E-00226-2002.R2 1 Specificity of Thyroid Hormone Receptor Subtype and Steroid Receptor Coactivator (SRC)-1 on Thyroid Hormone Action Peter M. Sadow 1, 2 , Olivier Chassande 3 , Karine Gauthier 3 , Jacques Samarut 3 , Jianming Xu 4 , Bert W. O’Malley 4 , and Roy E. Weiss 1,5 1 Depts. of Medicine and 2 Pathology, The University of Chicago, Chicago, IL 60637; 3 Laboratoire de Biologie Moléculaire et Cellulaire de l’Ecole Normale Supérieure de Lyon, Lyon, France; 4 Department of Molecular and Cellular Biology, Baylor College of Medicine, Houston, TX 77030. 5 Corresponding Author: Roy E. Weiss, MD, PhD Thyroid Study Unit, MC 3090 University of Chicago 5841 S. Maryland Ave Chicago, IL USA Email: [email protected] Telephone: 773-702-9266 Fax: 773-834-3966 Running Title: SRC-1 and TR Specificity in peripheral tissues Copyright 2002 by the American Physiological Society. AJP-Endo Articles in PresS. Published on September 17, 2002 as DOI 10.1152/ajpendo.00226.2002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

E-00226-2002.R2
1
Specificity of Thyroid Hormone Receptor Subtype and Steroid Receptor
Coactivator (SRC)-1 on Thyroid Hormone Action
Peter M. Sadow1, 2, Olivier Chassande3, Karine Gauthier3, Jacques Samarut3, Jianming Xu4, Bert
W. O’Malley4, and Roy E. Weiss1,5
1Depts. of Medicine and 2Pathology, The University of Chicago, Chicago, IL 60637;
3Laboratoire de Biologie Moléculaire et Cellulaire de l’Ecole Normale Supérieure de Lyon,
Lyon, France; 4Department of Molecular and Cellular Biology, Baylor College of Medicine,
Houston, TX 77030.
5Corresponding Author:
Roy E. Weiss, MD, PhDThyroid Study Unit, MC 3090University of Chicago5841 S. Maryland AveChicago, IL USAEmail: [email protected]: 773-702-9266Fax: 773-834-3966
Running Title: SRC-1 and TR Specificity in peripheral tissues
Copyright 2002 by the American Physiological Society.
AJP-Endo Articles in PresS. Published on September 17, 2002 as DOI 10.1152/ajpendo.00226.2002

E-00226-2002.R2
2
Abstract
Isoforms of the thyroid hormone receptor (TR) and TR genes mediate thyroid
hormone action. How TR isoforms modulate tissue-specific thyroid hormone (TH) action
remains largely unknown. The steroid receptor coactivator-1 (SRC-1) is among a group of
transcriptional coactivator proteins that bind to TRs, along with other members of the nuclear
receptor superfamily, and modulates the activity of genes regulated by TH. Mice deficient in
SRC-1 possess decreased tissue responsiveness to TH and many steroid hormones; however, it is
not known whether or not SRC-1-mediated activation of TH-regulated gene transcription in
peripheral tissues, such as heart and liver, is TR isoform specific. We have generated mice
deficient in TRα and SRC-1 (TRα0/0SRC-1-/-), as well as in TRβ and SRC-1 (TRβ-/-SRC-1-/-) and
investigated thyroid function tests and effects of TH deprivation and TH treatment compared to
wild type (WT) mice or those deficient in either TR or SRC-1 alone. The data show that (1) in
the absence of TRα or TRβ, SRC-1 is important for normal growth; (2) SRC-1 modulates TRα
and TRβ effects on heart rate; (3) two new TRβ-dependent markers of TH action in the liver
have been identified, osteopontin (up-regulated) and glutathione-S-transferase (down-regulated);
and (4) SRC-1 may mediate the hypersensitivity to TH seen in liver of TRα0/0 mice.
Keywords: knockout/ resistance to thyroid hormone/ SRC-1/ thyrotropin/ thyroid hormone
receptor

E-00226-2002.R2
3
Introduction
Thyroid hormone receptor (TR) α and TRβ function as nuclear transcription factors that
mediate thyroid hormone (TH) action. TRs bind to TH response elements (TREs) on thyroid
responsive genes in association with transcriptional coregulators. Corepressors form part of a
transcriptional complex and recruit histone deacetylases, reducing transcription (18, 40). The
conformational change of TR produced by TH binding releases the corepressor and permits the
recruitment and binding of a coactivator. The coactivator has a number of functions that may
include intrinsic histone acetyltransferase activity, recruitment of histone acetyltransferases and
recruitment of additional transcription factors and RNA polymerase (21, 23). Several classes of
nuclear coactivators have been described that are important in mediating the response of
mammalian cells to thyroid as well as steroid and retinoid hormones (2, 7, 14, 17, 41, 43) among
which is the steroid receptor coactivator (SRC)-1, a member of the p160 family of coactivators
(26).
Since some genes are up-regulated and others are down-regulated by TH in same cell,
various theories have been proposed to explain the mechanism(s) of TR- modulated gene
expression (see (44) for review). We propose that specificity of interaction among TR subtypes
with particular cofactors may influence whether there is stimulation or inhibition of mRNA
expression. Determination of the nature of interaction of TR subtypes with specific cofactors,
and how it affects gene transcription can be evaluated in vivo by using animals that are lacking
each of the two TR genes with or without SRC-1. It has been shown that TRβ knockout mice
(TRβ-/-) have resistance to thyroid hormone (RTH) (10-12, 22, 34, 37), while mice with
disruption of the TRα1 and 2 isoforms (TRα0/0) are hypersensitive to TH in several of the tissues
examined (22) or less prone to the effects of TH deprivation (24). On the other hand, mice

E-00226-2002.R2
4
completely deficient in both TRβ and TRα exhibit more severe resistance to TH than those
lacking TRβ only (16). Taken together, these data suggest that both isoforms play selective and
overlapping roles, both centrally and peripherally. Furthermore, coactivators are important in
TR-mediated TH action in vivo as demonstrated by a mouse model with disruption of the SRC-1
gene (SRC-1-/-) which has also been shown to produce a phenotype of reduced hormone
sensitivity (37, 42). Therefore we ask whether SRC-1 differentially modulates the functions of
TRα and TRβ, and if so, how does this effect influence TH action in the liver and heart?
For this purpose we generated mice deficient in TRα and SRC-1 (TRα0/0SRC-1-/-), as
well as mice deficient in the TRβ and SRC-1 (TRβ-/-SRC-1-/-) and compared these to wildtype
(WT) mice or mice with deficiency in TRβ, TRα and SRC-1 alone. Our data provide evidence
that in the absence of TRα or TRβ, SRC-1 is important for normal growth. We also show that
SRC-1 mediates TRα and TRβ action in the heart. This study identifies two novel TRβ-
dependent markers of TH action in the liver, osteopontin (up-regulated) and glutathione-S-
transferase (down-regulated). We have also demonstrated that SRC-1 may mediate the
hypersensitivity to TH seen in TRα0/0 mice.

E-00226-2002.R2
5
Materials and Methods
Generation and handling of animals
Mice deficient in SRC-1 (SRC-1-/-) were generated as reported previously (42). A
targeting vector disrupted the SRC-1 gene in 129sv ES cells by inserting an in-frame stop codon
at the Met-381 position and deleting approximately 9 kb of downstream genomic sequence that
contains 446 amino acids from Met-381 to Thr-826. This eliminated all functional domains for
transcriptional activation, histone acetyltransferase activity and interactions with nuclear
receptors, CBP, p300, and p/CAF (42). The SRC-1-/- construct was maintained in a C57BL/6J
mouse strain. The genotype of mice was confirmed by analysis of tail DNA as previously
described (42).
TRβ-/- mice were produced as previously described (12). These mice were produced by
insertion of the LacZ-NeoR cassette downstream to the splice site in exon 4, eliminating the
expression of the DNA and ligand binding domains of TRβ1 and TRβ2 (12). The TRα0/0 mice
were produced by insertion of the LacZ-NeoR cassette downstream from exon 3 and replacing
exons 5 through 7. This effectively abolished not only the generation of TRα1 and
TRα2 transcripts, but also that of TR∆α1 or TR∆α2 by removing a transcription start site in
intron 7 (13). The gene sequence for rev-erbA α protein encoded by the opposite strands for the
TRα (31) remains intact. In both sets of mice, the recombinant ES cells were derived from
129sv mice and were implanted into C57BL/6 recipient blastocysts. C57BL/6 mice were mated
to each chimeric mouse and then backcrossed at 3-9 times into the same strain, diluting the
129sv background.

E-00226-2002.R2
6
The individual knockout mice were backcrossed more than nine times on a C57BL/6
background to produce a uniform genetic background for wildtype and knockout animals. We
then crossed the SRC-1-/- mice with TRα0/0 and TRβ-/- mice to produce double heterozygous
animals, 8 times. The double heterozygous mice were mated to produced double homozygous
mice which were backcrossed 3-5 times to each other.
Mice were weaned on the 4th week after birth and were fed Purina Rodent Chow (0.8
ppm Iodine) ad libitum and tap water. They were housed, 5 or less mice per cage, in an
environment of controlled 19˚C temperature and 12 h alternating darkness and artificial light
cycles. All animal experiments were performed according to protocols approved by the
Institutional Animal Care and Use Committee at the University of Chicago.
Mice were 40-70 days old at the time of sacrifice. At various intervals, approximately
300 µl of blood were obtained by tail vein under light methoxyflurane (Pitman Moore,
Mundelein, IL) anesthesia. Experiments were terminated by exsanguination via retroorbital vein.
Whole blood was allowed to clot overnight at 4˚C and serum was collected following
centrifugation and stored at minus 20˚C until analyzed.
Induction of hypothyroidism and treatment with TH
TH deficiency was induced in male mice by feeding with low iodine (LoI) diet
supplemented with 0.15% propylthiouracil (PTU, Harlan Teklad, Madison, WI). On the 10th
day, one group of mice (>5 mice/genotype) was injected daily for 4 days with vehicle only (1X
PBS, controls) while another received 0.2 µg of L-T3/25g body wt/day, maintained on the
LoI/PTU diet. Fourteen to 16 h after the final injection, experiments were terminated by
exsanguination. L-T3, dissolved in phosphate buffered saline and 0.002% human serum albumin
as a vehicle, was given by intraperitoneal injection in a total volume of 0.1-0.3 ml. A stock of L-
T3 (Sigma, St. Louis, MO) at a concentration of 1 mg/ml was prepared in a solution of 50%
ethanol/50% 1X PBS containing 5 mM NaOH and kept at –20˚ C, protected from light.
Concentration of L-T3 was confirmed by RIA (Diagnostic Products, Los Angeles, CA). Blood

E-00226-2002.R2
7
samples were obtained at baseline, on the 10th day after the initiation of the LoI/PTU diet and at
the termination of the experiment on day 15.
The dose of L-T3 given to thyroid hormone deficient animals was derived from previous
experiments. It was optimized to achieve a partial suppression of serum TSH in order to make
evident the differences between wildtype and SRC-1-/- mice (22, 34, 38). This allowed us to
examine the effect of deficiency of receptor and coactivator under identical conditions of TH
supply. Metabolism of T3 was determined in each genotype by measuring serum T3 levels at 2,
4, 8, and 16 h following injection of L-T3 (22).
TH and TSH concentrations in serum
Serum TSH was measured in 50 µl of serum using a sensitive, heterologous,
disequilibrium double antibody precipitation radioimmunoassay as previously described (28).
Samples containing more than 200 mU TSH/L were 5- and 50-fold diluted with TSH-deficient
mouse serum.
Serum T4 and total T3 concentrations were measured by a double antibody precipitation
RIA (Diagnostic Products, Los Angeles, CA) using 25 and 50 µl of serum, respectively. The
sensitivities of these assays were 0.2 µg T4/dL and 5 ng T3/dL. The inter-assay coefficients of
variation were 5.4, 4.2 and 3.6% at 3.8, 9.4 and 13.7 µg/dl for T4 and 7.7, 7.1 and 6.2% at 32, 53
and 110 ng/dl for T3.
Serum Leptin
Samples were taken from frozen sera (obtained by retroorbital bleed and stored at –80˚C)
and leptin levels were determined by RIA (LINCO Research, St. Charles, MO) with 10 µL of
serum diluted into 100 µL in duplicate. Results are reported as serum leptin in ng/mL.

E-00226-2002.R2
8
Measurements of Growth, Heart Rate and Energy Expenditure
Mice were weighed on 200 gm balance on the first day of each week from age 2-9 weeks.
In addition, length of each mouse was determined from the tip of the nose to the base of the tail
under light gas anesthesia at the time of weighing. Heart rates of mice were determined at
baseline, following 14 days of a LoI/PTU diet, and on a PTU diet following 14-16 hours after the
fourth daily IP injection of T3 (0.2µg/25g). Animals were anesthetized with chloral hydrate (4
mg/10 g body weight, ip) and heart rate was determined using a Hewlett Packard
Monitor/Terminal Model 78534AA with a chart speed of 25 mm/second. Body temperature was
maintained by keeping mice on a heating pad during measurement. Energy expenditure (EE)
was determined at baseline by measurement of change in body weight and food consumption
over 4 days as previously described (5, 37). EE was calculated according to the formula:
EE(Kcal/day)=(food consumption (g/day) x 4.058* x 0.8**) ± (weight change (g/day) x 7***)
Above, * is the caloric value of the food (in Kcal/g); ** is adjustment for 20% food
wasted in litter as determined by bomb calorimetry in euthyroid wildtype mice; *** is caloric
value of 1 g body weight change. Loss of weight is added and weight gain subtracted.
Isolation of Liver mRNA
Livers from animals were immediately frozen on dry ice and stored at -80˚C. For RNA
extraction, approximately 80 mg of liver tissue from individual mice were homogenized in 1 mL
of TRIzol (Life Technologies, Rockville, MD) with a Polytron tissue homogenizer. Total RNA
was extracted according to the protocol provided with the TRIzol reagent. Concentration (A260)
of the total RNA was determined and RNA was stored in 1/10 volume 3M NaOAc and 3
volumes of 100% ethanol at -80˚C.
Microarray Analysis of Mouse Liver
Six mice (male, age 70 days) of three genotypes (WT, TRβ-/- or SRC-1-/-) were treated
with PTU and ip L-T3 as described above. Mice were killed on the 15th day and livers were

E-00226-2002.R2
9
removed and immediately frozen on dry ice (and stored at -80˚C). Livers from 3 mice were
pooled into one sample used to extract RNA and make cDNA. The cDNA was hybridized to
Clontech’s mouse Atlas Microarray V1.2 (1,172 genes) and resolved on a phosphorimager
performed by Clontech (Palo Alto, CA). Each microarray compared expression in TH-deprived
liver tissue to that in L-T3-treated mouse liver for each genotype. Using threshold ratios for
significance of 1.67 for each comparison (WT with and without L-T3, TRβ-/- with and without T3,
and SRC-1-/- with and without L-T3), genes were identified in each group that were either up- or
down-regulated in response to TH. In all 3 groups, 7.8-8.9% of the total 1,172 genes displayed a
potential response to TH. In WT and SRC-1-/- mice, the majority of responsive genes were up-
regulated by TH, while in the TRβ-/- mice, the majority of responsive genes were down-regulated
by TH (Table I).

E-00226-2002.R2
10
TaqMan RT PCR of Genes Expressed in Liver
In order to quantitate mRNA expression of various genes in livers of different genotypes,
2 µg of total RNA were reverse transcribed using the First-Strand Synthesis Superscript Kit (Life
Technologies, Rockville, MD) according to the provided protocol. Reverse transcription (RT)
was performed using random hexamers. cDNAs obtained from the RT reaction were diluted
with RNAse-free water to a concentration of 1 ng/µL. TaqMan fluorescent probe/primer sets
were designed using Primer Express 1.5 (Applied Biosystems, Foster City, CA) and mRNA
sequences taken from GenBank. Specificity was confirmed by BLAST search. Primer/probe
sets were then obtained for osteopontin, glutathione-S-transferase (GST), split hand/split foot
(SHSF), Ets-related transactivation factor (ERF), and 5’-deiodinase (MegaBases, Evanston, IL)
(Table II). Equal loading of wells was controlled using a commercially available probe/primer
set for 18S ribosomal RNA (Applied Biosystems, Foster City, CA). Detection of mRNA was
performed with Sequence Detector Software and the ABI 7000 Sequence Detection System
(Applied Biosystems, Foster City, CA), capable of reading two fluorophores simultaneously in
each well (sample probe and 18S ribosomal control probe).
10 ng of each reverse-transcribed cDNA sample were run in duplicate and the reaction
was performed with TaqMan Universal Mix and 96-well optical plates (Applied Biosystems,
Foster City, CA). Each duplicate sample represents reverse transcribed total RNA from
individual mouse liver samples. The threshold cycle (Ct) is the first cycle, in a 40-cycle reaction,
at which fluorescence is detected. For each sample, there are two recorded threshold cycles, the
first corresponding to amplification of 18S rRNA (VIC fluorophore), and the second to specific
gene of interest (FAM fluorophore). Normalization of data involved subtraction of rRNA Ct
from that of the specific gene being amplified per well, as amplification is logarithmic. For each
mouse genotype analyzed, at least five individual liver sample RNAs were run in duplicate. To
calculate results, the average Ct for wildtype mice was determined. Individual mouse liver data
was reported as fold increase or decrease from this wildtype average. Assays were repeated at
least 3 times and the data were normalized and merged.

E-00226-2002.R2
11
TaqMan quantitative RT PCR expression data for the four genes identified by microarray
analysis (osteopontin, glutathione S-transferase (GST), split hand split foot (SHSF), and Est-
related factor(ERF)) is reported as % change of PTU-treated mice from littermates on a PTU diet
treated with T3 (as described above). The mean fold change of PTU-treated animals of each
genotype was determined relative to wildtype mice on PTU. Percent change was calculated by
dividing each genotype’s T3-treated mean (calculated against WT PTU) by its own PTU mean (±
SE). Each group of animals (PTU and PTU+T3) had at least five mice.
Data presentation and statistics
Values are reported as mean ± SE. Initially a two-way or one-way ANOVA calculation
was done to determine if there was a significant interaction between treatment and genotype on
each of the parameters measured. If significant interaction was detected, the effect of treatment
was examined separately for each genotype and vice versa. The Tukey-Kramer method, at 5%
significance (Statview V 5.0, SAS Institute Inc.) was used to control for multiple comparisons.
To stabilize the variance of data for serum TSH and Leptin, these data only were analyzed on a
logarithmic scale.

E-00226-2002.R2
12
Results
Thyroid function tests
Thyroid function tests of different mouse genotypes at baseline, and following treatment with
PTU or PTU and ipT3 are shown in Table III. Experiments were performed in adult male mice
because of previously reported sex and age differences in TH levels in WT mice (5, 28). Serum total
T4 and T3 levels as well as TSH concentrations were significantly higher in TRβ-/- and SRC-1-/- as
compared to WT mice as previously reported (12, 34, 38). TRα0/0 mice demonstrated decreased
serum T4 with normal TSH compared to WT mice (p<0.005). In the combined TRβ-/-SRC-1-/- mice,
serum levels of T4, T3 and TSH were 1.7, 2.7 and 2.8 times greater, respectively, than in TRβ-/- or
2.7, 2.8 and 10 times greater, respectively, than in the SRC-1-/- mice. Mice deficient in both SRC-1
and TRα have serum T4 and T3 values that were 2.0 and 1.7 times greater than TRα0/0, respectively,
and TSH values were also increased by 2.3 times. Levels of free T3 and T4 showed similar
differences (35).
TH deprivation resulted in marked increases in serum TSH in all genotypes, although serum
TSH levels in TRα0/0 and TRα0/0SRC-1-/- mice did not increase as much as in WT, and TSH levels in
TRβ-/-SRC-1-/- mice increased 3-fold greater than WT (p<0.0001). T4 levels in all mice decreased to
<0.25 µg/dl following PTU/LoI diet. Treatment with TH resulted in variable suppression of absolute
serum TSH values depending on the genotype of the mouse. The TRβ-/-SRC-1-/- mice had the least
suppression, indicative of its highest degree of resistance to TH and TRα0/0 mice showed the highest
suppression of serum TSH; the latter did not reach statistical significance due to the large standard
deviation of WT mice.
Growth in mice of different genotypes
Length and body weight from age 2 to 9 weeks were measured in mice of different genotypes
(Fig. 1). SRC-1-/- mice grew at the same rate and achieved similar lengths at week 9 compared to
WT mice. At week 5, lengths of TRβ-/- and TRα0/0 mice were 14% less than WT mice (significance

E-00226-2002.R2
13
at the 5% confidence level) and remained from 5-10% smaller compared to WT mice at week 9.
Therefore, both TRα and TRβ are required for normal linear growth. TRβ is the major determinant
of overall body length, as in the absence of TRβ (with or without SRC-1) these animals have the
most stunted length (Fig. 1A). TRα0/0SRC-1-/- show more retarded linear growth than TRα0/0 mice
(Fig. 1B).
Weight gain remained similar for TRβ-/-, SRC-1-/- and WT mice through week 9. For animals
to achieve normal body weight, SRC-1 appears to be an important modifier in the absence of either
the TRα or TRβ. TRα0/0 mice have a 28% reduction in body weight at 3 weeks and by 9 weeks still
maintain a 17% reduction in body weight (Fig. 1D). In the absence of both SRC-1 and either the
TRα or TRβ, mice have greater reduction in growth. Specifically, TRβ-/-SRC-1-/- mice begin to have
a decline in body weight at 8 weeks (p<0.0001), a change not seen in the SRC-1-/- or TRβ-/- mice.
Energy Expenditure and Serum Leptin
Energy expenditure experiments were performed in mice 6-10 weeks of age (Table IV).
TRβ-/-SRC-1-/- mice have the highest energy expenditure, 1.4 times that of WT mice (significance at
the 5% confidence level). This, along with the high TH levels in these mice, imply the TH-
mediated energy expenditure occurs through TRα. This corresponds to the decrease in body weight
seen in the TRβ-/-SRC-1-/- mice (Fig. 1C). TRα0/0 tend to have lower energy expenditure, but it does
not reach significance. In the absence of SRC-1, TRβ and/or TRα also maintain normal energy
expenditure.
Serum leptin concentrations in nonfasting mice were 2.1 fold higher in SRC-1-/- mice
(p=.0472) and 2.5 fold higher in TRβ-/-SRC-1-/- mice (p=0.020) compared to WT (Table IV, Fig. 2).

E-00226-2002.R2
14
The higher leptin levels in these animals suggest that TH mediated increase in leptin occurs in the
absence of SRC-1 and TRβ.
The contrasting effect of TH and genotype in EE and leptin concentration is demonstrated in
Fig. 2. Note that EE did not increase very much despite significant increases in TH levels in the
different genotypes, except for the combined TRβ-/-SRC-1-/- mice. On the other hand, regulation of
serum leptin levels are less dependent on genotype, but more dependent on TH levels, where the
increase in TH is relatively independent of the genotype.
Heart Rate
Heart rates were measured in mice of different genotypes at baseline and after 14 days of a
LoI/PTU diet or LoI/PTU with T3 treatment (Table V). As previously reported at baseline (22), we
find that TRα0/0 mice have relative bradycardia compared to WT mice (371 ± 28 beats/min versus
524 ± 10 beats/min, respectively; p<0.0001). Unexpectedly, SRC-1-/- mice also had bradycardia
(388 ± 12; p<0.0001). Deletion of both TRα and SRC-1 resulted in the lowest mean heart rate.
However, this value was not significantly different from those in the TRα0/0 and SRC-1-/- above.
We also found baseline heart rates in TRβ-/- mice to be lower than WT (p=0.02). Mice showed
decreases in heart rate in response to TH deprivation (LoI/PTU, Table V), except for animals
deficient in TRα. Although all genotypes increased heart rate in response to TH treatment,
TRα0/0SRC-1-/- mice had a modest increase of 17.6 ± 3.8% compared to 27-43% observed in the
other genotypes. These data suggest that SRC-1 facilitates the TRα- mediated TH action in the
heart.

E-00226-2002.R2
15
Identification of TH-responsive genes by microarray analysis
TH responsive genes specifically mediated by TRβ or SRC-1 were identified by
microarray analysis. Microarrays of mRNA from liver of wildtype, SRC-1-/-, and TRβ-/- mice
were analyzed comparing each genotype in hypothyroid and TH-treated states. Analysis of over
1,100 genes revealed 92-105 genes that were affected by TH in each group (Table I). Using high
stringency (>2-fold change), we identified two genes that were TRβ-dependent (i.e., in which
there was no effect of TH treatment in TRβ-/- mice): osteopontin (up-regulated in WT mice) and
gluathione-S-transferase (GST) (down-regulated in WT mice). Also, we identified two genes
that were SRC-1-dependent (i.e., in which there was no effect of TH treatment in SRC-1-/- mice):
split hand split foot (SHSF) (up-regulated in WT mice) and Ets-related transactivation factor
(ERF) (down-regulated in WT mice) (Table VI).
Gene Expression in Liver with TH withdrawal and TH treatment
Confirmation of gene expression observed in the microarray analyses was done by
TaqMan quantitative real time PCR with the same and additional livers.
Osteopontin (J04806)
WT and SRC-1-/- mice had a 76 and 30% increase in osteopontin expression with T3
treatment, respectively. RNA from livers of TRβ-/- and TRβ-/-SRC-1-/- mice had no response to
T3, confirming a TRβ-dependence for TH-mediated upregulation of osteopontin demonstrated by
microarray. Although not evaluated on the microarray, TRα also appears to be necessary for
regulations of osteopontin expression, as TRα0/0 and TRα0/0SRC-1-/- mice failed to stimulate
expression with TH treatment (Table VII). SRC-1 is not absolutely required for induction of

E-00226-2002.R2
16
osteopontin by TH, though it may facilitate the response, 1.36 ± 0.16 fold (30.6 ± 0.16%)
increase in SRC-1-/- compared to 1.86 ± 0.39 fold increase (82.2 ± 50.7%) in WT.
Glutathione S-transferase (J03958)
GST expression appeared to be down-regulated by TH based on data from the microarray
in WT mice and was also TRβ-dependent. Results from quantitative analysis are consistent with
this data, showing an 0.21 ± 0.02 fold (81.7 ± 2.1%) decrease in GST expression in response to
TH. In the absence of TRβ, there was a small, paradoxical increase in GST expression in
response to TH (Table VII). Interestingly, in the absence of both TRβ and SRC-1, TH mediated
down-regulation is restored (0.23 ± 0.04 fold; -77.8 ± 5.7%). This result indicates that the two
receptor subtypes work together to facilitate down-regulation of GST in response to thyroid
hormone. However, in the presence of only the TRα (TRβ-/- mice), the additional presence of
SRC-1 inhibits TH-mediated down-regulation of GST. By eliminating the SRC-1 along with
TRβ, the inhibition of TH-mediated GST repression is removed.
Split Hand Split Foot (U41606)
In WT mice, SHSF expression was increased by 1.5 ± 0.08 fold with TH treatment, and
although a blunted response was seen in SRC-1-/- mice (1.19 ± 0.09), it was not significantly
different from WT mice. The other genotypes also showed reduced responses (Table VII).

E-00226-2002.R2
17
Ets Related Factor (U58533)
ERF was shown to be SRC-1-dependent by microarray, and we have confirmed this to be
the case by quantitative PCR (Table VII). However, unlike the TH-mediated down-regulation of
ERF seen by microarray in WT and TRβ-/- mice, quantitative RT PCR results demonstrate a
modest increase (1.59 ± 0.22 and 1.90 ± 0.38, respectively) in ERF expression. Furthermore,
TRα0/0 mice did not demonstration any effect of TH.
5’Deiodinase (5’ DI, NM_007860)
Although not included as part of the microarray, we chose to study 5’DI, a gene well
known to be up-regulated by TH. In order to evaluate the role of the TR subtypes and SRC-1 in
mediating 5’DI expression, we investigated the response of 5’DI to TH in mice deficient in either
TR subtype, SRC-1, or SRC-1 and each of the TR subtypes with TH deprivation and TH
treatment (Fig. 3). There was a 300% fold increase in 5’DI expression in WT, SRC-1-/-, and
TRα0/0SRC-1-/- mice in response to TH. An even greater response (>2,400%) was observed in
the TRα0/0 mice, consistent with the hyperresponsiveness observed with other markers of TH
action in these animals. This increased response was abrogated when SRC-1 was deleted along
with the TRα. This result indicates that the hypersensitivity seen in the TRα0/0 mice may be, in
part, due to the presence of SRC-1, as in the absence of both, the increased response is ablated.
TRβ-/- mice had a markedly reduced response (only about 2.6% of the wildtype with TH), a
reduction further compounded by deficiency in both TRβ and SRC-1 (1.3% of WT with TH).

E-00226-2002.R2
18
Discussion
These studies investigate the in vivo effect of TR subtypes in peripheral tissues,
specifically by focusing on animal growth, heart rates, energy expenditure and markers of TH
action in the liver and heart. To do this, we have bred mice deficient in the TRα or TRβ, SRC-1,
or each of the TR subtypes and SRC-1 together. These animals were generated in a mixed sv129
and C57BL/6J background. Therefore, single and double heterozygous mice were backcrossed
with wildtype littermates to generate a uniform genetic background closer to C57BL/6J. The
role of the TR subtypes in growth has been studied previously in TR-deficient animals by our
laboratory and others (10-12, 16, 30). In addition, mice deficient in SRC-1 have no reported
changes in linear growth and weight (42). Here, we show that at 5 weeks, mice deficient in TRβ
or SRC-1 have body weights similar to WT mice. However, we show somewhat decreased
linear growth in TRβ-/- mice relative to WT at 5 weeks (86.8%, significance at the 5% confidence
level) and linear growth at 9 weeks (90.9%, significance at the 5% confidence level). Although
we and others have previously reported no differences in body weight and linear growth in TRβ-/-
mice, the trends were there and with our larger number of mice, differences reach significance.
These changes are much milder than in mice deficient in TRα at 5 weeks (74.6%; 86.8%), which
is consistent with work showing that TRα1 is important for intestinal crypt development at
weaning, a phenotype that seems to improve (27). At 9 weeks, TRα0/0 mice still show decreased
linear and body weight growth (86.8%; 95.0%), similar to what has been shown previously
(12,13). However, following full maturity, both TRα- and TRβ- deficient mice have retarded
growth. Although SRC-1-/- mice achieve adult linear and body weights that are no different than
wildtype animals, loss of SRC-1 in combination with either TR subtype (TRα0/0SRC-1-/-/TRβ-/-
SRC-1-/-) results in linear and weight growth retardation compared to WT both at 5 weeks

E-00226-2002.R2
19
(86.7/92.2% and 69/76.9%, respectively) and at 9 weeks (77.7/73.0% and 90.8/90.0%,
respectively). Taken together, these data indicate that both TR subtypes play important roles in
governing linear and body weight growth. In addition, SRC-1 plays a cooperative role in growth
with each TR subtype in the absence of the other. In the absence of SRC-1, the presence of other
cofactors may compensate for its loss, but insufficiently when there is additional loss of either
TR. Although we did not investigate expression of growth hormone in TR subtype/SRC-1
deficient mice, we speculate that these levels may be normal, as has been shown in TRα0/0 (13),
TRα1-/- and TRβ-/- (13) and SRC-1-/- (32) mice. Additionally, animals deficient in both TRβ and
SRC-1 show increased energy expenditure over all other genotypes on a normal diet (p<0.001).
This corresponds with much higher baseline levels of thyroid hormone (Table III) and
progressive weight loss in these mice at 9 weeks. The use of a TRβ isoform specific ligand, GC-
1, failed to maintain core body temperature and reduced stimulation of uncoupling protein in
brown adipose tissue of hypothyroid mice (29). Taken together these data indicate that TRα
mediates the increased energy expenditure observed in response to TH supporting previous
observations (39) and that SRC-1 is not necessary for this action of TH. The increased energy
expenditure and weight loss in these animals does not correspond with illness, as these animals
demonstrate fertility and litter size at homozygosity that is similar to WT, without increased
mortality in adult animals, at least through 60 weeks, the longest we have maintained them.
That TRα is required for normal heart rate has been shown previously (15, 19, 20, 22,
30). In addition, it has also been reported by our laboratory and others that absence of TRβ does
not cause such a decline in basal heart rates, and in fact, results in a slight increase in heart rate
attributed to increased circulating levels of TH in these mice (Table III) (15, 19, 20, 33, 35, 36).
However, there appear to be a number of differences in baseline heart rates of WT mice

E-00226-2002.R2
20
dependent on strain, even in papers by the same laboratories. SRC-1 appears to be essential for
maintaining heart rate, as in its absence, alone and in combination with each of the receptor
subtypes, heart rate is decreased. In the absence of SRC-1, TRα and TRβ are unable to maintain
normal heart rates, presumably by competing for a limited supply of cofactors or an affinity of
the TRα2, which does not bind TH, for other cofactors. In the absence of TRβ alone, TRα and
SRC-1 are unable to maintain WT heart rate. However, in the absence of both TRβ and SRC-1,
there is a marked increase in serum TH (Table III) over that in TRβ-/- mice available to bind to
TRα1, which does not happen in the absence of SRC-1 alone. This seems likely in that TRβ-/-
SRC-1-/- mice achieve WT heart rates when treated with exogenous TH. Despite the proposed
role for SRC-1 in regulating heart rate inferred by these data, we have previously shown that
other TH-dependent genetic markers in heart, specifically SERCA2, MHCα, and MHCβ, are
unaffected by the absence of SRC-1 (32).
In order to discern the mechanism of TH action in the liver, we sought to identify genetic
markers that were TRβ-dependent vs. SRC-1-dependent. It has been shown in vitro that
coactivators are used by TRβ to mediate TH action (reviewed in (23)). We took livers from WT,
TRβ-/- and SRC-1-/- mice that were made hypothyroid with a LoI/PTU diet or hypothyroid or
treated with TH and subjected them to microarray analysis. From this analysis, we identified
four TH-dependent genes; two were determined to be TRβ-dependent (osteopontin; up-
regulated) and glutathione-S-transferase (GST; down-regulated) and two were SRC-1-dependent
(split hand/split foot, SHSF; up-regulated) and Ets-related transactivation factor (ERF, down-
regulated). Osteopontin is a secreted glycoprotein with an RGD domain characteristic of
integrin-binding proteins. It has been shown to be an important chemokine in inflammation, a
potential oncogene in renal cancers, a stable component in mineralized tissues (interacting with

E-00226-2002.R2
21
Vitamin D Receptor and the retinoid X receptor) and smooth muscle, with an additional presence
in the anterior pituitary (6, 25). A direct role for osteopontin regulation by TH has not been
shown. Data here confirmed microarray results that showed that osteopontin expression was
indeed TRβ-dependent, as mice deficient in TRβ (TRβ-/- and TRβ-/-SRC-1-/-) showed no response
to TH (Fig. 3A). We also showed that osteopontin may also be regulated by the TRα, as mice
deficient in TRα (TRα0/0 and TRα0/0SRC-1-/-) show a decrease in osteopontin in response to TH
(-16.9% and -%26.3, respectively), distinguishing the roles for TR subtypes in controlling this
gene. Flores-Morales et al (9) demonstrated that of 40% of TH responsive genes identified in an
expression profile were TRβ independent, also suggesting a role for TRα in modulating gene
expression. In addition, we investigated GST by RT PCR. Beckett et al showed that by
depriving rats of selenium in their diets (inhibiting T3 production in liver by 5’DI) or by PTU
diet, there was in increase in expression of GST (3, 4). However, there has been no detailed
study on the effects of administration of exogenous L-T3 on GST expression. Here, we show in
TRβ-/- mice, there is no down-regulation of GST as seen in other genotypes, which suppress GST
with TH treatment by about 75% (Table VII). Interestingly, in the absence of TRβ and SRC-1,
suppression by TH absent in TRβ-/- mice is restored. From this, we conclude that in TRβ-/- mice,
the presence of SRC-1 inhibits TRα-mediated suppression of GST, and when SRC-1 is also
removed, the TRα, possibly acting with another coregulatory molecule, mediates TH-induced
GST suppression.
Of the TH responsive genes identified to be SRC-1-dependent, we were unable to
confirm the microarray data with RT PCR for these 2 genes. In WT mice, SHSF increased by
40% without any response in SRC-1-/- mice (Table VII). However there were also blunted
responses to TH in the other genotypes investigated, indicating that SHSF will not be a good

E-00226-2002.R2
22
marker for further use. ERF was only partially SRC-1 dependent by RT PCR; however, we saw
differences between TaqMan and microarray in the response of WT and TRβ-/- mice to TH,
showing increases in ERF expression (Table VII), versus decreases seen by microarray (Table
VI). It is possible that ERF would be a good marker for future use, but it would require further
investigation. Importantly, we confirmed the viability of two new TRβ-dependent markers
(Osteopontin and GST) in the presence of TH. These genes have not been previously identified
in 2 other studies of TH responsive gene expression (8, 9). We have seen other genes reported
by microarray that were not reproduced by other methods (unpublished data) and note that the
previous studies only confirmed a small number of the genes with Northern analysis that were
identified by microarray (8, 9).
In addition to studying new markers identified by microarray, we also investigated liver
expression of 5’DI by RT PCR, an enzyme whose expression has been known to be up-regulated
by thyroid hormone via the TRβ (1). Interestingly, we saw sharp increases in 5’DI expression in
TRα0/0 mice, far greater than in any other genotype (Fig. 3). Macchia et al recently reported that
mice deficient in all known TRα isoforms have hypersensitivity to thyroid hormone (22). It was
speculated that a potential mechanism for hypersensitivity in these animals would be elimination
of the inhibitory TRα2. However, as 5’DI data would indicate, the hypersensitivity conferred
upon TRα-deficient mice (evidenced by vast TH-induced increases in 5’DI expression) is
ablated in the absence of both TRα and SRC-1, in which TRα0/0SRC-1-/- mice increase 5’DI in
response to TH to similar levels as WT and SRC-1-/- animals. This result indicates that the
hypersensitivity seen in TRα0/0 mice may be due to more than absence of the inhibitory TRα2.
In fact, it is likely that the hypersensitivity is due to an increased availability of SRC-1 to interact

E-00226-2002.R2
23
with the TRβ, an event that may be controlled in TR-competent mice by squelching of the
coactivator by the TRα.
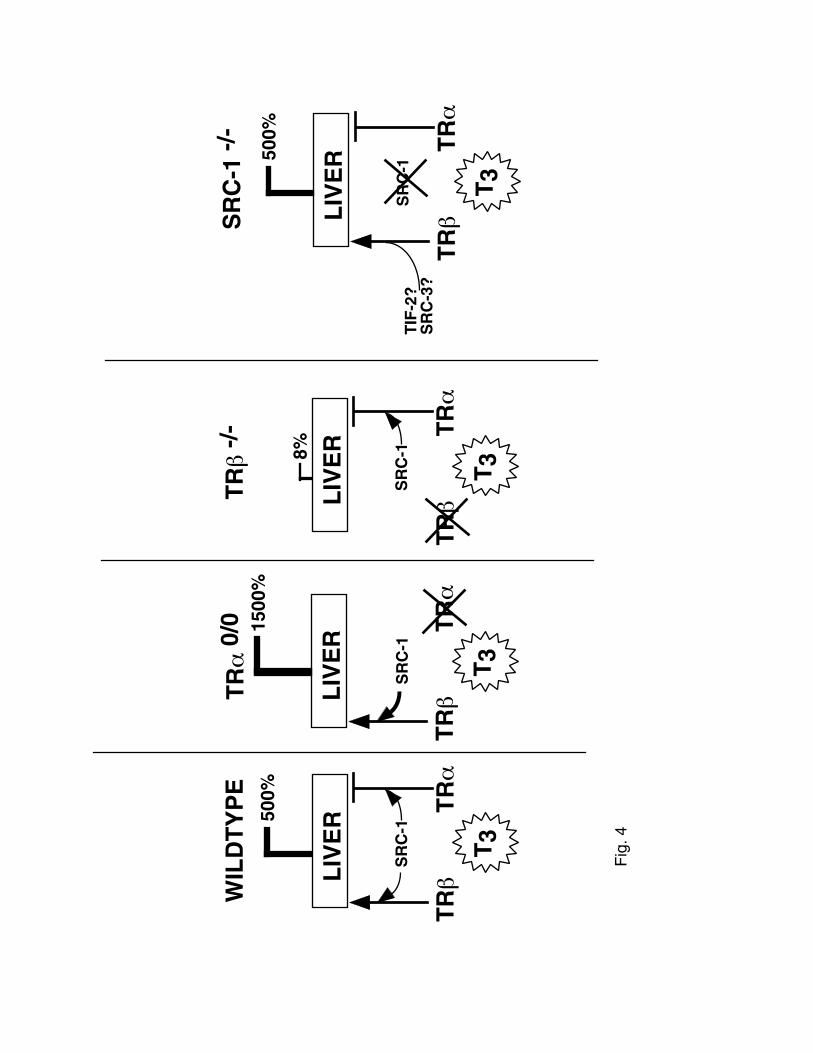
A model of TH action in liver is shown in figure 4. 5’deiodinase represents a liver gene
up-regulated by TH. TRβ1 appears to be the key isoform to mediate TH action on 5’DI in the
liver, as in its absence, there is a major reduction in 5’DI induction with TH. In TRα0/0 mice,
there is a hyperresponse in 5’DI to TH. We propose that this TH hypersensitivity is due to relief
of inhibition by TRα2, as originally hypothesized by Macchia et al (22). However, we further
this hypothesis to include that SRC-1 is necessary for this action in liver, and that SRC-1 inhibits
TRα2 activity, as TRα0/0SRC-1-/- mice are not hypersensitive to TH induced 5’DI expression.
This result might best be confirmed by overexpression of SRC-1 in vivo in liver and observing
the effect on gene expression. Additionally, we propose that SRC-1 facilitates TRβ-induced
5’DI expression, but is not necessary (Figure 5B). It is possible that in the absence of SRC-1,
alternative coactivators, such as TIF-2 or SRC-3 could compensate.
From these studies, we conclude that (1) in the absence of TRα or TRβ, SRC-1 is
important for normal growth; (2) SRC-1 partially mediates the TH effect on heart rate by TRα
and TRβ; (3) we have identified two new TH responsive markers in the liver mediated by TRβ;
osteopontin (up-regulated) and glutathione-S-transferase (down-regulated); and (4) we have
shown that SRC-1 may mediate the hypersensitivity to TH seen in TRα0/0 mice as demonstrated
by 5’deoiodinase expression in liver.

E-00226-2002.R2
24
Acknowledgements
The authors are indebted to Prof. Samuel Refetoff for advise and guidance during this project
and Dr. T. Karrision for help with statistical analyses. This work was supported in part by grants
from the National Institutes of Health DK 58281 (to R.E.W.) the Ministry of Research ACI 283 (to
J.S.), DK 58242 (to J.X.), and NICHD 078587 (to B.O’M.) and by the Seymour J. Abrams Thyroid
Research Center. The authors gratefully acknowledge the technical assistance of Kevin Cua with
preliminary energy expenditure and heart rate experiments.

E-00226-2002.R2
25
Figure Legends
Figure 1. Growth of mice of genotypes from two weeks until adulthood. Length of mice of
each genotype was determined from age 2 weeks to age 9 weeks (above). Body weight was
determined for the same time period of time. Comparisons are shown for WT, TRβ-/-, SRC-1-/-,
and TRβ-/-SRC-1-/- mice on the left (A, C) and WT, TRα0/0, SRC-1-/-, TRα0/0SRC-1-/- on the right
(B, D). Numbers of animals in each group (N) is shown in the legend. Values in the curves
represent the mean of each group ± SE. Where no SE is not present, values were sufficiently low
as to not appear using the graphing program. A table of significance is shown below the graphs.
Comparison made between WT mice and other genotypes at 5 weeks and 9 weeks by Tukey-
Kramer method at 5% confidence.
Figure 2. Relationship of serum T4 levels and energy expenditure (EE) (kcal/day/gm body
weight), (top); and nonfasting serum Leptin (ng/ml/gm body weight), (bottom). Values represent
means. Note that EE and Leptin and T4 levels were the highest in the TRβ-/-SRC-1-/- mice.
Figure 3. mRNA quantitation of 5’Deiodinase in liver. 5’Deiodinase expression was
determined by TaqMan quantitative real time PCR. Data is shown as percent increase (on a
logarithmic scale) from PTU-treated wildtype when treated with four days ip injection of T3
(0.2µg/25g/day). The percent increase is determined from the mean liver expression of these
genes from at least five mice in each treatment group and each genotype. SE determined from %
change of individual samples from PTU-treated WT mice. Five mice were used for each
hypothyroid and T3-treated group.
Figure 4. Model for thyroid hormone action in the liver. Arrows represent activation and
blockers represent inhibition.Thickness or lines or number of arrows represents potential strength
of interaction.

E-00226-2002.R2
26
References:
1. Amma LL, Campos Barros A, Wang Z, Vennstrom B, Forrest D Distinct tissue-
specific roles for thyroid hormone receptors beta and alpha1 in regulation of type 1
deiodinase expression. Molec Endocrinol 15:467-475,2001.
2. Anzick SL, Kononen J, Walker RL, Azorsa DO, Tanner MM, Guan XY, Sauter G,
Kallioniemi OP, Trent JM, Meltzer PS AIB1, a steroid receptor coactivator amplified
in breast and ovarian cancer. Science 277:965-8.,1997.
3. Beckett GJ, Nicol F, Proudfood D, Dyson K, Loucaides G, Arthur JR The changes in
heptaic enzyme expression by selenium deficiency and hypothyroidism in rats are
produced by independent mechanisms. Biochem J. 266:743-747,1990.
4. Beckett GJ, Russell A, Nicol F, Sahu P, Wolf CR, Arthur JR Effect of selenium
deficiency on hepatic type I 5-iodothyronine deiodinase activity and hepatic thyroid
hormone levels in the rat. Biochem J. 2821992.
5. Bergmann BM, Kushida CA, Everson CA, Gilliland MA, Obermeyer W,
Rechtschaffen A Sleep deprivation in the rat: II. Methodology. Sleep 12:5-12,1989.
6. Ehrchen J, Heuer H, Sigmund R, Schafer M-H, Bauer K Expression and regulation of
osteopontin and connective tissue growth factor transcripts in rat anterior pituitary. J.
Endocrinology 169:87-96,2001.
7. Eng FC, Barsalou A, Akutsu N, Mercier I, Zechel C, Mader S, White JH Different
classes of coactivators recognize distinct but overlapping binding sites on the estrogen
receptor ligand binding domain. J Biol Chem 273:28371-7,1998.

E-00226-2002.R2
27
8. Feng X, Jiang Y, Meltzer P, Yen PM Thyroid hormone regulation of hepatic genes in
vivo detected by complementary DNA microarray. Molec Endocrinol 14:947-955,2000.
9. Flores-Morales A, Gullberg H, Fernandez L, Stahlberg N, Lee NH, Vennström B,
Norstedt G Patterns of liver gene expression governed by TRβ. Molec Endocrinol
16:1257-1268,2002.
10. Forrest D, Erway LC, Ng L, Altschuler R, Curran T Thyroid hormone receptor β is
essential for development of auditory function. Nature Genet. 13:354-357,1996.
11. Forrest D, Hanebuth E, Smeyne RJ, Evereds N, Stewart CL, Wehner JM, Curran T
Recessive resistance to thyroid hormone in mice lacking thyroid hormone receptor β:
evidence for tissue-specific modulation of receptor function. EMBO J. 15:3006-
3015,1996.
12. Gauthier K, Chassande O, Plateroti M, Roux JP, Legrand C, Pain B, Rousset B,
Weiss R, Trouillas J, Samarut J Different functions for the thyroid hormone receptors
TRα and TRβ in the control of thyroid hormone production and post-natal development.
EMBO J 18:623-31,1999.
13. Gauthier K, Plateroti M, Harvey CB, Williams GR, Weiss RE, Refetoff S, Willott
JF, Sundin V, Roux JP, Malaval L, Hara M, Samarut J, Chassande O Genetic
analysis reveals different functions for the products of the thyroid hormone receptor alpha
locus. Mol Cell Biol 21:4748-60,2001.
14. Glass C, Rose D, Rosenfeld M Nuclear receptor coactivators. Curr Opin Cell Biol
9:222-232,1997.
15. Gloss B, Trost SU, Gluhm WF, Swanson EA, Clark R, Winkfein R, Janzen KM,
Giles W, Chassande O, Samarut J, Dillmann WH Cardiac ion channel expression and

E-00226-2002.R2
28
contractile function in mice with deletion of thyroid hormone receptor alpha or beta.
Endocrinology 142:544-550,2001.
16. Gothe S, Wang Z, Ng L, Kindblom JM, Campos Barros A, Ohlsson C, Vennstrom
B, Forrest D Mice devoid of all known thyroid hormone receptors are viable but exhibit
disorders of the pituitary-thyroid axis, growth and bone maturation. Genes and
Development 13:1329-1341,1999.
17. Horwitz KB, Jackson TA, Bain DL, Richer JK, Takimoto GS, Tung L Nuclear
receptor coactivators and corepressors. Molec. Endocrinol. 10:1167-1177,1996.
18. Jenster G Coactivators and corepressors as mediators of nuclear recpetor function: An
update. Molec Cell Endocrinol 143:1-7,1998.
19. Johansson C, Gothe S, Forrest D, Vennstrom B, Thoren P Cardiovascular phenotype
and temperature control in mice lacking thyroid hormne receptor β or both α1 and β. Am
J Physiol 276:H2006-H2012,1999.
20. Johansson C, Vennstrom B, Thoren P Evidence that decreased heart rate in thyroid
hormone receptor alpha1 deficient mice is an intrinsic defect. Am J Physiol 275:R640-
R646,1998.
21. Leo C, Li H, Chen JD Differential mechanisms of nuclear receptor regulation by
receptor associated Coactivator 3. J Biol Chem 275:5976-5982,2000.
22. Macchia PE, Takeuchi Y, Kawai T, Cua K, Gauthier K, Chassande O, Seo H,
Hayashi Y, Samarut J, Murata Y, Weiss RE, Refetoff S Increased sensitivity to
thyroid hormone in mice with complete deficiency of thyroid hormone receptor alpha.
Proc Natl Acad Sci U S A 98:349-54,2001.

E-00226-2002.R2
29
23. McKenna NJ, O'Malley BW Combinatorial control of gene expression by nuclear
receptors and coactivators. Cell 108:465-474,2002.
24. Morte B, Manzano J, Scanlan T, Vennström B, Bernal J Deletion of the thyroid
hormone receptor 1 prevents the structural alterations of the cerebellum induced by
hypothyroidism. Proc Natl Acad Sci 99:3985-3989,2002.
25. O'Regan AW, Nau GJ, Chupp GL, Berman JS Oteopontin (Eta-1) in cell mediated
immunity: Teaching an old dog new tricks. Immunology Today 21:475-478,2000.
26. Onate SA, Tsai SY, Tsai MJ, O'Malley BW Sequence and characterization of a
coactivator for the steroid hormone receptor superfamily. Science 270:1354-1357,1995.
27. Plateroti M, Gauthier K, Domon-Dell C, Freund JN, Samarut J, Chassande O
Functional interference between thyroid hormone receptor alpha (α) and natural
truncated TRα isoforms in the control of intestine development. Mol Cell Biol 21:4761-
72,2001.
28. Pohlenz J, Maqueem A, Cua K, Weiss RE, Van Sande J, Refetoff S Improved
radioimmunoassay for measurement of mouse thyrotropin in serum: Strain differences in
thyrotropin concentration and thyrotroph sensitivity to thyroid hormone. Thyroid 9:1265-
1271,1999.
29. Ribeiro MO, Carvalho SD, Schultz JJ, Chiellini G, Scanlan TS, Bianco AC, Brent
GA Thyroid hormone-sympathetic interaction and adaptive thermogenesis are thyroid
hormone receptor isoform-specific. J Clin Invest 108:97-105,2001.
30. Saltó C, Kindblom JM, Johansson C, Wang Z, Gullberg H, Norström K, Mansén A,
Ohlsson C, Thorén P, Forrest D, Vennström B Ablation of TRα2 and a concomitant

E-00226-2002.R2
30
overexpression of α1 yields a mixed hypo- and hyperthyroid phenotype in mice. Molec
Endocrinol 15:2115-2128,2001.
31. Spanjaard R, Nguyen V, Chin W Rat rev-erbA alpha, an orphan receptor related to
thyroid hormone receptor, binds to specific thyroid hormone response elements. Molec
Endocrinol 8:286-295,1994.
32. Takeuchi Y, Murata Y, Sadow PM, Hayashi Y, Seo H, Xu J, O'Malley BW, Weiss
RE, Refetoff S Steroid receptor coactivator-1 deficiency causes variable alterations in the
modulation of T3-regulated transcription of genes in vivo. Endocrinology 143:1346-
1352,2002.
33. Weiss RE, Chassande O, Koo EK, Macchia PE, Cua K, Samarut J, Refetoff S
Thyroid function and effect of aging in combined heter/homozygous mice deficient in
thyroid hormone receptors alpha and beta genes. J Endocrinol 172:177-185,2002.
34. Weiss RE, Forrest D, Pohlenz J, Cua K, Curran T, Refetoff S Thyrotropin regulation
by thyroid hormone in thyroid hormone receptor β-deficient mice. Endocrinology
138:3624-3629,1997.
35. Weiss RE, Gehin M, Xu J, Sadow PM, O'Malley BW, Chambon P, Refetoff S
Thyroid function in mice with compound heterozygous and homozygous disruption of
SRC-1 and TIF-2 coactivators: evidence for haploinsufficiency. Endocrinology
143:1554-1557,2002.
36. Weiss RE, Korcarz C, Chassande O, Cua K, Sadow PM, Koo E, Samarut J, Lang R
Thyroid hormone and cardiac function in mice deficient in thyroid hormone receptors
alpha or beta: An echocardiograph study. Am J Physiol in press2002.

E-00226-2002.R2
31
37. Weiss RE, Murata Y, Cua K, Hayashi Y, Forrest D, Seo H, Refetoff S Thyroid
hormone action on liver, heart and energy expenditure in thyroid hormone receptor β
deficient mice. Endocrinology 139:4945-4952,1998.
38. Weiss RE, Xu J, Ning G, J. P, O'Malley B, Refetoff S Mice deficient in the steroid
receptor co-activator 1 (SRC-1) are resistant to thyroid hormone. EMBO J. 18:1900-
1904,1999.
39. Wikström L, Johansson C, Salto C, Barlow C, Campos Barros A, Baas F, Forrest D,
Thorén P, Vennström B Abnormal heart rate and body temperature in mice lacking
thyroid hormone receptor α1. EMBO J. 17:455-461,1998.
40. Wolffe AP, Kurumizaka H The nucleosome: a powerful regulator of transcription. Prog
Nucleic Acid Res Mol Biol 61:379-422,1998.
41. Xu J, Liao L, Ning G, Yoshida-Komiya H, Deng C, O'Malley BW The steroid
receptor coactivator SRC-3 (p/CIP/RAC3/AIB1/ACTR/TRAM-1) is required for normal
growth, puberty, female reproductive function, and mammary gland development. Proc
Natl Acad Sci U S A 97:6379-84,2000.
42. Xu J, Qui Y, DeMayo FJ, Tsai SY, Tsai M-J, O'Malley BW Partial hormone
resistance in mice with disruption of the steroid receptor coactivator-1 (SRC-1) gene.
Science 279:1922-1925,1998.
43. Xu L, Glass CK, Rosenfeld MG Coactivator and corepressor complexes in nuclear
receptor function. Curr Opin Genet Dev 9:140-7,1999.
44. Yen PM Physiological and molecular basis of thyroid hormone action. Physiol Rev
81:1097-1142,2001.

Tab
le I
. G
enes
up
and
dow
n re
gula
ted
by T
H in
TR
β-/- a
nd S
RC
-1-/
-m
ice*
.G
enot
ype
#Gen
es R
espo
ndin
g (%
)#U
p R
egul
ated
(%
)#D
own
Reg
ulat
ed (
%)
WT
92
(7.8
)77
(83
.7)
15 (
16.3
)T
Rβ-/
-99
(8.
4)34
(34
.2)
65 (
65.6
)SR
C-1
-/-
105
(8.9
)97
(92
.3)
8 (
7.6)
*Thr
esho
ld r
atio
s of
1.6
7 w
ere
used
for
eac
h co
mpa
riso
n

TA
BLE
II.
Pro
be/P
rimer
Set
s fo
r T
aqM
an Q
uant
itativ
e R
eal T
ime
PC
R u
sed
for
mR
NA
qua
ntita
tion.
GE
NE
prim
ers
sequ
ence
sor
ient
atio
n
5'-TGCCTCCTCCCTCCCGGTGA
-3'
sens
e*O
steo
pont
in5'
-GATTTGCTTTTGCCTGTTTGG
-3'
sens
e5'
-TGAGCTGCCAGAATCAGTCACT
-3'
an
tisen
se
5'-CGTGCTTCACTACTTCAATGCCCGG
-3'
sens
e*G
ST
5'-GCAATGGCCGGGAAGC
-3'
sens
e5'
-ACCTGATGCACTCCATTCTGC
-3'
antis
ense
5'-TCTTGGCGCGGCGCATGTC
-3'
sens
e*S
HS
F5'
-GAGGTCCGAGCAGCTTGG
-3'
sens
e5'
-AAGTCGACCGGCTGCTTCTT
-3'
antis
ense
5'-CCAGCACGCCACAGCCCAACT
-3'
sens
e*E
RF
5'-CCACGACGGGTGAGCTCT
-3'
se
nse
5'-ATAGCTCTCAGGAATCTCGGTGC
-3'
antis
ense
5'-CTGCCTGAGAGGCTCTACGTGATACAGGA
-3'
sens
e*5’
DI
5'-AGCCAGCTCTACGCGGC
-3'
se
nse
5'-CCCTTGTAGCAGATCCTGCC
-3'
antis
ense
*FA
M/T
AM
RA
fluo
resc
ent T
aqM
an P
robe
(M
egaB
ases
, Eva
nsto
n, IL
).

Tab
le I
II.
Thy
roid
fun
ctio
n te
sts.
BA
SEL
INE
PT
U/L
oI1
PT
U/L
oI +
L-T
32
GE
NO
TY
PE
TT
4*T
T3*
TSH
*T
SH*
TSH
*
µg/d
lng
/dl
mU
/lm
U/l
mU
/l
Wild
type
3.7
± 0
.1 (
53)
73
± 5
(21
) 3
0 ±
5 (
65)
93
11 ±
811
(16
)
4
6 ±
21
(16)
TR
β-/-
7.6
± 0
.3 (
53)**
139
± 9
(23
)**25
8 ±
44
(57)
**
1491
6 ±
1,3
06 (
21) *
*
2941
± 3
01 (
17) *
*
TR
α0/0
3.2
± 0
.1 (
33) *
*96
± 6
(12
) 2
3 ±
4 (
34)
4754
± 5
29 (
17) *
*
3
2 ±
10
(18)
** SRC
-1-/
- 4
.7 ±
0.2
(19
) **
130
± 4
(12
) **
74
± 1
9 (1
9) *
*92
76 ±
840
(24
)
966
± 3
27 (
18) *
*
TR
β-/-SR
C-1
-/-
12.6
± 2
.6 (
11) *
*40
4 ±
39
(7) *
*73
3 ±
152
(19
) **
2346
8 ±
1,4
40 (
18) *
* 1
2645
± 1
,290
(12
) **
TR
α0/0 SR
C-1
-/-
6.5
± 0
.3 (
18) *
* 1
59 ±
8 (
14) *
* 5
2 ±
12
(18)
**
52
58 ±
591
(19
) **
10
25 ±
449
(12
) **
Mea
n ±
sta
ndar
d er
ror.
Num
bers
in p
aren
thes
es =
num
ber
of m
ice
1 Low
Iod
ine/
PTU
die
t for
14
days
2 LoI
/PT
U d
iet 1
4 da
ys w
ith I
P L
-T3 0
.2µg
/25g
bod
y w
t/day
*, O
ne-w
ay A
NO
VA
for
eff
ect o
f ge
noty
pe o
n th
yroi
d te
st e
xam
ined
, p<
0.00
01
**si
gnif
ican
ce a
t 5%
by
Tuk
ey/K
ram
er c
ompa
red
to w
ildty
pe
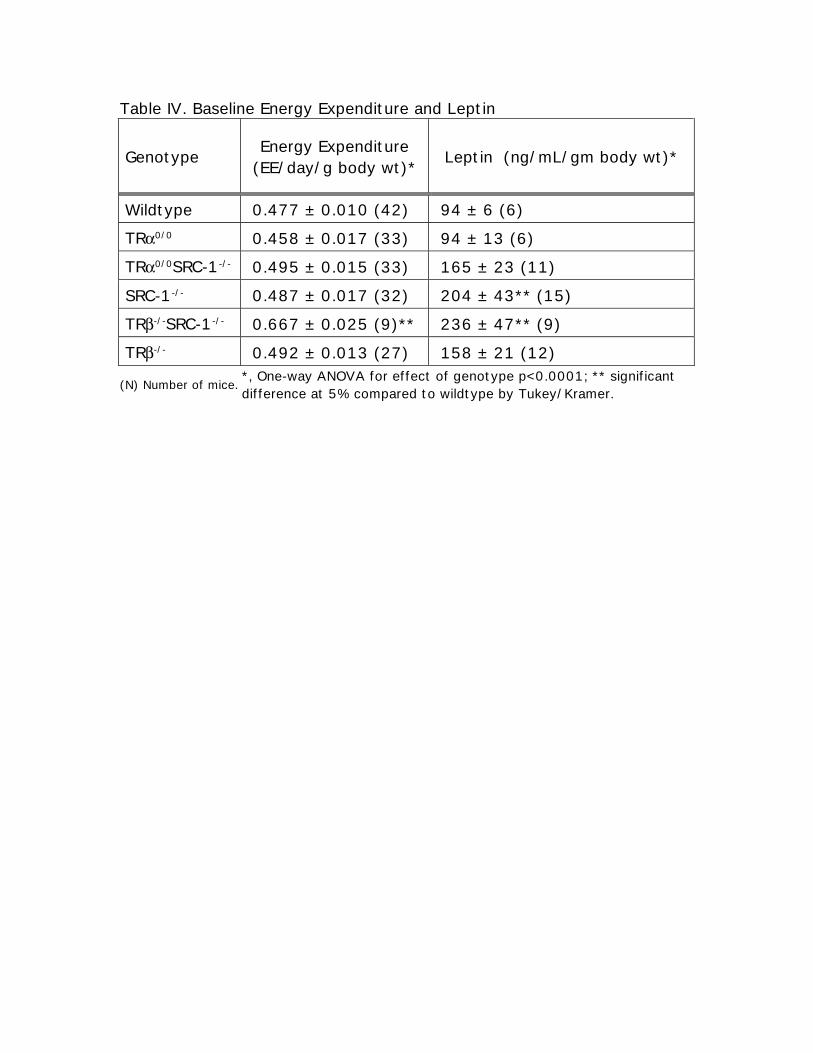
Table IV. Baseline Energy Expenditure and Leptin
Genotype Energy Expenditure(EE/day/g body wt)* Leptin (ng/mL/gm body wt)*
Wildtype 0.477 ± 0.010 (42) 94 ± 6 (6)
TRα0/0 0.458 ± 0.017 (33) 94 ± 13 (6)
TRα0/0SRC-1-/- 0.495 ± 0.015 (33) 165 ± 23 (11)
SRC-1-/- 0.487 ± 0.017 (32) 204 ± 43** (15)
TRβ-/-SRC-1-/- 0.667 ± 0.025 (9)** 236 ± 47** (9)
TRβ-/- 0.492 ± 0.013 (27) 158 ± 21 (12)
(N) Number of mice.*, One-way ANOVA for effect of genotype p<0.0001; ** significantdifference at 5% compared to wildtype by Tukey/Kramer.
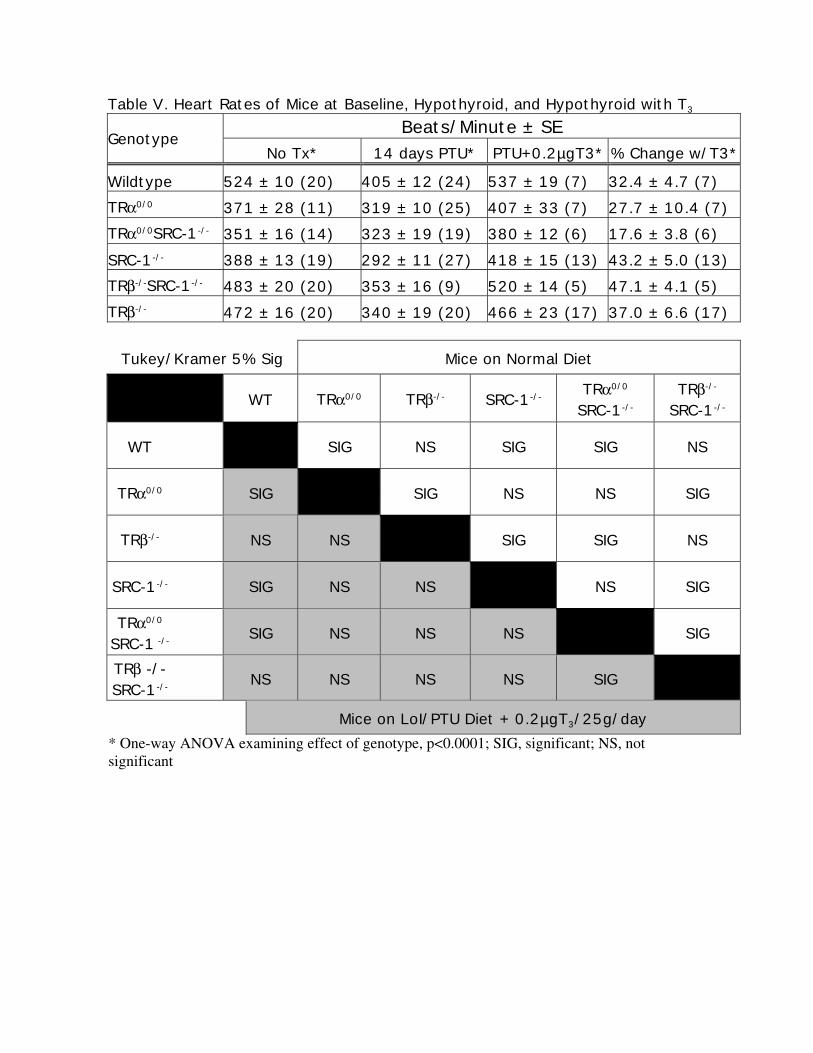
Table V. Heart Rates of Mice at Baseline, Hypothyroid, and Hypothyroid with T3
Beats/Minute ± SEGenotype
No Tx* 14 days PTU* PTU+0.2µgT3* % Change w/T3*
Wildtype 524 ± 10 (20) 405 ± 12 (24) 537 ± 19 (7) 32.4 ± 4.7 (7)TRα0/0 371 ± 28 (11) 319 ± 10 (25) 407 ± 33 (7) 27.7 ± 10.4 (7)TRα0/0SRC-1-/- 351 ± 16 (14) 323 ± 19 (19) 380 ± 12 (6) 17.6 ± 3.8 (6)
SRC-1-/- 388 ± 13 (19) 292 ± 11 (27) 418 ± 15 (13) 43.2 ± 5.0 (13)TRβ-/-SRC-1-/- 483 ± 20 (20) 353 ± 16 (9) 520 ± 14 (5) 47.1 ± 4.1 (5)TRβ-/- 472 ± 16 (20) 340 ± 19 (20) 466 ± 23 (17) 37.0 ± 6.6 (17)
Tukey/Kramer 5% Sig Mice on Normal Diet
WT TRα0/0 TRβ-/- SRC-1-/- TRα0/0
SRC-1-/-
TRβ-/-
SRC-1-/-
WT SIG NS SIG SIG NS
TRα0/0 SIG SIG NS NS SIG
TRβ-/- NS NS SIG SIG NS
SRC-1-/- SIG NS NS NS SIG
TRα0/0
SRC-1 -/- SIG NS NS NS SIG
TRβ -/-SRC-1-/- NS NS NS NS SIG
Mice on LoI/PTU Diet + 0.2µgT3/25g/day* One-way ANOVA examining effect of genotype, p<0.0001; SIG, significant; NS, notsignificant

Tab
le V
I. G
enes
iden
tifi
ed t
o be
up
or d
own
regu
late
d in
res
pons
e to
TH
and
dep
ende
nt o
n ei
ther
TR
or
SRC
-1.
G
enba
nk #
(Nam
e)E
ffec
t of
TH
WT
TR
-/-
SRC
-1-/
-
TR
Dep
ende
nt
J048
06 (
oste
opon
tin)
↑2.8
no e
ffec
t↑2
.0
J039
58 (
glut
athi
one
s-tr
ansf
eras
e)↓3
.5no
eff
ect
↓4.3
SRC
-1 D
epen
dent
U
4160
6 (s
plit
hand
/foo
t gen
e)↑2
.4↑2
.7no
eff
ect
U
5853
3 (E
ts-r
elat
ed tr
ans.
Fac
tor)
↓1.8
↓2.4
no e
ffec
t

Tab
le V
II.
Fold
indu
ctio
n w
ith L
-T3 t
reat
men
t of
vari
ous
gene
s m
easu
red
by T
aqM
an P
CR
(N
=5
anim
als
in e
ach
grou
p).
Gen
otyp
eO
steo
pont
inG
luta
thio
ne-S
-Tra
nsfe
rase
Split
Han
d Sp
lit F
oot
Ets
-Rel
ated
Fac
tor
Wild
type
1.86
± 0
.39*
0.21
± 0
.02*
1.45
± 0
.08*
1.59
± 0
.22*
TR
α0/0
0.89
± 0
.11∆
0.31
± 0
.04*
1.14
± 0
.04∆
0.96
± 0
.16
TR
β-/-
1.02
± 0
.13∆
1.34
± 0
.16∆
1.17
± 0
.09
1.9
± 0
.38*
SRC
-1-/
-1.
36 ±
0.1
6*0.
19 ±
0.0
4*1.
19 ±
0.0
91.
3 ±
0.2
1
TR
α0/0 /S
RC
-1-/
-0.
84 ±
0.0
6∆0.
25 ±
0.0
5*1.
03 ±
0.0
5∆0.
76 ±
0.1
1
TR
β-/- /S
RC
-1-/
-1.
00±
0.1
7∆0.
23 ±
0.0
4*0.
91 ±
0.0
2∆1.
01 ±
0.1
1
AN
OV
Ap
Val
ue0.
0062
<0.
0001
<0.
0001
0.00
35
*: 5
% s
igni
fica
nce
com
pare
d to
PT
U tr
eatm
ent,
sam
e ge
noty
pe∆:
5%
sig
nifi
canc
e co
mpa
red
to L
-T3 t
reat
men
t, w
ildty
pe
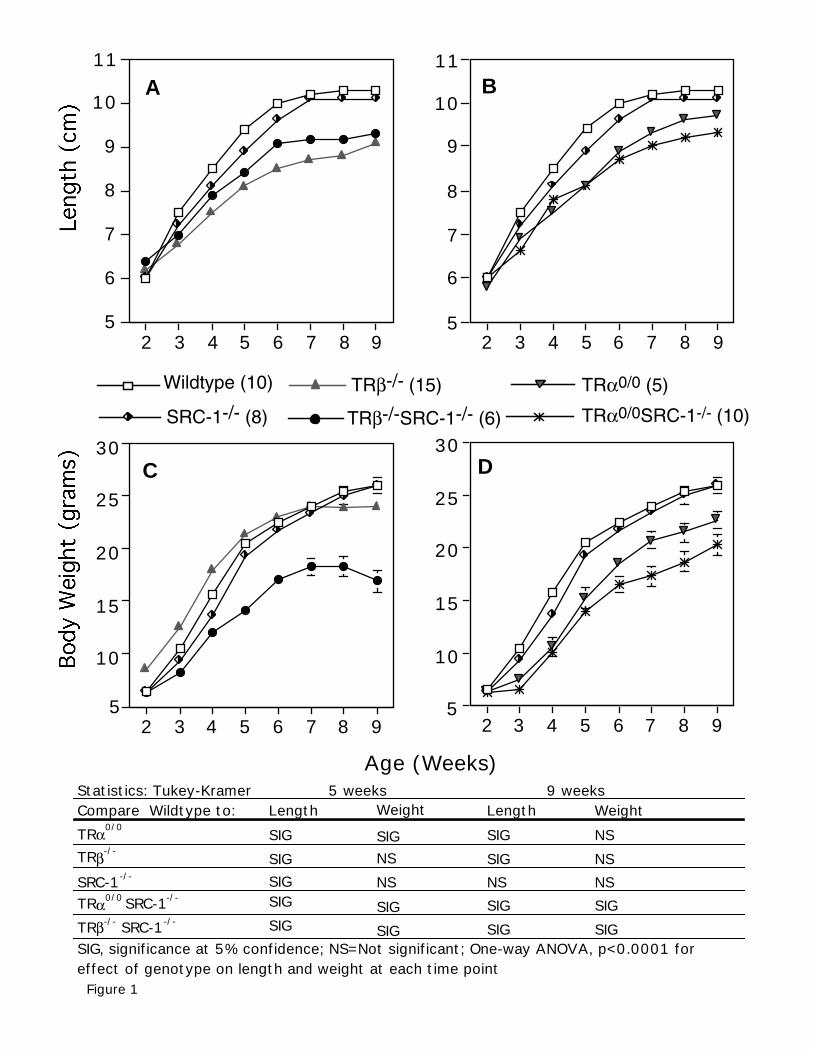
Figure 1
Age (Weeks)
5
10
15
20
25
30
2 3 4 5 6 7 8 95
10
15
20
25
30
2 3 4 5 6 7 8 9
TRβ-/- (15)
TRβ-/-SRC-1-/- (6)SRC-1-/- (8)
Wildtype (10)
TRα0/0SRC-1-/- (10)
5
6
7
8
9
10
11
2 3 4 5 6 7 8 95
6
7
8
9
10
11
2 3 4 5 6 7 8 9
A
Statistics: Tukey-KramerCompare Wildtype to: Length Weight Length Weight
SIG SIG SIG NSSIG NS SIG NSSIG NS NS NSSIG SIG SIG SIGSIG SIG SIG SIG
SIG, significance at 5% confidence; NS=Not significant; One-way ANOVA, p<0.0001 for effect of genotype on length and weight at each time point
TRα0/0
TRβ-/-
5 weeks 9 weeks
SRC-1-/-
TRα0/0 SRC-1-/-
TRβ-/- SRC-1-/-
C
B
D
TRα0/0 (5)

[T4]
0.4
0.5
0.6
0.7
4 6 8 10 12 14
WTTRβ-/-
TRαo/oSRC-1-/-
TRβ-/-/SRC-1-/-
TRαo/oSRC-1-/-
WT
TRβ-/-
TRαo/o
SRC-1-/-TRβ-/-/SRC-1-/-
TRαo/oSRC-1-/-
0
100
200
300
Fig. 2

WT TR 0/0 TR -/-SRC-1-/-TR 0/0
SRC-1-/-TR -/-
SRC-1-/-
Figure 3
1
10
100
1000
10000
LIV
ER
SR
C-1
TR
TR
T3
500%
WIL
DT
YP
E
LIV
ER
SR
C-1
TR
TR
T3
1500
%T
R0/
0
LIV
ER
SR
C-1
TR
TR
T38%
TR
-/-
LIV
ER
SR
C-1
TR
TR
T3
500%
SR
C-1
-/-
TIF
-2?
SR
C-3
?
Fig
. 4
Related Documents











