Hydrobiologia 440: 317–329, 2000. M.B. Jones, J.M.N. Azevedo, A.I. Neto, A.C. Costa & A.M. Frias Martins (eds), Island, Ocean and Deep-Sea Biology. © 2000 Kluwer Academic Publishers. Printed in the Netherlands. 317 Stable isotope evidence of benthic microalgae-based growth and secondary production in the suspension feeder Cerastoderma edule (Mollusca, Bivalvia) in the Marennes-Ol´ eron Bay Pierre-Guy Sauriau 1 & Chang-Keun Kang 2 1 Centre de Recherche en Ecologie Marine et Aquaculture de L’Houmeau (CNRS-IFREMER, UMR-10), BP 5, F-17137 L’Houmeau, France E-mail: [email protected] 2 National Fisheries Research & Development Institute (Pohang Regional laboratory), 616 Duhodong, Buk-gu, Pohang 791-110, South Korea Key words: growth, secondary production, Cerastoderma edule, stable isotope ratios, microphytobenthos, intertidal, Marennes-Ol´ eron Bay Abstract The contribution of natural food sources to the growth and secondary production of the suspension feeding bivalve Cerastoderma edule (L.) was estimated under field conditions in the Marennes-Oléron Bay (Atlantic coast, France). Monthly estimates of abundance, biomass and cockle growth were combined with seasonal analyses of δ 13 C and δ 15 N ratios of juvenile and adult cockles, together with their potential food sources [i.e. suspended particulate or- ganic matter (POM), microphytobenthos, macroalgae and seagrass] sampled at mid-tide level in a muddy sandflat. Adult cockles grew mainly in spring, whereas juveniles grew in summer and autumn, following spat recruitment in early summer. Total annual production and elimination of cockles were estimated to be 32.5 and 34.7 g AFDW m -2 yr -1 . Relative contributions of each year class to production were ca 40, 41, 11 and 6% for 0-group, 1-, 3- and 4-yr- old cockles in 1995, respectively. Quantitative assessment of proportions of food sources to the annual secondary production of cockles was obtained by using a simple carbon isotope-mixing model with microphytobenthos (δ 13 C = -16.0±0.6 ‰) and POM (δ 13 C= -22.2±1.1 ‰) as end-members. On average, more than 70% of the total annual cockle production originated from microphytobenthos, with a much higher contribution for the 0-group (88%) than for adult cockles (60%). The between-age difference was induced mainly by changes in the availability of food resources (benthic versus planktonic) during the non-synchronous growing seasons of juvenile and adult cockles. Introduction The edible cockle Cerastoderma edule (L.) is one of the most common benthic suspension feeders on tidal flats along the European Atlantic coast (Tebble, 1966). This species exhibits large year-to-year and between-site fluctuations in larval recruitment success, and of subsequent survival and growth of juveniles (Kristensen, 1957; Dörjes et al., 1986; Ducrotoy et al., 1991). Consequently, relative contributions of each year class to the total annual secondary production of cockle populations are highly variable, as repor- ted previously (Evans, 1977; Möller & Rosenberg, 1983). Amongst the main factors that may induce these variations are abnormal thermal conditions (Or- ton, 1933; Beukema, 1982; Guillou et al., 1992), predation by shore crabs, shrimps and fish (Reise, 1985; Sanchez-Salazar et al., 1987), intraspecific com- petition (André & Rosenberg, 1991; Bachelet et al., 1992; Jensen, 1993), and spatfall transport by near- bottom flows (Baggerman, 1953; de Montaudouin & Bachelet, 1996). However, little attention has been paid to the influence of changes in the diet available for suspension feeding cockles during critical periods of their life cycle. Jensen (1992) hypothesised that mis- match in growing seasons of juvenile and adult cockles may induce a differential use of food resources; this may minimise intraspecific competition within the

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hydrobiologia 440: 317–329, 2000.M.B. Jones, J.M.N. Azevedo, A.I. Neto, A.C. Costa & A.M. Frias Martins (eds), Island, Ocean and Deep-Sea Biology.© 2000Kluwer Academic Publishers. Printed in the Netherlands.
317
Stable isotope evidence of benthic microalgae-based growth andsecondary production in the suspension feederCerastodermaedule(Mollusca, Bivalvia) in the Marennes-Oleron Bay
Pierre-Guy Sauriau1 & Chang-Keun Kang21Centre de Recherche en Ecologie Marine et Aquaculture de L’Houmeau (CNRS-IFREMER, UMR-10),BP 5, F-17137 L’Houmeau, FranceE-mail: [email protected] Fisheries Research & Development Institute (Pohang Regional laboratory), 616 Duhodong,Buk-gu, Pohang 791-110, South Korea
Key words: growth, secondary production,Cerastoderma edule, stable isotope ratios, microphytobenthos,intertidal, Marennes-Oleron Bay
Abstract
The contribution of natural food sources to the growth and secondary production of the suspension feeding bivalveCerastoderma edule(L.) was estimated under field conditions in the Marennes-Oléron Bay (Atlantic coast, France).Monthly estimates of abundance, biomass and cockle growth were combined with seasonal analyses ofδ13C andδ15N ratios of juvenile and adult cockles, together with their potential food sources [i.e. suspended particulate or-ganic matter (POM), microphytobenthos, macroalgae and seagrass] sampled at mid-tide level in a muddy sandflat.Adult cockles grew mainly in spring, whereas juveniles grew in summer and autumn, following spat recruitment inearly summer. Total annual production and elimination of cockles were estimated to be 32.5 and 34.7 g AFDW m−2
yr−1. Relative contributions of each year class to production were ca 40, 41, 11 and 6% for 0-group, 1-, 3- and 4-yr-old cockles in 1995, respectively. Quantitative assessment of proportions of food sources to the annual secondaryproduction of cockles was obtained by using a simple carbon isotope-mixing model with microphytobenthos (δ13C= −16.0±0.6 ‰) and POM (δ13C = −22.2±1.1 ‰) as end-members. On average, more than 70% of the totalannual cockle production originated from microphytobenthos, with a much higher contribution for the 0-group(88%) than for adult cockles (60%). The between-age difference was induced mainly by changes in the availabilityof food resources (benthicversusplanktonic) during the non-synchronous growing seasons of juvenile and adultcockles.
Introduction
The edible cockleCerastoderma edule(L.) is oneof the most common benthic suspension feeders ontidal flats along the European Atlantic coast (Tebble,1966). This species exhibits large year-to-year andbetween-site fluctuations in larval recruitment success,and of subsequent survival and growth of juveniles(Kristensen, 1957; Dörjes et al., 1986; Ducrotoy etal., 1991). Consequently, relative contributions of eachyear class to the total annual secondary productionof cockle populations are highly variable, as repor-ted previously (Evans, 1977; Möller & Rosenberg,1983). Amongst the main factors that may induce
these variations are abnormal thermal conditions (Or-ton, 1933; Beukema, 1982; Guillou et al., 1992),predation by shore crabs, shrimps and fish (Reise,1985; Sanchez-Salazar et al., 1987), intraspecific com-petition (André & Rosenberg, 1991; Bachelet et al.,1992; Jensen, 1993), and spatfall transport by near-bottom flows (Baggerman, 1953; de Montaudouin &Bachelet, 1996). However, little attention has beenpaid to the influence of changes in the diet available forsuspension feeding cockles during critical periods oftheir life cycle. Jensen (1992) hypothesised that mis-match in growing seasons of juvenile and adult cocklesmay induce a differential use of food resources; thismay minimise intraspecific competition within the

318
cockle population (Kang et al., 1999). Nevertheless,the hypothesis was not supported by the literature des-pite several studies on the feeding physiology ofC.eduledescribing size-dependent variations (Hawkinset al., 1990; Iglesias & Navarro, 1991; Small et al.,1997) and selective ingestion of organic matter (Prinset al., 1991; Urrutia et al., 1996; Navarro & Widdows,1997).
Various potential food sources are available tobenthic suspension feeders in the seston of coastal andestuarine environments (Newell, 1979; Dame, 1996).It is, however, difficult to assess the relative contribu-tion of each food source to their growth and secondaryproduction because direct observation of feeding be-haviour is difficult over long time periods in the field(Bayne et al., 1977; Dame, 1996). Indirect methods,such as gut content analyses (e.g. Verwey, 1952; Iv-ell, 1981; Kamermans, 1994), are also not satisfactorysince they give snapshot information on food itemsthat have been ingested and not what is assimilatedover a long period of time. Stable isotope analysescan provide answers to this question (Fry & Sherr,1984), as the13C/12C and15N/14N ratios of an an-imal directly reflect the composition of food sourcesassimilated and incorporated over time (DeNiro &Epstein, 1978, 1981). Stable isotope analyses havebeen used successfully in many studies of spatial andtemporal variations of isotopic composition in inver-tebrates and their potential diets in various estuarineand saltmarsh food webs (see reviews by Fry & Sherr,1984; Michener & Schell, 1994). Only Junger &Planas (1994), however, have combined stable carbonisotope mixing models with quantitative inventories ofmacro-invertebrate biomasses to assess quantitativelythe trophic base of lotic ecosystems. Consequently,the contribution of various food sources to the annualgrowth and secondary production of intertidal marinespecies has never been estimatedin situ.
The purpose of the present study was to quantifythe relative contribution of natural food sourcesto the growth and secondary production of thecockleCerastoderma edulein muddy sandflats of theMarennes-Oléron Bay. Our latest results (Kang etal., 1999) indicated that juvenile and adult cockleswere able to preferentially ingest and/or assimilate mi-crophytobenthos and that there were only two majorsources of organic matter assimilated by cockles i.e.particulate organic matter (POM) advected on thesetidal flats through water channels and locally resus-pended microphytobenthos (MPB). Therefore, in thisstudy, we used a simple two-source mixing model of
stable carbon isotopes (Fry & Sherr, 1984) combinedwith growth and secondary production estimates toquantify the relative contribution of each food sourceto the total annual production of the mid-tide levelcockle population.
Materials and methods
Study site
The study took place on the muddy intertidal sand-flat of Ronce-les-Bains, at the southern end of theMarennes-Oléron Bay, halfway along the French At-lantic coast (Fig. 1a). The bay is characterised bysemidiurnal tides with a maximal range of 6.5 m. Mar-ine waters enter mostly via. the north entrance of thebay (Pertuis d’Antioche). They are also introducedthrough the south-west entrance (Pertuis de Maumus-son) but their influence on the southern part of thebay is restricted to flood tides. Interaction between thetidal regime and local bathymetric features induces asouthward residual transport of marine water masses,which are drained out of the bay through the Pertuis deMaumusson. The water residence time in the wholebay varies between 5 and 10 d depending on tides(Raillard & Ménesguen, 1994). The River Charente,on the east, is the major source of freshwater inputinto the bay (Ravail et al., 1988), whereas freshwaterinput from the River Seudre is poor (Soletchnick et al.,1998). Average salinity within the bay ranges from 28to 33 (Soletchnick et al., 1998). Seston dynamics areaffected strongly by advection of marine water masses,and by wind- and tide-induced resuspension of bothmuddy sediments (Raillard et al., 1994; Raillard &Ménesguen, 1994) and benthic microalgae (Razet etal., 1990; Zurburg et al., 1994). Additional informa-tion on seasonal hydrobiology, sedimentary conditionsand shellfish rearing activities in the bay is given byHéral et al. (1983), Sauriau et al. (1989) and Héral etal. (1990), respectively.
At Ronce-les-Bains, cockle beds extend over a 50ha surface area (Fig. 1b) and the cockle population atmid-tide level appears to be stable over time (Sauriau,1992). All lower parts of the shore, between MLWNand ELWS, are devoted to farming oysters and cocklesare scarce within the oyster parks (Fig. 1b). At mid-tide level, sediments are dominated by fine muddysands (Guillou et al., 1990) with an organic mattercontent of ca 1% (loss of weight on ignition at 450◦C).Dominant macrofauna areHydrobia ulvae(Pennant),

319
Figure 1a. Map of Marennes-Oleron Bay showing location of the three hydrological sampling stations in channels (�) and the sampling site(•) at Ronce-les-Bains.
Figure 1b. Aerial photograph of Ronce-les-Bains muddy sandflats in August 1993 showing oyster parks on lower parts of the shore and locationof the sampling site (•) at mid-tide level. Channels and seashore line are located on the top left-hand corner and the bottom right-hand corner,respectively.

320
Cerastoderma edule, Notomastus latericeus(M. Sars),Tharyx marioni(de Saint-Joseph),Scoloplos armiger(O.F. Müller) andArenicola marina(L.). In the past,the mid-tide level area was covered by a dense bed ofZostera noltii(Hornem), but it vanished in 1985–1986(Sauriau, 1992), possibly due to local changes in ba-thymetry (Soletchnick et al., 1998). The seagrass bedhas not recovered except for a fewZosteraspots (<0.1m2). Bottom roughness is characterised by wave andcurrent ripple marks of a few centimetres in height.Although several macroalgae species occur in Ronce-les-Bains, they are restricted mainly to the oysterparks. Greening of intertidal sandflats [i.e. summerdevelopment ofUlvae andEnteromorphaalgal matsas reported by Thiel & Watling (1998)], is never ob-served at Ronce-les-Bains. However, some strandsof Chondria dasyphylla(Woodward),EnteromorphaandUlva species have been observed attached to deadcockle shells, mainly on the lower muddy-sand area,throughout the year.
Sample of cockles
Cockles were collected monthly from February 1995to February 1996. Three replicate plots were sampledrandomly using a 0.25 m2 aluminium corer to de-termine mean monthly abundance and biomasses atmid-tide level. Sediment was sieved through a 1 mmaperture mesh, and all recovered animals were storedin 4% neutralised formalin and sorted in the laborat-ory. To determine monthly length weight relationship,cockles (10 for each year class, total>30 individu-als) were collected by hand at each sampling date andkept alive overnight in filtered seawater to evacuate gutcontents. Shell lengths were measured with Verniercallipers to the nearest 0.05 mm, prior to dissectionof individuals. Flesh was separated from shells, storedat−18◦C for at least 48 h and freeze-dried to determ-ine flesh dry weight (DW). Subsamples of dry fleshwere combusted in a muffle furnace at 450◦C for6 h to estimate ash contents and ash-free dry weight(AFDW). Flesh organic carbon content was determ-ined using a CHN elemental analyser (Perkin Elmer2400) after acidification (10% HCl) to remove car-bonates. The allometric relationships at each samplingdate were expressed as logW = log a + b log L (W= flesh weight andL = shell length). Length data ofthe samples collected to estimate production were con-verted into flesh weight and ash-free dry weight usingthese allometric equations at each sampling date. Theage of each cockle was determined by counting the
clearly distinguishable annual winter rings, as reportedby Sauriau (1992) for this cockle population.
Production and elimination
Production (P ) and elimination (E) terms of each yearclass were calculated according to incremental and re-moval summation methods reviewed by Crisp (1984)for populations with non continuous recruitment anddistinguishable year classes:
P = t=n6t=0[(Nt + Nt+1)/2 · (Wt+1−Wt )] and
E = t=n6t=0[(Wt +Wt+1)/2 · (Nt − Nt+1)],
whereNt = abundance of a cohort at timet ,Wt= meanindividual flesh weight in a cohort at timet andn = thenumber of sampling occasions at different samplingtimest .
The mean biomass was calculated by:
B= [t=n6t=1(Nt ·Wt )]/n,
wheren = number of sampling dates in a year.Previous studies have expressed production estim-
ates ofCerastoderma edulein ash-free dry weightunits (Warwick & Price, 1975; Hibbert, 1976; Evans,1977; Jensen, 1993) or in organic carbon content(Warwick et al., 1979). To convert flesh dry weight(DW) into ash-free dry weight (AFDW) and organiccarbon content (C), we obtained the following tworegression equations:
AFDW = 0.915 DW – 0.003 (R2 = 0.999,n = 729)
C = 0.321 DW + 0.001 (R2 = 0.987,n = 578)
Sampling and analyses of stable isotopes in cocklesand food sources
Cockles for stable isotope analyses were collectedseasonally (March, June, September and December)at Ronce-les-Bains. To determine the isotopic valuefor POM introduced on tidal flats through centralchannels of the bay, spring and neap tidal samplingswere conducted seasonally at three stations: Pertuis deMaumusson, the mouth of the River Seudre and LeChapus (Fig. 1a), and prepared for analysis as indic-ated previously by Kang et al. (1999). Macroalgae andseagrass were collected seasonally at Ronce-les-Bains.They were cleaned of epibionts, washed with 10%HCL to remove carbonates, rinsed with Milli-Q water,

321
freeze-dried, ground to powder and kept frozen (−80◦C) until analysis. Microphytobenthos isotope valueswere given by Riera et al. (1996) and Riera & Richard(1996, 1997) from microphytobenthos samples collec-ted on various occasions on mudflats in the Bay ofMarennes-Oléron.
Stable isotope analyses followed the standard pro-tocols used by Riera & Richard (1996, 1997). Sampleswere combusted at 900◦C using CuO as an oxidant inevacuated quartz tubes. The resulting CO2 was pur-ified using a cryogenic distillation method similar tothat described by Boutton (1991). Before the purifica-tion of CO2, N2 was trapped on silica gel granules in astop-cock sample ampoule and analysed immediatelyafter CO2 collection. The carbon and nitrogen isotoperatios were measured using a Sigma 200 (CJS Sci-ences) double inlet, triple collector isotope ratio massspectrometer. Stable isotope ratios are reported in thestandardδ unit notation as follows:
δX = [(Rsample/Rreference)− 1] × 1000,
whereX is 13C or 15N, R is 13C/12C or 15N/14Nfor carbon and nitrogen, respectively. Values are ex-pressed as units of permil ( ‰) relative to the Pee DeeBelemnite standard (PDB) for carbon and to atmo-spheric N2 for nitrogen. The typical precision of thecomplete analysis (preparation, combustion and massspectrometric analysis) was± 0.1 ‰ for carbon and± 0.2 ‰ for nitrogen.
Statistical analyses of isotope data were performedusing 2-way Analysis of Variance and/or non para-metric Kruskal-Wallis test in the case of compar-ison of samples with a low number of observations(Sokal & Rohlf, 1981). The exact distribution table ofthe Kruskal–Wallis statistic was used when necessary(Siegel, 1956).
Results
Recruitment, abundance and biomass
At mid-tide level, the mean annual density ofCerasto-derma edulewas 259±277 individuals m−2 (Fig. 2a).Recruitment started in May/June (Figs 2 and 3), butthe main spat fall occurred in July–August with a max-imum spat density over 1000 individuals m−2 (Fig.2a). A secondary recruitment occurred during lateautumn (November/December), but it was 10 timessmaller than during the summer. Densities of cocklesdecreased quickly from August onwards due to high
spat mortality, which appeared to be higher in autumnthan in early winter (Fig. 2a). Two cohorts (1994 and1995 year classes) dominated the abundance duringthe study period (Fig. 2a). The 1994 year class (1-yr-old cockles) was the more numerous cohort from thebeginning of the study (February 1995) to early sum-mer, but was then dominated by the 1995 recruitmentfrom summer to early winter (Fig. 2a).
The 1994 year class attained the highest propor-tion (over 50%) of total annual biomass during thestudy period (Fig. 2b). Maximum biomass of the 1994year class occurred in August (30 g AFDW m−2),but 6 months later, biomass of this year class wasless than 2 g AFDW m−2. The contribution of the1995 recruitment to the total annual biomass peakedin September/October (6 g AFDW m−2), whereas thecontribution of the 1992 year class (3-yr-old cockles)was more stable from May to September (Fig. 2b).Age structure of the cockle population in 1995 wascharacterised by the relative absence of the 1993 yearclass (2-yr-old cockles), whereas, the 1992 year class(3-yr-old cockles) was well established. Some cocklesolder than 4 years (1991 year class) were also re-corded during this study, but their densities remainedlower than 5 – 10 individuals m−2, except for Augustand September with biomasses ranging from 4 to 5g AFDW m−2. No cockles older than 5 years weresampled at mid-tide level in 1995 (Fig. 2).
Shell growth
Shell growth of year classes varied with seasons (Fig.3). The 0-group (1995 year class) grew from May/June(settlement) to November. At the beginning of theirfirst winter, shell length of 0-group cockles averaged16.2± 1.3 mm. Growth rate of the secondary recruit-ment, that occurred in autumn and early winter, waslow and most of these cockles did not survive duringtheir first winter.
The main growing season of 1-yr-old cockles(1994 year class) was from spring to early summer(April until July) and the 1-yr-old cockle growth ratewas much lower from July to October (Fig. 3). At theend of their second growing season, shell length of the1994 year class reached 26.6± 1.8 mm. Similar trendswere shown by both 1993 year class and cockles olderthan 3 years. Shell growth of year classes appearedto diminish with increasing age, with an obvious de-crease of growth rate in cockles over 2 years (Fig. 3).No cockles more than 34 mm shell length were foundduring this study.
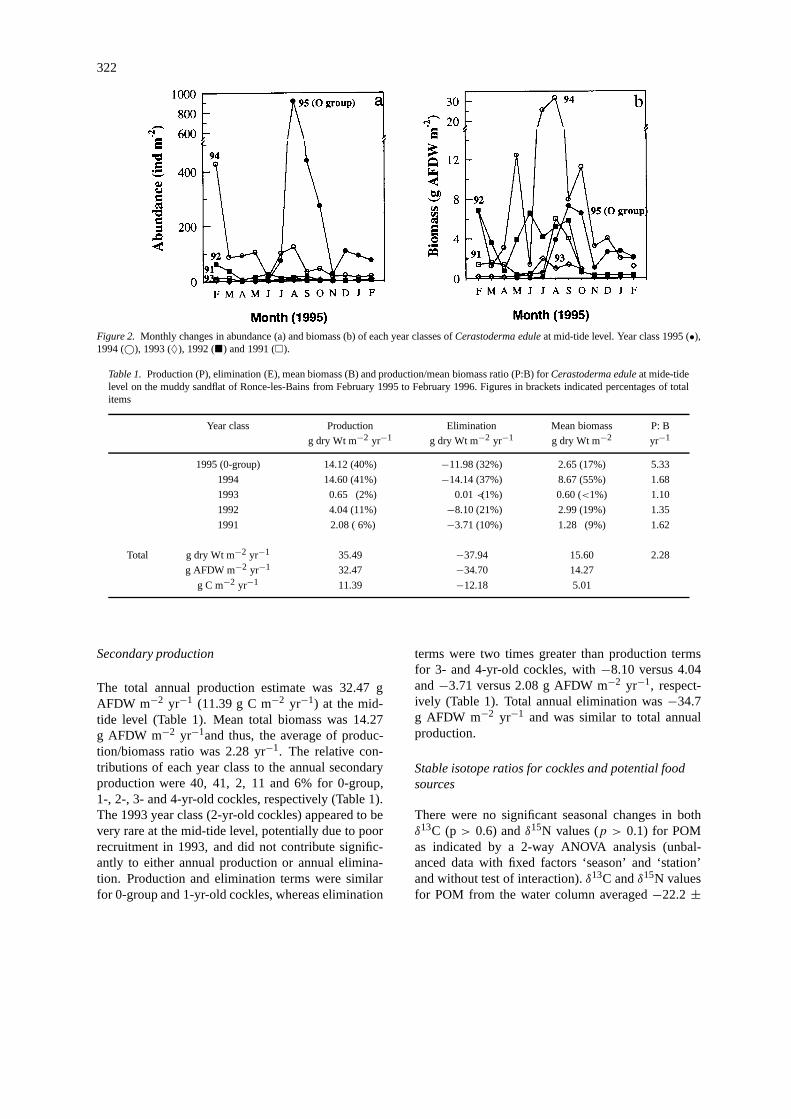
322
Figure 2. Monthly changes in abundance (a) and biomass (b) of each year classes ofCerastoderma eduleat mid-tide level. Year class 1995 (•),1994 (©), 1993 (♦), 1992 (�) and 1991 (�).
Table 1. Production (P), elimination (E), mean biomass (B) and production/mean biomass ratio (P:B) forCerastoderma eduleat mide-tidelevel on the muddy sandflat of Ronce-les-Bains from February 1995 to February 1996. Figures in brackets indicated percentages of totalitems
Year class Production Elimination Mean biomass P: B
g dry Wt m−2 yr−1 g dry Wt m−2 yr−1 g dry Wt m−2 yr−1
1995 (0-group) 14.12 (40%) −11.98 (32%) 2.65 (17%) 5.33
1994 14.60 (41%) −14.14 (37%) 8.67 (55%) 1.68
1993 0.65 (2%) 0.01 (<1%) 0.60 (<1%) 1.10
1992 4.04 (11%) −8.10 (21%) 2.99 (19%) 1.35
1991 2.08 ( 6%) −3.71 (10%) 1.28 (9%) 1.62
Total g dry Wt m−2 yr−1 35.49 −37.94 15.60 2.28
g AFDW m−2 yr−1 32.47 −34.70 14.27
g C m−2 yr−1 11.39 −12.18 5.01
Secondary production
The total annual production estimate was 32.47 gAFDW m−2 yr−1 (11.39 g C m−2 yr−1) at the mid-tide level (Table 1). Mean total biomass was 14.27g AFDW m−2 yr−1and thus, the average of produc-tion/biomass ratio was 2.28 yr−1. The relative con-tributions of each year class to the annual secondaryproduction were 40, 41, 2, 11 and 6% for 0-group,1-, 2-, 3- and 4-yr-old cockles, respectively (Table 1).The 1993 year class (2-yr-old cockles) appeared to bevery rare at the mid-tide level, potentially due to poorrecruitment in 1993, and did not contribute signific-antly to either annual production or annual elimina-tion. Production and elimination terms were similarfor 0-group and 1-yr-old cockles, whereas elimination
terms were two times greater than production termsfor 3- and 4-yr-old cockles, with−8.10 versus 4.04and−3.71 versus 2.08 g AFDW m−2 yr−1, respect-ively (Table 1). Total annual elimination was−34.7g AFDW m−2 yr−1 and was similar to total annualproduction.
Stable isotope ratios for cockles and potential foodsources
There were no significant seasonal changes in bothδ13C (p> 0.6) andδ15N values (p > 0.1) for POMas indicated by a 2-way ANOVA analysis (unbal-anced data with fixed factors ‘season’ and ‘station’and without test of interaction).δ13C andδ15N valuesfor POM from the water column averaged−22.2±
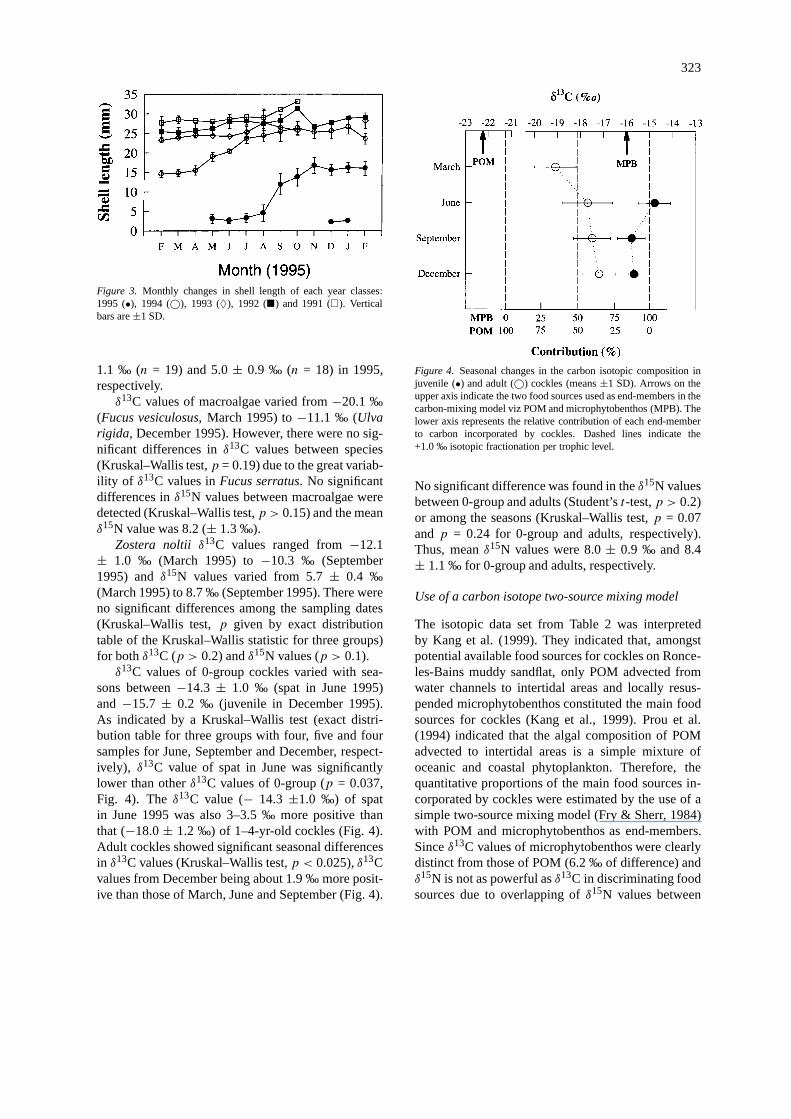
323
Figure 3. Monthly changes in shell length of each year classes:1995 (•), 1994 (©), 1993 (♦), 1992 (�) and 1991 (�). Verticalbars are±1 SD.
1.1 ‰ (n = 19) and 5.0± 0.9 ‰ (n = 18) in 1995,respectively.δ13C values of macroalgae varied from−20.1 ‰
(Fucus vesiculosus, March 1995) to−11.1 ‰ (Ulvarigida, December 1995). However, there were no sig-nificant differences inδ13C values between species(Kruskal–Wallis test,p = 0.19) due to the great variab-ility of δ13C values inFucus serratus. No significantdifferences inδ15N values between macroalgae weredetected (Kruskal–Wallis test,p > 0.15) and the meanδ15N value was 8.2 (± 1.3 ‰).
Zostera noltii δ13C values ranged from−12.1± 1.0 ‰ (March 1995) to−10.3 ‰ (September1995) andδ15N values varied from 5.7± 0.4 ‰(March 1995) to 8.7 ‰ (September 1995). There wereno significant differences among the sampling dates(Kruskal–Wallis test,p given by exact distributiontable of the Kruskal–Wallis statistic for three groups)for bothδ13C (p > 0.2) andδ15N values (p > 0.1).δ13C values of 0-group cockles varied with sea-
sons between−14.3 ± 1.0 ‰ (spat in June 1995)and−15.7± 0.2 ‰ (juvenile in December 1995).As indicated by a Kruskal–Wallis test (exact distri-bution table for three groups with four, five and foursamples for June, September and December, respect-ively), δ13C value of spat in June was significantlylower than otherδ13C values of 0-group (p = 0.037,Fig. 4). Theδ13C value (− 14.3 ±1.0 ‰) of spatin June 1995 was also 3–3.5 ‰ more positive thanthat (−18.0± 1.2 ‰) of 1–4-yr-old cockles (Fig. 4).Adult cockles showed significant seasonal differencesin δ13C values (Kruskal–Wallis test,p < 0.025),δ13Cvalues from December being about 1.9 ‰ more posit-ive than those of March, June and September (Fig. 4).
Figure 4. Seasonal changes in the carbon isotopic composition injuvenile (•) and adult (©) cockles (means±1 SD). Arrows on theupper axis indicate the two food sources used as end-members in thecarbon-mixing model viz POM and microphytobenthos (MPB). Thelower axis represents the relative contribution of each end-memberto carbon incorporated by cockles. Dashed lines indicate the+1.0 ‰ isotopic fractionation per trophic level.
No significant difference was found in theδ15N valuesbetween 0-group and adults (Student’st-test,p > 0.2)or among the seasons (Kruskal–Wallis test,p = 0.07and p = 0.24 for 0-group and adults, respectively).Thus, meanδ15N values were 8.0± 0.9 ‰ and 8.4± 1.1 ‰ for 0-group and adults, respectively.
Use of a carbon isotope two-source mixing model
The isotopic data set from Table 2 was interpretedby Kang et al. (1999). They indicated that, amongstpotential available food sources for cockles on Ronce-les-Bains muddy sandflat, only POM advected fromwater channels to intertidal areas and locally resus-pended microphytobenthos constituted the main foodsources for cockles (Kang et al., 1999). Prou et al.(1994) indicated that the algal composition of POMadvected to intertidal areas is a simple mixture ofoceanic and coastal phytoplankton. Therefore, thequantitative proportions of the main food sources in-corporated by cockles were estimated by the use of asimple two-source mixing model (Fry & Sherr, 1984)with POM and microphytobenthos as end-members.Sinceδ13C values of microphytobenthos were clearlydistinct from those of POM (6.2 ‰ of difference) andδ15N is not as powerful asδ13C in discriminating foodsources due to overlapping ofδ15N values between
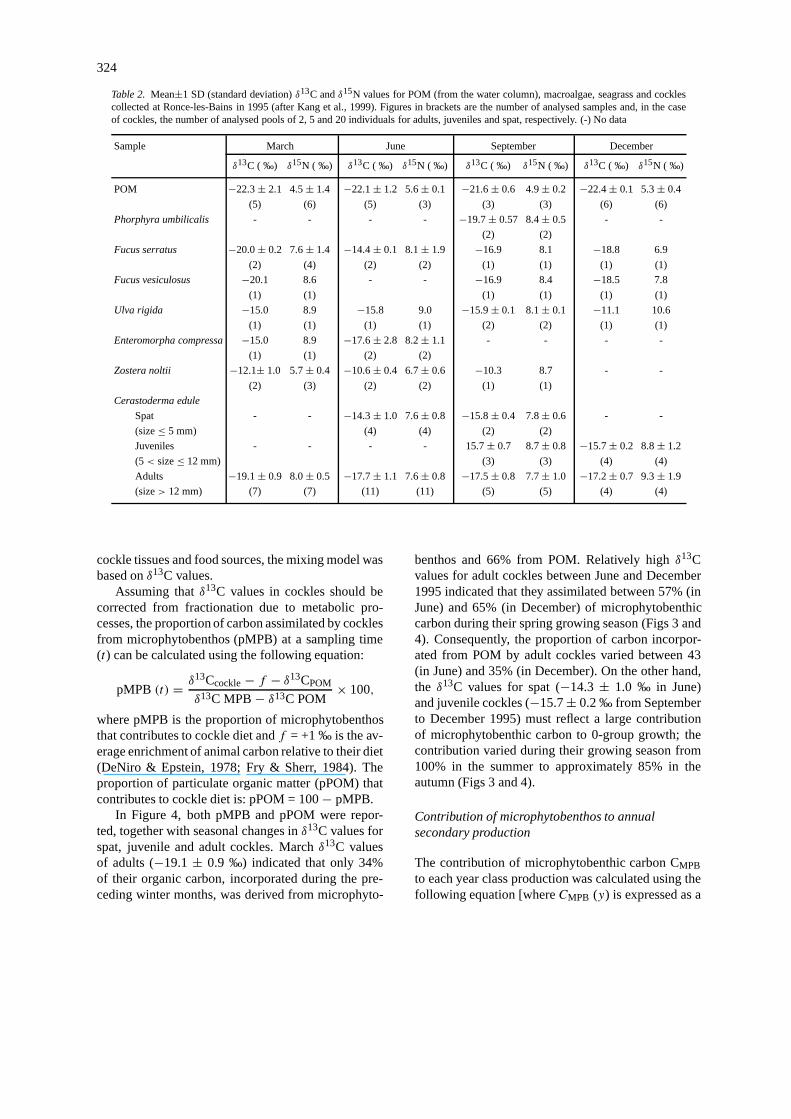
324
Table 2. Mean±1 SD (standard deviation)δ13C andδ15N values for POM (from the water column), macroalgae, seagrass and cocklescollected at Ronce-les-Bains in 1995 (after Kang et al., 1999). Figures in brackets are the number of analysed samples and, in the caseof cockles, the number of analysed pools of 2, 5 and 20 individuals for adults, juveniles and spat, respectively. (-) No data
Sample March June September December
δ13C ( ‰) δ15N ( ‰) δ13C ( ‰) δ15N ( ‰) δ13C ( ‰) δ15N ( ‰) δ13C ( ‰) δ15N ( ‰)
POM −22.3± 2.1 4.5± 1.4 −22.1± 1.2 5.6± 0.1 −21.6± 0.6 4.9± 0.2 −22.4± 0.1 5.3± 0.4
(5) (6) (5) (3) (3) (3) (6) (6)
Phorphyra umbilicalis - - - - −19.7± 0.57 8.4± 0.5 - -
(2) (2)
Fucus serratus −20.0± 0.2 7.6± 1.4 −14.4± 0.1 8.1± 1.9 −16.9 8.1 −18.8 6.9
(2) (4) (2) (2) (1) (1) (1) (1)
Fucus vesiculosus −20.1 8.6 - - −16.9 8.4 −18.5 7.8
(1) (1) (1) (1) (1) (1)
Ulva rigida −15.0 8.9 −15.8 9.0 −15.9± 0.1 8.1± 0.1 −11.1 10.6
(1) (1) (1) (1) (2) (2) (1) (1)
Enteromorpha compressa −15.0 8.9 −17.6± 2.8 8.2± 1.1 - - - -
(1) (1) (2) (2)
Zostera noltii −12.1± 1.0 5.7± 0.4 −10.6± 0.4 6.7± 0.6 −10.3 8.7 - -
(2) (3) (2) (2) (1) (1)
Cerastoderma edule
Spat - - −14.3± 1.0 7.6± 0.8 −15.8± 0.4 7.8± 0.6 - -
(size≤ 5 mm) (4) (4) (2) (2)
Juveniles - - - - 15.7± 0.7 8.7± 0.8 −15.7± 0.2 8.8± 1.2
(5< size≤ 12 mm) (3) (3) (4) (4)
Adults −19.1± 0.9 8.0± 0.5 −17.7± 1.1 7.6± 0.8 −17.5± 0.8 7.7± 1.0 −17.2± 0.7 9.3± 1.9
(size> 12 mm) (7) (7) (11) (11) (5) (5) (4) (4)
cockle tissues and food sources, the mixing model wasbased onδ13C values.
Assuming thatδ13C values in cockles should becorrected from fractionation due to metabolic pro-cesses, the proportion of carbon assimilated by cocklesfrom microphytobenthos (pMPB) at a sampling time(t) can be calculated using the following equation:
pMPB (t) = δ13Ccockle− f − δ13CPOM
δ13C MPB− δ13C POM× 100,
where pMPB is the proportion of microphytobenthosthat contributes to cockle diet andf = +1 ‰ is the av-erage enrichment of animal carbon relative to their diet(DeNiro & Epstein, 1978; Fry & Sherr, 1984). Theproportion of particulate organic matter (pPOM) thatcontributes to cockle diet is: pPOM = 100− pMPB.
In Figure 4, both pMPB and pPOM were repor-ted, together with seasonal changes inδ13C values forspat, juvenile and adult cockles. Marchδ13C valuesof adults (−19.1± 0.9 ‰) indicated that only 34%of their organic carbon, incorporated during the pre-ceding winter months, was derived from microphyto-
benthos and 66% from POM. Relatively highδ13Cvalues for adult cockles between June and December1995 indicated that they assimilated between 57% (inJune) and 65% (in December) of microphytobenthiccarbon during their spring growing season (Figs 3 and4). Consequently, the proportion of carbon incorpor-ated from POM by adult cockles varied between 43(in June) and 35% (in December). On the other hand,the δ13C values for spat (−14.3± 1.0 ‰ in June)and juvenile cockles (−15.7± 0.2 ‰ from Septemberto December 1995) must reflect a large contributionof microphytobenthic carbon to 0-group growth; thecontribution varied during their growing season from100% in the summer to approximately 85% in theautumn (Figs 3 and 4).
Contribution of microphytobenthos to annualsecondary production
The contribution of microphytobenthic carbon CMPBto each year class production was calculated using thefollowing equation [whereCMPB (y) is expressed as a

325
Table 3. Relative contribution of carbon derived from micro-phytobenthos to the annual secondary production and eliminationof Cerastoderma edule. 0-group (1995 cohort) and adult cockles(1991 to 1994 cohorts)
Year class Production Elimination Contribution of(age group) g C m−2 yr−1 g C m−2 yr−1 microphytobenthos
1995 (0-group) 4.53 −3.85 88%1994–1991 (adults) 6.86 −8.33 60%
Total 11.39 −12.18 71%
percentage of the production P(y) of a given year class(y)]:
CMPB(y) = 100· SUM/P(y)
with
SUM =t=n∑t=0
[(Nt +Nt+1)/2 · (Wt+1−Wt)] ·
pMPB(t + 1).
Nt , Wt andn are the terms used in the computation ofthe production P(y) of a year class (y) and pMPB (t+1)is the proportion of microphytobenthos that contrib-uted to the year class diet between the two samplingdatest andt+1.
Calculation of the termsCMPB (y) for the 0-groupand adult cockles are given in Table 3 and indicate thatthe relative contribution of microphytobenthos carbonto the annual secondary production of cockle carbonon average is 88% and 60% for the 0-group and adultcockles, respectively. The weighted contribution ofmicrophytobenthos to the annual production of thewhole population was more than 70% (Table 3).
Discussion
Seasonal recruitment patterns
The first spawning period of cockles at Ronce-les-Bains occurs in late spring, following winter andspring gonadal development in adult cockles, but asecond spawning period can also occur in late summer(Guillou et al., 1990; Sauriau, 1992). As a con-sequence, the major spatfall in early summer contrib-utes to the maintenance of the cockle stock, whereaslater recruitments are insignificant. These seasonalpatterns are in agreement with results of most studiesperformed along the North European coasts (Boyden,1971; Seed & Brown, 1975; Newell & Bayne, 1980;
Möller & Rosenberg, 1983; Navarro et al., 1989).However, Guillou et al. (1992) and Guillou & Tartu(1994) found opposite results in an intertidal popu-lation in northern Brittany. They reported, from a 5year study, that summer spawnings were followed byearly autumn to winter recruitment and that, in spiteof high post-larval mortality during winter, juvenilesborn during the foregoing autumn largely contributedto the population biomass during the following spring.It is also interesting to note that Seed & Brown (1978)in Northern Ireland found both opposite recruitmentpatterns in the same cockle population, but duringdifferent years: autumnal to early winter recruitmentin 1972, 1973 and 1975 and spring recruitment in1974. Year-to-year changes in local thermal and foodconditions, competition and predation pressures couldexplain such variabilities.
Production and elimination estimates
Total annual production and elimination of cockles inRonce-les-Bains fell within the range 0–200 g AFDWm−2 yr−1, as reported by Warwick & Price (1975),Hibbert (1976), Evans (1977), Ivell (1981) and Jensen(1993) for unexploited stocks. In the sandy mud-flat of Ronce-les-Bains, the cockle population suffershuman predation on cockles larger than 30 mm inshell length, mainly during the summer tourist season(Sauriau, 1992). Such a selective mortality rate leadsto a decrease in density and elimination exceedingproduction in all oldest cohorts (Table 1). This alsoinduces underestimation of growth rate estimates andlongevity. Hibbert (1976) reported 6-year-old cocklesin Southampton Water and the species is known to livefor up to 10 years in Northern Europe (Hancock &Urquhart, 1965; Jones, 1979). Consequently, total pro-duction estimates, calculated here, are underestimatesbecause methods used for estimating annual produc-tion (Crisp, 1984) assume that mortality is not sizedependent. It should, however, be pointed out thatselective removal of the oldest adult cohorts may fa-vour growth and survival of remaining juveniles andadults (de Montaudouin & Bachelet, 1996) and re-cruits of the following year (Bachelet et al., 1992)through intraspecific density-dependent relationships.Since P:B ratios of the youngest cohorts are generallymuch higher than those of oldest cohorts (e.g. Hibbert,1976; see also Brey, 1990), selective removal of oldestcohorts may also result, by feedback, in a relative over-contribution of production of youngest cohorts in totalannual production estimates.

326
Age-related differences in growing season and linkswith microphytobenthos availability
Due to recruitment patterns of the cockle popula-tion, a shift in growth seasons between juveniles andadults (> 1 yr old) was found at Ronce-les-Bains(Fig. 3). A similar pattern has been reported for othercockle populations by various studies (Farrow, 1971;Barnes, 1973; Jensen, 1992). Jensen (1992) hypothes-ised that juvenile and adult cockles exploit differentfood resources during their different growth periodsin relation to seasonal food availability. A similarmismatch-hypothesis was made by Ankar (1980) inthe case ofMacoma balthica. Hydrological surveysperformed within the Marennes-Oléron Bay (Héralet al., 1983; Razet et al., 1990; Soletchnik et al.,1998; Kang et al., 1999) and our stable carbon andnitrogen isotope results (Kang et al., 1999) supportthis hypothesis. From long-term monitoring in theMarennes-Oléron Bay, Soletchnik et al. (1998) in-dicated that maximum values of chlorophylla in thewater column appear in May and June. Results of the1995 hydrological survey (performed at three loca-tions surrounding the Ronce-les-Bains sandy mudflat,Figure 1), were also in agreement with this seasonalpattern: values up to 3.7± 1.7µg l−1 of chlorophyllawere found in POM during June 1995, whereas POMchlorophyll a values ranged from 0.5 to 0.9µg l−1
during the other seasons (Kang et al., 1999). However,surveys of sedimentary chlorophylla on the muddysandflat at Ronce-les-Bains in 1995 (Kang et al., 1999)indicated that sedimentary chlorophylla values werenot significantly different from 10.9± 3.5 µg g−1
dry sediment, all year round. Since seston dynamicswithin the Marennes-Oléron Bay are largely governedby the resuspension of intertidal sediments (Razet etal., 1990; Zurburg et al., 1994) under the influence oftidal currents and wind-driven waves (Raillard et al.,1994), it may be expected that resuspended benthicdiatoms are available for suspension feeding speciesmost of the time. Therefore, Zurburg et al. (1994)showed that, in the central water channel, benthic di-atom species formed about 50% of chlorophylla inthe water column at spring tide in May, and most ofthe chlorophylla in winter during both spring andneap tides. These observations are also in agreementwith various reports indicating that resuspended mi-crophytobenthos from large intertidal areas exposed tocurrent and wave actions becomes a major algal com-ponent in the water column (Baillie & Welsh, 1980;Delgado et al., 1991; De Jonge & Van Beusekom,
1995). These can be ingested subsequently by sus-pension feeders (De Jonge & Van Beusekom, 1992;Kamermans, 1994).
The age-related differences in the incorporationof microphytobenthos carbon by cockles in theMarennes-Oléron Bay is partly explained by the mis-match hypothesis. After settlement, 0-group cocklesgrew from summer to early autumn, a period with lowphytoplankton but high microphytobenthos abund-ance, whereas growth of adults mostly occurred inspring during phytoplankton blooms. Furthermore,juveniles have incorporated a higher proportion of mi-crophytobenthos in their tissues than adults. However,significant differences in theδ13C signature betweenspat in June (size≤5 mm) and juveniles (size>5mm) from the other seasons are not satisfactorily ex-plained. Similarly, the spring to autumnδ13C signatureof adults still appeared to be intermediate betweenPOM and microphytobenthos, and also significantlydifferent from theδ13C signature of juvenile evenafter a 6 month exposure to similar microphytobenthicfood source. Therefore, several possible explanationswere proposed by Kang et al. (1999): (1) size-relatedchanges in particle capture mechanism within thebenthic boundary layer, (2) size-related selective feed-ing capabilities, (3) size-related differential assimil-ation of dissolved organic matterversusparticulateorganic matter and (4) higher lipid contents of adultsduring reproductive winter to spring periods. Due tothe lack of evidence, to our knowledge, in the liter-ature onCerastoderma edule, for testing the validityof the three latter hypotheses, only the first explana-tion will be dealt with here. A closer connection ofjuvenile cockles to near-bottom layers compared withadult cockles, which have potentially longer inhalantsiphon and higher strength of inhalant/exhalant jets(André et al., 1993), may explain that spat and juven-iles are fueled mainly by microphytobenthos, whileadult growth is sustained by a higher proportion ofplanktonic items.
Role of microphytobenthos in cockle production
Estimates of the microphytobenthos proportion in theannual production of both 0-group and adult cocklesindicated that over 70% of the secondary productionof the whole population was fueled by microphyto-benthos. The fact thatCerastoderma edulefeed onepibenthic diatoms has been established by gut contentanalyses (e.g. Verwey, 1952; Ivell, 1981; Kamer-mans, 1994). However, our results suggest a much

327
stronger trophic link between microphytobenthos andthe infaunal suspension feederCerastoderma edulethan would be expected from these studies. On the onehand, examination of cockle gut contents generallygave qualitative results and, for instance, Ivell (1981)indicated that “in addition to their normal diet ofphytoplankton, in the Limfjord the cockles were alsotaking in benthic diatoms and other material includingsand grains, bivalve larvae and Foraminifera”. On theother hand, quantitative results of Kamermans (1994)indicated that cockle stomach contents appeared to beslightly enriched in benthic algae compared to algalcomposition of water samples taken at 1 cm above thebottom (70% pelagic algae, 30% benthic algae). Theover-proportion of benthic algae in cockle stomachsremained small and highly variable (between−30 and+50%) amongst cockles (Kamermans, 1994).
Similarly, our results appear to contradict the gen-erally accepted paradigm that marine phytoplanktonis a major component of the diet of suspension feed-ing cockles on sandy habitats (Newell, 1979; Loo& Rosenberg, 1989). It is not known whether thestrength of the trophic links between benthic diat-oms and cockles in the Marennes-Oléron Bay de-pends on ecophysiological capabilities of the speciesand/or trophic/hydrodynamic characteristics of thelocal muddy sandflats.Cerastoderma eduleis reportedto be able to selectively ingest microflora from a widevariety of food items (Iglesias et al., 1992; Navarroet al., 1992). The importance of microphytobenthosto the epifaunal suspension feederCrassostrea gigas(Thunberg), living in the vicinity of the River Char-ente plume in the north-east of the Marennes-OléronBay, has also been proven by isotope techniques (Ri-era & Richard, 1996, 1997). Microphytobenthos is,therefore, expected to be the major component ofthe total annual primary production in the Marennes-Oléron Bay (Guarini et al., 1998), where primaryproduction of phytoplankton is limited by water tur-bidity (Ravail et al., 1988; Raillard & Ménesguen,1994). Other primary producers, such as macroalgae,seagrass and marsh vascular plants, are restricted torelative small areas (Guillaumont, 1991) and most or-ganic matter is provided by resuspension of intertidalmuddy sediments (Razet et al., 1990; Zurburg et al.,1994).
In conclusion, this study highlights new hypo-theses on age-related processes that may influencethe suspension feedingCerastoderma eduletrophicbase primarily driven by temporal changes in theavailability of food resources in coastal ecosystems.
Acknowledgements
This study was supported by CNRS and IFREMER.C.-K. K. was supported by a grant from the FrenchEmbassy in the framework of a French-Korean co-operation programme between IFREMER and NFRDI(National Fisheries Research & Development Insti-tute). Thanks to M. Héral, G. Blanchard and P. Richardfor their support and to A. Knutsen for her commentson the English. We gratefully acknowledge two an-onymous referees for their helpful comments on themanuscript.
References
André, C. & R. Rosenberg, 1991. Adult-larval interactions in thesuspension-feeding bivalvesCerastoderma edule(L.) and MyaarenariaL. Mar. Ecol. Prog. Ser. 71: 227–234.
André, C., P. R. Jonsson & M. Lindegarth, 1993. Predation on set-tling bivalve larvae by benthic suspension feeders: the role ofhydrodynamics and larval behaviour. Mar. Ecol. Prog. Ser. 97:183–192.
Ankar, S., 1980. Growth and production ofMacoma balthica(L.) ina Northern Baltic soft bottom. Ophelia, Suppl. 1: 31–48.
Bachelet, G., J. Guillou & P. J. Labourg, 1992. Adult-larvaland juvenile interactions in the suspension-feeding bivalve,Cerastoderma edule(L.): field observations and experiments. InColombo, G., I. Ferrari, V. U. Ceccherelli & R. Rossi (eds), Mar-ine Eutrophication and Population Dynamics. Olsen & Olsen,Fredensborg: 175–182.
Baillie, P. W. & B. L. Welsh 1980. The effect of tidal resuspen-sion on the distribution of intertidal epipelic algae in an estuary.Estuar. coast. shelf Sci. 10: 165–180.
Baggerman, B., 1953. Spatfall and transport ofCardium eduleL.Arch. Néerl. Zool. 10: 315–342.
Barnes, R. S. K., 1973. The intertidal lamellibranchs of Southamp-ton Water, with particular reference toCerastoderma eduleandC. glaucum. Proc. malac. Soc. Lond. 40: 413–433.
Bayne, B. L., J. Widdows & R. I. E. Newell, 1977. Physiolo-gical measurements on estuarine bivalve molluscs in the field.In Keegan, B. F., P. O. Ceidigh & P. J. S Boaden (eds), Bio-logy of Benthic Organisms. Proc. 11th Europ. Mar. Biol. Symp.,Pergamon Press, Oxford: 57–68.
Beukema, J. J., 1982. Annual variation in reproductive success andbiomass of the major macrozoobenthic species living in a tidalflat area of the Wadden Sea. Neth. J. Sea Res. 16: 37–45.
Boyden, C. R., 1971. A comparative study of the reproductive cyclesof the cocklesCerastoderma eduleandC. glaucum. J. mar. biol.Ass. U.K. 51: 605–622.
Boutton, T. W., 1991. Stable carbon isotope ratios of natural ma-terials: I. Sample preparation and mass spectrometric analysis.In Coleman, D. C. & B. Fry (eds), Carbon Isotope Techniques.Academic Press, San Diego: 155–171.
Brey, T., 1990. Estimating productivity of macrobenthic inverteb-rates from biomass and mean individual weight. Meeresforsch32: 329–343.
Crisp, D. J., 1984. Energy flow measurements. In Holme, N. A. &A. D. McIntyre (eds), Methods for the Study of Marine Benthos,IBP Handbook 16. Blackwell Sci. Publ., Oxford: 284–372.

328
Dame, R. F., 1996. Ecology of marine bivalves. An ecosystem ap-proach. CRC Marine Science Series, CRC Press, Boca Raton,Florida: 254 pp.
De Jonge, V. N. & J. E. E. Van Beusekom, 1992. Contribution of re-suspended microphytobenthos to total phytoplankton in the Emsestuary and its possible role for grazers. Neth. J. Sea Res. 30:91–105.
De Jonge, V. N. & J. E. E. Van Beusekom, 1995. Wind- and tide-induced resuspension of sediment and microphytobenthos fromtidal flats in the Ems estuary. Limnol. Oceanogr. 40: 766–778.
Delgado, M., V. N. De Jonge & H. Peletier, 1991. Experimentson resuspension of natural microphytobenthos populations. Mar.Biol. 108: 321–328.
De Montaudouin, X. & G. Bachelet, 1996. Experimental evidenceof complex interactions between biotic and abiotic factors in thedynamics of an intertidal population of the bivalveCerastodermaedule. Oceanol. Acta. 19: 449–463.
DeNiro, M. J. & S. Epstein, 1978. Influence of diet on the distribu-tion of carbon isotopes in animals. Geochim. Cosmochim. Acta.42: 495–506.
DeNiro, M. J. & S. Epstein, 1981. Influence of diet on the distri-bution of nitrogen isotopes in animals. Geochim. Cosmochim.Acta. 45: 341–351.
Dörjes, J., H. Michaelis & B. Rhode, 1986. Long-term studiesof macrozoobenthos in intertidal and shallow subtidal habitatsnear the island of Norderney (East Frisian coast, Germany).Hydrobiologia 142: 217–232.
Ducrotoy, J. P., H. Rybarczyk, J. Souprayen, G. Bachelet, J. J.Beukema, M. Desprez, J. Dörjes, K. Essink, J. Guillou, H. Mi-chaelis, B. Sylvand, J. G. Wilson, B. Elkaïm & F. Ibanez, 1991. Acomparison of the population dynamics of the cockle (Cerasto-derma edule,L.) in North-Western Europe. In Elliott, M. &J. P. Ducrotoy (eds), Estuaries and Coasts: Spatial and Tem-poral Intercomparisons. ECSA 19 Symposium, Olsen & Olsen,Fredensborg: 173–184.
Evans, S., 1977. Growth, production and biomass release of a non-stable population ofCardium eduleL. (Bivalvia). Zoon 5: 133–141.
Farrow, G. E., 1971. Periodicity structures in the bivalve shell: ex-periments to establish growth controls inCerastoderma edulefrom the Thames Estuary. Ibid 14: 571–588.
Fry, B. & E. B. Sherr, 1984.δ13C measurements as indicators ofcarbon flow in marine and freshwater ecosystems. Contrib. mar.Sci. 27: 13–47.
Guarini, J.-M., G. F. Blanchard, C. Bacher, P. Gros, P. Riera, P.Richard, D. Gouleau, R. Galois, J. Prou & P.-G. Sauriau, 1998.Dynamics of spatial patterns of microphytobenthic biomass: in-ferences from a geostatistical analysis of two comprehensivesurveys in Marennes-Oléron Bay (France). Mar. Ecol. Prog. Ser.166: 131–141.
Guillaumont, B., 1991. Utilisation de l’imagerie satellitaire pourles comparaisons spatiales et temporelles en zone intertidale. InElliott, M. & J. P. Ducrotoy (eds), Estuaries and Coasts: Spatialand Temporal Intercomparisons. ECSA 19 Symposium, Olsen &Olsen, Fredensborg: 63–68.
Guillou, J., G. Bachelet, M. Desprez, J.-P. Ducrotoy, I. Madani,H. Rybarczyk, P.-G. Sauriau, B. Sylvand, B. Elkaim & M.Glémarec, 1990. Les modalités de la reproduction de la coque(Cerastoderma edule) sur le littoral français de la Manche et del’Atlantique. Aquat. Living Resour. 3: 29–41.
Guillou, J., G. Bachelet & M. Glémarec, 1992. Influence des fluctu-ations de température sur la reproduction et le recrutement de lacoqueCerastoderma edule(L.). Ann. Inst. Océanogr. Paris 68:65–74.
Guillou, J. & C. Tartu, 1994. Post-larval and juvenile mortality ina population of the edible cockleCerastoderma edule(L.) fromnorthern Brittany. Neth. J. Sea Res. 33: 103–111.
Hancock, D. A. & A. E. Urquhart, 1965. The determination of nat-ural mortality and its causes in an exploited population of cockles(Cardium eduleL.). M. A. F. F., Fish. Invest. 24: 1–40.
Hawkins, A. J. S., E. Navarro & J. I. P. Iglesias, 1990. Comparativeallometries of gut-passage time, gut content and metabolic faecalloss inMytilus edulisandCerastoderma edule. Mar. Biol. 105:197–204.
Héral, M., D. Razet, J.-M. Deslous-Paoli, J.-P. Berthomé & J.Garnier, 1983. Caractéristiques saisonnières de l’hydrobiologiedu complexe estuarien de Marennes-Oléron (France). Rev. Trav.Inst. Pêches. Marit. 46: 97–119.
Héral, M., C. Bacher & J.-M. Deslous-Paoli, 1990. La capacité bi-otique des bassins ostréicoles. In Troadec, J.-P. (ed.), L’hommeet les Ressources Halieutiques. IFREMER, Brest: 225–259.
Hibbert, C. J., 1976. Biomass and production of a bivalve com-munity on an intertidal mud-flat. J. exp. mar. Biol. Ecol. 25:249–261.
Iglesias, J. I. P. & E. Navarro, 1991. Energetics of growth andreproduction in cockles (Cerastoderma edule): seasonal andage-dependent variations. Mar. Biol. 111: 359–368.
Iglesias, J. I. P., E. Navarro, P. Alvarez Jorna & I. Armentia,1992. Feeding, particle selection and absorption in cocklesCerastoderma edule(L.) exposed to variable conditions of foodconcentration and quality. J. exp. mar. Biol. Ecol. 162: 177–198.
Ivell, R., 1981. A quantitative study of aCerastoderma– Nephthyscommunity in the Limfjord, Denmark, with special reference toproduction ofCerastoderma edule.J. Moll. Stud. 47: 147–170.
Jensen, K. T., 1992. Dynamics and growth of the cockle,Cerasto-derma edule, on an intertidal mud-flat in the Danish WaddenSea: effects of submersion time and density. Neth. J. Sea Res.28: 335–345.
Jensen, K. T., 1993. Density-dependent growth in cockles (Cerasto-derma edule): evidence from interannual comparisons. J. mar.biol. Ass. U.K. 73: 333–342.
Jones, A. M., 1979. Structure and growth of a high-level populationof Cerastoderma edule(Lamellibranchiata). J. mar. biol. Ass.U.K., 59: 277–287.
Junger, M. & D. Planas, 1994. Quantitative use of stable carbonisotope analysis to determine the trophic base of invertebratecommunities in a boreal forest lotic system. Can. J. Fish. aquat.Sci. 51: 52–61.
Kamermans, P., 1994. Similarity in food source and timing of feed-ing in deposit- and suspension-feeding bivalves. Mar. Ecol. Prog.Ser. 104: 63–75.
Kang, C. K., P.-G. Sauriau, P. Richard & G. F. Blanchard,1999. Food sources of the infaunal suspension-feeding bivalveCerastoderma edulein a muddy sandflat of Marennes-OléronBay, as determined by analyses of carbon and nitrogen stableisotopes. Mar. Ecol. Prog. Ser. 187: 147–158.
Kristensen, I., 1957. Differences in density and growth in a cocklepopulation in the Dutch Wadden sea. Arch. Néerl. Zool. 12: 350–453.
Loo, L. O. & R. Rosenberg, 1989. Bivalve suspension-feeding dy-namics and benthic-pelagic coupling in an eutrophicated marinebay. J. exp. mar. Biol. Ecol. 130: 253–276.
Michener, R. H. & D. M. Schell, 1994. Stable isotope ratios astracers in marine aquatic food webs. In Lajtha, K. & R. H.Michener (eds), Stable Isotopes in Ecology and EnvironmentalScience. Blackwell Scientific Publications, Oxford: 138–157.

329
Möller, P. & R. Rosenberg, 1983. Recruitment, abundance and pro-duction ofMya arenariaandCardium edulein marine shallowwaters, western Sweden. Ophelia 22: 33–35.
Navarro, E., J. I. P. Iglesias & A. Larranaga, 1989. Interannual vari-ation in the reproductive cycle and biochemical composition ofthe cockleCerastoderma edulefrom Mundaca Estuary (Biscay,North Spain). Mar. Biol. 101: 503–511.
Navarro, E., J. I. P. Iglesias & M. M. Ortega, 1992. Natural sedimentas a food source for the cockleCerastoderma edule(L.): effectof variable particle concentration on feeding, digestion and thescope for growth. J. exp. mar. Biol. Ecol. 156: 69–87.
Navarro, J. M. & J. Widdows, 1997. Feeding physiology ofCerasto-derma edulein response to a wide range of seston concentrations.Mar. Ecol. Prog. Ser. 152: 175–186.
Newell, R. C., 1979. Biology of Intertidal Animals. 3rd edn. MarineEcological Surveys Ltd., Faversham, Kent: 781 pp.
Newell, R. I. E. & B. J. Bayne, 1980. Seasonal changes inthe physiology, reproductive condition and carbohydrate con-tent of the cockleCardium (= Cerastoderma) edule (Bivalvia:Cardiidae). Mar. Biol. 56: 11–19.
Orton, J. H., 1933. Summer mortality of cockles on some Lan-cashire and Cheshire Dee beds in 1933. Nature, London 132:314–315.
Prins, T. C., A. C. Smaal & A. J. Pouwer, 1991. Selective in-gestion of phytoplankton by the bivalvesMytilus edulisL. andCerastoderma edule(L.). Hydrobiol. Bull. 25: 93–100.
Raillard, O. & A. Ménesguen, 1994. An ecosystem box model forestimating the carrying capacity of a macrotidal shellfish system.Mar. Ecol. Prog. Ser. 115: 117–130.
Raillard, O., P. Le Hir & P. Lazure, 1994. Transport de sédimentsfins dans le bassin de Marennes-Oléron: mise en place d’unmodèle mathématique. Houille Blanche 4: 63–71.
Ravail, B., M. Héral, S. Maestrini & J. M. Robert, 1988. Incid-ence du débit de la Charente sur la capacité biotique du bassinostréicole de Marennes-Oléron. J. Rech. Océanogr. 13: 48–52.
Razet, D., M. Héral, J. Prou, J. Legrand & J.-M. Sornin, 1990.Variations des productions de biodépôts (feces et pseudofeces)de l’huîtreCrassostrea gigasdans un estuaire macrotidal : baiede Marennes-Oléron. Haliotis 10: 143–161.
Reise, K., 1985. Tidal flat ecology. An experimental approach tospecies interactions. Springer-Verlag, Berlin: 191 pp.
Riera, P. & P. Richard, 1996. Isotopic determination of food sourcesof Crassostrea gigasalong a trophic gradient in the estuarine bayof Marennes-Oléron. Estuar. coast. shelf Sci. 42: 347–360.
Riera, P. & P. Richard, 1997. Temporal variation ofδ13C in particu-late organic matter and oysterCrassostrea gigasin Marennes-Oléron Bay (France): effect of freshwater inflow. Mar. Ecol.Prog. Ser. 147: 105–115.
Riera, P., P. Richard, A. Grémare & G. Blanchard, 1996. Foodsource of intertidal nematodes in the Bay of Marennes-Oléron(France), as determined by dual stable isotope analysis. Mar.Ecol. Prog. Ser. 142: 303–309.
Sanchez-Salazar, M. E., C. L. Griffiths & R. Seed, 1987. The effectof size and temperature on the predation of cocklesCerasto-derma edule(L.) by the shore crabCarcinus maenas(L.). J. exp.mar. Biol. Ecol. 111: 181–193.
Sauriau, P.-G., 1992. Les mollusques benthiques du bassin deMarennes-Oléron : estimation et cartographie des stocks non cul-tivés, compétition spatiale et trophique, dynamique de popula-tion deCerastoderma edule(L.). Unpub. PhD. Thesis, Universitéde Bretagne Occidentale, Brest: 309 pp.
Sauriau, P.-G., V. Mouret & J.-P. Rincé, 1989. Organisationtrophique de la malacofaune benthique non cultivée du bassinostréicole de Marennes-Oléron. Oceanol. Acta. 12: 193–204.
Seed, R. & R. A. Brown, 1975. The influence of reproductivecycle, growth and mortality on population structure inModiolusmodiolus(L.), Cerastoderma edule(L.) and Mytilus edulisL.,(Mollusca: Bivalvia). In Barnes, H. (ed.), The Biochemistry,Physiology and Behaviour of Marine Organisms in Relation totheir Ecology. Proc. 9th Europ. Mar. Biol. Symp., AberdeenUniversity Press, Aberdeen: 257–274.
Seed, R. & R. A. Brown, 1978. Growth as a strategy for sur-vival in two marine bivalves,Cerastoderma eduleandModiolusmodiolus. J. anim. Ecol. 47: 283–292.
Siegel, S., 1956. Nonparametric statistics for the behavioral sci-ences. McGraw-Hill, New York: 312 pp.
Smaal, A. C., A. P. M. A. Vonck & M. Bakker, 1997. Season-nal variation in physiological energetics ofMytilus edulisandCerastoderma eduleof different size classes. J. mar. biol. Ass.U. K. 77: 817–838.
Sokal, R. & F. J. Rohlf, 1981. Biometry. 2nd edn. W.H. Freeman &Co, New York: 859 pp.
Soletchnik, P., N. Faury, D. Razet & Ph. Goulletquer, 1998.Hydrobiology of the Marennes-Oléron bay. Seasonal indicesand analysis of trends from 1978 to 1995. Hydrobiologia 386:131–146.
Tebble, N., 1966. British bivalve seashells. A handbook for iden-tification. Trustees of the British Museum (Natural History),London: 212 pp.
Thiel, M. & L. Watling, 1998. Effects of green algal mats on in-faunal colonization of a New England mud flat – long-lasting buthighly localized effects. Hydrobiologia 375/376: 177–189.
Urrutia, M. B., J. I. P. Iglesias, E. Navarro & J. Prou, 1996. Feedingand absorption inCerastoderma eduleunder environmental con-ditions in the Bay of Marennes-Oléron (western France). J. mar.biol. Ass. U. K. 76: 431–450.
Verwey, J., 1952. On the ecology of distribution of cockle and mus-sel in the Dutch Wadden Sea, their role in sedimentation and thesource of their food supply. Arch. Néerl. Zool. 10: 171–239.
Warwick, R. M., I. R. Joint & P. J. Radford, 1979. Secondary pro-duction of the benthos in an estuarine environment. In Jefferies,R. L. & A. J. Davy (eds), Ecological Processes in Coastal Envir-onments. Blackwell Scientific Publications, Oxford: 429–450.
Warwick, R. M. & R. Price, 1975. Macrofauna production in anestuarine mud-flat. J. mar. biol. Ass. U. K. 55: 1–18.
Zurburg, W., A. A. D. Smaal, M. Héral & N. Danker, 1994. Sestondynamics and bivalve feeding in the bay of Marennes-Oléron(France). Neth. J. aquat. Ecol. 26: 459–466.
Related Documents











