NATURE BIOTECHNOLOGY VOLUME 29 NUMBER 5 MAY 2011 453 LETTERS The capacity of conventional breeding to simultaneously improve the yield and quality of cotton fiber is limited 1 . The accumulation of the plant hormone indole-3-acetic acid (IAA) in cotton fiber initials prompted us to investigate the effects of genetically engineering increased IAA levels in the ovule epidermis. Targeted expression of the IAA biosynthetic gene iaaM, driven by the promoter of the petunia MADS box gene Floral Binding protein 7 (FBP7) 2 , increased IAA levels in the epidermis of cotton ovules at the fiber initiation stage. This substantially increased the number of lint fibers, an effect that was confirmed in a 4-year field trial. The lint percentage of the transgenic cotton, an important component of fiber yield, was consistently higher in our transgenic plants than in nontransgenic controls, resulting in a >15% increase in lint yield. Fiber fineness was also notably improved. Cotton (Gossypium hirsutum L.) is the most important textile fiber crop in the world. Approximately 100 million families rely on cotton production as a major source of their income and 150 countries either import or export cotton lint 3 . The rapid increase of the world’s population and the severe decrease in arable land raise serious con- cerns about our continued ability to meet global demands for cotton. However, efforts to use conventional breeding to improve yields have been thwarted by a narrow germplasm base and technical limitations, such as the extended time frames needed to improve quantitative traits and the difficulty of introducing traits from allopolyploids or diploids of other Gossypium species 1 . It is even more challenging to improve both yield and fiber quality concurrently; the yield of cotton fibers, usually known as cotton lint, is usually negatively associated with fiber quality 4 . Fiber fineness is one of the most important quality properties of cotton fibers, and is usually measured in units of micronaire 5 , a parameter that also describes the maturity of fibers. Whereas micronaire values from 3.5–4.9 are desirable in the textile industry 6 , values >5.0 denote coarse fibers. Coarse fibers mean fewer fibers in a cross-section of yarn, result- ing in weaker spun yarn, which commands lower market values 5 . In traditional breeding, the selection for high yield of cotton fib- ers has usually been accompanied by higher micronaire values 7 . Consequently, micronaire has become a key quality parameter in many cotton-producing regions. Cotton fiber, a highly elongated cell derived from the ovule epider- mis, is characteristic of the genus Gossypium 8 . The development of cotton fiber includes four well-defined, yet overlapping, stages: initia- tion, elongation, secondary cell wall accumulation and maturation 9 . Fiber initiation and elongation have a great impact on the number, length and fineness of fibers, which are major factors determining lint yield and quality. Phytohormones, such as auxin 10–15 , gibberel- lins 10–13 , cytokinins 11,16 and ethylene 17 play important roles in fiber development. IAA, the most important natural auxin, is required for fiber production from unfertilized ovules in culture 10 . Application of IAA in vitro promoted fiber initiation 12 and increased total fiber units 11 . Under field conditions, it has been reported that the exo- genous application of IAA or gibberellin (GA 3 ) to the flower buds of the cotton plant, known as squares, and flowers resulted in significant increases in the fiber number per ovule 13 . However, as the interval when fields of cotton are flowering may span >3 months, the targeted application of growth regulators to individual flowers is impractical due to labor cost and potential environmental risk 18 . Genetic modification has provided a powerful means for cotton improvement as exemplified by transgenic cotton engineered for resist- ance to insects and herbicides 19 . An attractive strategy to enhance the intrinsic yield and/or to improve the fiber quality of cotton is to geneti- cally manipulate the plant hormone biosynthesis or signaling genes associated with cotton fiber cell development. Such a strategy has not yet succeeded in cotton 15 . Expression of the IAA biosynthetic genes iaaM and iaaH from Agrobacterium tumefaciens in cotton under the regulation of the fiber-specific E6 promoter did not affect fiber length, strength or micronaire, despite significant increases in the free IAA content in transgenic fibers 15 . Nevertheless, the well-documented role of auxin in cell development inspired us to further explore this strategy in the hope of enhancing cotton fiber yield or quality. Here, we report that the spatiotemporal regulation conferred by other promoters can Spatiotemporal manipulation of auxin biosynthesis in cotton ovule epidermal cells enhances fiber yield and quality Mi Zhang 1,4 , Xuelian Zheng 1,4 , Shuiqing Song 1 , Qiwei Zeng 1 , Lei Hou 1 , Demou Li 1 , Juan Zhao 1 , Yuan Wei 1 , Xianbi Li 1 , Ming Luo 1 , Yuehua Xiao 1 , Xiaoying Luo 1 , Jinfa Zhang 2 , Chengbin Xiang 3 & Yan Pei 1 1 Key Laboratory of Biotechnology and Crop Quality Improvement of Ministry of Agriculture, Biotechnology Research Center, Southwest University, Chongqing, P.R. China. 2 Department of Plant and Environmental Sciences, New Mexico State University, Las Cruces, New Mexico, USA. 3 School of Life Sciences, University of Science and Technology of China, Hefei, Anhui, P.R. China. 4 These authors contributed equally to this work. Correspondence should be addressed to Y.P. ([email protected]). Received 25 January; accepted 14 March; published online 10 April 2011; doi:10.1038/nbt.1843 © 2011 Nature America, Inc. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

nature biotechnology VOLUME 29 NUMBER 5 MAY 2011 453
l e t t e r s
The capacity of conventional breeding to simultaneously improve the yield and quality of cotton fiber is limited1. The accumulation of the plant hormone indole-3-acetic acid (IAA) in cotton fiber initials prompted us to investigate the effects of genetically engineering increased IAA levels in the ovule epidermis. Targeted expression of the IAA biosynthetic gene iaaM, driven by the promoter of the petunia MADS box gene Floral Binding protein 7 (FBP7)2, increased IAA levels in the epidermis of cotton ovules at the fiber initiation stage. This substantially increased the number of lint fibers, an effect that was confirmed in a 4-year field trial. The lint percentage of the transgenic cotton, an important component of fiber yield, was consistently higher in our transgenic plants than in nontransgenic controls, resulting in a >15% increase in lint yield. Fiber fineness was also notably improved.
Cotton (Gossypium hirsutum L.) is the most important textile fiber crop in the world. Approximately 100 million families rely on cotton production as a major source of their income and 150 countries either import or export cotton lint3. The rapid increase of the world’s population and the severe decrease in arable land raise serious con-cerns about our continued ability to meet global demands for cotton. However, efforts to use conventional breeding to improve yields have been thwarted by a narrow germplasm base and technical limitations, such as the extended time frames needed to improve quantitative traits and the difficulty of introducing traits from allopolyploids or diploids of other Gossypium species1.
It is even more challenging to improve both yield and fiber quality concurrently; the yield of cotton fibers, usually known as cotton lint, is usually negatively associated with fiber quality4. Fiber fineness is one of the most important quality properties of cotton fibers, and is usually measured in units of micronaire5, a parameter that also describes the maturity of fibers. Whereas micronaire values from 3.5–4.9 are desirable in the textile industry6, values >5.0 denote coarse fibers. Coarse fibers mean fewer fibers in a cross-section of yarn, result-ing in weaker spun yarn, which commands lower market values5.
In traditional breeding, the selection for high yield of cotton fib-ers has usually been accompanied by higher micronaire values7. Consequently, micronaire has become a key quality parameter in many cotton-producing regions.
Cotton fiber, a highly elongated cell derived from the ovule epider-mis, is characteristic of the genus Gossypium8. The development of cotton fiber includes four well-defined, yet overlapping, stages: initia-tion, elongation, secondary cell wall accumulation and maturation9. Fiber initiation and elongation have a great impact on the number, length and fineness of fibers, which are major factors determining lint yield and quality. Phytohormones, such as auxin10–15, gibberel-lins10–13, cytokinins11,16 and ethylene17 play important roles in fiber development. IAA, the most important natural auxin, is required for fiber production from unfertilized ovules in culture10. Application of IAA in vitro promoted fiber initiation12 and increased total fiber units11. Under field conditions, it has been reported that the exo-genous application of IAA or gibberellin (GA3) to the flower buds of the cotton plant, known as squares, and flowers resulted in significant increases in the fiber number per ovule13. However, as the interval when fields of cotton are flowering may span >3 months, the targeted application of growth regulators to individual flowers is impractical due to labor cost and potential environmental risk18.
Genetic modification has provided a powerful means for cotton improvement as exemplified by transgenic cotton engineered for resist-ance to insects and herbicides19. An attractive strategy to enhance the intrinsic yield and/or to improve the fiber quality of cotton is to geneti-cally manipulate the plant hormone biosynthesis or signaling genes associated with cotton fiber cell development. Such a strategy has not yet succeeded in cotton15. Expression of the IAA biosynthetic genes iaaM and iaaH from Agrobacterium tumefaciens in cotton under the regulation of the fiber-specific E6 promoter did not affect fiber length, strength or micronaire, despite significant increases in the free IAA content in transgenic fibers15. Nevertheless, the well-documented role of auxin in cell development inspired us to further explore this strategy in the hope of enhancing cotton fiber yield or quality. Here, we report that the spatiotemporal regulation conferred by other promoters can
spatiotemporal manipulation of auxin biosynthesis in cotton ovule epidermal cells enhances fiber yield and qualityMi Zhang1,4, Xuelian Zheng1,4, Shuiqing Song1, Qiwei Zeng1, Lei Hou1, Demou Li1, Juan Zhao1, Yuan Wei1, Xianbi Li1, Ming Luo1, Yuehua Xiao1, Xiaoying Luo1, Jinfa Zhang2, Chengbin Xiang3 & Yan Pei1
1Key Laboratory of Biotechnology and Crop Quality Improvement of Ministry of Agriculture, Biotechnology Research Center, Southwest University, Chongqing, P.R. China. 2Department of Plant and Environmental Sciences, New Mexico State University, Las Cruces, New Mexico, USA. 3School of Life Sciences, University of Science and Technology of China, Hefei, Anhui, P.R. China. 4These authors contributed equally to this work. Correspondence should be addressed to Y.P. ([email protected]).
Received 25 January; accepted 14 March; published online 10 April 2011; doi:10.1038/nbt.1843
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.
454 VOLUME 29 NUMBER 5 MAY 2011 nature biotechnology
l e t t e r s
manipulate auxin levels in ovule epidermal cells at the stage of fiber cell initiation in a manner that improves both cotton fiber yield and quality.
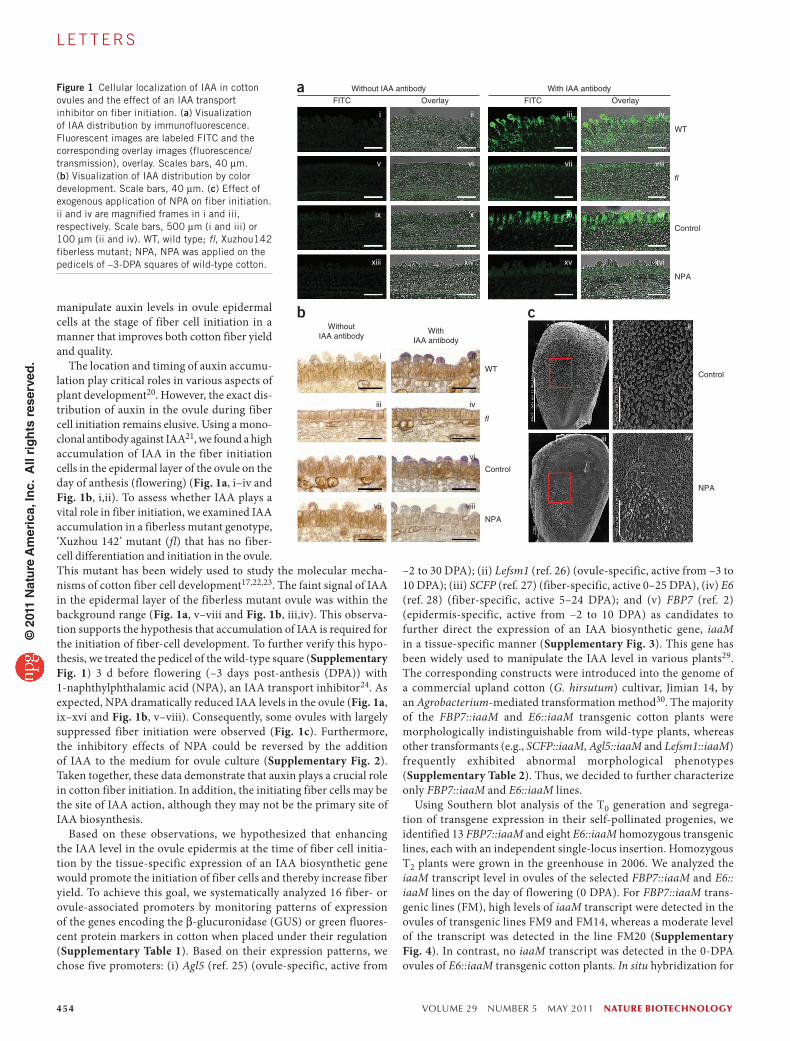
The location and timing of auxin accumu-lation play critical roles in various aspects of plant development20. However, the exact dis-tribution of auxin in the ovule during fiber cell initiation remains elusive. Using a mono-clonal antibody against IAA21, we found a high accumulation of IAA in the fiber initiation cells in the epidermal layer of the ovule on the day of anthesis (flowering) (Fig. 1a, i–iv and Fig. 1b, i,ii). To assess whether IAA plays a vital role in fiber initiation, we examined IAA accumulation in a fiberless mutant genotype, ‘Xuzhou 142’ mutant (fl) that has no fiber-cell differentiation and initiation in the ovule. This mutant has been widely used to study the molecular mecha-nisms of cotton fiber cell development17,22,23. The faint signal of IAA in the epidermal layer of the fiberless mutant ovule was within the background range (Fig. 1a, v–viii and Fig. 1b, iii,iv). This observa-tion supports the hypothesis that accumulation of IAA is required for the initiation of fiber-cell development. To further verify this hypo-thesis, we treated the pedicel of the wild-type square (Supplementary Fig. 1) 3 d before flowering (–3 days post-anthesis (DPA)) with 1-naphthylphthalamic acid (NPA), an IAA transport inhibitor24. As expected, NPA dramatically reduced IAA levels in the ovule (Fig. 1a, ix–xvi and Fig. 1b, v–viii). Consequently, some ovules with largely suppressed fiber initiation were observed (Fig. 1c). Furthermore, the inhibitory effects of NPA could be reversed by the addition of IAA to the medium for ovule culture (Supplementary Fig. 2). Taken together, these data demonstrate that auxin plays a crucial role in cotton fiber initiation. In addition, the initiating fiber cells may be the site of IAA action, although they may not be the primary site of IAA biosynthesis.
Based on these observations, we hypothesized that enhancing the IAA level in the ovule epidermis at the time of fiber cell initia-tion by the tissue-specific expression of an IAA biosynthetic gene would promote the initiation of fiber cells and thereby increase fiber yield. To achieve this goal, we systematically analyzed 16 fiber- or ovule-associated promoters by monitoring patterns of expression of the genes encoding the β-glucuronidase (GUS) or green fluores-cent protein markers in cotton when placed under their regulation (Supplementary Table 1). Based on their expression patterns, we chose five promoters: (i) Agl5 (ref. 25) (ovule-specific, active from
–2 to 30 DPA); (ii) Lefsm1 (ref. 26) (ovule-specific, active from –3 to 10 DPA); (iii) SCFP (ref. 27) (fiber-specific, active 0–25 DPA), (iv) E6 (ref. 28) (fiber-specific, active 5–24 DPA); and (v) FBP7 (ref. 2) (epidermis-specific, active from –2 to 10 DPA) as candidates to further direct the expression of an IAA biosynthetic gene, iaaM in a tissue-specific manner (Supplementary Fig. 3). This gene has been widely used to manipulate the IAA level in various plants29. The corresponding constructs were introduced into the genome of a commercial upland cotton (G. hirsutum) cultivar, Jimian 14, by an Agrobacterium-mediated transformation method30. The majority of the FBP7::iaaM and E6::iaaM transgenic cotton plants were morphologically indistinguishable from wild-type plants, whereas other transformants (e.g., SCFP::iaaM, Agl5::iaaM and Lefsm1::iaaM) frequently exhibited abnormal morphological phenotypes (Supplementary Table 2). Thus, we decided to further characterize only FBP7::iaaM and E6::iaaM lines.
Using Southern blot analysis of the T0 generation and segrega-tion of transgene expression in their self-pollinated progenies, we identified 13 FBP7::iaaM and eight E6::iaaM homozygous transgenic lines, each with an independent single-locus insertion. Homozygous T2 plants were grown in the greenhouse in 2006. We analyzed the iaaM transcript level in ovules of the selected FBP7::iaaM and E6::iaaM lines on the day of flowering (0 DPA). For FBP7::iaaM trans-genic lines (FM), high levels of iaaM transcript were detected in the ovules of transgenic lines FM9 and FM14, whereas a moderate level of the transcript was detected in the line FM20 (Supplementary Fig. 4). In contrast, no iaaM transcript was detected in the 0-DPA ovules of E6::iaaM transgenic cotton plants. In situ hybridization for
a
b c
Without IAA antibody
WithoutIAA antibody
FITC
i ii iii iv
i ii
iii iv
i ii
iii iv
v vi vii viii
v vi
vii viii
ix x xi xii
xiii xiv xv xvi
Overlay
With IAA antibody
WithIAA antibody
FITC Overlay
WT
Control
NPA
Control
NPA
fl
WT
Control
NPA
fl
Figure 1 Cellular localization of IAA in cotton ovules and the effect of an IAA transport inhibitor on fiber initiation. (a) Visualization of IAA distribution by immunofluorescence. Fluorescent images are labeled FITC and the corresponding overlay images (fluorescence/transmission), overlay. Scales bars, 40 µm. (b) Visualization of IAA distribution by color development. Scale bars, 40 µm. (c) Effect of exogenous application of NPA on fiber initiation. ii and iv are magnified frames in i and iii, respectively. Scale bars, 500 µm (i and iii) or 100 µm (ii and iv). WT, wild type; fl, Xuzhou142 fiberless mutant; NPA, NPA was applied on the pedicels of –3-DPA squares of wild-type cotton.
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.

nature biotechnology VOLUME 29 NUMBER 5 MAY 2011 455
l e t t e r s
FBP7::iaaM cotton (Fig. 2a) showed that the FBP7 promoter could direct the expression of iaaM specifically in the epidermal cells, which was in agreement with the results of GUS staining for FBP7::GUS cotton (Fig. 2b). Quantitative real-time PCR was used to investigate the temporal expression pattern of iaaM in FM9. The iaaM transcript level was high in the –2-DPA to 5-DPA ovules (during the stages of fiber cell differentiation and ini-tiation) and then gradually declined. At 15 DPA, the iaaM transcript was nearly undetectable (Fig. 2c). To further quantify the IAA levels, we analyzed the free IAA content in 0-, 1-, 2- and 3-DPA ovules by liquid chromatography/mass spectrometry. An increase of IAA level was detected in the ovules of FM9 relative to that in wild-type ovules. On average, IAA levels in FM9 ovules increased by 128% relative to those of ovules from wild-type plants (Fig. 2d). As a comparison, no increase of IAA was found in the 0-DPA ovules of the E6::iaaM transgenic lines (Supplementary Fig. 5).
In addition, we examined the NPA effects on IAA biosynthesis in FM9 using IAA monoclonal antibody. Although the IAA levels in initiating fiber cells of FM9 treated with NPA were lower than those in the untreated FBP7::iaaM cotton, levels in FM9 were con-siderably higher than those in wild-type plants treated with NPA (Fig. 2e). This result confirmed the FBP7 promoter–targeted expres-sion of iaaM-enhanced IAA content in ovule epidermal cells.
Next, we assessed the effects of the enhanced IAA accumulation in the ovule epidermis on the development of fiber cells in the FM lines. The number of initiated fiber cells (initials) on the 0-DPA ovules, as quantified by scanning electron microscopy, had a 28.6%, 26.6% and 13.5% increase in the transgenic line FM9, FM14 and FM20, respectively, relative to the wild type (Fig. 3a). This result suggests that fiber cell initiation was promoted by the FBP7::iaaM-mediated elevation of IAA in the epidermis of cotton ovules. Consequently, the transgenic lines FM9, FM14 and FM20 produced 39.8%, 38.9% and 19.9% more mature lint fibers per seed, respectively, than the wild
type (Fig. 3b). The finding is consistent with the increase in fiber cell initiation after treatment of the pedicels of –3-DPA squares with IAA (Supplementary Fig. 6). However, no significant changes in the number of fiber initials and mature fibers were found in E6::iaaM transgenic lines (Fig. 3a,b). To further verify the role of IAA in fiber cell initiation, we generated FBP7::iaaL transgenic cotton plants. iaaL encodes an enzyme converting IAA to IAA-lysine31, a biologically inactive auxin. Reduced IAA accumulation was observed in the ovules of FBP7::iaaL plants, and the number of fiber initials on the 0-DPA ovules was dramatically decreased (Supplementary Fig. 7).
Fiber quality and yield components of the transgenic cotton lines (T2) and their nontransgenic segregants (control) grown in the greenhouse were measured in 2006. Lint percentages [(weight of lint fibers/weight of seed cotton) × 100] were 45.0%, 44.4% and 40.5% for transgenic lines FM9, FM14 and FM20, respectively, which were much higher than that for the control plants (37.5%). Interestingly, micronaire values of the fibers were considerably lower in the trans-genic lines (4.5, 4.3 and 4.8 for FM9, FM14 and FM20, respectively) compared with the control plants (6.1–6.3). We observed paraffin-embedded sections of the fiber and found that the cross-section perimeter of the transgenic cotton was smaller than that of the wild type (Supplementary Fig. 8a,b), demonstrating that the fibers of the transgenic cotton were finer.
The agronomic performance of the transgenic lines in terms of fiber yield and quality was further evaluated in field trials in the experi-mental farm at Southwest University (Chongqing, China) in 2007, 2008, 2009 and 2010 (transgenic trial permit number: 2007T019). The lint percentage of transgenic lines FM9 and FM14 were consistently
e
a c
d
bf nfv vi
Sense probe
i ii
WT
FM
iii
f
nf
iv
Antisense probe
Rel
ativ
e ex
pres
sion
leve
l
8
7
6
5
4
3
2
1
0–2 0 1 2 3 5 10 15 20 30
DPA
IAA
con
tent
(ng
/g F
W) Wild type
FBP7::iaaM975
100
50
25
00 DPA 1 DPA 2 DPA 3 DPA
i ii iii iv
v vi vii viii
Without IAA antibody With IAA antibody
FITC Overlay FITC Overlay
FM
NPA(FM)
NPA(WT)
ix x xi xii
Figure 2 Targeted expression of iaaM driven by FBP7 promoter in the epidermis of ovules increases IAA accumulation in transgenic cotton. (a) Localization of iaaM mRNA in cotton ovules of FBP7::iaaM cotton, as determined by in situ hybridization. (b) Histochemical localization of GUS activity in FBP7::GUS transgenic cotton. Stained 0-DPA ovules of FBP7::GUS cotton were observed in paraffin-embedded sections. The magnified image (vi) is from the red square in the original (v). Triangles point to the ovule epidermal layer. Arrows point to the fiber initial (f) or the uninitiated cell (nf). WT, wild type; FM, FBP7::iaaM. Scale bars, 50 µm. (c) FBP7::iaaM expression pattern in transgenic cotton ovules (with fibers). Bars represent the s.d. of three repeated PCRs. (d) Endogenous IAA levels in ovules of line FM9 and wild type during fiber initiation. The bar represents the s.d. of three samples. (e) Cellular localization of IAA distribution in 0-DPA ovules visualized by immunofluorescence. Fluorescent images are marked as FITC and corresponding overlay images (fluorescence/transmission), as overlay. FM, FBP7::iaaM cotton treated with DMSO; NPA (FM), FBP7::iaaM cotton treated with NPA; NPA (WT), wild type treated with NPA. Scale bars, 40 µm.
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.

456 VOLUME 29 NUMBER 5 MAY 2011 nature biotechnology
l e t t e r s
higher and micronaire values of the two lines were lower than for the nontransgenic control in the 3-year trials (2007, 2008 and 2009; Table 1). The combined analysis of variance (ANOVA) for the results of 2007, 2008 and 2009 confirmed significant genotype effects on lint percentage (P = 4.71×10−10), lint yield (P = 3.32×10−10) and micro-naire value (P = 2.24×10−7) (Supplementary Tables 3a,b). Averaged
over the 3 years, the lint percentages of transgenic lines FM9 (49.8%) and FM14 (48.3%) were significantly higher than those of the controls (40.6%), resulting in a 23.7% and 14.7% increase, respectively, in lint yield. The combined ANOVA also showed that there is no significant genotype × year interaction for lint percentage, lint yield and micro-naire value. This result demonstrates that the field performance of the
a
b
c
d
Mic
rona
ire (
2008
)
Mic
rona
ire (
2009
)
5.21 ± 0.52
4.48 ± 0.494.42 ± 0.48
5.19 ± 0.15
4.56 ± 0.214.21 ± 0.16
7
6
5
4
3
2
7
6
5
4
3
2
Aug. 2
1–31
Sept.
1–10
Sept.
11–2
0
Sampling time
Sept.
21–3
0
Oct. 1
–10
Aug. 2
1–31
Sept.
1–10
Sept.
11–2
0
Sampling time
Sept.
21–3
0
Oct. 1
–10
WT
FM9
FM14
FM20
EM23
10,000
12,000
** ** *
Initi
al d
ensi
ty (
per
mm
2 )
8,000
6,000
4,000
2,000
WT
FM9
FM14
FM20
EM23
24,000
20,000
16,000
12,000
8,000
4,000
0
* *
Num
ber
of m
atur
e �b
ers
(per
see
d)
56
48
40
32
24
56
48
40
32
24
Lint
per
cent
age
(200
8)
Lint
per
cent
age
(200
9)
Aug. 2
1–31
Sept.
1–10
Sept.
11–2
0
Sampling time
Sept.
21–3
0
Oct. 1
–10
Aug. 2
1–31
Sept.
1–10
Sept.
11–2
0
Sampling time
Sept.
21–3
0
Oct. 1
–10
49.9 ± 1.148.3 ± 1.1
41.3 ± 1.3
50.1 ± 0.848.6 ± 1.2
39.7 ± 1.2
Control
FM9
FM14
Control
FM9
FM14
ControlFM9FM14
ControlFM9FM14
Table 1 Mean comparisons for micronaire, lint percentage, seed cotton and lint yield in transgenic (FBP7::iaaM) cotton and the control within and over years (2007, 2008 and 2009) in Chongqing, ChinaYear Line Micronaire Lint (%) Seed cotton yield (kg/plot) Lint yield (kg/plot)
2007 FM9 4.6a 49.4a 3.57a 1.76a
FM14 4.5a 48.1a 3.25a 1.57a
FM20 5.2b 43.7b 3.38a 1.48a
Control 5.2b 40.7c 3.35a 1.37a
2008 FM9 4.5a 50.0a 3.97a 1.98a
FM14 4.4a 48.2a 3.87a 1.87ab
FM20 5.2b 43.8b 3.95a 1.73bc
Control 5.3b 41.3b 3.90a 1.61c
Wild type 5.2b 40.4b 3.75a 1.51c
2009 FM9 4.6a 50.1a 4.10b 2.05a
FM14 4.2b 48.6a 3.96b 1.93b
Control 5.2c 39.7b 4.25a 1.69c
Average over 2 years (2007 & 2008) FM9 4.5a 49.7a 3.77a 1.87a
FM14 4.5a 48.2b 3.56a 1.72ab
FM20 5.2b 43.7c 3.67a 1.60ab
Control 5.3b 41.0d 3.63a 1.49b
Average over 3 years (2007, 2008 & 2009) FM9 4.5a 49.8a 3.88a 1.93a
FM14 4.4b 48.3b 3.69a 1.79a
Control 5.2c 40.6c 3.83a 1.56b
Lint (%) = (lint weight/seed cotton weight) × 100. Seed cotton = seed + lint. Control, segregated nontransgenic plants derived from T1 of the transformants FM9, FM14 and FM20. Because FM20 did not show significant improvements in lint yield and micronaire compared with the control in 2007 and 2008 trials, this line was not grown in the 2009 trial. Within each column, means that are not followed by the same letter are significantly different according to Tukey’s range test at 0.05 level. In 2007, the plot means of lint percentage and micronaire value were based on the average over three sampling times every 10 d (September 1 to September 30). For 2008 and 2009 trials, the plot means of lint percentage and micronaire value were based on the average over five sampling times every 10 d (August 21 to October 10).
Figure 3 Comparison of fiber initial density, number of mature fibers per seed, and variation of lint percentage and micronaire value over five sampling times in transgenic cotton plants and nontransgenic controls. (a) Fiber initial densities on the surface of ovules at 0 DPA. Error bars represent s.d. of nine ovules. (b) Number of mature fibers per seed. Bars represent s.d. of six measurements. Asterisks indicate statistically significant differences between transgenic lines and wild type, as determined by Student’s t-test (*, P < 0.05; **, P < 0.01). (c) Variation of lint percentage over five sampling times in the trials of 2008 and 2009. (d) Variation of micronaire value over five sampling times in the trials of 2008 and 2009. Bars represent s.d. of three replications. Dotted lines with corresponding colors show the average throughout the harvest season in each year. WT, wild type; FM, FBP7::iaaM; EM, E6::iaaM; Control, segregated, nontransgenic plants derived from FBP7::iaaM cotton.
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.

nature biotechnology VOLUME 29 NUMBER 5 MAY 2011 457
l e t t e r s
transgenic lines was stable over years (Supplementary Tables 3a,b). The yield component analysis showed that lint index (fiber weight in grams per 100 seeds) was significantly (P = 1.27×10−2) higher in FM9 and FM14, which is accompanied by a lower seed index (seed weight in grams per 100 seeds, Supplementary Table 4). As there was no difference in seed cotton yield between transgenic lines and the nontransgenic control (Table 1), the greater lint yield is mainly due to the enhanced lint percentage. The lower seed weight of transgenic cotton might result from a changed distribution of carbohydrates in the developing seed. Nonetheless, the smaller seed size is not of con-cern because the seed index was still in the range of most commercial cotton cultivars (9–12 g).
Within each single year (2007, 2008 and 2009), time of sampling substantially influenced lint percentage and micronaire (Fig. 3c,d and Supplementary Table 3c), indicating that both traits, espe-cially micronaire, were affected by the weather, time of the year when the plants were grown and developmental stage of the plants. Nevertheless, the FBP7::iaaM transgenic lines always outperformed the nontransgenic control in each sampling time (Fig. 3c,d), con-firming that the FBP7::iaaM transgene improves both traits. Interestingly, as the amount of lint fibers increased, the amount of fuzz fiber on the seed, which renders material unusable in the textile industry, was reduced in the transgenic lines FM9 and FM14 (Supplementary Fig. 9). In addition, no significant alteration in fiber strength and fiber length was observed (Supplementary Table 5). The 2010 field trial was carried out with the same random-ized complete block experimental design as in previous years, but a much larger plot size (100 m2 per plot) was used. The transgenic lines consistently showed a higher lint percentage (47.8% for FM9 and 44.8% for FM14) than that of the nontransgenic control (37.3%), leading to a 34.6% (FM9) and 17.5% (FM14) increase in lint yield. Meanwhile, the micronaire values of the transgenic lines remained significantly (P = 5.51×10−5) lower than those of control plants (Supplementary Table 6). Overall, the consecutive 4-year field trials demonstrated that the transgenic traits, including significantly higher lint percentage and significantly lower micronaire, were consistent across multiple growing seasons.
Transgenic approaches have enabled the manipulation of hormone concentration or tissue sensitivity to hormones in plants for improv-ing crop yields and quality29. We attribute the different phenotypes seen in FBP7::iaaM and E6::iaaM plants to the need to precisely tune the spatial and temporal regulation of endogenous hormone levels with the processes of fiber initiation and development. Approximately 10% of the epidermal cells on cotton ovules develop into fibers32. Only those cells that initiate within 3 d post-anthesis (0–3 DPA) become lint fibers, whereas cells that initiate 4–10 DPA become fuzz fibers9. As the FBP7 promoter is active specifically in the epidermal layer from −2 to 10 DPA, an interval that spans the lint fiber initiation stage, the expression of FBP7::iaaM leads to an evident enhancement of IAA levels in the epidermal layers during fiber initiation. Consequently, the enhanced IAA level resulted in a significant (P = 1.22×10−2) increase of lint fibers, accompanied by a large decrease of fuzz fibers (Supplementary Fig. 9) as more ovule epidermal cells developed into lint fibers. In contrast, the E6 promoter is active in the 5- to 24-DPA fiber cells28. By then, the fate of epidermal cells to develop into lint fibers has already been deter-mined. To further confirm that enhancing IAA accumulation at the initiation stage of lint fiber cells in the ovule is critical for a higher lint yield, we generated new transgenic cotton plants using another epidermis-specific promoter BAN33. The GUS expression profile of this Arabidopsis promoter resembles that of the FBP7 promoter
(Supplementary Fig. 10a). A significant improvement of lint per-centage and micronaire was observed in the BAN::iaaM transgenic plants (Supplementary Fig. 10b,c). This corroborates what we found in the FBP7::iaaM transgenic cotton plants.
In addition to spatial and temporal specificity, the level of the target gene expression is also critical for fiber improvement. We found that most of the transgenic cotton plants we tested (e.g., SCFP::iaaM, Agl5::iaaM and Lefsm1::iaaM) showed abnormal growth (Supplementary Table 2). The transgenic plants that demonstrated normal growth were those with weaker promoters, such as E6 and FBP7. These results demonstrate that to avoid growth abnormalities in iaaM transgenic cotton, it is advisable to use a weaker promoter rather than a stronger promoter, because the amount of the hormone produced by the trans-gene should be confined to the target tissue at an appropriate level.
Most agronomic traits related to fibers, such as lint yield, fiber length, strength and fineness, are quantitatively inherited. Moreover, the frequency with which lint yield and fiber quality traits are unfavo-rably associated, has greatly limited the efficiency of conventional cot-ton breeding4. In the past several decades, conventional breeding has developed many high-yielding cotton varieties, but it has also inad-vertently increased micronaire values in these commercial varieties7. Consequently, high micronaire values (>5.0) have become one of the main concerns of fiber quality in the cotton industry. The present study demonstrated that both fiber yield and micronaire value can be simulta-neously improved. The ovule epidermis–specific expression of iaaM produces more lint fibers, and the greater fiber density on the ovule may lead to finer fibers.
Our demonstration that ovule epidermis–specific expression of an auxin biosynthetic gene can improve both cotton fiber yield and quality appears to have resolved a longstanding challenge for cotton improvement.
MeThoDSMethods and any associated references are available in the online version of the paper at http://www.nature.com/nbt/index.html.
Note: Supplementary information is available on the Nature Biotechnology website.
ACknoWLeDgMentSWe are indebted to X. Jiang (Beijing Forestry University, China) for technical assistance on plant hormone analysis, and to D. Zhang (USDA/ARS, BARC/PSI/SPCL) for his kind help in the statistical analysis of field data. We are grateful to X. Qiu (Department of Food & Bioproduct Sciences, University of Saskatchewan, Canada), J. Huang (Monsanto Company, USA) and J. Jacobs (Bayer CropScience, Belgium), Z. Pei (Department of Biology, Duke University, USA) and C. Lin (Department of Molecular, Cell, and Developmental Biology, University of California, USA), G. Xia (Institute of Microbiology, Chinese Academy Of Science) for their critical comments on the manuscript. This work has been supported by grant 2010CB126000 (to Y.P.) from the Chinese Ministry of Science and Technology, and by grant 30530490 (to Y.P.) from the National Natural Sciences Foundation of China.
AUtHoR ContRIBUtIonSM.Z. performed field trials, in situ hybridization and quantified IAA content; X.Z. performed greenhouse experiments and quantified IAA content; S.S. performed cotton transformation and field trials; Q.Z. and Y.W. performed cotton transformation; J.Z. and X.L. performed field trials; L.H., D.L., M.L., Y.X. and X.L. performed vector construction and promoter analysis; J.Z. performed field data analysis; C.X. designed the hybridization experiment; Y.P. designed experiments and wrote the paper.
CoMPetIng FInAnCIAL InteReStSThe authors declare no competing financial interests.
Published online at http://www.nature.com/nbt/index.html. reprints and permissions information is available online at http://www.nature.com/reprints/index.html.
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.

458 VOLUME 29 NUMBER 5 MAY 2011 nature biotechnology
l e t t e r s
1. Meredith, W.R. Cotton yield progress: why has it reached a plateau? Better Crops 84, 6–9 (2000).
2. Colombo, L. et al. Downregulation of ovule-specific MADS box genes from petunia results in maternally controlled defects in seed development. Plant Cell 9, 703–715 (1997).
3. Chen, Z.J. et al. Toward sequencing cotton (Gossypium) genomes. Plant Physiol. 145, 1303–1310 (2007).
4. Meredith, W.R. Quantitative genetics. in Cotton (eds. Kohel, R.J. & Lewis, C.F.) 131–150 (American Society of Agronomy, Madison, Wisconsin, USA, 1984).
5. Gordon, S. Cotton fibre quality. in Cotton: Science and Technology (eds. Gordon, S. & Hsieh, Y.L.) 68–100 (CRC Press, Boca Raton, FL, USA, 2007).
6. Benedict, C.R., Kohel, J.R. & Lewis, H.L. Cotton fiber quality. in Cotton (eds. Smith, C.W. & Cothren, J.T.) 270 (John Wiley & Sons, New York, USA, 1999).
7. Zhang, J.F., Lu, Y., Adragna, H. & Hughs, E. Genetic improvement of New Mexico acala cotton germplasm and their genetic diversity. Crop Sci. 45, 2363–2373 (2005).
8. Stewart, J.M. Fiber initiation on the cotton ovule (Gossypium hirsutum). Am. J. Bot. 62, 723–730 (1975).
9. Basra, A.S. & Saha, S. Growth regulation of cotton fibers. in Cotton Fibers: Developmental Biology, Quality Improvement, and Textile Processing (ed. Basra, A.S.) 47–58 (The Haworth Press, New York, USA, 1999).
10. Beasley, C.A. Hormonal regulation of growth in unfertilized cotton ovules. Science 179, 1003–1005 (1973).
11. Beasley, C.A. & Ting, I.P. Effects of plant growth substances on in vitro fiber development from unfertilized cotton ovules. Am. J. Bot. 61, 188–194 (1974).
12. Gialvalis, S. & Seagull, R.W. Plant hormones alter fiber initiation in unfertilized, cultured ovules of Gossypium hirsutum. J. Cotton Sci. 5, 252–258 (2001).
13. Seagull, R.W. & Giavalis, S. Pre- and post-anthesis application of exogenous hormones alters fiber production in Gossypium hirsutum L. cultivar Maxxa GTO. J. Cotton Sci. 8, 105–111 (2004).
14. Guinn, G. & Brummett, D.L. Changes in abscisic acid and indoleacetic acid before and after anthesis relative to changes in abscission rates of cotton fruiting forms. Plant Physiol. 87, 629–631 (1988).
15. John, M.E. Genetic engineering strategies of cotton fiber modification. in Cotton Fibers: Developmental Biology, Quality Improvement and Textile Processing (ed. Basra, A.S.) 271–292 (The Haworth Press, New York, USA, 1999).
16. Chen, J.G., Du, X.M., Zhou, X. & Zhao, H.Y. Levels of cytokinins in the ovules of cotton mutants with altered fiber development. J. Plant Growth Regul. 16, 181–185 (1997).
17. Shi, Y. et al. Transcriptome profiling, molecular biological, and physiological studies reveal a major role for ethylene in cotton fiber cell elongation. Plant Cell 18, 651–664 (2006).
18. Li, Y. et al. Transgenics of plant hormones and their potential application in horticultural crops. in Genetically Modified Crops, their Development, Uses and Risks (eds. Liang, G.H. & Skinner, D.Z.) 101–117 (The Haworth Press, New York, USA, 2004).
19. Wilkins, T.A., Rajasekaran, K. & Anderson, D.M. Cotton biotechnology. Crit. Rev. Plant Sci. 19, 511–550 (2000).
20. Vanneste, S. & Friml, J. Auxin: a trigger for change in plant development. Cell 136, 1005–1016 (2009).
21. Hou, Z. & Huang, W. Immunohistochemical localization of IAA and ABP1 in strawberry shoot apexes during floral induction. Planta 222, 678–687 (2005).
22. Ji, S. et al. Isolation and analyses of genes preferentially expressed during early cotton fiber development by subtractive PCR and cDNA array. Nucleic Acids Res. 31, 2534–2543 (2003).
23. Tu, L. et al. Genes expression analyses of sea-island cotton (Gossypium barbadense L.) during fiber development. Plant Cell Rep. 26, 1309–1320 (2007).
24. Morgan, D.G. Influence of 1-naphthylphthalamic acid on the movement of indolyl-3-acetic acid in plants. Nature 201, 476–477 (1964).
25. Savidge, B., Rounsley, S.D. & Yanofsky, M.F. Temporal relationship between the transcription of two Arabidopsis MADS box genes and the floral organ identity genes. Plant Cell 7, 721–733 (1995).
26. Barg, R. et al. The tomato early fruit specific gene Lefsm1 defines a novel class of plant-specific SANT/MYB domain proteins. Planta 221, 197–211 (2005).
27. Hou, L. et al. SCFP, a novel fiber-specific promoter in cotton. Chin. Sci. Bull. 53, 2639–2645 (2008).
28. John, M.E. & Crow, L.J. Gene expression in cotton (Gossypium hirsutum L.) fiber: cloning of the mRNAs. Proc. Natl. Acad. Sci. USA 89, 5769–5773 (1992).
29. Phillips, A.L. Genetic and transgenic approaches to improving crop performance via hormones. in Plant Hormones: Biosynthesis, Signal Transduction, Action, edn 2 (ed. Davies, P.J.) 582–609 (Springer, Dordrecht, The Netherlands, 2007).
30. Luo, M. et al. GhDET2, a steroid 5α-reductase, plays an important role in cotton fiber cell initiation and elongation. Plant J. 51, 419–430 (2007).
31. Glass, N.L. & Kosuge, T. Cloning of the gene for indoleacetic acid-lysine synthetase from Pseudomonas syringae subsp. savastanoi. J. Bacteriol. 166, 598–603 (1986).
32. Ryser, U. Cotton fiber initiation and histodifferentiation. in Cotton Fibers: Developmental Biology, Quality Improvement, and Textile Processing (ed. Basra, A.S.) 1–34 (The Haworth Press, New York, USA, 1999).
33. Debeaujon, I. et al. Proanthocyanidin-accumulating cells in Arabidopsis testa: Regulation of differentiation and role in seed development. Plant Cell 15, 2514–2531 (2003).
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.

nature biotechnologydoi:10.1038/nbt.1843
oNLINe MeThoDSPlasmid construction. The FBP7 gene promoter2 was amplified with flank-ing KpnI and XbaI restriction sites, and the iaaM fragment was cloned with flanking XbaI and EcoRI restriction sites. The fragments were ligated and inserted to a modified binary vector pBI121-M by KpnI and EcoRI sites. The construction contains GUS and NPTII genes driven by CaMV 35S promoter. A similar strategy was used for construction of iaaM driven by E6, SCFP, Lefsm1 and Agl5 promoters. The resulting plasmids were inserted into Agrobacterium tumefaciens (LBA4404) by electroporation.
Plant transformation. Cotton transformation was performed as previously described30. A commercial upland cotton (Gossypium hirsutum) cultivar ‘Jimian 14’ was used as transformation material. Kanamycin-resistant and GUS-positive regeneration plants were grown in the greenhouse at Southwest University, Chongqing, China.
Molecular analyses. RNA extraction was conducted according to the method previously described34. cDNA was synthesized by RevertAid First Strand cDNA Synthesis Kit (MBI). For RT-PCR analysis, the cDNA sample was used to amplify an iaaM segment with primers: IAAM1, 5′-AAGGTA GCAGTTCTCTCCGC-3′ and IAAM2, 5′-TCGGCTTAGGAACATCCT CC-3′. HISTONE3 (AF024716) was used as the internal control with primers GhHIS1 (5′-GAAGCCTCATCGATACCGTC-3′) and GhHIS2 (5′-CT ACCACTACCATCATGGC-3′). Real-time quantitative RT-PCR performed in an iCycler Multicolor Real-Time PCR Detection System (Bio-Rad).
IAA content determination. IAA extraction was performed as the modified method35. Ovules (~0.5 g) were frozen with liquid nitrogen, homogenized and extracted with 80% (vol/vol) methanol containing 10 ng 13C6-IAA (CIL) as the internal standard. The supernatant was evaporated then redissolved in the aqueous phase, further purified on Sep-Pak Plus tC18 cartridge (Waters). The eluant was dried and stored at −20 °C. IAA was determined in the College of Life Science and Biotechnology, Beijing Forestry University, China.
Microscopic observations. Ovules at mid-position in the cotton boll were col-lected for microscopic observations. Sample preparation of paraffin-embedded sections was done as described36. For scanning electron micrography, sample pretreatment followed a published method37. The samples were observed by an S-3000N scanning electron microscope (Hitachi).
In situ hybridization. A 300 bp segment was amplified (primer: IAAM3, 5′-TGTTTTTCTTCCTCGAGCGT-3′; IAAM4, 5′-ATGTGTGCCCAGAAA GATCC-3′) from iaaM and subcloned into the pGEM-T Easy vector (Promega). The plasmid was used for preparing probes according to the manual of DIG RNA Labeling Kit (SP6/T7) (Roche). Paraffin sections were treated according to the method described36. Hybridization was carried out overnight at 50 °C with the probes (10 µg/ml). Signals were detected by using the anti-DIG-AP conjugate (Roche), which had been diluted to 1:1,000.
Immunohistochemical localization of IAA. The IAA immunolocalization procedure was performed as described21. For immunofluorescence, the sec-tions were incubated with FITC-labeled secondary antibody (Abcam) instead. After rinsing twice in RSR and once in PBS (pH7.4), the slides were mounted and photographed on a confocal scanning system (TCP SP2, Leica).
Fiber count. Fiber initials were counted at the similar region on 0-DPA ovules with an area of 250 × 250 µm2 of scanning electron micrograph. Nine ovules from three bolls (three ovules per boll) were counted for each line. For the counting of lint fibers per seed (N1), fibers from 20 mature seeds were gath-ered and weighed (W1). A subsample of ~10 mg fiber was randomly taken, then divided into six bundles, and each bundle was weighed (W2). The fiber
bundles were put in boiled water for 10 min and then cooled to 25 °C. The mid-dle part of the bundle was cut into three segments (~1 mm for each). Snippets of each segment was separated in six drops of 45% (vol/vol) acetic acid on glass slides then photographed. The snippet number (N2) in each drop was counted. According to the number/weight ratio of the bundles, the number of lint fibers per seed was determined by the formula: N1 = (W1/20)/(W2/N2).
Field trial. The experimental design of the field trial followed the requirements of National Cotton Variety Regional Test Project (China, http://pg.natesc.gov.cn/sites/pg/). Plants were grown in the experimental farm at Southwest University (Chongqing, China) in 2007, 2008, 2009 and 2010, using a rand-omized complete block design with three replications. In the 2007, 2008 and 2009 trials, each plot contained 40 plants grown in four rows with 1 m space (10 plants in a row with 0.45 m space). All mature/open bolls in each plot were harvested every 10 d from August 21 to October 10. After ginning, fibers and cotton seeds were weighed, and the lint percentage (fiber weight/seed cotton weight) of the cotton bolls in each harvest was determined. Fiber samples for the measurement of fiber quality were taken (>10 g each) randomly from the fibers of each harvest. The fiber samples were then sent to the Center of Cotton Fiber Quality Inspection and Testing, Chinese Ministry of Agriculture (Anyang, Henan province, China) for quality measurement. In 2010, the plot size of the field trial was increased to 100 m2 with the same plant density.
Statistical analysis. The data of lint percentage and micronaire value within each year were first treated as repeated measurements as multiple samples were taken on the same experimental plot over time. A univariate split-plot model was used for the analysis of variance (ANOVA). F-tests for effect of Time and Genotype × Time interaction were reexamined using conservative degrees of freedom. Then the data of lint percentage and micronaire taken from multiple sampling were averaged for each year. Combined ANOVA over years were performed based on the mean of lint percentage and micronaire value (over sampling times), lint yield and seed cotton yield using PROC ANOVA of the Statistical Analysis System software (SAS Institute). Because the number of genotypes differed over years, the combined ANOVA were analyzed separately for the 2-year data (2007 and 2008; three transgenic lines and one control) and 3-year data (2007, 2008 and 2009; two transgenic lines and one control). For both analyses, the following mixed model, with genotype as fixed effects, and year as a random effect, was assumed:
Y M S R S G GS eijkl i j i k ik ijkl= + + + + + ( ) ( )( )
where:Yijkl = the measurement on plot (l) in year (i), block (j), containing geno-type (k)M = the overall mean of all plots in all yearsSi = the effect of year (i)R(S)j(i) = the effect of replication (j) within year (i)Gk = the effect of genotype (k)GEik = the interaction of genotype (k) with year (i)eijkl = the plot residualMean separation tests used Tukey’s range test (SAS Institute).
34. Wan, C.Y. & Wilkins, T.A. A modified hot borate method significantly enhances the yield of high-quality RNA from cotton (Gossypium hirsutum L.). Anal. Biochem. 223, 7–12 (1994).
35. Gou, J. et al. Gibberellins regulate lateral root formation in Populus through interactions with auxin and other hormones. Plant Cell 22, 623–639 (2010).
36. Marrison, J.L. & Leech, R.M. The subcellular and intra-organelle recognition of nuclear and chloroplast transcripts in developing leaf cells. Plant J. 6, 605–614 (1994).
37. Sun, Y. et al. Brassinosteroid regulates fiber development on cultured cotton ovules. Plant Cell Physiol. 46, 1384–1391 (2005).
© 2
011
Nat
ure
Am
eric
a, In
c. A
ll ri
gh
ts r
eser
ved
.
Related Documents