Spaceflight regulates ryanodine receptor subtype 1 in portal vein myocytes in the opposite way of hypertension Fabrice Dabertrand, 1,2 Yves Porte, 1,2 Nathalie Macrez, 1,2 and Jean-Luc Morel 1,2 1 Universite de Bordeaux, Institut des Maladies Neurodégénératives, and 2 CNRS, Institut des Maladies Neurodégénératives, Bordeaux, France Submitted 15 June 2011; accepted in final form 16 November 2011 Dabertrand F, Porte Y, Macrez N, Morel JL. Spaceflight regulates ryanodine receptor subtype 1 in portal vein myocytes in the opposite way of hypertension. J Appl Physiol 112: 471– 480, 2012. First published November 17, 2011; doi:10.1152/japplphysiol.00733.2011.—Gravity has a structural role for living systems. Tissue development, architec- ture, and organization are modified when the gravity vector is changed. In particular, microgravity induces a redistribution of blood volume and thus pressure in the astronaut body, abolishing an upright blood pressure gradient, inducing orthostatic hypotension. The present study was designed to investigate whether isolated vascular smooth muscle cells are directly sensitive to altered gravitational forces and, second, whether sustained blood pressure changes act on the same molecular target. Exposure to microgravity during 8 days in the International Space Station induced the decrease of ryanodine recep- tor subtype 1 expression in primary cultured myocytes from rat hepatic portal vein. Identical results were found in portal vein from mice exposed to microgravity during an 8-day shuttle spaceflight. To evaluate the functional consequences of this physiological adaptation, we have compared evoked calcium signals obtained in myocytes from hindlimb unloaded rats, in which the shift of blood pressure mimics the one produced by the microgravity, with those obtained in myo- cytes from rats injected with antisense oligonucleotide directed against ryanodine receptor subtype 1. In both conditions, calcium signals implicating calcium-induced calcium release were signifi- cantly decreased. In contrast, in spontaneous hypertensive rat, an increase in ryanodine receptor subtype 1 expression was observed as well as the calcium-induced calcium release mechanism. Taken to- gether, our results shown that myocytes were directly sensitive to gravity level and that they adapt their calcium signaling pathways to pressure by the regulation of the ryanodine receptor subtype 1 expres- sion. ryanodine receptor; calcium signaling; microgravity; hindlimb sus- pension; hypertension DURING SPACEFLIGHT, THE MICROGRAVITY (G) induces blood volume redistribution and concomitant modification of the upright blood pressure gradient in the astronaut body from 70 mmHg in the head and 200 mmHg in the lowest resistance arteries to a constant pressure near 100 mmHg in the whole body (23). Thus a decrease in blood pressure below the heart seems to be one of the first consequences of G, which can precede and trigger the adaptation of the vascular system (56). Many factors known to regulate blood pressure likely coexist in the vascular adaptation to G: 1) the control of fluid volume and associated decrease in plasma volume (45), 2) the cardiac hypofunction and the alteration of baro- and cardiopulmonary reflexes (24), and 3) the adaptation of vascular responses at endothelial and vascular smooth muscle cell (VSMC) levels. Indeed, simulated G, using the hindlimb unloaded (HU) rat model, can modify the vascular reactivity in infero-posterior vessels like abdominal aorta, mesenteric, femoral arteries, and portal vein (8, 20, 40, 47). In the animal model of hypertension, a dysregulation of vasoreactivity encoded by Ca 2 signals has been reported. Indeed, the contraction induced by depolarization is greater in spontaneously hypertensive rats (SHR) than in control rats, and a change in Ca 2 signaling pathways implicating intracellular Ca 2 stores has been evidenced (1, 29, 30). More recently, a global adaptation of Ca 2 signaling pathways was suggested in this hypertensive model (51). The vascular adaptation to changes in blood pressure is due to smooth muscle contractile activity and its regulation by endothelium, hormones, and neurotransmitters (13). In fact, the myogenic response can be considered as the equilibrium be- tween contraction and relaxation pathways. Both pathways involve Ca 2 signaling. In VSMC, ryanodine receptors (RyRs) are reticulum Ca 2 channels likely implicated in both relax- ation and contraction processes (5). Indeed, RyRs can be activa- ted via the Ca 2 -induced Ca 2 mechanism (CICR) that amplifies InsP3R and Ca 2 entry primary signals and thus participates to the cytoplasmic Ca 2 increase that can trigger contraction (22, 27). However, RyRs produce localized Ca 2 events (Ca 2 sparks), which activate Ca 2 -activated K currents leading to membrane hyperpolarization and thus relaxation through inhi- bition of L-type channels (28). Three RyR genes have been described, and, although their expression pattern in VSMC varies between species and studies, the common expression of the three subtypes (RyR1–3) has been reported in rat aorta, superior and small mesenteric arteries, and hepatic portal vein (9, 44). Each RyR subtype had a specific function in Ca 2 signals. In smooth muscle, it has been shown that both RyR1 and RyR2 are able to generate Ca 2 sparks and Ca 2 waves (9). RyR expression is modulated in physiopathological con- ditions like muscular dystrophy and after blood redistribution following a chronic position change as hindlimb unloading (8, 10, 39, 40). We designed the present study to investigate the effect of real G on RyR expression in portal veins from mice and on rat primary cultured VSMC exposed to G. To evaluate the functional effect of G exposure, we have measured Ca 2 signals observed in HU rat model. Second, we have evaluated whether the regulation observed in HU (hypotension model) could be the opposite of those observed in SHR (hypertension model). In portal vein, the expression pattern of RyR1 was decreased by exposure to G in both cultured VSMC and mouse portal vein as well as in portal vein from HU rat. We mimicked altered Ca 2 signals measured in portal vein from HU rats by in vivo injections of antisense oligonucleotide Address for reprint requests and other correspondence: J. L. Morel, IMN, UMR5293, Ave. des Facultés, 33405 Talence Cedex, France (e-mail: jean-luc.morel @u-bordeaux2.fr). J Appl Physiol 112: 471–480, 2012. First published November 17, 2011; doi:10.1152/japplphysiol.00733.2011. 8750-7587/12 Copyright © 2012 the American Physiological Society http://www.jappl.org 471 on February 2, 2012 jap.physiology.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Spaceflight regulates ryanodine receptor subtype 1 in portal vein myocytes inthe opposite way of hypertension
Fabrice Dabertrand,1,2 Yves Porte,1,2 Nathalie Macrez,1,2 and Jean-Luc Morel1,2
1Universite de Bordeaux, Institut des Maladies Neurodégénératives, and 2CNRS, Institut des Maladies Neurodégénératives,Bordeaux, France
Submitted 15 June 2011; accepted in final form 16 November 2011
Dabertrand F, Porte Y, Macrez N, Morel JL. Spaceflight regulatesryanodine receptor subtype 1 in portal vein myocytes in the opposite wayof hypertension. J Appl Physiol 112: 471–480, 2012. First publishedNovember 17, 2011; doi:10.1152/japplphysiol.00733.2011.—Gravityhas a structural role for living systems. Tissue development, architec-ture, and organization are modified when the gravity vector ischanged. In particular, microgravity induces a redistribution of bloodvolume and thus pressure in the astronaut body, abolishing an uprightblood pressure gradient, inducing orthostatic hypotension. The presentstudy was designed to investigate whether isolated vascular smoothmuscle cells are directly sensitive to altered gravitational forces and,second, whether sustained blood pressure changes act on the samemolecular target. Exposure to microgravity during 8 days in theInternational Space Station induced the decrease of ryanodine recep-tor subtype 1 expression in primary cultured myocytes from rathepatic portal vein. Identical results were found in portal vein frommice exposed to microgravity during an 8-day shuttle spaceflight. Toevaluate the functional consequences of this physiological adaptation,we have compared evoked calcium signals obtained in myocytes fromhindlimb unloaded rats, in which the shift of blood pressure mimicsthe one produced by the microgravity, with those obtained in myo-cytes from rats injected with antisense oligonucleotide directedagainst ryanodine receptor subtype 1. In both conditions, calciumsignals implicating calcium-induced calcium release were signifi-cantly decreased. In contrast, in spontaneous hypertensive rat, anincrease in ryanodine receptor subtype 1 expression was observed aswell as the calcium-induced calcium release mechanism. Taken to-gether, our results shown that myocytes were directly sensitive togravity level and that they adapt their calcium signaling pathways topressure by the regulation of the ryanodine receptor subtype 1 expres-sion.
ryanodine receptor; calcium signaling; microgravity; hindlimb sus-pension; hypertension
DURING SPACEFLIGHT, THE MICROGRAVITY (!G) induces bloodvolume redistribution and concomitant modification of theupright blood pressure gradient in the astronaut body from 70mmHg in the head and 200 mmHg in the lowest resistancearteries to a constant pressure near 100 mmHg in the wholebody (23). Thus a decrease in blood pressure below the heartseems to be one of the first consequences of !G, which canprecede and trigger the adaptation of the vascular system (56).Many factors known to regulate blood pressure likely coexistin the vascular adaptation to !G: 1) the control of fluid volumeand associated decrease in plasma volume (45), 2) the cardiachypofunction and the alteration of baro- and cardiopulmonaryreflexes (24), and 3) the adaptation of vascular responses atendothelial and vascular smooth muscle cell (VSMC) levels.
Indeed, simulated !G, using the hindlimb unloaded (HU) ratmodel, can modify the vascular reactivity in infero-posteriorvessels like abdominal aorta, mesenteric, femoral arteries, andportal vein (8, 20, 40, 47).
In the animal model of hypertension, a dysregulation ofvasoreactivity encoded by Ca2" signals has been reported.Indeed, the contraction induced by depolarization is greater inspontaneously hypertensive rats (SHR) than in control rats, anda change in Ca2" signaling pathways implicating intracellularCa2" stores has been evidenced (1, 29, 30). More recently, aglobal adaptation of Ca2" signaling pathways was suggested inthis hypertensive model (51).
The vascular adaptation to changes in blood pressure is dueto smooth muscle contractile activity and its regulation byendothelium, hormones, and neurotransmitters (13). In fact, themyogenic response can be considered as the equilibrium be-tween contraction and relaxation pathways. Both pathwaysinvolve Ca2" signaling. In VSMC, ryanodine receptors (RyRs)are reticulum Ca2" channels likely implicated in both relax-ation and contraction processes (5). Indeed, RyRs can be activa-ted via the Ca2"-induced Ca2" mechanism (CICR) that amplifiesInsP3R and Ca2" entry primary signals and thus participates tothe cytoplasmic Ca2" increase that can trigger contraction (22,27). However, RyRs produce localized Ca2" events (Ca2"
sparks), which activate Ca2"-activated K" currents leading tomembrane hyperpolarization and thus relaxation through inhi-bition of L-type channels (28). Three RyR genes have beendescribed, and, although their expression pattern in VSMCvaries between species and studies, the common expression ofthe three subtypes (RyR1–3) has been reported in rat aorta,superior and small mesenteric arteries, and hepatic portal vein(9, 44). Each RyR subtype had a specific function in Ca2"
signals. In smooth muscle, it has been shown that both RyR1and RyR2 are able to generate Ca2" sparks and Ca2" waves(9). RyR expression is modulated in physiopathological con-ditions like muscular dystrophy and after blood redistributionfollowing a chronic position change as hindlimb unloading (8,10, 39, 40).
We designed the present study to investigate the effect ofreal !G on RyR expression in portal veins from mice and onrat primary cultured VSMC exposed to !G. To evaluate thefunctional effect of !G exposure, we have measured Ca2"
signals observed in HU rat model. Second, we have evaluatedwhether the regulation observed in HU (hypotension model)could be the opposite of those observed in SHR (hypertensionmodel). In portal vein, the expression pattern of RyR1 wasdecreased by exposure to !G in both cultured VSMC andmouse portal vein as well as in portal vein from HU rat. Wemimicked altered Ca2" signals measured in portal vein fromHU rats by in vivo injections of antisense oligonucleotide
Address for reprint requests and other correspondence: J. L. Morel, IMN,UMR5293, Ave. des Facultés, 33405 Talence Cedex, France (e-mail: [email protected]).
J Appl Physiol 112: 471–480, 2012.First published November 17, 2011; doi:10.1152/japplphysiol.00733.2011.
8750-7587/12 Copyright © 2012 the American Physiological Societyhttp://www.jappl.org 471
on February 2, 2012jap.physiology.org
Dow
nloaded from

directed against RyR1. Finally, the increased pressure observedin SHR induced the opposite effect: the increase of RyR1expression and the increase of Ca2"-induced Ca2" release(CICR). Taken together, these findings suggest that VSMCadapt themselves to the forces that they experience, gravita-tional as hemodynamic, by modulating their expression ofRyR1.
METHODS
Animals. These investigations conform to the European Commu-nity and French guiding principles for the care and use of animals.Authorization to perform animal experiments (A-33-063-003) wasobtained from the Prefecture de la Gironde (France). Wistar male ratsas well as spontaneous hypertensive rats (SHR) and their controlWistar Kyoto rats (WKR) (120–150 g) were euthanized by cervicaldislocation.
Hindlimb unloading rat. Briefly, rats were hindlimb unloaded (HU;8 days, angle 30–40°) by tail attachment as described previously (40)and in accordance with recommendations published by the NationalAeronautics and Space Administration (NASA) (42). The hindlimbunloading time was chosen because 1) it was compatible to !Gexposure during space flight missions (Soyuz TMA-8 transport mis-sion and STS-118) and 2) we have previously shown that portal veinadaptation was significant after this HU duration. If there is nomodification of the body weight, the HU effects calibrated on soleuswet weight are significant (52). Here, soleus wet weight from HU ratswas 20% decreased compared with control rats. Moreover, vasculareffects of !G such as orthostatic disorders can appear after only a fewdays of !G exposure (55).
Mice in STS118. European Space Agency (ESA) gave us theopportunity to retrieve hepatic portal vein from mice that wereexposed to !G during the STS-118 mission. The C57Bl6/J breedingwas assumed by NASA. Animals were separated in three groups (n #6 mice/group): conditioned in classical breeding environment baseline(BL); in cage used for the flight and stayed on Earth [ground control(GC)] and boarded on STS-118 (fly). Portal veins were removed,stored in 4% PFA for immunolabeling and RNA later solution(Qiagen) for RT-qPCR, and shipped to our laboratory for analysis.
Culture of VSMC from hepatic portal vein. The dissociation ofportal vein VSMC was detailed previously (41). Briefly, tissues wereincubated twice in a cocktail of 0.8 mg/ml collagenase (EC 3.4.24.3)and 0.2 mg/ml pronase E (EC 3.4.24.31) in a low Ca2" (40 !M)physiological solution (Hanks’ balanced salt solution) at 37°C. VSMCwere obtained by mechanical dispersion. VSMC used for Ca2"
measurements were seeded on glass slides and maintained in short-term primary culture in medium M199 containing 5% fetal calf serum(FCS), 20 U/ml penicillin, and 20 !g/ml streptomycin. Cells werekept in an incubator gassed with 95% air and 5% CO2 at 37°C andused during 24 h.
Packaging of VSMC to spaceflight. Cells were packed in bagsspecific for cell culture in space flight. The packaging and the functionof the hardware may be obtained from the CNES and ESA. Briefly,VSMC were in culture in appropriate medium, and the hardwarecontained programmed sequence to release angiotensin II (AII) solu-tion (100 nM final) to verify their contractile phenotype and afterformaldehyde fixative solution (3.7% final) to stop cell life after spaceexposure. The time profile was made to perform these experimentsafter either 24 or 200 h of !G exposure. Bags were placed into sixMAMBA containers (Dutch-Space): three in !G conditions and threein the centrifuge to reproduce 1 G with space environment in theKUBIK incubator (COMAT). Finally, to measure eventual effects oftravel or space environment, simultaneous and identical experimentswere performed on cells kept on Earth. Cells were sent to the ISS withthe Soyuz TMA-8 transport mission and returned with the SoyuzTMA-7 transport mission.
Immunostaining. We used protocols that were previously described(12). Briefly, fixed VSMC were washed with PBS and permeabilizedin PBS containing 2% bovine serum albumin and 1 mg/ml saponin.Anti-RyR antibody (34C, Santa Cruz Biotechnology, 1:1,000), anti-AT1 receptor antibody (SC-579, 1:500), or anti-smooth muscle actinantibody (SC-8432; 1:500) was added, and cells were incubatedovernight at 4°C. Fixed portal veins from mice were treated as VSMCbut were incubated with anti-RyR1 (AB9078, Millipore, 1:200),anti-RyR2 (C3.33, Millipore, 1:200), and anti-RyR3 (AB9082, Mil-lipore, 1:200) antibodies. Samples were washed three times andincubated with the secondary Fluoprobe-488 antibody (1:250, Inter-chim) during 45 min at room temperature. After three washes in PBS,slides were mounted in Vectashied (Abcys). We have evaluated thenon-specific fluorescence of secondary antibody by incubating cellsonly with the Fluoprobe-488 antibody (1:250), namely the primaryantibody was omitted in the first steps. Microscope settings weredefined with the non-specific preparation, and the acquisition param-eters were kept constant to compare fluorescence emitted in eachcondition. The fluorescence counting was made only with non-satu-rated images and analyzed as described in Ref. 12.
RT-PCR. Total RNA was extracted from cultured VSMC takenfrom hepatic portal vein using the RNA preparation kit from Epicentrefollowing the instructions of the supplier. The reverse transcriptionreaction was performed on 50 ng of RNA using the Sensiscript-RT kit(Qiagen). Two different sets of sense and antisense primer pairsspecific for RyR1, RyR2, and RyR3 were used as previously de-scribed (9). The first PCR protocol used with cutured portal vein cellswas detailed in Ref. 12. In the second one, amplicons produced withCFX96 q-PCR machine (Biorad) were directly quantified by usingCFX96 software. The normalization of RyR expression was realizedwith rat samples after verification of the stability of reference geneexpression with GeNorm algorithms; the normalization was madewith actin and GAPDH. The normalization used for the sampleexposed to !G was made with Alien strategy (Agilent Technologies).It was consisting in addition to 1 pg of Alien RNA in RNA sample,and it was reverse-transcripted during PCR experiments with specificprimer pair.
Antisense oligonucleotide treatment. To mimic the decrease inRyR1 expression, we injected intraperitoneally a solution containingphosphorothioate antisense oligonucleotides (ASON, denoted withthe prefix “as”) and JetPEI in vivo (Polyplus). Glucose solutions (5%),containing either asRyR1 associated to the JetPEI in vivo or only thevector, were injected following the recommendations of the supplier.After 4 days, a second injection was performed. Eight days after thefirst injection (D8), the rats were euthanized, and VSMC were isolatedfrom hepatic portal vein as described above. To detect cells containingasRyR1, we used 5=-Cy5 indocarbocyanin-labeled ASON (Eurogen-tec). The fluorescence of Cy5 indocyanin was measured at 680 $ 32nm (excitation 647 nm). The sequences of asRyR1 and the scrambledsequence of asRyR1 (asSCR) were previously described (9). Thetreated rats acted as control rats without visible troubles.
Contraction of portal vein strips. Portal vein was dissected andopened to remove the endothelium. One muscle strip was taken in themiddle of portal vein from rats, calibrated (mass 1 $ 0.2 mg), placedin an unloaded condition to minimize spontaneous contractions on aglass slide, and fully visualized with LSI macroscope setup (LeicaMicrosystems). The contraction was evaluated by the shortening ofstrip in both x and y directions.
Western blot analysis. The protocols used were previously de-scribed (10). The muscle hepatic portal vein were homogenized in 50!l of PBS 10% SDS solution. Supernatant was collected, and theprotein content was measured according to the method of Bradford(6). Two different systems of revelations were used and described insupplementary methods. Briefly, in both cases, membranes wereincubated (overnight, 4°C) with anti-RyR1 (AB9078), anti-RyR2(C3.33), and anti-RyR3 (AB9082) primary antibodies (1:1,000). Byusing a stripping step, generally two different RyR subtypes have been
472 GRAVITY AND HYPERTENSION REGULATE RyR1 IN PORTAL VEIN
J Appl Physiol • doi:10.1152/japplphysiol.00733.2011 • www.jappl.org
on February 2, 2012jap.physiology.org
Dow
nloaded from
probed per membrane. The secondary antibodies were coupled to ahorseradish peroxydase and detected using an enhanced chemolumis-cence kit (GE Healthcare) or coupled to Q-dot fluorophores (%em, 655nm), and signal was detected using a ChemiGenius 2 system (Syn-gene).
Cytosolic Ca2" measurements on VSMC from hepatic portal vein.Physiological solution was composed as followed (in mM): 140 NaCl,5.6 KCl, 1 MgCl2, 2 CaCl2, 11 glucose, 10 hepes, pH # 7.4 at 24°Cwith NaOH. Cells were loaded by incubation in a physiologicalsolution containing 2 !M Fluo-4-AM or Fluo8L-AM, and cells wereloaded for 20 min at 37°C. Probes were excited at 488 nm, andemitted fluorescence was filtered and measured at 540 $ 30 nm. Twodifferent modes of image acquisition were used with the confocalsystem MRC1024ES (Bio-Rad) connected to a Nikon Diaphot micro-scope: image series were acquired at 1.2-s intervals and line-scanmode images were composed at a rate of 2 ms/scan. In this image,time increases from left to right. Image processing was performed byusing Lasersharp 2000 software (Bio-Rad) and analysis by using IDLsoftware as previously described (12). Background fluorescence wasminimized by specific adjustment of MRC1024ES acquisition param-eters. Caffeine, KCl, AII, and endothelin-1 (ET-1) were applied bypressure ejection from a glass pipette for the period indicated on therecords, whereas other pharmacological agents were added to theperfused physiological solution. All experiments were carried out at26 $ 1°C.
Flash photolysis. Cells from portal vein were loaded with thepermeant form of InsP3 (cag-iso-2-145-InsP3, 2 !M in M199) during30 min at 37°C before the Fluo8-AM loading. Photolysis produce bythe xenon flash lamp (1-ms pulse) has been described previously (40).The flash intensity was adjusted from 150 to 250 V in these experi-ments.
Chemicals and drugs. Collagenase was obtained from Worthington(Freehold, NJ). Fluo-4-AM, Fluo8L-AM, PBS, Sybr Green, TRITC-phalloidin, GeBa-Gels, and fluoprobe-488 secondary antibodies werefrom Interchim (Montluçon, France). Caffeine was from Merck(Darmstadt, Germany). Medium M199, streptomycin, and penicillinwere from Invitrogen (Cergy Pontoise, France). As described in thissection, antibodies were from Millipore (Saint-Quentin en Yvelines,France) and Santa-Cruz Biotechnologies (Tebu-Bio, Le Perray enYvelines, France). Both used FCS were from PAA Laboratories (Linz,Austria). Cag-iso-2-145-InsP3 [D-2,3-O-isopropylidene-6-O-(2-nitro-4,5-dimethoxy)benzyl-myo-inositol 1,4,5-trisphosphate-hexakis(propionoxym-ethyl)ester] was synthesized by SiChem (Bremen, Germany). AII andET-1 were synthesized by Genepep (Montpellier, France). All otherchemicals were from Sigma (Lyon, France).
Data analysis. Data are expressed as means $ SE; n represents thenumber of tested cells. Significances between different conditionswere tested with one-way ANOVA test with a Tukey’s post test orwith Student’s t-test. P values of &0.05 were considered significant.
RESULTS
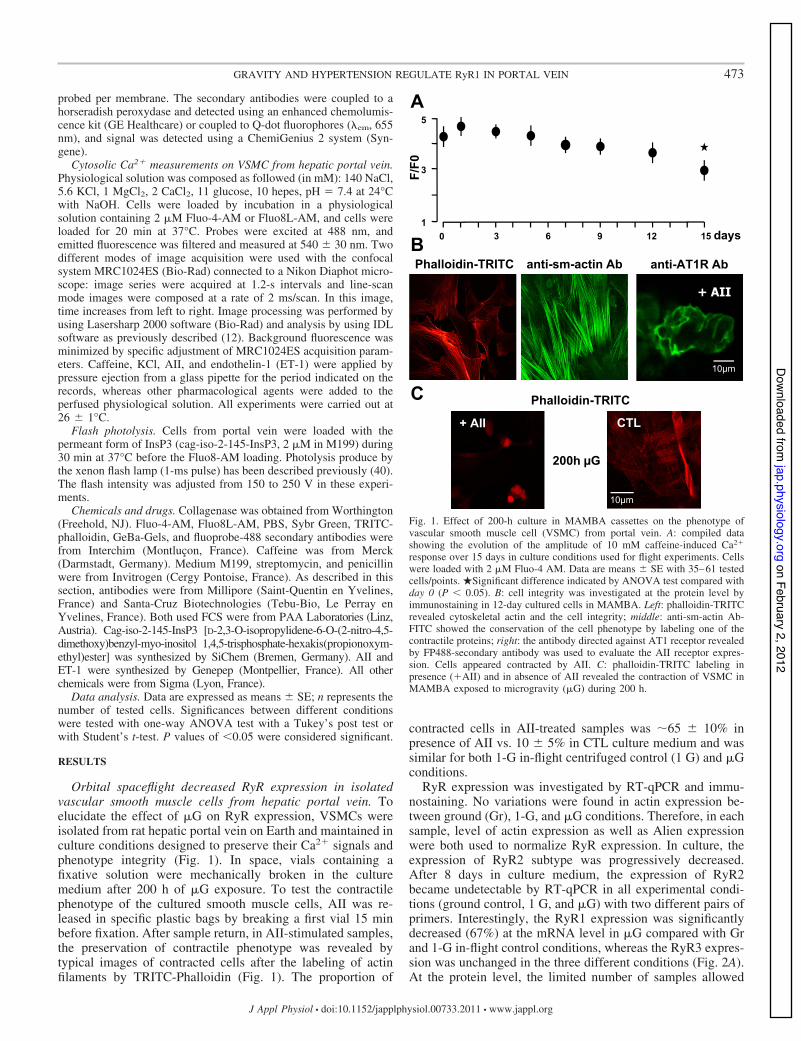
Orbital spaceflight decreased RyR expression in isolatedvascular smooth muscle cells from hepatic portal vein. Toelucidate the effect of !G on RyR expression, VSMCs wereisolated from rat hepatic portal vein on Earth and maintained inculture conditions designed to preserve their Ca2" signals andphenotype integrity (Fig. 1). In space, vials containing afixative solution were mechanically broken in the culturemedium after 200 h of !G exposure. To test the contractilephenotype of the cultured smooth muscle cells, AII was re-leased in specific plastic bags by breaking a first vial 15 minbefore fixation. After sample return, in AII-stimulated samples,the preservation of contractile phenotype was revealed bytypical images of contracted cells after the labeling of actinfilaments by TRITC-Phalloidin (Fig. 1). The proportion of
contracted cells in AII-treated samples was '65 $ 10% inpresence of AII vs. 10 $ 5% in CTL culture medium and wassimilar for both 1-G in-flight centrifuged control (1 G) and !Gconditions.
RyR expression was investigated by RT-qPCR and immu-nostaining. No variations were found in actin expression be-tween ground (Gr), 1-G, and !G conditions. Therefore, in eachsample, level of actin expression as well as Alien expressionwere both used to normalize RyR expression. In culture, theexpression of RyR2 subtype was progressively decreased.After 8 days in culture medium, the expression of RyR2became undetectable by RT-qPCR in all experimental condi-tions (ground control, 1 G, and !G) with two different pairs ofprimers. Interestingly, the RyR1 expression was significantlydecreased (67%) at the mRNA level in !G compared with Grand 1-G in-flight control conditions, whereas the RyR3 expres-sion was unchanged in the three different conditions (Fig. 2A).At the protein level, the limited number of samples allowed
Fig. 1. Effect of 200-h culture in MAMBA cassettes on the phenotype ofvascular smooth muscle cell (VSMC) from portal vein. A: compiled datashowing the evolution of the amplitude of 10 mM caffeine-induced Ca2"
response over 15 days in culture conditions used for flight experiments. Cellswere loaded with 2 !M Fluo-4 AM. Data are means $ SE with 35–61 testedcells/points. !Significant difference indicated by ANOVA test compared withday 0 (P & 0.05). B: cell integrity was investigated at the protein level byimmunostaining in 12-day cultured cells in MAMBA. Left: phalloidin-TRITCrevealed cytoskeletal actin and the cell integrity; middle: anti-sm-actin Ab-FITC showed the conservation of the cell phenotype by labeling one of thecontractile proteins; right: the antibody directed against AT1 receptor revealedby FP488-secondary antibody was used to evaluate the AII receptor expres-sion. Cells appeared contracted by AII. C: phalloidin-TRITC labeling inpresence ("AII) and in absence of AII revealed the contraction of VSMC inMAMBA exposed to microgravity (!G) during 200 h.
473GRAVITY AND HYPERTENSION REGULATE RyR1 IN PORTAL VEIN
J Appl Physiol • doi:10.1152/japplphysiol.00733.2011 • www.jappl.org
on February 2, 2012jap.physiology.org
Dow
nloaded from

only an analysis based on the quantification of immunostainingwith an antibody directed against a common RyR epitope (34Cmonoclonal antibody). First, we have verified that the nonspe-cific fluorescence was not modified in the three studied condi-tions. And then a significant decrease in RyR specific fluores-cence (47%) was revealed in cells exposed to !G (Fig. 2, B andC). The !G is a unique environment because there is no forceapplied on the system. Therefore, by culturing VSMC in a !Genvironment, it allows one to study the influence of oneparticular variable, in this case, blood pressure.
We have also verified whether the transport of samples wasdeleterious for cell viability and RyR subtype expression byperforming RT-qPCR on two different specific additive cas-settes fixed exactly at the same time but stored in laboratory
and on the launch area just before the launch. There was nodifference in the level of expression of RyR subtypes betweencell samples stored in laboratory and those that have traveled(RyR1/actin # 0.61 $ 0.09 vs. 0.48 $ 0.13; RyR2/actin #0.20 $ 0.08 vs. 0.26 $ 0.07; RyR3/actin # 0.70 $ 0.05 vs.0.74 $ 0.07 in laboratory vs. traveled samples, respectively).
RyR1 expression was also decreased in portal vein frommouse exposed to orbital spaceflight. With a similar protocol,we have investigated the expression of RyR subtypes in portalvein from mice exposed to !G for the same duration during theSTS-118 mission. The RT-qPCR experiments revealed that theexpression of RyR1 was decreased ((61%), whereas RyR2and RyR3 expressions were not changed (Fig. 3A). This resultwas also confirmed at the protein level by analysis of confocalmicroscopy stacks of RyR immunostaining because a Westernblot analysis was not possible because the size of sample was
Fig. 3. Effect of !G on VSMC from mouse portal vein. Mice were exposed to!G during orbital flight of STS118 mission. A: expression of RyR subtypes asrevealed by RT-qPCR. cDNA fragments corresponding to RyR1–3 wereamplified from hepatic portal vein from mice maintained in classical cages(baseline, Bl), in experimental device on ground (Gr.), and in experimentaldevice in the Shuttle (!G). The quantification was realized by using theconcomitant amplification of RNA Alien added in RNA samples before RT.Each experiment was repeated five times in three different preparations.B: nuclei (red) as revealed by propidium iodide and RyR1 (green) as revealedby immunostaining in each condition. Scale bar: 10 !m. C: expression levelsof each RyR subtype protein were evaluated by the quantification of fluores-cence observed with subtype-specific antibodies in all conditions. Compileddata are expressed as the mean of fluorescence divided by the number ofpropidium iodure tagged nucleus $ SE; the number of tested cells is indicatedin parentheses. Data are means $ SE. !Significant difference by ANOVA test(P & 0.05).
Fig. 2. Effect of !G on cultured VSMC. A: expression of ryanodine receptor(RyR) subtypes 1 and 3 as revealed by RT-qPCR. cDNA fragments corre-sponding to RyR1 and RyR3 were amplified from primary cultured VSMCexposed to ground (Gr), 1-G force (1G), or !G conditions in the InternationalSpace Station (ISS). The quantification was realized by using the concomitantamplification of RNA Alien added in RNA samples before RT. Each experi-ment was repeated three times in five different cell preparations. Data aremeans $ SE. *Significant difference by ANOVA test (P & 0.05). B: expres-sion of RyR as revealed by immunostaining. Typical confocal sections ob-tained with anti-RyR antibody (1:1,000; C34 clone) in ground, 1-G force, or!G conditions. The same correction factors were applied on all images. Nstands for nucleus. One example of region of interest (ROI) used for fluores-cence measurement is symbolized in white line in each condition. Experimentwas performed on the three different pairs of cassettes. Scale bar: 10 !m.C: expression level of RyR protein evaluated by the quantification of fluores-cence observed with anti-RyR antibody in ground (Gr), 1-G force, or !Gconditions. Data are means $ SE; the number of tested cells is indicated inparentheses. !Significant difference by ANOVA test (P & 0.05).
474 GRAVITY AND HYPERTENSION REGULATE RyR1 IN PORTAL VEIN
J Appl Physiol • doi:10.1152/japplphysiol.00733.2011 • www.jappl.org
on February 2, 2012jap.physiology.org
Dow
nloaded from

too small. The immunolabeling using an anti-RyR1 antibodywas decreased ((43%), whereas the immunostainnings ofRyR2 and RyR3 revealed with specific antibodies againstRyR2 and RyR3, respectively, were not changed (Fig. 3, Band C).
RyR1 expression is decreased by both HU and intraperito-neal injection of anti-RyR1 antisense oligonucleotides in portalvein. The HU rat model has been widely used during the lasttwo decades to mimic the fluid volume redistribution inducedby !G. In portal vein, we previously demonstrated that the[3H]ryanodine binding was decreased by HU (40). To com-pare the effect of the decrease in gravitational force with thedecrease in hemodynamic force on VSMC, we investigated theRyR expression pattern in hepatic portal vein from CTL andHU rats. The RT-qPCR revealed a significant decrease ofRyR1 expression in HU rats compared with CTL rats, whereasthe expression levels of RyR2 and RyR3 were identical in bothconditions (Fig. 4A). To assess the RyR1 expression variationat the protein level, Western blots were made with proteinextracted from portal vein from CTL and HU rats (Fig. 4B).The intensity of anti-RyR bands was normalized against ananti-actin antibody. The ratio of RyR1 to actin was signifi-cantly lower in portal vein taken from HU rats than from CTL,whereas the ratio of RyR2 or RyR3 to actin was not modified(Fig. 4C). RyR1 expression is thus decreased by the reducedinfero-posterior blood pressure induced by HU.
To mimic the decrease of RyR1 expression, we developedan in vivo approach by antisense oligonucleotide (ASON)directed against RyR1 (asRyR1). Localization of the ASON inthe VSMC was assessed by fluorescence microscopy using aCy5-tagged asRyR1. Inhibition of the RyR1 expression wasinvestigated by RT-qPCR following the same methodology asdescribed earlier (Fig. 4A). In rats injected with the cationicvector only, the RyR1-to-actin ratio was similar to the ratiomeasured in CTL rat. Injection of asRyR1 causes similarreduction of the RyR1-to-actin ratio as observed in HU animalswithout effect on RyR2-to-actin and RyR3-to-actin ratios.Western analysis showed that ASON treatment reduced thedensity of the RyR1 protein band (Fig. 4D). The RyR1-to-actinratio was (58.5 $ 18.7% less in ASON-treated than invehicle-treated animals. The asRyR1 in vivo injection had noeffect on RyR2 and RyR3 expression (Fig. 4E).
RyR1 has a functional role in portal vein VSMC adaptationto HU conditions. To assess the role of RyR1 in hepatic portalvein adaptation to infero-posterior decreased blood pressure,different Ca2" signals were compared between myocytes iso-lated from CTL, HU, and asRyR1-treated rats.
To measure the RyR-dependent Ca2" release, caffeine (10mM), known to activate RyRs by increasing their sensitivity toCa2" activation, was applied by puff on isolated VSMC fromportal vein. Amplitudes and upstroke velocity of induced Ca2"
responses were identical between myocytes isolated from CTLand vehicle-treated rats and were thus pooled in the followingexperiments. Line-scan mode with a line acquired each 2 msconfirmed these differences and revealed a decreased upstrokevelocity in HU and asRyR1-treated myocytes compared withcontrol cells (Fig. 5A). Mean amplitude of caffeine-inducedCa2" responses was decreased in HU and asRyR1-treated ratswithout any significant difference between these two last con-ditions (Fig. 5B). A SERCA inhibitor was used to check theeffect of HU and asRyR1 injections on Ca2" store refilling.
The use of 1 !M thapsigargin and the reproducibility of theresponses between CTL, HU, and asRyR1-treated rats sug-gested that the SR Ca2" loading was unchanged (not shown).
In 1997, we showed a reduced cytosolic Ca2" responseevoked by AII in hepatic VSMC from HU rats (40). Then AII(100 nM) was first tested on VSMC from CTL, HU, andasRyR1-injected rats. As expected, HU decreased the ampli-tude of Ca2" signal induced by application of AII comparedwith amplitude obtained in control condition (Fig. 6A, top).Interestingly, intraperitoneal injections of asRyR1 also reducedthe AII-evoked Ca2" response (Fig. 6A, bottom). In the pres-ence of 10 !M ryanodine for 30 min, the amplitude ofAII-evoked Ca2" responses was decreased to a similar level in
Fig. 4. Effect of hindlimb unloading (HU) and intraperitoneal injection ofantisense oligonucleotides directed against RyR1 (asRyR1) on RyR1 expres-sion in hepatic portal vein. Animals were hindlimb suspended during 8 daysand treated twice, and VSMC were prepared 8 days after the first injection.A: expression of RyR1–3 was investigated by RT-qPCR in isolated VSMCfrom hepatic portal vein. Quantification of the relative mRNA expression ofRyR1, RyR2, and RyR3 from vehicle injected (CTL; open bars), HU (filledbars), and asRyR1-injected rats (shaded bars). B: typical Western blot reveal-ing the expression of protein RyR1 in hepatic portal vein from CTL and HUrats. C: compiled data of the ratio of RyR1–3 to actin in CTL (open bars) andHU conditions (filled bars). D: expression of RyR1 and actin in hepatic portalfrom CTL and asRyR1-injected (as) rats as revealed by Western blot withanti-RyR1 and anti-actin antibodies and detected by immunofluorescence. Theladder lane was indicated by L. E: compiled data of the ratio of RyR1–3 toactin in CTL (open bars) and asRyR1 (shaded bars) conditions. Experimentswere repeated five times, and two animals were used each time for eachcondition. Data are means $ SE. !Significant difference by ANOVA test(P & 0.05).
475GRAVITY AND HYPERTENSION REGULATE RyR1 IN PORTAL VEIN
J Appl Physiol • doi:10.1152/japplphysiol.00733.2011 • www.jappl.org
on February 2, 2012jap.physiology.org
Dow
nloaded from

CTL, HU, and asRyR1-injected rats, suggesting that onlychanges in RyR function were responsible of the decreasedAII-induced Ca2" response in HU and asRyR1 conditions (Fig.6B). We also evaluated the effect of asRyR1 and HU condi-tions on the AII-induced contraction (100 nM) measuring theshortening of portal vein strip placed on microscope setup usedfor Ca2" measurement. The contraction of portal vein wasdecreased by HU from 22 $ 3% (n # 11) in CTL conditionsto 12 $ 1% (n # 12) in HU conditions, and it was decreasedfrom 17 $ 1% (n # 15) to 11 $ 2% (n # 20) in asRyR1-treated animals. This last result indicated that the decrease ofCa2" signal possibly reduced the contraction induced by AII.
Since RyRs can be physiologically activated by a localincrease in intracellular Ca2", we tested the ability of the HUand asRyR1 treatment to modify this mechanism called Ca2"-induced Ca2" release (CICR). The CICR mechanism can alsobe activated by membrane depolarization, leading to the open-ing of L-type Ca2" channels and thus intracellular Ca2"
elevation. KCl (140 mM) was applied by puff on isolatedVSMC (Fig. 7A); as a result, the induced Ca2" responses werelower in HU and asRyR1 groups than in CTL group, whereas
this difference was ablated by 10 !M ryanodine (Fig. 7, Band C).
We completed the investigation by the analysis of Ca2"
signal induced by ET1 (100 nM), known to activate InsP3production (29). The amplitude of ET1-induced Ca2" re-sponses was decreased from 5.4 $ 0.3 (n # 39, CTL) to 4.2 $0.3 ratio units (n # 41, HU; P & 0.05, one-way ANOVA test)and to 3.9 $ 0.3 ratio units (n # 33, asRyR1-treated rats; P &0.05, one-way ANOVA test). The mean amplitudes of theET1-evoked Ca2" response in HU and asRyR1-injected ratswere similar. In the presence of 10 !M ryanodine, the meanamplitude of ET1-evoked Ca2" responses was similar in allconditions [CTL: 1.6 $ 0.3 (n # 16); HU: 2.1 $ 0.4 (n # 25);asRyR1-injected rats: 1.8 $ 0.3 (n # 10)], suggesting that, ininfero-posterior VSMCs, the InsP3 expression is not modifiedby HU condition. Taken together, these results clearly show thedecrease in the RyR expression is sufficient to mimic thealteration of Ca2" signaling induced by HU in VSMC fromhepatic portal.
RyR1 was also regulated in portal vein VSMC from hyper-tensive model. Finally, the SHR model could be used as theopposite model to !G and hindlimb suspension because ifthese last conditions induced a decrease of portal vein bloodpressure (1), on the contrary, the blood pressure was increasedin SHR model. Then, to complete our study, we wanted to
Fig. 6. Effect of hindlimb unloading and intraperitoneal injection of oligonu-cleotides antisense asRyR1 on AII-evoked Ca2" response in VSMC isolatedfrom hepatic portal vein. A: typical Ca2" responses induced by AII (100 nM)in portal vein VSMC from CTL and HU rats (top) and vehicle-injected andasRyR1-injected (bottom) rats. B: compiled data showing the mean amplitudesof Ca2" responses evoked by AII (100 nM) in left CTL (open bar) and HU(filled bar) rats and right vehicle-injected (open bar) and asRyR1-injected(shaded bars) rats. Shaded bars represent the Ca2" responses evoked by AII inpresence of ryanodine (10 !M). Data are means $ SE with the number oftested cells indicated in parentheses. VSMC were loaded with 2 !M Fluo8L-AM. !Significant difference by t-test (effect of HU or asRyR1 injection; P &0.05). Shaded star indicates significant difference by t-test (effect of ryanodine;P & 0.05).
Fig. 5. Effect of hindlimb unloading and intraperitoneal injection of oligonu-cleotides antisense asRyR1 on caffeine-evoked Ca2" response in VSMCisolated from hepatic portal vein. A: typical 2 ms/line-scan mode imagesobtained by confocal microscopy of the Ca2" signals evoked by application of10 mM caffeine, observed in VSMC from CTL, HU, and asRyR1-injected rats.B: compiled data showing the amplitude of Ca2" signals evoked by 10 mMcaffeine. Data are means $ SE with the number of tested cells indicated inparentheses. VSMC were loaded with 2 !M Fluo8L-AM. !Significant dif-ference by ANOVA test (P & 0.05).
476 GRAVITY AND HYPERTENSION REGULATE RyR1 IN PORTAL VEIN
J Appl Physiol • doi:10.1152/japplphysiol.00733.2011 • www.jappl.org
on February 2, 2012jap.physiology.org
Dow
nloaded from

analyze the RyR-dependent Ca2" signaling in VSMC fromSHR and WKR portal veins. First, the caffeine-induced Ca2"
release was significantly increased in VSMC from SHR portalvein compared with those obtained from WKR (Fig. 8A;)F/F0 # 4.48 $ 0.26, n # 51 in SHR vs. 2.94 $ 0.15, n # 23,in WKR; P & 0.05, t-test). Because the expression patterns ofthe ET-1 receptors were regulated in SHR model, the implica-tion of RyR into the CICR induced by InsP3 pathway usingET-1 was not possible. Then, we used the photolysis ofcag-iso-2-145-IP3 to activate IP3-dependent Ca2" signals. Inportal vein VSMC loaded with cag-iso-2–145-InsP3, the ultra-violet flash induced a Ca2" signal whose amplitude was sig-nificantly increased in SHR compared with WKR (Fig. 8B;)F/F0 # 3.50 $ 0.13, n # 53 in SHR vs. 1.77 $ 0.03, n # 48,in WKR at 100-V intensity; and )F/F0 # 5.17 $ 0.47, n # 53,in SHR vs. 2.37 $ 0.20, n # 48, in WKR at 200-V intensity).As also illustrated in Fig. 8C, the KCl-induced Ca2" responsewas significantly increased in SHR compared with WKR. Theapplication of 10 !M ryanodine induced a more importantdecrease of KCl-induced Ca2" response in SHR than in WKR((68.3 $ 7.1% vs. (29.9 $ 4.2%, n # 6 different experi-ments), suggesting that the participation of CICR was moreimportant in SHR than in WKR.
Finally, the expression of RyR subtype was investigated inSHR and WKR by RT-qPCR. Only the expression of RyR1subtype was increased in SHR (Fig. 9), indicating an oppositeeffect to those observed in VSMC from HU rats or !G exposedmice and !G exposed VSMC.
DISCUSSION
The molecular basis of physiological effects of vascularadaptation to !G must be investigated at several levels. Indeed,vascular reactivity could be modified by changes in cell inter-actions, contractile proteins, and transduction pathways. Themodifications could affect the expression level and also thefunctional state of proteins implicated in regulation of contrac-tion. Vascular reactivity is due to the VSMC contraction/relaxation balance. The regulation of this balance is assumed
Fig. 7. Effect of hindlimb unloading and intraperitoneal injection of oligonu-cleotides antisense asRyR1 on KCl-evoked Ca2" response in VSMC isolatedfrom hepatic portal vein. A: typical Ca2" responses induced by KCl (140 mM)in portal vein VSMC from CTL, HU, and asRyR1-injected rats. B: compileddata showing the mean amplitudes of Ca2" responses evoked by KCl (140mM) in CTL (open bar), HU (filled bar), and asRyR1-injected (shaded bars)rats. Shaded bars represented the Ca2" responses evoked by KCl in presenceof ryanodine (10 !M). Data are means $ SE with the number of tested cellsindicated in parentheses. VSMC were loaded with 2 !M Fluo8L-AM. !Sig-nificant difference by one-way ANOVA (effect of HU and asRyR1 injection;P & 0.05). Shaded stars indicate significant difference by t-test (effect ofryanodine; P & 0.05).
Fig. 8. Typical Ca2" responses in VSMC isolated from SHR and WKR hepaticportal vein induced by caffeine (10 mM; A), by ultraviolet flash (100 V; B), andby KCl (140 mM; C) VSMC were loaded with 2 !M Fluo8L-AM.
Fig. 9. RyR expression in VSMC isolated from SHR and WKR hepatic portalvein. Quantification by RT-qPCR of the relative mRNA expression of RyR1,RyR2, and RyR3 from WKR (open bars) and SHR (filled bars) rats. Thequantification was realized by using the concomitant amplification of RNAAlien added in RNA samples before RT. Data are means $ SE; 6 differentanimals were used. !Significant difference by t-test (P & 0.05).
477GRAVITY AND HYPERTENSION REGULATE RyR1 IN PORTAL VEIN
J Appl Physiol • doi:10.1152/japplphysiol.00733.2011 • www.jappl.org
on February 2, 2012jap.physiology.org
Dow
nloaded from

by endothelial function principally via NO synthesis and by theregulation of Ca2" signaling pathway of the VSMC. Here, wehave investigated the regulation of Ca2" signal encoding by!G and vessel pressure.
VSMC are directly sensitive to the real !G. First, to delin-eate the relative contribution of vascular smooth muscle fromthe endothelial component in terms of the vascular adaptationto !G, experiments were performed in cultured VSMC. Theexpressions of RyR protein and RyR1 mRNA were decreasedin cultured VSMC exposed to !G during TMA-8 spaceflight.These results suggested that Ca2" signaling pathways wereaffected by !G. But one of the important challenges of this partof our work was to maintain the closest possible phenotype ofthe initial situation in the cultured VSMC. The presence of calfserum in culture medium is necessary to maintain the VSMCviability, but it also induced a dedifferentiation from a con-tractile to a proliferative phenotype, characterized by decreasedSERCA2A and RyR expression and increased InsP3R, SOC,and CaV3 (26, 33, 34, 53). As we described previously, the useof calf serum decomplementation limits the VSMC prolifera-tion (36), and the expression of RyR subtypes was not signif-icantly modified during 4 days (9), but after 8 days only theRyR2 expression decreased to a level undetectable by RT-PCR. However, the RyR-dependent Ca2" response and thecontractile phenotype remained after 8 days, showing a pre-served VSMC integrity. These culture conditions are usable tofollow the effect of !G on Ca2" signaling, despite a possibleeffect on RyR2 expression. There is no evidence in our workthat !G induced drastic modifications of VSMC survival andproliferation, whereas RyR2 could be necessary to Ca2" sig-nals implicated in portal vein VSMC proliferation. In adherentcells, both real and simulated !G (clinorotation) decreased cellmitosis (37, 54). The differences between our work and thesestudies are due to the experimental procedures. Indeed, wedefined particular conditions to maximally inhibit the cellproliferation. By indirect measurement, we have also shownthat the contractile phenotype was preserved in !G becauseAII was still able to induce contraction of VSMC, and therewas no modification in the proportion of contracted cellsbetween 1-G and !G samples. Finally, at this stage of inves-tigation, we cannot exclude that the decrease in RyR1 expres-sion in !G can also be due, at least partly, to a decreasedVSMC metabolism as observed in cultured cells in clinorota-tion (7). In the vascular physiology, it will be important to usethe !G environment to characterized the pressure sensor andthe level of gravity change that is perceptible by the VSMC asperformed in vegetal root cells (16).
In !G conditions, the VSMCs have been packaged inEarth atmosphere, and only the gravity was not applied oncells in the ISS environment; blood and adjacent tissuepressures were then absent. The use of animals living inspace environment has been necessary to verify whether theeffect of !G observed in cultured VSMC made sense invascular physiology.
In portal vein from mice exposed to !G during theSTS-118 Endeavour mission, the expression of RyR1 wasdecreased as observed in cultured VSMC, indicating amolecular adaptation to real !G and potentially to changesin applied forces.
In a specific situation, the forces applied on cells change andmodify the gene expression pattern as observed in myometrium
near parturition (10, 25), in portal hypertension (48), or in renalhypertonic stress (31). Taken together, these results suggestthat VSMC can rapidly (8 days) adapt their Ca2" signalingpathways to !G during spaceflight and probably to bloodpressure as observed in hypertensive situation.
Our study is the first direct demonstration that shows theabsence of force obtained by !G exposure during spaceflightinduces a modulation of VSMC and that reinforces the “pres-sure effect theory,” indicating that the intravascular pressure isa key regulator of vascular reactivity.
The asRyR1 in vivo treatment induces the decrease invascular reactivity. To investigate the significance of !G-induced RyR1 decrease in Ca2" signaling, we have induced thedecrease of RyR1 expression in rat by intraperitoneal injectionof asRyR1. The specific decrease in RyR1 expression wasfollowed at mRNA and protein levels and was responsible fordecreases in Ca2" responses induced by AII, ET1, and depo-larization. This result indicates diverse Ca2" signals are mod-ified to likely affect many cell functions such as contractionand proliferation. First, the decrease in contraction was sug-gested by the decrease in Ca2" signals evoked by vasocon-strictors such as depolarization, ET1, AII, and NE in ourlaboratory’s previous work (40). In the same way, we haveverified in confocal microscopy that AII-induced contraction ofportal vein strips significantly decreased asRyR1 treatments.These experiments showed that the level of RyR1 expressionwas determinant to encode the magnitude and upstroke veloc-ity of CICR in VSMC, confirming other studies using RyR1KO (19, 32). In a previous work, our laboratory determinedthat the 80% inhibition of RyR1 expression by asRyR1 incultured VSMC from the same model decreased the amplitudeand the upstroke velocity of caffeine-activated Ca2" responsein the range of 60% and 80%, respectively (9). Here, the effectsof asRyR1 and HU were similar, probably because the effectsof the used concentration of asRyR1 inhibits RyR1 expressionin the same range to those observed with HU.
But at this stage of the discussion, we cannot link togetherthe decrease of RyR1 expression level as an adaptation to thedecrease of pressure. To have some arguments to discuss thispoint, we have compared results obtained on asRyR1-treatedanimals and HU rats. This last model was admitted to mimicvascular adaptation to !G because HU reproduces the hypo-reactivity of infero-posterior vessels.
The HU decreases the RyR1 expression. The observed de-crease of AII-induced contraction of portal vein strips in HUcould be considered as a confirmation to the decrease invasoconstriction that was also found in other peripheral vesselsfrom HU rats, including abdominal aorta and femoral, pulmo-nary, and mesenteric arteries (4, 8, 15, 46, 47). In infero-posterior veins, the effect of HU is less clear than in arteries.HU also decreased, in a few days, the NE-induced contractionin vena cava, but it was due to the modifications of transduc-tion pathways via *1-adrenoceptor desensitization (50). Thedecrease of contraction in mesenteric veins from Sprague-Dawley rats was also observed (4, 17), but on the contraryPurdy and coworkers concluded that jugular and femoral veinswere globally not sensitive to HU (47). In these last models, theeffects also could be analyzed by focusing on the nature ofvasoconstrictors: NE-induced contraction was unchanged,whereas KCl-induced contraction was inhibited in HU rats.This last point would be consistent with our study, indicating
478 GRAVITY AND HYPERTENSION REGULATE RyR1 IN PORTAL VEIN
J Appl Physiol • doi:10.1152/japplphysiol.00733.2011 • www.jappl.org
on February 2, 2012jap.physiology.org
Dow
nloaded from

the evidence that InsP3R Ca2" release is not affected by HU(NE pathway), whereas CICR is affected (CICR betweenVDCC-induced Ca2" influx and RyR).
However, the molecular basis of vascular adaptation to HUcould be due to GPCR-activated pathway as suggested (3, 50),but it was not sufficient to explain the observed decrease indepolarization-induced and caffeine-induced contraction evenif the activation of GPCR by depolarization was possible (35).In 1997, our laboratory showed for the first time that RyRexpression is modified by HU conditions and may explain thedecrease in vasoreactivity of portal veins (40). In 2008, Col-leran and coworkers associated a decrease in RyR2 expressionin HU rats with diminished vasoconstrictor responses to KCl,NE, and caffeine in mesenteric resistance arteries (8). Theapparent discordance of their results with ours might be ex-plained in that they only detected RyR2 and RyR3 mRNAexpression. We have shown that RyR1 and RyR2 functionsseemed to be interchangeable in VSMC (9). So it is notsurprising to observe a HU-induced decrease in RyR2 expres-sion in the absence of RyR1 protein. The origin of this differentRyR subtype expression pattern could lie in the tissue used (10,11, 38).
Finally, the present study might explain the medical obser-vation during bed-rest experiments indicating a significantsplanchnic blood stagnation. Indeed, a decreased contractilityof the portal vein appears to be linked to a decreased RyR1function (2). With mesenteric vessels, portal vein is the mostimportant blood reserve; a loss of VSMC contractile ability in!G can induce the decrease in portal vein ability to transportthe blood and then becomes essential in spaceflight-consecu-tive orthostatic intolerance. The second consequence of thedecrease in Ca2" signals can be the remodeling of the vascularwall. In fact, HU-induced decreased contraction was associatedwith vascular hypotrophy in aorta and skeletal muscle arteries(14, 21), but this point must be studied more precisely byfollowing the proliferative pathway at the molecular level.
Regulation of RyR1 expression was also observed in SHRhypertension model. The decrease in RyR1 expression ob-served in HU model or after !G exposure is possibly due to thedrop of pressure applied on cells or blood pressure in vessels.Our results obtained from SHR showed that the chronic in-crease in blood pressure produces the opposite effect. Ourresults could also explain several effects observed in SHRmodel. In fact, the increase of RyR1 expression and theassociated increase of Ca2" signaling could explain the in-crease of the contraction induced by depolarization, caffeine,and agonists (29, 30, 43, 49) but also the effect of the inhibitorof sarcoplasmic reticulum Ca2" stores (18).
In conclusion, our study shows that, during spaceflight, !Gdirectly affects cultured VSMC by modulating the same mo-lecular target as hemodynamic changes. Thus !G-inducedblood pressure changes should induce the regulation of RyR1expression in the same way. Nevertheless, it is necessary toinvestigate as soon as possible the sensor of gravity and/orblood pressure variation and pathways activated to regulategene expression to adapt the Ca2" pathways to understand thebasis of vascular adaptation to spaceflight and also to give anew target to control and regulate blood pressure in hyperten-sive and hypotensive situations.
ACKNOWLEDGMENTS
The authors thank the people with whom !G experiments have beenprepared: G. Gauquelin-Koch (CNES); G. Gasset (GSBMS); L. Stodieck andV. Ferguson (BioServe Space Technologies, University of Colorado); D.Chaput (CADMOS, CNES); J. Hatton (ESA); all astronauts of the TMA-8 andTMA-7 soyouz missions and STS-118 mission; J. Mironneau for the helpfuldiscussion; and A. Prevot for manuscript editing.
Present address of F. Dabertrand: Department of Pharmacology, Universityof Vermont, UVM College of Medicine, Burlington, VT 05405.
GRANTS
This work was supported by grants from CNES (1999–2011), AgenceNationale pour la Recherche (AdapHyG, no. ANR-09-BLAN-0148), and apostdoctoral fellowship from CNES to F. Dabertrand. Equipment was financedby Centre National de la Recherche Scientifique (CNRS) and Conseil régionald’Aquitaine.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
Author contributions: F.D., N.M., and J.-L.M. conception and design ofresearch; F.D., Y.P., and J.-L.M. performed experiments; F.D., Y.P., andJ.-L.M. analyzed data; F.D. and J.-L.M. interpreted results of experiments;F.D. and J.-L.M. prepared figures; F.D. and J.-L.M. drafted the manuscript;F.D. and J.-L.M. edited and revised the manuscript; F.D., Y.P., N.M., andJ.-L.M. approved the final version of the manuscript.
REFERENCES
1. Aiello EA, Grassi de Gende AO. Uptake and release of Ca2" inchemically skinned aortic strips from spontaneously hypertensive (SHR)and normotensive (WKY) rats. Jpn Heart J 36: 377–388, 1995.
2. Arbeille PP, Besnard SS, Kerbeci PP, Mohty DM. Portal vein cross-sectional area and flow and orthostatic tolerance: a 90-day bed rest study.J Appl Physiol 99: 1853–1857, 2005.
3. Bao J, Zhang L, Ma J. Angiotensinogen and AT1R expression incerebral and femoral arteries during hindlimb unloading in rats. AviatSpace Environ Med 78: 852–858, 2007.
4. Behnke BJ, Zawieja DC, Gashev AA, Ray CA, Delp MD. Diminishedmesenteric vaso- and venoconstriction and elevated plasma ANP and BNPwith simulated microgravity. J Appl Physiol 104: 1273–1280, 2008.
5. Berridge MJ. Smooth muscle cell calcium activation mechanisms. JPhysiol 586: 5047–5061, 2008.
6. Bradford MM. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 72: 248–254, 1976.
7. Coinu R, Chiaviello A, Galleri G, Franconi F, Crescenzi E, PalumboG. Exposure to modeled microgravity induces metabolic idleness inmalignant human MCF-7 and normal murine VSMC cells. FEBS Lett 580:2465–2470, 2006.
8. Colleran PN, Behnke BJ, Wilkerson MK, Donato AJ, Delp MD.Simulated microgravity alters rat mesenteric artery vasoconstrictor dy-namics through an intracellular Ca2" release mechanism. Am J PhysiolRegul Integr Comp Physiol 294: R1577–R1585, 2008.
9. Coussin F, Macrez N, Morel JL, Mironneau J. Requirement of ryano-dine receptor subtypes 1 and 2 for Ca2"-induced Ca2" release in vascularmyocytes. J Biol Chem 275: 9596–9603, 2000.
10. Dabertrand F, Fritz N, Mironneau J, Macrez N, Morel J. Role ofRYR3 splice variants in calcium signaling in mouse nonpregnant andpregnant myometrium. Am J Physiol Cell Physiol 293: C848–C854, 2007.
11. Dabertrand F, Mironneau J, Henaff M, Macrez N, Morel J. Compar-ison between gentamycin and exon skipping treatments to restore ryano-dine receptor subtype 2 functions in mdx mouse duodenum myocytes. EurJ Pharmacol 628: 36–41, 2010.
12. Dabertrand F, Morel J, Sorrentino V, Mironneau J, Mironneau C,Macrez N. Modulation of calcium signalling by dominant negative splicevariant of ryanodine receptor subtype 3 in native smooth muscle cells. CellCalcium 40: 11–21, 2006.
13. Davis MJ, Hill MA. Signaling mechanisms underlying the vascularmyogenic response. Physiol Rev 79: 387–423, 1999.
479GRAVITY AND HYPERTENSION REGULATE RyR1 IN PORTAL VEIN
J Appl Physiol • doi:10.1152/japplphysiol.00733.2011 • www.jappl.org
on February 2, 2012jap.physiology.org
Dow
nloaded from

14. Delp MD, Colleran PN, Wilkerson MK, McCurdy MR, Muller-Delp J.Structural and functional remodeling of skeletal muscle microvasculatureis induced by simulated microgravity. Am J Physiol Heart Circ Physiol278: H1866–H1873, 2000.
15. Delp MD, Holder-Binkley T, Laughlin MH, Hasser EM. Vasoconstric-tor properties of rat aorta are diminished by hindlimb unweighting. J ApplPhysiol 75: 2620–2628, 1993.
16. Driss-Ecole D, Legué V, Carnero-Diaz E, Perbal G. Gravisensitivityand automorphogenesis of lentil seedling roots grown on board theInternational Space Station. Physiol Plant 134: 191–201, 2008.
17. Dunbar SL, Berkowitz DE, Brooks-Asplund EM, Shoukas AA. Theeffects of hindlimb unweighting on the capacitance of rat small mesentericveins. J Appl Physiol 89: 2073–2077, 2000.
18. Feng JJ, Arendshorst WJ. Calcium signaling mechanisms in renalvascular responses to vasopressin in genetic hypertension. Hypertension30: 1223–1231, 1997.
19. Fritz N, Morel J, Jeyakumar LH, Fleischer S, Allen PD, Mironneau J,Macrez N. RyR1-specific requirement for depolarization-induced Ca2"
sparks in urinary bladder smooth muscle. J Cell Sci 120: 3784–3791,2007.
20. Fu Z, Xie M, Zhang L, Cheng H, Ma J. Differential activation ofpotassium channels in cerebral and hindquarter arteries of rats duringsimulated microgravity. Am J Physiol Heart Circ Physiol 287: H1505–H1515, 2004.
21. Gao F, Bao JX, Xue JH, Huang J, Huang WQ, Wu SX, Zhang L.Regional specificity of adaptation change in large elastic arteries ofsimulated microgravity rats. Acta Physiol Hung 96: 167–187, 2009.
22. Garcha RS, Hughes AD. Action of ryanodine on neurogenic responses inrat isolated mesenteric small arteries. Br J Pharmacol 122: 142–148, 1997.
23. Hargens AR, Watenpaugh DE. Cardiovascular adaptation to spaceflight.Med Sci Sports Exerc 28: 977–982, 1996.
24. Hasser EM, Moffitt JA. Regulation of sympathetic nervous systemfunction after cardiovascular deconditioning. Ann NY Acad Sci 940:454–468, 2001.
25. Helguera G, Eghbali M, Sforza D, Minosyan TY, Toro L, Stefani E.Changes in global gene expression in rat myometrium in transition fromlate pregnancy to parturition. Physiol Genomics 36: 89–97, 2009.
26. House SJ, Potier M, Bisaillon J, Singer HA, Trebak M. The non-excitable smooth muscle: calcium signaling and phenotypic switchingduring vascular disease. Pflügers Arch 456: 769–785, 2008.
27. Iino M, Kasai H, Yamazawa T. Visualization of neural control ofintracellular Ca2" concentration in single vascular smooth muscle cells insitu. EMBO J 13: 5026–5031, 1994.
28. Jaggar JH, Nelson MT. Differential regulation of Ca2" sparks and Ca2"
waves by UTP in rat cerebral artery smooth muscle cells. Am J PhysiolCell Physiol 279: C1528–C1539, 2000.
29. Kanagy NL, Ansari MN, Ghosh S, Webb RC. Recycling and bufferingof intracellular calcium in vascular smooth muscle from geneticallyhypertensive rats. J Hypertens 12: 1365–1372, 1994.
30. Kojima M, Aoki K, Asano M, Fujimoto S, Matsuda T. Malfunction ofarterial sarcoplasmic reticulum leading to faster and greater contractioninduced by high-potassium depolarization in young spontaneously hyper-tensive rats. J Hypertens 9: 783–788, 1991.
31. Lanaspa MA, Andres-Hernando A, Rivard CJ, Dai Y, Berl T. Hyper-tonic stress increases claudin-4 expression and tight junction integrity inassociation with MUPP1 in IMCD3 cells. Proc Natl Acad Sci USA 105:15797–15802, 2008.
32. Li X, Zheng Y, Rathore R, Ma J, Takeshima H, Wang Y. Geneticevidence for functional role of ryanodine receptor 1 in pulmonary arterysmooth muscle cells. Pflügers Arch 457: 771–783, 2009.
33. Lipskaia L, Hulot J, Lompré A. Role of sarco/endoplasmic reticulumcalcium content and calcium ATPase activity in the control of cell growthand proliferation. Pflügers Arch 457: 673–685, 2009.
34. Lipskaia L, Lompré A. Alteration in temporal kinetics of Ca2" signalingand control of growth and proliferation. Biol Cell 96: 55–68, 2004.
35. Liu Q, Zheng Y, Korde AS, Yadav VR, Rathore R, Wess J, Wang Y.Membrane depolarization causes a direct activation of G protein-coupledreceptors leading to local Ca2" release in smooth muscle. Proc Natl AcadSci USA 106: 11418–11423, 2009.
36. Macrez-Leprêtre N, Kalkbrenner F, Morel JL, Schultz G, MironneauJ. G protein heterotrimer Galpha13beta1gamma3 couples the angiotensin
AT1A receptor to increases in cytoplasmic Ca2" in rat portal veinmyocytes. J Biol Chem 272: 10095–10102, 1997.
37. Mariotti M, Maier JAM. Gravitational unloading induces an anti-angio-genic phenotype in human microvascular endothelial cells. J Cell Biochem104: 129–135, 2008.
38. Morel J, Dabertrand F, Fritz N, Henaff M, Mironneau J, Macrez N.The decrease of expression of ryanodine receptor sub-type 2 is reversed bygentamycin sulphate in vascular myocytes from mdx mice. J Cell MolMed 13: 3122–3130, 2009.
39. Morel J, Rakotoarisoa L, Jeyakumar LH, Fleischer S, Mironneau C,Mironneau J. Decreased expression of ryanodine receptors alterscalcium-induced calcium release mechanism in mdx duodenal myocytes.J Biol Chem 279: 21287–21293, 2004.
40. Morel JL, Boittin FX, Halet G, Arnaudeau S, Mironneau C, Miron-neau J. Effect of a 14-day hindlimb suspension on cytosolic Ca2"
concentration in rat portal vein myocytes. Am J Physiol Heart Circ Physiol273: H2867–H2875, 1997.
41. Morel JL, Macrez-Leprêtre N, Mironneau J. Angiotensin II-activatedCa2" entry-induced release of Ca2" from intracellular stores in rat portalvein myocytes. Br J Pharmacol 118: 73–78, 1996.
42. Morey-Holton E, Globus RK, Kaplansky A, Durnova G. The hindlimbunloading rat model: literature overview, technique update and compari-son with space flight data. Adv Space Biol Med 10: 7–40, 2005.
43. Moriyama K, Osugi S, Shimamura K, Sunano S. Caffeine-inducedcontraction in arteries from stroke-prone spontaneously hypertensive rats.Blood Vessels 26: 280–289, 1989.
44. Neylon CB, Richards SM, Larsen MA, Agrotis A, Bobik A. Multipletypes of ryanodine receptor/Ca2" release channels are expressed in vas-cular smooth muscle. Biochem Biophys Res Commun 215: 814–821, 1995.
45. Norsk P. Cardiovascular and fluid volume control in humans in space.Curr Pharm Biotechnol 6: 325–330, 2005.
46. Nyhan D, Kim S, Dunbar S, Li D, Shoukas A, Berkowitz DE. Impairedpulmonary artery contractile responses in a rat model of microgravity: roleof nitric oxide. J Appl Physiol 92: 33–40, 2002.
47. Purdy RE, Duckles SP, Krause DN, Rubera KM, Sara D. Effect ofsimulated microgravity on vascular contractility. J Appl Physiol 85:1307–1315, 1998.
48. Reiberger T, Angermayr B, Schwabl P, Rohr-Udilova N, Mitter-hauser M, Gangl A, Peck-Radosavljevic M. Sorafenib attenuates theportal hypertensive syndrome in partial portal vein ligated rats. J Hepatol51: 865–873, 2009.
49. Samain E, Bouillier H, Miserey S, Perret C, Renaud JF, Safar M,Dagher G. Extracellular signal-regulated kinase pathway is involved inbasic fibroblast growth factor effect on angiotensin II-induced Ca2"
transient in vascular smooth muscle cell from Wistar-Kyoto and sponta-neously hypertensive rats. Hypertension 35: 61–67, 2000.
50. Sayet I, Neuilly G, Mironneau J, Mironneau C. Influence of spaceflight,hindlimb suspension, and venous occlusion on alpha 1-adrenoceptors inrat vena cava. J Appl Physiol 78: 1882–1888, 1995.
51. Sekiguchi F, Kawata K, Komori M, Sunano S. Caffeine-induced con-tracture in oesophageal striated muscle of normotensive and hypertensiverats. Eur J Pharmacol 465: 153–161, 2003.
52. Stevens L, Sultan KR, Peuker H, Gohlsch B, Mounier Y, Pette D.Time-dependent changes in myosin heavy chain mRNA and proteinisoforms in unloaded soleus muscle of rat. Am J Physiol Cell Physiol 277:C1044–C1049, 1999.
53. Vallot O, Combettes L, Jourdon P, Inamo J, Marty I, Claret M,Lompré AM. Intracellular Ca2" handling in vascular smooth muscle cellsis affected by proliferation. Arterioscler Thromb Vasc Biol 20: 1225–1235,2000.
54. Vassy J, Portet S, Beil M, Millot G, Fauvel-Lafève F, Karniguian A,Gasset G, Irinopoulou T, Calvo F, Rigaut JP, Schoevaert D. The effectof weightlessness on cytoskeleton architecture and proliferation of humanbreast cancer cell line MCF-7. FASEB J 15: 1104–1106, 2001.
55. Yates BJ, Kerman IA. Post-spaceflight orthostatic intolerance: possiblerelationship to microgravity-induced plasticity in the vestibular system.Brain Res Brain Res Rev 28: 73–82, 1998.
56. Zhang LF. Vascular adaptation to microgravity: what have we learned?.J Appl Physiol 91: 2415–2430, 2001.
480 GRAVITY AND HYPERTENSION REGULATE RyR1 IN PORTAL VEIN
J Appl Physiol • doi:10.1152/japplphysiol.00733.2011 • www.jappl.org
on February 2, 2012jap.physiology.org
Dow
nloaded from
Related Documents











