Staleya guttiformis attachment on poly(tert-butylmethacrylate) polymeric surfaces Elena P. Ivanova a, *, Natasa Mitik-Dineva a , James Wang a , Duy K. Pham a , Jonathan P. Wright a , Dan V. Nicolau b , Radu C. Mocanasu a , Russell J. Crawford a a Swinburne University of Technology, P.O. Box 218 Hawthorn, Vic. 3122, Australia b University of Liverpool, UK 1. Introduction Attachment of bacteria to solid surfaces is critical in the formation of a biofilm, a phenomenon that prompted extensive research for many decades due to its diverse implications in natural environments, industrial processes, biotechnological applications, and clinical situations (Beech and Gaylard, 1991; Bruinsma et al., 2001a,b; Denes et al., 2001; Sutherland, 2001; Pasmore and Costerton, 2003; Francolini et al., 2004; Hall-Stoodley et al., 2004; Beech et al., 2005; Senaratne et al., 2005). Recently non-specific binding interactions received significant amount of attention over the course of the development of novel nanofab- rication techniques, e.g. Senaratne et al. (2005). The role of solid material surface chemistry in bioelectronics and biosensors is particularly critical as the resistance to non-specific binding significantly reduces background interferences. Resistance to non- specific binding is also of concern when designing surfaces for implants, for biomedical diagnostics and drug release (Hall- Stoodley et al., 2004). The initial interaction between the bacterial cell and the solid surface through non-specific binding has been described by classical physico-chemical theories (Marshall et al., 1971; Norde and Lyklema, 1989; Rijnaarts et al., 1995; Bos et al., 1999; Razatos, 2001; Dong et al., 2002; Eboigbodin et al., 2006) which, however, do not take into account the inherent properties of bacteria, e.g. the formation of fimbriae, pili, or production of EPS. Notwithstanding numerous studies performed in this area, the process of bacterial adhesion remains far from being fully understood mainly because of the enormous diversity of both, the surfaces of the bacterial cells and the surfaces of the substrata. Our previous work in this area has demonstrated that common polymers such as poly(tert-butylmethacrylate), poly(styrene maleic acid) (PSMA) (Ivanova et al., 2002a) (as well as a combination of both in a bilayer arrangement) can be used as a platform for patterning of biological species via, e.g. the discrimination of differences in hydrophobicity (Wright et al., 2003). In this study we investigated a non-specific attachment of bacterial cells Staleya guttiformis DSM 11458 T on P(tBMA) polymeric surfaces. Bacteria of this taxonomic lineage, namely the marine a-proteobacteria of ‘Roseobacter– Sulfitobacter–Silicibacter’ group (Staley, 1968; Sorokin, 1995; Vogler, 1998; Wagner-Do ¨bler et al., 2003), represent the second most abundant 16S rRNA gene clone type in marine environments (Rappe ´ et al., 2000). Bacteria of these taxa play an important role in nutrient Micron 39 (2008) 1197–1204 ARTICLE INFO Article history: Received 15 April 2008 Received in revised form 28 April 2008 Accepted 28 April 2008 Keywords: Bacterial attachment AFM SEM Extracellular polymeric substances Poly(tert-butylmethacrylate) polymeric surfaces ABSTRACT The attachment behaviour of Staleya guttiformis DSM 11458 T on poly(tert-butyl methacrylate) (P(tBMA)) polymeric surfaces has been studied. The electrostatic charge of the S. guttiformis cell surface (measured as zeta potential via microelectrophoresis) was 43.18 mV. S. guttiformis cells appeared weakly hydrophilic as the water contact angle measured on lawns of bacterial cells was found to be 55 4.98. It was found that while attaching on P(tBMA) surfaces, S. guttiformis cells produced extracellular polymeric substances (EPS) as observed from atomic force microscopy (AFM) and scanning electron microscopy (SEM) analysis. The AFM high resolution imaging revealed the nano-topography of the ‘free’ (the EPS that is produced by the bacterial cells, but no longer directly attached to the cells) EPS associated on the cell surface and also found on P(tBMA) surface. The ‘free’ EPS exhibited granular structure with lateral dimensions of 30– 50 nm and a vertical nano-roughness of 7–10 nm. Another type of the EPS secreted by S. guttiformis cells appeared as a hydogel substance, presumably polysaccharide that formed a biopolymer network that facilitated bacterial attachment. ß 2008 Elsevier Ltd. All rights reserved. * Corresponding author. Tel.: +61 3 9214 5137; fax: +61 3 9214 5050. E-mail address: [email protected] (E.P. Ivanova). Contents lists available at ScienceDirect Micron journal homepage: www.elsevier.com/locate/micron 0968-4328/$ – see front matter ß 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.micron.2008.04.009

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Staleya guttiformis attachment on poly(tert-butylmethacrylate) polymericsurfaces
Elena P. Ivanova a,*, Natasa Mitik-Dineva a, James Wang a, Duy K. Pham a, Jonathan P. Wright a,Dan V. Nicolau b, Radu C. Mocanasu a, Russell J. Crawford a
a Swinburne University of Technology, P.O. Box 218 Hawthorn, Vic. 3122, Australiab University of Liverpool, UK
Micron 39 (2008) 1197–1204
A R T I C L E I N F O
Article history:
Received 15 April 2008
Received in revised form 28 April 2008
Accepted 28 April 2008
Keywords:
Bacterial attachment
AFM
SEM
Extracellular polymeric substances
Poly(tert-butylmethacrylate) polymeric
surfaces
A B S T R A C T
The attachment behaviour of Staleya guttiformis DSM 11458T on poly(tert-butyl methacrylate) (P(tBMA))
polymeric surfaces has been studied. The electrostatic charge of the S. guttiformis cell surface (measured
as zeta potential via microelectrophoresis) was �43.18 mV. S. guttiformis cells appeared weakly
hydrophilic as the water contact angle measured on lawns of bacterial cells was found to be 55 � 4.98. It
was found that while attaching on P(tBMA) surfaces, S. guttiformis cells produced extracellular polymeric
substances (EPS) as observed from atomic force microscopy (AFM) and scanning electron microscopy (SEM)
analysis. The AFM high resolution imaging revealed the nano-topography of the ‘free’ (the EPS that is
produced by the bacterial cells, but no longer directly attached to the cells) EPS associated on the cell surface
and also found on P(tBMA) surface. The ‘free’ EPS exhibited granular structure with lateral dimensions of 30–
50 nm and a vertical nano-roughness of 7–10 nm. Another type of the EPS secreted by S. guttiformis cells
appeared as a hydogel substance, presumably polysaccharide that formed a biopolymer network that
facilitated bacterial attachment.
� 2008 Elsevier Ltd. All rights reserved.
Contents lists available at ScienceDirect
Micron
journal homepage: www.elsevier .com/locate/micron
1. Introduction
Attachment of bacteria to solid surfaces is critical in theformation of a biofilm, a phenomenon that prompted extensiveresearch for many decades due to its diverse implications innatural environments, industrial processes, biotechnologicalapplications, and clinical situations (Beech and Gaylard, 1991;Bruinsma et al., 2001a,b; Denes et al., 2001; Sutherland, 2001;Pasmore and Costerton, 2003; Francolini et al., 2004; Hall-Stoodleyet al., 2004; Beech et al., 2005; Senaratne et al., 2005). Recentlynon-specific binding interactions received significant amount ofattention over the course of the development of novel nanofab-rication techniques, e.g. Senaratne et al. (2005). The role of solidmaterial surface chemistry in bioelectronics and biosensors isparticularly critical as the resistance to non-specific bindingsignificantly reduces background interferences. Resistance to non-specific binding is also of concern when designing surfaces forimplants, for biomedical diagnostics and drug release (Hall-Stoodley et al., 2004).
* Corresponding author. Tel.: +61 3 9214 5137; fax: +61 3 9214 5050.
E-mail address: [email protected] (E.P. Ivanova).
0968-4328/$ – see front matter � 2008 Elsevier Ltd. All rights reserved.
doi:10.1016/j.micron.2008.04.009
The initial interaction between the bacterial cell and the solidsurface through non-specific binding has been described byclassical physico-chemical theories (Marshall et al., 1971; Nordeand Lyklema, 1989; Rijnaarts et al., 1995; Bos et al., 1999; Razatos,2001; Dong et al., 2002; Eboigbodin et al., 2006) which, however,do not take into account the inherent properties of bacteria, e.g. theformation of fimbriae, pili, or production of EPS. Notwithstandingnumerous studies performed in this area, the process of bacterialadhesion remains far from being fully understood mainly becauseof the enormous diversity of both, the surfaces of the bacterial cellsand the surfaces of the substrata.
Our previous work in this area has demonstrated that commonpolymers such as poly(tert-butylmethacrylate), poly(styrene maleicacid) (PSMA) (Ivanova et al., 2002a) (as well as a combination of bothin a bilayer arrangement) can be used as a platform for patterning ofbiological species via, e.g. the discrimination of differences inhydrophobicity (Wright et al., 2003). In this study we investigated anon-specific attachment of bacterial cells Staleya guttiformis DSM11458T on P(tBMA) polymeric surfaces. Bacteria of this taxonomiclineage, namely the marine a-proteobacteria of ‘Roseobacter–
Sulfitobacter–Silicibacter’ group (Staley, 1968; Sorokin, 1995; Vogler,1998; Wagner-Dobler et al., 2003), represent the second mostabundant 16S rRNA gene clone type in marine environments (Rappeet al., 2000). Bacteria of these taxa play an important role in nutrient

E.P. Ivanova et al. / Micron 39 (2008) 1197–12041198
cycling, e.g. by oxidation or degradation of sulfite (Rijnaarts et al.,1999), dimethyl sulfoniopropionate (Gonzalez et al., 2003), methy-lamine (Doronina et al., 2000), lignin (Gonzalez et al., 1997),aromatic compounds (Buchan et al., 2000), etc. Despite theirsignificant ecological functions and apparent applications inbiotechnology, these microorganisms remain poorly studied inthe context of their attachment capabilities. The present study is anextension of our previous investigation to probe the attachment ofmarine a-proteobacteria and their biofilm formation on polymericsurfaces (Ivanova et al., 2002b). We previously found thatSulfitobacter brevis, Sulfitobacter mediterraneus, and Sulfitobacter
pontiacus, all belonging to the same taxonomic lineage as Staleya,expressed different attachment behaviour on P(tBMA). In particular,vegetative cells of S. mediterraneus underwent morphologicalconversion into coccoid forms, while S. pontiacus and S. brevis failedto attach onto poly(tert-butyl methacrylate) polymeric surfaces. Incontrast, S. guttiformis was able to successfully sustain its existenceon this surface and form a biofilm (Ivanova et al., 2002b). In light ofthis finding, it was of interest to study in more detail the S. guttiformis
attachment pattern on a P(tBMA) surface. To fulfill the aims of thisstudy, we undertook a comprehensive characterisation of thepolymeric surfaces; both SEM and the various operating modes ofAFM (including the friction force mode) were particularly useful ininvestigation of the cell surface and ultrastructure of the EPSreleased by bacteria during their attachment to polymeric surfaces.
2. Material and methods
2.1. Polymer film preparation
The P(tBMA) polymeric films were prepared as describedelsewhere (Ivanova et al., 2002b). Briefly, a 4 wt% solution ofP(tBMA) (MW � 170,000) in propylene glycol methyl ether acetate(PGMEA), (Sigma–Aldrich Co.) 99% was used to prepare thepolymer film. Polymer films were prepared on 30 mm � 50 mmglass substrates (glass cover slips, #1, 0.17 mm thickness,24 mm � 24 mm, Knittel) that had been sonicated in isopropanol(PriOH) for 30 min, washed with copious amounts of filtered(0.2 mm) nanopure water (18.2 M cm�1, Barnstead/ThermolyneNANOpure1 Infinity water purification system), and dried under astream of high purity nitrogen prior to priming with hexamethyl-disilazane (HMDS) (Sigma–Aldrich Co.). The primer was spun at1000 rpm and polymers at 3000 rpm for 40 s using a SpecialtyCoating Systems spin coater (Model P6708). Finally, polymercovered slides were baked for 60 min at 95 8C in an oven and storedin a desiccator prior to use.
2.2. Polymer film characterisation
The polymeric films were analysed for their surface wettabilityusing contact angle measurements. Static contact angles weremeasured using sessile drops (2 mL) of nanopure water at roomtemperature (ca. 23–25 8C) in air using a Rohm–Hart contact anglegoniometer (Model 100-00-115). Measured contact angles wereaveraged over six different readings. Polymer film thicknessmeasurements were performed using a standard AutoEl-IIIellipsometer (software version 3.9, Rudolph Research, USA),equipped with a He–Ne (632.8 nm) laser source as describedelsewhere (Jonsson et al., 1985; Ivanova et al., 2006).
Polymer films roughness measurements were performed usinga Solver P7LS scanning probe microscope (SPM) from NT-MDT, RF.The focused ion beam carbon ‘‘whisker’’ type silicon cantilevers(NSC05), manufactured by NT-MDT, with a spring constant of11 N/m, curvature radius of the tip 10 nm, aspect ratio 10:1 andresonance frequency of 150 kHz were used to obtain the best
morphological resolution. Scanning was performed perpendicularto the axis of the cantilever at a typical rate of 1 Hz. Imageprocessing of the raw topographical data was performed with firstorder horizontal and vertical levelling, and the topography andsurface profile of the samples were obtained simultaneously. SPMSolver P7LS and its accompanying instrument software (LS7-SPMv.8.58) allow measurement of sample features with resolution of afraction of nanometer by means of statistical analysis of the surfaceroughness and grain size for the investigated areas.
The surface features of polymer films was also observed using aconventional type of scanning electron microscope (SEM) ZEISSSUPRA 40VP (Fig. 2A). Before the sample was mounted on pin typealuminium SEM mounts with double-sided conducting carbontape, it was coated a 10 nm thick gold using a magnetronsputtering system (Kurt J. Lesker CMS-18). The working distance(WD) varied between 6 and 7 mm, and images were mostlycaptured on 500�, 1000� and 10,000� magnification.
The chemical composition of the polymer was analysed by XPSusing Axis Ultra spectrometer (Kratos Analytical Ltd., UK),equipped with a monochromatised X-ray source (Al Ka,hn = 1486.6 eV) operating at 150 W. The spectrometer energyscale was calibrated using the Au 4f7/2 photoelectron peak atbinding energy (EB = 83.98 eV). Photoelectrons emitted at 908 tothe surface from an area of 700 mm � 300 mm were using 160 eVsurvey spectra and 20 eV for high resolution region spectra forselected elements (O 1s, C 1s, K 2p, Si 2p).
2.3. Bacterial sample preparation
S. guttiformis DSM 11458T originally isolated from hypersaline,heliothermal and meromistic Ekho Lake (Vestfold Hills, EastAntarctica), was a generous gift from Dr. P. Hirsch. The bacteriumwas routinely cultured on marine agar 2216 and PYGV agar platesand stored at �80 8C in marine broth 2216 (Difco) supplementedwith 20% (v/v) of glycerol. For bacterial adhesion experiments theglass cover slips coated by polymer were placed in wells (NuncMultidishes (12 wells) in duplicate). The wells were inoculatedwith log-phase culture (2 mL) of cell density adjusted to OD650
0.13. Sterile marine broth 2216 (2 mL) was used as a negativecontrol. Multidishes were incubated at 28 8C for 24–72 h. Afterincubation, the biofilms formed by statically grown bacteria, wererinsed three times with sterilized nanopure H2O (18.2 MV cm�1).Biofilms were handled under sterile conditions until just prior tolabelling and/or imaging. For AFM imaging all slides were kepthydrated during experiments.
2.4. Cell surface wettability
Cell surface wettability was determined by contact anglesmeasurements on the lawns of bacteria using a First TenAngstroms (FTA200). Bacterial cells in a buffer (OD450 = 0.4) weredeposited on cellulose acetate membrane filters (Sartorius, porediameter 0.2 mm). The wet filters were air dried at ambienttemperature until a ‘‘plateau state’’ was reached. Droplets of waterwere deposited to each surface. The droplet contact angle wasobserved with a charge-coupled device (CCD) camera. The dropwas allowed to settle for 2 s without needle contact (for staticcontact angle measurements), images were digitally saved andcontact angle values obtained by processing the image with theimage analysis software (v 1.961; Folio Instruments Inc.).
2.5. Electrophoretic mobility
The electrophoretic mobility (EPM) was measured using a zetapotential analyzer (ZetaPALS, Brookhaven Instruments Corp.) as

E.P. Ivanova et al. / Micron 39 (2008) 1197–1204 1199
described elsewhere (Mitik-Dineva et al., 2008). Briefly, thebacterial cell suspension was freshly prepared before themeasurement. After 24 h of growth in marine broth, the cellswere harvested by centrifugation for 5 min at 5000 rpm. Harvestedcell pellet was re-suspended in 10 mM potassium chloride (KCl)followed by further washing and centrifugation. This step wasrepeated four times and finally the cells were re-suspended in10 mM KCl solution to OD(600 nm) = 1. This cell solution was thendiluted 1000 times in 5 mL of 10 mM KCl for the EPM measure-ments. Measurements were conducted in electric field 2.5 V cm�1
and 2 Hz. All measurements were done in triplicate and for eachsample the final EPM quoted represented the average of 5successive ZetaPALS readings, each of which consisted of 14 cyclesper run.
2.6. Atomic force microscopy
The pattern of bacterial attachment was evaluated using AFM.AFM imaging was carried out using an Explorer system (ThermoMicroscopes) (Model No. 4400-11) in normal contact mode. Lateralforce images were obtained from the difference in the forward andreverse scans using AFM software. Images were obtained using an8 mm-Z, linearised liquid scanner (100 mm � 100 mm), under air-ambient conditions (ca. 23 8C and 45% relative humidity). Employ-ment of these conditions helped to avoid drastic cell dehydration,hence minimizing their deformation when obtaining high resolu-tion AFM images. Pyramidal silicon nitride tips attached tocantilevers with a spring constant of 0.032 N/m (Gibson et al.,1996; Watson et al., 2003) were used in the contact mode. In thenon-contact mode silicon tips (Microscopes SFM Probes Model No.1650-00 HFR) of radius < 20 nm (the size was verified using theSEM micrograph (data not shown)) and cantilevers with a springconstant of 42 N/m and resonance frequency of 320 kHz were used.Scanning was performed perpendicular to the axis of the cantileverat a typical rate of 5 Hz. Image processing of the raw topographicaldata was performed with first order horizontal and verticalleveling using the accompanying instrument software (TopoMe-trix SPMLab v.3.06.06). Before AFM imaging all samples werechecked by visual observation and scanned using an opticalOlympus IX71/IX51 microscope.
2.7. Confocal microscopy
Cells and EPS were labelled by utilizing two dyes, thefluorescein-derived dye CFDA SE (Vybrant CFDA SE Cell Tracer,Molecular Probes) and concanavalin A 488 (Molecular Probes) aspreviously described (Mitik-Dineva et al., 2008). For visualisationof viable bacterial cells, a carboxyfluorescein diacetate succinimi-
Fig. 1. Typical AFM topographical images taken from poly(tert-butyl methacrylate) polym
dyl ester (CFDA SE) stock solution (10 mM) was diluted to 20 mM inPBS and was further used as working solution. Concanavalin Astock solution was prepared according to the manufacturerprotocol. The working solution was prepared by diluting the stocksolution to 1:20 using the same buffer to avoid changes in pH. Afterincubation (total of 24 h) the samples were washed and carefullytransferred to be observed with Olympus Fluoview FV1000Spectroscopic Confocal System (CSLM) including an invertedMicroscope System OLYMPUS IX81 (20�, 40� (oil), 100� (oil)UIS objectives). The system was equipped with a transmitted lightdifferential interference contract attachment and a CCD camera(Cool view FDI). Excitation and emission wavelengths forconcanavalin A and for CFDA SE are 495/519 and 492/517 nm,respectively.
3. Results and discussion
3.1. P(tBMA) surface characterisation
P(tBMA) is a commonly used polymer for various biomedicalapplications because of its characteristics, e.g. transparency (>90%transmission), stiffness, low water absorption and high abrasionresistance. The polymeric films used in this work were of about54.8 � 3 nm thick as estimated by ellipsometry. This thickness iswithin a range of the polymeric films thickness and sufficientlyrepresents the chemical species on the surface (Ivanova et al., 2006).The P(tBMA) surface was hydrophobic with measured water contactangles (u) ranging from 798 to 848. The hydrophobic nature of P(tBMA)is conferred by the surface methyl groups and the bondingarrangement around the oxygen atoms (Raczkowska et al., 2004).The relative atomic concentration of elements detected by XPS wasquantified from the area of peaks in the survey spectra sensitivityfactors appropriate for the Kratos instrument. The results indicated(data not shown) that carbon and oxygen were the most abundantelements present on the surface, representing more than 90% of theelemental composition on the polymer surface. The polymer surfacetopography was analysed by SEM (Fig. 2A) and AFM imaging (Fig. 1).Polymer surface roughness was inferred by AFM employing threeconventional roughness parameters. The average surface roughnessof the poly(tert-butyl methacrylate) polymeric surfaces wasRa = 5.523 nm, the root mean square was Rq = 8.546 nm, and themaximum roughness Rmax = 53.65 nm.
3.2. S. guttiformis cell surface characterization
The physico-chemical properties of the cell surfaces wereassessed as these properties are known to control cell–surfaceinteractions (Marshall et al., 1971; Beech et al., 1991, 2005; Parsek
eric surfaces. The imaged areas are 30 mm � 30 mm and 5 mm � 6 mm, respectively.
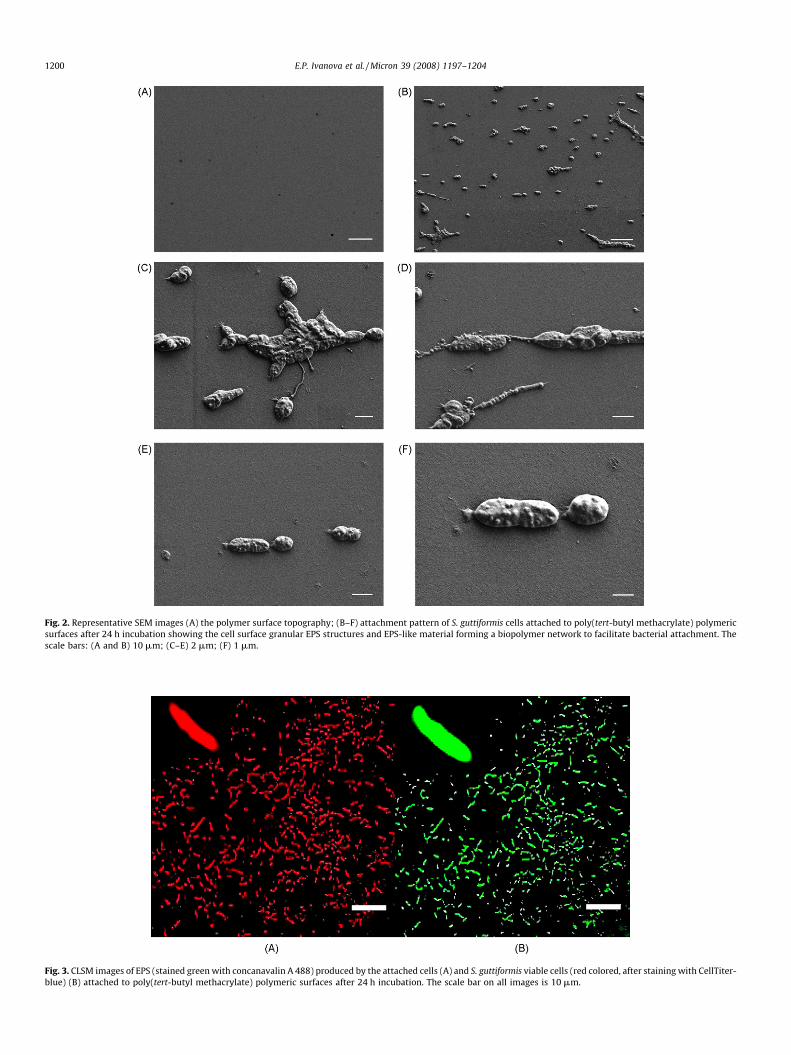
Fig. 2. Representative SEM images (A) the polymer surface topography; (B–F) attachment pattern of S. guttiformis cells attached to poly(tert-butyl methacrylate) polymeric
surfaces after 24 h incubation showing the cell surface granular EPS structures and EPS-like material forming a biopolymer network to facilitate bacterial attachment. The
scale bars: (A and B) 10 mm; (C–E) 2 mm; (F) 1 mm.
Fig. 3. CLSM images of EPS (stained green with concanavalin A 488) produced by the attached cells (A) and S. guttiformis viable cells (red colored, after staining with CellTiter-
blue) (B) attached to poly(tert-butyl methacrylate) polymeric surfaces after 24 h incubation. The scale bar on all images is 10 mm.
E.P. Ivanova et al. / Micron 39 (2008) 1197–12041200
E.P. Ivanova et al. / Micron 39 (2008) 1197–1204 1201
and Singh, 2003). The electrostatic surface charge of S. guttiformis
cells was evaluated by the measurements of the cells’ electro-phoretic mobility. S. guttiformis cells were found to be negativelycharged with an estimated zeta potential of �43.18 mV, a valuethat is in the range of values previously reported for g-protobacteria (van der Mei and Busscher, 2001; Li and Logan,2004, 2005; Jones and Velegol, 2006). Cell surface wettability(quantified by water contact angle measurements on lawns onbacterial cells) was generally in the range of 55 � 4.98. This contactangle shows that the cells are on the borderline between exhibitinghydrophobic and hydrophilic properties given that angles greaterthan 60–658 denote hydrophobicity (Wagner-Dobler et al., 2003;Korenevsky and Beveridge, 2007). Therefore, S. guttiformis cells can beregarded as only slightly hydrophilic.
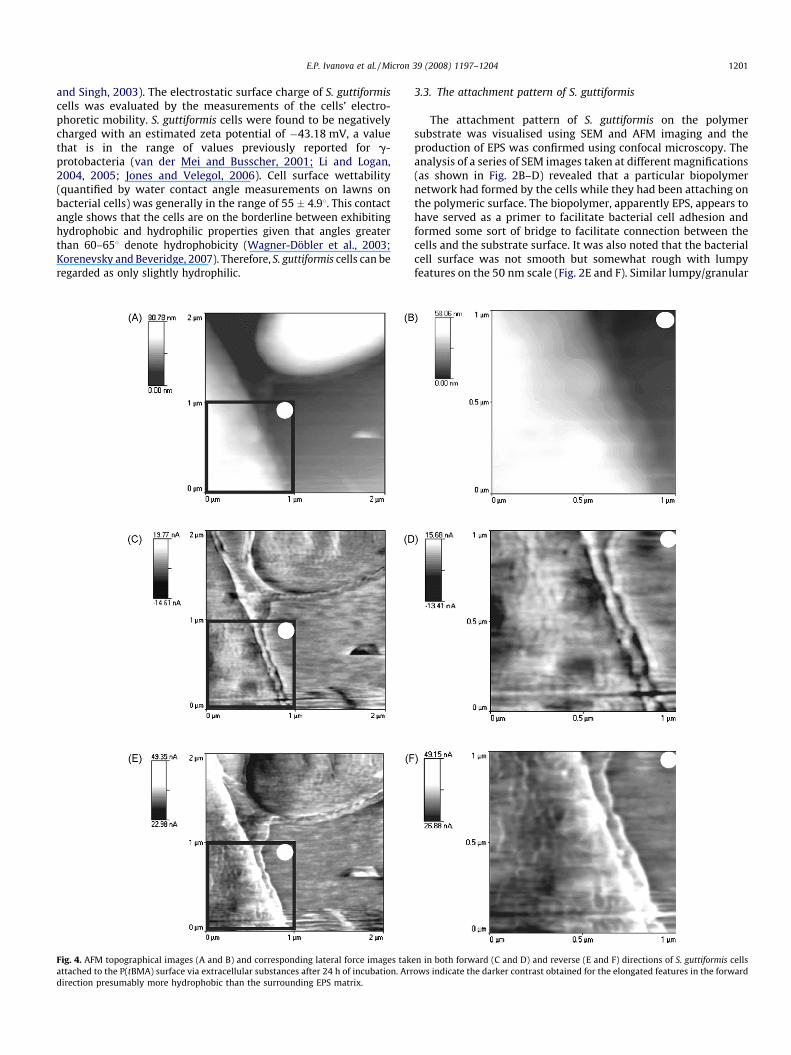
Fig. 4. AFM topographical images (A and B) and corresponding lateral force images tak
attached to the P(tBMA) surface via extracellular substances after 24 h of incubation. Arr
direction presumably more hydrophobic than the surrounding EPS matrix.
3.3. The attachment pattern of S. guttiformis
The attachment pattern of S. guttiformis on the polymersubstrate was visualised using SEM and AFM imaging and theproduction of EPS was confirmed using confocal microscopy. Theanalysis of a series of SEM images taken at different magnifications(as shown in Fig. 2B–D) revealed that a particular biopolymernetwork had formed by the cells while they had been attaching onthe polymeric surface. The biopolymer, apparently EPS, appears tohave served as a primer to facilitate bacterial cell adhesion andformed some sort of bridge to facilitate connection between thecells and the substrate surface. It was also noted that the bacterialcell surface was not smooth but somewhat rough with lumpyfeatures on the 50 nm scale (Fig. 2E and F). Similar lumpy/granular
en in both forward (C and D) and reverse (E and F) directions of S. guttiformis cells
ows indicate the darker contrast obtained for the elongated features in the forward
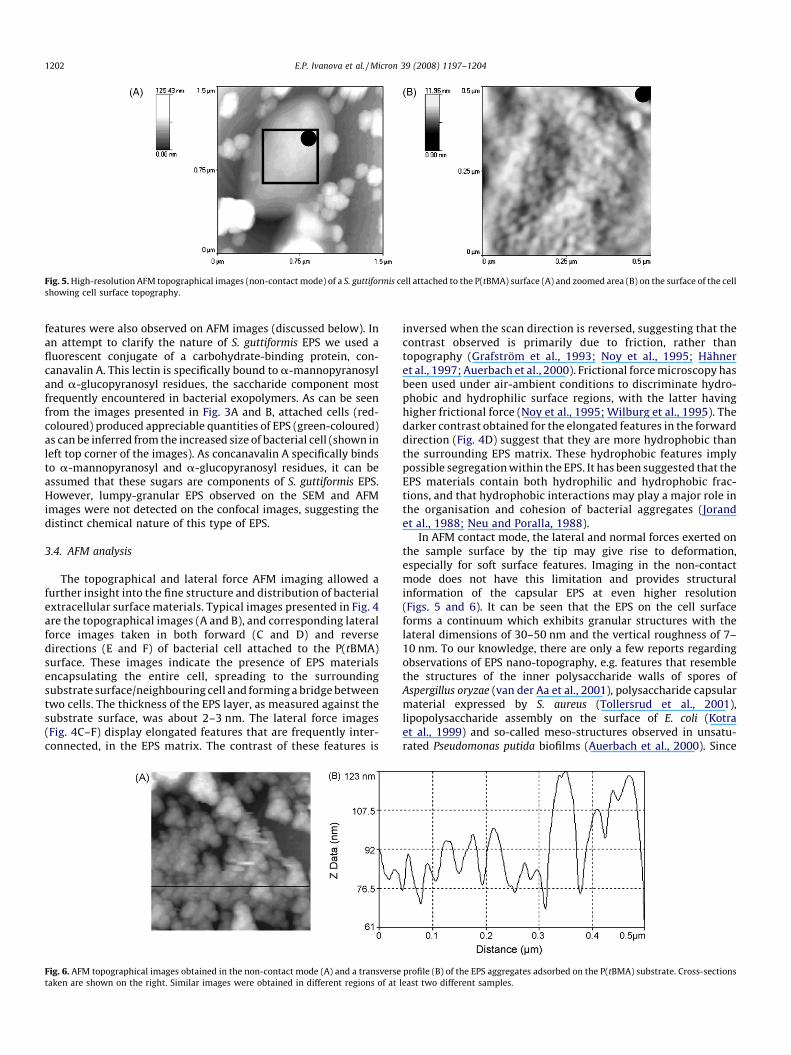
Fig. 5. High-resolution AFM topographical images (non-contact mode) of a S. guttiformis cell attached to the P(tBMA) surface (A) and zoomed area (B) on the surface of the cell
showing cell surface topography.
E.P. Ivanova et al. / Micron 39 (2008) 1197–12041202
features were also observed on AFM images (discussed below). Inan attempt to clarify the nature of S. guttiformis EPS we used afluorescent conjugate of a carbohydrate-binding protein, con-canavalin A. This lectin is specifically bound to a-mannopyranosyland a-glucopyranosyl residues, the saccharide component mostfrequently encountered in bacterial exopolymers. As can be seenfrom the images presented in Fig. 3A and B, attached cells (red-coloured) produced appreciable quantities of EPS (green-coloured)as can be inferred from the increased size of bacterial cell (shown inleft top corner of the images). As concanavalin A specifically bindsto a-mannopyranosyl and a-glucopyranosyl residues, it can beassumed that these sugars are components of S. guttiformis EPS.However, lumpy-granular EPS observed on the SEM and AFMimages were not detected on the confocal images, suggesting thedistinct chemical nature of this type of EPS.
3.4. AFM analysis
The topographical and lateral force AFM imaging allowed afurther insight into the fine structure and distribution of bacterialextracellular surface materials. Typical images presented in Fig. 4are the topographical images (A and B), and corresponding lateralforce images taken in both forward (C and D) and reversedirections (E and F) of bacterial cell attached to the P(tBMA)surface. These images indicate the presence of EPS materialsencapsulating the entire cell, spreading to the surroundingsubstrate surface/neighbouring cell and forming a bridge betweentwo cells. The thickness of the EPS layer, as measured against thesubstrate surface, was about 2–3 nm. The lateral force images(Fig. 4C–F) display elongated features that are frequently inter-connected, in the EPS matrix. The contrast of these features is
Fig. 6. AFM topographical images obtained in the non-contact mode (A) and a transverse
taken are shown on the right. Similar images were obtained in different regions of at
inversed when the scan direction is reversed, suggesting that thecontrast observed is primarily due to friction, rather thantopography (Grafstrom et al., 1993; Noy et al., 1995; Hahneret al., 1997; Auerbach et al., 2000). Frictional force microscopy hasbeen used under air-ambient conditions to discriminate hydro-phobic and hydrophilic surface regions, with the latter havinghigher frictional force (Noy et al., 1995; Wilburg et al., 1995). Thedarker contrast obtained for the elongated features in the forwarddirection (Fig. 4D) suggest that they are more hydrophobic thanthe surrounding EPS matrix. These hydrophobic features implypossible segregation within the EPS. It has been suggested that theEPS materials contain both hydrophilic and hydrophobic frac-tions, and that hydrophobic interactions may play a major role inthe organisation and cohesion of bacterial aggregates (Jorandet al., 1988; Neu and Poralla, 1988).
In AFM contact mode, the lateral and normal forces exerted onthe sample surface by the tip may give rise to deformation,especially for soft surface features. Imaging in the non-contactmode does not have this limitation and provides structuralinformation of the capsular EPS at even higher resolution(Figs. 5 and 6). It can be seen that the EPS on the cell surfaceforms a continuum which exhibits granular structures with thelateral dimensions of 30–50 nm and the vertical roughness of 7–10 nm. To our knowledge, there are only a few reports regardingobservations of EPS nano-topography, e.g. features that resemblethe structures of the inner polysaccharide walls of spores ofAspergillus oryzae (van der Aa et al., 2001), polysaccharide capsularmaterial expressed by S. aureus (Tollersrud et al., 2001),lipopolysaccharide assembly on the surface of E. coli (Kotraet al., 1999) and so-called meso-structures observed in unsatu-rated Pseudomonas putida biofilms (Auerbach et al., 2000). Since
profile (B) of the EPS aggregates adsorbed on the P(tBMA) substrate. Cross-sections
least two different samples.

E.P. Ivanova et al. / Micron 39 (2008) 1197–1204 1203
the observed extracellular structures were larger than theglycolipids and glycoproteins, those authors assumed that suchstructures were aggregates of smaller molecules. Similar to the cellsurface granular EPS structures, ‘free’ EPS-like material, sometimeswithout any association with bacterial cells, was also detected. Asshown in the non-contact mode images in Figs. 5 and 6, thesematerials are of various sizes and shapes and often adhere to eachother to form larger aggregates. The latter have granular structureswith lateral dimensions of 30–50 nm, apparently bound togetherto form stacks which can be up to 150 nm in height. The nanometerscale granular structures (Figs. 5 and 6) as seen in the non-contactmode are not observed in the contact mode, indicating that theywere made of very soft material and that both the lateral andnormal forces at the tip–sample surface caused considerablesurface deformation. As well, there is no significant variation in thelateral force contrast that can be attributed to frictional differenceson the EPS surface. The lateral force contrast at the boundaries ofthe EPS aggregates is likely due to topographical effects. This is incontrast to the capsular-like EPS that exhibited elongated featureshaving lower friction than the surrounding EPS matrix as discussedabove.
3.5. Concluding remarks
If only theoretical thermodynamic properties are considered,the net negative surface charge and weak hydrophobic/hydro-philic nature of S. guttiformis would not be expected to show apropensity for attachment to the P(tBMA) surfaces (Bruinsmaet al., 2001a,b; Li and Logan, 2004). However, it was found thatafter 12 h of incubation, S. guttiformis cells were able to attachonto an apparently ‘unfriendly surface’ by the production of EPS. Afew supporting arguments are worth noting. For example,Korenevsky and Beveridge (2007) while studying the surfaceand the adhesive properties of a number of Shewanella species,suggested that the cells adhered to hydrophobic surfaces due tothe presence of different surface proteins and/or EPS. The authorssuggested the possibility of the surface molecules undergoingconstant dynamic motion. In the light of numerous observationsthat not only the cell surface characteristics, the substrate surfacewettability and other parameters are important factors affectingbacterial adhesion as they dictate the composition of adsorbedmaterials (Norde and Lyklema, 1989; Beech et al., 1991, 1999;Skilman et al., 1999; Joseph and Wright, 2004), it can be arguedthat P(tBMA) surface might influence the S. guttiformis cells’metabolic activity where over the 12 h period S. guttiformis cellswere able to initiate the production of some specific EPS. Since in S.
guttiformis case, one type of EPS was effectively labeled byconcanavalin A, it is inferred that this type of S. guttiformis EPScontained mannose or glucose or both. Therefore, this EPS couldpositively contribute to the attachment of S. guttiformis cells inlight of finding that Vibrio cholerae and Pseudomonas aeruginosa
EPSs as composed of neutral sugars, glucose and galactose,increased the propensity of attachment of bacterial cells (Yildizand Schoolnik, 1999).
In summary, our data provide evidence that the physicochem-ical cell surface characteristics such as cell surface hydrophobicityand electrostatic charge did not substantially reduce the attach-ment propensity of S. guttiformis to P(tBMA). The EPS produced byS. guttiformis modulated the attachment pattern by enablingpolymeric surface colonization.
Acknowledgements
We thank Mr. Hans Brinkies for SEM analysis. This study wasfunded in part by Australian Research Council (ARC).
References
Auerbach, I.D., Sorensen, C., Hansma, H.G., Holden, P.A., 2000. Physical morphologyand surface properties of unsaturated Pseudomonas putida biofilms. J. Bacteriol.182 (13), 3809–3815.
Beech, I.B., Gaylard, C.C., 1991. Microbial polysaccharides and corrosion. Int. Bio-deterior. 27, 95–107.
Beech, B.I., Gaylard, C.C., Smith, J.J., Geesey, G.G., 1991. Extracellular polysaccharidesfrom Desulfovibrio desulfuricans and Pseudomonas fluorescens in the presence ofmild and stainless steel. Appl. Microbiol. Biotechnol. 25, 65–71.
Beech, B.I., Hanjagsit, L., Kalaji, M., Neal, A.L., Zinkevich, V., 1999. Chemical andstructural characterization of exopolymers produced by Pseudomonas sp.NCIMB 2021 in continuous culture. Microbiology 145 (6), 1491–1497.
Beech, B.I., Sunner, A.J., Hiraoka, K., 2005. Microbe–surface interactions in biofould-ing and biocorrosion processes. Int. Microbiol. 8, 157–168.
Bos, R., van der Mei, H.C., Busscher, H.J., 1999. Physico-chemistry of initial microbialadhesive interactions—its mechanisms and methods for study. FEMS Microbiol.Rev. 23, 179–230.
Bruinsma, G.M., Rustema-Abbing, M., van der Mei, H.C., Busscher, H.J., 2001a. Effectsof cell surface damage on surface properties and adhesion of Pseudomonasaeruginosa. J. Microbiol. Methods 45 (2), 95–101.
Bruinsma, G.M., van der Mei, H.C., Busscher, H.J., 2001b. Bacterial adhesion tosurface hydrophilic and hydrophobic contact lenses. Biomaterials 22, 3217–3224.
Buchan, A., Collier, L.S., Neidle, E.L., Moran, M.A., 2000. Key aromatic-ring-cleavingenzyme, protocatechuate 3,4-dioxygenase, in the ecologically important mar-ine Roseobacter lineage. Appl. Environ. Microbiol. 66 (11), 4662–4672.
Denes, A.R., Somers, E.B., Wong, A.C.L., Denes, F., 2001. 12-Crown-4-ether andtri(ethylene glycol) dimethyl-ether plasma-coated stainless steel surfacesand their ability to reduce bacterial biofilm deposition. J. Appl. Polym. Sci.81, 3425–3438.
Dong, H., Onstotta, T.C., Kob, C.-H.A., Hollingsworth, A.D., Brown, D.G., Mailloux, B.J.,2002. Theoretical prediction of collision efficiency between adhesion-deficientbacteria and sediment grain surface. Colloids Surf. B: Biointerfaces 24 (3–4),229.
Doronina, N.V., Trotsenko, Y.A., Tourova, T.P., 2000. Methylarcula marina gen. nov.,sp. nov. and Methylarcula terricola sp. nov.: novel aerobic, moderately halo-philic, facultatively methylotrophic bacteria from coastal saline environments.Int. J. Syst. Evol. Microbiol. 50 (5), 1849–1859.
Eboigbodin, E.K., Newton, A.R.J., Routh, F.A., Biggs, A.C., 2006. Bacterial quorumsensing and cell surface electrokinetic properties. Appl. Microbiol. Biotechnol.73, 669–675.
Francolini, I., Norris, P., Piozzi, A., Donelli, G., Stoodley, P., 2004. Usnic acid, a naturalantimicrobial agent able to inhibit bacterial biofilm formation on polymersurfaces. Antimicrob. Agents Chemother. 48 (11), 4360–4365.
Gibson, C.T., Watson, G.S., Myhra, S., 1996. Determination of the spring constants ofprobes for force microscopy/spectroscopy. Nanotechnology 7 (259–262).
Gonzalez, J.M., Mayer, F., Moran, M.A., Hodson, R.E., Whitman, W.B., 1997. Sagittulastellata gen. nov., sp. nov., a lignin-transforming bacterium from a coastalenvironment. Int. J. Syst. Bacteriol. 47 (3), 773–780.
Gonzalez, J.M., Covert, J.S., Whitman, W.B., Henriksen, J.R., Mayer, F., Scharf, B.,Schmitt, R., Buchan, A., Fuhrman, J.A., Kiene, R.P., Moran, M.A., 2003. Silicibacterpomeroyi sp. nov. and Roseovarius nubinhibens sp. nov., dimethylsulfoniopro-pionate-demethylating bacteria from marine environments. Int. J. Syst. Evol.Microbiol. 53 (5), 1261–1269.
Grafstrom, S., Neitezt, M., Hagen, T., Ackermann, J., Neumann, R., Probst, O., Wortge,M., 1993. The role of topography and friction for the image contrast in lateralforce microscopy. Nanotechnology 4, 143–151.
Hahner, A., Marti, A., Spencer, N.D., 1997. The influence of pH on friction betweenoxide surfaces in electrolytes studied with lateral force microscopy: applicationas a nanochemical imaging technique. Tribol. Lett. 3, 359–365.
Hall-Stoodley, L., Costerton, J.W., Stoodley, P., 2004. Bacterial biofilms: from thenatural environment to infectious diseases. Nat. Rev. Microbiol. 2, 95–108.
Ivanova, E.P., Papiernik, M., Oliveira, A., Sbarski, I., Smekal, T., Grodzinski, P., Nicolau,D.V., 2002a. Feasibility of using carboxylic-rich polymeric surfaces for thecovalent binding of oligonucleotides for micropcr applications. Smart Mater.Struct. 11 (5), 783–791.
Ivanova, E.P., Pham, D.K., Wright, P.J., Nicolau, D.V., 2002b. Detection of coccoidforms of Sulfitobacter mediterraneus using atomic force microscopy. FEMSMicrobiol. Lett. 214 (2), 177–181.
Ivanova, E.P., Wright, J.P., Pham, D.K., Brack, N., Pigram, P., Alekseeva, Y.V., Demya-shev, G.M., Nicolau, D.V., 2006. A comparative study between the adsorptionand covalent binding of human immunoglobulin and lysozyme on surface-modified poly(tert-butyl methacrylate). Biomed. Mater. 1, 24–32.
Jones, J.F., Velegol, D., 2006. Laser trap studies of end-on E. coli adhesion to glass.Colloids Surf. B: Biointerfaces 50 (1), 66–71.
Jonsson, U., Malmqvist, M., Ronnberg, I., 1985. Adsorption of immunoglobulin g,protein a, and fibronectin in the submonolayer region evaluated by a combinedstudy of ellipsometry and radiotracer techniques. J. Colloid Interface Sci. 103(2), 360–372.
Jorand, F., Boue-Bigne, F., Block, J.C., Urbain, V., 1988. Hydrophobic/hydrophilicproperties of activated sludge exopolymeric substances. Water Sci. Technol. 37,307–315.
Joseph, L.A., Wright, A.C., 2004. Expression of Vibrio vulnificus capsular polysacchar-ide inhibits biofilm formation. J. Bacteriol. 186 (3), 889–893.

E.P. Ivanova et al. / Micron 39 (2008) 1197–12041204
Korenevsky, A., Beveridge, T.J., 2007. The surface physicochemistry and adhesive-ness of Shewanella are affected by their surface polysaccharides. Microbiology153 (6), 1872–1883.
Kotra, L.P., Golemi, D., Amro, N.A., Liu, G.-Y., Mobashery, S., 1999. Dynamics of thelipopolysaccharide assembly on the surface of Escherichia coli. J. Am. Chem. Soc.121, 8707–8711.
Li, B., Logan, B.E., 2004. Bacterial adhesion to glass and metal-oxide surfaces.Colloids Surf. B: Biointerfaces 36 (2), 81–90.
Li, B., Logan, B.E., 2005. The impact of ultraviolet light on bacterial adhesion to glassand metal oxide-coated surface. Colloids Surf. B: Biointerfaces 41 (2–3), 153–161.
Marshall, K.C., Stout, R., Mitchell, R., 1971. Mechanisms of the initial events in thesorption of marine bacteria to surfaces. J. Gen. Microbiol. 68, 337–348.
Mitik-Dineva, N., Wang, J., Mocanasu, C.R., Stoddart, R.P., Crawford, J.R., Ivanova,E.P., 2008. Impact of nano-topography on bacterial attachment. Biotechnol. J. 3,1–9.
Neu, T.M., Poralla, K., 1988. An amphiphilic polysaccharide from adhesive Rhodo-coccus strain. FEMS Microbiol. Lett. 49, 389–392.
Norde, W., Lyklema, J., 1989. Protein adsorption and bacterial adhesion to solidsurfaces: a colloid-chemical approach. J. Colloid Surf. 38, 1–13.
Noy, A., Frisbie, C.D., Rozsnyaim, L.F., Wrighton, M.S., Lieber, C.M., 1995. Chemicalforce microscopy—exploiting chemically modified tips to quantify adhesion,friction, and functional-group distributions in molecular assemblies. J. Am.Chem. Soc. 117, 7943–7951.
Parsek, M.R., Singh, P.K., 2003. Bacterial biofilms: an emerging link to diseasepathogenesis. Ann. Rev. Microbiol. 57 (1), 677–701.
Pasmore, M., Costerton, J.W., 2003. Biofilms, bacterial signaling, and their ties tomarine biology. J. Ind. Microbiol. Biotechnol. 30, 407–413.
Raczkowska, J., Bernasik, A., Budkowski, A., Sajewicz, K., Penc, B., Lekki, J., Lekka, M.,Rysz, J., Kowalski, K., Czuba, P., 2004. Structures formed in spin-cast films ofpolystyrene blends with poly(butyl methacrylate) isomers. Macromolecules 37(19), 7308–7315.
Rappe, M.S., Vergin, K., Giovannoni, S.J., 2000. Phylogenetic comparisons of a coastalbacterioplankton community with its counterparts in open ocean and fresh-water systems. FEMS Microbiol. Ecol. 33 (3), 219–232.
Razatos, A., 2001. Application of atomic force microscopy to study initial events ofbacterial adhesion. Methods Enzymol. 337, 276–285.
Rijnaarts, H.H.M., Norde, W., Bouwer, E.J., Lyklema, J., Zehnder, A.J.B., 1995. Rever-sibility and mechanism of bacterial adhesion. Colloids Surf. B: Biointerfaces 4,5–22.
Rijnaarts, H.H.M., Norde, W., Lyklema, J., Zehnder, A.J.B., 1999. Dlvo and stericcontributions to bacterial deposition in media of different ionic strengths.Colloids Surf. B: Biointerfaces 14, 179–195.
Senaratne, W., Andruzzi, L., Ober, C.K., 2005. Self-assembled monolayers andpolymer brushes in biotechnology: current applications and future perspec-tives. Biomacromolecules 6 (5), 2427–2448.
Skilman, L.C., Suthereland, I.W., Jones, M.V., 1999. The role of exopolysaccharides indual species biofilm development. J. Appl. Microbiol. 85, S13–S18.
Sorokin, D.Y., 1995. Sulfitobacter pontiacus gen. nov., sp. nov.—a new heterotrophicbacterium from the black sea, specialized on sulfite oxidation. Mikrobiologiya64, 354–365.
Staley, J.T., 1968. Prosthecomicrobium and Ancalomicrobium: new prosthecate fresh-water bacteria. J. Bacteriol. 95 (5), 1921–1942.
Sutherland, I.W., 2001. Biofilm exppolysaccharides: a strong and sticky framework.Microbiol. Mol. Biol. Rev. 147, 3–9.
Tollersrud, T., Berge, T., Andersen, S.R., Lund, A., 2001. Imaging the surface ofStaphylococcus aureus by atomic force microscopy. APMIS 109, 541–545.
van der Aa, B.C., Michel, R.M., Asther, M., Torrez Zamora, M., Rouxhet, P.G., Dufrene,Y.F., 2001. Stretching cell surface macromolecules by atomic force microscopy.Langmuir 17 (11), 3116–3119.
van der Mei, H.C., Busscher, H.J., 2001. Electrophoretic mobility distributions ofsingle-strain microbial populations. Appl. Environ. Microbiol. 67 (2), 491–494.
Vogler, E.A., 1998. Structure and reactivity of water at biomaterial surfaces. Adv.Colloid Interface Sci. 74 (1–3), 69–117.
Wagner-Dobler, I., Rheims, H., Felske, A., Pukall, R., Tindall, B.J., 2003. Jannaschiahelgolandensis gen. nov., sp. nov., a novel abundant member of the marineRoseobacter clade from the north sea. Int. J. Syst. Evol. Microbiol. 53, 731–738.
Watson, G.S., Blach, J.A., Nicolau, D.V., Pham, D.K., Wright, J.P., Myhra, S., 2003.Surface topography and surface chemistry of radiation-patterned p(tBuMA)-Analysis by AFM. Polym. Int. 52, 1408–1414.
Wilburg, J.L., Biebuyck, H.A., MacDonald, J.C., Whitesides, G.M., 1995. Scanning forcemicroscopies can image patterned self-assembled monolayers. Langmuir 11,825–831.
Wright, J., Ivanova, E.P., Pham, D., Filipponi, L., Viezzoli, A., Suyama, K., Shirai, M.,Tsunooka, M., Nicolau, D.V., 2003. Positive and negative tone protein patterningon a photobase generating polymer. Langmuir 19 (2), 446–452.
Yildiz, F.H., Schoolnik, G.K., 1999. Vibrio cholerae o1 el tor: identification of a genecluster required for the rugose colony type, exopolysaccharide production,chlorine resistance, and biofilm formation. Proc. Natl. Acad. Sci. U.S.A. 96,4028–4033.
Related Documents











