Specificity of catecholamine-induced growth in Escherichia coli O157:H7, Salmonella enterica and Yersinia enterocolitica Primrose P.E. Freestone 1 , Richard D. Haigh 1 and Mark Lyte 2* Department of Infection, Immunology and Inflammation, University of Leicester-Warwick School of Medicine, Leicester, United Kingdom 1 and Department of Pharmacy Practice, School of Pharmacy, Texas Tech University Health Sciences Center, Lubbock, TX 79430, USA 2 * Corresponding author: Dr. Mark Lyte Department of Pharmacy Practice Texas Tech University Health Sciences Center 3601 4 th Street, STOP 8182 Lubbock, TX 79430-8162, USA Telephone: (806) 743-4200, ext. 262 FAX: (806) 743-4209 Email: [email protected] 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Specificity of catecholamine-induced growth in Escherichia coli O157:H7,
Salmonella enterica and Yersinia enterocolitica
Primrose P.E. Freestone1, Richard D. Haigh1 and Mark Lyte2*
Department of Infection, Immunology and Inflammation, University of Leicester-Warwick
School of Medicine, Leicester, United Kingdom1 and Department of Pharmacy Practice,
School of Pharmacy, Texas Tech University Health Sciences Center, Lubbock, TX 79430,
USA2
* Corresponding author:
Dr. Mark Lyte
Department of Pharmacy Practice
Texas Tech University Health Sciences Center
3601 4th Street, STOP 8182
Lubbock, TX 79430-8162, USA
Telephone: (806) 743-4200, ext. 262
FAX: (806) 743-4209
Email: [email protected]
1

Abstract
The present study demonstrates that catecholamine responsiveness in Yersinia
enterocolitica, a bacterial pathogen whose infectious spectrum is principally limited to the gut,
is limited to norepinephrine and dopamine, and not epinephrine; this behavior contrasts with
observations for two pathogens with a wider extra-gastrointestinal spectrum, Escherichia coli
O157:H7 and Salmonella enterica, which respond to all three catecholamines. Epinephrine
showed lower potency than norepinephrine and dopamine in inducing growth of E. coli and S.
enterica, and was a potent antagonist of norepinephrine and dopamine growth
responsiveness in Y. enterocolitica. Given that only norepinephrine and dopamine and not
epinephrine containing neurons are found with the enteric nervous system, our results
suggest that certain of the more exclusive enteric pathogens may have developed response
systems preferentially for those neuroendocrine hormones that are produced by the enteric
nervous system as host-derived signals by which to sense the environment and initiate
pathogenic processes.
Keywords: norepinephrine, epinephrine, dopamine, iron, enteric pathogens
2

Introduction
The intersection of the fields of microbiology and neuroendocrinology represents an
interdisciplinary approach to the study of the infectious disease process that has been termed
microbial endocrinology (Lyte, 1993; Lyte, 2004). This field has as its central tenet that
microbes can use host hormones as environmental cues to initiate growth and pathogenic
processes. Microbial endocrinology research has focussed primarily on the role in the
infectious disease process of the catecholamines, a group of organic compounds responsible
for many different neuroendocrine signalling phenomena in multicellular organisms. The
catecholamine hormones epinephrine (Epi) and norepinephrine (NE) play an integral part of
the acute stress response in metazoa.
A role for catecholamine stress hormones in the infectious disease process was first
suggested in the early 1930’s (Lyte, 2004), but was not clearly elucidated until 1992 when the
first demonstration of the direct action of catecholamines on bacterial growth was shown by
Lyte and co-workers (Lyte & Ernst, 1992) and a theory proposed for a direct role of
catecholamine-bacterial interaction in the pathogenesis of infectious disease (Lyte, 1992,
Lyte, 1993). More recent interest in the role that catecholamines may play in bacterial
pathogenesis has been heightened by reports that NE, Epi and dopamine (Dop), can directly
stimulate bacterial growth and elaboration of virulence-associated factors, as well as induce
the production of autoinducer-like substances (Lyte & Ernst, 1992; Freestone, et al., 1999;
Kinney, et al., 1999; Kinney, et al., 2000; Neal, et al., 2001; Belay & Sonnenfeld, 2002;
Freestone, et al., 2002; Reissbrodt, et al., 2002; Lyte, et al., 2003).
The biochemical pathway for the synthesis of catecholamines is L-dopa (most
commonly from food-borne sources) → dopamine → norepinephrine → epinephrine. NE and
Dop containing sympathetic nerve terminals are distributed widely throughout the body,
3

including the intestinal tract where they make up part of the enteric nervous system (ENS)
(Costa, et al., 2000). Indeed, half of the NE present within the mammalian body is
synthesized and utilized within the ENS. Importantly, Epi is not synthesized within the ENS
and so far appreciable levels of Epi have not been detected within the gastrointestinal tract
(Costa, et al., 2000).
We, and others (Lyte & Ernst, 1992; Freestone, et al., 1999; Kinney, et al., 1999;
Kinney, et al., 2000; Neal, et al., 2001; Belay & Sonnenfeld, 2002; Freestone, et al., 2002;
Reissbrodt, et al., 2002; Lyte, et al., 2003) have demonstrated that the in vitro growth of a
number of bacterial species can be increased in the presence of one or more of the
catecholamines NE, Epi and Dop. However, there have been conflicting reports concerning
the ability of any one specific catecholamine to affect bacterial growth (Belay & Sonnenfeld,
2002; Belay, et al., 2003). Additionally, Sperandio and co-workers have reported that certain
aspects of the virulence of E. coli O157:H7 were regulated preferentially by Epi (Walters &
Sperandio, 2006), and suggested Epi may be a specific host hormonal cue for
enteropathogenic bacteria. In marked contrast, Lyte and Ernst (Lyte & Ernst, 1992) had
earlier found that another enteric pathogen, Y. enterocolitica, showed no apparent
responsiveness to Epi. However, the Lyte and Ernst study was performed with a single Y.
enterocolitica isolate, while the Walters and Sperandio report investigated Epi effects on the
virulence of high cell density cultures of E. coli O157:H7. To clarify whether there exists
preferences for catecholamine stress hormones amongst enteric bacteria that are reflective of
the host environment in which they are normally found, we undertook a comparative analysis
of the specificity of catecholamine responsiveness of three enteric pathogens characterized
by either their propensity to primarily inhabit the gut (Y. enterocolitica, or to colonize extra-
intestinal sites (E. coli O157:H7 and S. enterica).
4

Materials and methods
Bacterial strains and growth conditions
Recent clinical and reference (NCTC) isolates of Y. enterocolitica isolates were obtained from
Dr. Paddy Kimmit of the Leicester Public Health Laboratory, Leicester Royal Infirmary,
Leicester UK. S. enterica strain SL1344 was obtained from Dr. Jay Hinton, Institute of Food
Research, Norwich, UK. E. coli O157:H7 strain NCTC 12900 was used previously (Freestone,
et al., 2003). Serum-SAPI medium was prepared as described previously (Lyte & Ernst, 1992;
Freestone, et al., 1999), and had the following composition: 6.25 mM NH4NO3, 1.84 mM
KH2PO4, 3.35 mM KCl, 1.01 mM MgSO4 and 2.77 mM glucose, pH 7.5, supplemented with
30% (v/v) adult bovine serum) (Sigma, Poole, UK). DMEM medium, apo-forms of human
transferrin (Tf), epinephrine, dopamine and norepinephrine were all purchased from Sigma,
Poole, UK. 55FeCl3 (IES, specific activity 5 mCi/mg Fe), 3H-NE (TRK584,l-[7,8-3H]
norepinephrine) were obtained from Amersham Life Science, UK.
Catecholamine growth response and Epi antagonism assays
A serum-based medium was employed to more closely approximate the growth restrictive in
vivo conditions within a mammalian host (such as complement, antibodies, low nutrient
availability and Fe-restriction due to the presence of host iron sequestering proteins) (Lyte,
2004). Catecholamine (Dop, Epi and NE) dose response and antagonism assays were
performed in serum-SAPI supplemented with concentrations of the compounds as shown in
the text. Controls comprised equivalent volumes of the solvent used to dissolve the
catecholamine. To demonstrate that Epi was not directly inhibitory to Y. enterocolitica growth,
Epi antagonism of catecholamine-growth induction assays were also performed in the
presence of a concentration of Fe which overcomes the Fe-limitation of serum-SAPI (100 µM
5

Fe(NO3)3) (Burton, et al., 2002). Bacteria were inoculated into serum-SAPI at approximately
50-100 CFU per ml and incubated statically at 37°C in a 5% CO2 humidified incubator for 18
hrs in the case of E. coli O157:H7 and S. enterica, and 40 hours in the case of Y.
enterocolitica (which is slower growing in serum; Lyte & Ernst, 1992).
Dependence of catecholamine responsiveness on bacteria cell density
To examine correlations between bacterial cell density and specificity of catecholamine
responsiveness, one ml aliquots of overnight cultures of E. coli O157:H7, S. enterica and Y.
enterocolitica were pelleted by centrifugation at 5000 x g for 5 minutes, washed twice in warm
serum-SAPI, re-suspended in 1 ml of serum-SAPI and serially diluted to a nominal cell
density of less than 1 CFU per ml in serum-SAPI containing no additions (control), or 50 µM
NE, 50 µM Dop or 100 µM Epi. Cultures were incubated and enumerated for growth as
described above.
Analysis of effects of Epinephrine on catecholamine-mediated Y. enterocolitica
incorporation of iron from transferrin and uptake of NE
55Fe-Tf was prepared as described previously (Freestone, et al., 2000). Exponential bacteria
were inoculated at 108 CFU per ml into serum-SAPI supplemented with 2 x 105 counts per
minute (cpm) of 55Fe-Tf plus catecholamines added at the concentrations indicated in the
legend to Table 1. Cultures were incubated at 37°C in a 5% CO2 humidified incubator for 6
hrs, harvested by centrifugation at 5000 x g for 5 minutes, washed in PBS and assayed for
cell numbers and 55Fe incorporation using pour plate analysis and scintillation counting as
described previously (Freestone, et al., 2000). Assays were performed in triplicate on at least
two occasions; variation within individual assay sets was usually less than 5%, and between
experiments less than 10%.
6

To examine the effects of Epi on uptake of NE, exponential cultures of Y. enterocolitica
were harvested, washed twice in DMEM, and added at 2 x 108 CFU per ml to fresh DMEM
containing 50 μM NE plus 5 x 105 cpm per ml of 3H-NE and either no additions (control), or
100-300 μM Epi. Cultures were incubated statically for 6 hrs at 37°C in a 5% CO2 humidified
incubator (this time point was determined by conducting a prior time course of 3H-NE uptake
and represents the time required for maximal uptake), and then analyzed for growth and 3H-
NE uptake as described for the 55Fe incorporation assays above. 3H-NE uptake assays were
performed in duplicate on at least two occasions; variation within individual assay sets was
5% or less, and between experiments no more than 15%.
Statistical analysis
Where appropriate, statistical analysis was performed using an unpaired t-test in which a two-
tailed P value was calculated (Instat program, GraphPad Software, San Diego, CA, USA).
Statistical significance was defined as a P value of <0.05.
7

Results
Analysis of the specificity of catecholamine growth responsiveness in E. coli O157:H7
and Y. enterocolitica
During our initial investigations of the growth responsiveness of clinical bacterial isolates to
NE we occasionally found strains that had acquired mutations rendering them deficient in NE
recognition (Freestone, et al., 1999). In addition, we had also determined that bacterial
responses to catecholamine stress hormones are markedly concentration dependent (Lyte &
Ernst, 1992; Freestone, et al., 2002). We therefore investigated whether the lack of Epi
growth induction previously observed in a single Y. enterocolitica isolate (Lyte & Ernst, 1992)
was due either to a strain specific mutation, or simply because the concentrations of Epi
utilized were too low, by analyzing the responses of 14 Y. enterocolitica strains (including 5
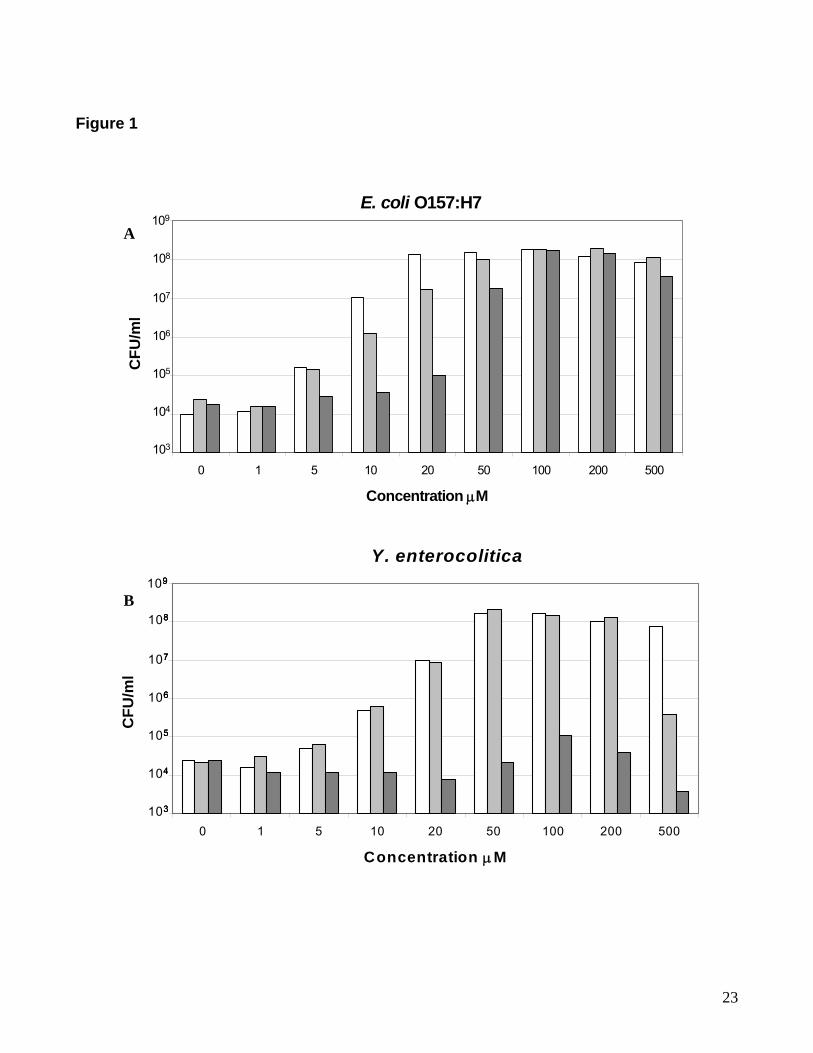
NCTC strains) to a range of Epi concentrations. Figure 1A shows a typical growth response
profile of one of these 14 strains (NCTC 551) to increasing concentrations of Epi, compared
with similar concentrations of NE and Dop; an equivalent profile from E. coli O157:H7 is
shown in Figure 1B. It can clearly be seen that there is no growth responsiveness to
concentrations of Epi that are stimulatory to E. coli, indicating that the inability of Epi to affect
the growth of Y. enterocolitica does not appear to be a consequence of catecholamine
potency. The dose-response growth effect of the catecholamines for both Y. enterocolitica
and E. coli O157:H7 (Figure 1) also shows that on a concentration-dependent basis NE
exposure had the most potent ability among the catecholamines to stimulate bacterial growth.
8

Epinephrine acts as an antagonist of norepinephrine and dopamine growth induction
in Y. enterocolitica
It has previously been shown that catecholamines can act in a synergistic manner to promote
bacterial growth (Freestone, et al., 2002). The response of Y. enterocolitica to
catecholamines was further investigated by examining whether Epi could synergistically affect
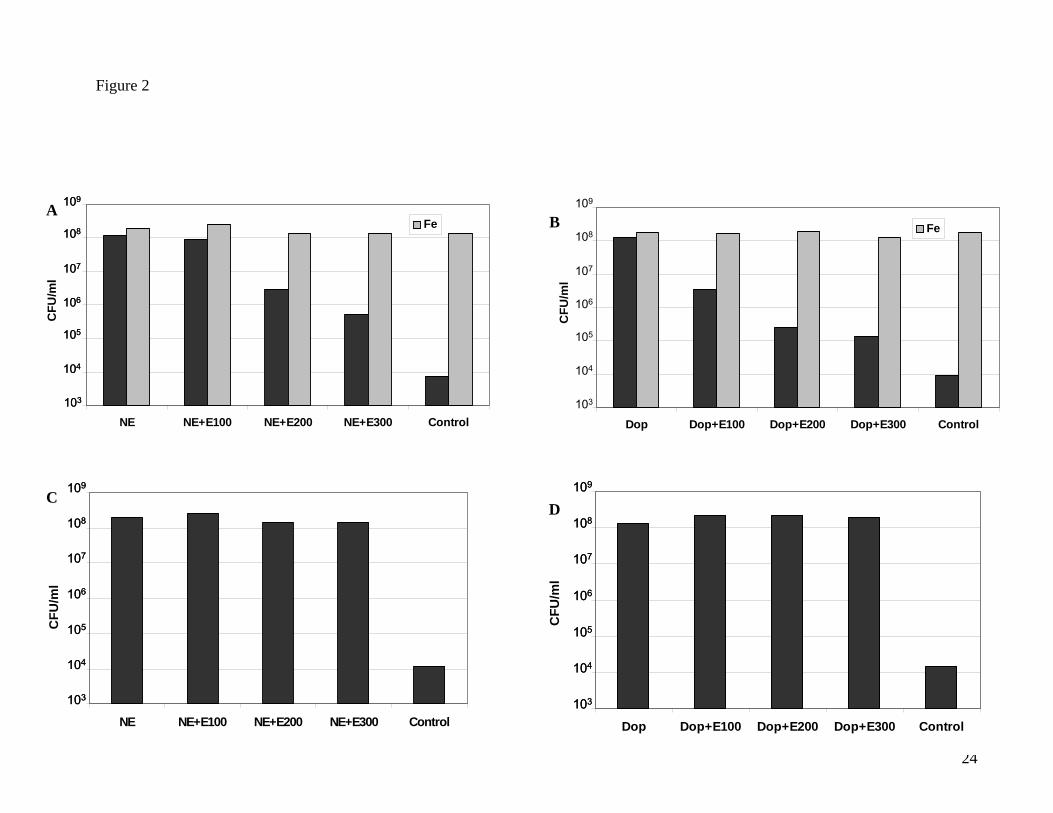
NE and Dop-mediated growth enhancement. Figures 2A and 2B show that addition of Epi to
serum-SAPI assays of Y. enterocolitica supplemented with either NE or Dop resulted in a
concentration-dependent inhibition of NE and Dop growth induction (P<0.0001). This
unexpected inhibitory action of Epi appears to be due to a specific antagonism of both NE
and Dop-responsiveness and not Epi cytotoxicity, since addition of 100 µM Fe(NO3)3 to Epi-
supplemented Y. enterocolitica NE and Dop cultures relieved the Epi-induced growth
inhibition (Figures 2A and 2B). In contrast, inclusion of Epi into NE and Dop supplemented
cultures of E. coli O157:H7 (Figures 2C and 2D) or S. enterica (data not shown) acted only to
enhance growth as earlier reported (Freestone, et al., 2002).
Previously, we demonstrated that NE is internalized by bacteria during the NE-growth
induction process (Freestone, et al., 2000), prompting us to investigate whether Epi was
antagonizing Y. enterocolitica responses to NE by blocking entry of NE into the bacterial cell.
We found that uptake of 3H-NE in high cell density cultures of Y. enterocolitica treated with
growth inhibitory concentrations of Epi as used in Figure 2A) (100, 200 and 300 μM Epi) was
not significantly reduced (P>0.05). Internalisation of 3H-NE in control cultures was 1356
CPM/ml and in cells treated with 100-300 μM Epi 1284, 1222 and 1470 CPM/ml respectively
(standard deviations for each set of cultures were 64, 55, 70 and 90 CPM/ml) We also
analysed whether Epi might reduce uptake of 3H-NE in a active growth context using a low
cell density initial inoculum (around 102 CFU per ml) combined with 24 hrs incubation (to
9

enable the culture to grow to a cell density similar to that employed in the high cell density
uptake assay). Using this approach, we overall found slightly more 3H-NE internalisation
(around 2000 CPM/ml), but once again observed no significant effects of Epi on 3H-NE
cellular incorporation. We did find that Y. enterocolitica needs to be metabolically active to
assimilate NE, as a high cell density suspension of bacteria treated with 6 mM sodium azide
internalised very low levels of 3H-NE (approximately 50 CPM/ml of culture)). Similarly low cell-
associated levels of 3H-NE were also observed with heat-killed bacteria (around 40 CPM/ml).
Fractionation of 3H-NE-labelled bacteria into cytoplasm and membrane fractions also
revealed that over 95 % of the 3H-NE associated with the cell was cytoplasmic/periplasmic in
location (data not shown).
The data shown in Figures 1 and 2 illustrating Y. enterocolitica growth responses to
Epi were obtained using low cell density inocula, (around 102 CFU per ml). A previous report
(Walters & Sperandio, 2006) demonstrating apparent Epi specificity in E. coli O157:H7 used
very high cell density cultures (around 108 CFU per ml) that are not reflective of actual in vivo
infective doses (Tarr & Neill, 2001), causing us to question whether increasing the Y.
enterocolitica population density would affect its responsiveness to Epi. Because Y.
enterocolitica does not show growth responsiveness to Epi at low cell numbers, we chose a
high cell density iron-uptake assay to investigate this possibility since previous work from our
laboratories had demonstrated that Epi, NE and Dop stimulated bacterial growth via their
ability to facilitate iron removal from the host iron binding proteins Tf and lactoferrin
(Freestone, et al., 2000; Neal, et al., 2001; Freestone, et al., 2002; Lyte, et al., 2003). Table 1
shows that at higher cell densities (108 CFU per ml), Y. enterocolitica responds to Epi in the
same manner as E. coli and S. enterica, specifically utilizing it to access Fe bound to Tf. For
all 3 bacterial species increasing the concentration of Epi resulted in enhancement of 55Fe
10

incorporation, a trend also observed when cultures were supplemented with higher
concentrations of NE and Dop. This striking reversal in the response of Y. enterocolitica to
Epi suggests that the effect of Epi on Y. enterocolitica physiology may vary between cultures
of different population densities.
Catecholamine specificity in E. coli O157:H7, S. enterica and Y. enterocolitica is
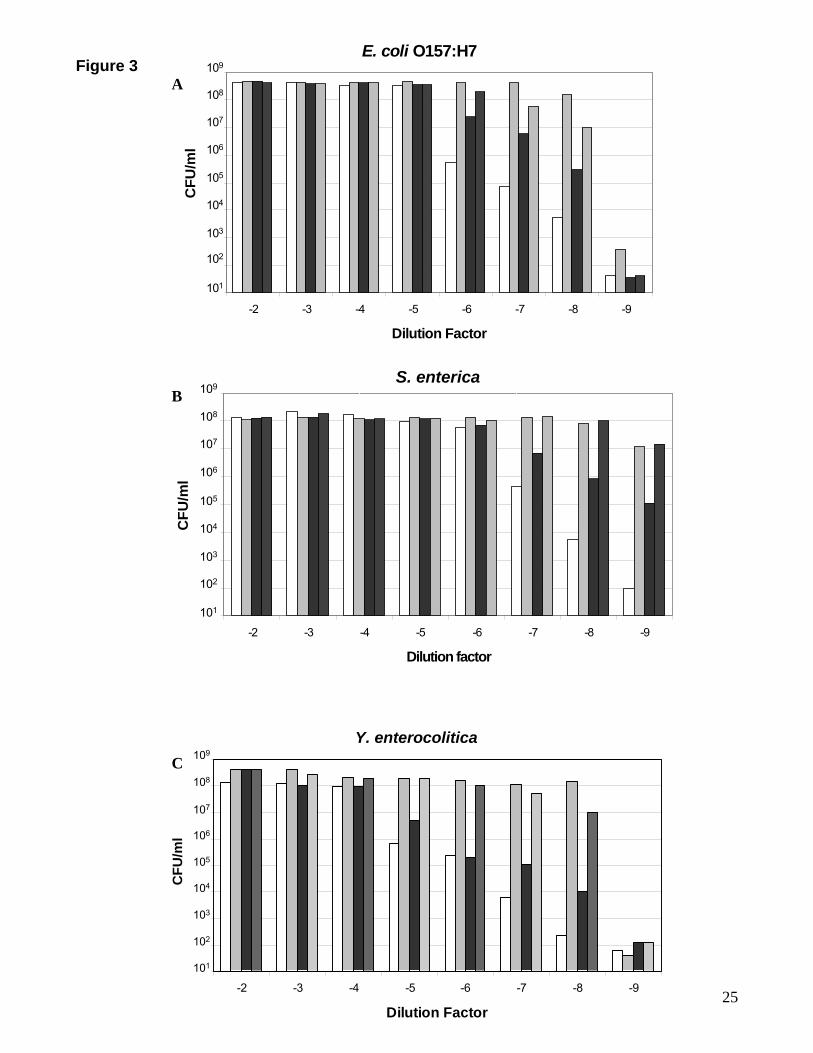
dependent on population density
To determine if bacterial population density does influence the specificity of catecholamine
responsiveness, we examined growth of E. coli O157:H7, S. enterica and Y. enterocolitica
over an 8-log dilution curve using concentrations of Dop, NE and Epi optimal for growth
induction (Figure 1). Figure 3 shows the ability of the catecholamines to affect bacterial
growth in a serum-based medium is initially evident at low (<104 CFU per ml) cell densities
with the greatest differences observed at very low (<102 CFU per ml) cell densities. Although
Figure 3 shows that both E. coli O157:H7 and S. enterica are able to respond to Epi, at very
low population densities, which are reflective of the bacterial numbers that are likely to be
present at the initial stages of an infection, there exists an order of catecholamine preference
with growth responses to NE and Dop being at least a log-fold greater than those to Epi
(P<0.0001).
11

Discussion
The lack of Y. enterocolitica growth responsiveness to Epi is of great interest in that it
is impossible to reconcile this observation with a model of growth induction where
catecholamines are merely siderophore-like in their interaction with Tf/lactoferrin to provide
iron for bacterial growth. Intriguingly, the results shown in Figure 2 demonstrated that not
only did Epi fail to induce growth but that it could also antagonize the growth inducing effects
of both NE and Dop in Y. enterocolitica. To investigate how Epi was blocking NE and Dop
responsiveness in Y. enterocolitica, we used a high cell density iron uptake assay and found
that Y. enterocolitica could use Epi in a manner similar to that employed by E. coli O157:H7
and S. enterica, specifically, to mediate access to iron from Tf (Table 1). Indeed, Epi
concentrations markedly inhibitory to NE and Dop growth induction of Y. enterocolitica were
now able to significantly enhance acquisition of iron from Tf. Using 3H-NE we also found that
Epi does not block NE uptake thereby indicating that the inhibitory action of Epi is also not at
the level of uptake of catecholamines. Furthermore, the evidence from Epi antagonism of
both NE and Dop growth induction demonstrates that, in Y. enterocolitica at least, the
response pathways to NE and Dop must possess a common element that can be blocked by
Epi. This data also has experimental design implications; it suggests that, in the context of
analysis of microbe:host catecholamine interactions, caution should be exercised before
assuming that conclusions reached from in vitro observations of responses of high density
cultures can be directly extrapolated to the low population numbers typically present during
the initial stage of a foodborne infection (Tarr & Neill, 2001). Our data also indicates that the
bacterial growth response to catecholamines involves more than the simple provision of host-
derived iron, and suggests that a specific response pathway(s) may exist for each
catecholamine.
12

Importantly, the primary anatomical site of production and degradation of NE and Dop
in the body are the mesenteric organs which account for over 50% of the whole body total
(Eisenhofer, et al., 1996). While the level of catecholamines information is not available
within the enteric neuroepithelial synapses of the gastrointestinal tract, making it difficult to
estimate appropriate in vitro concentrations that are comparable to those that could exist in
vivo. Additionally, the use of plasma concentrations of catecholamines is not valid as
concentrations in the plasma reflect spillover from the tissues and therefore grossly
underestimate the local production that might be present within any specific target organ
(Leinhardt, et al., 1993). Nevertheless, it is emphasized that the level of catecholamines
tested in the bacterial growth studies reported in the present report are similar to those used
in other fields such as neurochemistry [e.g., to demonstrate the phosphorylation of synapsin I
by NE in the rat frontal cortex (Mobley & Greengard, 1985)] and in immunology [(e.g. the
identification of β-adrenergic receptor mediation of antibody production ((Sanders & Munson,
1984) and the inhibition of gamma interferon synthesis (Andrade-Mena, 1997)]. As the
gastrointestinal tract is also the principal anatomical site in which bacteria can routinely come
into contact with endogenously produced catecholamines (Eisenhofer, et al., 1996), the
convergence of these two systems, one neuroendocrine, one bacterial, suggests that an
interrelationship may exist that can contribute to the development of enteric bacterial disease.
What does the lack of Epi responsiveness in Y. enterocolitica, which is almost
exclusively an enteric pathogen with little invasion of extra-intestinal sites, tell us about the
potential role of neuroendocrine hormones in the infectious disease process in the gut?
While the presence of adrenergic and dopaminergic containing neurons has been well
demonstrated (Costa, et al., 2000) within the ENS, as well as local production of NE and Dop
13

(Meirieu, et al., 1986), there have been no reports of Epi or Epi-containing neurons. It has
been proposed (Walters & Sperandio, 2006) that Epi is the key host-derived hormonal signal
in the pathogenesis of enteric pathogens such as E. coli O157:H7; however if Epi were
present in concentrations equivalent to those of NE and Dop, our observation that Epi
antagonizes Y. enterocolitica growth responsiveness to these catecholamines would suggest
that this species should have significant problems in growing within the gut environment,
which it clearly does not. A recent report (Clarke, et al., 2006) which used in vitro constructs
to demonstrate that NE and Epi could bind to the E. coli O157:H7 two component regulator
sensor kinase QseC, proposed that this is the bacterial receptor for these catecholamines.
Interestingly, Y. enterocolitica (in common with most other pathogenic Yersinia species) does
not contain homologs for QseC (or the related receptor QseE). Indeed, BLAST searches for
matches to QseC/QseE in the microbial genome database revealed that of the Yersinia
species only Y. mollaretii possesses the qseC gene. If QseC does not exist in Y.
enterocolitica our data indicates that a different signal transduction system specific for NE
must exist in Y. enterocolitica.
Although it has been suggested (Walters & Sperandio, 2006) that sufficient quantities
of Epi could spill over from the circulatory system and somehow transverse the physical and
biological barriers separating the gut lumen from the host to affect bacteria, there are
currently no published clinical or animal studies of enteric neurophysiology to support this
assertion. Norepinephrine was the most potent catecholamine growth stimulator for all of the
three bacterial species examined, and we therefore further speculate that it is NE, rather than
Epi, that is likely to be the cross-communicating adrenergic signal molecule between host and
enteric pathogens and that the bacterial response to Epi observed in E. coli O157:H7
(Walters & Sperandio, 2006) is more likely related to its structural similarity to NE.
14
Acknowledgements
This work was supported by grant 064488/Z/01/Z from the Wellcome Trust (to P.P.E.F.) and
NIH grant MH-50431 (to M.L.).
15

References
Andrade-Mena CE (1997) Inhibition of gamma interferon synthesis by catecholamines. J
Neuroimmunol 76: 10-14.
Belay T & Sonnenfeld G (2002) Differential effects of catecholamines on in vitro growth of
pathogenic bacteria. Life Sci 71: 447-456.
Belay T, Aviles H, Vance M, Fountain K & Sonnenfeld G (2003) Catecholamines and in vitro
growth of pathogenic bacteria: enhancement of growth varies greatly among bacterial
species. Life Sci 73: 1527-1535.
Burton CL, Chhabra SR, Swift S, Baldwin TJ, Withers H, Hill SJ & Williams P (2002) The
growth response of Escherichia coli to neurotransmitters and related catecholamine drugs
requires a functional enterobactin biosynthesis and uptake system. Infect Immun 70: 5913-
5923.
Clarke MB, Hughes DT, Zhu C, Boedeker EC & Sperandio V (2006) The QseC sensor
kinase: a bacterial adrenergic receptor. Proc Natl Acad Sci U S A 103: 10420-10425.
Costa M, Brookes SJ & Hennig GW (2000) Anatomy and physiology of the enteric nervous
system. Gut 47 Suppl 4: iv15-19; discussion iv26.
Eisenhofer G, Aneman A, Hooper D, Rundqvist B & Friberg P (1996) Mesenteric organ
production, hepatic metabolism, and renal elimination of norepinephrine and its metabolites in
humans. J Neurochem 66: 1565-1573.
Freestone PP, Haigh RD, Williams PH & Lyte M (1999) Stimulation of bacterial growth by
heat-stable, norepinephrine-induced autoinducers. FEMS Microbiol Lett 172: 53-60.
Freestone PP, Haigh RD, Williams PH & Lyte M (2003) Involvement of enterobactin in
norepinephrine-mediated iron supply from transferrin to enterohaemorrhagic Escherichia coli.
FEMS Microbiol Lett 222: 39-43.
16

Freestone PP, Lyte M, Neal CP, Maggs AF, Haigh RD & Williams PH (2000) The mammalian
neuroendocrine hormone norepinephrine supplies iron for bacterial growth in the presence of
transferrin or lactoferrin. J Bacteriol 182: 6091-6098.
Freestone PP, Williams PH, Haigh RD, Maggs AF, Neal CP & Lyte M (2002) Growth
stimulation of intestinal commensal Escherichia coli by catecholamines: a possible
contributory factor in trauma-induced sepsis. Shock 18: 465-470.
Kinney KS, Austin CE, Morton DS & Sonnenfeld G (1999) Catecholamine enhancement of
Aeromonas hydrophila growth. Microb Pathog 26: 85-91.
Kinney KS, Austin CE, Morton DS & Sonnenfeld G (2000) Norepinephrine as a growth
stimulating factor in bacteria--mechanistic studies. Life Sci 67: 3075-3085.
Leinhardt DJ, Arnold J, Shipley KA, Mughal MM, Little RA & Irving MH (1993) Plasma NE
concentrations do not accurately reflect sympathetic nervous system activity in human sepsis.
Am J Physiol 265: E284-288.
Lyte M (1992) The role of catecholamines in gram-negative sepsis. Med Hypotheses 37: 255-
258.
Lyte M (1993) The role of microbial endocrinology in infectious disease. J Endocrinol 137:
343-345.
Lyte M (2004) Microbial endocrinology and infectious disease in the 21st century. Trends
Microbiol 12: 14-20.
Lyte M & Ernst S (1992) Catecholamine induced growth of gram negative bacteria. Life Sci
50: 203-212.
Lyte M, Freestone PP, Neal CP, Olson BA, Haigh RD, Bayston R & Williams PH (2003)
Stimulation of Staphylococcus epidermidis growth and biofilm formation by catecholamine
inotropes. Lancet 361: 130-135.
17

Meirieu O, Pairet M, Sutra JF & Ruckebusch M (1986) Local release of monoamines in the
gastrointestinal tract: an in vivo study in rabbits. Life Sci. 38: 827-834.
Mobley P & Greengard P (1985) Evidence for widespread effects of noradrenaline on axon
terminals in the rat frontal cortex. Proc Natl Acad Sci U S A 82: 945-947.
Neal CP, Freestone PP, Maggs AF, Haigh RD, Williams PH & Lyte M (2001) Catecholamine
inotropes as growth factors for Staphylococcus epidermidis and other coagulase-negative
staphylococci. FEMS Microbiol Lett 194: 163-169.
Reissbrodt R, Rienaecker I, Romanova JM, et al. (2002) Resuscitation of Salmonella enterica
serovar typhimurium and enterohemorrhagic Escherichia coli from the viable but
nonculturable state by heat-stable enterobacterial autoinducer. Appl Environ Microbiol 68:
4788-4794.
Sanders VM & Munson AE (1984) Beta adrenoceptor mediation of the enhancing effect of
norepinephrine on the murine primary antibody response in vitro. J Pharmacol Exp Ther 230:
183-192.
Tarr PI & Neill MA (2001) Escherichia coli O157:H7. Gastroenterol Clin North Am 30: 735-
751.
Walters M & Sperandio V (2006) Quorum sensing in Escherichia coli and Salmonella. Int J
Med Microbiol 296: 125-131.
18

Figure legends
Figure 1. Dose-response effect of catecholamines on E. coli O157:H7 and Y.
enterocolitica.
E. coli O157:H7 and Y. enterocolitica NCTC strain 551 were inoculated at approximately 102
CFU per ml into duplicate 1-mL aliquots of serum-SAPI containing the concentrations of the
catecholamines shown and incubated for either 40 hours (Y. enterocolitica) or 18 hours (E.
coli O157:H7), and enumerated for growth (CFU per ml) as described in Materials and
methods. NE (grey bar), Dop (white bar), Epi (black bar). The results shown are
representative data from two separate experiments; data points showed variation of less than
3%. Results similar to those shown for E. coli O157:H7 were also obtained for S. enterica
(data not shown).
Figure 2. Ability of epinephrine to inhibit Y. enterocolitica growth induction by
norepinephrine and dopamine.
Y. enterocolitica NCTC strain 551 (panels A and B) and E. coli O157:H7 (panels C and D)
were inoculated at approximately 102 CFU per ml into duplicate 1 mL aliquots of serum-SAPI
containing the combination of catecholamines shown, and incubated for either 40 hours (Y.
enterocolitica) or 18 hours (E. coli O157:H7), and enumerated for growth (CFU per ml) as
described in Materials and Methods. The results shown are representative data from four
separate experiments; data points typically showed variation of no more than 3%; similar
result to those shown for Y. enterocolitica NCTC strain 551 were also seen with a Y.
enterocolitica clinical isolate and NCTC 1477 (data not shown).
Black bar, catecholamine/catecholamine combination only; light gray bar,
catecholamine/catecholamine combinations plus 100 μM Fe(NO3)3.
19

NE, 50 μM NE; NE+E100, 50 μM NE plus 100 μM Epi; NE+E200, 50 μM NE plus 200 μM Epi;
NE+E300, 50 μM NE plus 300 μM Epi; Dop, 50 μM Dop; Dop+E100, 50 μM Dop plus 100 μM
Epi; D+E200, 50 μM Dop plus 200 μM Epi; Dop+E300, 50 μM Dop plus 300 μM Epi. The
results shown are representative data from three separate experiments; data points typically
showed variation of no more than 3%.
Figure 3.
Bacterial population density influences catecholamine specificity.
Histograms A-C show the effects of varying inoculum size on the specificity of growth
response to catecholamines NE, Epi and Dop. Cultures were diluted in 10-fold steps into
serum-SAPI medium, incubated for either 18 hours (E. coli O157:H7 and S. enterica) or 40
hours (Y. enterocolitica NCTC 551) (and NCTC 1477, data not shown), and enumerated for
growth (CFU per ml) as described in Materials and Methods. The inoculum size of the E. coli
O157:H7 culture (panel A) was 5.29 x 108 CFU per ml; S. enterica (panel B) 2.84 x 108 CFU
per ml and Y. enterocolitica (panel C) 4.44 x 108 CFU per ml. The results shown are
representative data from four separate experiments; individual data points showed variation
of no more than 5%.
White bar, no additions (control); light grey bar, 50 µM NE; black bar, 100 µM Epi; diagonal
hatch, 50 µM Dop.
20
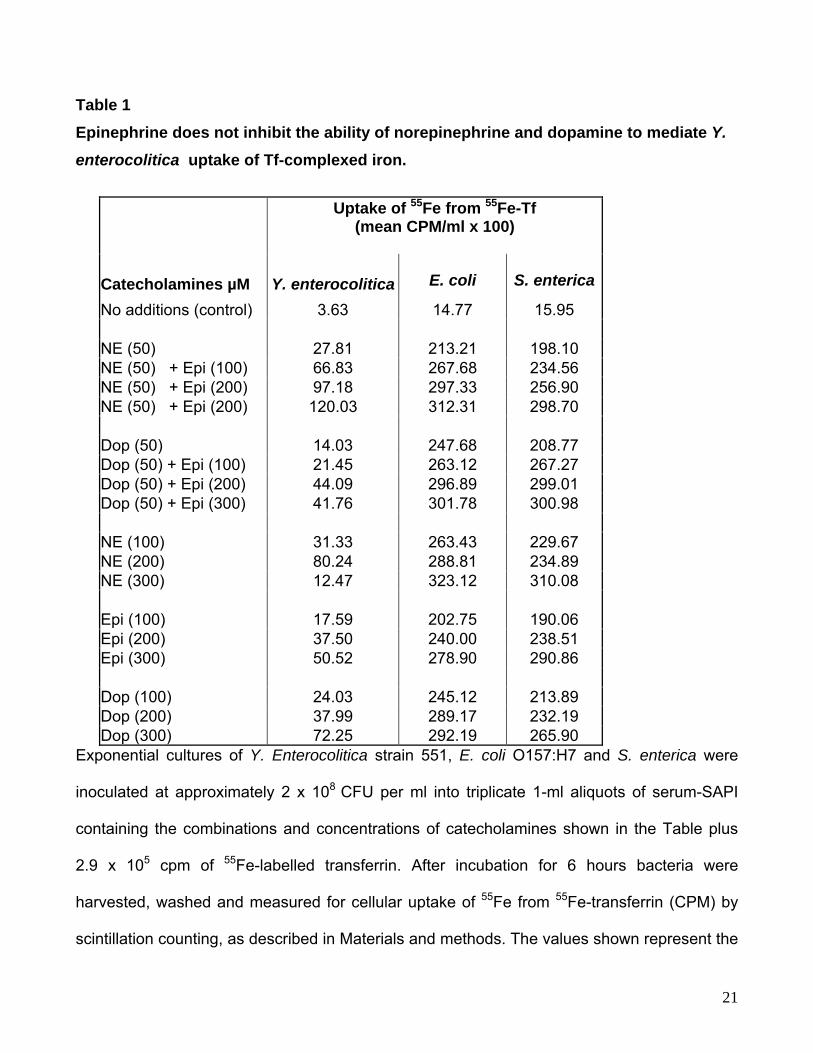
Table 1 Epinephrine does not inhibit the ability of norepinephrine and dopamine to mediate Y.
enterocolitica uptake of Tf-complexed iron.
Uptake of 55Fe from 55Fe-Tf (mean CPM/ml x 100)
Catecholamines µM Y. enterocolitica E. coli S. enterica
No additions (control) 3.63 14.77 15.95 NE (50) 27.81 213.21 198.10 NE (50) + Epi (100) 66.83 267.68 234.56 NE (50) + Epi (200) 97.18 297.33 256.90 NE (50) + Epi (200) 120.03 312.31 298.70 Dop (50) 14.03 247.68 208.77 Dop (50) + Epi (100) 21.45 263.12 267.27 Dop (50) + Epi (200) 44.09 296.89 299.01 Dop (50) + Epi (300) 41.76 301.78 300.98 NE (100) 31.33 263.43 229.67 NE (200) 80.24 288.81 234.89 NE (300) 12.47 323.12 310.08 Epi (100) 17.59 202.75 190.06 Epi (200) 37.50 240.00 238.51 Epi (300) 50.52 278.90 290.86 Dop (100) 24.03 245.12 213.89 Dop (200) 37.99 289.17 232.19 Dop (300) 72.25 292.19 265.90
Exponential cultures of Y. Enterocolitica strain 551, E. coli O157:H7 and S. enterica were
inoculated at approximately 2 x 108 CFU per ml into triplicate 1-ml aliquots of serum-SAPI
containing the combinations and concentrations of catecholamines shown in the Table plus
2.9 x 105 cpm of 55Fe-labelled transferrin. After incubation for 6 hours bacteria were
harvested, washed and measured for cellular uptake of 55Fe from 55Fe-transferrin (CPM) by
scintillation counting, as described in Materials and methods. The values shown represent the
21

means of bacterial 55Fe incorporation from triplicate 1 ml uptake assays; standard deviations
were between 1 and 6 % of the mean values shown in the table.. Similar results to those
shown for NCTC strain 551 were also observed with Y. enterocolitica NCTC strain 1477 (data
not shown). Analysis of cell numbers revealed no significant differences in growth levels
between control and catecholamine/antagonist supplemented cultures (data not shown).
22
23
E. coli O157:H7
1.E
1.E
1.E
1.E+06
1.E+07
1.E+08
.E+09
0 1 5 10 20 50 100 200 500
Concentration μM
+03
+04
+05
1 109
108
107
106
105
104
103
CFU
/ml
E. coli O157:H7E. coli O157:H7
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
.E+09
0 1 5 10 20 50 100 200 500
Concentration μM
1 109
108
107
106
105
104
103
CFU
/ml
E. coli O157:H7
Y. enterocolitica
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
1.E+09
0 1 5 10 20 50 100 200 500
Concentration μM
109
108
107
106
105
104
103
CFU
/ml
Y. enterocoliticaY. enterocolitica
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
1.E+09
0 1 5 10 20 50 100 200 500
Concentration μM
109
108
107
106
105
104
103
CFU
/ml
Y. enterocolitica
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
1.E+09
0 1 5 10 20 50 100 200 500
Concentration μM
109
108
107
106
105
104
103
CFU
/ml
109
108
107
106
105
104
103
109
108
107
106
105
104
103
CFU
/ml
Y. enterocolitica
Figure 1
A
B
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+07
24
1.0E+08
1.0E+09
Dop Dop+E100 Dop+E200 Dop+E300 Control
Fe
109
108
107
106
105
104
103
CFU
/ml
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+07
0E+08
1.0E+09
Dop Dop+E100 Dop+E200 Dop+E300 Control
1.Fe
109
108
107
106
105
104
1031.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+07
0E+08
1.0E+09
Dop Dop+E100 Dop+E200 Dop+E300 Control
1.Fe
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+07
1. +08
1.0E+09
Dop Dop+E100 Dop+E200 Dop+E300 Control
109
108
107
106
105
104
103
CFU
/ml
1.
1.
1.
1.
1.
1.
1.
E+03
E+04
E+05
E+06
E+07
E+08
E+09
NE NE+E100 NE+E200 NE+E300 Control
109
108
107
106
105
104
103
CFU
/ml
1.
1.
1.
1.
1.
1.
1.
E+03
E+04
E+05
E+06
E+07
E+08
E+09
NE NE+E100 NE+E200 NE+E300 Control
109
108
107
106
105
104
103
CFU
/ml
109
108
107
106
105
104
103
109
108
107
106
105
104
103
CFU
/ml
0E
109
108
107
106
105
104
103
CFU
/ml
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+07
1. +08
1.0E+09
Dop Dop+E100 Dop+E200 Dop+E300 Control
0E
109
108
107
106
105
104
103
109
108
107
106
105
104
103
CFU
/ml
B
D
Figure 2
1.0E
1.0E
1.0E
1.0E
1.0E
1.0E
1.0E
+03
+04
+05
+06
+07
+08
+09
NE NE+E100 NE+E200 NE+E300 Control
Fe
109
108
107
106
105
104
103
CFU
/ml
1.0E
1.0E
1.0E
1.0E
1.0E
1.0E
1.0E 09
NE NE+E100 NE+E200 NE+E300 Control+03
+04
+05
+06
+07
+08Fe
+109
108
107
106
105
104
1031.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+07
1.0E+08
1.0E 09
NE NE+E100 NE+E200 NE+E300 Control
Fe
+109
108
107
106
105
104
103
109
108
107
106
105
104
103
CFU
/ml
A
C
25
S. enterica
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
1.E+09
-2 -3 -4 -5 -6 -7 -8 -9
Dilution factor
109
108
107
106
105
104
103
102
101
S. enterica109
108
107
106
105
104
103
102
101
CFU
/ml
S. enterica
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
E+09
-2 -3 -4 -5 -6 -7 -8 -9
Dilution factor
1. 109
108
107
106
105
104
103
102
101
109
108
107
106
105
104
103
102
101
S. enterica
E. coli O157:H7
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
E+09
-2 -3 -4 -5 -6 -7 -8 -9
Dilution Factor
109
108
107
106
105
104
103
102
101
CFU
/ml
1.E. coli O157:H7
109
108
107
106
105
104
103
102
101
CFU
/ml
E. coli O157:H7
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
E+09
-2 -3 -4 -5 -6 -7 -8 -9
Dilution Factor
1.E. coli O157:H7
109
108
107
106
105
104
103
102
101
Figure 3 109
108
107
106
105
104
103
102
101
CFU
/ml
A
B
Y. enterocolitica
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
1 09
-2 -3 -4 -5 -6 -7 -8 -9
Dilution Factor
.E+
Y. enterocolitica109
108
107
106
105
104
103
102
101
CFU
/ml
Y. enterocolitica
1.E+01
1.E+02
1.E+03
1.E+04
1.E+05
1.E+06
1.E+07
1.E+08
1 09
-2 -3 -4 -5 -6 -7 -8 -9
Dilution Factor
.E+
Y. enterocolitica
109
108
107
106
105
104
103
102
101
CFU
/ml
C
Related Documents











