Somatic embryogenesis through pseudo-bulblet transverse thin cell layer of Lilium longiflorum Duong Tan Nhut 1, * , Bui Van Le 2 , Nguyen Tri Minh 1 , Jaime Teixeira de Silva 3 , Seiichi Fukai 4 , Michio Tanaka 4 and K. Tran Thanh Van 5 1 Institute of Biology in Dalat, 116 Xo Viet Nghe Tinh, Dalat, Lam Dong, Vietnam; 2 Department of Biotechnology, Faculty of Biology, College of Natural Science, Vietnam National University-Ho Chi Minh City, 227 Nguyen Van Cu, Distrist 5, Ho Chi Minh City, Vietnam; 3 Departamento de Biologia Vegetal, Faculdade de Ciências de Lisboa, C-2 Campo Grande, 1780 Lisboa, Portugal; 4 Faculty of Agriculture, Kagawa University, 761-0795 Miki-cho, Kagawa, Japan; 5 Institut de Biotechnologie des Plantes, Universitê de Paris-Sud, UMR 0569, F-91405 Orsay Cedex, France; *Author for correspondence (e-mail: [email protected]; phone: 84-63-831056; fax: 84-63-831028) Received 6 July 2001; accepted in revised form 1 November 2001 Key words: Globular embryo, Lilium longiflorum, Pseudo-bulblet, Regeneration, Sucrose, Thin cell layer Abstract Somatic embryogenesis was achieved directly from pseudo-bulblet transverse thin cell layers (tTCL) of Lilium longiflorum. Embryo-like structures (globular embryos) were obtained from different size explants of pseudo- bulblet tTCLs after 45 days culture on Murashige and Skoog, 1962 (MS) medium containing 5.4 M naphtha- lene acetic acid (NAA) and 1.1 M thidiazuron (TDZ). The embryo-like structures were then isolated and mass proliferated on MS medium, containing 5.4 M NAA and 0.4 M TDZ, every 45 days. A 0.8–1.0 mm thick explant was shown to be optimal for obtaining the highest number of embryo-like structures. For plant regen- eration these structures were transferred to hormone-free MS medium with 30 g/l sucrose. All of these structures formed plantlets after 90 days culture. Abbreviations: ELS – Embryo like structure, MS – Murashige and Skoog (1962), NAA – Naphthalene-acetic acid, SEM – Scanning Electron Microscopy, TDZ – Thidiazuron, tTCL – Transverse Thin Cell Layer Introduction Lilies are one of the most important bulbous crops and are used throughout the world in the flower in- dustry as cut flowers and potted plants. The success of breeding programs of Lilium species depends on having a procedure for producing plants rapidly. For this purpose an in vitro technique is frequently used. Such in vitro procedures are used increasingly for in vitro selection, protoplast fusion or genetic transfor- mation, as well as for regenerating plants by somatic embryogenesis. Thin sections made transversely from either pseu- do-bulblets, young stems, stem segments or recepta- cles have also been used for manipulation of the mor- phogenetic pathways and bud regeneration in L. longiflorum (Bui et al. 1999; Nhut 1998; Nhut et al. (2001, 2001, 2001)). Recently, somatic embryogenesis was shown to occur from bulb scales in some lily hybrids (Haensch 1996), and in L. longiflorum by using styles and flower pedicels (Tribulato et al. 1997), but in these procedures somatic embryogenesis was induced via callus culture. In this research, however, somatic embryogenesis was achieved directly from pseudo-bulblet thin cell layers of L. longiflorum. 193 Plant Growth Regulation 37: 193–198, 2002. © 2002 Kluwer Academic Publishers. Printed in the Netherlands.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Somatic embryogenesis through pseudo-bulblet transverse thin cell layerof Lilium longiflorum
Duong Tan Nhut1,*, Bui Van Le2, Nguyen Tri Minh1, Jaime Teixeira de Silva3, SeiichiFukai4, Michio Tanaka4 and K. Tran Thanh Van5
1Institute of Biology in Dalat, 116 Xo Viet Nghe Tinh, Dalat, Lam Dong, Vietnam; 2Department ofBiotechnology, Faculty of Biology, College of Natural Science, Vietnam National University-Ho Chi MinhCity, 227 Nguyen Van Cu, Distrist 5, Ho Chi Minh City, Vietnam; 3Departamento de Biologia Vegetal,Faculdade de Ciências de Lisboa, C-2 Campo Grande, 1780 Lisboa, Portugal; 4Faculty of Agriculture,Kagawa University, 761-0795 Miki-cho, Kagawa, Japan; 5Institut de Biotechnologie des Plantes, Universitêde Paris-Sud, UMR 0569, F-91405 Orsay Cedex, France; *Author for correspondence (e-mail:[email protected]; phone: 84-63-831056; fax: 84-63-831028)
Received 6 July 2001; accepted in revised form 1 November 2001
Key words: Globular embryo, Lilium longiflorum, Pseudo-bulblet, Regeneration, Sucrose, Thin cell layer
Abstract
Somatic embryogenesis was achieved directly from pseudo-bulblet transverse thin cell layers (tTCL) of Liliumlongiflorum. Embryo-like structures (globular embryos) were obtained from different size explants of pseudo-bulblet tTCLs after 45 days culture on Murashige and Skoog, 1962 (MS) medium containing 5.4 �M naphtha-lene acetic acid (NAA) and 1.1 �M thidiazuron (TDZ). The embryo-like structures were then isolated and massproliferated on MS medium, containing 5.4 �M NAA and 0.4 �M TDZ, every 45 days. A 0.8–1.0 mm thickexplant was shown to be optimal for obtaining the highest number of embryo-like structures. For plant regen-eration these structures were transferred to hormone-free MS medium with 30 g/l sucrose. All of these structuresformed plantlets after 90 days culture.
Abbreviations: ELS – Embryo like structure, MS – Murashige and Skoog (1962), NAA – � Naphthalene-aceticacid, SEM – Scanning Electron Microscopy, TDZ – Thidiazuron, tTCL – Transverse Thin Cell Layer
Introduction
Lilies are one of the most important bulbous cropsand are used throughout the world in the flower in-dustry as cut flowers and potted plants. The successof breeding programs of Lilium species depends onhaving a procedure for producing plants rapidly. Forthis purpose an in vitro technique is frequently used.Such in vitro procedures are used increasingly for invitro selection, protoplast fusion or genetic transfor-mation, as well as for regenerating plants by somaticembryogenesis.
Thin sections made transversely from either pseu-do-bulblets, young stems, stem segments or recepta-cles have also been used for manipulation of the mor-
phogenetic pathways and bud regeneration in L.longiflorum (Bui et al. 1999; Nhut 1998; Nhut et al.(2001, 2001, 2001)).
Recently, somatic embryogenesis was shown tooccur from bulb scales in some lily hybrids (Haensch1996), and in L. longiflorum by using styles andflower pedicels (Tribulato et al. 1997), but in theseprocedures somatic embryogenesis was induced viacallus culture.
In this research, however, somatic embryogenesiswas achieved directly from pseudo-bulblet thin celllayers of L. longiflorum.
193Plant Growth Regulation 37: 193–198, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.

Materials and methods
Plant materials
In vitro pseudo-bulblets of L. longiflorum were ob-tained from stem nodes as reported by Nhut (1998).Four-week-old pseudo-bulblets were used as an ex-plant source for the experiments.
Thin cell layer explant culture and induction ofembryo-like structures
In order to induce embryo-like structures (ELS), dif-ferent sized pseudo-bulblets (0.5–0.7, 0.8–1.0, 1.2–1.5 mm thick) transverse thin cell layers (tTCLs)were used as explants. These were excised under astereo microscope in a laminar flow cabinet and cul-tured on MS medium (Murashige and Skoog 1962)containing 30 g/l sucrose, 0.8% agar (Difco) and sup-plemented with NAA (5.4 �M) and TDZ (0.4, 1.1 or2.2 �M).
Embryo-like structure proliferation
To maintain the ELS type, ELS clusters were cut intosmall clusters and subcultured on MS medium con-taining NAA (5.4 �M) and TDZ (0.2, 0.4, 0.7, 0.9 or1.1 �M).
Plant regeneration
All ELS clusters were cultured on hormone-free MSmedium containing different sucrose concentrations(0, 10, 20, 30 or 40 g/l).
Statistical analysis
Each treatment was repeated 3 times and data wererecorded at 45 or 90 days after culture. Explants inexperiments were arranged in a randomized completeblock design with 10 explants per treatment and threeblocks. Data were analyzed for significance by analy-sis of variance with the mean separation by Duncan’smultiple range test (Duncan 1995).
Scanning electron microscopy (SEM)
Specimens for scanning electron microscope (SEM)observations were fixed with FAA (formalin: aceticacid: 70% ethanol, 5: 5: 90), then dehydrated in anethanol-acetone series and critical point dried. After
coating with platinum, specimens were observed withSEM (Hitachi, S-2150, Japan).
Culture conditions
In all three experiments 100 ml media was dispensedinto 500 ml bottles and capped with a plastic cap.Medium was adjusted to pH 5.8 before autoclaving at121 °C at 105 Kpa for 15 min. All cultures were in-cubated at 25 ± 1 °C with a photoperiod of 16 h perday at a light intensity of 40 �mol/m/s fluorescentlight.
Results
Effects of tTCL explant thickness on somatic embryoinduction
The highest survival rate (100%) of tTCL explantswas obtained with a 0.8–1.0 or 1.2–1.5 mm thickness(Table 1). Pseudo-bulblet tTCL thickness has an ef-fect on the induction of ELS when pseudo-bulblettTCLs are cultured on the medium with all NAA andTDZ combinations (Table 1). MS medium containing5.4 �M NAA and 1.1 �M TDZ showed the highestsomatic embryo induction as compared to the othercombinations of either 5.4 �M NAA and 0.4 �MTDZ or 5.4 �M NAA and 2.2 �M TDZ (Table 1).The number of somatic embryos per tTCL explantwas higher with a tTCL thickness of 0.8–1.0 mm (2.7embryos) than with other thickness (1.9 or 2.1 em-bryos with 0.5–0.7 mm or 1.2–1.5 mm tTCL thick-ness) (Table 1). Callus with somatic embryo forma-tion and the number of embryos per tTCL explant of0.5–0.7 mm and 1.2–1.5 mm thick explants were al-most equal (Table 1). The 0.8–1.0 mm thick tTCL ex-plants gave the highest number of ELSs when com-pared to the 0.5–0.7 mm and 1.2–1.5 mm thick. TheELS can be easily identified from surrounding cal-luses by its more intense yellow colour (Figure 1B).
Effect of NAA and TDZ ratio on proliferatingsomatic embryogenic masses
Subculturing of an ELS gave rise to new ELS on MSmedium containing 5.4 �M NAA and TDZ varyingfrom 0.2, 0.4, 0.7, 0.9 or 1.1 �M (Table 2, Figure 1C).The ELS cluster weight was higher at either 5.4 �MNAA and �M 0.4 TDZ or 5.4 �M NAA and 0.7 �MTDZ after 90 days of culture, when compared to other
194
combinations (Table 2). The ELS cluster weight waslowest at 5.4 �M NAA and 1.1 �M TDZ.
Effect of sucrose concentration on plantregeneration
Transfer of ELS to hormone-free MS medium re-sulted in embryo development. Whole plantletsformed on hormone-free MS medium after 3 monthsof culture. The number of plantlets (31) per ELS clus-ter was highest at 30 mg/l sucrose, as compared tothat at 20 mg/l or 40 mg/l sucrose, and the plantletswere very uniform (Table 3, Figure 1G). The ELSclusters did not form any plantlets at 0 or 10 mg/l su-crose and became necrotic after 90 days culture (Ta-ble 3).
Morphogenetic response
ELSs were observed to initiate from the surface ofpseudo-bulblet tTCL explants after 90 days of incu-bation on MS medium containing 5.4 �M NAA and1.1 �M TDZ with a 0.8–1.0 mm thickness. ELSswere initiated directly from pseudo-bulblet tTCL ex-plants. A more intense yellow colour was easy toclearly identify and separate among surrounding cal-luses (Figure 1B). SEM allowed clear visualization ofindividual ELSs (Figure 1D). This is a close-up of asingle ELS (Figure 1E).
SEM clearly showed shoot primordium formationon hormone-free MS medium with 30 g/l sucrose(Figure 1F) as well as root formation from ELS. Thesurrounding calluses of embryo-like structures did not
regenerate into whole plants, while very rarely wasthere shoot formation without rooting after 3 months(Figure 1H). SEM also confirmed that embryos de-veloped directly from the surface of tTCL explants(Figure 1D) and that no intermediate callus wasformed.
Discussion
Recently, somatic embryogenesis was shown to occurfrom bulb scales in some lily hybrids (Haensch 1996).However, there is no report describing this regenera-tion process from undifferentiated tissues of lily spe-cies and hybrids. Somatic embryogenesis is the mostpromising technique for plant multiplication, becauseof its high proliferation potential and the ability ofminimizing or eliminating the risk of chimeric plantdevelopment.
Somatic embryogenensis was reported to occur inL. longiflorum when dicamba and picloram was used(Tribulato et al. 1997), but these somatic embryoswere derived from calluses by style and pedicel cul-ture. On the other hand, Saiju et al. (1994) mentionedthe regeneration of embryoids in lilies, but they usedthem only for later multiplication of shoots with sub-sequent rooting of the developed shoots (Saiju et al.1994). Haensch (1996) reported the induction of so-matic embryogenesis on bulb scales of Lilium hybridsand the regeneration of the whole plants (Haensch1996). The experimental design was developed uponthe basis of the results on Gasteria and Haworthia(also Liliaceae), where somatic embryogenesis was
Table 1. Effect of L. longiflorum pseudo-bulblet explant thickness on the induction of embryo-like structures after 45 days in culture
Explant thickness
(mm)
Plant hormone (�M) Survival rate (%) Callus with somatic
embryo formation
(%)
Number of ELSs per
explanta
NAA TDZ
0.5–0.7 5.4 0.4 70 40 1.3c
5.4 1.1 75 60 1.9b
5.4 2.2 70 10 0.5f
0.8–1.0 5.4 0.4 100 60 1.5c
5.4 1.1 100 85 2.7a
5.4 2.2 100 15 1.1d
1.2–1.5 5.4 0.4 100 40 1.0d
5.4 1.1 100 65 2.1b
5.4 2.2 100 15 0.8e
a Different letters within a column indicate significant differences at P = 0.05 by Duncan’s multiple range test
195
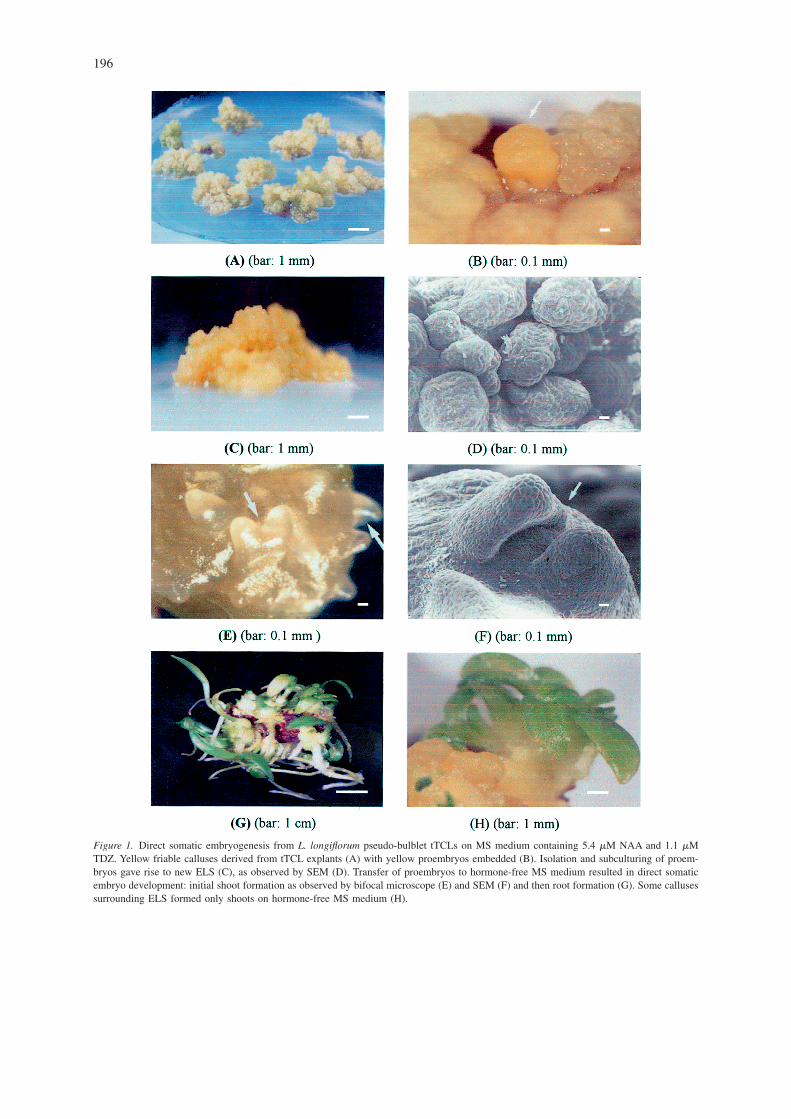
Figure 1. Direct somatic embryogenesis from L. longiflorum pseudo-bulblet tTCLs on MS medium containing 5.4 �M NAA and 1.1 �MTDZ. Yellow friable calluses derived from tTCL explants (A) with yellow proembryos embedded (B). Isolation and subculturing of proem-bryos gave rise to new ELS (C), as observed by SEM (D). Transfer of proembryos to hormone-free MS medium resulted in direct somaticembryo development: initial shoot formation as observed by bifocal microscope (E) and SEM (F) and then root formation (G). Some callusessurrounding ELS formed only shoots on hormone-free MS medium (H).
196

successfully induced in leaf fragments on media byusing 2,4-dichlorophenoxyacetic acid (2,4-D) or4-amino-3,5,6-tricloro-picolinic acid (picloram) with-out subculturing on the growth regulator-free media(Beyl and Sharma 1983).
Most calluses formed were compact (type I) andoccurred from pseudo-bulblet layers on MS mediumcontaining NAA (data not shown). Only the presenceof TDZ induced friable calluses (type II) and somaticembryos. Friable calluses have been previously re-ported in L. longiflorum (Loffler et al. 1990) and L.japonicum Thunb. (Mizuguchi et al. 1994) on NAA-and BA-containing media. Famaelaer et al. (1996)demonstrated dicamba to be toxic for bulb scales in aconcentration ranging from 5–50 �M (Famaelaer etal. 1996). In the present study, the presence of TDZpromoted ELS formation at all concentrations in com-bination with NAA, clearly visualized by SEM.
The combination of NAA with TDZ or BA inducedcalluses and bulblets, calluses and somatic embryos,or calluses with bulblets and somatic embryo forma-tion (Nhut et al. 2001). Our previous study showedthat the combination of NAA and TDZ induced cal-luses with somatic embryogenesis (Nhut et al. 2001).Our results are in agreement with Nhut et al. (2001),who found that tTCLs of L. longflorum young stem,
cultured on MS agar medium supplemented withNAA in combination with TDZ, induced somatic em-bryogenesis (Nhut et al. 2001). However, the percent-age of pseudo-bulblet tTCL explants forming visiblesomatic embryogensis was higher (85%) as comparedto that of young stem tTCLs (45%) (Nhut et al. 2001).Earlier reports showed that friable calluses can beobtained from flower pedicels and styles of L. longi-florum ‘Snow Queen’ or the Oriential lily hybrid ‘StarGazer’, on media containing either dicamba or piclo-ram at a low concentration. When cell suspensioncultures were initiated from this callus type and se-lected during subculturing for small dense embryo-genic cells, ELS were formed (Tribulato et al. 1997).Our results showed that NAA and TDZ stimulated theinduction of callus on pseudo-bulblet tTCLs and thedirect formation of somatic embryos.
The in vitro direct somatic embryo formation of L.longiflorum through tTCLs of pseudo-bulblets has notpreviously been reported. In this study, by using pseu-do-bulblet tTCLs (0.8–1.0 mm thick) and NAA incombination with TDZ, a simple and highly effectivemethod for successful somatic embryo formation wasachieved.
In conclusion, ELSs can be obtained from L. longi-florum pseudo-bulbet tTCLs on media containing 5.4�M NAA and 1.1 �M TDZ. These ELSs germinatedon MS hormone-free medium with both a root and ashoot, and we conclude that they are in fact somaticembryos. These somatic embryos may be used for themass propagation of lily and are a valuable target tis-sue for genetic transformation of this ornamental spe-cies.
Acknowledgements
The authors wish to thank DL Oluf L. Oamborg forcritical reading of the manuscript and many valuablesuggestions.
References
Beyl C.A. and Sharma G.C. 1983. Picloram induced somatic em-bryogenensis in Gasteria and Haworthia. Plant Cell Tiss. Org.Cult. 2: 123–132.
Bui V.L., Nhut D.T. and Tran Thanh Van K. 1999. Plant produc-tion via shoot regeneration from thin cell layer pseudo-bulbletexplants of Lilium longiflorum in vitro. C. R. Acad. Sci. Paris322: 303–310.
Table 2. Effect of NAA and TDZ on proliferation of L. longiflo-rum ELS after 45 days in culture
Plant hormone (�M) Fresh weight (mg) of ELS clustersa
NAA TDZ
5.4 0.2 1989.2c
5.4 0.4 2401.1a
5.4 0.7 2420.4a
5.4 0.9 2039.3b
5.4 1.1 204.5d
a Different letters within a column indicate significant differencesat P = 0.05 by Duncan’s multiple range test
Table 3. Effect of sucrose concentration on L. longiflorum plantletregeneration after 90 days in culture
Sucrose concentration (g/l) Number of plantlets/ELS clustersa
0 0
0 10
20 20.2b
30 31.3a
40 14.2c
a Different letters within a column indicate significant differencesat P = 0.05 by Duncan’s multiple range test
197

Duncan D.B. 1995. Multiple range and multiple F test. Biometrics11: 1–42.
Famaelaer I., Ennik E., Van Tuyl J.M., Meijer H. and Creemers-Molenaar C. 1996. The establishment of suspension and mer-istem cultures for the development of a protoplasts and fusionin lily. Acta. Hort. 414: 161–168.
Haensch K.T. 1996. Plant regeneration through somatic embryo-genesis in different genotypes of Lilium hybrids. Gartenbauw61: 214–218.
Loffler H.J.M., Mouris J.R. and Van Harmelen M.J. 1990. In vitroselection for resistance against Fusarium oxysporum in lily:prospects. Lily Yearb. N. Am. Lily Soc. :
Mizuguchi S., Ohkawa M. and Ikekawa F. 1994. Effect of naph-thaleneacetic acid and benzyladenine on the growth of whitecallus and formation of bulblets from callus induced frommother-scale of Lilium japonicum Thunb. J. Soc. Hort. Sci. 63:131–137.
Murashige T. and Skoog F. 1962. A revised medium for a rapidgrowth and bioassay with tobacco tissue culture. Physiol. Plant15: 473–479.
Nhut D.T. 1998. Micropropagation of lily (Lilium longiflorum) viain vitro stem node and pseudo-bulblet culture. Plant Cell Rep.17: 913–916.
Nhut D.T., Bui V.L., Tanaka M. and Tran Thanh Van K. 2001.Shoot induction and plant regeneration from receptacle tissueof Lilium longiflorum. Sci. Hort. 87: 131–138.
Nhut D.T., Bui V.L. and Tran Thanh Van K. 2001. Manipulation ofthe morphogenetic pathways of Lilium longiflorum transversethin cell layer explants by auxin and cytokinin. In Vitro CellBio. Dev. 37: 44–49.
Nhut D.T., Bui V.L., Fukai S., Tanaka M. and Tran Thanh Van K.2001. Effects of activated charcoal, explant size, explant posi-tion and sucrose concentration on plant and shoot regenerationof Lilium longiflorum via young stem culture. Plant GrowthReg. 33: 59–65.
Saiju H.K., Rajbhandary S.B. and Malla S.B. 1994. In vivo rootingof in vitro produced microshoots of Lilium longiflorum. Ab-stracts of VIIIth Intern. Congress of Plant Tissue and Cell cul-ture, Firenze, June 12-17, Abstract S1-125.
Tribulato A., Remotti P.C., Loffler H.J.M. and Van Tuyl J.M. 1997.Somatic embryogenesis and plant regeneration in Lilium longi-florum Thumb. Plant Cell Rep. 17: 113–118.
198
Related Documents











