J. Anat. (1998) 192, pp. 1–12, with 8 figures Printed in the United Kingdom 1 Species-specificity of growth-promoting effects of prolactin during rat embryogenesis AHMET KAGAN KARABULUT 1 AND MARGARET K. PRATTEN 2 " Department of Anatomy, University of Selcuk, Konya, Turkey and # Department of Human Anatomy and Cell Biology, Queen’s Medical Centre, Nottingham, UK (Accepted 29 July 1997) In the early stages of embryonic development, many growth-promoting molecules must be provided by the maternal system. The molecules involved in growth processes may be either hormones or growth factors, or molecules that interact with such factors. The pregnancy related hormone, prolactin (PRL, MW 23 kDa) has been implicated in the control of embryonic growth. The growth-promoting potential of PRL and its species-specificity was investigated by culturing 9–5 d rat embryos in vitro for 48 h in depleted serum in the presence and absence of PRL from 3 different species. The growth-supporting capacity of the serum was reduced by removal of low molecular weight molecules by prolonged filtration of the serum using filters with a molecular weight exclusion of 30 kDa. This method provided a ‘ semidefined ’ medium (retenate) in which embryonic growth and development was significantly reduced, demonstrating that the low molecular weight fraction of serum may contain some growth-promoting factors. Addition of PRL (0–4–25–6 ng}ml) from different species (human, sheep and rat) to retenate significantly improved embryonic growth and development, suggesting that the developing embryo may utilise PRL. Amongst PRLs, rat PRL was found to be active at much lower concentrations than either of the other molecules, and human PRL had more effect in low concentrations than sheep PRL suggesting a species-specificity for this hormone. It may be that the PRL receptors of the rat embryos have greater affinity for the rat hormone as different responses for hormones from different species have been shown. These findings suggest that embryos may be able to utilise maternally derived PRL during organogenesis. Key words : Whole embryo culture ; rat ; organogenesis ; development. It has been well known that a variety of hormones play an important role in the development of the mammalian embryo, from the moment of conception. The hormonal milieu which bathes the embryo is the product of a maternal–placental–embryonic inter- action. The hormones that play a critical role in the regulation of postnatal growth, such as growth hormone, thyroid hormone, glucocorticoids and sex steroids, appear to be less important in embryonic and fetal growth, since the absence or deficiency of these hormones is not accompanied by fetal growth failure (Freemark & Handwerger, 1989). Since growth hor- Correspondence to Dr Margaret K. Pratten, Department of Human Anatomy and Cell Biology, Queen’s Medical Centre, Nottingham, NG7 2UH, UK. Tel: ›44-115 9709429 ; fax : ›44-115 9709732 ; e-mail : Margaret.Pratten!nottingham.ac.uk mone (GH) appears to be of minimal importance for the stimulation of embryonic and fetal growth, GH- like peptides synthesised by the placenta such as placental lactogen (PL) and prolactin (PRL) have been postulated to play roles in the regulation of embryonic and fetal growth processes due to their circulation in fetal serum in mid-late gestation (Ogren & Talamantes, 1988), direct metabolic effects in fetal tissues (Freemark & Handwerger, 1984 a, b) and receptor expression in embryonic and fetal tissues (Underwood & D’Ercole, 1984 ; Hill et al. 1988 ; Hill, 1992 ; Fowlkes & Freemark, 1993 ; Freemark et al. 1993). However, the role of these hormones in early embryonic development is still not clear.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J. Anat. (1998) 192, pp. 1–12, with 8 figures Printed in the United Kingdom 1
Species-specificity of growth-promoting effects of prolactin
during rat embryogenesis
AHMET KAGAN KARABULUT1 AND MARGARET K. PRATTEN2
"Department of Anatomy, University of Selcuk, Konya, Turkey and #Department of Human Anatomy and Cell Biology,
Queen’s Medical Centre, Nottingham, UK
(Accepted 29 July 1997)
In the early stages of embryonic development, many growth-promoting molecules must be provided by the
maternal system. The molecules involved in growth processes may be either hormones or growth factors, or
molecules that interact with such factors. The pregnancy related hormone, prolactin (PRL, MW 23 kDa) has
been implicated in the control of embryonic growth. The growth-promoting potential of PRL and its
species-specificity was investigated by culturing 9±5 d rat embryos in vitro for 48 h in depleted serum in the
presence and absence of PRL from 3 different species. The growth-supporting capacity of the serum was
reduced by removal of low molecular weight molecules by prolonged filtration of the serum using filters with
a molecular weight exclusion of 30 kDa. This method provided a ‘semidefined’ medium (retenate) in which
embryonic growth and development was significantly reduced, demonstrating that the low molecular weight
fraction of serum may contain some growth-promoting factors. Addition of PRL (0±4–25±6 ng}ml) from
different species (human, sheep and rat) to retenate significantly improved embryonic growth and
development, suggesting that the developing embryo may utilise PRL. Amongst PRLs, rat PRL was found
to be active at much lower concentrations than either of the other molecules, and human PRL had more
effect in low concentrations than sheep PRL suggesting a species-specificity for this hormone. It may be that
the PRL receptors of the rat embryos have greater affinity for the rat hormone as different responses for
hormones from different species have been shown. These findings suggest that embryos may be able to
utilise maternally derived PRL during organogenesis.
Key words : Whole embryo culture ; rat ; organogenesis ; development.
It has been well known that a variety of hormones
play an important role in the development of the
mammalian embryo, from the moment of conception.
The hormonal milieu which bathes the embryo is the
product of a maternal–placental–embryonic inter-
action. The hormones that play a critical role in the
regulation of postnatal growth, such as growth
hormone, thyroid hormone, glucocorticoids and sex
steroids, appear to be less important in embryonic and
fetal growth, since the absence or deficiency of these
hormones is not accompanied by fetal growth failure
(Freemark & Handwerger, 1989). Since growth hor-
Correspondence to Dr Margaret K. Pratten, Department of Human Anatomy and Cell Biology, Queen’s Medical Centre, Nottingham, NG7
2UH, UK. Tel : 44-115 9709429; fax: 44-115 9709732; e-mail : Margaret.Pratten!nottingham.ac.uk
mone (GH) appears to be of minimal importance for
the stimulation of embryonic and fetal growth, GH-
like peptides synthesised by the placenta such as
placental lactogen (PL) and prolactin (PRL) have
been postulated to play roles in the regulation of
embryonic and fetal growth processes due to their
circulation in fetal serum in mid-late gestation (Ogren
& Talamantes, 1988), direct metabolic effects in fetal
tissues (Freemark & Handwerger, 1984a, b) and
receptor expression in embryonic and fetal tissues
(Underwood & D’Ercole, 1984; Hill et al. 1988; Hill,
1992; Fowlkes & Freemark, 1993; Freemark et al.
1993). However, the role of these hormones in early
embryonic development is still not clear.

Several studies demonstrated the importance of the
hormone for fetal development. Treatment of preg-
nant sheep with bromocriptine, a well known an-
tagonist for PRL, reduces both maternal and fetal
PRL levels, resulting in decreased fetal weight (Taka-
hashi et al. 1979), thus suggesting that there is a
possible link between PRL and fetal growth. PRL has
also been postulated as an important embryological
and neonatal growth hormone (Nicoll, 1978; Under-
wood & D’Ercole, 1984). It has been suggested that
hormones with lactogenic activity play an important
role in the enhancement of insulin secretion by
pancreatic islets of Langerhans that occurs during
pregnancy. Recent in vitro studies have confirmed
that both PLs and PRLs, but not GHs, are potent
stimulators of both insulin secretion and beta cell
proliferation in pancreatic islets of Langerhans iso-
lated from mice, rat, and humans (Nielsen, 1982;
Sorenson et al. 1987; Brelje & Sorenson, 1988, 1991;
Brelje et al. 1989, 1993, 1994). The importance of
insulin for growth and development of the embryo
and fetus has been demonstrated (Travers et al. 1989,
1992).
Another possible mechanism for the effect of
prolactin may be in the regulation of fetal insulin-like
growth factor (IGF) concentrations during pregnancy
(Francis & Hill, 1975; Hill et al. 1977; Underwood &
D’Ercole, 1984). It is known that the IGFs are anabolic
hormones with both proliferative and differentiative
functions (D’Ercole et al. 1980; D’Ercole & Under-
wood, 1981; Adams et al. 1983a, b ; Hill et al. 1985;
Sara & Hall, 1990).
It is clear that PRL may be involved in the
regulation of embryonic and fetal growth. However,
apart from a limited study (Calvert, 1985; Calvert et
al. 1986), there are no published data observing the
direct effect of PRLs on embryonic growth and
development using the embryo culture system.
Prolactin is a single 23 kDa polypeptide chain of
199 amino acid residues with 3 intramolecular
disulphide bridges. The sequence identity of human
prolactin (hPRL) with PRL of sheep (sPRL), pig
(pPRL) and rat (rPRL) is 73, 77 and 60% respectively,
and it shows 16% homology to GH. Of the 199 amino
acid residues of prolactins 99 residues are identical in
sequence position in all 4 species (Basudev & Parlow,
1977). There is also 63% overall homology between
rat and sheep prolactin (Nicoll et al. 1986). Therefore,
considering the species-specificity, we investigated the
influence of the addition of wide range concentrations
of human (hPRL), sheep (sPRL) and rat (rPRL)
prolactin to depleted serum on early embryonic
growth and development. Depletion of serum was
achieved by prolonged filtration of serum in a
centrifugal concentrator, which has been shown to
reduce its growth-supporting capacity by removing
low molecular weight molecules (Karabulut &
Pratten, 1995). We have also shown that depletion
of whole rat serum by prolonged filtration does not
have any detrimental effect on serum components as
reconstitution of the retenate (high molecular weight
fraction) with the filtrate (low molecular weight
fraction) completely restored the growth supporting
capacity of the serum molecules (Karabulut & Pratten,
1995).
Chemicals
Rat prolactin was supplied by Biogenesis Ltd (Poole,
Dorset, UK). Human and sheep PRL, and all other
chemicals were supplied by Sigma Chemical Company
(Poole, Dorset, UK).
Animals
Wistar rats (Rattus norvegicus) were used in this study
and were obtained from the breeding colony of the
Queen’s Medical Centre, University of Nottingham.
Male and female rats were placed together in the
evening (1 pair per cage), and the presence of a vaginal
plug the following morning was taken to indicate
mating had occurred. The female rat would be
considered to be 0±5 d pregnant at noon on that day,
since copulation was assumed to have occurred within
2 h either side of midnight.
Explanation and the culture of embryos
Female Wistar rats were anaesthetised, using ether, on
d 9±5 and the conceptuses explanted according to a
standard method of embryo culture (New, 1978). The
uterus was quickly removed from pregnant rats and
transferred to a dish containing Hanks’ balanced salt
solution. It was then opened with jeweller’s forceps to
expose pear-shaped masses of decidual tissue which
were teased away from the contained egg cylinder
under the dissecting microscope. Reichert’s membrane
was removed, and the 9±5 d egg cylinders were placed
in a sterile glass bottle which contained 1 ml of
immediately centrifuged, heat-inactivated rat serum
per embryo (New, 1978). The bottles containing
medium were then stoppered and placed horizontally
on rollers in an incubator at 37 °C at 30–40 rev}min
for 48 h. Embryos were gassed immediately after
2 A. K. Karabulut and M. K. Pratten

explantation with a gas mixture containing 5% O#,
5% CO#, 90% N
#. After 24 h this was replaced with
20% O#, 5% CO
#, 75% N
#, and after 44 h with 40%
O#, 5% CO
#, 55% N
#. Embryos were harvested at
48 h.
Endpoint measurement
After 48 h of culture, the growth and development of
embryos was evaluated under a dissecting microscope
using the morphological scoring system of Van Maele-
Fabry et al. (1990). In this system the following
endpoint measurements were assessed. For growth,
crown-rump length was measured; for differentiation,
somite number was recorded; and for development,
the flexion of the embryos, as well as the development
of the fore, mid and hind brain, caudal neural tube,
optic, otic and olfactory systems, maxillary and
mandibular processes, branchial bars, and fore and
hind limbs were examined. In addition the size and
vascularisation of the yolk sac and development of
allantois were also recorded. Protein assay (Lowry et
al. 1951) was performed to assess the total protein
content of yolk sac and embryo, using bovine serum
albumin as reference protein.
Filtration of serum
Filtration of homologous heat inactivated serum was
achieved using a Macrosep centrifugal concentrator
with a nominal molecular weight exclusion of 30 kDa
(Filtron, Northborough, USA). Pooled serum (15 ml)
was added to each concentrator and centrifuged for a
total of 8 h at 3500 rpm at 4 °C, with 5 ml retained as
control serum for each experiment. After 2 h of
centrifugation the low molecular weight filtrate was
removed and stored at ®20 °C. The high molecular
weight fraction retained by the filter membrane
(retenate) was resuspended with distilled water to
15 ml to prevent it becoming too viscous to be filtered
efficiently. Serum was recentrifuged and the above
procedure repeated every 2 h. At the end of the
procedure the filtrate was removed and stored. During
the filtration retenate loses many of the low molecular
weight molecules it originally contained; therefore, it
has a very low osmolarity. The retenate was recon-
stituted with double-strength Hanks’ balanced salt
solution (600 mOsm) to a final volume of 15 ml. The
osmolarity of the retenate was then corrected to
within the normal range for whole rat serum (290–330
mOsm) with distilled water. Also the pH was adjusted
to 7±8 with 0±1 sodium hydroxide or 0±1 hy-
drochloric acid if required. Prior to use the retenate
was sterile filtered with a Minisart NML disposable
syringe filter holder, pore size 0±2 µm (Sartorius AG,
Go$ ttingen, Germany) and supplemented with 2 mg}ml glucose and 10 µl}ml minimal essential medium
(MEM) vitamin solution (Sigma, Poole, UK).
Experiments undertaken
In order to investigate the influence of PRL from
different species on mammalian embryonic growth
and development rat embryos were cultured in
retenate in the presence of human, sheep and rat PRL.
Human and sheep PRL were used at concentrations
between 0±4 and 25±6 ng}ml. In order to determine the
effect of rat PRL a wider range of concentrations
(0±1–25±6 ng}ml) of hormone was added to retenate.
In each experiment retenate (supplemented with
glucose and MEM vitamins) and whole rat serum
controls were included. At least 10 embryos were used
for each experimental condition. Embryos were then
cultured for 48 h with appropriate gassing, assessed
morphologically and embryonic and yolk sac protein
contents determined.
Statistics
Differences between experimental and control groups
were analysed statistically as follows. The yolk sac
diameter, crown–rump length and protein contents of
the embryos and yolk sacs were analysed using 1-way
analysis of variance (ANOVA) to determine the
presence of significant difference within the data, and
a parametric Duncan’s multiple range test was
performed in order to demonstrate where the exact
significant differences lay. Analysis of differences in
morphological score and somite number was per-
formed using a nonparametric Kruskal–Wallis test to
demonstrate the presence of a significant difference
within the data and this was followed by subsequent
Mann–Whitney U tests as the use of parametric tests
on such data was inappropriate. All tests were
performed on the statistics package, SPSS-X. Signi-
ficance was assumed at P! 0±05, P! 0±01, and
P! 0±001.
The embryos cultured in retenate supplemented with
glucose and vitamins showed severe growth retar-
dation in all scoring criteria (total morphological
Effects of prolactin during rat organogenesis 3

Fig. 1. Effects of supplementation with human PRL (0±4–25±6 ng}ml) on the growth supporting capacity of retenate. WRS, whole rat serum;
30 KR, retenate. Results are the mean of the values³... for at least 10 embryos. *, **, *** Significantly different from retenate control
values at P! 0±05, P! 0±01, P! 0±001.
4 A. K. Karabulut and M. K. Pratten

Fig. 2. 11±5 d rat embryos following 48 h culture (a) in normal rat
serum, (b) in serum depleted of low molecular weight molecules
(MW! 30 kDa). Note severe growth retardation in embryo grown
in retenate. Bars, 500 µm. Bb, branchial bars ; H, hindbrain; He,
heart ; F, forebrain; M, midbrain; O, optic system; Ot, otic system;
S, somites.
score, yolk sac diameter, crown–rump length, somite
number, protein contents of embryo and yolk sac)
compared with those cultured in whole rat serum
(Figs 1, 2).
The addition of hPRL to retenate caused an
increase in embryonic development when compared
with development of embryos cultured in retenate
supplemented with glucose and vitamins only (Fig. 1).
Morphological score and somite number were signifi-
cantly increased in the presence of 0±4 and 0±8 ng}ml
hPRL. Addition of higher levels (1±6–25±6 ng}ml) of
hPRL significally improved all parameters, especially
morphological score and somite number, and embryo
protein content when 6±4 ng}ml and higher concen-
Fig. 3. A left lateral view of an 11±5 d rat embryo following 48 h
culture in the presence of 12±8 ng}ml human PRL. Bar, 500 µm.
trations of hPRL were added (Fig. 3). Indeed, there
was no significant difference between the values
obtained for somite number at 12±8 ng}ml and for
yolk sac protein content at 1±6–25±6 ng}ml and those
observed in whole rat serum.
All embryos cultured in the presence of low
concentrations (0±4–1±6 ng}ml) of sPRL did not show
any significant difference in growth and development
in all parameters compared with retenate control
embryos. Significant improvement was seen when
retenate was supplemented with higher levels (3±2–25±6ng}ml) of sPRL in morphological score, somite
number, crown–rump length, embryo and yolk sac
protein contents, and yolk sac diameter when 3±2 and
12±8 ng}ml sPRL was added (Fig. 4). The extent of
growth and development of the embryos in this group
did not reach that of those grown in whole rat serum
at any concentration of sPRL (Fig. 5).
The outcome of all parameters of growth and
development was significantly improved on the ad-
dition of 0±4–25±6 ng}ml rPRL compared with ret-
enate only (Figs 6, 7). There was no significant
difference between the results obtained with rPRL-
supplemented retenate and those obtained for whole
rat serum for embryonic protein (0±4–3±2 ng}ml), yolk
sac protein (1±6–6±4 ng}ml), and somite number
(0±8–1±6 ng}ml). It was noticeable that at higher
concentrations both yolk sac and embryo protein
levels tended to be decreased. It was observed that the
growth-promoting effects of rPRL started at very low
concentrations and increased dose dependently (Fig.
8).
Effects of prolactin during rat organogenesis 5

Fig. 4. Effects of supplementation with sheep PRL (0±4–25±6 ng}ml) on the growth-supporting capacity of retenate. WRS, whole rat serum;
30 KR, retenate. Results are the mean of the values³... for at least 10 embryos. *, **, *** Significantly different from retenate control
values at P! 0±05, P! 0±01, P! 0±001.
6 A. K. Karabulut and M. K. Pratten
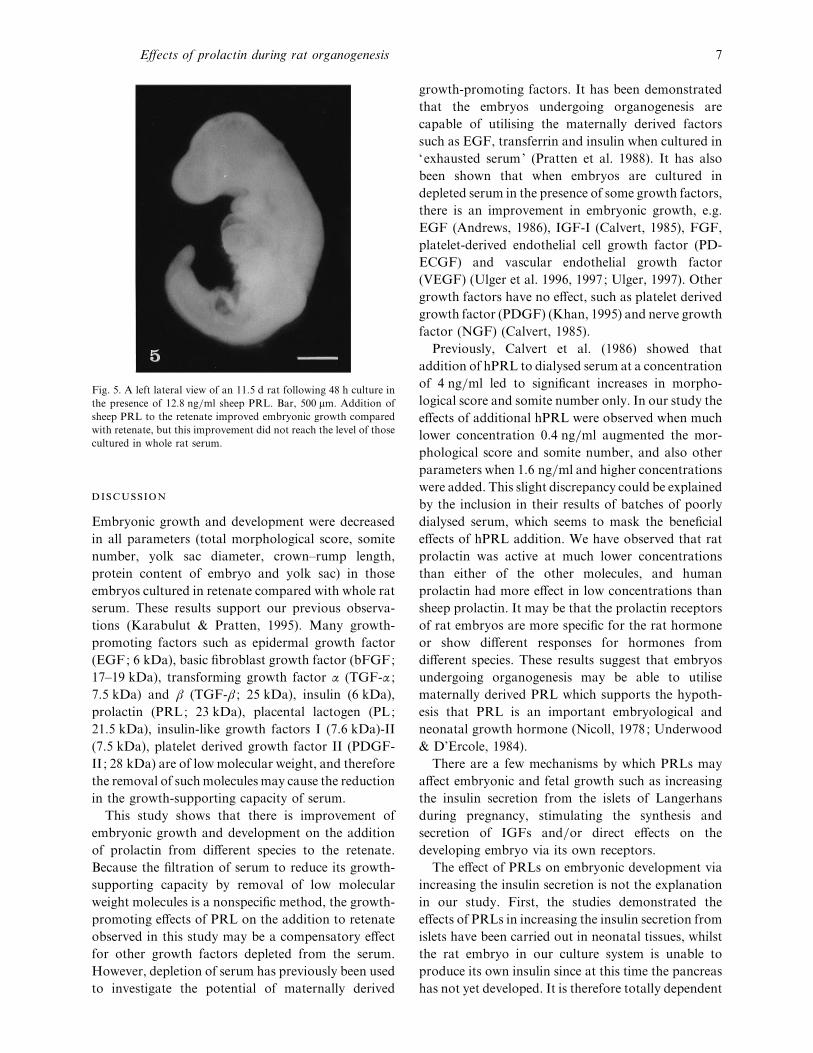
Fig. 5. A left lateral view of an 11±5 d rat following 48 h culture in
the presence of 12±8 ng}ml sheep PRL. Bar, 500 µm. Addition of
sheep PRL to the retenate improved embryonic growth compared
with retenate, but this improvement did not reach the level of those
cultured in whole rat serum.
Embryonic growth and development were decreased
in all parameters (total morphological score, somite
number, yolk sac diameter, crown–rump length,
protein content of embryo and yolk sac) in those
embryos cultured in retenate compared with whole rat
serum. These results support our previous observa-
tions (Karabulut & Pratten, 1995). Many growth-
promoting factors such as epidermal growth factor
(EGF; 6 kDa), basic fibroblast growth factor (bFGF;
17–19 kDa), transforming growth factor α (TGF-α ;
7±5 kDa) and β (TGF-β ; 25 kDa), insulin (6 kDa),
prolactin (PRL; 23 kDa), placental lactogen (PL;
21±5 kDa), insulin-like growth factors I (7±6 kDa)-II
(7±5 kDa), platelet derived growth factor II (PDGF-
II ; 28 kDa) are of low molecular weight, and therefore
the removal of such molecules may cause the reduction
in the growth-supporting capacity of serum.
This study shows that there is improvement of
embryonic growth and development on the addition
of prolactin from different species to the retenate.
Because the filtration of serum to reduce its growth-
supporting capacity by removal of low molecular
weight molecules is a nonspecific method, the growth-
promoting effects of PRL on the addition to retenate
observed in this study may be a compensatory effect
for other growth factors depleted from the serum.
However, depletion of serum has previously been used
to investigate the potential of maternally derived
growth-promoting factors. It has been demonstrated
that the embryos undergoing organogenesis are
capable of utilising the maternally derived factors
such as EGF, transferrin and insulin when cultured in
‘exhausted serum’ (Pratten et al. 1988). It has also
been shown that when embryos are cultured in
depleted serum in the presence of some growth factors,
there is an improvement in embryonic growth, e.g.
EGF (Andrews, 1986), IGF-I (Calvert, 1985), FGF,
platelet-derived endothelial cell growth factor (PD-
ECGF) and vascular endothelial growth factor
(VEGF) (Ulger et al. 1996, 1997; Ulger, 1997). Other
growth factors have no effect, such as platelet derived
growth factor (PDGF) (Khan, 1995) and nerve growth
factor (NGF) (Calvert, 1985).
Previously, Calvert et al. (1986) showed that
addition of hPRL to dialysed serum at a concentration
of 4 ng}ml led to significant increases in morpho-
logical score and somite number only. In our study the
effects of additional hPRL were observed when much
lower concentration 0±4 ng}ml augmented the mor-
phological score and somite number, and also other
parameters when 1±6 ng}ml and higher concentrations
were added. This slight discrepancy could be explained
by the inclusion in their results of batches of poorly
dialysed serum, which seems to mask the beneficial
effects of hPRL addition. We have observed that rat
prolactin was active at much lower concentrations
than either of the other molecules, and human
prolactin had more effect in low concentrations than
sheep prolactin. It may be that the prolactin receptors
of rat embryos are more specific for the rat hormone
or show different responses for hormones from
different species. These results suggest that embryos
undergoing organogenesis may be able to utilise
maternally derived PRL which supports the hypoth-
esis that PRL is an important embryological and
neonatal growth hormone (Nicoll, 1978; Underwood
& D’Ercole, 1984).
There are a few mechanisms by which PRLs may
affect embryonic and fetal growth such as increasing
the insulin secretion from the islets of Langerhans
during pregnancy, stimulating the synthesis and
secretion of IGFs and}or direct effects on the
developing embryo via its own receptors.
The effect of PRLs on embryonic development via
increasing the insulin secretion is not the explanation
in our study. First, the studies demonstrated the
effects of PRLs in increasing the insulin secretion from
islets have been carried out in neonatal tissues, whilst
the rat embryo in our culture system is unable to
produce its own insulin since at this time the pancreas
has not yet developed. It is therefore totally dependent
Effects of prolactin during rat organogenesis 7
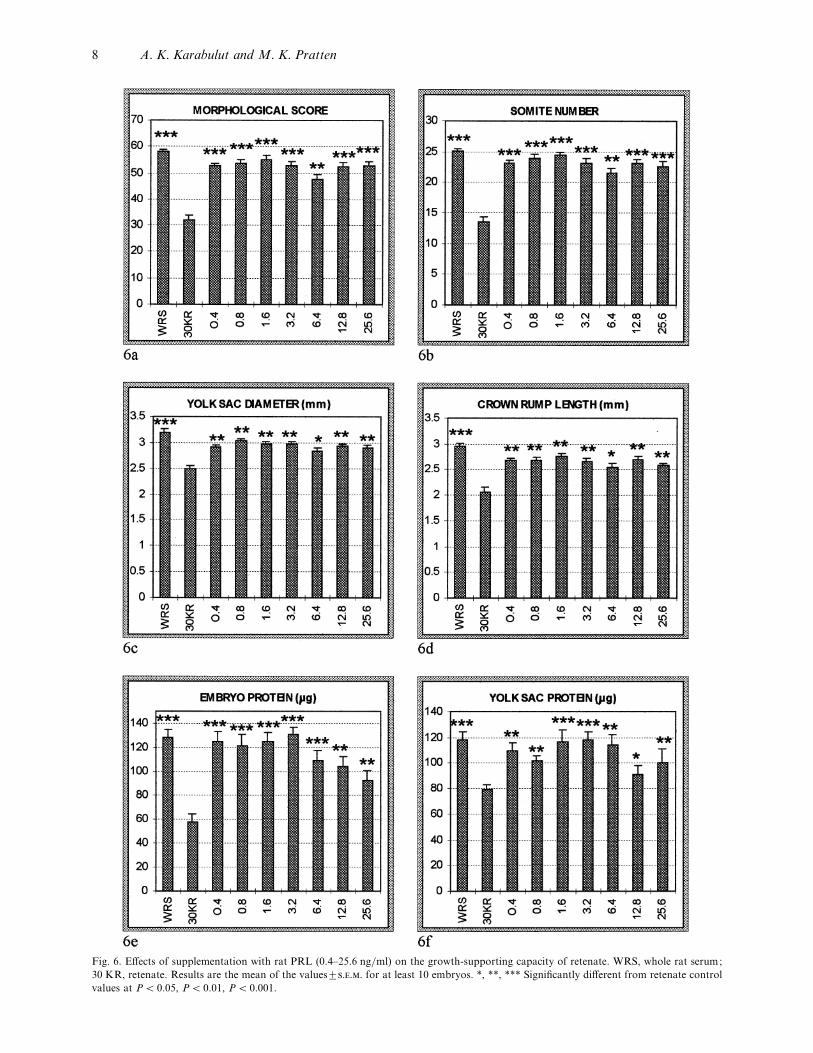
Fig. 6. Effects of supplementation with rat PRL (0±4–25±6 ng}ml) on the growth-supporting capacity of retenate. WRS, whole rat serum;
30 KR, retenate. Results are the mean of the values³... for at least 10 embryos. *, **, *** Significantly different from retenate control
values at P! 0±05, P! 0±01, P! 0±001.
8 A. K. Karabulut and M. K. Pratten
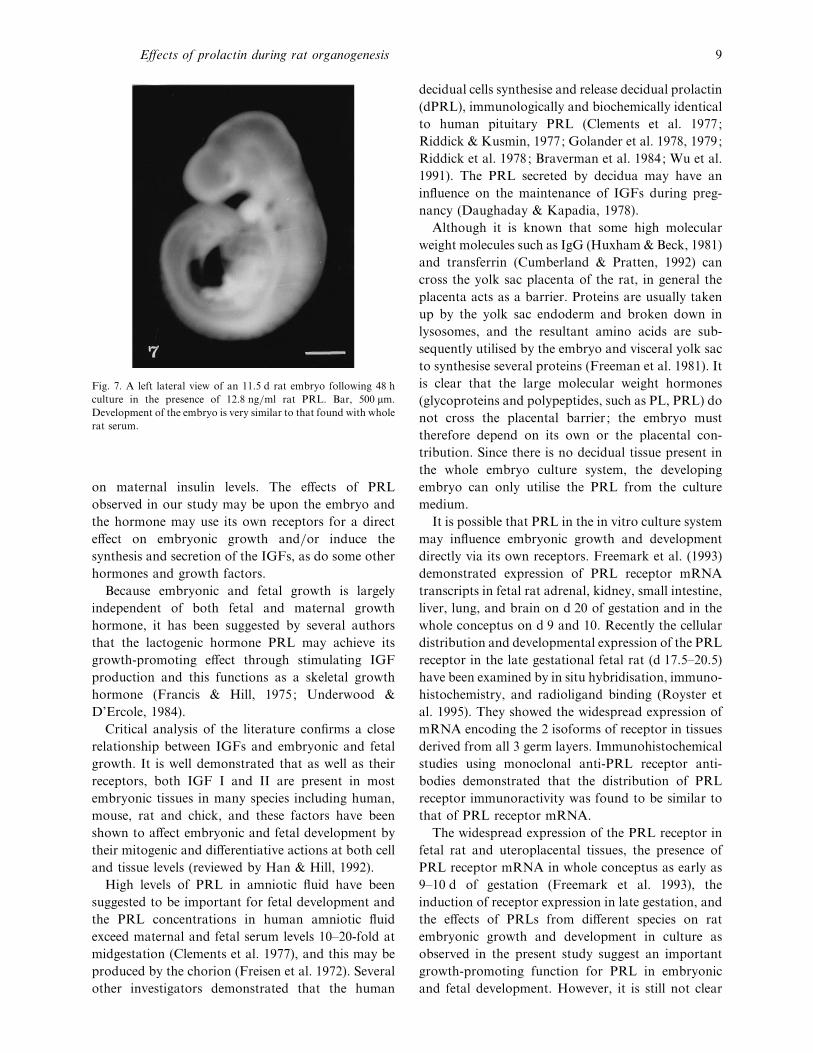
Fig. 7. A left lateral view of an 11±5 d rat embryo following 48 h
culture in the presence of 12±8 ng}ml rat PRL. Bar, 500 µm.
Development of the embryo is very similar to that found with whole
rat serum.
on maternal insulin levels. The effects of PRL
observed in our study may be upon the embryo and
the hormone may use its own receptors for a direct
effect on embryonic growth and}or induce the
synthesis and secretion of the IGFs, as do some other
hormones and growth factors.
Because embryonic and fetal growth is largely
independent of both fetal and maternal growth
hormone, it has been suggested by several authors
that the lactogenic hormone PRL may achieve its
growth-promoting effect through stimulating IGF
production and this functions as a skeletal growth
hormone (Francis & Hill, 1975; Underwood &
D’Ercole, 1984).
Critical analysis of the literature confirms a close
relationship between IGFs and embryonic and fetal
growth. It is well demonstrated that as well as their
receptors, both IGF I and II are present in most
embryonic tissues in many species including human,
mouse, rat and chick, and these factors have been
shown to affect embryonic and fetal development by
their mitogenic and differentiative actions at both cell
and tissue levels (reviewed by Han & Hill, 1992).
High levels of PRL in amniotic fluid have been
suggested to be important for fetal development and
the PRL concentrations in human amniotic fluid
exceed maternal and fetal serum levels 10–20-fold at
midgestation (Clements et al. 1977), and this may be
produced by the chorion (Freisen et al. 1972). Several
other investigators demonstrated that the human
decidual cells synthesise and release decidual prolactin
(dPRL), immunologically and biochemically identical
to human pituitary PRL (Clements et al. 1977;
Riddick & Kusmin, 1977; Golander et al. 1978, 1979;
Riddick et al. 1978; Braverman et al. 1984; Wu et al.
1991). The PRL secreted by decidua may have an
influence on the maintenance of IGFs during preg-
nancy (Daughaday & Kapadia, 1978).
Although it is known that some high molecular
weight molecules such as IgG (Huxham & Beck, 1981)
and transferrin (Cumberland & Pratten, 1992) can
cross the yolk sac placenta of the rat, in general the
placenta acts as a barrier. Proteins are usually taken
up by the yolk sac endoderm and broken down in
lysosomes, and the resultant amino acids are sub-
sequently utilised by the embryo and visceral yolk sac
to synthesise several proteins (Freeman et al. 1981). It
is clear that the large molecular weight hormones
(glycoproteins and polypeptides, such as PL, PRL) do
not cross the placental barrier ; the embryo must
therefore depend on its own or the placental con-
tribution. Since there is no decidual tissue present in
the whole embryo culture system, the developing
embryo can only utilise the PRL from the culture
medium.
It is possible that PRL in the in vitro culture system
may influence embryonic growth and development
directly via its own receptors. Freemark et al. (1993)
demonstrated expression of PRL receptor mRNA
transcripts in fetal rat adrenal, kidney, small intestine,
liver, lung, and brain on d 20 of gestation and in the
whole conceptus on d 9 and 10. Recently the cellular
distribution and developmental expression of the PRL
receptor in the late gestational fetal rat (d 17±5–20±5)
have been examined by in situ hybridisation, immuno-
histochemistry, and radioligand binding (Royster et
al. 1995). They showed the widespread expression of
mRNA encoding the 2 isoforms of receptor in tissues
derived from all 3 germ layers. Immunohistochemical
studies using monoclonal anti-PRL receptor anti-
bodies demonstrated that the distribution of PRL
receptor immunoractivity was found to be similar to
that of PRL receptor mRNA.
The widespread expression of the PRL receptor in
fetal rat and uteroplacental tissues, the presence of
PRL receptor mRNA in whole conceptus as early as
9–10 d of gestation (Freemark et al. 1993), the
induction of receptor expression in late gestation, and
the effects of PRLs from different species on rat
embryonic growth and development in culture as
observed in the present study suggest an important
growth-promoting function for PRL in embryonic
and fetal development. However, it is still not clear
Effects of prolactin during rat organogenesis 9

Fig. 8. Effects of supplementation with lower concentrations of rat PRL (0±1–0±4 ng}ml) on the growth-supporting capacity of retenate.
WRS, whole rat serum; 30 KR, retenate. Results are the mean of the values³... for at least 10 embryos. *, **, *** Significantly different
from retenate control values at P! 0±05, P! 0±01, P! 0±001.
10 A. K. Karabulut and M. K. Pratten

what exact role PRL plays in tissue development, and
the mechanism of action of PRL is poorly understood.
Ahmet K. Karabulut acknowledges the generosity of
the University of Selcuk, Turkey for the financial
support of this project.
A SO, N SP, K M, F TP, R MM
(1983a) Receptors for insulin-like growth factors and growth
effects of multiplication-stimulating activity (rat insulin-like
growth factor-II) by rat embryo fibroblasts. Endocrinology 112,
971–978.
A SO, N SP, G LA, Y YWH, R
MM (1983b) Synthesis of multiplication-stimulating activity (rat
insulin-like growth factor-II) by rat embryo fibroblasts. Endo-
crinology 112, 979–987.
A KE (1986) Epidermal growth factor:growth promotion
and pinocytosis in rat embryonic tissue. B.Sc. thesis, Leicester
University.
B S, P AF (1977) Human pituitary prolactin (hPRL):
the entire linear amino acid sequence. Journal of Clinical
Endocrinology and Metabolism 45, 1112–1115.
B MB, B A, D Z, T D, G E
(1984) Isolation of prolactin-producing cells from first and
second trimester decidua. Journal of Endocrinolology and Ex-
perimental Morpholology 58, 521–525.
B TC, S RL (1988) Nutrient and hormonal regulation
of the threshold of glucose-stimulated insulin secretion in isolated
rat pancreases. Endocrinology 123, 1582–1590.
B TC, A P, H O, S LR (1989) Effect of
prolactin versus growth hormone on islet function and the
importance of using homologous mammosomatotropic hor-
mones. Endocrinology 125, 2392–2399.
B TC, S RL (1991) Role of prolactin versus growth
hormone on islet B-cell proliferation in vitro: implications for
pregnancy. Endocrinology 128, 45–57.
B TC, S DW, L PE, O L, T F,
R M et al. (1993) Effect of homologus placental
lactogens, prolactins, and growth hormones on islet B-cell
division and insulin secretion in rat, mouse, and human islets :
implication for placental lactogen regulation of islet function
during pregnancy. Endocrinology 132, 879–887.
B TC, P JA, S RL (1994) Regulation of islet β-
cell proliferation by prolactin in rat islets. Diabetes 43, 263–273.
C NIR (1985) Trophic factors in rat serum and their effects on
embryonic development. B.Sc thesis, Leicester University.
C NIR, P MK, B F (1986) Trophic factors in rat
serum and embryonic development. Biochemical Society Trans-
actions 14, 980–982.
C JA, R FI, W JSD, F C (1977) Studies on
human sexual development. IV. Fetal pituitary and serum, and
amniotic fluid concentrations of prolactin. Journal of Clinical
Endocrinology and Metabolism 44, 408–413.
C PFT, P MK (1992) Differences in binding
characteristics of rat and human transferrin by rat visceral yolk
sac placenta. Placenta 14, 287–307.
D’E AJ, A GT, U LE (1980) Evidence
that somatomedin is synthesized by multiple tissues in the fetus.
Developmental Biology 75, 315–328.
D’E AJ, U LE (1981) Growth factors in fetal
growth and development. In Fetal Endocrinology: ORPC
Symposia on Reproductive Biology, vol. 1 (ed. Novy MJ, Resco
JA), pp. 155–182. New York: Academic Press.
D WH, K M (1978) Maintenance of serum
somatomedin activity in hypophysectomised pregnant rats.
Endocrinology 102, 1317–1320.
F J, F M (1993) Placental lactogen-binding sites in
isolated fetal fibroblasts : characterization, processing and regu-
lation. Endocrinology 132, 2477–2483.
F MJO, H DJ (1975) Prolactin-stimulated production of
somatomedin by rat liver. Nature 255, 167–168.
F SJ, B F, L JB (1981) The role of the visceral yolk
sac in mediating protein utilization by rat embryos cultured in
vitro. Journal of Embryology and Experimental Morphology 66,
223–234.
F M, H S (1984a) Ovine placental lactogen
stimulates glycogen synthesis in fetal rat hepatocytes. American
Journal of Physiology 246, E21–E24.
F M, H S (1984b) Synergistic effects of oPL
and insulin on glycogen metabolism in fetal rat hepatocytes.
American Journal of Physiology 247, E714–E718.
F M, H S (1989) The role of placental lactogen
in the regulation of fetal metabolism and growth. Journal of
Pediatric Gastroenterology and Nutrition 8, 281–287.
F M, K K, P J, R MJ, S RPC,
D P (1993) Pregnancy lactogens in the rat conceptus and
fetus : circulating levels, distribution of binding, and expression
of receptor messenger ribonucleic acid. Endocrinology 133,
1830–1842.
F HG, H P, G H, T G, T J, M R
(1972) A radioimmunoassay for human prolactin. In Prolactin
and Carcinogenesis (ed. Boyns AR, Griffiths K), pp. 64–80.
Cardiff: Fourth Tenovus Workshop, Alpha Omega Alpha.
G A, H T, B J, H A, H S
(1978) Prolactin synthesis by human chorion–decidual tissue: a
possible source of prolactin in the amniotic fluid. Science 202,
311–313.
G A, B J, H T, B S, H S
(1979) Failure of bromocriptine, dopamine, and thyrotropin-
releasing hormone to affect prolactin secretion by human
decidual tissue in vitro. Journal of Clinical Endocrinology and
Metabolism 49, 787–789.
H VKM, H DJ (1992) The involvement of insulin-like growth
factors in embryonic and fetal development. In The Insulin-like
Growth Factors (ed. Schofield PN), pp. 179–219. Oxford: Oxford
University Press.
H DJ (1992) What is the role of growth hormone and related
peptides in implantation and the development of the embryo and
fetus? Journal of Hormone Research 38, 28–34.
H DJ, F BJO, M RDG (1977) Action of rat
prolactin on plasma somatomedin levels in the rat and on
somatomedin release from perfused rat liver. Journal of Endo-
crinology 75, 137–143.
H DJ, C CJ, M RDG (1985) Incorporation of
[$H]thymidine by isolated human fetal myoblasts and fibroblasts
in response to human placental lactogen; possible mediation of
hPL action by release of somatomedin-C. Journal of Cellular
Physiology 125, 337–344.
H DJ, F M, S AJ, H S, M RDG
(1988) Placental lactogen and growth hormone receptors in
human fetal tissues : relationship to fetal plasma human placental
lactogen concentrations and fetal growth. Journal of Clinical
Endocrinology and Metabolism 66, 1283–1290.
H IM, B F (1981) Receptor mediated coated vesicle
transport of rat IgG across the 11"#day in vitro rat yolk sac. Cell
Biology International Reports 5, 1073–1081.
K AK, P MK (1995) The growth promoting
effects of human placental lactogen and human prolactin in
Effects of prolactin during rat organogenesis 11

mammalian embryos during early organogenesis. Acta Anatomica
152, 299.
K K (1995) Investigation into the role of plateled-derived growth
factor in the control of normal embryonic development. B.Sc.
thesis, Leicester University.
L OH, R NJ, F AL, R RJ (1951)
Protein measurement with the folin phenol reagent. Journal of
Biological Chemistry 193, 265–275.
N DAT (1978) Whole embryo culture and the study of
mammalian embryos during organogenesis. Biological Reviews
53, 81–122.
N CS (1978) Comparative aspects of prolactin physiology: is
prolactin the initial growth hormone in mammalian species also?
In Progress in Prolactin Physiology and Pathology (ed. Robyn C,
Harter M), pp. 175–188. Amsterdam: Elsevier}North Holland
Biomedical Press.
N CS, M GL, R SM (1986) Structural features of
prolactins and growth hormones that can be related to their
biological properties. Endocrine Reviews 7, 169–203.
N JH (1982) Effects of growth hormone, prolactin, and
placental lactogen on insulin content and release, and deoxyribo-
nucleic acid synthesis in cultured pancreatic islets. Endocrinology
110, 600–606.
O L, T F (1988) Prolactins of pregnancy and their
cellular source. International Review of Cytology 112, 1–65.
P MK, B AM, B SC, B F (1988) The effect
of epidermal growth factor, insulin and transferrin on the
growth-promoting properties of serum depleted by repeated
culture of postimplantation rat embryos. Development 104,
137–145.
R DH, K WF (1977) Decidua: a possible source of
amniotic fluid prolactin. American Journal of Obstetrics and
Gynecology 127, 187–190.
R DH, L AA, K WF, M IA (1978) De
novo synthesis of prolactin by human decidua. Life Sciences 23,
1913–1921.
R M, D P, K PA, F M (1995) The
prolactin receptor in the fetal rat : cellular localization of
messenger ribonucleic acid, immunoreactive protein, and ligand-
binding activity and induction of expression in late gestation.
Endocrinology 136, 3892–3900.
S VR, H K (1990) Insulin like growth factors and their
binding proteins. Physiological Reviews 70, 591–614.
S RL, B TC, H OD, M PA, S
JD (1987) Prolactin (in vitro) decreases the glucose stimulation
threshold, enhances insulin secretion, and increases dye coupling
among islets B cells. Endocrinology 121, 1447–1453.
T K, B LI, S A, A FJ (1979) Is
prolactin the growth hormone of the ovine fetus? Proceedings of
the Endocrinology Society 61st Annual Meeting 225, abstract 610.
T JP, P MK, B F (1989) Effects of low insulin
levels on rat embryonic growth and development. Diabetes 38,
773–778.
T JP, E L, H B, T E, L MJ, P
MK et al. (1992) Insulin and insulin-like growth factors in
embryonic development : effects of biologically inert insulin
(guinea pig) on rat embryonic growth and development in vitro.
Diabetes 41, 318–324.
U H (1997) The growth promoting effects of bFGF, VEGF and
PD-ECGF on embryonic development and yolk sac vascularisation.
PhD thesis, Nottingham University.
U H, K AK, P MK (1996) The effects of
fibroblast growth factor on embryonic development and yolk sac
vascularisation. Teratology 53, 32A.
U H, K AK, P MK (1997) The growth
promoting effects of platelet-derived endothelial cell growth
factor on early embryonic development and yolk sac vascularisa-
tion. Journal of Anatomy 191, 142.
U LE, D’E AJ (1984) Insulin and insulin-like
growth factors}somatomedins in fetal and neonatal development.
Clinical Endocrinology and Metabolism 13, 169–189.
V M-F G, D F, P JJ (1990) Morpho-
genesis and quantification of the development of post-implan-
tation mouse embryos. Toxicology In Vitro 4, 149–156.
W WX, M M, B J, L P, G A,
MN AS (1991) Localization by immunohistochemistry
and in situ hybridization of the site of production and uptake of
prolactin within the amino-placental unit in human pregnancy.
Journal of Molecular Endocrinology 7, 241–247.
12 A. K. Karabulut and M. K. Pratten
Related Documents











