P.S.Z.N.: Marine Ecology, 19 (I): 5 3 4 6 (1998) 0 1998 Blackwell Wissenschafts-Verlag, Berlin ISSN 01 73-9565 Accepted: January 16, 1998 Spatial Distribution of Shelf- and Slope- Dwelling Fishes Collected by Bottom Longline off Lanzarote and Fuerteventura, Canary Islands FRANZ UIBLEIN', FERNANDO BORDES~, RICARDO CASTILLO~ & ANTONIO G. RAMOS~ ' Institut fiir Zoologie der Universitat Salzburg, Hellbrunnerstr. 34, A-5020 Salzburg, Austria. E-mail: [email protected] 'Institute Canario de Ciencias Marinas, Aptdo. de Correos 56, E-35200 Telde, Gran Canaria, Spain. 'Departamento de Biologia, Faculdad de Ciencias del Mar, Universidad de Las Palmas de Gran Canaria, P.B. 550, Las Palmas de Gran Canaria, Spain. With 6 figures and 2 tables Key words: Depth distribution, depth-size, spawning aggregation, reproductive timing, Mora moro, topography, hydrography, local upwelling, satellite imagery. Abstract. A total of 1789 fish belonging to 38 families and 73 species were collected at depths between 18 and 1 102 m during 216 bottom longline operations off Lanzarote and Fuerteventura, Canary Islands, between February 1994 and December 1995. For each species the depth distribution is provided. Length-weight and depth-size relationships are reported for three shelf-dwelling species. The bigger- deeper relationship found in two of them contrasts with the bigger-shallower pattern of the deeper living trichiurid Lepidopus caudurus. In November 1997, nine additional bottom longline operations were carried out of€ eastern Fuerteventura at depths between 805 and 1217 m. In this area, after earlier studies in October 1995, a spawning aggregation of the morid Mom moro was encountered for the second time. The catches of 1997 revealed a strongly male-biased sex ratio. Also, the males showed a significantly lower gonadosomal index than two years earlier. These findings indicate slight interannual variations in reproductive timing and an earlier arrival of male Mora moro at the spawning grounds. Clear variations in the number of fish collected at adjacent sites possibly reflect a preference for distinct microhabitats. Preliminary evidence of local upwelling of cold water above the spawning grounds is provided by satellite imagery. Problem The Canary Islands region, situated in the northernmost part of the Eastern Central Atlantic, shows distinctive topographic and hydrographic conditions which may affect the spatial distribution of both shallow- and deep-water fishes to a con- siderable extent. The rather narrow shelf and the steep upper slope, typical for oceanic islands of volcanic origin, set spatial limits to the demersal fish assemblage. The hydrography of this region is characterized by countercurrents at different U. S. Copyright Clearance Center Code Statement: 0173-9565/98/1901-0053!$14.00/0

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

P.S.Z.N.: Marine Ecology, 19 (I): 5 3 4 6 (1998) 0 1998 Blackwell Wissenschafts-Verlag, Berlin ISSN 01 73-9565
Accepted: January 16, 1998
Spatial Distribution of Shelf- and Slope- Dwelling Fishes Collected by Bottom Longline off Lanzarote and Fuerteventura, Canary Islands FRANZ UIBLEIN', FERNANDO BORDES~, RICARDO CASTILLO~ & ANTONIO G. RAMOS~
' Institut fiir Zoologie der Universitat Salzburg, Hellbrunnerstr. 34, A-5020 Salzburg, Austria. E-mail: [email protected]
'Institute Canario de Ciencias Marinas, Aptdo. de Correos 56, E-35200 Telde, Gran Canaria, Spain.
'Departamento de Biologia, Faculdad de Ciencias del Mar, Universidad de Las Palmas de Gran Canaria, P.B. 550, Las Palmas de Gran Canaria, Spain.
With 6 figures and 2 tables
Key words: Depth distribution, depth-size, spawning aggregation, reproductive timing, Mora moro, topography, hydrography, local upwelling, satellite imagery.
Abstract. A total of 1789 fish belonging to 38 families and 73 species were collected at depths between 18 and 1 102 m during 216 bottom longline operations off Lanzarote and Fuerteventura, Canary Islands, between February 1994 and December 1995. For each species the depth distribution is provided. Length-weight and depth-size relationships are reported for three shelf-dwelling species. The bigger- deeper relationship found in two of them contrasts with the bigger-shallower pattern of the deeper living trichiurid Lepidopus caudurus. In November 1997, nine additional bottom longline operations were carried out of€ eastern Fuerteventura at depths between 805 and 1217 m. In this area, after earlier studies in October 1995, a spawning aggregation of the morid Mom moro was encountered for the second time. The catches of 1997 revealed a strongly male-biased sex ratio. Also, the males showed a significantly lower gonadosomal index than two years earlier. These findings indicate slight interannual variations in reproductive timing and an earlier arrival of male Mora moro at the spawning grounds. Clear variations in the number of fish collected at adjacent sites possibly reflect a preference for distinct microhabitats. Preliminary evidence of local upwelling of cold water above the spawning grounds is provided by satellite imagery.
Problem
The Canary Islands region, situated in the northernmost part of the Eastern Central Atlantic, shows distinctive topographic and hydrographic conditions which may affect the spatial distribution of both shallow- and deep-water fishes to a con- siderable extent. The rather narrow shelf and the steep upper slope, typical for oceanic islands of volcanic origin, set spatial limits to the demersal fish assemblage. The hydrography of this region is characterized by countercurrents at different
U. S. Copyright Clearance Center Code Statement: 0173-9565/98/1901-0053!$14.00/0

54 UIBLEIN, BORDES, CASTILL0 & RAMOS
depth zones with different temperature and salinity regimes and a strong upwelling activity off the adjacent north-west African coast (e.g., MANN & LAZIER, 1991). Compared to topography, the effects imposed by water temperature, salinity or current velocity on fish distribution patterns may be less constraining. However, many fish species have been observed to migrate vertically or horizontally in response to local or temporal variations of water temperature (e.g., NEILSON &
During the last years, efforts have been increased in the Canary Islands to study the ecology of shelf- and slope-dwelling fish in detail (GONZ~LEZ et al., 1988; BORDES et al., 1991; BRITO el al., 1993; BORDES et al., 1995; UIBLEIN et al., 1996). In one of these studies, the results of four bottom longline cruises off Lanzarote and Fuerteventura with special emphasis on the deep-water ichthyofauna from 100 to 1 100 m depth were reported and discussed with respect to possible environmental effects on diversity, depth distribution and local abundance (UIBLEIN et al., 1996). At depths of less than 100 m, an additional 48 bottom longline operations were carried out during these cruises. The present study analyses the results from these shallow-water catches and provides a complete overview of the depth distribution and abundance of fish species collected at a total of 216 localities between 18 and 1 102 m. Furthermore, depth-size relationships of the most abundant shelf-dwelling species are determined and compared with the findings from deeper zones.
A special effort was made to more closely investigate a locality off Gran Tarajal, eastern Fuerteventura. In October 1995, a spawning aggregation of the morid Mora rnoro was discovered there at about 1000 m depth (UIBLEIN et al., 1996). Two years later, in early November 1997, nine bottom longline operations were carried out to examine if this spawning aggregation occurs consistently in the same area and during the same season. The results from this fishing experiment were compared with those obtained in October 1995. To detect possible inter- relationships between the occurrence of the spawning aggregation and local hydrography, satellite-derived sea-surface temperature images were examined.
PERRY, 1990; UIBLEIN & JAGSCH, 1994; h M O S et d., 1996).
Material and Methods
The collections were made during four cruises with the local fishing vessel Alkartasuna, a boat of 12 m length and an engine power of 85 kW, It is equipped with a Furuno 50 kHz sounder, a Furuno GPS/Plotter as position indicator, and an OSA hydraulic winch. Between February 1994 and December 1995 a total of 216 localities off Lanzarote and Fuerteventura (Fig. I ) were sampled by bottom longline at depths between 18 and 1102 m. The gear was of the following type: the main line, a 2 mm diameter nylon rope (600 m length) was anchored at intervals of 50 m by stones weighing 500-600 g and at its ends by stones with 3-4 kg weight. Every two meters a 0.9 mm diameter snood, 100 cm long and terminating in a MUSTAD 3/0 hook, was attached to the main line by a swivel, giving a total of 300 hooks. The bait used was imported Cololabis saira (Scomberesocidae; 20 cm total length), cut into four longitudinal pieces of fillet. The time that the longline was set at the bottom ranged between 23 and 45 min (mean 30.9 min).
On 6-8 November 1997, during cruise “La Bocaina 11-97” a bottom longline experiment consisting of nine hauls was carried out off eastern Fuerteventura at depths between 805 and 1217 m (Fig. I ) . In contrast to the longline operations carried out earlier, a slightly modified gear was used which allowed sampling of a larger bottom area: the main line had a total length of 1000 m, resulting in larger intervals for the anchoring stones and swivels. Size and number of hooks were identical with the gear described above. The bottom resting time of the longline ranged between 45 and 46 min.

Spatial distribution of demersal fishes 55
1sw 45' 30' 15 14' 45' 30'
30'
15'
0 100 - 3W m A 3W - 500 m 0 500 - 700 m 0 'IW - 800 m
28"
30 IS' _ _
15.w 45' 30' 15' 14'
Fig. 1. Chart of the survey area off Fuerteventura and Lanzarote, Canary Islands, indicating longline station positions for six different depth zones sampled during the four earlier cruises (1994-1995) and the stations of the nine longline operations carried out during cruise "La Bocaina 11-97".
The exact station position, initial and final depth and duration of the respective longline operation, species identity and number of individuals were recorded for each operation. The samples were stored in ice. Family identity was determined according to NELSON (1 994) and species identity was re-examined (based on WHITEHEAD et al., 1984-86; BRITO, 1991; NAKAMURA & PARIN, 1993; FRANQUET & BRITO, 1995; among others). In addition, total length and total weight were deteimined.
An additional depth zone of less than 100 m was added to the five depth zones already described by UIBLEIN ef al. (1996). This resulted in a total of six different depth zones, with Zone 0 ranging from 0- 100 m, Zone 1 from 100-300 m, Zone 2 from 300-500 m, Zone 3 from 500-700 m, Zone 4 from 700- 900 m and Zone 5 from 900-1 100 m.
Length-weight relationships were calculated using the regression function
where W is the total weight and L is the total length. To detect significant depth-size relationships, the most abundant species (n > 60) were analysed using
two different methods, SPEARMAN rank correlation based on the raw data set and linear regression

56 UIBLEIN, BORDES, CASTILLO & RAMOS
function based on the geometric mean weight and length per haul. Hauls consisting of only one individual were excluded from the latter analysis.
The species collected in the longline experiment were identified and of all M o m moro collected, sex, total length, total weight and gonad weight were determined.
The gonadosomal index (IG)
10 = 1oo'WG*W-'
was calculated, where WG is the gonad weight and W is the total weight. In addition, ovarian maturity stage based on macroscopic characteristics was determined (c.f. UIBLEIN et a/., 1996).
Between August 1996 and November 1997, satellite imagery of the Advanced Very High Resolution Radiometer (AVHRR/NOAA) was received from a ground station (UTIS-ULPGC) and processed. Radiance values of channels 4 (10.5-11.3 nm) and 5 (11.5-12.5 nm) were converted to sea-surface temperatures and corrected for atmospheric effects using a multichannel algorithm. SST maps were then geometrically corrected using orbital parameters and fixed points on the ground and completed with a legend and frame. Due to frequent cloud accumulations in the study area only a relatively small sample of good quality images appropriate to show the surface temperature conditions was obtained. From this sample four representative images from different seasons or years were selected.
Results
Of the total 216 bottom longline operations carried out between February 1994 and December 1995, 175 (81%) were successful. A total of 1789 fishes were collected, resulting in 1627.9 kg of fish and means of 10.2 individuals and 9.3 kg fish per successful haul. Of the total 48 hauls carried out in Zone 0 (0-100 m depth), 47 (97.9%) resulted in the capture of at least one fish specimen. Six hundred twenty-two individuals belonging to 38 species and 19 families were collected in this depth zone, with an average of 13 individuals and 1 1.38 kg of fish per successful haul. Compared to the catches from the deeper zones studied earlier (UIBLEIN et al., 1996), the results from Zone 0 closely resemble those of Zone 1 and clearly differ from the much lower capture success at depths between 500 and 900 m (Zone 3, Zone 4). They also differ from the increasingly lower numbers of species and families collected with increasing depth from Zone 2 (300-500 m) to Zone 5 (900- 1100 m).
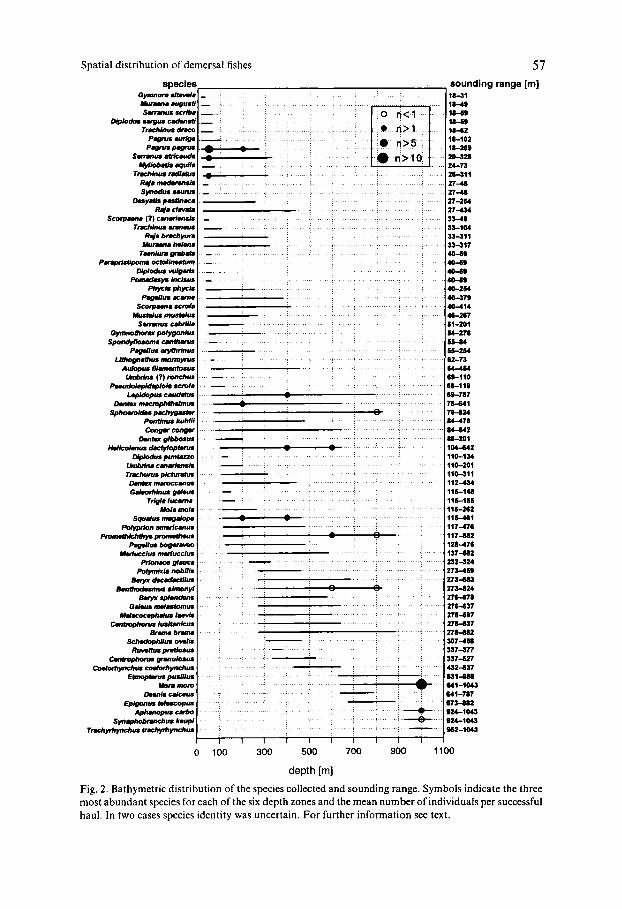
Four families are new among those 34 reported by UIBLEIN et al. (1996): Gym- nuridae (Gymnura altavela), Myliobatidae (Myliobatis aquila), Synodontidae (Syn- odus saurus) and Haemulidae (Parapristipoma octolineatum). The depth distri- bution of all 73 species collected in the six depth zones, with the most abundant ones emphasized, is shown in Fig. 2.
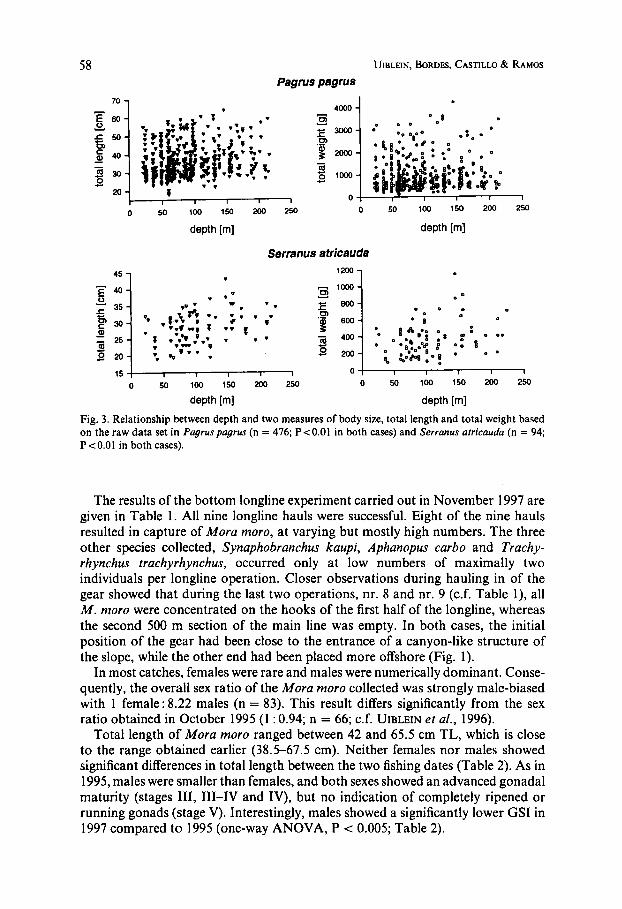
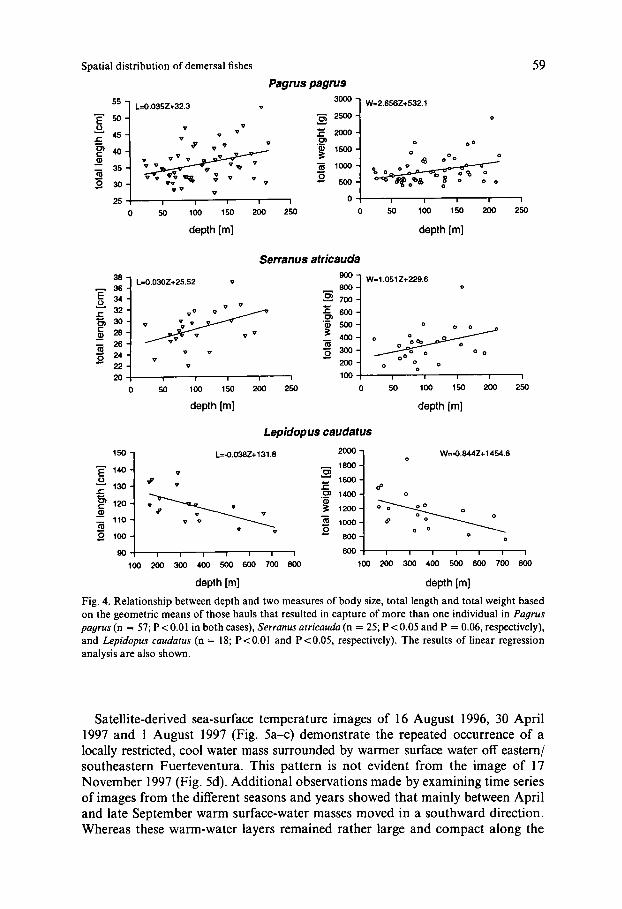
The capture of a great number of Serranus atricauda, Trachinus radiatus and Pagrus pagrus in Zone 0 enabled a detailed analysis of length-weight and depth- size relationships; this supplements earlier statistical treatments of the data from deep-water fish (UIBLEIN et al., 1996). The following length-weight relationships were obtained: S. arricauda: W = 0.0081 L3.'602 (n = 94, 19-43 cm TL); T . radiatus: W = 0.0035 L3.2923 (n = 93,23-54 cm TL); P. pagrus: W = 0.0184 L2.969' (n = 476, 18-64 cm TL). Both methods of correlation analysis revealed a clear bigger-deeper pattern in S. atricauda and P . pagrus (Figs 3 and 4). For T. radiatus no significant correlations were discovered. These findings contrast with the depth-body size relationship of Lepidopus caudatus, which indicates a bigger-shallower pattern (Fig. 4; see also UIBLEIN et al., 1996).
depth [m]
Fig. 2. Bathymetric distribution of the species collected and sounding range. Symbols indicate the three most abundant species for each of the six depth zones and the mean number of individuals per successful haul. In two cases species identity was uncertain. For further information see text.
1200 - 45 - . a loO0- .. E 800-
$ Q 400-
s 200-
Y . * w 7s 40- .* s
cn 600- c 35- V. 8x3 -. >: T V .-
25- . 8 .:.::.; v - ' c
... V W I : .. g - F? 30- ,o 20- - b ' V . .
a . - M
15 I I 1 I t 0
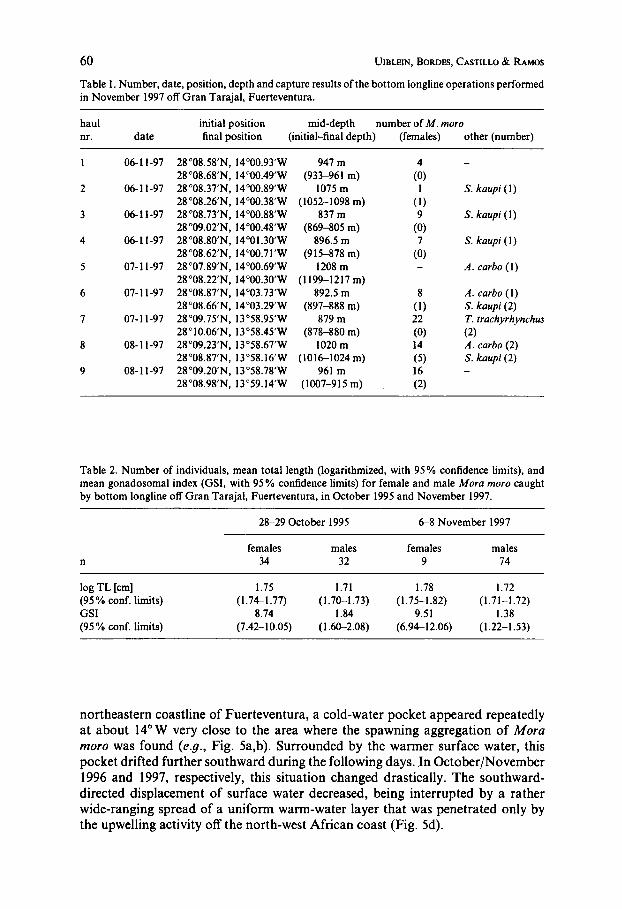
The results of the bottom longline experiment carried out in November 1997 are given in Table 1. All nine longline hauls were successful. Eight of the nine hauls resulted in capture of Mora moro, at varying but mostly high numbers. The three other species collected, Synaphobranchus kaupi, Aphanopus carbo and Trachy- rhynchus trachyrhynchus, occurred only at low numbers of maximally two individuals per longline operation. Closer observations during hauling in of the gear showed that during the last two operations, nr. 8 and nr. 9 (c.f. Table l), all M . moro were concentrated on the hooks of the first half of the longline, whereas the second 500 m section of the main line was empty. In both cases, the initial position of the gear had been close to the entrance of a canyon-like structure of the slope, while the other end had been placed more offshore (Fig. 1).
In most catches, females were rare and males were numerically dominant. Conse- quently, the overall sex ratio of the Mora moro collected was strongly male-biased with 1 female: 8.22 males (n = 83). This result differs significantly from the sex ratio obtained in October 1995 (1 : 0.94; n = 66; c.f. UIBLEIN et al., 1996).
Total length of Mora moro ranged between 42 and 65.5 cm TL, which is close to the range obtained earlier (38.5-67.5 cm). Neither females nor males showed significant differences in total length between the two fishing dates (Table 2). As in 1995, males were smaller than females, and both sexes showed an advanced gonadal maturity (stages 111, 111-IV and IV), but no indication of completely ripened or running gonads (stage V). Interestingly, males showed a significantly lower GSI in 1997 compared to 1995 (one-way ANOVA, P < 0.005; Table 2).
0 0 -
0 8
D " i ; :8 :: 0 0 0
0 ~ .&y : 0 0 I 0 0 0 B & - . O !
I I 1 1 I
Spatial distribution of demersal fishes
Pagrus pagrus
.- 0 0 0 g 1500- 0
- 35 m - 500-
25 0 I I I I I
0 50 100 150 200 250 0 50 100 150 200 250
depth [m] depth [m]
38- - 38- 3 2 -
El 34-
F ": 3 = - (1, 28
- 22,
- ij 24 -
L&O3OZ+25.52 V
P P P
P
Serranus atricauda
2 0 1 0 50 100 150 200 250
59
0
- 800-
W=1.051Z+229.6 - 9 700-
.- (1,500
m 5 300- 0 0 0 c
100 1 1 1 I 1
0 50 100 150 200 250
depth [m] depth [m]
Lepidopus caudatus
'50 1 L=4.038Z+131.8 1 - 1 4 0 - - 130- 5
z 110 - s l o o -
E p 120- (1,
m c
0
+ v
W=4.844Z+1454.6 0
100 200 300 400 500 600 700 800 100 200 300 400 500 600 700 800
depth [m] depth [m]
Fig. 4. Relationship between depth and two measures of body size, total length and total weight based on the geometric means of those hauls that resulted in capture of more than one individual in Pagrus pagrus (n = 57; P i 0.01 in both cases), Serranus atricauda (n = 25; P <0.05 and P = 0.06, respectively), and Lqidopus caudatus (n = 18; P<O.OI and P<0.05, respectively). The results of linear regression analysis are also shown.
Satellite-derived sea-surface temperature images of 16 August 1996, 30 April 1997 and 1 August 1997 (Fig. 5a-c) demonstrate the repeated occurrence of a locally restricted, cool water mass surrounded by warmer surface water off eastern/ southeastern Fuerteventura. This pattern is not evident from the image of 17 November 1997 (Fig. 5d). Additional observations made by examining time series of images from the different seasons and years showed that mainly between April and late September warm surface-water masses moved in a southward direction. Whereas these warm-water layers remained rather large and compact along the
60 UIBLEIN, Bowes, CASTILLO & RAMOS
Table I . Number, date, position, depth and capture results of the bottom longline operations performed in November 1997 off Gran Tarajal, Fuerteventura.
haul initial position mid-depth number of M . moro nr. date final position (initial-final depth) (females) other (number)
06-1 1-97
06-1 1-97
06-1 1-97
06- I 1-97
07-1 1-97
07-1 1-97
07-1 1-97
08-1 1-97
08-1 1-97
28"08.58'N, 14"00.93'W 28 "08.68'N. 14"00.49'W 28"08.37'N, 14"00.89'W 28"08.26", 14"00.38'W 28"08.73'N, 14"00.88'W 28"09.02'N, 14"00.48'W 28 "08.80", I4 "0 1.30'W 28 "08.62'N. l4OOO.7 1 'W 28 "07,89'N, 14"00.69'W 28 "08.22". 14"00.30'W 2So08.87'N, 14"03.73'W 28 "08.66". 14 "03.29'W 28 "09.75'N, 13 "58.95'W 28"10.06", 13"58.45'W 28O09.23". 13"58.67'W 28 "08.87", 13"58.16W 28O09.20". 1338.78'W 28'08.98". 13"59.14W
947 m (933-961 m)
1075 m (1052-1098 m)
837 m (869-805 m)
896.5 m (915-878 m)
1208 m (1199-1217 m)
892.5 m (897-888 m)
879 m (878-880 m)
1020 m (1016-1024 m)
961 m (1 007-9 15 m)
4
S. kaupi ( I )
S. kaupi ( I )
S. kaupi ( 1)
A . carbo ( I )
A . carbo ( I ) S. kaupi (2) T. trachyrhynchus (2) A . carbo (2) S. kaupi (2) -
Table 2. Number of individuals, mean total length (logarithmized, with 95% confidence limits), and mean gonadosomal index (GSI, with 95% confidence limits) for female and male Mora moro caught by bottom longline off Gran Tarajal, Fuerteventura, in October 1995 and November 1997.
28-29 October 1995 6-8 November 1997
n females males females males
34 32 9 74
log TL [cm] 1.75 1.71 1.78 1.72
GSI 8.74 1.84 9.51 1.38 (95% conf. limits) (1.741.77) (1.70-1.73) (1.75482) (1.71-1.72)
(95% conf. limits) (7.42-10.05) (1.60-2.08) (6.94-12.06) (1.22-1.53)
northeastern coastline of Fuerteventura, a cold-water pocket appeared repeatedly at about 14" W very close to the area where the spawning aggregation of Mora moro was found (e.g., Fig. 5a,b). Surrounded by the warmer surface water, this pocket drifted further southward during the following days. In October/November 1996 and 1997, respectively, this situation changed drastically. The southward- directed displacement of surface water decreased, being interrupted by a rather wide-ranging spread of a uniform warm-water layer that was penetrated only by the upwelling activity off the north-west African coast (Fig. 5d).

Spatial distribution of demersal fishes 61
Fig. 5. Satellite-derived sea-surface temperature in the region of Lanzarote and Fuerteventura on 16 August 1996 (a), 30 April 1997 (b), 01 August 1997 (c) and 17 November 1997 (d).
Discussion
Among the 73 fish species collected, four major bathymetric groups can be roughly distinguished: shelf dwellers with rather narrow depth ranges (species occurring exclusively at less than about 200 m depth, e.g., Serranus scriba), shelf and upper slope dwellers with rather wide depth ranges (e.g., Squalus megalops), upper slope dwellers (e.g., Beryx decadactyfus) and middle slope dwellers (e.g., Trachyrhynchus trachyrhynchus). Further representatives of the three latter bathymetric groups
UIBLEIN, BORDES, CASTILLO & RAMOS 62
0 100 200 300 400 500 600 700 800 800
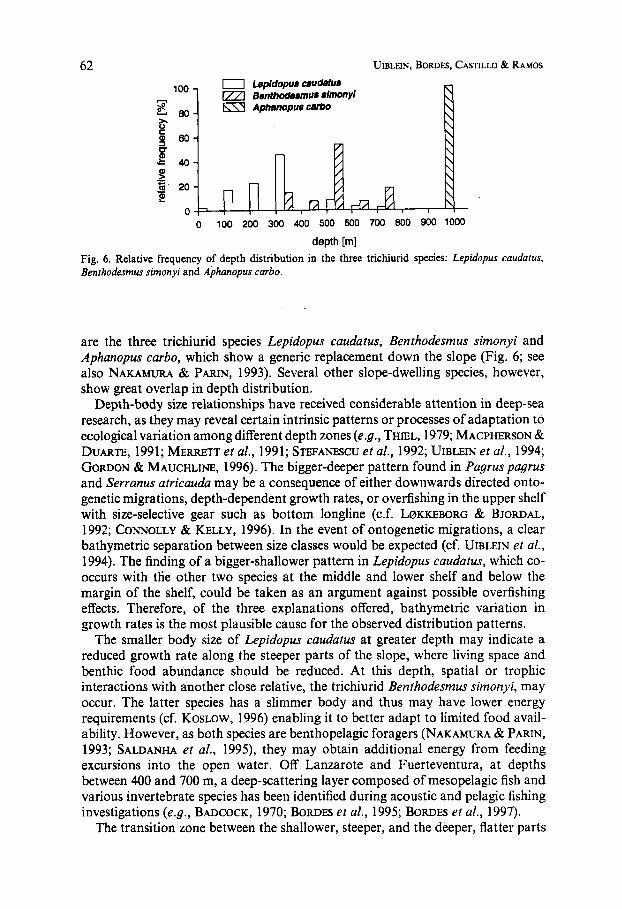
depth [m] Fig. 6. Relative frequency of depth distribution in the three trichiurid species: Lepidopus caudafus, Benthodesmus simonyi and Aphanopus carbo.
are the three trichiurid species Lepidopw caudatus, Benthodesmus simonyi and Aphanopus carbo, which show a generic replacement down the slope (Fig. 6; see also NAKAMURA & Pmm, 1993). Several other slope-dwelling species, however, show great overlap in depth distribution.
Depth-body size relationships have received considerable attention in deep-sea research, as they may reveal certain intrinsic patterns or processes of adaptation to ecological variation among different depth zones (e.g., THIEL, 1979; MACPHERSON & DUARTE, 1991; MERRETT et al., 1991; STEFANESCU et al., 1992; UIBLEIN et al., 1994; GORDON & MAUCHLINE, 1996). The bigger-deeper pattern found in Pagrus pagrus and Serranus atricauda may be a consequence of either downwards directed onto- genetic migrations, depth-dependent growth rates, or overfishing in the upper shelf with size-selective gear such as bottom longline (c.f. L ~ K K E B ~ R G & BJORDAL, 1992; CONNOLLY & KELLY, 1996). In the event of ontogenetic migrations, a clear bathymetric separation between size classes would be expected (cf. UIBLEIN et al., 1994). The finding of a bigger-shallower pattern in Lepidopus caudatus, which co- occurs with tlie other two species at the middle and lower shelf and below the margin of the shelf, could be taken as an argument against possible overfishing effects. Therefore, of the three explanations offered, bathymetric variation in growth rates is the most plausible cause for the observed distribution patterns.
The smaller body size of Lepidopus caudatus at greater depth may indicate a reduced growth rate along the steeper parts of the slope, where living space and benthic food abundance should be reduced. At this depth, spatial or trophic interactions with another close relative, the trichiurid Benthodesmus sinioiiyi, may occur. The latter species has a slimmer body and thus may have lower energy requirements (cf. KOSLOW, 1996) enabling it to better adapt to limited food avail- ability. However, as both species are benthopelagic foragers (NAKAMURA & PARIN, 1993; SALDANHA et al., 1995), they may obtain additional energy from feeding excursions into the open water. Off Lanzarote and Fuerteventura, at depths between 400 and 700 m, a deep-scattering layer composed of mesopelagic fish and various invertebrate species has been identified during acoustic and pelagic fishing investigations (e.g., BADCOCK, 1970; BORDES et al., 1995; BORDES et al., 1997).
The transition zone between the shallower, steeper, and the deeper, flatter parts

Spatial distribution of dernersal fishes 63
of the slope - as it occurs off eastern Fuerteventura at ca. 850 to 1000 m depth - may be a particularly interesting habitat for demersal fishes to settle. This zone provides more living space and probably also an increased food intake via slumps, turbidity currents, or other ways of rapid transport from the shelf and the steeper parts of the upper slope (c.f. GORDON, 1979). In this zone, the spawning aggregation of Mora moro was encounted for the second time in November 1997. However, as the present study suggests, this aggregation is not uniformly distributed, but shows considerable fluctuations in fish density among adjacent sites. Such a “patchy” distribution pattern may reflect the selection of distinct microhabitats within the same depth zone and area. In this respect the observation of an increased abundance close to the entrance of a submarine canyon is of particular interest.
In November 1997, as in October 1995, Mora moro showed an advanced gonadal maturity. However, the lower gonadosomal index of the males indicates a delay in the timing of reproduction in 1997. In contrast to such slight interannual changes in reproductive timing within one area, Mora moro may show considerable vari- ations in spawning seasonality among different regions of the Atlantic. In a study on the biology of three morid species living on the continental slope of the Rockall Trough (north-east Atlantic, 53”N), GORDON & DUNCAN (1985) found clear indi- cations of a spawning peak for Mora moro in June/July. Such geographical vari- ation in spawning seasonality could reflect an environmentally induced, phenotypically plastic response (WINTERS & WHEELER, 1996) or may be genetically based (BYE, 1984). In the north-east Atlantic there is considerable latitudinal variation in seasonal food input from the surface which - apart from effects on species composition, foraging behaviour, growth and fecundity (MERRETT, 1987) - may also influence the timing of reproduction in deep-sea fishes.
Whereas males were highly abundant in the 1997 longline catches, females were much less frequent compared to the findings of two years before. As bottom longline operations carried out in the same depth zone and area during other seasons led to the capture of only a few M. moro (UIBLEIN et al., 1996), the following scenario of two temporally separated, sex-specific spawning migrations may be hypothesized: first, the males travel to this area, select certain microhabitats, and terminate the last phase of gonad growth; later, the females arrive at these sites and then spawning takes place over a period of a few weeks. In another gadiform species, the macrourid Coryphaenoides acrolepis, it has been suggested that the males hold territories and females move up the slope to these areas to spawn (see GORDON, 1979).
This study provides preliminary evidence for the formation of a cold-water pocket off eastern Fuerteventura. This indicates a local upwelling activity just above the spawning grounds of Mora moro. Several factors have been identified to cause upwelling such as opposing undercurrents, shelf breaks, shelf-edge canyons, strong winds, capes and coastal mountains (HUTHNANCE, 1995). Most of these factors also deserve consideration for the area under study.
In gadiform fish, local hydrography has been suggested to be associated with the formation of spawning aggregations. Hoki (Macruronus novaezelandiae), for instance, spawn in the eastern Cook Strait, New Zealand, in dense concentrations in deep canyons at more than 200 m depth; here deep currents and local upwelling carry the eggs and larvae into shallow coastal regions of Clifford Bay (LIVINGSTON, 1990; MURDOCH er al., 1990). Mora moro spawn at much greater depths than hoki,

64 UIBLEIN, BORDES, CASTILLO & RAMOS
where upwelling may be less influential or exert no effects at all. Furthermore, the upwelling activity off eastern Fuerteventura seems to diminish before the start of the spawning period. Fine-scaled investigations of the hydrography, topography and nutrient input at and in the immediate surroundings of the spawning grounds are needed to identify the environmental cues these deep-sea fish may use to locate the spawning area and to aggregate and reproduce there during a limited period of the year.
Summary
(1) An analysis of 48 bottom longline operations from depths of less than 100 m is accompanied by a complete overview of the results of four bottom longline cruises carried out off Lanzarote and Fuerteventura between February 1994 and December 1995 at 216 localities between 18 and 1102 m depth. A total of 1789 fishes belonging to 38 families and 73 species were collected. The depth distribution for each species is provided, with the most abundant ones being emphasized. (2) For the species encountered at high frequency at less than 100 m, the length- weight and depth-size relationships are reported: Two inhabitants of the shelf and uppermost parts of the upper slope, Pagruspagrus and Serranus atricauda, show a bigger-deeper pattern, whereas in the trichiurid Lepidopus caudatus a bigger-shal- lower pattern had been discovered during an earlier study of the deep-water fishes collected during these cruises (UIBLEIN et al., 1996). The possible effects of the narrow shelf and a steep upper slope typical for the Canary Islands on the depth distribution and local abundance of demersal fish are discussed. (3) In November 1997, nine bottom longline operations were carried out off Gran Tarajal, eastern Fuerteventura, at depths between 805 and 1217 m. In this area, a spawning aggregation of the morid Mora moro had been discovered at about 1000 m depth during an earlier cruise in October 1995. Two years later, this spawning aggregation was encountered again. Interestingly, the 1997 catches revealed a strongly male-biased sex-ratio (1 female : 8.22 males versus 1 : 0.94 in 1995). Fur- thermore, the males showed a significantly lower gonadosomal index in 1997. These findings indicate slight interannual variations in reproductive timing and an earlier arrival of male Mora moro at the spawning grounds. Clear variations in the numbers of fish collected at adjacent sites within the study area may reflect a patchy distribution pattern within the spawning aggregation. (4) Sea-surface temperature data derived from satellite imagery indicate a zone of local upwelling of cold water just above the spawning grounds of this species. This upwelling activity, however, seems to diminish before the start of the spawning season.
Acknowledgements
We thank the crew of the fishing vessel Alkuriusunu for assistance in the field, ANTONIO BARRERA L U J ~ N and Josk A. G ~ M E Z BETHENCOURT for indispensable help in data collection, Josk CARILLO MOLINA and Feme F%REZ ARTILFS for their collaboration in the analysis of catches, and JOSEP COCA for help in image processing. We thank NIGEL MERRETT, JOHN D. M. GORDON and J ~ G E N G. NIELSEN for many

Spatial distribution of demersal fishes
valuable comments on earlier versions of the manuscript. Financial support was provided by the Gobierno de Canarias.
65
References
BADCOCK, J . , 1970: The vertical distribution of mesopelagic fishes collected on the SOND cruise. J. Mar. Biol. Assoc. U.K., 50: 1001-1044.
BORDES, F., A. BARRERA, R. CASTILLO, J . A. GOMEZ, A. OJEDA & F. PEREZ, 1991: Cartografia y evaluacion de recursos pesqueros de la plataforma y talud de Gran Canaria (Mas Canarias). Consejeria de Pesca y Transportes, Gobierno de Canarias, Telde, Las Palmas GC; 28 pp.
--, A. BARRERA, J. CARILLO, R. CASTILLO, J . A. GOMEZ, K. HANSEN, F. PEREZ & F. UIBLEIN, 1995: Cartografia y evaluacion de 10s recursos pesqueros en la plataforma y talud de Fuerteventura y Lanzarote (Mas Canarias). Consejeria de Agricultura, Pesca y Alimentacion, Telde, Las Palmas GC; 56 pp.
--, A. BARRERA, J. CARILLO, R. CASTILLO, J. J. CASTRO, J. A. GOMEZ, K. HANSEN, V. HERNANDEZ, T. MORENO, F. PEREZ & F. UIBLEIN, 1997: Evaluacion acustica de 10s recursos epipelagicos y estudio de la capa profunda de reflexion en Lanzarote, Fuerteventura y Gran Canaria (Mas Canarias). Viceconsejeria de Pesca. Consejeria de Agricultura. Pesca y Alimentacibn, Telde, Las Palmas GC;
BRITO, A., 1991: Catalog0 de 10s peces de la Mas Canarias. Ed. Francisco Lemus, La Laguna TF; 230 PP.
--, J. 1. LOZANO, R. RABANAL, P. PASCUAL, M. HERN~NDEZ, J. M. FALCON, J. A. GONZALEZ, J . 1. SANTANA, J. BARQUiN, G . GONZ~LEZ, F. I. PEREZ & V. FERN~NDEZ, 1993: Prospeccion de las poblaciones de condrictios (Tiburones) de 10s fondos batiales profundos de Canarias, con especial atencion a su posible aprovechamiento pesquero. Consejeria de Pesca y Transportes, Gobierno de Canarias, Las Palama GC; 63 pp.
BYE, V. J., 1984: The role of environmental factors in the timing of reproductive cycles. In: G . W. P o ~ r s & R. J . WOTTON (Eds.), Fish reproduction: strategies and tactics. Academic Press, London: 187- 222.
CONOLLY, P. L. & C. J. KELLY, 1996: Catch and discards from experimental trawl and longliiie fishing in the deep water of the Rockall Trough. J . Fish. Biol.. 49A: 132-144.
FRANQUET, F. & A. BRITO, 1995: Especies de interes pesquero de Canarias. Consejeria de Pesca y Transportes, Gobierno des Canarias, Telde, Las Palmas GC; 143 pp.
GONZ~LEZ. J. A,, I. J. LOZANO, M. A. CALDENTEY, J. I . SANTANA, J. A. GOMEZ & R. CASTILLO, 1988: Resultados de la campafia de prospeccion pesquera “Canarias 85”. Inf. Tic. Inst. Esp. Oceanogr.,
GORDON, J. D. M., 1979: Lifestyle and phenology in deep sea anacanthine teleosts. Synip. Zool. SOC. London, 44: 327-359.
-- & J. A. R. DUNCAN, 1985: The biology of fish of the family Moridae in the deep-water of the Rockall Trough. J. Mar. Biol. Assoc. U.K., 65: 475485.
-- & J. MAUCHLINE, 1996: The distribution and diet of the dominant, slope-dwelling eel, Syn- apliobranchus kaupi, of the Rockall Trough. J. Mar. Biol. Assoc. U.K., 7 6 493-503.
HAEDRICH, R. L. & N. R. MERRETT, 1990: Little evidence for faunal zonation or communities in deep sea demersal fish faunas. Prog. Oceanogr., 2 4 239-250.
HUTHNANCE, J. M., 1995: Circulation, exchange and water masses at the ocean margin: the role of physical processes at the shelf edge. Prog. Oceanogr., 35: 353431.
KOSLOW, J. A,, 1996: Energetic and life-history patterns of deep-sea benthic, benthopelagic and sea- mount-associated fish. J. Fish. Biol., 49A: 54-74.
LIVINGSTON, M. E., 1990: Spawning hoki (Macruronus notmzelandiue HECTOR) concentrations in Cook Strait and off the east coast of South Island, New Zealand, August-September 1987. N. Z. J . Mar. Freshwater. Res., 2 4 503-517.
LBKKEBORG, s. & A. BJORDAL, 1992: Species and size selectivity in longline fishing: a review. Fish. Res.,
MACPHERSON, E. & C. M. DUARTE, 1991: Bathynietric trends in demersal fish size: is there a general
MANN, K. H. & J . R. N. LAZIER, 1991: Dynamics of marine ecosystems. Blackwell Scientific Publi-
MERRETT, N. R. , 1987: A zone of faunal change in assemblages of abyssal demersal fish in the eastern
MURDOCH. R.. R. Guo & A. MCCRONE, 1990: Distribution of hoki (Macruronus nooaezela17diae) eggs
63 PP.
57: 1-93.
1 3 31 1-322.
relationship? Mar. Ecol. Prog. Ser., 71: 103-1 12.
cations, Boston; 466 pp.
north atlantic: a response to seasonality in production? Biol. Oceanogr., 5: 137-151.

66 UIBLEIN, BORDES, CASTILL0 & RAMOS
and larvae in relation to hydrography in eastern Cook Strait, September 1987. N . Z. J. Mar. Freshwater. Res., 2 4 529-539.
NAKAMURA, I. & N. V. PARIN, 1993: F A 0 species catalogue vol. 15. Snake mackerels and cutlass fishes of the world (Families Gempylidae and Trichiuridae). FA0 Fish. Synop., 15 1-1 36.
NEILSON, J. D. & R. 1. PERRY, 1990: Die1 vertical migrations of marine fishes: an obligate or facultative process? Adv. Mar. Biol., 26: 115-168.
NELSON, J. S . , 1994: Fishes of the World. 3rd edition. John Wiley & Sons, NY; 600 pp. RAMOS, A. G., J. SANTIAGO, P. SANGRA & M. CANTON, 1996: An application of satellite-derived sea
surface temperature data to the skipjack (Katsuwonus pelamis LINNAEUS, 1758) and albacore tuna (Thunnus alalunga BONATERRE, 1788) fisheries in the north-east Atlantic. Int. J. Remote Sensing, 4 749-759.
SALDANHA, L., A. J. ALMEIDA, F. ANDRADE & J . GUERREIRO, 1995: Observations on the diet of some slope dwelling fishes of Southern Portugal. Int. Rev. ges. Hydrobiol., 80: 217-234.
STEFANESCU, C., J. RUCABADO & D. LLORIS, 1992: Depth-size trends in western Mediterranean demersal deep-sea fishes. Mar. Ecol. Prog. Ser., 81: 205-213.
THIEL, H., 1979: Structural aspects of the deep-sea benthos. Ambio (Special Report), 6: 25-31. U m m , F. & A. JAGSCH, 1994: Wassertemperatur, Laichaktivittit und Laichplatzwahl-Beobachtungen
an der Salzach-Asche, Thyniallus rhymallus (Pisces; Salmonidae). Artenschutzreport, 4 3841. --, J. G . NlELsEN & W. KLAUSEWITZ, 1994: Depth dependent morphological variation in two ophi-
diiform fishes from the deep Red Sea: evidence for species-specific structure in vertical distribution. Cybium, 18: 15-23.
--, F. BORDES & R. CASTILLO, 1996: Diversity, abundance and depth distribution of demersal deep- water fishes off Lanzarote and Fuerteventura, Canary Islands. J. Fish Biol., 49A 75-90.
WHITEHEAD, P. J. P., M.-L. BAUCHOT, J.-C. HUREAU, J. NlELSEN & E. TORTONESE (Eds.), 1984-86: Fishes of the north-eastern Atlantic and the Mediterranean. Vol 1-111. UNESCO, Paris; 1473 pp.
WiNTERS, G . H. & J. P. WHEELER, 1996: Environmental and phenotypic factors affecting the reproductive cycle of Atlantic herring. ICES J . Mar. Sci., 53: 73-88.
Related Documents











