Stable transfection of Eimeria tenella: Constitutive expression of the YFP-YFP molecule throughout the life cycle Wenchao Yan a , Xianyong Liu a , Tuanyuan Shi a , Lili Hao a , Fiona M. Tomley b , Xun Suo a, * a Parasitology Laboratory, College of Veterinary Medicine, China Agricultural University, Beijing 100193, PR China b Division of Microbiology, The Institute for Animal Health, Compton, Berkshire RG20 7NN, UK article info Article history: Received 28 January 2008 Received in revised form 15 May 2008 Accepted 3 June 2008 Keywords: Eimeria tenella Stable transfection Reproduction Immunogenicity abstract The obligate intracellular apicomplexan parasite Eimeria tenella, one of seven species of Eimeria that infect chickens, elicits protective cell-mediated immunity against challenge infection. For this reason, recombinant E. tenella parasites could be utilised as an effective vaccine vehicle for expressing foreign antigens and inducing immunity against heterologous intracellular microbes. A stable line of E. tenella expressing foreign genes is a prerequisite, and in this work an in vivo stable transfection system has been developed for this parasite using restriction enzyme-mediated integration (REMI). Two transgenic popu- lations of E. tenella have been obtained that express YFP-YFP constitutively throughout the parasite life cycle. Southern blotting and plasmid rescue analyses show that the introduced exogenous DNA was inte- grated at random into the parasite genome. Although the life cycle of the transgenic populations was delayed by at least 12 h and the output of oocysts was reduced 4-fold relative to the parental BJ strain of E. tenella, the transgenic parasites were sufficiently immunogenic to protect chickens against challenge with either transgenic or parental parasites. These results are encouraging for the development of trans- genic E. tenella as a vaccine vector and for more detailed investigation of the biology of the genus Eimeria. Ó 2008 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved. 1. Introduction Avian coccidiosis is considered to be one of the most economi- cally important diseases of domestic poultry, inflicting economic losses in excess of £2 billion annually to the world poultry industry (Shirley, 2000). Conventional disease control strategies rely pri- marily on prophylactic chemotherapy; unfortunately the Eimeria parasites that cause coccidiosis have developed resistance to most of the anticoccidial drugs that are used in chicken flocks (Williams, 1998). Alternative means of control have been focused on vaccina- tion with either virulent (Coccivac Ò and Immucox Ò ) or attenuated (Paracox Ò and Livacox Ò ) live parasite formulations and these have been used successfully to protect breeder, layer and broiler flocks (Danforth, 1998; Shirley, 2000; Suo et al., 2006). The immunogenic components of current coccidiosis vaccines are the developing intracellular parasites that multiply in the intestinal epithelial cells of chickens and which elicit solid protec- tive immunity against challenge infection (Lillehoj and Choi, 1998; Yun et al., 2000). Previous studies have indicated that the cellular effectors of protective immune responses to primary and challenge coccidial infections are primarily CD4 + and CD8 + T cells residing in gut-associated lymphoid tissues. Furthermore, CD8 + T cells can kill parasite-infected cells and protective immunity can be abolished by depletion of CD8 + T cells in challenge infections (Trout and Lil- lehoj, 1996; Lillehoj and Choi, 1998). CD4 + Th1 lymphocytes pro- vide help for the CD8 + T cell-dependent protective response to coccidiosis (Trout and Lillehoj, 1996; Shedlock and Shen, 2003). Eimeria infections also induce high levels of IFN-c, IL-2 and IL-12 (Hong et al., 2006; Digby and Lowenthal, 1995), and IFN-c stimu- lates macrophages to produce nitric oxide (NO), which inhibits replication of Eimeria tenella within host cells (Lillehoj and Choi, 1998). Accumulated evidence shows that Th1 cytokines and CD8 + cytotoxic cells are required for efficient protection against various intracellular pathogens (Sharma, 1991). Based on the above prop- erties, transgenic E. tenella as a live vaccine vector might be ex- pected to induce strong Th1 cytokines and CD8+ T cell responses against heterologous antigens that are expressed, and could be a powerful live vaccine vector system for heterologous intracellular gastroenteric microorganisms. A stable line of parasites expressing target foreign genes is a prerequisite for establishing a live vaccine vector system based on E. tenella for the control of other major avian diseases. Genetic manipulation is also a powerful tool for investigating the biology of the apicomplexa, including E. tenella. Transient and stable trans- fection systems are well established in Toxoplasma gondii and Plas- modium spp. (Goonewardene et al., 1993; Soldati and Boothroyd, 1993; Sibley et al., 1994; van Dijk et al., 1995) and in vitro transient transfection systems have been developed for E. tenella (Kelleher and Tomley, 1998; Hao et al., 2007). Now we report an in vivo sta- 0020-7519/$34.00 Ó 2008 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.ijpara.2008.06.013 * Corresponding author. Tel./fax: +86 10 62734325. E-mail address: [email protected] (X. Suo). International Journal for Parasitology 39 (2009) 109–117 Contents lists available at ScienceDirect International Journal for Parasitology journal homepage: www.elsevier.com/locate/ijpara

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

International Journal for Parasitology 39 (2009) 109–117
Contents lists available at ScienceDirect
International Journal for Parasitology
journal homepage: www.elsevier .com/locate / i jpara
Stable transfection of Eimeria tenella: Constitutive expression of the YFP-YFPmolecule throughout the life cycle
Wenchao Yan a, Xianyong Liu a, Tuanyuan Shi a, Lili Hao a, Fiona M. Tomley b, Xun Suo a,*
a Parasitology Laboratory, College of Veterinary Medicine, China Agricultural University, Beijing 100193, PR Chinab Division of Microbiology, The Institute for Animal Health, Compton, Berkshire RG20 7NN, UK
a r t i c l e i n f o
Article history:Received 28 January 2008Received in revised form 15 May 2008Accepted 3 June 2008
Keywords:Eimeria tenellaStable transfectionReproductionImmunogenicity
0020-7519/$34.00 � 2008 Australian Society for Paradoi:10.1016/j.ijpara.2008.06.013
* Corresponding author. Tel./fax: +86 10 62734325.E-mail address: [email protected] (X. Suo).
a b s t r a c t
The obligate intracellular apicomplexan parasite Eimeria tenella, one of seven species of Eimeria thatinfect chickens, elicits protective cell-mediated immunity against challenge infection. For this reason,recombinant E. tenella parasites could be utilised as an effective vaccine vehicle for expressing foreignantigens and inducing immunity against heterologous intracellular microbes. A stable line of E. tenellaexpressing foreign genes is a prerequisite, and in this work an in vivo stable transfection system has beendeveloped for this parasite using restriction enzyme-mediated integration (REMI). Two transgenic popu-lations of E. tenella have been obtained that express YFP-YFP constitutively throughout the parasite lifecycle. Southern blotting and plasmid rescue analyses show that the introduced exogenous DNA was inte-grated at random into the parasite genome. Although the life cycle of the transgenic populations wasdelayed by at least 12 h and the output of oocysts was reduced 4-fold relative to the parental BJ strainof E. tenella, the transgenic parasites were sufficiently immunogenic to protect chickens against challengewith either transgenic or parental parasites. These results are encouraging for the development of trans-genic E. tenella as a vaccine vector and for more detailed investigation of the biology of the genus Eimeria.
� 2008 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Avian coccidiosis is considered to be one of the most economi-cally important diseases of domestic poultry, inflicting economiclosses in excess of £2 billion annually to the world poultry industry(Shirley, 2000). Conventional disease control strategies rely pri-marily on prophylactic chemotherapy; unfortunately the Eimeriaparasites that cause coccidiosis have developed resistance to mostof the anticoccidial drugs that are used in chicken flocks (Williams,1998). Alternative means of control have been focused on vaccina-tion with either virulent (Coccivac� and Immucox�) or attenuated(Paracox� and Livacox�) live parasite formulations and these havebeen used successfully to protect breeder, layer and broiler flocks(Danforth, 1998; Shirley, 2000; Suo et al., 2006).
The immunogenic components of current coccidiosis vaccinesare the developing intracellular parasites that multiply in theintestinal epithelial cells of chickens and which elicit solid protec-tive immunity against challenge infection (Lillehoj and Choi, 1998;Yun et al., 2000). Previous studies have indicated that the cellulareffectors of protective immune responses to primary and challengecoccidial infections are primarily CD4+ and CD8+T cells residing ingut-associated lymphoid tissues. Furthermore, CD8+ T cells can killparasite-infected cells and protective immunity can be abolished
sitology Inc. Published by Elsevier
by depletion of CD8+ T cells in challenge infections (Trout and Lil-lehoj, 1996; Lillehoj and Choi, 1998). CD4+ Th1 lymphocytes pro-vide help for the CD8+ T cell-dependent protective response tococcidiosis (Trout and Lillehoj, 1996; Shedlock and Shen, 2003).Eimeria infections also induce high levels of IFN-c, IL-2 and IL-12(Hong et al., 2006; Digby and Lowenthal, 1995), and IFN-c stimu-lates macrophages to produce nitric oxide (NO), which inhibitsreplication of Eimeria tenella within host cells (Lillehoj and Choi,1998). Accumulated evidence shows that Th1 cytokines and CD8+
cytotoxic cells are required for efficient protection against variousintracellular pathogens (Sharma, 1991). Based on the above prop-erties, transgenic E. tenella as a live vaccine vector might be ex-pected to induce strong Th1 cytokines and CD8+ T cell responsesagainst heterologous antigens that are expressed, and could be apowerful live vaccine vector system for heterologous intracellulargastroenteric microorganisms.
A stable line of parasites expressing target foreign genes is aprerequisite for establishing a live vaccine vector system basedon E. tenella for the control of other major avian diseases. Geneticmanipulation is also a powerful tool for investigating the biologyof the apicomplexa, including E. tenella. Transient and stable trans-fection systems are well established in Toxoplasma gondii and Plas-modium spp. (Goonewardene et al., 1993; Soldati and Boothroyd,1993; Sibley et al., 1994; van Dijk et al., 1995) and in vitro transienttransfection systems have been developed for E. tenella (Kelleherand Tomley, 1998; Hao et al., 2007). Now we report an in vivo sta-
Ltd. All rights reserved.

110 W. Yan et al. / International Journal for Parasitology 39 (2009) 109–117
ble transfection of E. tenella using a restriction enzyme-mediatedintegration strategy.
2. Materials and methods
2.1. Parasite and cell culture
The BJ strain of E. tenella was used in this study (Hao et al.,2007) and was maintained by passage through coccidia-free 2-week-old Arbor Acre (AA) broiler chickens. Oocysts were collectedfrom the faeces of chickens 6–9 days after oral infection. Proce-dures for isolation, purification and sporulation of oocysts of E.tenella have been described previously (Long et al., 1976). Sporozo-ites were excysted with a modified excystation buffer (PBS pH 7.6,containing 0.75% trypsin and 10% chicken bile extract) and purifiedby DE-52 anion-exchange chromatography (Schmatz et al., 1984).
Primary chicken kidney (PCK) cells were used for the propaga-tion of E. tenella sporozoites in in vitro transient transfection exper-iments. Kidneys were removed from 1- to 3-week-old AA broilerchickens and PCK cells prepared using a procedure described pre-viously (Doran, 1970; Hofmann and Raether, 1990; Tierney andMulcahy, 2003). PCK cells in nutrient DMEM culture medium (pH7.4) supplemented with L-glutamine, 10% FCS, penicillin(200 U mL�1) and streptomycin (20 mg mL�1) were incubated at41 �C in an atmosphere of 5% CO2 for 48–72 h. After the PCK cellsreached approximately 80% confluency, the medium in each flaskwas removed, and 1 mL of fresh medium (pH 7.4) containing1 � 107 sporozoites was added.
All chicken experimentation was carried out in compliance withthe current Chinese legislation and due attention was paid to thewelfare of experimental chickens.
2.2. Plasmid constructs
Plasmids for expression of a tandem yellow fluorescent protein(YFP-YFP) and dihydrofolate reductase thymidylate synthase de-rived from Toxoplasma gondii (TgDHFR-TS) (kindly provided by Pro-
Fig. 1. Schematic representation of plasmids used for expression of YFP-YFP and mutangene and the EtACTIN gene were used to drive transcription of foreign genes.
Table 1Oligonucleotides used in vector constructs
Name of primers (forward primer/reverseprimer)
Forward primera
P1/P2 50aatGGATCCAAACGAACGAGCTCGTAG30
P3/P4 50atgGCGGCCGCGTTTGCAGCAGAGTAGT30
P5/P6 50cggACTAGTGCCCGTGATCCTGTCG 30
P7/P8 50acgcGGATCCTTCGTTTTCTCAAATGC30
P9/P10 50acccAAGCTTATGCAGAAACCGGTGTGTC30
a Underlined bp designate restriction endonuclease sites within primer sequences; loprimers.
fessor David Roos) are shown in Fig. 1 and the oligonucleotidesused in PCR are listed in Table 1.
Approximately 1000 bp of 50 flanking sequence and 90 bp of thetranslated sequence of the HISTONE4 gene of E. tenella (EtHI-STONE4) were amplified using a forward primer P1 that created aBamHI site and a reverse primer P2 that added a SpeI site. Theresulting PCR product was digested with BamHI and SpeI andcloned into BamHI/SpeI-digested plasmid pUC19-M to obtain anintermediate vector pEtH. Approximately 800 bp of the 30 untrans-lated region (UTR) of the ACTIN gene of E. tenella (EtACTIN) wereamplified with primers P3/P4 that created NotI and SacII sitesand the digested product was cloned into NotI and SacII-digestedpEtH to make another intermediate plasmid pEtHA. A tandemYFP-YFP sequence was amplified from plasmid pHYA (Hao et al.,2007) with primers P5/P6 that contained SpeI and NotI sites andthe product cloned into SpeI and NotI-digested pEtHA to producethe final expression vector pEtHEA. The EtHISTONE4 promoterand the EtACTIN 30 UTR in pEtHEA were shorter than in the previ-ously described vector pHYA (Hao et al., 2007).
The EtHISTONE4 promoter in pEtHEA was excised with BamHIand HindIII, and replaced with a 50 flanking region of EtACTINamplified with primers P7/P8 that contained BamHI and HindIIIsites to make another reporter vector pEtAEA.
To generate the selectable vector pEtADA, the TgDHFR-TS openreading frame (ORF) was amplified from pDHFR-TSc3 (kindly pro-vided by Professor David Roos) using primers P9/P10 that createdHindIII and NotI sites. The amplification product was digested withHindIII and NotI and cloned to replace the YFP-YFP ORF of pEtAEA.All PCR amplifications were performed using the high fidelity ther-mostable Pfu DNA polymerase to reduce mutation frequency.
2.3. Parasite transfection
For transfection, freshly purified E. tenella sporozoites wereresuspended in cytomix buffer supplemented with 2 mM ATPand 5 mM glutathione (van den Hoff and Lamers, 1992). Transfec-tions were performed using the restricted enzyme-mediated inte-
t TgDHFR-TS molecules in Eimeria tenella. The 50 flanking regions of the EtHISTONE4
Reverse primera Fragment size(bp)
Name offragment
50cgtACTAGTGATACCCTGGATGTTGTC30 1090 EtHISTON 4 50
50atcCCGCGGCTTCACATGGAACCCCTGG30 800 EtACTIN 30
50attGCGGCCGCCTAAAGC30 1496 YFP-YFP50acccAAGCTTATCATCACCTGCAACTC30 1200 EtACTIN 50
50ataagaatGCGGCCGCCTAGACAGCCATCTCCATC30 1833 TgDHFR-TS
wercase bp show protective bp for restriction enzyme sites at the 50 ends of these

W. Yan et al. / International Journal for Parasitology 39 (2009) 109–117 111
gration (REMI) technique, as described previously (Black et al.,1995). Briefly, a 40–60 lg aliquot of plasmid DNA, linearised withBamHI and phenol extracted to eliminate residual enzymatic activ-ity, was mixed with 2 � 107 sporozoites in a total volume of800 lL. In REMI transfections, 200 U of BamHI were immediatelyadded to the electroporation cuvette containing the parasites andlinearised DNA fragments possessing corresponding enzymic endsprior to electroporation. All electroporations were performed usinga Gene Pulser X cellTM.
Electroporation System (Bio-Rad, Hercules, California) set at2.0 kV, 25 mF, resulting in a pulse time of 0.3–0.4 ms. Electropora-ted parasites were allowed to recover for 20 min at room temper-ature and then transferred to a confluent monolayer of PCK cells fortransient transfection, or inoculated into the ileocaecal opening of4-day-old chickens via the cloacal route for the in vivo stabletransfection.
2.4. Microscopic observation of YFP- expressing parasites
For YFP-expressing sporozoites in vitro, parasites were incu-bated at 41 �C in 5% CO2 in PCK cells for 16 h after electroporationand then examined under a fluorescent microscope (OlympusIX71, Tokyo, Japan). In the in vivo transfection experiments, thecaecal mucous membranes of infected chickens were scraped andviewed under a fluorescent microscope to find various fluorescentendogenous stages of E. tenella at 120 h p.i. Oocysts in faeces ex-creted 6–9 days p.i. were further checked by fluorescencemicroscopy.
To determine the transfection efficiency of E. tenella in vivo, thetotal numbers of oocysts collected from faeces were counted usingthe McMaster method (Jeffers, 1975). The total oocysts were ad-justed to a standard volume, a 5 mL sample added to a35 mm � 10 mm cell culture dish (BD FalconTM, Boston, US) andthe numbers of fluorescent oocysts counted using a fluorescentmicroscope. When large numbers of oocysts were present furtherdilutions were required. The transfection efficiency was calculatedas a percentage as ffollows:
f½Number of oocysts expressing YFP-YFP in 5mL sample
� 5� Total volume of oocyst suspensionðmLÞ�=Number of total oocysts collected from faecesg � 100
2.5. Pyrimethimine selection and fluorescence activated cell sorting
Approximately 2.0 � 106 transfected sporozoites were inocu-lated into the ileocaecal opening of 4-day-old chickens via the clo-aca. After 24 h p.i., these chickens were treated with 100 ppmpyrimethamine (Sigma–Aldrich Co., St. Louis, Mo., USA) in feedfor 8 days. Oocysts from faeces excreted 6–10 days p.i. were col-lected, surface sterilized by incubation with 10% sodium hypochlo-rite solution (Norton et al., 1983), filtered through a 40 lm nyloncell strainer (BD FalconTM, Boston, US) and resuspended in PBS.The fluorescent oocysts collected from faeces were sorted using aMoFlo Cell Sorter (Dako Cytomation, Fort Collins, CO, USA) usingthe single cell mode, and propagated in chickens. The transgenicE. tenella population obtained by co-transfection with the reportervector pEtHEA and drug selectable vector pEtADA was referred toas TE1, another obtained by transfection with a single plasmid pE-tAEA as TE2.
To accurately analyse the fluorescence intensity of TE1 and TE2populations, both 2 � 104 fluorescent oocysts of the sixth passageTE1 and the third passage TE2 populations were first obtained bysorting with a MoFlo Cell Sorter using the single cell mode. The rel-ative intensity of fluorescent protein expression in the sorted fluo-rescent oocysts of these two transgenic populations was then
estimated by the flow cytometer (BD FACSCalibur, Columbus, OH,USA).
2.6. Parasite ‘‘cloning” using a single oocyst isolation technique
Transgenic parasite ‘‘cloning” by the single oocyst isolationtechnique was performed from the fifth and tenth passage oocystsof the TE1 population as previously described (Lee, 1979). Briefly,the oocyst suspension was diluted in sterile PBS at pH 7.6 to a dilu-tion containing <1 oocyst/2 lL. A single fluorescent sporulated oo-cyst in each 2 lL of PBS was selected under the fluorescentmicroscope on the sterile agar plate and given to a 4-day-old chick-en via the crop. Faeces were examined for oocysts 7–10 days p.i.The isolated fluorescent oocysts from each chicken were consecu-tively propagated in five 7-day-old chickens to obtain enough oo-cysts for genomic DNA analysis by Southern blotting and theplasmid rescue method.
2.7. Genomic DNA analysis of transgenic parasites
An aliquot of 107 oocysts were ground in a mortar with liquidnitrogen and then suspended in 1 mL DNAzol reagent (Bioteke, Bei-jing, China). Genomic DNA was isolated from transgenic popula-tions following the manufacturer’s instructions. Genomic DNAs oftransgenic populations TE1, TE2 and of the parental BJ strain wereused as templates for PCR amplification to detect the presence ofintroduced DNA fragments in the transgenic parasites using thespecific primers for YFP and TgDHFR-TS, respectively.
Southern blotting analysis was carried out using a probe spe-cific for the YFP sequence. The probe preparation, hybridization,washing and detection followed the manufacturer’s guide of DIGHigh Prime DNA Labeling and Detection Starter Kit II (Roche, Basel,Switzerland). The YFP-specific probe was prepared in a 20 lL reac-tion system, including 1 lg denatured PCR product of YFP gene and4 lL DIG-High Prime. The mixture reacted at 37 �C overnight andthe resultant probe solution was heated at 65 �C for 10 min afteradding 2 lL 0.2 M EDTA (pH 8.0) to stop the reaction. Approxi-mately 3 lg genomic DNA samples of transgenic populations(TE1.1, TE1.2 and TE1.3) cloned using a single oocyst isolation tech-nique were digested with restriction enzyme EcoRV which doesnot cleave in the pEtHEA plasmid, overnight, electrophoresed in0.8% agarose gels, and transferred with 20-fold standard sodiumcitrate buffer (SSC) on positively charged nylon membrane (Amer-sham Biosciences, NJ, USA) by capillary as described before (Sam-brook and Russell, 2001). The transferred DNAs were fixed to themembranes by u.v.-crosslinking for 10 min. Then the membraneswere rinsed briefly in double distilled water. After 30 min prehy-bridization at 42 �C, the probe-hybridization mixture was preparedby adding 10 lL heat-denatured probe solution to 15 mL pre-heated DIG Easy Hyb solution. This mixture was then hybridizedat 42 �C for 6 h by rotating at 4 rpm. The resultant membraneswere washed for 2 � 5 min in 2� SSC (containing 0.1% SDS) atambient temperature, followed by another washing of 2 � 15 minin 0.5� SSC (containing 0.1% SDS) at 65 �C. Subsequently, immuno-logical detection was performed on the washing membranes in thefollowing steps: (i) 5 min rinse in washing buffer (0.1 M maleicacid, 0.15 M NaCl and 0.3% Tween 20, pH 7.5); (ii) 30 min incuba-tion in 20 mL blocking solution (provided by the kit); (iii) 30 minincubation in 20 mL antibody solution (prepared by adding 2 lLAnti-Digoxigenin-AP conjugate to 20 mL blocking solution); (iv)2 � 15 min washing in 50 mL washing buffer; (v) 5 min equilibra-tion in 20 mL detection buffer (0.1 M Tris–HCl and 0.1 M NaCl, pH9.5); (vi) apply 1-2 mL disodium 3-(4-methoxyspiro {1,2-dioxe-tane-3; 20-(50-chloro) tricyclo [3.3.1.1] decan}-4-yl) phenyl phos-phate (CSPD) ready-to-use to the side of membranes with DNA,and incubate the damp membrane for 10 min at 37 �C; (vii) multi-

112 W. Yan et al. / International Journal for Parasitology 39 (2009) 109–117
ple exposures of the membrane to X-ray film for 1–30 min at ambi-ent temperature to achieve the desired signal strength.
The plasmid rescue method was applied to further study theintegration events in the genome of the TE1 population and oneof the TE1 populations cloned (TE1.2) by single oocyst isolationtechnique. Genomic DNAs were digested overnight with therestriction enzyme EcoRV, which does not cleave in plasmidspEtHEA and pEtADA. The digested purified DNA was diluted 10-fold and religated, the ligated products were used to transformcompetent DH5a cells and selected transformants by growth in100 lg/mL ampicillin in Luria–Bertani (LB) culture medium. Theclones were identified by colony PCR and digestion with EcoRV. Fi-nally, the positive clones for pEtHEA or pEtADA were sequencedwith inverted specific primers within the transfected plasmids(S1: AAGTAGGAGGACCTTAGTGCGT for pEtHEA clones, S2: TAGCTGGAGTGGGTTTGAGT for pEtADA clones, S3: AGCATTTATCAGGGTTATTGTC for both pEtHEA and pEtADA clones), the se-quences obtained by sequencing the flanking regions of the trans-fected plasmids were analysed by DNAStar 5.01 and Clustal X 1.8softwares, and the insertion sites in the genome were identifiedby performing a BLAST search in the E. tenella DB database (Kimand Weiss, 2004).
2.8. Measurement of reproduction of transgenic parasites
To check the effect of over-expression and/or disruption of for-eign genes on the reproduction of transgenic parasites, the devel-opmental rate and reproductive potential of a transgenic E.tenella population (TE1) obtained in the stable transfection exper-iment (see Section 3.2) were compared with that of its parentalstrain BJ in three replicate experiments. In each replicate, differ-ences in the rate of development of endogenous stages were as-sessed by microscopic examination of caecal wall scrapings takenfrom the gut of four 10-day-old AA chickens sacrificed at 6 h inter-vals from 120 to 138 h following inoculation with 1 � 103 sporu-lated oocysts/bird. A further six chickens in each replicateexperiment were used to determine the rate of oocyst productionand fecundity of the transgenic population TE1, by counting thenumber of oocysts recovered in the faeces at 12 h intervals be-tween 120 and 240 h p.i., using the McMaster method as describedpreviously (Jeffers, 1975; Haug et al., 2006).
2.9. Immunogenicity test
AA broiler chickens were reared coccidia-free in isolators andthen transferred to wire-floored cages for experiments at 4 or 7days of age. The immunogenicity of a transgenic parasite popula-tion (TE1) was evaluated with two different immunization sched-ules (Table 3). In experiment A, chickens were inoculated oncewith 1 � 103 sporulated oocysts of the TE1 population or the BJstrain at 7 days of age. In experiment B, 4-day-old chickens wereinoculated with 2 � 102 oocysts of the TE1 population or the BJstrain, and boosted with the same dose at 18 days of age. Fourteendays after the primary or secondary inoculation of oocysts, thechickens and uninfected age-matched controls in experiments Aand B were challenged with 1 � 103 oocysts of BJ strain or TE1 par-asites. The immunogenicity of transgenic population TE1 was mea-sured by comparing the oocyst output per bird from immunizedand control birds after challenge with the parent or transgenic par-asites (Long and Rowell, 1958; Haug et al., 2006).
2.10. Statistical analysis
Statistical analysis was performed between the control andtreated groups and also amongst the different experimental groupsusing the Student’s t-test or one-way ANOVA in SPSS 11.5 for win-
dows software. Differences with a P value of <0.05 were consideredto be statistically significant.
3. Results
3.1. Transient expression of YFP-YFP in E. tenella
To demonstrate the feasibility of foreign gene expression in E.tenella, in vitro transient transfections were performed with vec-tors pEtHEA or pEtAEA. The correctness of the constructs was ver-ified by PCR, restriction enzyme digestion and sequencing.Electroporation conditions were based on values required for 40–65% survival of E. tenella sporozoites, determined by viabilitycounts using trypan blue dye exclusion (Kelleher and Tomley,1998).
At 19–72 h p.i., sporozoites and first generation schizontsexpressing YFP were observed in parasites transfected with bothpEtHEA and pEtAEA and both found to display high fluorescence.Sporozoites electroporated with pEtHEA expressed YFP-YFP whichwas located within the nucleus of the parasite (Fig. 2A), whilstYFP-YFP driven by the EtACTIN promoter produced uniform fluores-cence in sporozoites and developing schizonts, suggestingcytoplasmic localisation of the reporter protein (Fig. 2G). Similarlocalisations were observed in the in vivo transfection experi-ments. We suggest that N-terminal fusion of the 90 bp nuclearlocalisation sequence of EtHISTONE 4 or the 69 bp encoding theN-terminal sequence of EtACTIN results in the differential localisa-tion of YFP within the parasites (Liu et al., unpublished data). Inaddition, different localisation of YFP might be due to the timingof expression from the two different promoters (Bonilla et al.,2007). These data demonstrated that both transfection vectorspEtHEA and pEtAEA can successfully express YFP in E. tenella. Inparasites transfected with linearised plasmids, enumeration offluorescent organisms from two separate electroporations withpEtHEA and pEtAEA revealed that approximately 1.6 � 10�4 and4.04 � 10�5 of the parasite populations expressed YFP, respec-tively. Transfection efficiency using the REMI method is muchhigher than that of transfection with circular plasmid (Liu et al.,unpublished data).
3.2. Stable transfection of E. tenella
To establish stable transfection of E. tenella, a T. gondii DHFR-TSallele that confers resistance to pyrimethamine was cloned into theE. tenella transfection plasmid under transcriptional control of theEtACTIN promoter. The resulting vector pEtADA was confirmed byrestriction enzyme digestion and sequencing. The purified spor-ozoites were cotransfected with 40 lg pEtADA and 40 lg pEtHEA;transfected parasites were inoculated into 4-day-old chickens andthen 100 ppm pyrimethamine in feed was administered to infectedbirds from 24 h p.i. The microscopic observation revealed that0.04% of the E. tenella oocysts in faeces from these birds expressedYFP (Table 2). After the fluorescent oocysts were FACs sorted andfurther propagated in chickens, 95.7% of oocysts in the faeces ofinoculated chickens were found to highly express YFP-YFP at thefourth passage (Fig. 3A–C). At this point, pyrimethamine selectionwas withdrawn, although FACs sorting continued, and it was foundthat >95% of the shed oocysts expressed YFP-YFP at the fifth andsixth passage (Table 2). This transfected population of E. tenellawas referred to as TE1. No fluorescent oocysts were detected inthe faeces of chickens infected with mock transfected parasites.During initial experiments to try and generate cloned lines oftransgenic parasites, single oocyst infections were performed threetimes using oocysts from the fifth passage, into a total of 45 birdsbut unfortunately we failed to obtain a population. More recently,
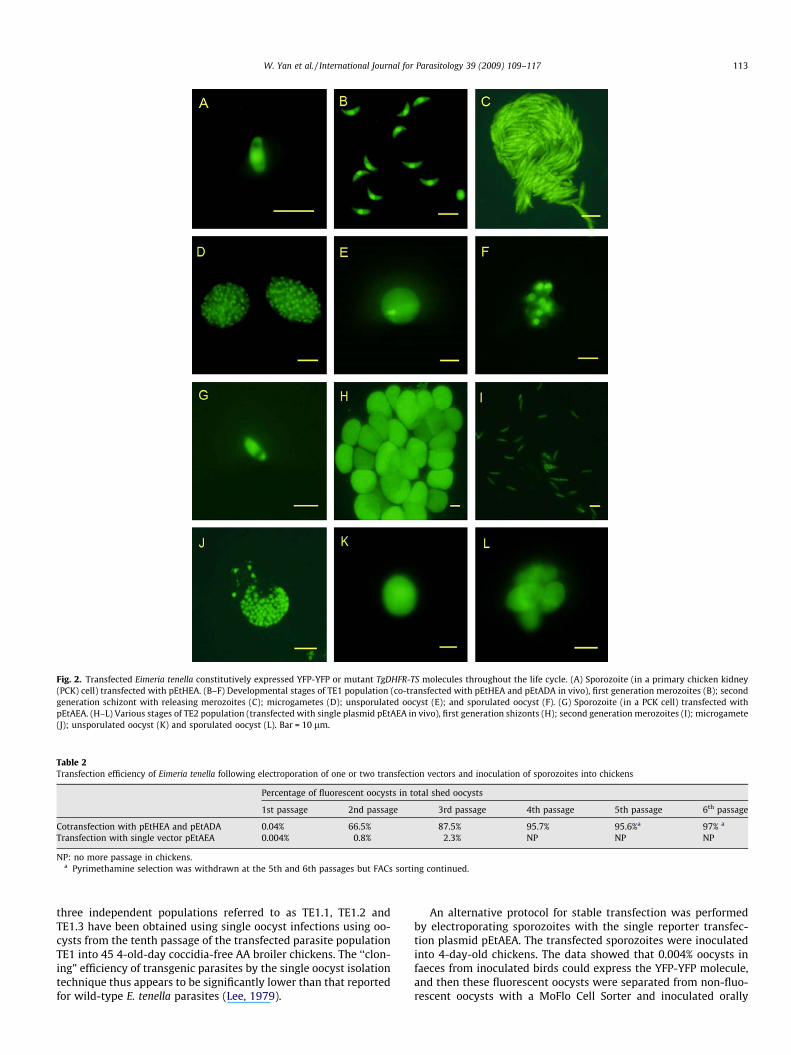
Fig. 2. Transfected Eimeria tenella constitutively expressed YFP-YFP or mutant TgDHFR-TS molecules throughout the life cycle. (A) Sporozoite (in a primary chicken kidney(PCK) cell) transfected with pEtHEA. (B–F) Developmental stages of TE1 population (co-transfected with pEtHEA and pEtADA in vivo), first generation merozoites (B); secondgeneration schizont with releasing merozoites (C); microgametes (D); unsporulated oocyst (E); and sporulated oocyst (F). (G) Sporozoite (in a PCK cell) transfected withpEtAEA. (H–L) Various stages of TE2 population (transfected with single plasmid pEtAEA in vivo), first generation shizonts (H); second generation merozoites (I); microgamete(J); unsporulated oocyst (K) and sporulated oocyst (L). Bar = 10 lm.
Table 2Transfection efficiency of Eimeria tenella following electroporation of one or two transfection vectors and inoculation of sporozoites into chickens
Percentage of fluorescent oocysts in total shed oocysts
1st passage 2nd passage 3rd passage 4th passage 5th passage 6th passage
Cotransfection with pEtHEA and pEtADA 0.04% 66.5% 87.5% 95.7% 95.6%a 97% a
Transfection with single vector pEtAEA 0.004% 0.8% 2.3% NP NP NP
NP: no more passage in chickens.a Pyrimethamine selection was withdrawn at the 5th and 6th passages but FACs sorting continued.
W. Yan et al. / International Journal for Parasitology 39 (2009) 109–117 113
three independent populations referred to as TE1.1, TE1.2 andTE1.3 have been obtained using single oocyst infections using oo-cysts from the tenth passage of the transfected parasite populationTE1 into 45 4-old-day coccidia-free AA broiler chickens. The ‘‘clon-ing” efficiency of transgenic parasites by the single oocyst isolationtechnique thus appears to be significantly lower than that reportedfor wild-type E. tenella parasites (Lee, 1979).
An alternative protocol for stable transfection was performedby electroporating sporozoites with the single reporter transfec-tion plasmid pEtAEA. The transfected sporozoites were inoculatedinto 4-day-old chickens. The data showed that 0.004% oocysts infaeces from inoculated birds could express the YFP-YFP molecule,and then these fluorescent oocysts were separated from non-fluo-rescent oocysts with a MoFlo Cell Sorter and inoculated orally
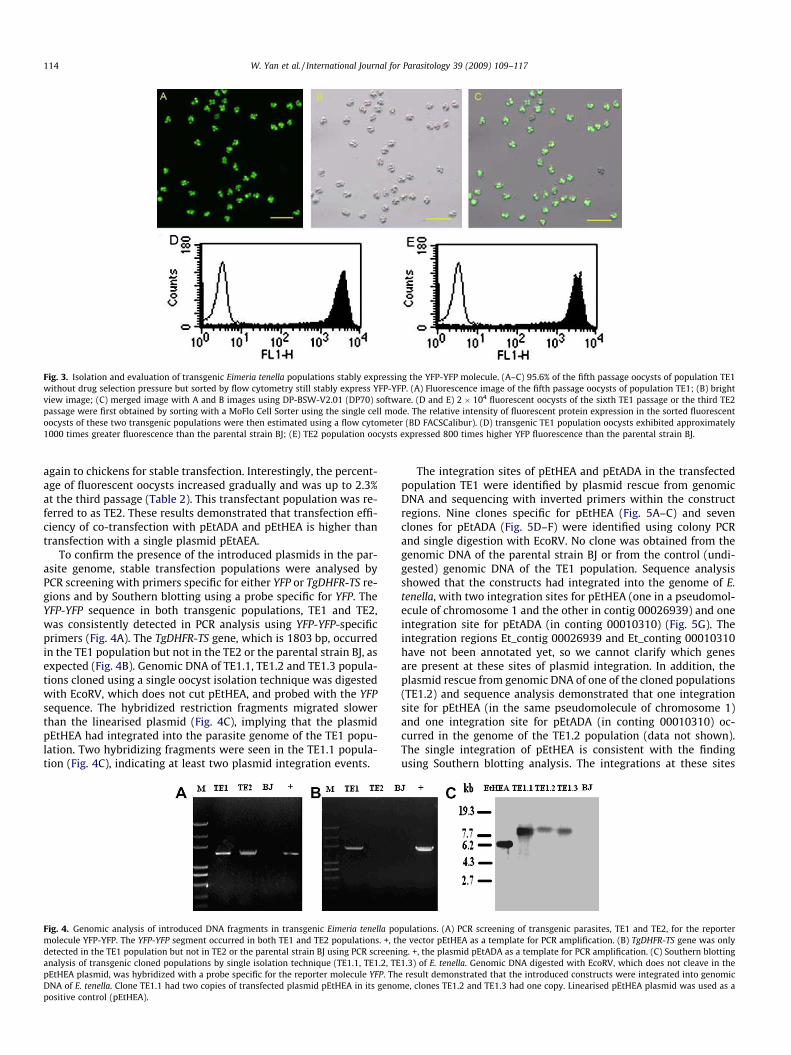
Fig. 3. Isolation and evaluation of transgenic Eimeria tenella populations stably expressing the YFP-YFP molecule. (A–C) 95.6% of the fifth passage oocysts of population TE1without drug selection pressure but sorted by flow cytometry still stably express YFP-YFP. (A) Fluorescence image of the fifth passage oocysts of population TE1; (B) brightview image; (C) merged image with A and B images using DP-BSW-V2.01 (DP70) software. (D and E) 2 � 104 fluorescent oocysts of the sixth TE1 passage or the third TE2passage were first obtained by sorting with a MoFlo Cell Sorter using the single cell mode. The relative intensity of fluorescent protein expression in the sorted fluorescentoocysts of these two transgenic populations were then estimated using a flow cytometer (BD FACSCalibur). (D) transgenic TE1 population oocysts exhibited approximately1000 times greater fluorescence than the parental strain BJ; (E) TE2 population oocysts expressed 800 times higher YFP fluorescence than the parental strain BJ.
114 W. Yan et al. / International Journal for Parasitology 39 (2009) 109–117
again to chickens for stable transfection. Interestingly, the percent-age of fluorescent oocysts increased gradually and was up to 2.3%at the third passage (Table 2). This transfectant population was re-ferred to as TE2. These results demonstrated that transfection effi-ciency of co-transfection with pEtADA and pEtHEA is higher thantransfection with a single plasmid pEtAEA.
To confirm the presence of the introduced plasmids in the par-asite genome, stable transfection populations were analysed byPCR screening with primers specific for either YFP or TgDHFR-TS re-gions and by Southern blotting using a probe specific for YFP. TheYFP-YFP sequence in both transgenic populations, TE1 and TE2,was consistently detected in PCR analysis using YFP-YFP-specificprimers (Fig. 4A). The TgDHFR-TS gene, which is 1803 bp, occurredin the TE1 population but not in the TE2 or the parental strain BJ, asexpected (Fig. 4B). Genomic DNA of TE1.1, TE1.2 and TE1.3 popula-tions cloned using a single oocyst isolation technique was digestedwith EcoRV, which does not cut pEtHEA, and probed with the YFPsequence. The hybridized restriction fragments migrated slowerthan the linearised plasmid (Fig. 4C), implying that the plasmidpEtHEA had integrated into the parasite genome of the TE1 popu-lation. Two hybridizing fragments were seen in the TE1.1 popula-tion (Fig. 4C), indicating at least two plasmid integration events.
Fig. 4. Genomic analysis of introduced DNA fragments in transgenic Eimeria tenella pomolecule YFP-YFP. The YFP-YFP segment occurred in both TE1 and TE2 populations. +, thdetected in the TE1 population but not in TE2 or the parental strain BJ using PCR screenianalysis of transgenic cloned populations by single isolation technique (TE1.1, TE1.2, TEpEtHEA plasmid, was hybridized with a probe specific for the reporter molecule YFP. ThDNA of E. tenella. Clone TE1.1 had two copies of transfected plasmid pEtHEA in its genompositive control (pEtHEA).
The integration sites of pEtHEA and pEtADA in the transfectedpopulation TE1 were identified by plasmid rescue from genomicDNA and sequencing with inverted primers within the constructregions. Nine clones specific for pEtHEA (Fig. 5A–C) and sevenclones for pEtADA (Fig. 5D–F) were identified using colony PCRand single digestion with EcoRV. No clone was obtained from thegenomic DNA of the parental strain BJ or from the control (undi-gested) genomic DNA of the TE1 population. Sequence analysisshowed that the constructs had integrated into the genome of E.tenella, with two integration sites for pEtHEA (one in a pseudomol-ecule of chromosome 1 and the other in contig 00026939) and oneintegration site for pEtADA (in conting 00010310) (Fig. 5G). Theintegration regions Et_contig 00026939 and Et_conting 00010310have not been annotated yet, so we cannot clarify which genesare present at these sites of plasmid integration. In addition, theplasmid rescue from genomic DNA of one of the cloned populations(TE1.2) and sequence analysis demonstrated that one integrationsite for pEtHEA (in the same pseudomolecule of chromosome 1)and one integration site for pEtADA (in conting 00010310) oc-curred in the genome of the TE1.2 population (data not shown).The single integration of pEtHEA is consistent with the findingusing Southern blotting analysis. The integrations at these sites
pulations. (A) PCR screening of transgenic parasites, TE1 and TE2, for the reportere vector pEtHEA as a template for PCR amplification. (B) TgDHFR-TS gene was only
ng. +, the plasmid pEtADA as a template for PCR amplification. (C) Southern blotting1.3) of E. tenella. Genomic DNA digested with EcoRV, which does not cleave in the
e result demonstrated that the introduced constructs were integrated into genomice, clones TE1.2 and TE1.3 had one copy. Linearised pEtHEA plasmid was used as a
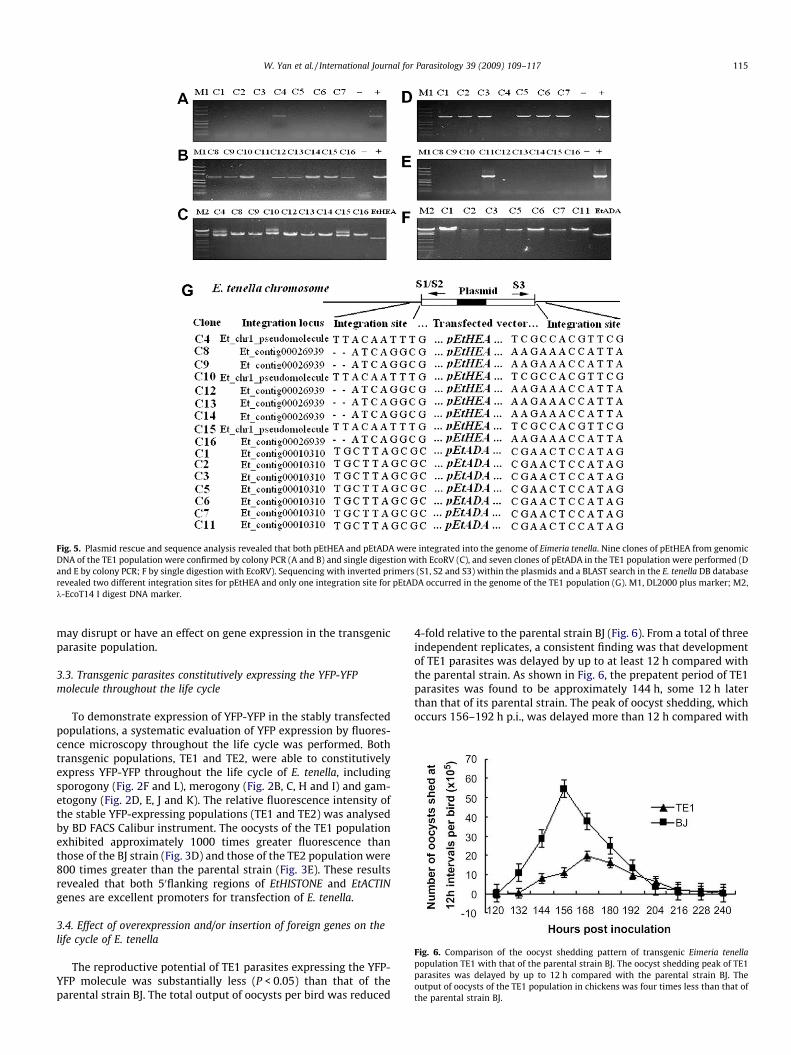
Fig. 6. Comparison of the oocyst shedding pattern of transgenic Eimeria tenellapopulation TE1 with that of the parental strain BJ. The oocyst shedding peak of TE1parasites was delayed by up to 12 h compared with the parental strain BJ. Theoutput of oocysts of the TE1 population in chickens was four times less than that ofthe parental strain BJ.
Fig. 5. Plasmid rescue and sequence analysis revealed that both pEtHEA and pEtADA were integrated into the genome of Eimeria tenella. Nine clones of pEtHEA from genomicDNA of the TE1 population were confirmed by colony PCR (A and B) and single digestion with EcoRV (C), and seven clones of pEtADA in the TE1 population were performed (Dand E by colony PCR; F by single digestion with EcoRV). Sequencing with inverted primers (S1, S2 and S3) within the plasmids and a BLAST search in the E. tenella DB databaserevealed two different integration sites for pEtHEA and only one integration site for pEtADA occurred in the genome of the TE1 population (G). M1, DL2000 plus marker; M2,k-EcoT14 I digest DNA marker.
W. Yan et al. / International Journal for Parasitology 39 (2009) 109–117 115
may disrupt or have an effect on gene expression in the transgenicparasite population.
3.3. Transgenic parasites constitutively expressing the YFP-YFPmolecule throughout the life cycle
To demonstrate expression of YFP-YFP in the stably transfectedpopulations, a systematic evaluation of YFP expression by fluores-cence microscopy throughout the life cycle was performed. Bothtransgenic populations, TE1 and TE2, were able to constitutivelyexpress YFP-YFP throughout the life cycle of E. tenella, includingsporogony (Fig. 2F and L), merogony (Fig. 2B, C, H and I) and gam-etogony (Fig. 2D, E, J and K). The relative fluorescence intensity ofthe stable YFP-expressing populations (TE1 and TE2) was analysedby BD FACS Calibur instrument. The oocysts of the TE1 populationexhibited approximately 1000 times greater fluorescence thanthose of the BJ strain (Fig. 3D) and those of the TE2 population were800 times greater than the parental strain (Fig. 3E). These resultsrevealed that both 50flanking regions of EtHISTONE and EtACTINgenes are excellent promoters for transfection of E. tenella.
3.4. Effect of overexpression and/or insertion of foreign genes on thelife cycle of E. tenella
The reproductive potential of TE1 parasites expressing the YFP-YFP molecule was substantially less (P < 0.05) than that of theparental strain BJ. The total output of oocysts per bird was reduced
4-fold relative to the parental strain BJ (Fig. 6). From a total of threeindependent replicates, a consistent finding was that developmentof TE1 parasites was delayed by up to at least 12 h compared withthe parental strain. As shown in Fig. 6, the prepatent period of TE1parasites was found to be approximately 144 h, some 12 h laterthan that of its parental strain. The peak of oocyst shedding, whichoccurs 156–192 h p.i., was delayed more than 12 h compared with
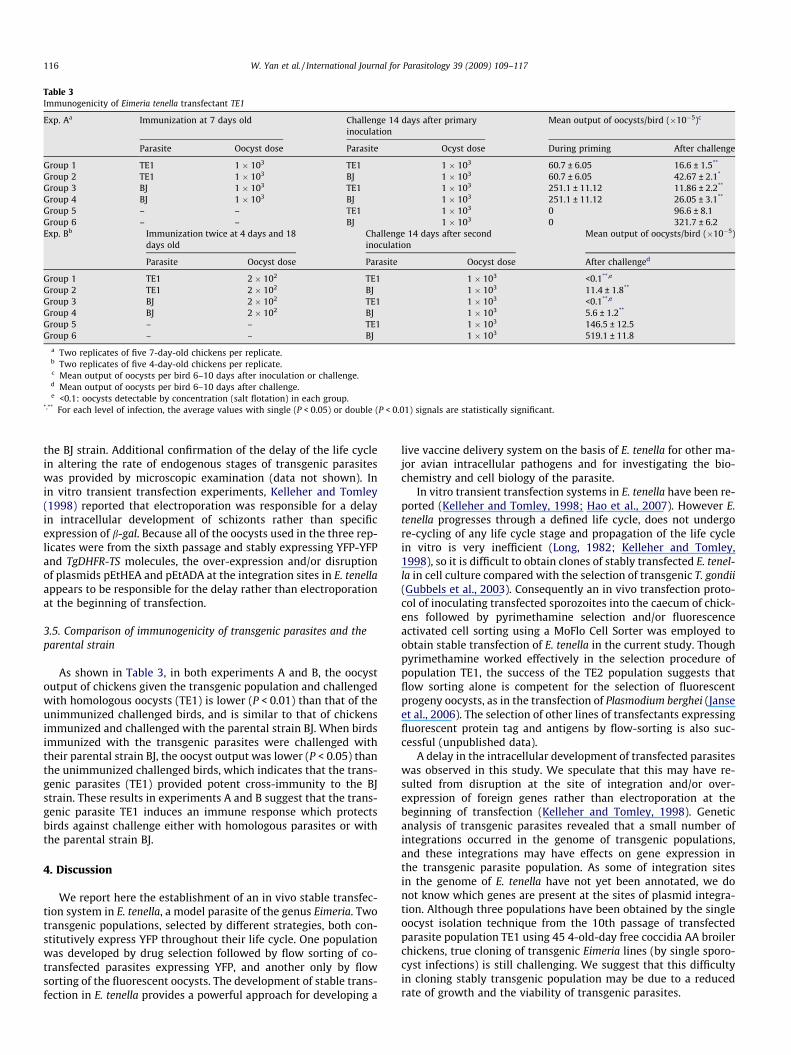
Table 3Immunogenicity of Eimeria tenella transfectant TE1
Exp. Aa Immunization at 7 days old Challenge 14 days after primaryinoculation
Mean output of oocysts/bird (�10�5)c
Parasite Oocyst dose Parasite Ocyst dose During priming After challenge
Group 1 TE1 1 � 103 TE1 1 � 103 60.7 ± 6.05 16.6 ± 1.5**
Group 2 TE1 1 � 103 BJ 1 � 103 60.7 ± 6.05 42.67 ± 2.1*
Group 3 BJ 1 � 103 TE1 1 � 103 251.1 ± 11.12 11.86 ± 2.2**
Group 4 BJ 1 � 103 BJ 1 � 103 251.1 ± 11.12 26.05 ± 3.1**
Group 5 – – TE1 1 � 103 0 96.6 ± 8.1Group 6 – – BJ 1 � 103 0 321.7 ± 6.2Exp. Bb Immunization twice at 4 days and 18
days oldChallenge 14 days after secondinoculation
Mean output of oocysts/bird (�10�5)
Parasite Oocyst dose Parasite Oocyst dose After challenged
Group 1 TE1 2 � 102 TE1 1 � 103 <0.1**,e
Group 2 TE1 2 � 102 BJ 1 � 103 11.4 ± 1.8**
Group 3 BJ 2 � 102 TE1 1 � 103 <0.1**,e
Group 4 BJ 2 � 102 BJ 1 � 103 5.6 ± 1.2**
Group 5 – – TE1 1 � 103 146.5 ± 12.5Group 6 – – BJ 1 � 103 519.1 ± 11.8
a Two replicates of five 7-day-old chickens per replicate.b Two replicates of five 4-day-old chickens per replicate.c Mean output of oocysts per bird 6–10 days after inoculation or challenge.d Mean output of oocysts per bird 6–10 days after challenge.e <0.1: oocysts detectable by concentration (salt flotation) in each group.
*,** For each level of infection, the average values with single (P < 0.05) or double (P < 0.01) signals are statistically significant.
116 W. Yan et al. / International Journal for Parasitology 39 (2009) 109–117
the BJ strain. Additional confirmation of the delay of the life cyclein altering the rate of endogenous stages of transgenic parasiteswas provided by microscopic examination (data not shown). Inin vitro transient transfection experiments, Kelleher and Tomley(1998) reported that electroporation was responsible for a delayin intracellular development of schizonts rather than specificexpression of b-gal. Because all of the oocysts used in the three rep-licates were from the sixth passage and stably expressing YFP-YFPand TgDHFR-TS molecules, the over-expression and/or disruptionof plasmids pEtHEA and pEtADA at the integration sites in E. tenellaappears to be responsible for the delay rather than electroporationat the beginning of transfection.
3.5. Comparison of immunogenicity of transgenic parasites and theparental strain
As shown in Table 3, in both experiments A and B, the oocystoutput of chickens given the transgenic population and challengedwith homologous oocysts (TE1) is lower (P < 0.01) than that of theunimmunized challenged birds, and is similar to that of chickensimmunized and challenged with the parental strain BJ. When birdsimmunized with the transgenic parasites were challenged withtheir parental strain BJ, the oocyst output was lower (P < 0.05) thanthe unimmunized challenged birds, which indicates that the trans-genic parasites (TE1) provided potent cross-immunity to the BJstrain. These results in experiments A and B suggest that the trans-genic parasite TE1 induces an immune response which protectsbirds against challenge either with homologous parasites or withthe parental strain BJ.
4. Discussion
We report here the establishment of an in vivo stable transfec-tion system in E. tenella, a model parasite of the genus Eimeria. Twotransgenic populations, selected by different strategies, both con-stitutively express YFP throughout their life cycle. One populationwas developed by drug selection followed by flow sorting of co-transfected parasites expressing YFP, and another only by flowsorting of the fluorescent oocysts. The development of stable trans-fection in E. tenella provides a powerful approach for developing a
live vaccine delivery system on the basis of E. tenella for other ma-jor avian intracellular pathogens and for investigating the bio-chemistry and cell biology of the parasite.
In vitro transient transfection systems in E. tenella have been re-ported (Kelleher and Tomley, 1998; Hao et al., 2007). However E.tenella progresses through a defined life cycle, does not undergore-cycling of any life cycle stage and propagation of the life cyclein vitro is very inefficient (Long, 1982; Kelleher and Tomley,1998), so it is difficult to obtain clones of stably transfected E. tenel-la in cell culture compared with the selection of transgenic T. gondii(Gubbels et al., 2003). Consequently an in vivo transfection proto-col of inoculating transfected sporozoites into the caecum of chick-ens followed by pyrimethamine selection and/or fluorescenceactivated cell sorting using a MoFlo Cell Sorter was employed toobtain stable transfection of E. tenella in the current study. Thoughpyrimethamine worked effectively in the selection procedure ofpopulation TE1, the success of the TE2 population suggests thatflow sorting alone is competent for the selection of fluorescentprogeny oocysts, as in the transfection of Plasmodium berghei (Janseet al., 2006). The selection of other lines of transfectants expressingfluorescent protein tag and antigens by flow-sorting is also suc-cessful (unpublished data).
A delay in the intracellular development of transfected parasiteswas observed in this study. We speculate that this may have re-sulted from disruption at the site of integration and/or over-expression of foreign genes rather than electroporation at thebeginning of transfection (Kelleher and Tomley, 1998). Geneticanalysis of transgenic parasites revealed that a small number ofintegrations occurred in the genome of transgenic populations,and these integrations may have effects on gene expression inthe transgenic parasite population. As some of integration sitesin the genome of E. tenella have not yet been annotated, we donot know which genes are present at the sites of plasmid integra-tion. Although three populations have been obtained by the singleoocyst isolation technique from the 10th passage of transfectedparasite population TE1 using 45 4-old-day free coccidia AA broilerchickens, true cloning of transgenic Eimeria lines (by single sporo-cyst infections) is still challenging. We suggest that this difficultyin cloning stably transgenic population may be due to a reducedrate of growth and the viability of transgenic parasites.

W. Yan et al. / International Journal for Parasitology 39 (2009) 109–117 117
Compared with its parental strain, the transgenic populationTE1 reduced fecundity but maintained immunogenicity, and thesetraits were similar to that of the precocious strain of E. tenella (Jef-fers, 1975; McDonald and Ballingall, 1983; McDonald et al., 1986).These data strongly indicate that a transgenic parasite expressingprotective antigens could be a promising eukaryotic vaccine vector.This presumption using eukaryotic protozoa as vaccine vehicle hasbeen tested in T. gondii. Recombinant T. gondii expressing the Plas-modium yoelii circumsporozoite protein provided highly effectivepriming for CD8+ T cell-dependent protective immunity againstmalaria (Charest et al., 2000). Based on this intriguing report, weexpect that transgenic E. tenella, a close relative of T. gondii, couldalso elicit potent cellular immune response of the host, by express-ing antigenic proteins of other intracellular pathogens in chickens.One study, to generate transgenic strains of E. tenella expressingthe NP or HA gene originating from Highly Pathogenic Avian Influ-enza virus (H5N1) together with YFP is under way.
In conclusion, we have developed two transgenic populations ofE. tenella through an in vivo transfection protocol based on drugselection and/or flow sorting, and investigated the biological char-acteristics of one population, especially focusing on its reproduc-tivity and immunogenicity. Establishment of a stable transfectionsystem in the genus Eimeria is also of considerable significanceas the availability of such a tool will greatly facilitate the investiga-tion of the biology of Eimeria.
Acknowledgements
We thank Professor D.S. Roos (University of Pennsylvania, USA)for kindly providing plasmid pDHFR-TSc3. We are grateful to Prof.Qijun Chen (Swedish Institute for Infectious Disease Control, Swe-den), and Dr. Jin Zhu (the Therapeutic Goods Administration, Aus-tralia) for helpful advice and review of the paper. We alsoacknowledge our colleagues Yonggen Jia and Guangwen Yin fortheir help during experiments and Julie Clark for technical advice.This study is supported by the National Natural Science Foundationof China (Project Nos. 30540003 and 30471298), the National HighTechnology Research and Development Program of China (ProjectNo. 2006AA02Z458), and the Beijing Natural Science Foundation(No. 6073027). Interactions between the authors’ laboratories arefacilitated by a BBSRC China Partnering Award.
References
Black, M., Seeber, F., Soldati, D., Kim, K., Boothroyd, J.C., 1995. Restriction enzyme-mediated integration elevates transformation frequency and enables co-transfection of Toxoplasma gondii. Mol. Biochem. Parasitol. 74, 55–63.
Bonilla, J.A., Moura, P.A., Bonilla, T.D., Yowell, C.A., Fidock, D.A., Dame, J.B., 2007.Effects on growth, hemoglobin metabolism and paralogous gene expressionresulting from disruption of genes encoding the digestive vacuole plasmepsinsof Plasmodium falciparum. Int. J. Parasitol. 37, 317–327.
Charest, H., Sedegah, M., Yap, G.S., Gazzinelli, R.T., Caspar, P., Hoffman, S.L., Sher, A.,2000. Recombinant attenuated Toxoplasma gondii expressing the Plasmodiumyoelii circumsporozoite protein provides highly effective priming for CD8+ Tcell-dependent protective immunity against malaria. J. Immunol. 165, 2084–2092.
Danforth, H.D., 1998. Use of live oocyst vaccines in the control of avian coccidiosis:experimental studies and field trials. Int. J. Parasitol. 28, 1099–1109.
Digby, M.R., Lowenthal, J.W., 1995. Cloning and expression of the chickeninterferon-gamma gene. J. Interferon Cytokine Res. 15, 939–945.
Doran, D.J., 1970. Effect of age and freezing on development of Eimeria adenoeidesand E. tenella sporozoites in cell culture. J. Parasitol. 56, 27–29.
Goonewardene, R., Daily, J., Kaslow, D., Sullivan, T.J., Duffy, P., Carter, R., Mendis, K.,Wirth, D., 1993. Transfection of the malaria parasite and expression of fireflyluciferase. Proc. Natl. Acad. Sci. USA 90, 5234–5236.
Gubbels, M.J., Li, C., Striepen, B., 2003. High-throughput growth assay forToxoplasma gondii using yellow fluorescent protein. Antimicrob. AgentsChemother. 47, 309–316.
Hao, L., Liu, X., Zhou, X., Li, J., Suo, X., 2007. Transient transfection of Eimeria tenellausing yellow or red fluorescent protein as a marker. Mol. Biochem. Parasitol.153, 213–215.
Haug, A., Williams, R.B., Larsen, S., 2006. Counting coccidial oocysts in chickenfaeces: a comparative study of a standard McMaster technique and a new rapidmethod. Vet. Parasitol. 136, 233–242.
Hofmann, J., Raether, W., 1990. Improved techniques for the in vitro cultivation ofEimeria tenella in primary chick kidney cells. Parasitol. Res. 76, 479–486.
Hong, Y.H., Lillehoj, H.S., Lillehoj, E.P., Lee, S.H., 2006. Changes in immune-relatedgene expression and intestinal lymphocyte subpopulations following Eimeriamaxima infection of chickens. Vet. Immunol. Immunopathol. 114, 259–272.
Janse, C.J., Franke-Fayard, B., Waters, A.P., 2006. Selection by flow-sorting ofgenetically transformed, GFP-expressing blood stages of the rodent malariaparasite, Plasmodium berghei. Nat. Protoc. 1, 614–623.
Jeffers, T.K., 1975. Attenuation of Eimeria tenella through selection forprecociousness. J. Parasitol. 61, 1083–1090.
Kelleher, M., Tomley, F.M., 1998. Transient expression of beta-galactosidase indifferentiating sporozoites of Eimeria tenella. Mol. Biochem. Parasitol. 97, 21–31.
Kim, K., Weiss, L.M., 2004. Toxoplasma gondii: the model apicomplexan. Int. J.Parasitol. 34, 423–432.
Lee, E.H., 1979. Single and low-level oocyst infections of drug-resistant field strainsof Eimeria tenella in medicated birds. Can. Vet. J. 20, 102–104.
Lillehoj, H.S., Choi, K.D., 1998. Recombinant chicken interferon-gamma-mediatedinhibition of Eimeria tenella development in vitro and reduction of oocystproduction and body weight loss following Eimeria acervulina challengeinfection. Avian Dis. 42, 307–314.
Long, P.L., Millard, B.J., Joyner, L.P., Norton, C.C., 1976. A guide to laboratorytechniques used in the study and diagnosis of avian coccidiosis. Folia. Vet. Lat. 6,201–217.
Long, P.L., 1982. The Biology of the Coccidia. Edward Arnold Press, London.Long, P.L., Rowell, J.G., 1958. Counting oocysts of chicken coccidia. Lab. Pract. 7,
515–519.McDonald, V., Ballingall, S., 1983. Further investigation of the pathogenicity,
immunogenicity and stability of precocious Eimeria acervulina. Parasitology 86(Pt. 3), 361–369.
McDonald, V., Shirley, M.W., Bellatti, M.A., 1986. Eimeria maxima: characteristics ofattenuated lines obtained by selection for precocious development in thechicken. Exp. Parasitol. 61, 192–200.
Norton, C.C., Marshall, S.H.S.N., Joyner, L.P., Parsons, G., 1983. Freedom of coccidialoocysts from Newcastle disease virus. Res. Vet. Sci. 35, 367–369.
Sambrook, J., Russell, D., 2001. Molecular Cloning: A Laboratory Manual, thirdedition. Cold Spring Harbor Labor Press, New York.
Schmatz, D.M., Crane, M.S., Murray, P.K., 1984. Purification of Eimeria sporozoites byDE-52 anion exchange chromatography. J. Protozool. 31, 181–183.
Sharma, J.M., 1991. Avian Cellular Immunology. CRC Press, Boca Raton.Shedlock, D.J., Shen, H., 2003. Requirement for CD4 T cell help in generating
functional CD8 T cell memory. Science 300, 337–339.Shirley, M.W., 2000. The genome of Eimeria spp., with special reference to Eimeria
tenella-a coccidium from the chicken. Int. J. Parasitol. 30, 485–493.Sibley, L.D., Messina, M., Niesman, I.R., 1994. Stable DNA transformation in the
obligate intracellular parasite Toxoplasma gondii by complementation oftryptophan auxotrophy. Proc. Natl. Acad. Sci. USA 91, 5508–5512.
Soldati, D., Boothroyd, J.C., 1993. Transient transfection and expression in theobligate intracellular parasite Toxoplasma gondii. Science 260, 349–352.
Suo, X., Zhang, J.X., Li, Z.G., Yang, C.T., Min, Q.R., Xu, L.T., Liu, Q., Zhu, X.Q., 2006. Theefficacy and economic benefits of Supercox, a live anticoccidial vaccine in acommercial trial in broiler chickens in China. Vet. Parasitol. 142, 63–70.
Tierney, J., Mulcahy, G., 2003. Comparative development of Eimeria tenella(Apicomplexa) in host cells in vitro. Parasitol. Res. 90, 301–304.
Trout, J.M., Lillehoj, H.S., 1996. T lymphocyte roles during Eimeria acervulina andEimeria tenella infections. Vet. Immunol. Immunopathol. 53, 163–172.
van den Hoff, J.B.M.F.M., Lamers, W.H., 1992. Electroporation in intracellular buffersincreases cell survival. Nucleic Acid Res. 20, 2902.
van Dijk, M.R., Waters, A.P., Janse, C.J., 1995. Stable transfection of malaria parasiteblood stages. Science 268, 1358–1362.
Williams, R.B., 1998. Epidemiological aspects of the use of live anticoccidialvaccines for chickens. Int. J. Parasitol. 28, 1089–1098.
Yun, C.H., Lillehoj, H.S., Choi, K.D., 2000. Eimeria tenella infection induces localgamma interferon production and intestinal lymphocyte subpopulationchanges. Infect. Immun. 68, 1282–1288.
Related Documents






![Edinburgh Research Explorer - University of Edinburgh...Plasmodium chabaudi [19], Toxoplasma gondii [20], and Eimeria tenella [21], opening up the possibility of using classical genetic](https://static.cupdf.com/doc/110x72/61176d6fb41b2444541111f0/edinburgh-research-explorer-university-of-edinburgh-plasmodium-chabaudi-19.jpg)




