dhugga, K.S., Tiwari, S.C. and Ray, P.M. (1997) A reversiblyglycosylated polypeptide (RGP1) possibly involved in plant cellwall synthsis: Puri®cation, gene cloning and trans-Golgilocalization. Proc. Nat. Acad. Sci. USA 94, 7679±7684.
Dinges, J.R., Colleoni, C., Myers, A.M. and James, M.G. (2001)Molecular structure of three mutations at the maize sugary1locus and their allele-speci®c phenotypic effects. Plant Physiol.125, 1406±1418.
Doll, H. (1983) Barley seed proteins and possibilities for theirimprovement. In: Seed Proteins: Biochemistry, Genetics,Nutritive Value (Gottschalk, W. and MuÈ ller, H.P., eds), TheHague: Martinus-Nijhoff, pp. 207±223.
Edwards, A., Fulton, D., Hylton, C.M., Jobling, S., Gidley, M.,RoÈ ssner, U., Martin, C. and Smith, A.M. (1999) A combinedreduction in activity of starch synthases II and III of potato hasnovel effects on the starch of tubers. Plant J. 17, 251±261.
Feinberg, A.P. and Vogelstein, B. (1983) A technique forradiolabelling DNA restriction fragments to high speci®cactivity. Anal Biochem. 132, 6±13.
Frohman, M., Dush, M. and Martin, G. (1988) Rapid ampli®cationof full-length cDNAs from rare transcripts: Ampli®cation using asingle gene-speci®c oligonucleotide primer. Proc. Natl Acad.Sci. USA 85, 8998±9002.
Gautam, M., Sood, K., Prakash, N. and Mehta, S.L. (1994)Amyloplasts and starch characterization in high lysine barleymutant. J. Plant Biochem. Biotech 3, 131±136.
Gross, P. and ap Rees, T. (1986) Alkaline inorganicpyrophosphatase and starch synthesis in amyloplasts. Planta167, 140±145.
James, M.J., Robertson, D.S. and Myers, A.M. (1995)Characterization of the maize gene sugary1, a determinant ofstarch composition in kernels. Plant Cell 7, 417±429.
Jane, J.-L., Kasemsuwan, T., Leas, S., Ames, I.A., Zobel, H.,Darien, I.L. and Robyt, J.F. (1994) Anthology of starch granulemorphology by scanning electron microscopy. Starch 46, 121±129.
Jenner, C.F., Denyer, K. and Hawker, J.S. (1994) Caution on theuse of the generally accepted methanol precipitation techniquefor the assay of soluble starch synthase in crude extracts ofplant tissues. Aust. J.Plant Physiol. 21, 17±22.
Jensen, J. (1979) Chromosomal location of one dominant andfour recessive high-lysine genes in barley mutants. In: SeedProtein Improvement in Cereals and Grain Legumes, Vol. 1.Vienna: IAEA, pp. 89±96.
Kubo, A., Fujita, N., Harada, K., Matsuda, T., Satoh, H. andNakamura, Y. (1999) The starch-debranching enzymesisoamylase and pullulanase are both involved in amylopectinbiosynthesis in rice endosperm. Plant Physiol. 121, 399±409.
Laemmli, U.K. (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680±685.
Langridge, P., Karakousis, A., Collins, N., Kretschmer, J. andManning, S. (1995) A consensus linkage map of barley. Mol.Breed. 1, 389±395.
Lloyd, J.R. (1995) Effect and Interactions of Rugosus Genes on Pea(Pisum sativum) Seeds. PhD Thesis, University of East Anglia,Norwich, UK.
Lomarko, J., Lomarko, W.M. and Whelan, W.J. (1992) Thesubstrate speci®city of isoamylase and the preparation ofapo-glycogenin. Carbohydr. Res. 227, 331±338.
Lowry, O.H. (1972) A Flexible System of Enzymatic Analysis. NewYork: Academic Press.
Luetticke, S., Abel, G., Loerz, H. and Genschel, U. (2000) NucleicAcid Molecules Which Code for Enzymes Derived from Wheatand Which Are Involved in the Synthesis of Starch. Patent: WO9958690-A.
MacGregor, E.A., Bazin, S.L., Ens, E.W., Lahnstein, J., Macri, L.J.,Shirley, N.J. and Macgregor, A.W. (2000) Structural models oflimit dextrinase inhibitors from barley. J.Cereal Sci. 31, 79±90.
Macri, L.J., Macgregor, A.W., Schroder, S.W. and Bazin, S.L.(1993) Detection of a limit dextrinase inhibitor in barley. J.Cereal Sci. 18, 103±106.
Manly, K.F. and Cudmore, R.H. (1997) Map manager QT, softwarefor mapping quantitative trait loci. Abstracts of the 11thInternational Mouse Genome Conference. St Petersburg, FL,USA, pp. 75.
Mouille, G., Maddelein, M.L., Libessart, N., Talaga, P., Decq, A.,Delrue, B. and Ball, S. (1996) Preamylopectin processing: amandatory step for starch biosynthesis in plants. Plant Cell 8,1353±1366.
Myers, A.M., Morell, M.K., James, M.G. and Ball, S.G. (2000)Recent progress toward understanding biosynthesis of theamylopectin crystal. Plant Physiol. 122, 989±997.
Nakamura, Y., Kubo, A., Shimamune, T., Matsuda, T., Harada, K.and Satoh, H. (1997) Correlation between activities of starchdebranching enzymes and a-polyglucan structure inendosperms of sugary-1 mutants of rice. Plant J. 12, 143±153.
Nakamura, Y., Umemoto, T., Takahata, Y., Komae, K., Amano, E.and Satoh, H. (1996) Changes in structure of starch and enzymeactivities affected by sugary mutations in developing riceendosperm. Possible role of starch debranching enzyme (R-enzyme) in amylopectin biosynthesis. Physiol. Plant. 97, 491±498.
O'Shea, M. and Morell, M.K. (1996) High-resolution slab gelelectrophoresis of 8-amino-1,3,6-pyrenetrisulfonic acid (APTS)tagged oligosaccharides using a DNA sequencer.Electrophoresis 17, 681±688.
Pan, D. and Nelson, O.E. (1984) A debranching enzyme de®ciencyin endosperms of the sugary-1 mutants of maize. Plant Physiol.74, 324±328.
Rahman, S., Kosar-Hashemi, B., Samuel, M.S., Hill, A., Abbott,D.C., Skerritt, J.H., Preiss, J., Appels, R. and Morell, M.K. (1995)The major proteins of wheat endosperm starch granules. Aust.J. Plant Physiol. 22, 793±803.
Roach, P.J. and Skurat, A.V. (1997) Self-glucosylating initiatorproteins and their role in glycogen biosynthesis. Prog. Nucl.Acid Res. Mol. Biol. 57, 290±316.
Rogowsky, P.M., Guidet, F.L.Y., Langridge, P., Shepherd, K.W.and Koebner, R.M.D. (1991) Isolation and characterization of
Isoamylase mutants of barley 111
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112

wheat-rye recombinants involving chromosome arm 1DS ofwheat. Theor. Appl. Genet. 82, 537±544.
Schulman, A.H. and Ahokas, H. (1990) A novel shrunkenendosperm mutant of barley. Physiol. Plant. 78, 583±589.
Shannon, J.C., Pien, F.M. and Liu, K.C. (1996) Nucleotides andnucleotide sugars in developing maize endosperms Ðsynthesis of ADP-glucose in brittle-1. Plant Physiol. 110, 835±843.
Singletary, G.W., Banisadr, R. and Keeling, P.L. (1997) In¯uence ofgene dosage on carbohydrate synthesis and enzymaticactivities in endosperm of starch-de®cient mutants of maize.Plant Physiol. 113, 293±304.
Smith, A.M. (1990) Enzymes of starch synthesis. Meth PlantBiochem. 3, 93±102.
Smith, A.M., Bettey, M. and Bedford, I.D. (1989) Evidence that therb locus alters the starch content of developing pea embryos
through an effect on ADP glucose pyrophosphorylase. PlantPhysiol. 89, 1279±1284.
Sood, K., Prakash, N. and Mehta, S.L. (1992) Synthesis anddevelopment of starch granules in high lysine barley grains. J.Plant Biochem. Biotech 1, 55±60.
Sun, C., Sathish, P., Ahlandsberg, S. and Jansson, C. (1999)Analyses of isoamylase gene activity in wild-type barleyindicate its involvement in starch synthesis. Plant. Mol. Biol.40, 431±443.
Tester, R.F., Morrison, W.R. and Schulman, A.H. (1993) Swellingand gelatinization of cereal starches. 5. Risù mutants of Bomiand Carlsberg-II barley cultivars. J. Cereal Sci. 17, 1±9.
Zeeman, S.C., Umemoto, T., Lue, W.-L., Au-Yeung, P., Martin, C.,Smith, A.M. and Chen, J. (1998) A mutant of Arabidopsislacking a chloroplastic isoamylase accumulates both starch andphytoglycogen. Plant Cell 10, 1699±1711.
Accession numbers for GenBank database: AF490375, AF490376, AF490377
112 Rachel A. Burton et al.
ã Blackwell Science Ltd, The Plant Journal, (2002), 31, 97±112
Related Documents











![Functions of Heteromeric and Homomeric Isoamylase-Type ... · Functions of Heteromeric and Homomeric Isoamylase-Type Starch-Debranching Enzymes in Developing Maize Endosperm1[W][OA]](https://static.cupdf.com/doc/110x72/600557e5a427f91eee0161c8/functions-of-heteromeric-and-homomeric-isoamylase-type-functions-of-heteromeric.jpg)