SORBITOL CLEARANCE AND ITS EFFECTS ON FEEDLOT PERFORMANCE AND CARCASS CHARACTERISTICS OF STEERS by DENNIS WELDON BOYLES, JR., B.S., M.S. A DISSERTATION IN ANIMAL SCIENCE Submitted to the Graduate Faculty of Texas Tech University in Partial Fulfillment of the Requirements for the Degree of DOCTOR OF PHILOSOPHY Approved December, 1993

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

SORBITOL CLEARANCE AND ITS EFFECTS ON FEEDLOT PERFORMANCE
AND CARCASS CHARACTERISTICS OF STEERS
by
DENNIS WELDON BOYLES, JR., B.S., M.S.
A DISSERTATION
IN
ANIMAL SCIENCE
Submitted to the Graduate Faculty of Texas Tech University in
Partial Fulfillment of the Requirements for
the Degree of
DOCTOR OF PHILOSOPHY
Approved
December, 1993

M
ACKNOWLEDGMENTS C7^^ ""
J) 1 would like to express my gratitude to Dr. C. Reed
Richardson for his support during my graduate studies. He
has helped instill in me a deep appreciation of the
metabolic processes involved in nutrition.
I am grateful to have had distinguished men serve on my
committee. A sincere thank you goes to Drs. R. L. Preston,
R. C. Albin, D. Oberleas and N. A. Cole for their support
and encouragement.
I would like to thank Dr. G. S. Cameron (Texas Tech
Health Sciences Center, Department of Dermatology) for his
vital service to me while laboring through this research.
His guidance, assistance, friendship and the use of his
laboratory are deeply appreciated. I would also like to
thank the staff at the Burnett Center, Victor M. Montalvo-
Lugo and fellow graduate students for their assistance and
friendship.
The support of family and close friends is acknowledged
and appreciated. But the support and years of sacrifice
unselfishly supplied by my wife Janie and our children,
Colton, Caylen and Kaycie, have made the completion of this
research possible.
11

TABLE OF CONTENTS
ACKNOWLEDGMENTS ii
LIST OF TABLES v
LIST OF FIGURES vii
LIST OF ABBREVIATIONS ix
CHAPTERS
I. LITERATURE REVIEW 1
Introduction 1
Animal Performance 1
Metabolism of Sorbitol 2
Properties of Sorbitol 4
Rumen Fermentation of Sorbitol 5
Monosaccharide-Amino Acid Interactions 6 Summary 7
II. EFFECTS OF SORBITOL ON FEEDLOT PERFORMANCE AND CARCASS CHARACTERISTICS OF STEERS FED A STEAM-FLAKED GRAIN SORGHUM BASED DIET 9
Abstract 9
Introduction 10
Materials and Methods 10
Results and Discussion 12
Implications 14
III. EFFECTS OF PROTEIN SOURCE AND SORBITOL SUPPLEMENTION ON PERFORMANCE OF INCOMING FEEDLOT STEERS 28
Abstract 28
Introduction 28
Materials and Methods 29
Results and Discussion 30
Implications. 31
• • •
111

IV. EFFECTS OF SORBITOL ON SODIUM DEPENDENT AND SODIUM INDEPENDENT GLYCINE AND LEUCINE UPTAKE BY CULTURED BOVINE KIDNEY CELLS 38
Abstract 38
Introduction 38
Materials and Methods 40
Results and Discussion 4 3
Implications 44
V. CLEARANCE OF INTRAVENOUSLY
ADMINISTERED SORBITOL IN STEERS 49
Abstract 49
Introduction 50
Materials and Methods 51
Results and Discussion 53
Implications 57
VI. INTEGRATED SUMMARY 66
LITERATURE CITED 69
APPENDIX
A. CELL SUBCULTURE TECHNIQUE 75
B. CELL FREEZING TECHNIQUE 77
C. EXPERIMENTAL PROCEDURE FOR TREATMENT OF
CELLS IN COSTAR CLUSTER WELLS 78
D. PROCEDURE FOR PROTEIN DETERMINATION 80
E. CONSTRUCTION OF EXPERIMENTAL WASH TRAYS 81
F. CONSTRUCTION OF EXPERIMENTAL UPTAKE TREATMENT TRAYS 82
G. FRUCTOSE DETERMINATION IN BOVINE PLASMA 83
IV

LIST OF TABLES
2.1 Composition of initial backgrounding diet 15
2.2 Composition of second backgrounding diet 16
2. 3 Composition of third backgrounding diet 17
2.4 Composition and analysis of final basal diet 18
2.5 One hundred-nineteen day feedlot performance 19
2.6 Initial twenty-eight day feedlot performance 20
2 .7 Initial fifty-six day feedlot performance 21
2.8 Initial eighty-four day feedlot performance 22
2.9 One hundred-twelve day feedlot performance 23
2 .10 Carcass characteristics 24
2.11 Carcass quality grades by treatment 25
3.1 Composition of basal diet 33
3.2 Treatment means for twenty-eight day feedlot performance 34
4.1. Effect of sorbitol on sodium dependent and independent glycine uptake by bovine kidney cells 45
4.2. Effect of sorbitol on sodium dependent and independent leucine uptake by bovine kidney cells 46
5.1. Composition and analysis of basal diet 58
5.2. Mean plasma glucose of steers receiving an intrajugular infusion of either glucose, sucrose, sorbitol or propionate in 50% solutions at 2.2 g kg"^ metabolic weight or equal volume of saline 59
5.3. Mean plasma sorbitol of steers receiving an intrajugular infusion of sorbitol in a 50% solution at 2.2 g kg"^ metabolic weight or equal volume of saline 60

5.4. Mean plasma fructose of steers receiving an intrajugular infusion of sorbitol in a 50% solution at 2.2 g kg"l metabolic weight or equal volume of saline 61
VI

LIST OF FIGURES
2.1. Gain efficiency of steers fed for 119 d 26
2.2. Average daily gain of steers fed for 119 d 27
3.1 Dry matter intake of steers fed either low ruminally degradable protein (LD Protein) mixture or readily ruminally degradable protein (RD Protein) supplement 35
3.2. Average daily gain of steers fed either low ruminally degradable protein (LD Protein) mixture or readily ruminally degradable protein (RD Protein) supplement 3 6
3.3. Gain efficiency of steers fed either low ruminally degradable protein (LD Protein) mixture or readily ruminally degradable protein (RD Protein) supplement 37
4.1. Effects of sorbitol on glycine uptake by cultured bovine kidney cells in the presence of sodium (glycine NA) or in the absence of sodium (glycine WO) 47
4.2. Effects of sorbitol on leucine uptake by-cultured bovine kidney cells in the presence of sodium (leucine NA) or in the absence of sodium (leucine WO) 48
5.1. Plasma glucose levels of steers receiving an intrajugular infusion of either glucose, sucrose, sorbitol or propionate in 50% solutions at 2.2 g kg~^ metabolic weight or equal volume of saline 62
5.2. Plasma glucose levels of steers receiving an intrajugular infusion of either sucrose, sorbitol or propionate in 50% solutions at 2.2 g kg~^ metabolic weight or equal volume of saline 63
5.3. Plasma sorbitol levels of steers receiving an intrajugular infusion of sorbitol in a 50% solution at 2.2 g kg"^ metabolic weight or equal volume of saline 64
Vll

5.4. Plasma fructose levels of steers receiving an intrajugular infusion of sorbitol in a 50% solution at 2.2 g kg~^ metabolic weight or equal volume of saline 65
Vlll

LIST OF ABBREVIATIONS
ADG Average daily gain
BRSV Bovine respiratory syncytical virus
BVD Bovine viral diarrhea
Ca Calcium
cm Centimeters
CP Crude protein
CV Coefficient of variation
d Day(s)
DMI Dry matter intake
DM Dry matter
F:G Feed to gain ratio
g Grams
G:F Gain efficiency (g of gain / kg of feed)
h Hour(s)
HCWT Hot carcass weight
IBR Infectious bovine rhinotracheitis
K Potassium
kg Kilogram(s)
Meal Megacalories
w^5 Metabolic weight (body weight in kg*^5)
NEg Net energy for gain
NEm Net energy for maintenance
P Phosphorus when referring to diet composition
P Probability when referring to statistics
PI3 Parainfluenza
REA Ribeye area
SEM Standard error of the mean
VFA Volatile fatty acid(s)
IX

CHAPTER I
REVIEW OF LITERATURE
Introduction
Sorbitol, a natural six-carbon polyalcohol, is found in
various human foods such as cherries, plums, pears and
apples (Merck, 1989). It is also used as a sweetener. This
compound is regarded as safe for human consumption and is
classified as GRAS (generally recognized as safe) by the
Food and Drug Administration.
Supplementation of sorbitol has been shown to improve
performance of veal calves (Thivend, 1982; Bauchart et al.,
1985). Sorbitol addition to a corn silage-based diet has
also been shown to improve feedlot performance of steers and
bulls. Fontenot and Huchette (1993) reported improvements
in feed efficiency by steers without improved daily gains
after Geay et al. (1992) described improvements in both feed
efficiency and daily gains by bulls. Since cattle feeders
use feed additives and implants to improve animal
performance, sorbitol could be viewed as a natural, safe
compound that could be used as a livestock feed additive
provided it increases profits.
Animal Performance
When orally administered to ruminants, sorbitol has
been shown to improve feed efficiency in calves from birth
to three weeks of age (Daniels et al., 1981) and between 8
and 16 weeks of age in veal calves (Thivend, 1982; Bauchart
et al., 1985). Thivend et al. (1984) have reported an
increase in biliary organic matter flow of young calves when
consuming sorbitol. Improvements in feed efficiency when
supplementing sorbitol to ruminants used for beef production
has also been documented in two reports. Geay et al. (1992)
reported that sorbitol supplied at a concentration of 50 g

d~^ improved ADG and feed efficiency by finishing bulls.
Recent feedyard performance trials conducted by Fontenot and
Huchette (1993) have shown that feeding 20 to 4 0 g of
sorbitol d~^ to finishing steers improves feed efficiency
while not affecting ADG, in two of three studies, with no
marked effect on carcass characteristics. Supplementary
studies conducted by Fontenot and Huchette (1993) indicate
that the improvement in feed conversion by ruminants is not
due to differences in rate of passage or digestibility.
Bauchart et al. (1985) reported a 7% improvement in feed
efficiency from sorbitol supplementation to young veal
calves with no differences in digestibility.
Recent work conducted in France (Fostier, 1992) has
shown that large doses (one kg per animal) of sorbitol given
orally via the drinking water reduced the incidence of dark
cutting carcasses from Holstein cows (n = 2000) by 30%.
Also increased muscle glycogen concentrations in the
longissimus muscle of bulls was reported following sorbitol
consumption. Sorbitol's involvement in muscle glycogen
metabolism has been shown earlier by Johnston and Deuel
(1943) using rats.
Metabolism of Sorbitol
The metabolism of sorbitol is closely related to that
of fructose and glucose (Bye, 1969). Sorbitol dehydrogenase
and aldose reductase are the two enzymes involved in the
metabolism of sorbitol. Large amounts of sorbitol
dehydrogenase are found in hepatocytes and in very low
concentrations in other tissues (Blakley, 1951; Bye, 1969;
Seeberg et al., 1955). Aldose reductase is largely located
in the kidney (Stribling et al., 1989) and plays a minor
role in sorbitol metabolism (Bye, 1969). Early research
indicated that sorbitol dehydrogenase is highly specific for
D-sorbitol, but more recent reports suggest that other

polyols are metabolized by sorbitol dehydrogenase (Malone
and Lowitt, 1992). Sorbitol dehydrogenase catalyses the
following reactions in the presence of NAD or NADH
(Bergmeyer, 1974):
D-Sorbitol 4-> D-Fructose
L-Iditol <-> L-Sorbose
Ribitol 4-> D-Ribulose
Xylitol <^ D-Xylulose Allitol <- D-Allulose
L-Gala-D-glucoheptide <-> L-Galaheptulose
D-Altro-D-glucoheptide <- D-Altroheptulose.
This enzymatic activity involving other polyols could
possibly be related to the improvement in cattle feedlot
performance when cattle consume carrots that contain
mannitol (Richardson, 1993).
Evidence that fructose is the primary intermedate of
hepatic sorbitol oxidation leading to glucose production was
reported by Embden and Greesbach (1914). The enzymatic
oxidation of sorbitol to fructose leading to glucose
production was confirmed by Blakley (1951) in rats.
Sorbitol was rapidly oxidized by rat liver slices;
therefore, Blakley (1951) concluded that in liver slices the
oxidation of sorbitol to fructose with subsequent conversion
of fructose to glucose proceeded much more rapidly than
other hexose oxidation. Chemical reactions involved include
two pathways (Lehninger, 1982). The major pathway in
muscles and the kidney is as follows: sorbitol <-> fructose
<-> fructose-6-phosphate <-> glucose-6-phosphate <-> glucose.
In the liver, the pathway includes three more intermediates
according to the following: sorbitol <-> fructose <->
fructose-1-phosphate <-> glyceraldehyde-3-phosphate <->
fructose-l,6-diphosphate <-> fructose-6-phosphate <-> glucose-
6-phosphate <r> glucose.

Properties of Sorbitol
Sorbitol is (1) rapidly removed from the blood (Stetter
and Stetter, 1951; Hoshi, 1963). Molino et al. (1987) have
described the hepatic clearance of sorbitol to be such that
sorbitol is essentially metabolized and completely extracted
by the hepatocytes. Gianpaolo (1991) states that the
hepatic uptake of sorbitol is by passive transport and the
transport mechanism is unsaturable in the absence of any
substrate competition. Bye (1969) found that sorbitol was
rapidly removed from the blood of 16 humans during and
following intravenous infusion of sorbitol with only 3% of
administered sorbitol escaping via the kidneys. Sorbitol is
presently used in medicine as a compound for the non
invasive test in evaluating liver plasma flow because of its
biologic and kinetic (first-order) features. For a compound
to be used in the non-invasive test in evaluating liver
plasma flow, it must be a substance which is essentially
metabolized by the hepatocytes and completely extracted at
any passage through the liver (Waldstein and Arcilla, 1958;
Winkler et al., 1979; Winkler et al., 1973). The hepatic
clearance of sorbitol largely exceeds liver blood flow
(Molino et al., 1987; Bass and Winkler, 1980). The liver is
a highly vascular organ. In sheep the fraction of cardiac
output received by the liver is 30-40%. The liver contains
an enormous capillary bed which aids in the special effect
that the hepatic parenchyma cells have on substances
entering the organ (Smith and Hamlin, 1977). The amount of
cardiac output flowing through the liver and the enormous
vascular system it possesses sheds light on the rapid
clearance of sorbitol. (2) Sorbitol has been reported to
exhibit protein-sparing properties (Kaufmann, 1929; Griem
and Lang, 1960). Bye (1969) reported that sorbitol was used
as the main source of calories when human patients were fed
intravenously for three months without alteration in

nitrogen balance. Adcock and Gray (1956) concluded that
sorbitol does provide calories to both normal and diabetic
humans based on data collected after an oral dose of 35
grams of ^^c sorbitol. (3) Sorbitol possesses antiketogenic
properties that have been reported to be superior to
fructose. Early work reported by Edson (1936) showed that
sorbitol was more effective than fructose or glucose in
suppressing spontaneous ketogenesis in liver slices obtained
from starved rats.
A vitamin-sparing action of sorbitol has also been
reported. Morgan and Yudkin (1957) have shown that sorbitol
produces a change in the intestinal flora of rats so that
thiamin is synthesized and is available to the host. The
tissues of rats surviving on thiamin-free diets plus
sorbitol contained more thiamin than those rats on thiamin-
free diets when glucose was supplemented. Thus thiamin was
made available to the tissues, though not from the diet.
Supplementary studies conducted by Morgan and Yudkin (1957)
suggested that the administration of sorbitol appears to
make the rat independent of dietary sources not only of
thiamin but also of the other B-complex vitamins. This
reported action of sorbitol's involvement in thiamin
metabolism in rats if functional in ruminants could possibly
play a preventive role when cattle suffer from
polioencephalomalacia (Blood and Henderson, 1974).
Rumen Fermentation of Sorbitol
Little information concerning rumen fermentation of
sorbitol exists. No differences in VFA molar proportions 6
h after finishing bulls were fed 0 or 50 g of sorbitol d~l
were reported by Geay et al. (1992). However, they did find
an increase (P < .001) in propionic acid production in vitro
with a decrease in butyric and isovaleric acids along with a
reduction (P < .001) in the acetic:propionic ratio.

Improvements in feed efficiency elicited by sorbitol appears
to decline with time. The decline has occurred after 56 to
70 d when ruminants were fed 20 to 50 g of sorbitol animal"^
d"l without altering rate of passage or digestibility
(Fontenot and Huchette, 1993; Geay et al., 1992,
respectively). Geay et al. (1992) postulated that the
decline in the beneficial effect of sorbitol in improving
performance by rumanints could be attributed to two factors:
a decreasing growth potential of of the host and an
increasing capacity of ruminal microorganisms to metabolize
sorbitol.
Monosaccharide-Amino Acid Interaction
Monosaccharides such as glucose, galactose and fructose
derived from dietary sources have been reported to alter
amino acid transport (Reiser and Hallfrisch, 1987). Glucose
and galactose have generally inhibited the transport of
neutral amino acids, while fructose, a derivative of
sorbitol, has been documented to increase neutral amino acid
transport. In vitro, cellular uptake of leucine,
isoleucine, valine, phenylalanine, tryptophan and histidine
were increased 24 to 57% by the presence of fructose in the
incubation media (Reiser and Christiansen, 1971; Reiser et
al., 1975; Reiser and Hallfrisch, 1977). In vivo, fructose
has also stimulated neutral amino acid transport by intact
intestine (Reiser and Christiansen, 1969; Alvarado, 1968).
The effectiveness of fructose in stimulating leucine
transport appears to exist whether fructose occupies
intracellular or extracellular space (Reiser and Hallfrisch,
1987). They also have stated that the component of leucine
transport stimulated by intracellular fructose is dependent
on metabolic energy and the presence of a sodium gradient.
Sorbitol is a monosaccharide closely related to fructose.
Sorbitol and fructose are enzymaticaly interconverted by

sorbitol dehydrogenase (SDH; EC 1.1.1.14). Fructose is
reduced according to the following reaction:
Sorbitol + NAD"*" <- SDH _ Fructose + NADH + H"*".
Since sorbitol is a monosaccharide closely related to
fructose, and fructose has been reported to affect amino
acid transport, the effect of sorbitol on amino acid
transport should be examined.
Summary
In summary, data suggest that sorbitol is oxidized more
rapidly than other hexoses (Blakley, 1951), it is
antiketogenic (Edson, 1936) and its primary role in
metabolism pertains to energy utilization. Whether this
effect results from conversion of sorbitol into energy
intermediates or due to gastrointestinal tract changes that
promote better energy utilization such as the vitamin-
sparing action or due to some protein-sparing effect or to
all of the above is at present unknown.
Improvements in feed efficiency and sometimes ADG when
feeding sorbitol to ruminants has been reported by several
researchers. One could hypothesize that sorbitol improves
performance of cattle by providing added substrate used in
gluconeogenesis because sorbitol is quickly converted to
fructose and then to glucose. Since ruminants depend upon
gluconeogenic precursors (i.e., propionate and amino acids)
for their glucose needs, the gluconeogenic property of
sorbitol could possibly provide ruminants an additional
gluconeogenic precursor. Additionally, monosaccharides
closely related to sorbitol differentially affect amino acid
transport. It has not been determined if sorbitol affects
amino acid transport.
The mechanism and(or) mechanisms involved, associated
with sorbitol, in improving performance of ruminants is

presently unknown. Therefore, it was the intent of the
following research to determine the effects of sorbitol
supplementation to a steam-flaked grain sorghum-based diet
fed to steers; to determine the effects of sorbitol on amino
acid uptake by cultured bovine kidney cells; and to
determine sorbitol clearance in steers.
8

CHAPTER II
EFFECTS OF SORBITOL ON FEEDLOT PERFORMANCE
AND CARCASS CHARACTERISTICS OF
STEERS FED A STEAM-FLAKED
GRAIN SORGHUM-BASED DIET
Abstract
A feedlot experiment with growing/finishing steers was
conducted to determine the effects of supplementing a steam-
flaked grain sorghum-based diet with sorbitol on voluntary
feed intake, rate of gain, feed efficiency and carcass
characteristics. One hundred twelve crossbred steers
(Angus/Hereford; average initial shrunk weight = 337.3 ±
17.4 kg) were randomly assigned to treatment and fed a
steam-flaked grain sorghum-based diet with or without
sorbitol. Treatments were: A - control (basal diet); B -
basal diet plus 30 g sorbitol steer"^ d"^; C - basal diet
plus sorbitol at a variable rate (20 g sorbitol steer"^ d~^
first 28 d, 30 g second 28 d, then 40 g until termination of
the study); D - basal diet for first 69 d then addition of
30 g of sorbitol steer"^ d~^ until termination of the study.
Supplementation of sorbitol did not statistically improve
steer feedlot performance (P > .05) over the 119 d feeding
period. However, throughout the feeding period,
supplementing steers with 30 g of sorbitol d"^ showed a 3.4%
numerical increase in ADG and a 4.0% numerical improvement
in feed efficiency over steers receiving no sorbitol.
Steers receiving sorbitol only for the final 50 d had lower
dressing percent (P < .002) than other steers. Furthermore,
the carcass lean color tended (P = .16) to be altered by a
treatment effect . Steers receiving 30 g of sorbitol d"^
appeared to exhibit a more youthful bright cherry red color
of the carcass lean (P = .03). In summary, sorbitol

supplementation to a steam-flaked grain sorghum-based diet
did not improve feedlot performance of steers in this study.
However, steers fed 30 g of sorbitol d~l for the last 50 d
did exhibit (P < .002) lower dressing percentages.
Furthermore, steers receiving 30 g of sorbitol d~^ appeared
to exhibit (P = .03) a more youthful bright cherry red color
of carcass lean.
Introduction
Sorbitol, a natural six-carbon alcohol found in many
fresh fruits, has been reported to improve feed conversion
in calves (Daniels et al., 1981; Thivend, 1982; Bauchart et
al., 1985) and in finishing bulls (Geay et al., 1991). Two
of three feedlot experiments reported by Fontenot and
Huchette (1993) have shown improvements in feed efficiency
by finishing steers when sorbitol was supplemented to a corn
silage-based diet alone or in combination with monensin.
Thus, the objectives of this study were to determine the
effect of sorbitol supplementation to a steam-flaked grain
sorghum-based diet with monensin on feedlot performance and
carcass characteristics of growing/finishing steers.
Materials and Methods
One hundred and twelve crossbred steers (Angus/
Hereford; average initial shrunk weight = 337.3 ±17.4 kg)
were randomly assigned to treatment and fed a steam-flaked
grain sorghum-based diet with or without sorbitol.
Treatments were: A - control (basal diet); B - basal diet
plus 30 g sorbitol^ steer"^ d~^; C - basal diet plus
sorbitol at a variable rate (20 g sorbitol steer"! d~! first
1 Neosorb^Sorbitol supplied by Roquette Corporation, Gurnee, IL 60031-2392.
10

28 d, 30 g second 28 d, then 40 g until termination of the
study); D - basal diet for first 69 d then addition of 30 g
of sorbitol steer"! d"! until termination of the study.
Upon arrival at the research facility steers were all
weighed, ear tagged, and vaccinated against BVD, BRSV, IBR,
PI32, and Clostridium perfringens Types C and D^. Steers
were gradually placed on the final basal diet using three
diets over 28 d by decreasing roughage source and increasing
the amount of steam-flaked grain sorghum. Each
backgrounding diet (Tables 2.1, 2.2 and 2.3) was fed for at
least five d. The steers were implanted with Synovex-S^ at
the start of the experiment and at d 56. Steers were
weighed initially and at 28 d intervals through 112 d.
Steers reached market weight by 112 d and remained on
treatment an additional 7 d until slaughter was scheduled.
At the beginning and end (119 d) of the feeding period two
consecutive weights were taken to obtain full and shrunk
weights. Shrunk weights were obtained after 24 h without
feed and overnight without water. Additionally, steers fed
sorbitol starting on d 70 (treatment D) were weighed on d 69
before the morning feeding.
All diets were formulated to meet or exceed NRC (1984)
recommended requirements for CP, Ca, P and K. The final
basal diet is shown in Table 2.4. Sorbitol was added (via a
premix using ground grain sorghum as the carrier) to the
diets at 1% of diet DM. The amount of sorbitol premix was
2 Horizon IV. Bovine Rhinotracheitis-Virus Diarrhea-Parainfluenza3-Respiratory Syncytial Virus Vaccine, Modified Live and Killed Virus, Diamond Scientific, Company, Des Moines, lA.
3 Clostridium perfringes Types C & D, Coopers Animal Health, Inc., Kansas City, KS 66103.
4 Syntex Animal Health, 4800 Westown Parkway, W. Des Moines, lA 40265.
11

adjusted daily if needed to ensure proper sorbitol
consumption. Steers were maintained on treatment for 119 d.
Two steers were removed from the experiment; one steer on
treatment D died from bloat 22 d after initiation of the
study, and the other steer was removed from treatment B due
to hoof injury on d 84. Carcass data were obtained at a
commercial beef packing plant by trained university meat
science personnel. Livers were scored using the Elanco
abscessed liver classification system^.
There were four replications (pens) of seven steers
pen"! on each of the four treatments. Pens of steers served
as the experimental units in a completely randomized design.
The model included treatment and data were analyzed by
analysis of variance using the General Linear Model
procedure of SAS (1990). Treatment means were separated by
Tukey's Studentized Range test.
Results and Discussion
Supplementation of sorbitol over the 119 d feeding
period tended to alter gain efficiency (g of gain kg"! of
feed; P = .09; Table 2.5) of steers while DMI (P = .59) and
ADG (P = .19) were not affected. Numerical improvements
were exhibited by steers fed 30 g of sorbitol d"! (treatment
B) throughout the feeding period in G:F (P > .05; Figure
2.1) and ADG (P > .05; Figure 2.2) compared to steers fed no
sorbitol (treatment A) (175 vs. 168 [4.0 %] and 1.46 vs.
1.51 [3.4 % ] ; G:F and ADG; A and B, respectively; Table
2.5). Data summarized throughout the feeding period (Tables
2.6, 2.7, 2.8 and 2.9) indicate the numerical improvements
in steer performance from supplementing 30 g of sorbitol.
Steers receiving sorbitol at the variable rate (treatment C)
5 Liver Abscess Technical Information. Elanco Products Company, Cattle Products Marketing, Lilly Corporate Center, Indianapolis, IN 46285.
12

had lower ADG (P < .05) and G:F (P < .05) for the initial 28
d when fed 20 g of sorbitol d"! (Table 2.6), and thereafter
their performance appeared to be adversely affected (P >
.05), as exhibited in Tables 2.6, 2.7, 2.8 and 2.9.
Carcass data are shown in Table 2.10. Steers receiving
sorbitol only after 69 d (treatment D) had lower dressing
percentages (P < .002) than steers receiving the three other
diets. Dressing percent (hot carcass weight/live weight x
100) pertains to the carcass yield. It is a function of
gastrointestinal fill and carcass fat; therefore, fatter
cattle will usually dress higher (Romans et al., 1985). The
steers in this study were managed in the same manner, so it
was unlikely that fill was a factor. Carcass data such as
ribeye area and fat measurements did not show strong
differences that would help explain why the steers fed 30 g
of sorbitol d"! for last 50 d had lower dressing
percentages.
Quality grades were similar (P = .60). When observing
the quality grades (Choice, Select or Standard) assigned to
carcasses within each treatment as shown Table 2.11, it
appears that steers fed sorbitol had fewer carcasses that
graded Choice. Overall, the percentage of carcasses that
graded choice was 19% which is undesirable. Based on
carcass value, the target set for carcasses grading Choice
is at least 60%. The genotype and weight of these steers
would indicate a higher percentage of carcasses grading
Choice. The reason for the low percent of carcasses grading
Choice in this experiment is unclear. An overall treatment
effect tended (P = .16) to alter the color of carcass lean
tisssue. Steers supplemented sorbitol at a constant rate of
30 g d"! (treatment B) appeared to exhibit (P = .03)
carcasses with a more youthful bright cherry red color of
lean tissue.
13

As shown in Table 2.10 sorbitol tended to affect the
severity (P = .17) and the incidence of liver abscesses
(P = .11) of steers. An important note to realize when
considering the liver abscess data in this experiment is the
absence of antibiotic formulated into the diets as requested
by the funding agency.
In conclusion, the addition of sorbitol to steam-flaked
grain sorghum-based diets when fed to growing/finishing
steers in this experiment did not improve feedlot
performance of the steers. This is in contrast to results
reported by Fontenot and Huchette (1993) when finishing
steers were fed a corn silage-based diet supplemented with
sorbitol. However, steers fed 30 g of sorbitol d"! for the
last 50 d did exhibit (P < .002) a lower dressing
percentage. Also steers receiving 30 g of sorbitol d~! for
119 d exhibited (P = .03) a more youthful bright cherry red
color of carcass lean.
Implications
Supplementing sorbitol to steam-flaked grain sorghum-
based diets in this experiment did not improve feedlot
performance of steers. This is in contrast to data reported
by Fontenot and Huchette (1993) and Geay et al. (1992).
They reported that feedlot performance was improved when
sorbitol was supplemented to high corn silage-based diets.
Carcass characteristics were not altered except for (1)
steers fed 30 g of sorbitol d"! for the last 50 d had lower
dressing percentages (P < .002), and (2) steers receiving 30
g of sorbitol d~! for 119 d appeared to exhibit (P = .03) a
more youthful bright cherry red color of carcass lean.
Except for these two carcass characteristics, carcass data
exhibited in this study support earlier work reported by
Fontenot and Huchette (1993) and Geay et al. (1992).
14

Table 2.1. Composition^ of initial backgrounding diet.
Item Percent
Corn silage 56.40
Cottonseed hulls 31.00
Cottonseed meal 7.11
Urea .16
Molasses, cane 2.50
Animal/vegetable fat 1.20
Calcium carbonate .36
Dicalcium phosphate .20
Sodium chloride .16
Trace mineral premix^ .17
Vitamin A premix^ .21
AS700 premix^ .53
^ As-fed basis.
b Contained (ppm) I 1,232, Mn 8,069, Zn 8,409, Cu 827, Co
51, Fe 4,056.
c Contained 660,000 lU vitamin A kg"!.
^ Contained 6.16 g aureomycin and sulfamethazine kg"!.
15

Table 2.2. Composition^ of second backgrounding diet.
Item Percent
Corn silage 40.00
Steam-flaked grain sorghum 20.60
Cottonseed hulls 25.00
Cottonseed meal 7.77
Urea .25
Molasses, cane 3.30
Animal/vegetable fat 1.20
Calcium carbonate .65
Dicalcium phosphate .15
Sodium chloride .11
Trace mineral premix^ .22
Vitamin A premix^ .25
AS7 00 premix^ :_50
^ As-fed basis.
b Contained (ppm) I 1,232, Mn 8,069, Zn 8,409, Cu 827, Co
51, Fe 4,056.
c Contained 660,000 lU vitamin A kg"!.
^ Contained 6.16 g aureomycin and sulfamethazine kg"!.
16

Table 2.3. Composition^ of third backgrounding diet
Item Percent
Steam-flaked grain sorghum 62 . 05
Cottonseed hulls 23.47
Cottonseed meal 6.63
Urea .56
Molasses, cane 3.85
Animal/vegetable fat 1.12
Calcium carbonate .99
Dicalcium phosphate .04
Sodium chloride .21
Potassium chloride .09
Trace mineral premix^ .24
Vitamin ADE premix^ .33
Rumens in premix^ .42
^ As-fed basis.
^ Contained (ppm) I 1 232, Mn 8 069, Zn 8 409, Cu 827, Co
51, Fe 4056.
^ Contained (lU/kg) Vitamin A acetate 634,480; Vitamin D 63,448; Vitamin E 1,813.
^ Contained monensin at 3,044.8 mg/kg.
17

Table 2.4. Composition^ and analysis of final basal diet.
Item Percent
Steam-flaked grain sorghum 73.65
Cottonseed hulls 9.34
Cottonseed meal 7.47
Urea .37
Molasses, cane 3.27
Animal/vegetable fat 2.12
Calcium carbonate 1.27
Sodium chloride .17
Trace mineral premix^ .24
Vitamin ADE premix^ .33
Rumensin premix^ .84
Control premix® .93
Sorbitol premix^ .00
Analysis^
DM, % 83.03
CP, % !3.03
.67
.32
.67
Ca, %
P, % K, % NEm, Meal/kg 3.78
NEg, Mcal/kg !'25 ^ As-fed basis. b Contained (ppm) I 1,232, Mn 8,069, Zn 8,409, Cu 827, Co 51, Fe 4056. c Contained (lU/kg) Vitamin A acetate 634,480; Vitamin D 63,448; Vitamin E 1,813. ^ Contained monensin at 3,044.8 mg/kg. e Contained 2.0% mineral oil and 98.0% ground grain sorghum f Contained 2.0 % mineral oil, 23.6% ground gram sorghum and 74.4% sorbitol. g Analyzed, except for NEm and NEg, which were calculated, on a DM basis except for DM.
18

Table 2.5. One hundred-nineteen day feedlot performance^
Treatment
A B C
Item
Variable Start 30
Control 30 g/d rate g on d 70 SEM^ P
Initial
wt^, kg
DMI, kg
ADG, kg
G:F^
F:Ge
Final
wt, kg
340.5
8.68
1.46
168
5.93
514.2
338.6
8.65
1.51
175
5.73
518.3
330.9
8.39
1.38
164
6.08
495.1
334.5
8.48
1.46
172
5.80
508.2
8.68
.174
.040
2.71
.094
16.15
.35
.59
.19
.09
.09
.19
^ 119 d performance using shrunk weights.
^ Standard error of the mean.
^ Average initial shrunk weight after one d off feed and
without water overnight.
^ Gain efficiency = g of gain kg"! of feed.
® Feed to gain ratio.
19

Table 2.6. Initial twenty-eight day feedlot performance^.
Item
DMI,
ADG,
G:F®
F:Gf
kg
kg
A
Control
8.09
2.39C
295C
3.39C
Tr€
B
30 g/d
7.94
2.35C
296C
3.37C
jatment
C
Variable
rate
7.62
1.95^
256^
3.91^
D
Start 30
g on d 70
7.79
2.I2C
272Cd
3.5lCd
SEM^
.159
.111
9.08
.115
P
.25
.04
.02
.02
^ 28 d performance summary.
^ Standard error of the mean.
C/d Means in same row with different superscript differ.
® Gain efficiency = g of gain kg"! of feed.
^ Feed to gain ratio.
20

Table 2.7. Initial fifty-six day feedlot performance^
Treatment
A B C D
Item
DMI,
ADG,
G:FC
F:G^
kg
kg
Control
8.50
1.97
232
4.32
30 g/d
8.33
1.97
236
4.23
Variable
rate
8.04
1.81
225
4.44
Start :
g on d
8.15
1.90
233
4.30
30
70 SEM^
.143
.049
4.77
.089
P
.16
.13
.45
.45
^ 56 d performance summary.
^ Standard error of the mean.
^ Gain efficiency = g of gain kg"! of feed.
^ Feed to gain ratio.
21

Table 2.8. Initial eighty-four day feedlot performance^
__ ^ ^ __ ^ Treatment
A
Item Control
B C D
Variable Start 30
30 g/d rate g on d 70 SEM
DMI,
ADG,
G : F C
F:G^
kg
kg
8 . 6 6
1 . 7 2
1 9 9
5 . 0 4
8 . 5 1
1 . 7 7
2 0 8
4 . 8 1
8 . 2 7
1 . 5 9
192
5 . 2 0
8 . 2 9
1 . 6 6
2 0 0
5 . 0 0
. 1 6 5
. 0 5 6
3 . 8 7
. 0 9 7
. 3 2
. 1 8
. 1 0
. 1 0
^ 84 d performance summary.
^ Standard error of the mean.
^ Gain efficiency = g of gain kg"! of feed.
^ Feed to gain ratio.
22

Table 2.9. One hundred-twelve day feedlot performance^
Treatment
A B C D
Item Control
Variable Start 30
30 g/d rate g on d 70 SEM
DMI,
ADG,
G : F C
F:G^
k g
k g
8 . 7 0
1 . 6 7
1 8 8
5 . 3 2
8 . 6 7
1 . 6 8
194
5 . 1 5
8 . 3 8
1 . 5 4
1 8 3
5 . 4 7
8 . 4 7
1 . 5 8
1 8 5
5 . 3 6
. 1 7 5
. 0 4 2
3 . 8 0
. 1 0 8
. 5 3
. 1 3
. 2 9
. 2 9
^ 112 d performance summary.
^ Standard error of the mean.
^ Gain efficiency = g of gain kg"! of feed.
^ Feed to gain ratio.
23

Table 2 . 1 0 . Carcass c h a r a c t e r i s t i c s .
Treatment B
Item Variable Start 30 g
Control 30 g/d ra te on d 70 SEM^
HCWT^*, k g
Dress ing % REA®, cm2
B a c k f a t ^ ,
cm
KPH, %
Y i e l d
graded
Marbling
score^
Quality
grade^
Lean
colorD
Liver
score^
318.4
62.1^
74.5
1.30
1.75
3.1
316.0
61.5C
76.5
1.24
1.69
2.9
307.2
61.9^
75.5
1.22
1.85
2.9
452.8 437.5 444.5
10.7
4.7
.70
10.3
5.3
.80
10.5
5.0
1.17
312.6
59.8«i
79.4
1.09
1.76
2.6
434.5
10.3
4.9
.64
4.91
.287
2.23
.102
.059
.199
11.7
.218
.165
.170
34.5 9.40
43
0004
49
85
36
44
,70
60
16
17
11
Liver abscess incidence!, %
39.3 38.6 66.7 ^ Standard error of the mean. ^ Hot carcass weight. C/d Means in same row with different superscript differ. ® Ribeye area in square, cm. f Backfat thickness, cm. g Calculated using USDA yield grade equation. h Marbling score; slight = 400; small = 500. i Quality grade; Choice"= 12; Select"*"=ll; Select"=10. J Lean color; light = 8; dark = 1. ^ Liver score using the Elanco abscess liver classification system. ! Liver abscess incidence by treatment; percent of steers with at least one liver abscess.
24

Item
Choice"
Select"*"
Select"
Standard"*"
A
Control
32.1
10.7
50.0
7.2
B
Treatment
30 g/d
15.4
15.4
57.7
11.5
C
Variable
rate
15.4
26.9
53.8
3.9
D
Start 30 g
on d 70
14.8
7.4
74.1
3.7
^ Percent of carcasses in each grade by treatment.
25

310 T
'S290 fSi ^ 2 7 0 o
^ 2 5 0 ••
1 2 3 0 ^ 2 1 0
^190
0 170
150 28 56 — \ —
84 Days
112 119
-•- Control -©-Variable
^ 30 g/d -^ 30 g/d last 50 d
Figure 2.1. Gain efficiency of steers fed either basal diet (Control), 30 g sorbitol steer"! d"! (30 g/d), variable rate (Variable) (20 g first 28 d. 30 g second 28 d, then 40 g of sorbitol steer"! (j-l until termination) , or 30 g sorbitol steer"! j-l only after 69 d (30 g/d last 50 d) (SEM = 2.71).
26

2.40 T
2.20 -
2.00 -^
01.80 -
^1.60 +
1.40 +
1.20 28 56
-•- Control -©-Variable
+ + 84 112 119
Days
^ 3 0 g/d -& 30 g/d last 50 d
Figure 2.2. Average daily gain of steers fed either basal diet (Control), 30 g sorbitol steer"! ^-1 (30 g/d), variable rate (Variable) (20 g first 28 d, 30 g second 28 d, then 40 g of sorbitol steer"! ^-1 until termination), or 30 g sorbitol steer"! d"! only after 69 d (30 g/d last 50 d) (SEM = .040).
27

CHAPTER III
EFFECTS OF PROTEIN SOURCE AND SORBITOL
SUPPLEMENTATION ON PERFORMANCE
OF INCOMING FEEDLOT
STEERS
Abstract
A 28 d experiment was conducted to evaluate the
effects of protein source and level of sorbitol
supplementation on performance of incoming crossbred steers
(262 ±21.7 kg; n = 260) using a randomized block design
with a 2 X 3 factorial arrangement of treatments. Protein
source was either a low ruminally degradable (LD) mixture or
a readily ruminally degradable (RD) protein supplement.
Sorbitol was fed at either 0, 30 or 60 g steer"! d"!.
Feedlot performance measures (DMI, ADG, gain efficiency,
G:F, g of gain kg"! feed) were not improved by feeding LD
protein or by sorbitol supplementation during- the 28 d
receiving period. However, there was a tendency (P = .18)
for an interaction between protein source and level of
sorbitol for gain efficiency.
Introduction
Sorbitol has been reported to improve feed conversion
in calves and finishing steers (Fontenot and Huchette, 1993)
and finishing bulls (Geay et al., 1992) when supplemented in
corn silage-based diets. Geay et al. (1992) fed sorbitol
(50 g d"!) plus low ruminally degradable protein to
finishing bulls and reported increased ADG (+ 18%) and feed
efficiency (+ 14%). However, when supplementing sorbitol to
a steam-flaked grain sorghum-based diet only numerical
improvements in feed efficiency and daily gains by finishing
steers have been reported (Boyles and Richardson, 1993).
28

Cattle arriving at feedlots are usually stressed
because of transport and from being without feed; thus these
animals are depleted of nutrients. This problem is
confounded by the low DMI of these cattle during the
receiving period. Thus, it is important to formulate
receiving diets properly to restore lost body nutrients.
Eck et al. (1988) suggested that incoming feedlot cattle
have a requirement for low ruminally degradable protein.
Therefore, because of the benefit of supplementing sorbitol
to veal calves, steers and bulls in conjunction with
evidence for improved 28 d performance of cattle fed a
source of low ruminally degradable protein, it seemed
fitting to conduct a 28 d receiving study to evaluate the
effects of protein source and level of sorbitol
supplementation on performance of incoming crossbred steers.
Materials and Methods
A 28 d study was conducted to evaluate the effects of
protein source and level of sorbitol supplementation on
performance of incoming crossbred steers! (262 ± 21.7 kg; n
= 260) using a randomized block design with a 2 x 3
factorial arrangement of treatments. Pen location within
the feedlot served as blocks. Protein source was either a
low ruminally degradable (LD) protein mixture (Preston and
Bartle, 1989; blood meal, 15%; corn gluten meal, 8%;
cottonseed meal, 22%; hydrolyzed feather meal, 33%; and meat
and bone meal, 22%) or a more readily ruminally degradable
(RD) source (soybean meal). Sorbitol was supplemented at 0,
30 or 60 g steer"! (j-l. Upon arrival at the Texas Tech
feedlot, steers were weighed, ear tagged, dewormed^ and
! Source: Oklahoma City Livestock Auction, OK.
2 ivomec-F, MSD-AGVET, Division of Merck and Company, Inc., Pathway, NJ 07065.
29

immunized against BVD, BRSV, IBR, PI33 and Clostridium
perfringens type C and D^. Steers were fed a steam-flaked
grain sorghum, cottonseed hull-based diet (Table 3.1) for 28
d. Sorbitol^ (granular form) was supplemented via a premix
with ground sorghum used as the carrier. Sorbitol premix
(85% sorbitol, 13% ground grain sorghum plus 2% mineral oil)
and a control premix (contained 98% ground grain sorghum
plus 2% mineral oil and no sorbitol) were adjusted daily if
needed based on pen feed intake to insure proper sorbitol
consumption.
There were five replications (pens) of eight or nine
steers pen"!, on each of the six treatments. One pen of
nine steers fed LD protein plus 30 g sorbitol d"! were
removed from analysis because the diet was inadvertently
changed to LD protein without sorbitol seven d after
initiation of the experiment. Pens of steers served as the
experimental units in a randomized block design with a 2 x 3
factorial arrangement of treatments. Pen location served as
the blocks. The model included blocks, protein source,
sorbitol level and protein source*sorbitol level
interaction. Data were analyzed by analysis of variance
using the GLM procedure of SAS (1990).
Results and Discussion
No interactions existed between protein source and
level of sorbitol for DMI (P = .75; Figure 3.1). There was
3 Horizon IV. Bovine Rhinotracheitis-Virus Diarrhea-Parainfluenza3-Respiratory Syncytial Virus Vaccine, Modified Live and Killed Virus. Diamond Scientific, Company, Des Moines, lA.
4 Clostridium perfringens Types C & D, Coopers Animal Health, Inc., Kansas City, KS 66103.
5 Neosorb^ Sorbitol supplied by Roquette Corporation, Gurnee, IL 60031-2392.
30

a tendency for an interaction between protein source and
level of sorbitol for ADG (P = .20; Figure 3.2) and gain
efficiency (P = .18; Figure 3.3). The low ruminally
degradable protein mixture tended to lower DMI, not affect
ADG and tended to improve G:F (Table 3.2; 8.0 vs. 8.21, P =
.20; 2.09 vs. 2.06, P = .69; 262 vs. 250, P = .21; DMI, ADG,
and G:F for LD vs. RD, respectively). Feedlot performance
of steers was not improved by sorbitol supplementation
(Table 3.2; 7.96, 8.30, 8.04, P = .27; 2.02, 2.16, 2.04, P =
.44; 254, 260, 255, P = .90; DMI, ADG and G:F, for 0, 30 and
60 g of sorbitol, respectively).
The overall means and corresponding coefficients of
variation (CV) for DMI, ADG and G:F were 8.11, CV = 5.58%;
2.01, CV = 11.16%; and 256, CV = 8.95%, respectively. The
average DMI of steers in this receiving study was 2.8% of
body weight (body weight = average of initial and 28 d
weights). Dry matter intake of this magnitude for incoming
steers is quite good. Therefore, a possible explanation for
the lack of statistical effect found in this experiment
could be associated with the DMI of these steers during the
28 d receiving period.
In conclusion, performance of steers during the 28 d
receiving period was not improved by feeding a low ruminally
degradable protein mixture or by sorbitol supplementation at
either 3 0 or 60 g steer"! day"!; however, there was a
tendency for an interaction between protein source and
sorbitol level for feed efficiency.
Implications
Under stressful conditions when cattle usually lower
their feed intake, a feed additive such as sorbitol,
theoretically should benefit the animal. Likewise, low
ruminally degradable protein has been shown to improve 28 d
31

incoming performance of cattle. However, this study did not
produce data supporting a benefit of sorbitol
supplementation or a low ruminally degradable protein source
to incoming feedlot steers. The lack of response by steers
in this experiment could be attributed to DMI.
32

Table 3.1. Composition of basal diets^.
Item
Readily ruminally degradable
Low ruminally degradable
% %
Steam-flaked grain sorghum Cottonseed hulls Molasses, cane Animal fat, #2 yellow grease Urea Sodium chloride Calcium carbonate Dicalcium phosphate Potassium chloride Trace mineral premix^ Vitamin premix^ AS700 premix^ Soybean meal Escape protein mixture® Sorbitol premix^ Control premixg
Analysis^ DM, % CP, % Ca, % P, % K, % NEm, Mcal/kg NEg, Mcal/kg
39.88 40.08 3.00 1.50 .17 .20 .80 .22 .10 .35 .25 .75 00 .00 .00
1.70
86.52 13.01
.50
.28 1.04 1.69 .86
11
39.88 40.08 3.00 1.50 .17 .20 .80 .22 .10 .35 .25 .75 4.5 5.4 .00
1.70
86 13
1 1
55 02 55 28 00 63 77
^ Dry matter basis. b Contained (ppm) I 1,232, Mn 8,069, Zn 8,409, Cu 827, Co 51, Fe 4,056. c Contained 660,000 lU kg"! vitamin A. ^ Contained 6.16 g aureomycin and sulfamethazine kg"!. 6 Contained 15 % blood meal, 8 % corn gluten meal, 22 % cottonseed meal, 33 % hydrolyzed feather meal, and 22 % meat and bone meal. f Contained 2.0 % mineral oil, 23.6 % ground grain sorghum and 74.4 % sorbitol. g Contained 2.0 % mineral oil and 98.0 % ground grain sorghum. h DM basis except for DM.
33

.
a) o c <c s u 0
<Mh
Q)
4-> 0
i H
feed
> <o
43 tr •H
1
• P
c 0)
*• u 0
(M
to c (0 0) S i r
.nally
rotein
rumi
le p:
^ 4 2 ^ <0
S« 1 ^ «-§
c • H
> . < " . ^ • P Zi 0
0)1 ^ TJ g 1 ^ (0
4J (0 0) v^ EH
• CM
. ro
0) f H A
Hi
^ & 0)
•0
to Q) 3
i H
>
(I4
T3
"5
0 -p • H
4:3 ^ 0 in
43 t/D
X
04
Sorb
4J 0 U a*
(d
w W
0 VO
0 CO
0
0 VO
0
en
0
g 0)
4-> H
•^ ' t
cn
n ^
^ CO
CO
CM
0 H
265.6
60.7
r
"(t •
H VO CM
• CM VO CM
in •
00 in CM
CM
CM
tn 0 r-i X u (d (1) T H «-
Si -P 6 -H 4J ;3 c :? a H
in
•
r CM
•
0 CM
•
0 CM CM
•
8.08
0 'St
.
CO
VO
8.1
0 0
. 00
0 CM
• 00
VO
• r
tr M
^ H
s Q
0 CM
•
^
•
OS VO
•
in CM H
• 1.92
in H
. CM
0
2.1
VO H
• CM
VO H
• CM
in OS
•
H
T3 •0 0)
t r 0) >J <w
• t
* C tJ -H Q (C < 0
00 H
•
0 ON
•
H CM
•
• H H
238
256
1 ^
in CM
H t^ CM
VO CM
H in CM
tJ> ^ ' ^ 0>
^ r^ H
•
4.20
H ON
.
0
3.9
0 t^
. en
0 00
n
00 OS
• m
0) c
rH 00
* CM H
319.4
20.9
n
CM •
0 CM
0
• n CM n
0 •
ON H
00 •
H CO
^ .
•H 4J (d IS 0 • • i H
TJ <d &* 0) c ^ 0) -H CL, (i<
. C <0 0) g
Q) 43 4->
0
V4 0 u u 0)
73 U (d
•0 c (d -p CO
<d
ion.
4J 0 Id
nte
• H
i H 0) > Q)
r H
r-i 0
4J • H 43 U 0 (0
X
Q) 0 M
SOU
c • H
Q) -P 0 VH CI4
43
• to
• H to
i H <d c (d
0 ^
(M
T3 0) > 0 g 0) VH
0)
0) :^
eers
+J to
ine
c < 4 - l
0.-
c 0) cu
4-> C 0) s +J (d (U
4->
U Q) a to u Q) 0)
+J to
0
^ 0)
43 g 3 ^
0
• 73 0) (1)
0
H 1 tr ^
c • H (d tp
0
tJN
^ > 1 0 c 0)
•H 0
• H (M (M Q)
C • H
<d 0
TJ
• 0
•H 4J (d
•H (d 0 • •
TJ 0) (U ELI
(U
3A

8.40
8.30
8.20
^ 8 . 1 0
^ 8.00
7.90
7.80
7.70 1 30
Sorbitol, g/d 60
LD Protein » RD Protein
Figure 3.1. Dry matter intake of steers fed either low ruminally degradable protein (LD Protein) mixture or readily ruminally degradable protein (RD Protein) supplement (SEM = .220).
35

2.5 -r
2.3 --
^2.1 I
< 1-9 +
1.7 --
1.5 30 60
Sorbitol, g/d
LD Protein » RD Protein
Figure 3.2. Average daily gain of steers fed either low ruminally degradable protein (LD Protein) mixture or readily ruminally degradable protein (RD Protein) supplement (SEM = .125).
36

275 J
-O 270 --
^ 265 --o 260 +
^
•1 255 +
•S250 +
^^245 +
O 240 --
: : 235 1 30
Sorbitol, g/d 60
LD Protein * RD Protein
Figure 3.3. Gain efficiency of steers fed either low ruminally degradable protein (LD Protein) mixture or readily ruminally degradable protein (RD Protein) supplement (SEM = 11.1).
37

CHAPTER IV
EFFECTS OF SORBITOL ON SODIUM DEPENDENT
AND SODIUM INDEPENDENT GLYCINE AND
LEUCINE UPTAKE BY CULTURED
BOVINE KIDNEY CELLS
Abstract
A series of in vitro experiments were conducted in the
presence or absence of sodium to determine if sorbitol at
concentrations ranging from 10"^ to 10"!^ molar affects
glycine and leucine uptake by cultured bovine kidney cells.
Sorbitol tended (P = .094) to improve glycine uptake by
cultured bovine kidney cells when sodium was present in the
incubation media. However, addition of sorbitol when sodium
was absent improved (P = .0001) glycine uptake under these
experimental conditions.
Concentrations of sorbitol ranging from 10"^ to 10"!^
molar did not alter sodium dependent (P = .36) leucine
uptake by cultured bovine kidney cells. But as was seen
with glycine, the addition of sorbitol in the absence of
sodium enhanced (P = .0001) leucine uptake. Results
indicate that the presence of sorbitol in the range of 10~!2
molar increases sodium independent glycine and leucine
uptake under conditions of this experiment using cultured
bovine kidney cells.
Introduction
Sorbitol, a natural six-carbon polyalcohol, is found in
various foods such as cherries and apples. This compound is
regarded as a safe for human consumption and is classified
as GRAS (generally recognized as safe) by the Food and Drug
Administration.
Sorbitol has been reported to both improve biological
response of young calves (Daniels et al., 1981; Thivend,
38

1982) and finishing steers (Fontenot and Huchette, 1993) and
bulls (Geay et al., 1992) fed a corn silage-based diet, and
to exhibit numerical improvements in steers fed a steam-
flaked grain sorghum-based diet (Boyles and Richardson,
1993). The response in feed efficiency to sorbitol
supplementation has ranged from +12 and +7 % for experiments
with cattle in the finishing stage fed corn silage-based
diets (Fontenot and Huchette, 1993; Geay et al., 1992,
respectively) and only a +4 % numerical improvement when
finishing steers were fed a steam-flaked grain sorghum-based
diet (Boyles and Richardson, 1993). The reported
improvements in feed conversion along with improved feed
efficiency and rate of gain (+18 and +14 %, respectively) by
bulls when fed a low ruminally degradable protein source
with sorbitol (Geay et al., 1992) lead to the hypothesis
that sorbitol could be altering protein metabolism. The
proteinaceous body tissue is dynamic due to the continuous
state of flux in protein synthesis and degradation (Maynard
et al., 1979). An important component in protein metabolism
involves cellular uptake of amino acids; therefore, it
seemed appropriate to target research in this area. There
are separate amino acid transport systems for large (L-
system) and small (A-system and ASC-system) neutral amino
acids as well as for basic and acidic amino acids (Sato et
al., 1991). Therefore, the objectives of this research were
to determine the effect of sorbitol on glycine and leucine
uptake by cultured bovine kidney cells.
Glycine
Glycine is a non-essential amino acid that is readily
synthesized from common metabolic intermediates in all
organisms (Bender, 1985). Glycine and serine are readily
interconverted, thus glycine can be converted to serine for
gluconeogenesis. Since glycine has the simplest structure
39

(H2NCH2CO2H) of any amino acid and is universal to all
organisms, it was the first amino acid of choice used in
this experiment. Glycine seems to be transported by both a
specific glycine carrier and an imino-glycine carrier that
also carries proline and hydroxyproline, as well as being a
substrate for the ASC-system (Bender, 1985). Maximum
glycine transport is dependent upon the presence of
extracellular sodium ions (Wheeler and Christensen, 1967;
Thomas and Christensen, 1971; Thomas et al., 1971; Hundal et
al., 1989).
Leucine
Leucine is a branched chain amino acid that is wholly
ketogenic and has an important role in mammalian metabolism
as an energy yielding substrate (Bender, 1985). Leucine is
also important as a regulator of protein turnover (Taruvigna
et al., 1979). Low levels of leucine have been shown to
stimulate protein synthesis and higher levels have been
shown to inhibit the catabolism of tissue proteins (Bender,
1985). The L-amino acid transport system possesses a
preference for branched, non-polar side chain amino acids
which includes leucine [(CH3)2CHCH2CH(NH2)CO2H] and this
transport system has been reported to be an sodium
independent process (Boerner and Saier, 1982).
Material and Methods
Bovine kidney cells! were purchased and subcultured in
Eagle's medium^ with 10% horse serum^ upon arrival
! ATCC CCL 22 MDBK (NBL), American Type Culture Collection, 12301 Parklawn Drive, Rockville, MD 20852.
2 Basal Medium Eagle, Sigma Chemical Company, P.O. Box 14508, St. Louis, MO 63178.
3 intergen Company, Two Manhattan Road, Purchase, NY 10577.
40

(Appendix A). Cells were incubated in a water-jacketed
incubator at 37 °C under humidified atmosphere of 95% O2
with a constant injection of 5% CO2. Once cells became
confluent, 10 vials containing cells were stored in liquid
nitrogen for future use (Appendix B). Treatment solutions
were applied after cells became confluent in 24-well Costar^
culture cluster flasks. Treatment solutions consisted of
Krebs-Ringer phosphate buffer (KRP) with and without sodium
plus one of the following: unlabeled amino acids (glycine or
leucine) or ([^HJglycine^ or [^H]leucine^) plus sorbitol at
10"4, 10-6, 10-8^ 10"!0, 10"!2, or 10"!^ molar. The final
amino acid concentration was 200 |iM. Composition of the
KRP-buffer was as follows: KCl 6.0 mM, MgS04-7H20 1.2 mM,
KHCO3 2.0 mM, CaCl2 0.5 mM, NaCl 118 mM, D-Glucose 5.5 mM,
Na2HP04 25 mM. Analysis of sodium independent uptake was
facilitated by replacing sodium chloride and sodium hydrogen
phosphate in the KRP-buffer with choline chloride and
choline hydrogen phosphate in equal molarity. Replacement
of sodium chloride with equimolar concentrations of choline
chloride, lithium chloride or potassium chloride has been
reported to result in a reduction of glycine and alanine
uptake by > 90% (Wheeler and Christensen, 1976). This shows
that glycine or alanine transport is not enhanced by the
presence of choline. However, alkali metal ions other than
sodium have been shown to inhibit neutral amino acid
transport while choline had no effect (Thomas and
Christensen, 1971; Thomas et al., 1971). Therefore, the
4 Costar, 7035 Commerce Circle, Pleasanton, CA 94588 8008.
5 [2-3H]Glycine, Amersham Corporation, 2636 South Clearbrook Drive, Arlington Heights, IL 60005.
6 L-[4, 5--^H]Leucine, Amersham Corporation, 2636 South Clearbrook Drive, Arlington Heights, IL 60005.
41

replacement of sodium chloride with choline chloride
provides a method by which the sodium dependence of amino
acid transport can be investigated without interfering with
transport measurements (Hundal et al., 1989).
Treatment solutions were administered following amino
acid depletion according to the methodology of Vadgama
(1989) (Appendix C). The plated cells were washed twice
with warm choline-KRP and depleted of intracellular amino
acids in 1 mL of choline-KRP for one hour at 37 °C in a gas
atmosphere 95% O2 and 5% CO2 before test solutions were
placed in the wells of the culture flasks. Test solutions
(250 jiL) were incubated at 37 °C (humidified atmosphere of
95% ©2 and 5% CO2) while gently shaking culture plates to
avoid unstirred aqueous layers. The uptake incubation was
terminated at exactly two minutes by decanting uptake medium
and immediate addition of one mL of ice-cold phosphate
buffered saline twice for 15 seconds. To facilitate rapid
washing and treatment application, construction of wash
trays and treatment application trays were done according to
procedures of Vadgama (1989) (Appendices E and F).
The intracellular labeled amino acid was extracted
from the cell layer in each well with 220 |iL of 5%
trichloroacetic acid for one hour and 200 |iL of the extract
was counted in 5.0 mL of scintillation fluid in a liquid
scintillation counter. The protein content was determined
using a modification of the methodology of Lowery et al.
(1957) (Appendix D).
Data from 6 experiments for each amino acid were pooled
and analyzed in a randomized block design using the GLM
procedure of SAS (1990) for both the sodium dependent
(sodium present) groups and the sodium independent (sodium
absent) groups. Days in which the 6 experiments were
conducted served as the blocks. The model included blocks
and sorbitol level. Treatment means were separated by the
42

protected least significance difference procedure (Steel and
Torrie, 1980) and all means are reported as least squares
means. Each treatment well (four treatments group"!
experiment"!) was analyzed for protein content after the
radioactivity data was collected in order to normalize the
radioactivity (disintegrations minute"! [DPM]) to mg of
protein contained within each well. The unlabeled amino
acid treatment served as the negative control and its mean
DPM mg"! protein was subtracted from the other five
treatments. Data is reported as DPM per mg of protein.
Results and Discussion
Sorbitol tended (P = .094) to improve glycine uptake by
cultured bovine kidney cells when sodium was present in the
incubation media (Table 4.1 and Figure 4.1). There appeared
to be a positive response in glycine uptake by cultured
bovine kidney cells when sorbitol was present at 10"!2 molar
(S12) concentration.
The addition of sorbitol when sodium was absent
improved (P = .0001) glycine (Table 4.1 and Figure 4.1). In
the absence of sodium, sorbitol at molar concentrations of
10~!2 and 10"!^ (SIO) enhanced glycine uptake by cells
compared to cells not exposed to sorbitol (controls) (P =
.0001 and P = .0013 for S12 and SIO, respectively).
Leucine uptake data are shown in Table 4.2 and Figure
4.2. Cellular uptake of leucine when sorbitol and sodium
were present was not different (P = .363) from cells
incubated without sorbitol. However, in the absence of
sodium, leucine uptake was improved by the presence of
sorbitol at S12 (P = .0001) and sorbitol at 10"!^ molar
(S14; P = .002).
The mechanism by which sorbitol is altering uptake of
these two particular amino acids is unclear. Sorbitol is
known to exert an osmotic effect in biological fluids
43

(Merck, 1989) until it is metabolized. Sorbitol is most
likely exerting some osmotic effect. This may partially
explain the response elicited by sorbitol at concentrations
above 10"!0 molar on glycine and leucine uptake as exhibited
in Figures 4.1 and 4.2. Perhaps the presence of sorbitol in
the range of 10"!2 molar alters the sodium gradient or
membrane potential, thereby modifying glycine and leucine
transport.
In conclusion, data indicate that the presence of
sorbitol increases sodium independent cellular uptake of
glycine and leucine by cultured bovine kidney cells under
these experimental conditions. Further research is needed
to elucidate the effect exhibited by sorbitol concentration
at 10"!2 molar on glycine and leucine uptake.
Implications
The possibility exists based on these data that
sorbitol alters glycine and leucine transport by cultured
bovine kidney cells. Bender (1985) states that mammalian
amino acid transport is assumed to be similar in other
tissues as it is in the renal epithelium. Therefore, it may
be possible that sorbitol increases skeletal muscle uptake
of glycine and leucine in vivo.
44

C • H O > i
rH tJ^
4J C 0)
T3 C (U CI4 (U
c • H
TJ C (d
+> c (U
TJ C 0) 0)
:3 • H •P • o<d to to
a*
43
s CO
c 0 *
0 4J •H
tH Q) U
>i
0) C
^ -o }-l •H 0 ^ to (M 0
4J
Q) C •H > 0
0 43
u (d
iH
o
to c o
• H 4-) (d
4J C 0) o c o o
o 4J • H
u o w
«w >J <M 4 3 U
(1) • X
H (d
. 4J
rH
(d
CM
o
O iH I O H
00 I
o H
V O I
o
I
o
6 0) 4J
ON O
cn CM
g :3
• H
o
^ ^
00 00 00
n CM V O iH
O O
00 CM VO
OS m V O
o O N in
O 4J c Q)
C Q)
0) • H T3 O to
o o o
o V O
0) T3 in O N 00
H C>J CM
00 CJN O N
o O N 00
Q)
CTN 00
0
ON
OS
VO CO CO
4J
c 0)
TJ C 0) CM Q)
T3
c •H
to +) c (U
•H
0) (d & Q)
Q)
X • H to
Q) 43 4J
g 0 u «4-l
(d 4J (d T3
T3 (U iH 0 0
0
u 0 M U Q)
-0 M (d •0 c (d +>
c • H 0) +) o
in o
V
u <4-l <t-l • H
to +> •H U o to
0)
a to +> c 0) M Q) (M (M •H 73
H 43 I 4J ON -H
I
c • H
to c o
•H 4J (d u t7
o
0) g (d to
c •H
to c 0) (d 0)
cu CO
4J
c •H to (M
•H » O (U
rd 43 0 T3
45

0) c
• H O 0)
4J C 0) TJ C 0) (X Q)
T3 C
• H
T3 C (d
+J c <u C 0) cu
e • H TJ O • tO(d
to C rH O r
0) .H O o
+J >1 • H Q) 43 C U TJ O -H to X
<4H 0 )
o c • H
+J >
^ 2 0) 13
(M <tH >i H 43
Q) CM <d
. +> :3
0) rH 43 (d
EH
cu
43 X H (0
^ tH
1
m so n •
CM • ID CM SO
C^ CD 00
O O O •
cn •
^ CD in
9 CM so m
CM
U (0
>H
o E
a c o
•H
«
c 0) o c o u
o -H i3
o (0
00 I o
so I o
I o
E
4J
CM
o in
CM
00
cn
H as in
0) •o in in CM
o n o
CM
SO
o
CM
00
00
CN
CM
CM
•o o
so OS
•o in so
T3
o
OS
E 3
• H T3 O M
U
c 0)
T3 C
s. •o
E •H TJ 0
CO
4J c 0)
•o c
8L 0) •o c
to +) c Q> g
•H
o (d cu Q) X g (U
X •H to g o
<4H
(d +J (d
T3
T3 Q)
r-i o o
j : : 4J
4H o
o u u 0)
T3 M <d
T3 c (d +)
cu CO
c •H
o u
c • H g
in o
V
cu
u Q)
(M (M •H TJ
to 4J a •H
0
to u Q) CU to 4J
c (U »H
0) 0) 4 J <4H
«4H • H
CU 73 H i : I +> DN - H g >
O
g (d to
to c o
•H 4J (d M tr 0)
c • H to <4H
• H » Q 0)
C •H
to c (d 0)
(d A O TJ
46

4 6 8 10 12
[Sorbitol], lO-molar
•- glycine NA -B- glycine WO
Figure 4.1. Effects of sorbitol on glycine uptake by cultured bovine kidney cells in the presence of sodium (glycine NA; SEM =123.4) or in the absence of sodium (glycine WO; SEM = 60.1).
47

14000 X
13000 --
2 12000 +
E 11000 --
10000 --
9000 6 8 10
[Sorbitol], lO-molar 12 14
-•- leucine NA -B- leucine WO
Figure 4.2. Effects of sorbitol on leucine uptake by cultured bovine kidney cells in the presence of sodium (leucine NA; SEM = 625.2) or in the absence of sodium (leucine WO; SEM = 584.3).
48

CHAPTER V
CLEARANCE OF INTRAVENOUSLY
ADMINISTERED SORBITOL
IN STEERS
Abstract
Ten crossbred steers (365 ± 17 kg) were used to
determine the effect of intravenous administration of
sorbitol. Steers were fed a steam-flaked grain sorghum,
cottonseed hull based diet for 14 d prior to receiving an
intravenous infusion. Steers were not offered feed 24 h
prior to receiving intravenous injections in each of two
experimental periods. Steers were randomly allotted to
treatments (two steers per treatment) and were intravenously
infused. Treatment solutions were: physiological saline
(.9% sodium chloride), glucose, sorbitol, propionate or
sucrose in 50% solutions (except saline) at 2.2 g kg"!
metabolic weight. The injection site was the jugular vein.
Serial blood samples were collected at the following times
(T) : 0, 2, 30, 60, 120, 240 and 360 minutes.
Sorbitol, intravenously infused, produced a 62%
increase (P = .008) in plasma glucose 120 minutes post
infusion compared to saline infused steers (122.7 vs. 76.3
mg dL"!) and a 22% numerical improvement (P = .19) over
steers infused with propionate (122.7 vs. 100.8 mg dL"!).
Sorbitol infusion produced a slight increase in plasma
sorbitol concentration to a peak of 2.94 mg dL"! 30 min post
infusion. The increase in plasma sorbitol over (P = .01)
saline infused steers was exhibited at 30 and 60 min post
infusion (.23 vs. 2.94; .35 vs. 2.73 mg dL"! foj- saline and
sorbitol infused steers at 30 and 60 min, respectively). In
summary, intravenously administered sorbitol was rapidly
cleared and elicited an increase in plasma glucose in
steers.
49

Introduction
Sorbitol, a polyhydroxy alcohol, has been shown to
improve performance by bovine animals ranging from young
calves to bulls and steers of market weight (Daniels et al.,
1981; Thivend, 1982; Bauchart et al., 1985; Thivend et al.,
1984; Geay et al., 1992; Fontenot and Huchette, 1993).
Recent work reported by Fostier (1992) showed that
sorbitol when administered in the drinking water to Holstein
cows at a slaughtering facility reduced (P < .05) the
percentage of dark cutting carcasses by 30% when compared to
cows not receiving sorbitol. Fostier (1992) also reported
that three kg of sorbitol per animal supplied in the
drinking water after induced stress increased (P < .03) the
glycogen level contained in biopsies from the longissimus
muscle obtained from young Holstein bulls. Shaw (1974)
postulated that sorbitol follows the same metabolic pathways
in ruminants as in non-ruminants. In non-ruminants the
metabolism of sorbitol depends primarily on a hepatic
enzymatic process. Fructose was shown to be the chief
product of the enzymatic oxidation of sorbitol leading to
glucose (Blakley, 1951; Seeberg et al., 1955).
Sorbitol administered intravenously to dogs (Todd et
al., 1939) and rabbits (Seeberg et al., 1955) has resulted
in the development of only moderate levels of sorbitol in
the blood, but caused a prompt elevation of blood reducing
sugar. Since sorbitol dehydrogenase, a hepatic enzyme, is
commercially available prepared from sheep liver and is used
as a diagnostic tool in liver disorders (Asada and Galambos,
1963; Wiesner et al., 1965), one could postulate that
sorbitol metabolism is similar in ruminants. Therefore, the
objective of this research was to determine the clearance
and possible glucogenic property of sorbitol in steers.
50

Materials and Methods
Ten crossbred steers (3 65 ±17 kg) were fed a steam-
flaked grain sorghum, cottonseed hull based diet (Table 5.1)
for 14 d prior to receiving an intravenous infusion. Steers
were housed in one pen, fed once daily and had ad libitum
access to clean fresh water. Steers were not offered feed
24 h prior to receiving intravenous injections. Steer were
randomly allotted to treatments (two steers per treatment).
Steers were intravenously injected with one of five
treatment solutions at a rate of 2.2 g kg"! of metabolic
weight. Treatment solutions (pH, 7.0) were: physiological
saline (.9 % sodium chloride), glucose!, sorbitol^,
propionate^ (500 mL of propionic acid plus sodium hydroxide
to adjust pH, diluted to 1000 mL) or sucrose^ in 50 %
solutions (except saline). The injection site was the
jugular vein. The time required for the infusion was
approximately five minutes. After a fourteen d adjustment
period, steers were randomly assigned to treatments and the
experiment was repeated.
The procedure for intravenous treatment administration
was as follows: the necks of steers were disinfected with a
chlorhexidine diacetate solution. A 12 gauge needle was
inserted into the jugular vein and the initial blood sample
was obtained. A sterile catheter^ (inside diameter: 1.57
! Dextrose. Fisher, P.O. Box 1307, Houston, TX 77251.
2 Supplied by Rocjuette FRERES, Gurnee, IL.
3 Propionic Acid, Sigma Chemical Company, P.O. Box 14508, St. Louis, MO 63178. pH adjusted with sodiun hydroxide pellets, Fisher, P.O. Box 1307, Houston, TX 77251.
4 Baxter Scientific, P.O. Box 534032, Grand Praire, TX 75053.
5 Clay Adams, Division of Becton Dickinson and Company, parsippany, NJ 07054.
51

mm; outside diameter: 2.08 mm) was inserted through the
needle into the vein. After inserting approximately 38 cm
of the catheter into the vein the needle was removed and the
treatment solution administered. The catheter was then
flushed with heparinized physiological saline and plugged
with a three mL syringe. Sufficient length of the catheter
tubing was glued along the neck (approximately 30 cm) with
livestock identification tag cement^ and taped to the
forehead of each steer.
After the first two blood samples were collected each
steer was removed from the hydraulic squeeze chute and
placed in an individual holding stanchion for 6 h. All
blood samples except the initial sample were collected post
infusion using the catheter. Following each sample
collection, 10 mL of heparinized physiological saline was
flushed through the catheter to prevent blockage. Prior to
each blood collection, the heparinized saline along with
some blood (5 mL) was removed and discarded. Samples were
collected, placed in 15 mL polystyrene centrifuge tubes
containing sodium-fluoride, potassium oxalate mixture (2.5
mg sodium fluoride and 2 mg potassium oxalate per mL of
blood) and immediately placed on ice until plasma
separation, then stored at -80 °C until analyzed.
Plasma was analyzed for glucose^, sorbitol (Bergmeyer
et al., 1974), and fructose (modification of the method of
Anderson et al., 1979, Appendix G).
Data were analyzed using repeated measures analysis of
variance in the General Linear Model procedure of SAS
(1990). There were five treatments with four replications
(steers) per treatment and blood samples were collected
6 Ruscoe, W.J., Ruscoe Company, Akron, OH 44301.
7 Glucose oxidase method, Sigma Diagnostic Kit No. 510, Sigma Chemical Company, P. O. Box 14508, St. Louis, MO 63178.
52

serially seven times from each steer. Treatment means were
separated using the protected least significant difference
procedure (Steel and Torrie, 1980) when there was no
treatment*time interaction. When an interaction existed,
treatment means were separated using the IML procedure so
the proper error mean square could be used for the
denominator of each F statistic. The F statistic for
orthogonal contrasts was calculated using the subplot error
mean square. The standard errors are reported using both
the main and subplot error terms (Steel and Torrie, 1980).
Results and Discussion
Average initial plasma glucose concentrations for
steers in this experiment was 73.4 ± 1.99 mg dL"! (Table
5.2). After intravenous infusion, steers receiving glucose
showed an increase (P < .0001) in plasma glucose
concentration to 378.6 mg dL"! nd remained higher (P <
.0001) than steers receiving other infusions for the initial
120 min. Plasma glucose levels returned to baseline
concentrations by 360 min post infusion which were similar
(P = .35) to saline infused steers. Response surface
analysis of the glucose infusion over time (Figure 5.1)
produced a quadratic effect (P < .05).
As depicted in Figure 5.2 the plasma glucose of steers
infused with both sorbitol and propionate began to show a
numerical increase over baseline values within 3 0 minutes.
By 120 min post infusion the steers infused with sorbitol
showed an increase in plasma glucose to 122.7 mg dL"! which
was similar (P = .08) to steers infused with glucose and
higher (P = .008) than steers infused with saline (122.7 vs
76.3 mg dL"!, for sorbitol and saline infused steers,
respectively). This constituted a 62 % increase in plasma
glucose 120 min after infusion of sorbitol. Meanwhile
steers that received propionate tended (P = .14) to exhibit
53

an increase in glucose concentration over saline infused
steers (100.8 vs. 76.3 mg dL"!, respectively). Therefore,
by 120 min the sorbitol infused animals demonstrated a 62%
increase over the saline infused steers and a 22% numerical
increase over steers infused with propionate.
The implication of this significant glucogenic effect
elicited by intrajugular infusion of sorbitol when compared
to saline infused steers coupled with the numerical increase
over propionate lends credence to the possibility of
sorbitol serving as a glucose precursor in ruminants.
Metabolites known to serve as significant gluconeogenic
precursors in ruminants are: (1) propionate, (2) glycerol,
(3) amino acids, and (4) lactate and pyruvate (Bergman,
1970; Exton, 1972; Krebs, 1964; Weiss and Loffler, 1970).
Bergman (1973) states that other glucogenic precursors could
be valerate, ribose, citrate, along with a variety of
organic compounds that are present in the body. Results of
this experiment indicate that sorbitol can serve as a
glucogenic precursor in ruminants.
Plasma from steers infused with saline and sorbitol
were analyzed for sorbitol and fructose. Plasma sorbitol
values are listed in Table 5.3 and are shown in Figure 5.3.
After sorbitol infusion, an increase (P = .01) in plasma
sorbitol concentration over saline infused steers was
exhibited at 30 and 60 min post infusion (.23 vs. 2.94; .35
vs. 2.73 mg dL"! for saline and sorbitol infused steers at
30 and 60 min, respectively). As a comparison, Rerat et al.
(1993) using swine recently reported peak sorbitol portal
blood concentration of 4.76 mg dL"! ^d arterial blood
concentration of .56 mg dL"!. These concentrations were
reached 135 min postprandial when feeding sorbitol as the
disaccharide, Lycasin.
The slight increase in plasma sorbitol in steers
receiving sorbitol can be explained by the rapid hepatic
54

metabolism of sorbitol. Bye (1969) found that sorbitol was
rapidly removed from the blood of 16 humans during and
following intravenous infusion of sorbitol with only 3% of
administered sorbitol escaping via the kidneys. The liver
is a highly vascular organ. In sheep, the fraction of
cardiac output received by the liver is 30-40%. The liver
contains an enormous capillary bed which functions in the
special effects that the hepatic parenchyma cells have on
substances entering the organ (Smith and Hamlin, 1977). The
amount of cardiac output flowing through the liver and the
enormous vascular system it possesses sheds light on the
rapid clearance of sorbitol by steers found in this study,
especially based on a (juick, one time administration of
sorbitol. Bye (1969) reported an increase in human plasma
sorbitol to 350 mg dL"! after 4 hr infusion of 30% sorbitol
solution administered at .6 g min"!. Therefore, to produce
a large increase in plasma sorbitol, sorbitol needs to be
administered at a constant rate over time in order to exceed
hepatic capacity for clearing this compound. *
Plasma from steers receiving sorbitol and saline were
analyzed for fructose concentration. Because fructose is a
metabolic intermediate in the metabolism of sorbitol to
glucose coupled with its reported effect on amino acid
transport, it was deemed important to ascertain plasma
fructose levels. Plasma fructose values are listed in Table
5.4 and shown in Figure 5.4. The mean baseline fructose
concentration for steers used in this experiment was 35.1 mg
dL"! ± 2.88. These values are considerably higher than
those reported in cows (5 mg dL"!; Rice 1953), ewes (3.3 ±
.3 mg dL"!; schreiner et al., 1981), baboons (.6 ± .09 mg
dL"!; Crossley and Macdonald, 1970) and humans (4.7 ± 1.9 mg
dL"!; Yoshioka et al., 1984). Kurz and Willett (1992) have
reported calf plasma fructose concentrations of 63 mg dL"!
55

at birth which declined to undetectable levels by 14 h after
birth.
Neither glucose nor sorbitol when added to standard
solutions in this analysis affected fructose determination
(data not presented). However, the possibility of other
serum components affecting fructose determination can not be
ruled out. Based on the difference between total reducing
sugar (90 - 110 mg dL"!) and glucose (65 - 70 mg dL"!)
concentrations found in lactating dairy cows (Willett,
1993), the fructose baseline concentration of 35.1 mg dL"! ±
2.88 could be attributing to the difference in total blood
reducing sugar and glucose. Further investigation is needed
to adequately document steer plasma fructose concentrations
in this experiment.
Plasma fructose levels were similar for saline and
sorbitol infused steers until 120 min post infusion (Table
5.4). At 120 min the plasma fructose of saline infused
steers were higher (P = .002) than steers that received
sorbitol (44.6 and 27.7 mg dL"!, respectively). At 240 min
plasma fructose were similar (P = .31, 24.5 and 29.8 mg
dL"!; for saline and sorbitol infused steers, respectively).
Then by 360 min post infusion, steers that received saline
possessed higher (P = .03) plasma fructose levels than those
steers infused with sorbitol (34.2 and 22.7, respectively).
In conclusion, only glucose and sorbitol when
intravenously administered produced statistical increases (P
< .05) in steer plasma glucose. Sorbitol was rapidly
cleared and elicited an increase in plasma glucose.
Therefore, data indicates that sorbitol possesses a
gluconeogenic property in steers.
Implications
Intravenous administration of sorbitol in beef steers
increases plasma glucose concentrations. These data support
56

earlier work of Bye (1969) in humans, Seeberg et al. (1955)
in rabbits and Todd et al. (1939) in dogs and also exhibits
that sorbitol when infused intrajugularly in steers
increases plasma glucose. Results of this experiment
indicate that sorbitol can serve as a glucogenic precursor
in ruminants.
57

Table 5.1. Composition^ and analysis of basal diet.
Item Percent
Steam-flaked grain sorghum 45.00
Cottonseed hulls 46.00
Cottonseed meal 6.00
Urea .10
Molasses, cane 1.00
Calcium carbonate 1.10
Dicalcium phosphate .05
Salt .18
Trace mineral premix^ .20
Vitamin ADE premix^ .37
Nutrient Analysis^
DM, % 86
CP, % 10.01
Ca, % .55
P, % .25
K, % *.83
NEm, Mcal/kg 1.56
NEg, Mcal/kg -75
^ Dry matter basis.
b Contained (ppm) I 1,232, Mn 8,069, Zn 8,409, Cu 827, Co
51, Fe 4,056. c Contained (lU/kg) Vitamin A acetate 634,480, Vitamin D
63,448, Vitamin E 1,813.
d Calculated on a DM basis except for DM.
58

u fH tl)
tJN (d :3 c
't-y O (d -H ^ 0*
4-> c
•H
o u 0*
<d iH o
tJN4-> C - H
•H 43 > U
•H O Q) to O 0) -U Q)
to to O U U 0) u
4-> to to
<d 3 tr 0)
o
43 tr •H 0)
o •H •H O Si <d
0)
g
^1
o to o
<D 0 tJN to 3 0 ^ u 3
f H
CT
V
rg •
fNj
t r (U 4-> 43
<d + J g to <d
•H 0)
•-ixi
<d
to c o •H
D44-> 4 J
c (d 0) S
* OJ
• in
0) rH 43 (d EH
•H ^
C 0
:3 rH
• o c •H
iH (d to
O <4H to
• H d B to O p in (H
c •H c:
•H
0
0) g 3
rH 0 >
•J
tJN
g
O to O U rH
o <d g to fO
iH CU
c O
•H to
c M
4-> to O CU to 0) 4J
c •H
o VO
o <N
O
O VO
O n
CM
<d
c o g (d 0)
VO
n t^
43 ' I '
* H t^
43
• VO
43 ^
• VO t^
43 in
in t^
43 ON
• in r-
^
VO t^
0) c -H
r-{ (d t/3
O
OS CO
0 t^
• OS O rH
T3 VO
.
15
2
73 o
• OS 00 H
0 OS
H n <N
0 VO
. 00 r-cn
00
m r
Q) tu to 0 U 3
rH
o
cn
t^ VO
43 VO
• 00 VO
43
• in
43 00
t
o 00
43 t^
00 r
43 CM
• cn t^
in
VO VO
0) to 0 u 0 3 tn
cn
<JN t^
0 43
• cn ON
T3 0
12
2.
u CM
• ^ H H
43 VO
o O H
43 ^
• r t^
H
cn
((H rH 0 4J •H 43 U 0 Ui
o in 00
0 43 in
• ON CO
0 43 00
10
0.
u 43
VO
10
4.
43 •^
cn o H
43 VO
CO
cn
t^ t^
4J Id c 0
•H a 0 u cu
in o
• CM in
• H H
^ c (d 0) g (U 43 4J
<H 0
u 0 u u 0)
T3 V4 (d •0 c (d 4-> t/3
^ Q) ((H (M •H T3
to +J Q
•H V4 u to M 0) a 3 to
4-> c (U u 0) IM I4H •H T3
x: •H >
c uin
r-{ 0 0
(U g (d to
c •H
to c <d 0) s
TJ ^
0
. ^-^ in o
•
V
cu
4J 0 0)
((H (M 0)
0 •H +J Id u TJ (d P a
•
in o
•
V
cu
4J U 0)
<M <4H 0)
o •H 43 3 U
Id 43 0) «4H
59

g iH o >
C (M
o o C - H (d -P 0) DN rH C O
•H to >
•H cX» Q) O rH o in Id Q) P
>H Id t r
to c >H -H (d 0) 0) rH 4J O to 4-)
•H 4J <W ^ 43 o VJ tr
O -H rH to <D
o > 4-J (H •H O U
•H C r-i O O
to -H ^ to Id
Id 3 -P g SH Q) to c g Id -H
V4 o
43 U O
(d
Id t r rH M
0) tr tr •I-NCM Q) Id * c
. >-( CM -H cn 4J r
. C 4J Id in -H Id to
0) rH 43 Id
tr g
o 4J •H 43 VH O to
Id g to Id fH
cu
to 0)
4-> 3 C
• H
O VO cn
o CM
o CM
t^
o VO
in ON
o cn
CM
Id
c 0) g 4J Id
cn
^ 00 in cn
0)
c • H
CM
CM
43 U in n cn t ^
* . CM
43 0 cn ' t CM ON
CN
c n ON ^ VO
CO cn CM r*
O +J •H 43
f-i U to o
CO CO
u 0)
(M (4H • H
to
•H
u to u (U a 3 to
4J C 0)
v^
VO <4H 00 -H
. 73
Id
g
Q) 43 4J
MH O
o u u 0)
43 4-» • H
g 3
H
o 0
0) g Id to
c • H
T3 U (d
73 c Id +J CO 0
to
(d Q)
Id A
in o
V
4J O 0) (M <4H Q)
o •H 43
V 3 U
CU w no
in o
60

c
C <M 0
• H Id +J
3 DNrH
c • H >
o (U g 3
0 r^ to
• H <^ 0) U <D U
to ^ 0)
O in
Id
c • H
Q) rH 4J 0 to 4->
0 >
rH Id 3
0)
Id
V4 0
•H 4J <W 43 43 o (U to
U 0 to
O <M 4J O 3 U
(M
(d
0
tr •H (U IJ
0 • H
C tH 0 0
•H 43 to Id 3 4J
g <H to Id
tH
C • H
1
0) g
H Cu U 1
c Id (U
s •
•«;r •
in
(U r-\ Xi Id
H
Id rH 3
tr ;^
tr tr 3 •
•»-\CM Q) Id M 4J
• c CM ' H
rH c 4-> Id
• H Id to
•-3
tri g 0) to o
4-> U 3 ^ Cm
Id g to Id rH CU
to
3 c •H
o VO
cn
o CM
O CM H
O VO
O cn
CM
O
Id +J c 0) g
4J Id 0) u tH
0 CM
• ^ cn
in •
CM
u VO
• ^ '^
CM •
^ en
VO •
ON cn
in •
CO cn
en •
ON cn
Xi 0) c
•H rH Id CO
•0 r • CM CM
00 •
ON CM
T3 t^
• t^ CM
cn •
cn cn
^ •
r» cn
cn •
CM m
00 •
o cn
XI tH 0 4J • H 43 U 0 CO
• CO CO VO
• in
^
c Id 0) g 0 4: 4-»
(M 0
u 0 u u <u T3 u Id T3 c Id +J CO
Id
. .«~« in o
•
V
cu ^-'
+J 0 (U
<4H <4H 0)
0 •H 43 3 O
A
V4 0) (M dH •H TJ
to +> CU
•H ^ 0 to u 0) cu 3 to
4J c 0) ^ 0) ((H <M •H 13
43 4J •H ^
C g 3
rH 0 U
0) g Id to
c •H .
to ^ C in Id o Q) £
V T3
- CU 0 ^
61
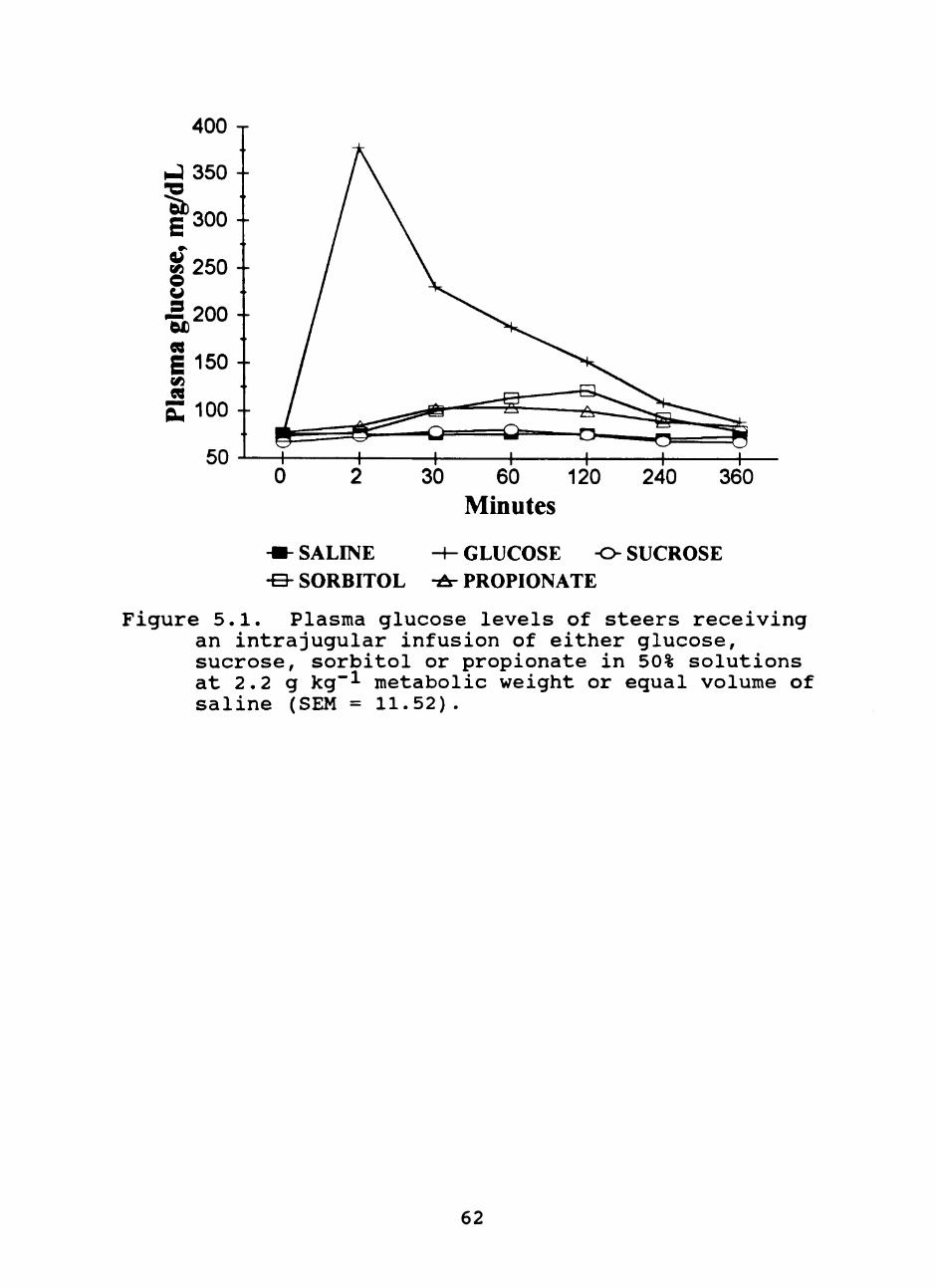
400 T
30 60 120 Minutes
240 360
SALINE SORBITOL
GLUCOSE O SUCROSE PROPIONATE
Figure 5.1. Plasma glucose levels of steers receiving an intrajugular infusion of either glucose, sucrose, sorbitol or propionate in 50% solutions at 2.2 g kg~^ metabolic weight or equal volume of saline (SEM = 11.52).
62

130 T
NJ 120
g110 +
2 100 o Tr 90
e 80
S- 70
60 0 30 60 120
Minutes 240 360
SALINE SORBITOL
oSUCROSE -^ PROPIONATE
Figure 5.2. Plasma glucose levels of steers receiving an intrajugular infusion of either sucrose, sorbitol or propionate in 50% solutions at 2.2 g kg~^ metabolic weight or equal volume of saline (SEM = 11.52). Same data as Figure 5.1 except glucose infusion data are not shown.
63

20 T
"51,15 E
o CA
E CA C9
10 --
5 -
60 120 Minutes
•m- SALINE -& SORBITOL
Figure 5.3. Plasma sorbitol levels of steers receiving an intrajugular infusion of sorbitol in a 50% solution at 2.2 g kg~^ metabolic weight -or equal volume of saline (SEM = .867).
64

50 T
^
E ^ CA
o "^l*
^ 9 u
sma
08
£
45 --.,
40 --< •
35 -
30 --• •
25 -
20 30 60 120
Minutes 240 360
SALINE SORBITOL Figure 5.4. Plasma fructose levels of steers receiving
an intrajugular infusion of sorbitol in a 50% solution at 2.2 g kg~^ metabolic weight or equal volume of saline (SEM = 5.67).
65

CHAPTER VI
INTEGRATED SUMMARY
Sorbitol, a polyhydroxy alcohol generally regarded as
safe (GRAS) by the FDA, has been reported to increase feed
efficiency by veal calves, finishing bulls, and steers. The
mode of action which elicits this effect in ruminants has
not been clearly defined. Therefore, it was the intent of
this research to identify the mechanism of action of
sorbitol in ruminants.
Two feedlot performance studies were conducted to
examine if various levels of sorbitol affected the feedlot
performance of steers fed a steam-flaked grain sorghum-based
diet. The initial feedlot study examined the effect of
supplementing sorbitol over a prolonged feeding period (119
d; n = 112; 337.3 ± 17 kg). Sorbitol was fed at various
levels: basal (no sorbitol), 30 g steer"^ d"^; a variable
rate (20 g first 28 d, 30 g second 28 d; then 40 g steer"^
d~^ the remaining time on feed); and 30 g steer'^ d~^ only
after d 69. Supplementation of sorbitol did not
statistically improve (P > .05) steer feedlot performance
over the 119 d feeding period. However, throughout the
feeding period, supplementing steers with 30 g of sorbitol
d"^ showed a 3.4% numerical increase in ADG and a 4.0%
numerical improvement in fed efficiency over steers
receiving no sorbitol. Steers receiving sorbitol only for
the final 50 d had lower dressing percent (P < .002) than
other steers. Furthermore, an overall treatment effect
tended (P = .16) to alter the carcass lean color. Steers
receiving 30 g of sorbitol d"l appeared to exhibit (P = .03)
a more youthful bright cherry red color of the carcass lean.
A subsequent 28 d feedlot receiving trial was conducted to
determine the response by newly received steers fed either
66

0, 30 or 60 g sorbitol steer"^ d~l in combination with
either a low ruminally degradable protein source or a more
readily ruminally available protein supplement. Performance
of incoming feedlot steers (262 ± 21.7 kg; n = 260) during a
28 day experiment was not improved by feeding a low
ruminally degradable protein or by sorbitol supplementation
at either 30 or 60 grams steer"^ day~l. However, there was
a tendency (P = .18) for an interaction between protein
source and level of sorbitol for gain efficiency.
Sorbitol has been documented to be glucogenic in
humans, rabbits, dogs and rats. Therefore, the possible
glucogenic property in steers was investigated. Sorbitol
intravenously infused in steers produced a 62% increase in
plasma glucose 120 minutes post infusion compared to saline
infused steers and a 22% numerical increase over steers
infused with propionate. Sorbitol was cleared and produced
a slight increase in plasma sorbitol concentration to a peak
of 2.94 mg dL~^ 30 min post infusion.
Reports that sorbitol improved feed efficiency and
daily gain of bulls when fed a low ruminally degradable
protein coupled with the magnitude of improvement shown in
steers fed a corn silage-based diet lead to the hypothesis
that sorbitol could be involved in protein metabolism.
Therefore, glycine and leucine uptake by cultured bovine
kidney cells was studied. Alterations in the sodium
independent uptake of these amino acids were exhibited by
the presence of sorbitol. Data indicate that the presence
of sorbitol at a concentration of 10~^2 molar increases
sodium independent cellular uptake of glycine and leucine by
cultured bovine kidney cells under these experimental
conditions.
In summary, sorbitol did not improve 119 d feedlot
performance of steers fed a steam-flaked grain sorghum-based
diet. However, steers receiving sorbitol only for the final
67

50 d had lower dressing percent (P < .002) than other
steers. Furthermore, steers receiving 30 g of sorbitol d"^
for 119 d appeared to exhibit (P = .03) a more youthful
bright cherry red color of the carcass lean when compared to
steers not fed sorbitol. The significant increase in steer
plasma glucose produced by intravenously infused sorbitol
points to a mode of action of sorbitol in energy
utilization. This may partially explain the improvements in
performance by ruminants when fed lower energy dense high
corn silage-based diets when compared to higher energy dense
diets based on steam-flaked grain sorghum. Sodium
independent uptake of glycine and leucine by cultured bovine
kidney cells was enhanced by sorbitol at a molar
concentration of 10~^2. therefore, sorbitol could possibly
increase skeletal muscle uptake of these two amino acids.
In conclusion, the mode of action of sorbitol includes
energy utilization and a possible role in amino acid
metabolism.
68

LITERATURE CITED
Adcock, L.H., and C. H. Gray/ 1956. Metabolism of sorbitol. Nature 177:329.
Alvarado, F. 1968. Amino acid transport in hamster small intestine; site of inhibition by galactose. Nature 219:276.
Anderson, R. A., J. M. Reddy, C. Oswald and L. J. D. Zaneveld. 1979. Enzymatic determination of fructose in seminal plasma. Clin. Chem. 25(10):1780.
Asada, M. , and J. T. Galambos. 1963. Sorbitol dehydrogenase and hepatocellular injury: An experimental and clinical study. Gastro. 44:578.
Bass, L., and K. Winkler. 1980. A method of determining intrinsic hepatic clearance from the first pass effect. Clin. Exp. Pharmacol. 7:339.
Bauchart, D. , B. Aurousseau, and E. Auclair. 1985. Addition of sorbitol to a milk-substitute for veal calves. I - Effect on health, growth and feed conversion. Reprod. Nutr. Dev. 25:399.
Bender, D. A. 1985. Amino Acid Metabolism. (2nd Ed.). John Wiley and Sons, New York.
Bergman, E. N. 1973. Glucose metabolism in ruminants as related to hypoglycemia and ketosis. Cornell Vet. 63:341.
Bergman, E. N. 1970. Disorders of carbohydrate and fat metabolism. In: Duke's Physiology of Domestic Animals, (Ed.) M. J. Swenson. Cornell Univ. Press, Ithaca, NY.
Bergmeyer, H. U., W. Gruber and I. Gutmann. 1974. D -Sorbitol: In Methods of Enzymatic Analysis. 2 New York.
Blakley, R. L. 1951. The metabolism and antiketogenic effects of sorbitol. Sorbitol dehydrogenase. Biochem. 49:17.
69

Blood, D. C., and J. A. Henderson. 1974. Diseases characterized by nervous system involvement. In: Veterinary Medicine (4th Ed.). D. C. Blood and J. A. Henderson (Eds.). Lea and Febiger, Philadelphia, PA., p. 924.
Boerner, P., and M. H. Saier, Jr. 1982. Growth regulation and amino acid transport in epithelial cells: Influence of culture conditions and transformation on A, ASC, and L transport activities. J. Cell. Physiol. 113:240.
Boyles, D. W., and C. R. Richardson. 1993. Effect of sorbitol on feedlot performance and carcass characteristics of steers fed a steam-flaked grain sorghum based diet. J. Amin. Sci. 71(Suppl. 1):294 (Abstr.).
Bye, P.A. 1969. The utilization and metabolism of intravenous sorbitol. Brit. J. Surg. 56:653.
Crossely, J. N., and I. Macdonald. 1970. The influence in male boboons, of a high sucrose diet on portal and arterial levels of glucose and fructose. Nutr. Metabol. 12:171.
Daniels, L. B., R. L. Peterson, E. L. Piper, and J. M. Rakes. 1981. Sorbitol in diet of young dairy calves. J. Dairy Sci. 64:449.
Eck, T. P., S. J. Bartle, R. L. Preston, R. T. Brandt, Jr., and C. R. Richardson. 1988. Protein source and level for incoming feedlot cattle. J. Anim. Sci. 66:1871.
Edson, N. L. 1936. Ketogenesis-Antiketogenis. IV. Substrate competition in liver. Biochem. J. 30:1862.
Embden, G. , and W. Griesbach. 1914. Z. Physiol. Chem. 91:251.
Exton, J. H. 1972. Gluconeogenesis. Metabolism. 21:945.
Fontenot, J. P., and H. M. Huchette. 1993. Feeding sorbitol alone or in combination with monensin to finishing cattle. J. Anim. Sci. 73:545.
Fostier, B. 1992. Reduction of DFD carcass percentage by supply of sorbitol before slaughtering. 38th ICoMST, Clermont-Ferrand France.
70

Gabbay, K. H. 1973. The sorbitol pathway and the complications of diabetes. N. Eng. J. Med. 288:831.
Geay, Y., E. Richet, S. Ba, and P. Thivend. 1992. Effect of feeding sorbitol associated with different sources and amounts of nitrogen, on growth, digestion and metabolism in young bulls. Anim. Feed Sci. Technol. 36:255.
Gianpaolo, M. 1991. The functioning liver mass. Res. Clin. Lab. 21:9.
Griem, W., and K. Lang. 1960. Klin. Wschr. 38:336
Hundal, H. S., M. J. Rennie, and P. W. Watt. 1989. Characteristics of acidic, basic and neutral amino acid transport in the perfused rat hindlimb. J. Physiol. (London), 408:93.
Johnston C., and H. J. Deul. 1943. Studies on ketosis XXI. The comparative metabolism of the hexitols. J. Biol. Chem. 149:117.
Kaufmann, E. 1929. Klin. Wschr. 8:66.
Krebs, H. A. 1964. Gluconeogenesis. Proc. Royal Soc. 159:545.
Kurz, M. M. and L. B. Willett. 1992. The clearance of carbon-14-fructose, carbon-14-glucose, and carbon-14-sorbitol by calves at birth and 7 days of age. J. Dairy Sci. 75:236.
Lehninger, A. L. 1982. Principles of Biochemistry. Worth Publishers, Inc., New York.
Lowery, C , J. Rosebrough, N. Farr, and R. Randall. 1957. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193:265.
Maynard, L. A., J. K. Loosli, H. F. Hintz, and R .G. Warner. 1979. The proteins and their metabolism. In: Animal Nutrition (7th ed.). McGraw-Hill., Inc. New York.
Merck Index. 1989. Sorbitol. In: The Merck Index (11th Ed.). Merck and Co., Inc., Pathway, NJ, p 1375.
Molino, G., A. Cavanna, P. Avagnina, M. Ballari, and M. Torchio. 1987. Hepatic clearance of D - sorbitol. Noninvasive test for evaluating functional liver plasma. Dig. Dis. Sci. 32:753.
71

Morgan, T. B., and J. Yudkin. 1957. The vitamin-sparing action of sorbitol. Nature 180:543.
NRC. 1984. Nutrient Requirements of Beef Cattle (6th Ed.). National Academy Press, Washington, DC.
Preston, R. L., and S. J. Bartle. 1989. Quantification of rumen escape protein and amino acid needs for new feedlot cattle. Texas Tech Univ. Agri. Sci. Rep. No. T-5-283. p 17.
Reiser, S., and J. Hallfrish. 1987. Metabolic Effects of Dietary Fructose. CRC Press, Inc., Boca Raton, FL.
Reiser, S., and P. A. Christiansen. 1971. The properties of the preferential uptake of L-leucine by isolated intestinal epithelial cells. Biochim. Biophys. Acta. 225:123.
Reiser, S., O. E. Michaelis, IV, and J. Hallfrish. 1975. Effects of sugars on leucine and lysine uptake by intestinal cells from rats fed sucrose and stock diets. Proc. Soc. Exp. Biol. Med. 150:110.
Reiser, S., and J. Hallfrish. 1977. Stimulation of neutral amino acid transport by fructose in epithelial cells isolated from rat intestine. J. Nutr. 107:767.
Reiser, S., and P. A. Christiansen. 1969. Intestinal transport of amino acids as affected by sugars. Am. J. Physiol. 216:915.
Rice, F. A. H. 1953. J. Amer. Chem. Soc. 75:2262.
Richardson, C. R. 1993. Personal communication. Dept. Anim. Sci. and Food Techn. Texas Tech Univ., Lubbock, Texas.
Romans, J. R., K. W. Jones, W. J. Costello, C. W. Carlson, and P. T. Ziegler. 1985. Cattle Slaughter. In: The Meat We Eat (12th Ed.). The Interstate Printers and Publishers, Inc. Danville, IL, p. 139.
SAS. 1990. SAS User's Guide: Statistics (Version 6). SAS Inst. Inc., Cary, NC.
Sato, H., T. Ishii, Y. Sugita, and S. Bannai. 1991. Induction of cationic amino acid transport activity in mouse peritoneal macrophages by lipoplysaccharide. Biochim. Biophys. Acta 1069:46.
72

Schriener, R. L., J. A. Lemons, and E. L. Gresham. 1981. Metabolic and hormonal responses to chronic maternal fasting in the ewe. Ann. Nutr. Metabol. 25:38.
Siimes, A. S. I., and R. K. Creasy. 1980. Maternal and fetal metabolic responses to ritodrine in the sheep. Acta. Obstet. Gynecol. Scand. 59:181.
Seeberg, V. P., E. B. McQuarrie, and C.C. Secor. 1955. Metabolism of intravenously-infused sorbitol. Proc. Soc. Exp. Bio. Med. 89:303.
Shaw, F. D. 1974. Sorbitol dehydrogenase in the diagnosis of liver disease of ruminants. Aust. Vet. J. 50:277.
Smith, C. R., and R. L. Hamlin. 1977. Regional Circulation. In: Duke's Physiology of Domestic Animals, (Ed.) M. J. Swenson. Cornell Univ. Press, Ithaca, NY.
Steel, R. G. D., and J. H. Torrie. 1980. Principles and Procedures of Statistics: A Biometrical Approach (2nd Ed.). McGraw-Hill Book Co., New York.
Stetten, M. R., and D. Stetten. 1951. J. Biol. Chem. 193:157.
Stribling, D. , F. M. Armstrong, and H. E. Harrison. 1989. Aldose reductase in the etiology of diabetic complications: 2. Nephropathy. J. Diabetic Compl. 2:70.
Taruvinga, M. , A. A. Jackson, and M. H. N. Golden. 1979. Comparison of ^^N-labeled glycine, aspartate, valine and leucine for measurement of whole body protein turnover. Clin. Sci. Molecular Med. 57:281.
Thivend, P. 1982. Influence du sorbitol dans 1'alimentation du veau de boucherie. Bull. Tech CRZV Theix, INRA, 50:47.
Thivend, P. M. Debarre, J. Lafaivre, and R. Toullec. 1984. Influence of sorbitol on biliary secretion in the preruminant calf. Can. J. Anim. Sci. 64(Suppl.):102.
Thomas, E. L., and H. N. Christensen. 1971. Nature of the cosubstrate action of Na"*" and neutral amino acids in a transport system. J. Biol. Chem. 246:1682.
73

Thomas, E. L., T. S. Shao, and H. N. Christensen. 1971. Structural selectivity in interaction of neutral amino acids and alkali metal ions with cationic amino acid transport system. J. Biol. Chem. 246:1677.
Todd, W. R., J. Myers, and E. S. West. 1939. On the metabolism of sorbitol and mannitol. J. Biol. Chem. 127:275.
Vadgama, J. V. 1989. Design of simple devices to measure solute fluxes and binding in monolayer cell cultures. Methods in Enzymology. 171:133.
Waldstein, S. S., and R. A. Arcilla. 1958. Measurement of hepatic blood flow by clearance methods. Am. J. Dig. Dis. 3:137.
Weiss, L. and G. Loffler. 1970. Interrelationship between adipose tissue and liver: gluconeogenesis and ketogenesis. Hormone and Metabolic Research. (Suppl. 2):196, NY: Academic Press.
Wiesner, I. S., H. M. Rawnsley, and F. P. Brooks. 1965. Sorbitol dehydrogenase in the diagnosis of liver disease. Am. J. Dig. Dis. 10:147.
Willett, L. B. 1993. Personal communication. Dep. Dairy Sci. Ohio State Univ. Ohio Agri. Res. Devel. Center, Wooster 44691.
Winkler, K., L. Bass, S. Keiding, and N. Tygstrup. 1979. The physiologic basis for clearance measurements in hepatology. Scand. J. Gastroenterol. 14:439.
Winkler, K, S. Keiding, and N. Tygstrup. 1973. Clearance as a quantitative measure of liver function. In: The Liver. Quantitative Aspects of Structure and Function. (Eds.) G. Paumgartner, R. Preisig. Basel, Karger, pp. 144-155.
Wheeler, K. P., and H. N. Christensen. 1967. Role of Na"*" in the transport of amino acids in rabbit red calls. J. Biol. Chem. 242:1450.
Yoshika, S., S. Saitoh, S. Seki, and K. Seki. 1984. Concentrations of non-glucose poloys in serum and cerebrospinal fluid from apparently healthy adults and children. Clin. Chem. 30:188.
74

APPENDIX A
CELL SUBCULTURE TECHNIQUE
1. Remove growth media from culture flasks with pipette.
2. Wash (rinse) twice with phosphate buffered saline (5 mL)
3. Add 2 mL Trypsin-EDTA solution to slightly cover bottom
of dish.
(a) Gently swirl the dish.
(b) Cells will become suspended by gently tapping on
side of dish. Be careful of the incubation time or
the Trypsin-EDTA will damage the cell membranes.
4. Add growth media (10 mL) when cells are off dish to stop
Trypsin-EDTA's action of dispersing cells.
(a) Draw and dispense solution several times.
5. Remove contents (cells and growth media) and place in 50
mL centrifuge tube.
(a) Draw and dispense several times to suspend cells.
6. Add additional growth media (5 mL) to solution in
centrifuge tube to aid in wash.
7) Centrifuge (180 x g) for ten minutes at 4 °C.
8) Pour off media.
9) Resuspend in new growth media.
(a) Remove aliquot (400 |il) to count cells.
10. For subculturing into flasks, add 15 mL growth media to
each flask to allow for 4 to 5 days growth of cells.
(a) To each flask add 2 mL of cell/growth media
solution.
(b) Gently swirl.
(c) Observe cells under microscope. Cells should
appear suspended and floating in solution.
11. Place culture dishes in a water-jacketed incubator at
37 °C in 95% O2 with a constant injection of 5% CO2.
75

12. For subculturing into 24 well Costar flasks plate out enough cell-growth media solution to give 250,000 cells mL~^ well-1.
76

APPENDIX B
CELL FREEZING TECHNIQUE
1. Remove growth media from culture flasks with pipette.
2. Wash (rinse) twice with phosphate buffered saline (5 mL).
3. Add 2 mL Trypsin-EDTA solution to slightly cover bottom
of dish.
(a) Gently swirl the dish.
(b) Cells will become suspended by gently tapping on
side of dish. Be careful of the incubation time or
the Trypsin-EDTA will damage the cell membranes.
4. Add growth media (10 mL) when cells are off dish to stop
Trypsin-EDTA's action of dispersing cells.
(a) Draw and dispense solution two or three times.
5. Remove contents (cells and growth media) and place in 50
mL centrifuge tube.
(a) Draw and dispense several times to suspend cells.
6. Add additional growth media (5 mL) to solution in
centrifuge tube to aid in wash.
7. Centrifuge (180 x g) for ten minutes at 4 °C.
8. Pour off media.
9. Mix new growth media plus 5% DMSO (dimethyl sulfoxide) to
equal 20 mL of solution: 19 mL growth media + 1 mL DMSO = 2 0
mL freezing media. Example: For 1:5 split, use only about
16 mL of freezing media to suspend cells. Use 3 culture
flasks.
10. Add 1 mL per chilled cryo-vial.
11. Place cryo-vials in freezing tank (cryo freezing tank
contains isopropyl alcohol at minus 70 °C, this lowers the
temperature 1 °C per minute.
(a) Freezing tank must be at room temperature.
(b) Place tank with vials in minus 70 °C freezing
container for more than 4 h.
(c) Store vials in liquid nitrogen.
77

APPENDIX C
EXPERIMENTAL PROCEDURE FOR TREATMENT
OF CELLS IN COSTAR CLUSTER WELLS
The incubation procedure is a modification of methodology
described by Vadgama (1989). Cells will be cultured in 24-
well Costar culture clusters. Once cells become confluent
the treatments will be administered. Krebs-Ringer Phosphate
buffer (KRP-buffer), pH 7.40 will be used as uptake medium.
The composition of this buffer will be as follows: KCl 6.0
mM, MgS04-7H20 1.2 mM, KHCO3 2.0 mM, CaCl2 0.5 mM, NaCl 118
mM, D-Glucose 5.5 mM, Na2HP04 25 mM. Analysis of sodium
independent uptake is facilitated by replacing sodium
chlorine and sodium hydrogen phosphate in the KRP-buffer
with choline chloride and choline hydrogen phosphate in
equal molarity.
Cells have been incubated in 24 well Costar cluster
flasks as described in Appendix A. Cells are washed with 1
mL choline Krebs ringer phosphate buffer (choline-KRP).
Before removing dishes from incubator two wash trays should
be filled with 2 mL of choline-KRP in each tube. Use one
tray to apply incubation solution (choline-KRP).
The above described steps will be conducted according
to the following steps:
(a) Pour off media in dishes.
(b) Wash one time with choline-KRP (1 mL).
(c) Incubate one hour at 37 °C with 1 mL choline-KRP.
This removes any free amino acids from cell by diffusion
(concentration gradient).
(d) Incubate with 250 ^L test solution for two minutes
while gently swirling the dishes.
(e) Wash twice with ice cold phosphate buffered saline
(one mL).
78

(f) Allow incubation wells to dry (15-30 minutes).
(g) Extract cells by applying 220 |iL 5 % (w/v)
trichloroacetic acid (TCA) for one hour.
(h) Pipette 200 jiL into scintillation tubes,
(i) Add 5 mL scintillation cocktail to each tube and
vortex.
(j) Count activity using scintillation counter.
In short:
1. Wash cells with one mL choline-KRP.
2. Incubate one hour at 37 °C with one mL choline-KRP.
3. Incubate with 250 |iL test solution for two minutes.
4. Wash twice with ice cold phosphate buffered saline (one
mL) .
5. Extract 22 0 |iL 5% (w/v) TCA for one hour.
6. Pipette 200 \xL and count.
79

APPENDIX D
PROCEDURE FOR PROTEIN DETERMINATION
The procedure used is a modification of the Lowery
method (1957) as described by Vadgama (1989). The 24-well
costar flasks were stored at 4 °C until protein
determination. The following methodology is based upon the
amount of protein contained in each well when the cells were
seeded at 250,000 cells per well and reached confluency in
two d.
1. To each well add 200 nL of 1 N sodium hydroxide.
The protein is completely dissolved in approximately 3 0
minutes. After 30 minutes, draw out 100 |iL of solution and
replace with 100 iiL of the 1 N sodium hydroxide thereby
making a 1:1 dilution.
2. Add one mL of Lowery Reagent to each well and
gently swirl the tray and allow to stand for 20 minutes.
The Lowery Reagent is composed of disodium cupric
ethylenedinitrotetraacetate (25 g/L), Na2C03 (20.0 g/L) ,
NaOH (4.0 g/L). If using primary culture refer to Vadgama
for additional component.
3. After 20 minutes, add 100 |iL of Folin-Ciocalteau
reagent diluted with an equal volume of distilled water.
Immediately swirl the cluster tray gently.
4. After 25-30 minutes, read absorbance at 500 nm. If
protein content is above 100 ng then read absorbance at 750
nm. This solution continues to darken over time, therefore
it is suggested that the addition of reagent solutions be
made systematically and absorbance readings recorded
accordingly.
5. Bovine serum albumin is used as the standard
(blank, 25, 50, 75 and 100 |ig) . This assay is linear up to
12 5 |ig of protein.
80

APPENDIX E
CONSTRUCTION OF EXPERIMENTAL WASH TRAYS
(Vadgama, 1989)
Cells in monolayer cultures can be washed rapidly in
the 24-well cluster using the device described by Vadgama
(1989). To construct this wash tray, take the top cover of
a 24-well Costar flasks (Costar 3424) and simply drill 24
12.5 mm holes through it. Smooth the rough edges around the
holes with a file or sandpaper. Now insert into each hole a
plastic test tube, 12 mm in diameter and 75 mm in length
(T1226-42, Baxter Scientific). The tubes are inserted so
that about 3.5 mm of the open end is exposed over the
surface of the tray, and it is then glued into position with
polystyrene cement. Several wash trays should be
constructed so sequential washes are possible. The bottom
of the 24-well culture flask is fitted over the wash tray
containing wash solution. The whole assembly is inverted
and the contents of the wash tray are rapidly transferred
into each well of the Costar tray.
81

APPENDIX F
CONSTRUCTION OF EXPERIMENTAL UPTAKE TREATMENT TRAYS
(Vadgama, 1989)
The construction of this uptake tray enables the
addition of small volumes of test substrate to the monolayer
cultures. This arrangement ensures quantitative transfer,
on subsequent inversion of the whole uptake assembly, of the
contents of each capsule simultaneously onto the adherent
cells in each of the 24 wells of the Costar flask.
The construction procedure is similar to that of the
wash tray (Appendix D), except instead of the long
polystyrene tubes, polypropylene embedding capsules (beam
size 00, Polysciences, Inc., No. 0224) are inserted into 9
mm holes drilled into another cover for the Costar 24-well
flask. The capsules can be attached with cyanoacrylate
adhesive (or any other suitable adhesive) so that the open
end of each capsule extends about 3 or 4 mm inside.
82

APPENDIX G
FRUCTOSE DETERMINATION IN BOVINE PLASMA
(Modification of the method described
by Anderson et al., 1979)
This procedure for fructose determination is based
upon the enzymatic reduction of fructose to sorbitol by
commercially available enzyme (sorbitol dehydrogenase, EC
1.1.1.14; Sigma Chemical Co., St. Louis, MO). The
concomitant oxidation of NADH to NAD provides a means of
determining fructose concentration using the conventional
two-point kinetic assay at 340 nm.
The following methodology was derived based upon the
method by Anderson et al. (1979).
1. Mix 300 laL of plasma with 300 |iL of distilled water.
2. Place in a boiling water bath for seven minutes.
3. Centrifuge for 30 minutes (could be modified as long as
denatured proteins are removed).
4. Dilute 250 |iL of supernate with 250 |iL of water and
centrifuge for an additional 15 minutes.
5. Add supernatant to reaction mixture.
6. Reaction mixture contains: 100 |iL of Na2P04*7 H2O (1 M,
pH 6.8), 150 |iL sorbitol dehydrogenase (100 ng of
protein/mL; final concentration of 4.5 Units), 300 |iL of
distilled water, and 300 nL of sample.
7. Zero spectrophotometer at 340 nm to this solution before
adding 150 |iL of NADH (32 mM) .
8. After three minutes from adding NADH to reaction mixture,
read absorbance (340 nm), and again at 23 minutes.
9. Compare total change in absorbance at 340 nm with that of
fructose standards.
10. Report data as change in absorbance per minute.
83
Related Documents











