© 2010 ISHAM DOI: 10.3109/13693780903496591 Medical Mycology August 2010, 48, 735–743 Splenic and lung response to nonlethal systemic Aspergillus fumigatus infection in C57BL/6 mice IVANA MIRKOV*, IVANA STOJANOVIC†, STANISLAVA STOSIC-GRUJICIC†, JASMINA GLAMOCLIJA*, LIDIJA ZOLOTAREVSKI‡, DRAGAN KATARANOVSKI*§ & MILENA KATARANOVSKI*# *Department of Ecology & †Department of Immunology, Institute for Biological Research ‘Sinisa Stankovic’, University of Belgrade, Belgrade, ‡Institute for Pathology, Military Medical Academy, Belgrade, §Institute of Zoology, Faculty of Biology, University of Belgrade, Belgrade, and #Institute of Physiology and Biochemistry, Faculty of Biology, University of Belgrade, Belgrade, Serbia In this study, we investigated splenic and lung cell responses to nonlethal systemic Aspergillus fumigatus infection in mice. Apart from basic indices of spleen and lung cell activity, IL-17 expression by cells from both tissues was determined and compared to the expression of IFN- and IL-4. Progressive decrease in tissue fungal burden cor- related with increased spleen and lung cell activity. Increased IL-17 message was noted in spleen cells at all time points (3, 7 and 15 days post-infection; p.i.), while a modest increase in IFN- mRNA expression was noted at day 3 p.i. Increased cytokine produc- tion at days 3 and 7 (IL-17) and throughout the experimental period (IFN-) was found. In contrast, spleen cell IL-4 expression was considerably lower during infection, result- ing in high IFN-/IL-4 and IL-17/IL-4 ratios in the spleen. Pro-inflammatory cytokine response was observed in the lungs as well, but primarily as the result of increased production of IFN- by lung cells in response to challenge with conidia and the absence of change in IL-4 response. Increased activity of cells from both tissues, as well as the pattern of cytokine production, created an optimal pro-inflammatory milieu for fungal eradication. Keywords nonlethal murine systemic aspergillosis, spleen, lungs, IL-17, IFN-, IL-4 Animal models of aspergillosis have been used exten- sively to study various aspects of infection caused by this fungus. Murine models have been used to study host resis- tance in the four main forms of invasive aspergillosis; pul- monary aspergillosis (IPA), bronchopulmonary aspergillosis (ABPA), systemic (disseminated) infection and central ner- vous system (CNS) infection [7]. The use of these models has been indispensable in enhancing our current under- standing of both pathogenesis and host resistance to Asper- gillus infections. Utilizing the intranasal or intratracheal route of inocu- lating mice with Aspergillus conidia (to simulate the natu- ral route of infection), the role of innate host defense and the generation of Th1 cytokines by interstitial lymphocytes was shown to play a critical role in the resistance to inva- sive pulmonary aspergillosis [6,8,9]. Immunosuppressive treatments (e.g., glucocorticoids, cyclophosphamide, etc), are required to increase the mouse’s susceptibility to infec- tion [10,11]. Inoculation of A. fumigatus by a systemic route (i.e., intravenous inoculation) leads to infection in Received 7 July 2009; Received in final revised form 22 October 2009; Accepted 18 November 2009 Correspondence: Milena Kataranovski, Department of Ecology, Institute for Biological Research ‘Sinisa Stankovic’, University of Belgrade, Belgrade, Despota Stefana 142, Serbia. Tel: +381 11 2078375; Fax +381 11 2761433; E-mail: [email protected] Introduction Aspergillus fumigatus is a ubiquitous saprophytic mould that forms airborne conidia [1–3]. The small diameter of the conidia, as well as environmental factors, make them easily accessible to the lungs [4,5]. Infections occur rarely in healthy, immunocompetent individuals, as conidia are efficiently eliminated by host immune defenses [1]. Thus, A. fumigatus is regarded as a weak pathogen, responsible in immunocompetent individuals for only allergic aspergillosis or aspergilloma. With the increasing number of immunocompromised/immunodeficient individuals, A. fumigatus has emerged as the second major cause of invasive fungal infections in such patients [1]. Med Mycol Downloaded from informahealthcare.com by University of Newcastle Upon Tyne on 02/03/11 For personal use only.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Medical Mycology August 2010, 48, 735–743
Med
Myc
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
02/
03/1
1Fo
r pe
rson
al u
se o
nly.
Splenic and lung response to nonlethal systemic
Aspergillus fumigatus infection in C57BL/6 mice
© 2010 ISHAM
IVANA MIRKOV*, IVANA STOJANOVIC†, STANISLAVA STOSIC-GRUJICIC†, JASMINA GLAMOCLIJA*,
LIDIJA ZOLOTAREVSKI‡, DRAGAN KATARANOVSKI*§ & MILENA KATARANOVSKI*#
*Department of Ecology & †Department of Immunology, Institute for Biological Research ‘Sinisa Stankovic’, University of Belgrade,
Belgrade, ‡Institute for Pathology, Military Medical Academy, Belgrade, §Institute of Zoology, Faculty of Biology, University of
Belgrade, Belgrade, and #Institute of Physiology and Biochemistry, Faculty of Biology, University of Belgrade, Belgrade, Serbia
Received 7 July 2009; Received i
Accepted 18 November 2009
Correspondence: Milena Katar
Institute for Biological Researc
Belgrade, Belgrade, Despota Stefa
Fax +381 11 2761433; E-mail: mil
In this study, we investigated splenic and lung cell responses to nonlethal systemic Aspergillus fumigatus infection in mice. Apart from basic indices of spleen and lung cell activity, IL-17 expression by cells from both tissues was determined and compared to the expression of IFN-� and IL-4. Progressive decrease in tissue fungal burden cor-related with increased spleen and lung cell activity. Increased IL-17 message was noted in spleen cells at all time points (3, 7 and 15 days post-infection; p.i.), while a modest increase in IFN-� mRNA expression was noted at day 3 p.i. Increased cytokine produc-tion at days 3 and 7 (IL-17) and throughout the experimental period (IFN-�) was found. In contrast, spleen cell IL-4 expression was considerably lower during infection, result-ing in high IFN-�/IL-4 and IL-17/IL-4 ratios in the spleen. Pro-infl ammatory cytokine response was observed in the lungs as well, but primarily as the result of increased production of IFN-� by lung cells in response to challenge with conidia and the absence of change in IL-4 response. Increased activity of cells from both tissues, as well as the pattern of cytokine production, created an optimal pro-infl ammatory milieu for fungal eradication.
Keywords nonlethal murine systemic aspergillosis, spleen, lungs, IL-17, IFN-�, IL-4
Introduction
Aspergillus fumigatus is a ubiquitous saprophytic mould
that forms airborne conidia [1–3]. The small diameter of
the conidia, as well as environmental factors, make them
easily accessible to the lungs [4,5]. Infections occur rarely
in healthy, immunocompetent individuals, as conidia are
effi ciently eliminated by host immune defenses [1]. Thus,
A. fumigatus is regarded as a weak pathogen, responsible
in immunocompetent individuals for only allergic
aspergillosis or aspergilloma. With the increasing number
of immunocompromised/immunodefi cient individuals,
A. fumigatus has emerged as the second major cause of
invasive fungal infections in such patients [1].
n fi nal revised form 22 October 2009;
anovski, Department of Ecology,
h ‘Sinisa Stankovic’, University of
na 142, Serbia. Tel: +381 11 2078375;
Animal models of aspergillosis have been used exten-
sively to study various aspects of infection caused by this
fungus. Murine models have been used to study host resis-
tance in the four main forms of invasive aspergillosis; pul-
monary aspergillosis (IPA), bronchopulmonary aspergillosis
(ABPA), systemic (disseminated) infection and central ner-
vous system (CNS) infection [7]. The use of these models
has been indispensable in enhancing our current under-
standing of both pathogenesis and host resistance to Asper-gillus infections.
Utilizing the intranasal or intratracheal route of inocu-
lating mice with Aspergillus conidia (to simulate the natu-
ral route of infection), the role of innate host defense and
the generation of Th1 cytokines by interstitial lymphocytes
was shown to play a critical role in the resistance to inva-
sive pulmonary aspergillosis [6,8,9]. Immunosuppressive
treatments (e.g., glucocorticoids, cyclophosphamide, etc),
are required to increase the mouse’s susceptibility to infec-
tion [10,11]. Inoculation of A. fumigatus by a systemic route (i.e., intravenous inoculation) leads to infection in
DOI: 10.3109/13693780903496591

© 2010 ISHAM, Medical Mycology, 48, 735–743
736 Mirkov et al.
Med
Myc
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
02/
03/1
1Fo
r pe
rson
al u
se o
nly.
immunocompetent mice with a relatively uniform pattern
of disease. This model of infection is used for disseminated
infection studies, infection-mortality ratio investigations
and is particularly useful in the determination of mecha-
nisms involved in host resistance or susceptibility to lethal
A. fumigatus infection [10,12–15]. Differential production
of cytokines that favor Th1 response (e.g., IFN-�) and Th2
cytokines (e.g., IL-4 and IL-10) by splenic CD4+ cells was
demonstrated in mice found to be resistant or susceptible
to lethal A. fumigatus infections. Although IFN-� was
largely responsible for the resistance to lethal dose of the
fungus, the production of Th2 cytokines was associated
with lethal outcome [13]. Relative resistance to lethal sys-
temic infection could be induced by IFN-� administration
[14] or IL-4 neutralization [13], as well as being found in
mice lacking a functional IL-10 [15].
Although these cited studies clearly demonstrated the
signifi cance of splenic Th1 and Th2 cytokines in the resis-
tance or susceptibility to lethal systemic A. fumigatus infec-
tion, there are few data regarding the relevance of these
cytokines in nonlethal infections. Although these infections
have some of the characteristics of systemic disease, little
attention has been paid to other target tissues. Additionally,
there is, to our knowledge, no data concerning the relevance
of the principal cytokine of Th17 subset, interleukin-17
(IL-17) during systemic infection with A. fumigatus. The
importance of activation of Th17 cells in immune response
to fungi was stressed recently [16,17] and the involvement
of IL-17 in innate host defense to pulmonary aspergillosis
with A. fumigatus has been demonstrated [18,19].
Our previous study demonstrated histopathological
changes in lungs and kidneys of mice that received lethal
or sublethal intravenous doses of A. fumigatus conidia [12].
Using this model of disseminated aspergillosis, we exam-
ined spleen and pulmonary cell responses to nonlethal
doses of A. fumigatus. Basic indices of spleen and lung
activity including changes in mass, cellularity and cell pro-
liferation, as well as leukocyte infi ltration and cell activa-
tion, were determined. Quantitative and temporal expression
of IL-17 in spleen and lung cells were compared to the
expression of IFN-� and IL-4. These data indicated that
resistance to systemic aspergillosis is associated with a
proinfl ammatory response, both in the spleen and the lungs.
Such a response coincided with a progressive decrease in
conidial burden, suggesting its contribution in effi cient
elimination of fungi from these tissues.
Materials and methods
Mice
Eight- to 12-week- old female C57BL/6 mice, four to six
per group, were used for the study. Animals were bred and
housed conventionally in a controlled environment at the
Institute for Biological Research ‘Sinisa Stankovic’
(Belgrade, Serbia) and provided with standard rodent chow
and water ad libitum. All experiments were approved by
the Ethical Committee of our Institute. Mice were eutha-
nized on days 1, 3, 7 and 15 post-infection (p.i.) for fungal
burden assessment, and at days 3, 7 and 15 for the other
analyses.
Fungal culture conditions and infection
A human isolate of A. fumigatus was obtained from the
Institute of Public Health of Serbia ‘Dr Milan Jovanovic-
Batut’. The fungus was grown on Sabouraud maltose agar
(SMA, Torlak, Belgrade, Serbia) for 7 days [20]. Conidia
were harvested by fl ooding the surface of agar slants with
nonpyrogenic sterile physiological saline and the suspen-
sion adjusted to a concentration of 107 conidia/ml. Anes-
thetized mice (Ketamidor, Richter Pharma, Austria) were
inoculated intravenously with 106 conidia in 0.1 ml of
pyrogen-free saline via the lateral tail vein. Control mice
received saline only. In restimulation experiments, conidia
that had been autoclaved at 121°C for 30 min
(heat-inactivated) were used.
Fungal burden assessment
Fungal burden in lungs, spleen and kidneys was deter-
mined by quantitative colony forming units (CFU) [21]
assay. At selected time points (i.e., 1, 3, 7 and 15 days) p.i., organs were removed aseptically and their wet masses
were determined by weighing them using a precision bal-
ance (� 0.01 g). Tissue specimens were homogenized
using an IKA T18 basic homogenizer (IKA Works INC.,
Wilmington, NC) in 5 ml of sterile saline, on ice. Serial
dilutions of the primary homogenates were inoculated on
SMA plates supplemented with streptomycin sulfate (ICN-
Galenika, Belgrade, Serbia), and incubated at 37°C for
24 to 48 h. The number of CFU per gram of tissue was
determined as described.
Spleen cell preparation and culture
Spleens were removed, blotted dry, and weighed. The rel-
ative spleen mass was calculated according to the follow-
ing formula: organ mass/body mass � 100. Spleen cells
were obtained aseptically by gentle teasing the spleen tis-
sue through a cell strainer (BD Falcon, BD Bioscience,
Bedford, USA). Cells were suspended in RPMI-1640 cul-
ture medium (Flow, ICN Pharmaceuticals) supplemented
with 2 mM glutamine, 20 μg/ml gentamicin (Galenika a.d.,
Serbia) with 5 % (v/v) heat-inactivated fetal calf serum

Proinfl ammatory response in systemic murine aspergillosis 737
Med
Myc
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
02/
03/1
1Fo
r pe
rson
al u
se o
nly.
(PAA laboratories, Austria) (complete medium) and
voriconazole (5 μg/ml) (Pfi zer PGM, France) and adjusted
to 3 � 106/ml cells per cell culture well.
The capacity of spleen cells to proliferate was deter-
mined ex vivo or following restimulation with
heat-inactivated conidia of A. fumigatus at a ratio of conidia
to spleen cells of 0.06:1. Briefl y, 0.3 � 106 cells/well
(96-well plate) were cultivated for 24 h (ex vivo prolifera-
tion) or 72 h (restimulated proliferation), with 0.5 μCi of
[3H]-thymidine (GE Healthcare, UK) per well during the
last 8 hours of culture. Incorporation of [3H]-thymidine
into cellular DNA was measured by liquid scintillation
counting (LKB, Wallac).
For cytokine production, 5 � 106/ml spleen cells/well
(24-well plate) were cultured for 72 h in medium without
conidia to assess spontaneous production or with
heat-inactivated conidia of A. fumigatus at a conidia:spleen
cell ratio of 2:1 (restimulation). Cytokine level in the con-
ditioned medium was determined by enzyme-linked immu-
nosorbent assay (ELISA). Cytokine production after in vitro restimulation is expressed as the index of stimulation,
calculated according to the formula of restimulated pro-
duction/spontaneous production.
RNA isolation and RT-PCR
Total RNA from splenocytes (5 � 106 cells) was isolated
with a mi-Total RNA Isolation Kit (Metabion, Martinsried,
Germany) according to the manufacturer’s instructions.
RNA (1 μg) was reverse transcribed using Moloney leuke-
mia virus reverse transcriptase and random hexamers (both
from Fermentas, Vilnius, Lithuania). PCR amplifi cation of
cDNA (1 μl per 20 μl of PCR reaction) was carried out in
a Real-Time PCR machine (Applied Biosystems, UK)
using SYBRGreen PCR master mix (Applied Biosystems)
as follows: 10 min at 50°C for dUTP activation, 10 min at
95°C for initial denaturation of cDNA followed by
40 cycles (15 s of denaturation at 95°C and 60 s for
primer annealing and chain extension step). Primer pairs
for IL-17 were 5′-GGGAGAGCTTCATCTGT-3′ and
5′-GACCCTGAAAGTGAAGGG-3′ (GenBank accession
no. NM 010552.3), IFN-� 5′-CATCAGCAACAA-
CATAAGCGTCA-3′ and 5′-CTCCTTTTCCGCTTC-
CTGA-3′ (GenBank accession no. NM 008337.2), IL-4
5′-ATCCTGC TCTTCTTTCTCG-3′ and 5′-GATGCTC
TTTAGGCTTTCC-3′ (GenBank accession no. NM
021283.1) and β-actin 5′-GGACCTGACAGACTACC-3′ and 5′-GGCATAGAGGTCTTTACGG-3′ (NM 007393.2).
The expression of these genes was calculated according
to the formula 2-(Cti-Cta) where Cti is the cycle threshold
of the gene of interest and Cta is the cycle threshold value
of β-actin.
© 2010 ISHAM, Medical Mycology, 48, 735–743
Pulmonary cell isolation and activity assays
Lungs were removed aseptically and their wet masses
determined. The relative lung mass was calculated by the
following formula, lung mass/body mass � 100. Lung leu-
kocytes were obtained by digestion with 1 mg/ml type IV
collagenase (Worthington Biochemical Corporation, Lake-
wood, USA) and 30 μg/ml Deoxyribonuclease I (DNase I,
Sigma, Sigma Chemical Co., St Louis, MO, USA) for half
an hour at 37°C. Cells were resuspended in the complete
medium and adjusted to 4 � 106 cells/ml.
The activation of lung cells was evaluated by a quanti-
tative cytochemical assay for the respiratory burst [22],
based upon the spontaneous or phorbol-12-myristate
13-acetate (PMA, Sigma, Sigma Chemical Co., St Louis,
MO, USA) stimulated capacity of the cells to reduce
nitroblue tetrazolium (NBT, ICN Pharmaceutical, Costa
Mesa, CA, USA). Briefl y, 0.2 � 106 cells/well (96-well
plate) were cultured with 500 μg/ml NBT, with or without
(spontaneous) 100 ng/ml PMA (PMA-stimulated NBT
reduction) for 30 min. Formazan produced by cells was
extracted overnight in 10% SDS-0.01N HCl, and optical
density (OD) measured by microplate spectrophotometer
(GRD, Rome, Italy) at 540 and 650 nm. For the measure-
ment of cytokine production, 0.2 � 106 cells/well (96-well
plate) were cultured for 72 h with heat-inactivated conidia
of A. fumigatus at a ratio of 2:1 conidia to cells (restimu-
lated production) or with medium only (spontaneous
production).
ELISA
Cytokine production was determined in the medium con-
ditioned by spleen or lung cells using commercially avail-
able ELISA sets for mouse IL-17 (BD Pharmingen, San
Diego, CA, USA), mouse IFN-� (eBioscience Inc, San
Diego, CA, USA) and mouse IL-4 (R&D Systems,
Minneapolis, USA). Tests were done according to the
manufacturers’ recommendations. Cytokine titer was cal-
culated by reference to a standard curve constructed using
known amounts of recombinant IFN-�, IL-4 or IL-17.
Lung histology
Lung tissue was excised, immediately fi xed in 4% formal-
dehyde (pH 6.9), and embedded in paraffi n wax for sec-
tioning at 5 μm. Hematoxylin and eosin (H&E)-stained
histological slides were analyzed subsequently.
Statistics
Results were pooled from at least three experiments and
expressed as mean values � SD. Results were analyzed

738 Mirkov et al.
Med
Myc
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
02/
03/1
1Fo
r pe
rson
al u
se o
nly.
statistically using a Mann-Whitney U test for fungal bur-
den and cell culture data, and a t-test for gene expression.
P-values less than 0.05 were considered signifi cant.
Results
Conidial burden in spleen and lungs following i.v. A. fumigatus
injection
Determination of tissue fungal burden at day 1 postinfec-
tion revealed the highest number of CFU/g in the spleen
(7400 � 5), followed by lungs (352 � 19) and kidneys
(123 � 10). A progressive decrease in the number of CFUs
in these tissues was noted, with conidial elimination at
days 7 and 15 in lungs and spleen, respectively (Fig. 1 A,
B and C). From day 7 onward, studies of tissue homoge-
nates of all animals indicated that they were free of infec-
tion. At day 15, no fungal growth was detected in any
spleen homogenates or in two of four kidney homogenates.
Given the more pronounced, as well as sharper, drop in
conidial burden in spleen and lungs compared to kidneys,
changes in splenocyte and pulmonary cell activity were
investigated further.
Spleen response to conidia application
Basic parameters relevant for antifungal response in the
spleen were examined and included changes in organ mass,
cellularity, cell proliferation rate and cytokine expression.
Increased relative spleen mass and cell numbers compared
to spleens of control animals were noted at all time points
following infection (Fig. 2A and 2B). An increase in
ex vivo proliferation of spleen cells was found at days
3 and 7 postinfection with a tendency (P � 0.07) towards
increases in cell proliferation at day 15 (Fig. 2C). Restim-
ulation with A. fumigatus conidia resulted in increased
spleen cell proliferation at days 3 and 7. However, this did
not reach statistical signifi cance at day 3 (Fig. 2D). Increase
in spleen cellularity correlated with a decrease in spleen
fungal burden (r � −0.84; P � 0.001; y � 229.8197 −
0.0792*x). A signifi cant positive correlation was noted
between spleen cellularity and spleen mass (r � 0.74; P �
0.001; y � 0.02 + 2.9365E-5*x) and between spontaneous
splenocyte proliferation and spleen cellularity (r � 0.61;
P � 0.001; y � 145.5 + 0.0019*x).
Examination of the kinetics of spleen cytokine gene-
expression after infection with conidia (Fig. 3) revealed
increased IL-17 mRNA levels in spleen cells throughout
the p.i. period, but did not reach statistical signifi cance at
day 7. Signifi cantly, more IFN-� message was noted on
day 3 only. In contrast, a decrease in IL-4 mRNA expres-
sion was found for all time points examined. High IL-17/
IL-4 mRNA ratio of 1.7 � 1.1 (P � 0.05), 2.9 � 2.5 and
1.8 � 1.1, (P � 0.05) at days 3, 7 and 15, respectively,
compared to 0.2 � 0.1 in controls was noted. In addition,
IFN-�/IL-4 ratios of 1473.7 � 812.4, (P � 0.05), 1258.9
� 805.0, (P � 0.05) and 933.1 � 306.5, (P � 0.05) at
days 3, 7 and 15, respectively, compared to 348.8 � 221.0
in controls, were also observed.
Determination of cytokine protein content in media
conditioned by spleen cells from mice infected with
A. fumigatus conidia revealed an increase in the spontane-
ous production of IL-17 at days 3 and 7 p.i. and increased
IFN-� production at all time points (Fig. 4A). Reduced
spontaneous production of IL-4 was found throughout the
period of examination. Calculation of pro-infl ammatory
cytokine versus IL-4 content demonstrated high IL-17/
Fig. 1 Time course of fungal burden in spleen, lungs and kidneys of animals
inoculated with Aspergillus fumigatus conidia. Fungal burdens were
determined by a colony forming unit (CFU) assay in homogenates of spleen
(A), lung (B) and kidney (C) tissue of four mice per time point. Results are
expressed as average values of CFU from triplicate determinations.
Signifi cance at **P � 0.01 and ***P � 0.001) from day 1 postinfection.
© 2010 ISHAM, Medical Mycology, 48, 735–743

Proinfl ammatory response in systemic murine aspergillosis 739
Med
Myc
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
02/
03/1
1Fo
r pe
rson
al u
se o
nly.
IL-4 ratios of 96.0 � 90 (P � 0.05) or 17.0 � 6.0
(P � 0.05) at days 3 and 7, respectively, as compared to 6.0 � 2.0 in controls, as well as IFN-�/IL-4 at all time
points, i.e., 338.1 � 361.4 (P � 0.05), 190.8 � 229.1 (P
� 0.05) and 151.2 � 74.7 (P � 0.01) versus 10.0 � 7.0
in controls.
Restimulation of splenocytes from mice infected with A. fumigatus conidia resulted in increased production of pro-
infl ammatory cytokines, with the highest stimulation index
for IFN-� at the beginning of infection (compared to later
time points), and at later time points for IL-17. Interest-
ingly, some responsiveness to secondary stimulation with
the conidia was noted regarding IL-4 production, although
without difference among time points p.i. (Fig. 4B).
Pulmonary cell activity
No changes in relative lung mass were noted between con-
trol and infected animals (not shown). Application of
A. fumigatus conidia resulted in histologically evident pul-
monary infl ammation expressed as perivascular leukocyte
infi ltration, with vascular changes including vessel wall
edema and narrowing of vascular lumen (Fig. 5A).
Increased lung cell recovery was noted at all time points
p.i. (Fig. 5B), mainly due to lymphocytes, and possibly
neutrophils at day 7, when a tendency to increase
© 2010 ISHAM, Medical Mycology, 48, 735–743
(P � 0.07) was noted. On day 15 p.i. a decrease in numbers
of recovered monocytes/macrophages and neutrophils was
found, whereas numbers of lymphocytes still remained
elevated. Increased activation (spontaneous NBT reduction
capacity), as well as PMA-induced reduction of NBT were
Fig. 2 Basic parameters of antifungal spleen response after inoculation of mice with Aspergillus fumigatus conidia. Spleens were isolated from infected
mice or from control uninfected mice (control) at indicated time points p.i. (A) Relative spleen mass. (B) Number of cells isolated from the spleen. (C)
Spontaneous (ex vivo) proliferation of spleen cells. (D) Spleen cell proliferation after restimulation with Aspergillus fumigatus conidia in vitro. Results
were pooled from at least three experiments with 4–6 mice per group and expressed as mean values � SD. Signifi cance at *P � 0.05, **P � 0.01 and
***P � 0.001 vs control animals.
Fig. 3 Cytokine gene expression kinetics in spleen cells after inoculation
of mice with Aspergillus fumigatus conidia. Cell RNA from spleens of
infected or control mice was extracted, reverse transcribed and a relative
expression of IL-17, IFN-� and IL-4 genes was analyzed usig real-time
(RT)-PCR. The expression was normalized to β-actin to account for
differences in total RNA content of each sample. Data are presented as
mean � SD of samples pooled from three independent experiments with
three to four animals per group for each time-point and expressed as
percentages of mRNA expression in spleen cells of infected mice relative
to cells from control noninfected mice. Signifi cance at *P � 0.05 vs control
animals.

© 2010 ISHAM, Medical Mycology, 48, 735–743
740 Mirkov et al.
Med
Myc
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
02/
03/1
1Fo
r pe
rson
al u
se o
nly.
noted in cultures of lung cells of mice exposed to A. fumig-atus conidia at almost all time points examined (except day
3 p.i. for spontaneous activation) (Fig. 5C). A signifi cant
correlation was noted between the stimulated lung cell
activity and decrease in conidial burden (r � −0.82; P �
0.05; y � 3.57 − 15.6*x).
Examination of cytokine production by lung cells
revealed no changes in spontaneous cytokine production.
However, increased proinfl ammatory cytokine production
in response to restimulation with A. fumigatus conidia was
found in cultures of lung cells at day 15 p.i (IL-17) and at
all time points (IFN-�) (Fig. 6A & B). Neither spontaneous
nor stimulated IL-4 production by lung cells was noted
throughout the period of examination, remanining at low
levels, ranging from 10–20 pg/ml (data not shown).
Increased ratio of stimulated IFN-� to IL-4 production was
noted at day 7 (217.4 � 210.7, P � 0.01) and day 15 (271.1
� 417.6, P � 0.01), compared to the ratio calculated in
controls (32.2 � 25.6), while numerically increased IL-17/
IL-4 ratio of stimulated cytokine production was observed
only on day 15 p.i. (119.2 � 96.9 vs 79.9 � 20.5 in con-
trols; P � 0.05).
Discussion
Spleen and lung cell activity in systemic nonlethal
A. fumigatus infection were evaluated in this study. Both
Fig. 4 Production of cytokines by spleen cells after inoculation of mice
with Aspergillus fumigatus conidia. Cells from spleens of infected or control
mice (5 � 106/well) were cultured for 72 h in medium only (spontaneous
production) (A) or with heat-inactivated conidia of Aspergillus fumigatus
(restimulated production) (B). The production of IL-17, IFN-� and IL-4 was
measured in cell culture supernatants by using ELISA. Data are presented
as mean � SD of samples pooled from three independent experiments
with four to six animals per group for each time point. Results are
expressed as cytokine protein concentration (pg/ml) (A) or index of
stimulation (restimulated/spontaneous cytokine production) in infected
mice (B). Signifi cance at *P � 0.05 and **P � 0.01 vs control animals (A)
or vs stimulation index at day 3 postinfection (IFN-� B).
Fig. 5 Pulmonary infl ammation in mice after inoculation with Aspergillus fumigatus conidia. Lungs were isolated from immunized mice or from
control uninfected mice (control) at indicated time points. (A) Histological
appearance of lungs of infected mice by day 7 p.i. Multifocal perivascular
leukocyte infi ltration. Blood vessel wall edema and narrowing of vascular
lumen (insert). (B,C) Activity of leukocytes obtained by collagenase/
DNase digestion from lungs of infected mice or control mice. Total and
differential number of recovered leukocytes (B). Spontaneous and PMA-
induced respiratory burst of lung leukocytes evaluated by quantitative
NBT reduction assay (C). Signifi cance at *P � 0.05, **P � 0.01 and
***P � 0.001 vs control animals.

Proinfl ammatory response in systemic murine aspergillosis 741
Med
Myc
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
02/
03/1
1Fo
r pe
rson
al u
se o
nly.
spleen cell and stimulated lung cell activity correlated sig-
nifi cantly with a decrease in tissue CFUs, suggesting the
importance of these cells in the elimination of fungi.
Increased spleen cellularity may have accounted for the
increase in relative spleen mass following infection, and
may indicate splenocyte proliferation ex vivo, as judged by
the correlation between relational parameters. However,
the contribution of the infl ux of leukocytes into the spleen
to the increased spleen mass/cellularity ratio could not be
ruled out, since the spleen is a site of intense leukocyte
migration in the setting of infections [23].
Exposure to A. fumigatus conidia resulted in increased
expression of pro-infl ammatory (IL-17 and IFN-�) cytok-
ines by splenocytes. Although splenic IFN-� and IL-4 pro-
duction in systemic (lethal) A. fumigatus infection have
been already reported [13], our data are novel concerning
the IL-17 expression in systemic nonlethal A. fumigatus
infection. Increased gene expression, as well as production
of this cytokine nearly throughout the infection indicates
the relevance of this cytokine in systemic nonlethal Asper-gillus infection. Difference in the dynamics of expression,
as well as in the magnitude of the increase of IL-17 message
(2–3 times versus control) compared to the modest 30% for
IFN-� may potentially imply differential importance of
these two cytokines in systemic aspergillosis. However,
increased production of IFN-� by spleen cells in culture
indicates differential regulation of these two cytokines dur-
ing Aspergillus infection. In this regard, recent data demon-
strated importance of IL-17 production by draining lymph
node cells under conditions of relative IFN-� defi ciency in
murine pulmonary Aspergillus infections [18]. A lack of an
increase in IL-17 protein expression at day 15 may have
been related to regulation of its production, as it is subject
to tight control [19]. Splenic IL-17 production occurs mainly
in CD4+ T cells, as demonstrated in host response to pul-
monary Aspergillus infection [18]. However, CD8+ cells, as
well as NK cells, have shown a capacity to produce this
cytokine under certain conditions [24], and thus, may have
contributed to the total amount of the cytokine produced.
Recent data have demonstrated the importance of IL-17
in lethal systemic infections caused by Cryptococcus
neoformans [25] and Candida albicans [26] in mice. Our
results also showed the relevance of IL-17 in hosts that have
successfully recovered from Aspergillus infection. Various
IL-17-related effector activities in pathogen killing [27], as
well as the T-cell activating properties of IL-17 suggested in
experimental hypersensitivity in mice [28], might be rele-
vant for its antifungal activity in the spleen.
Supplementary to the gene expression data, increased
spontaneous IFN-� production demonstrated the relevance
of this cytokine in the spleen in a systemic nonlethal model
of infection with A. fumigatus. Increased IFN-� protein
content, compared to negligible mRNA expression, might
© 2010 ISHAM, Medical Mycology, 48, 735–743
have resulted in the 72-h conditioned medium from various
cell-derived products, such as IL-12 or nitric oxide with
IFN-�-stimulating potential. Responsiveness of spleen
cells to secondary stimulation with A. fumigatus conidia
observed at time points when no increase in IFN-� mRNA
supports such an assumption. Both CD4+ [13] and CD8+ T
cells, as well as NK cells, are well known sources of IFN-�
in settings of various microbial infections [29] and may be
a source of splenic IFN-� during A. fumigatus infection.
Much higher spontaneous (throughout the p.i. period
examined), as well as conidia-restimulated production
(particularly at day 3 p.i.) of IFN-� compared to IL-17 by
spleen cells implies stringent requirements for IFN-� in
nonlethal A. fumigatus infections. In accord with this
assumption are our preliminary data in which signifi cant
IFN-� production (compared to IL-17) was noted in
infected BALB/c mice (known as low IFN-� producers in
lethal parasitic and some bacterial infections; data not
shown). Such considerations are in line with data that dem-
onstrated higher ConA-stimulated IFN-� production by
spleen cells from mice that survived systemic lethal A. fumigatus infection as compared to non-survivors [13].
Protective effects of this cytokine in lethal A. fumigatus
infection in mice [14] might be of relevance for antifungal
activity of the spleen demonstrated in this study. IFN-� is
a known stimulator of antifungal activity by phagocytes
[30–32] and activated splenic macrophages were shown to
be responsible for initial defense against invasive aspergil-
losis by their phagocytosis of conidia and conidiacidal
activity [33].
A decrease in IL-4 expression noted in our study is in
accord with data from a study of systemic lethal murine
aspergillosis which showed lower IL-4 expression by
spleens of survivors as compared to increased expression
of this cytokine by spleens of non-survivors [13]. Decreased
IL-4 expression might be benefi cial in nonlethal A. fumig-atus infection as IL-4 has been demonstrated to enhance
susceptibility to invasive pulmonary aspergillosis [34].
Although decreased gene transcription, as well as protein
expression of IL-4 was noted in the spontaneous response,
an increased production was observed in the restimulation
response. This could be explained by the demonstrated
stimulation of IL-4 response with heat-inactivated A. fumigatus conidia [35], which were used in restimulation
experiments in our study.
Progressive elimination of A. fumigatus conidia from
lungs of infected mice was associated with pulmonary
infl ammation. Similar to the pulmonary response to conidia
that have gained access to lungs by the respiratory route
[8,36], we noted increased activation of lung cells by conidia
deposited in the lungs following systemic infection.
In contrast to response in the spleen, IL-17 in the lungs
is produced in recall response only during the later course
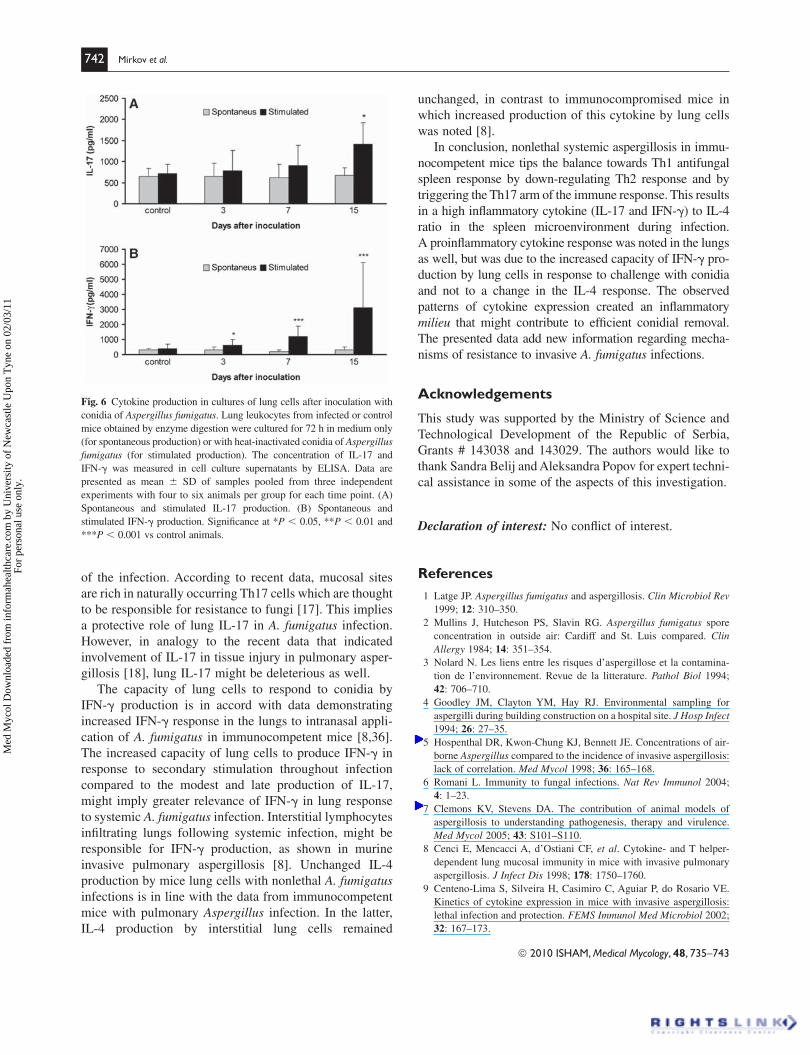
742 Mirkov et al.
Med
Myc
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
02/
03/1
1Fo
r pe
rson
al u
se o
nly.
of the infection. According to recent data, mucosal sites
are rich in naturally occurring Th17 cells which are thought
to be responsible for resistance to fungi [17]. This implies
a protective role of lung IL-17 in A. fumigatus infection.
However, in analogy to the recent data that indicated
involvement of IL-17 in tissue injury in pulmonary asper-
gillosis [18], lung IL-17 might be deleterious as well.
The capacity of lung cells to respond to conidia by
IFN-� production is in accord with data demonstrating
increased IFN-� response in the lungs to intranasal appli-
cation of A. fumigatus in immunocompetent mice [8,36].
The increased capacity of lung cells to produce IFN-� in
response to secondary stimulation throughout infection
compared to the modest and late production of IL-17,
might imply greater relevance of IFN-� in lung response
to systemic A. fumigatus infection. Interstitial lymphocytes
infi ltrating lungs following systemic infection, might be
responsible for IFN-� production, as shown in murine
invasive pulmonary aspergillosis [8]. Unchanged IL-4
production by mice lung cells with nonlethal A. fumigatus
infections is in line with the data from immunocompetent
mice with pulmonary Aspergillus infection. In the latter,
IL-4 production by interstitial lung cells remained
unchanged, in contrast to immunocompromised mice in
which increased production of this cytokine by lung cells
was noted [8].
In conclusion, nonlethal systemic aspergillosis in immu-
nocompetent mice tips the balance towards Th1 antifungal
spleen response by down-regulating Th2 response and by
triggering the Th17 arm of the immune response. This results
in a high infl ammatory cytokine (IL-17 and IFN-�) to IL-4
ratio in the spleen microenvironment during infection.
A proinfl ammatory cytokine response was noted in the lungs
as well, but was due to the increased capacity of IFN-� pro-
duction by lung cells in response to challenge with conidia
and not to a change in the IL-4 response. The observed
patterns of cytokine expression created an infl ammatory
milieu that might contribute to effi cient conidial removal.
The presented data add new information regarding mecha-
nisms of resistance to invasive A. fumigatus infections.
Acknowledgements
This study was supported by the Ministry of Science and
Technological Development of the Republic of Serbia,
Grants # 143038 and 143029. The authors would like to
thank Sandra Belij and Aleksandra Popov for expert techni-
cal assistance in some of the aspects of this investigation.
Declaration of interest: No confl ict of interest.
References
Latge JP. 1 Aspergillus fumigatus and aspergillosis. Clin Microbiol Rev
1999; 12: 310–350.
Mullins J, Hutcheson PS, Slavin RG. 2 Aspergillus fumigatus spore
concentration in outside air: Cardiff and St. Luis compared. Clin Allergy 1984; 14: 351–354.
Nolard N. Les liens entre les risques d’aspergillose et la contamina-3
tion de l’environnement. Revue de la litterature. Pathol Biol 1994;
42: 706–710.
Goodley JM, Clayton YM, Hay RJ. Environmental sampling for 4
aspergilli during building construction on a hospital site. J Hosp Infect 1994; 26: 27–35.
Hospenthal DR, Kwon-Chung KJ, Bennett JE. Concentrations of air-5
borne Aspergillus compared to the incidence of invasive aspergillosis:
lack of correlation. Med Mycol 1998; 36: 165–168.
Romani L. Immunity to fungal infections. 6 Nat Rev Immunol 2004;
4: 1–23.
Clemons KV, Stevens DA. The contribution of animal models of 7
aspergillosis to understanding pathogenesis, therapy and virulence.
Med Mycol 2005; 43: S101–S110.
Cenci E, Mencacci A, d’Ostiani CF, 8 et al. Cytokine- and T helper-
dependent lung mucosal immunity in mice with invasive pulmonary
aspergillosis. J Infect Dis 1998; 178: 1750–1760.
Centeno-Lima S, Silveira H, Casimiro C, Aguiar P, do Rosario VE. 9
Kinetics of cytokine expression in mice with invasive aspergillosis:
lethal infection and protection. FEMS Immunol Med Microbiol 2002;
32: 167–173.
Fig. 6 Cytokine production in cultures of lung cells after inoculation with
conidia of Aspergillus fumigatus. Lung leukocytes from infected or control
mice obtained by enzyme digestion were cultured for 72 h in medium only
(for spontaneous production) or with heat-inactivated conidia of Aspergillus fumigatus (for stimulated production). The concentration of IL-17 and
IFN-� was measured in cell culture supernatants by ELISA. Data are
presented as mean � SD of samples pooled from three independent
experiments with four to six animals per group for each time point. (A)
Spontaneous and stimulated IL-17 production. (B) Spontaneous and
stimulated IFN-� production. Signifi cance at *P � 0.05, **P � 0.01 and
***P � 0.001 vs control animals.
© 2010 ISHAM, Medical Mycology, 48, 735–743

Proinfl ammatory response in systemic murine aspergillosis 743
Med
Myc
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
02/
03/1
1Fo
r pe
rson
al u
se o
nly.
Schmidt A. Animal models of aspergillosis – also useful for vaccina-10
tion strategies? Mycoses 2002; 45: 38–40.
Polak A. Experimental models in antifungal chemotherapy. 11 Mycoses
1998; 41: 1–30.
Mirkov I, Zolotarevski L, Glamoclija J, Kataranovski D, Kataranovski 12
M. Experimental disseminated aspergillosis in mice: histopathologi-
cal study. J Med Mycol 2008; 18: 75–82.
Cenci E, Perito S, Enssle KH, 13 et al. Th1 and Th2 cytokines in mice
with invasive aspergillosis. Infect Immun 1997; 65: 564–570.
Nagai H, Guo J, Choi HY, Kurup V. Interferon-14 � and tumor necrosis
factor-α protect mice from invasive aspergillosis. J Infect Dis 1995;
172: 1554–1560.
Clemons KV, Grunig G, Sobel RA, 15 et al. Role of IL-10 in invasive
aspergillosis: increased resistance of IL-10 gene knockout mice to le-
thal systemic aspergillosis. Clin Exp Immunol 2000; 122: 186–191.
Palm NW, Medzhitov R. Antifungal defense turns 17. 16 Nat Immunol 2007; 8: 549–551.
Zelante T, De Luca A, D’Angelo C, Moretti S, Romani L. IL-17/17
Th17 in anti-fungal immunity: what’s new? Eur J Immunol 2009;
39: 645–648.
Zelante T, De Luca A, Bonifazi P, 18 et al. IL-23 and the Th17 pathway
promote infl ammation and impair antifungal immune resistance. Eur J Immunol 2007; 37: 2695–2706.
Zelante T, Bozza S, De Luca A, 19 et al. Th17 cells in the setting of Asper-gillus infection and pathology. Med Mycol 2009; 47: S162–S169.
Booth C. Fungal culture media. In: Norris JR, Ribbons DW (eds). 20
Methods in Microbiology, 4th edn. London & New York: Academic
Press, 1971: 49–94.
Sheppard DC, Marr KA, Fredricks DN, 21 et al. Comparison of three
methodologies for the determination of pulmonary fungal burden
in experimental murine aspergillosis. Clin Microbiol Infect 2006;
12: 376–380.
Monboisse JC, Garnotel R, Randoux A, Dufer J, Borel JP. Adhesion 22
of human neutrophils to and activation by type-I collagen involving a
β2 integrin. J Leukoc Biol 1991; 50: 373–380.
Mebius RE, Kraal G. Structure and function of the spleen. 23 Nat Rev Immunol 2005; 5: 606–616.
Weaver CT, Hatton RD, Mangan PR, Harrington LE. IL-17 family of 24
cytokines and the expanding diversity of effector T cell lineages. Annu Rev Immunol 2007; 25: 821–852.
© 2010 ISHAM, Medical Mycology, 48, 735–743
Kleinschek MA, Muller U, Brodie S, 25 et al. IL-23 enhances the in-
fl ammatory cell response in Cryptococcus neoformans infection and
induces a cytokine pattern distinct from IL-12. J Immunol 2006;
176: 1098–1106.
Huang W, Na L, Fidel PL, Schwarzenberger P. Requirement of inter-26
leukin-17A for systemic anti-Candida albicans host defense in mice.
J Infect Dis 2004; 190: 624–631.
Matsuzaki G, Umemura M. Interleukin-17 as an effector molecule of 27
innate and acquired immunity against infections. Microbiol Immunol 2007; 51: 1139–1147.
Nakae S, Komiyama Y, Nambu A, 28 et al. Antigen-specifi c T cell
sensitization is impaired in IL-17-defi cient mice, causing suppres-
sion of allergic cellular and humoral responses. Immunity 2002;
17: 375–387.
Abbas AK, Lichtman AH, Pillai S. Cytokines. In: Schmitt W, Gruliow 29
R (eds). Cellular and Molecular Immunology, 6th edn. Philadelphia:
Saunders Elsevier, 2007: 267–303.
Bernhisel-Broadbent J, Camargo EE, Jaffe HS, Lederman HM. Re-30
combinant human interferon-gamma as adjunct therapy for Aspergil-lus infection in a patient with chronic granulomatous disease. J Infect Dis 1991; 163: 908–911.
Rex JH, Bennett JE, Gallin JI, 31 et al. In vivo interferon-� therapy aug-
ments the in vitro ability of chronic granulomatous disease neutrophils
to damage Aspergillus hyphae. J Infect Dis 1991; 163: 849–852.
Roilides E, Uhlig K, Venzon D, Pizzo PA, Walsh TJ. Enhancement 32
of oxidative response and damage caused by human neutrophils to
Aspergillus fumigatus hyphae by granulocyte colony-stimulating fac-
tor and �-interferon. Infect Immun 1993; 61: 1185–1193.
de Repentigny L, Petitbois S, Boushira M, 33 et al. Acquired immunity in
experimental murine aspergillosis is mediated by macrophages. Infect Immun 1993; 61: 3791–3802.
Cenci E, Mencacci A, Del Sero G, 34 et al. Interleukin-4 causes sus-
ceptibility to invasive pulmonary aspergillosis through suppression of
protective type I responses. J Infect Dis 1999; 180: 1957–1968.
Rivera A, Van Epps HL, Hohl TM, Rizzuto G, Pamer EG. Distinct 35
CD4+-T-cell responses to live and heat-inactivated Aspergillus fumig-atus conidia. Infect Immun 2005; 73: 7170–7179.
Brieland JK, Jackson C, Menzel F, 36 et al. Cytokine networking in lungs
of immunocompetent mice in response to inhaled Aspergillus fumiga-tus. Infect Immun 2001; 69: 1554–1560.
This paper was fi rst published online on Early Online on 25 January 2010.
Related Documents











