ORIGINAL PAPER Soil carbon mineralisation responses to alterations of microbial diversity and soil structure Sabrina Juarez & Naoise Nunan & Anne-Claire Duday & Valérie Pouteau & Claire Chenu Received: 27 November 2012 / Revised: 11 February 2013 / Accepted: 14 February 2013 # Springer-Verlag Berlin Heidelberg 2013 Abstract Soil organisms are of fundamental importance for many soil functions, such as organic matter decomposition, nutrient cycling and energy flow. Most research suggests that soil microbial communities are functionally redundant, mean- ing that there is little relationship between microbial diversity and soil functions. However, the activity of biological com- munities is known to be affected by their physical environ- ment. Here, the effects of changes in microbial diversity and soil structure on organic C (OC) mineralisation were investi- gated. Sterile soil samples that had been subjected to different physical perturbations were inoculated with microbial com- munities with different levels of diversity. The samples were incubated for a period of 127 days and the mineralisation of native and added ( 13 C-labelled substrates, fructose and vanil- lin) OC was measured. It was hypothesised that the magnitude of the effect of changes in soil structure on OC mineralisation would increase as diversity decreased. The diversity treatment had a small but significant effect on the mineralisation of SOC and of the added substrates. The soil structure treatment had a significant effect only on the mineralisation of the added substrate C. There was no interaction between diversity and soil structure treatments, indicating that the relationship be- tween diversity and OC decomposition was not dependent on the soil physical environment. Keywords Soil microbial diversity . Soil structure . Soil organic carbon mineralisation . Carbon dynamics Introduction Global biodiversity is declining worldwide, mainly because of human-induced disturbances in the form of land use and climate change (Chapin et al. 2000). As a result, an interest in the relationship between biodiversity and ecosystem functioning has emerged in ecological (Loreau et al. 2003). Numerous studies on macro-organisms have demonstrated that the biotic composition of communities within an eco- system can significantly affect ecosystem process rates, such as primary production, N cycling or decomposition (e.g., Hector et al. 1999; Cardinale et al. 2006; Tilman et al. 2012). Although the relationship between diversity and functioning has only recently become a topic of interest in soil microbial ecology, a number of studies have investigated the relationship between the diversity of microbial commu- nities and many soil functions such as OM decomposition (Griffiths et al. 2000; Wertz et al. 2006), nitrification (Griffiths et al. 2000; Wertz et al. 2006), potential denitrifi- cation (Attard et al. 2011), resistance to disturbance or resilience after disturbance (Griffiths et al. 2001; Wertz et al. 2007), soil porosity (Martin et al. 2012) and also plant productivity (Chaparro et al. 2012). In contrast to what is seen for higher organisms, little or no consistent relationship has been found between soil microbial diversity and many soil functions, leading to the conclusion that soil microbial communities are characterised by high functional redundancy with respect to many functions (Nannipieri et al. 2003; Nielsen et al. 2011). Electronic supplementary material The online version of this article (doi:10.1007/s00374-013-0784-8) contains supplementary material, which is available to authorized users. S. Juarez : C. Chenu AgroParisTech, UMR 7618 BioEMCo, Bâtiment EGER, 78850 Thiverval-Grignon, France N. Nunan (*) : A.-C. Duday CNRS, UMR 7618 Bioemco, Bâtiment EGER, 78850 Thiverval Grignon, France e-mail: [email protected] V. Pouteau INRA, UMR 7618 Bioemco, Bâtiment EGER, 78850 Thiverval Grignon, France Biol Fertil Soils DOI 10.1007/s00374-013-0784-8

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
Soil carbon mineralisation responses to alterationsof microbial diversity and soil structure
Sabrina Juarez & Naoise Nunan & Anne-Claire Duday &
Valérie Pouteau & Claire Chenu
Received: 27 November 2012 /Revised: 11 February 2013 /Accepted: 14 February 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Soil organisms are of fundamental importance formany soil functions, such as organic matter decomposition,nutrient cycling and energy flow. Most research suggests thatsoil microbial communities are functionally redundant, mean-ing that there is little relationship between microbial diversityand soil functions. However, the activity of biological com-munities is known to be affected by their physical environ-ment. Here, the effects of changes in microbial diversity andsoil structure on organic C (OC) mineralisation were investi-gated. Sterile soil samples that had been subjected to differentphysical perturbations were inoculated with microbial com-munities with different levels of diversity. The samples wereincubated for a period of 127 days and the mineralisation ofnative and added (13C-labelled substrates, fructose and vanil-lin) OCwas measured. It was hypothesised that the magnitudeof the effect of changes in soil structure on OC mineralisationwould increase as diversity decreased. The diversity treatmenthad a small but significant effect on the mineralisation of SOCand of the added substrates. The soil structure treatment had asignificant effect only on the mineralisation of the addedsubstrate C. There was no interaction between diversity and
soil structure treatments, indicating that the relationship be-tween diversity and OC decomposition was not dependent onthe soil physical environment.
Keywords Soil microbial diversity . Soil structure . Soilorganic carbon mineralisation . Carbon dynamics
Introduction
Global biodiversity is declining worldwide, mainly becauseof human-induced disturbances in the form of land use andclimate change (Chapin et al. 2000). As a result, an interestin the relationship between biodiversity and ecosystemfunctioning has emerged in ecological (Loreau et al. 2003).Numerous studies on macro-organisms have demonstratedthat the biotic composition of communities within an eco-system can significantly affect ecosystem process rates, suchas primary production, N cycling or decomposition (e.g.,Hector et al. 1999; Cardinale et al. 2006; Tilman et al. 2012).Although the relationship between diversity and functioninghas only recently become a topic of interest in soilmicrobial ecology, a number of studies have investigatedthe relationship between the diversity of microbial commu-nities and many soil functions such as OM decomposition(Griffiths et al. 2000; Wertz et al. 2006), nitrification(Griffiths et al. 2000; Wertz et al. 2006), potential denitrifi-cation (Attard et al. 2011), resistance to disturbance orresilience after disturbance (Griffiths et al. 2001; Wertz etal. 2007), soil porosity (Martin et al. 2012) and also plantproductivity (Chaparro et al. 2012). In contrast to what isseen for higher organisms, little or no consistent relationshiphas been found between soil microbial diversity and manysoil functions, leading to the conclusion that soil microbialcommunities are characterised by high functional redundancywith respect to many functions (Nannipieri et al. 2003;Nielsen et al. 2011).
Electronic supplementary material The online version of this article(doi:10.1007/s00374-013-0784-8) contains supplementary material,which is available to authorized users.
S. Juarez : C. ChenuAgroParisTech, UMR 7618 BioEMCo, Bâtiment EGER,78850 Thiverval-Grignon, France
N. Nunan (*) :A.-C. DudayCNRS, UMR 7618 Bioemco, Bâtiment EGER,78850 Thiverval Grignon, Francee-mail: [email protected]
V. PouteauINRA, UMR 7618 Bioemco, Bâtiment EGER,78850 Thiverval Grignon, France
Biol Fertil SoilsDOI 10.1007/s00374-013-0784-8

It is now well established that interactions between micro-bial communities and their local environmental (pore networkcharacteristics, chemical properties, oxygen gradients…) notonly have significant effects on microbial community struc-ture (Ruamps et al. 2011; Sleutel et al. 2012; Chaparro et al.2012) but also regulate many of the soil functions carried outby microorganisms, including OM decomposition (Hassink1992; Strong et al. 2004), potential denitrification (Attard et al.2011) and enzyme activity (Allison 2005). The effects thatchanges in microbial diversity might have on these interac-tions are completely unknown, however. The objectives of thestudy presented here were to determine how concomitantchanges in the physical structure of soil and in microbialdiversity affect the decomposition of organic matter. It washypothesised that differences amongst physical structure treat-ments would increase as diversity decreased. This hypothesiswas based on the idea that diversity provides an insuranceagainst fluctuations in ecosystem functioning under differentconditions due to the fact that different species have differentabilities to carry out a given function under different environ-mental conditions (Loreau et al. 2001). It was alsohypothesised that the ability of microbial communities todecompose organic substrate would decrease as microbialdiversity decreased and as complexity of the substrate in-creased. Underlying this second hypothesis is the assumptionthat reductions in diversity levels are accompanied by loss (orsignificant reduction) of rarer catabolic functions.
In order to test these hypotheses, an incubation experimentwas established in which three soil structure treatments(undisturbed, sieved and dispersed) and three diversity levelswere imposed. The different diversity levels were obtained bythe removal approach used in Wertz et al. (2006). In addition,soluble 13C-labelled substrates of different complexity wereadded at the beginning of the incubation. Fructose and vanillinwere chosen because the first is a sugar and the other anaromatic aldehyde and, therefore, differ in their inherent bio-degradability. The production of CO2 and
13C-labelled CO2
were then monitored during a 4-month incubation. At the endof the incubation, the structure of microbial communities wasdetermined by phospholipid fatty acide (PLFA) profiling.
Materials and methods
Soil sampling
In the spring of 2010, undisturbed soil cores (5 cm diam-eter) were sampled from the surface 15 cm of the con-ventional cropping system in the “La Cage” long-termfield trial at the INRA research centre of Versailles(France) and stored at 4 °C until use. The organic C(OC) and total N contents were 9.3±0.7 and 1.0±0.07 mgg!1 dry soil, respectively. The soil is classified
as an Eutric Cambisol (FAO) with 17 % clay, 56 % siltand 27 % sand, with a pH (water: soil=1) of 7.4 and aCEC of 11.5 cmol+ kg!1 dry soil.
Soil core preparation, sterilisation and inoculation
After sampling, undisturbed subsamples were excised fromthe field cores at a depth equivalent to 6–10 cm. The excisedsamples were used to prepare the different structure treat-ments. A third of the samples were left intact and used forthe “undisturbed structure” treatment. A second third of thesamples was sieved to <5 mm for the “sieved structure”treatment. The final third of the samples was disaggregatedby shaking in water with glass beads according to theprotocol proposed by Balesdent et al. (1998). The soilsamples were suspended in deionised water with 5-mmdiameter glass beads and the aggregates ruptured by me-chanical shaking overnight. This ensured that all aggregateswere disrupted to <50 μm. Soil suspension was then air-dried and sieved (<5 mm) to form artificial aggregates. Thisfinal treatment was named “dispersed structure” treatment.The soil in both disturbed treatments was compacted with ahydraulic press to obtain the same bulk density as that of theundisturbed treatment (1.4 gcm!3). To ensure that there wasno bulk density gradient, the soil was packed in layers.Twenty-seven samples for each soil structure (81 soil coresin total) were then sterilised by gamma radiation (75 kGy,with a minimum exposure of 45 kGy, from a 60Co source).The sterilized samples were stored at 4 °C for 1 month inorder to ensure that there were no free radicals remainingafter the exposure to gamma radiation (McNamara et al.2003; Berns et al. 2008). The exposure levels were sufficientto eliminate virtually all microbial species (McNamara et al.2003). The sterile samples were then inoculated with micro-bial suspensions obtained from non-sterile soil sampledfrom the same site. The microbial suspensions were pre-pared by adding 180 mL 1/4 strength Ringer solution to 20 gof non-sterile soil and shaking for 30 min with glass beads.Serial dilutions (10!4, 10!6 and 10!8) of the suspensionswere used to inoculate the sterile soil cores (10 mL suspen-sion per core). The 10!4 dilution was termed high diversitytreatment, the 10!6 dilution intermediate diversity and the10!8 dilution, low diversity. Three independent microbialsuspensions were prepared and the three replicate cores pertreatment were inoculated with one of the microbial suspen-sions. In all, nine samples were prepared for each dilutionand each structure and were incubated as described below.The samples were then pre-incubated for 7 weeks at 20 °Cand at a matrix potential of !31.5 kPa to allow the inocu-lated communities to become established and to avoid thedifferent flushes of mineralisation that occur as the commu-nities consume (1) the labile C released as a result of thesterilisation and (2) the C released during the alterations in
Biol Fertil Soils

soil structure. Baumann et al. (2013) showed that after a 6week pre-incubation microbial densities in microcosms in-oculated with different diversity treatments had stabilised ata similar level. During the pre-incubation, the respirationrates of all treatments were stable between the fourth andseventh weeks, suggesting that the flushes of mineralisationhad passed (data not shown).
Addition of two 13C-labelled substrates (fructoseand vanillin) and incubation
At the end of the 7-week pre-incubation, 27 microcosms(three replicate microcosms for each level of microbialdiversity and each structure) were amended with a solutionof either 13C-labelled fructose or 13C-labelled vanillin (δ13Cfructose=1722 ‰ and δ13C vanillin=1272 ‰) at a concen-tration equivalent to 15 mg substrate C g!1 soil C. Unlikethe native SOM, the added substrates had a defined initiallocation in the samples: the substrate solutions (or water inthe case of the control samples) were added in order to bringthe matric potential of the samples to !31.5 kPa (or pF 2.5).The samples were then incubated at this matric potential. Thesubstrates had to be soluble to facilitate the amendment ofundisturbed samples. These treatments allowed us to deter-mine the effects of soil microbial diversity and soil structureon the mineralisation of different types of organic C.
Immediately after the amendment, the samples were re-placed on sample holders in 1-L airtight jars with springlock catches. The jars were sealed with rubber gaskets and aglass lid fitted with a septum for headspace sampling. Thejars had a wide opening to enable the incubation and easyremoval of the cores. In order to maintain constant humidityduring the incubation, 20 mL water was placed at the bottomof each jar. The microcosms were incubated at 20 °C in thedark for 127 days. In total, 81 microcosms were incubated(three microbial diversities! three structures!three amend-ments! three replicates).
Measurements of CO2 concentration and δ13C
Organic C mineralisation was monitored during the incuba-tion by measuring the CO2 concentration in the microcosmheadspaces on days 1, 3, 7, 14, 21, 28, 49, 70, 91 and 127. TheCO2 concentration was determined by gas chromatographywith a Micro GC (Agilent 3000A, Santa Clara, CA, USA).The headspace CO2 concentration was determined in parts permillion and then converted to milligram C-CO2 per kilogramdry soil equivalent. The δ13C of the headspace CO2 wasmeasured using an automatic unit that combined a 5890 GCgas chromatograph (Hewlett-Packard, Palo Alto, California,USA) coupled to an isotope ratio mass spectrometer(Isochrome III, Micromass, GVI Optima) equipped with a 3-m column filled with a Prorapak QS stationary phase (80–100
mesh). At the beginning of the incubation, the jars' atmospherewas CO2-free, and after each measurement, all jars wereflushed with reconstituted and moistened CO2-free air.
Quantification of labelled substrate and native organiccarbon mineralisation
SOC mineralisation in the samples that had received la-belled substrate (fructose or vanillin) was calculated usingthe following equation:
SOC mineralisation ! a " Qsample #1$
where !=(Afructose or vanillin!Asample) / (Afructose or vanillin!Acontrol),Q is the amount of CO2 in the flask's atmosphere in milligramCO2-C per kilogram dry soil equivalent and Afructose or vanillin,Asample and Acontrol represent the isotopic abundance offructose or vanillin, of CO2 in the sample headspaceand of CO2 in headspace of control samples (i.e., sam-ples to which water was added rather than labelledsubstrate), respectively. The mineralisation of labelledsubstrate was calculated using the following equation:
Fructose or vanillin mineralisation ! 1% a# $ " Qsample:
Phospholipid fatty acid analysis and microbial biomass
Lipid profiles were established after extraction from 10 gsoil using the method described by Frostegård et al. (1993).Phospholipids were separated from neutral and glycolipidson solid-phase extraction columns (Extract-Clean® Silica,Grace). After methylation, fatty acid methyl esters (FAME)were identified using the retention time of chromatogramsand mass spectral comparison on a Hewlett-Packard 6890gas chromatograph equipped with either a Flame IonisationDetector (GC-FID) or an Agilent 5973 Electronic Impact(70 eV) quadruple Mass Spectrometer (GC–MS). A stan-dard qualitative bacterial acid methyl esters mix (Sigma-Aldrich) that ranged from C11:0 to C20:0 was used for theidentification of sample FAME based on retention time.
The relative abundance of each fatty acid was used tocompare the community structure between the samples. Allfatty acids contributing less than 1 % to the fatty acids poolwere removed before analysis. Standard fatty acid nomencla-ture was used, as described previously in Frostegård et al.(1993). Mono-unsaturated and cyclopropyl fatty acids(C16:1w9, C18:1w9c, C18:1w9t, cycC17:0 and cycC19:0)were taken as Gram-negative bacterial biomarkers (Zelles1999), iso- and anteiso-fatty acids (iC15:0, aC15:0, iC16:0,iC17:0 and aC17:0) as Gram-positive bacterial biomarkers(Zelles 1999), C18:2w(9,12) as a fungal biomarker(Frostegård et al. 1993; Zelles 1997). The total bacterialcommunity was represented by the sum of the Gram-
#2$
Biol Fertil Soils

positive and Gram-negative bacterial biomarkers as well asC15:0 and C17:0. Fungi/bacteria and Gram-positive/Gram-negative ratios were calculated by dividing the respectivesums of biomarkers. The total microbial biomass was estimat-ed as the total PLFA peaks areas (Frostegård and Bååth 1996).
Statistical analysis
Statistical analyses were performed using R version 2.14.0(R Development Core Team, 2009. R: a language and envi-ronment for statistical computing. Available at http://www.Rproject.org). A two-compartment, first-order model(Eq. 3) was used to describe the mineralisation kinetics ofthe two added substrates:
CO2 ! C fast 1% e%at# $ & Cslow 1% e%bt! "#3$
where t is time in days,Cfast is the pool of substrate C (fructoseor vanillin) that was mineralised rapidly, Cslow is the pool ofsubstrate C that was mineralised more slowly and ! and " arethe associated first-order rate constants. The effect of thevarious treatments on the mineralisation of fructose and van-illin was verified by analysis of variance (ANOVA) of the fourmodel parameters. The rate parameters (! and ") were logtransformed before ANOVA in order to normalise the data.The use of this model to describe the mineralisation of theadded substrates can be justified on the basis that a portion ofsubstrate C is mineralised directly whilst the remaining sub-strate C is mineralised only after microbial processing. Thismeans that the molecular form of part of the labelled C wasaltered before being mineralised. It is also probable that someof the original substrate was mineralised more slowly becauseit was located in pores that did not contain degraders andtherefore, had to diffuse to sites of degradation activity.
The model did not converge for SOC mineralisationcurves. Therefore, differences in total SOC mineralisationat the end of the incubation were analysed by three-wayANOVA using diversity, structure and substrate treatmentsas factors (diversity ! structure ! substrate amendment).Differences in microbial biomass were analysed by the sameANOVA structure. The impact of the treatments (diversity,structure and substrate amendment) on microbial communi-ty structure (PLFA) was determined after ordination byPrincipal Components Analysis (PCA) using the “vegan:Community Ecology Package” package in R version 2.14.0.
Results
SOC mineralisation
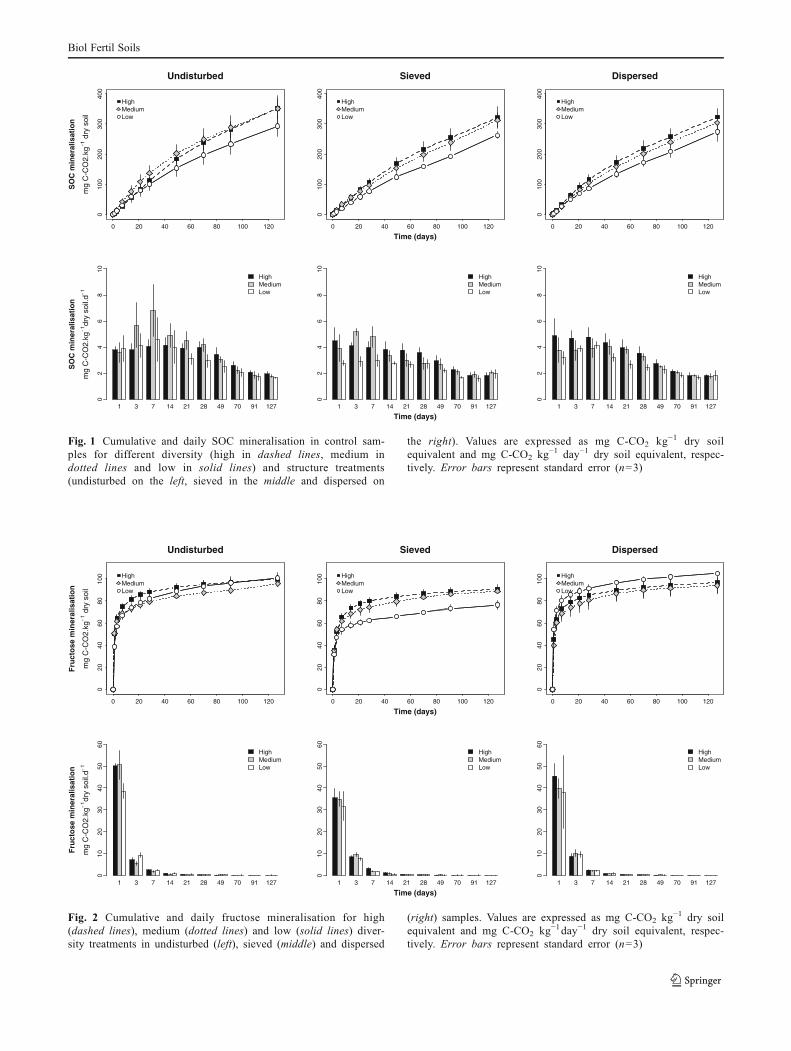
The shape of cumulative SOC mineralisation curves wasalways the same, regardless of treatment (Figs. 1 and S1).
The mineralisation rate was highest at the beginning of theincubation and decreased gradually to reach a constant rateafter about 2 months (Fig. 1). At the end of the incubation,there were significant differences in the amount of Cmineralised amongst diversity levels, with the least diversetreatment mineralising significantly less (P<0.05) than themost diverse treatment (Figs. 1 and S1). There were nosignificant differences amongst structure treatments andthe addition of fresh substrate (fructose or vanillin) did nothave a significant effect on the mineralisation of SOC (i.e.,there was no priming effect).
Substrate mineralisation
The mineralisation dynamics of the two substrates were clear-ly different: more fructose C than vanillin C was mineralisedduring the incubation and the rate of fructose C mineralisationat the beginning of the incubation was higher (Figs. 2 and 3).All the model parameters (sizes of the fast and slow pools andmineralisation rates of both pools) were significantly greaterfor fructose mineralisation than for vanillin mineralisation(Table 1). In samples amended with fructose, alterations inthe physical structure of the soil significantly affected the size(P<0.05) and mineralisation rate of the Cfast pool (P<0.01)but had no effect on theCslow pool. TheCfast pool in the sievedsamples was smaller than in the dispersed samples, and thispool was mineralised more slowly in sieved samples than inundisturbed samples (Table 1). There were also diversityeffects: the mineralisation rates of both C pools were consis-tently the lowest across treatments, but the differences wereonly significant in the case of the Cslow pool rate of thefructose amended samples (Table 1). The size of both C poolsdid not vary as a function of diversity level. There were nosignificant interactions between the structure and diversitytreatments for any of the parameters, suggesting that the Cmineralisation activity of the different microbial diversitylevels was affected in the same way by the alterations in soilstructure.
Microbial biomass and PLFA profiles
No treatment effects on the total microbial biomass, esti-mated as the total PLFA peaks areas, were observed. Theratio of C18:2w(9,12) to bacterial PLFA was used as anindex of the fungal/bacterial ratio. This ratio was signifi-cantly higher in undisturbed samples (P<0.01) becausesignificantly more fungal biomarkers were found inundisturbed samples (P<0.01). This ratio was, however,not significantly affected by the diversity or substrate treat-ments. The ratio of Gram+/Gram!was significantly higherin the dispersed samples (P<0.01) and also slightly lower insamples inoculated with the medium diversity treatment(P<0.05) (data not shown).
Biol Fertil Soils
0 20 40 60 80 100 120
010
020
030
040
0
SO
C m
iner
alis
atio
n
mg
C-C
O2.
kg1 d
ry s
oil
Undisturbed
HighMediumLow
0 20 40 60 80 100 120
010
020
030
040
0
Time (days)
Sieved
HighMediumLow
0 20 40 60 80 100 120
010
020
030
040
0
Dispersed
HighMediumLow
1 3 7 14 21 28 49 70 91 127
SO
C m
iner
alis
atio
n
02
46
810
mg
C-C
O2.
kg1 dr
y so
il.d
1
HighMediumLow
1 3 7 14 21 28 49 70 91 127
02
46
810
Time (days)
HighMediumLow
1 3 7 14 21 28 49 70 91 127
02
46
810
HighMediumLow
Fig. 1 Cumulative and daily SOC mineralisation in control sam-ples for different diversity (high in dashed lines, medium indotted lines and low in solid lines) and structure treatments(undisturbed on the left, sieved in the middle and dispersed on
the right). Values are expressed as mg C-CO2 kg!1 dry soilequivalent and mg C-CO2 kg!1 day!1 dry soil equivalent, respec-tively. Error bars represent standard error (n=3)
0 20 40 60 80 100 120
020
4060
8010
0
Fruc
tose
min
eral
isat
ion
mg
C-C
O2.
kg1 d
ry s
oil
Undisturbed
HighMediumLow
0 20 40 60 80 100 120
020
4060
8010
0
Time (days)
Sieved
HighMediumLow
0 20 40 60 80 100 120
020
4060
8010
0
Dispersed
HighMediumLow
1 3 7 14 21 28 49 70 91 127
Fruc
tose
min
eral
isat
ion
010
2030
4050
60
mg
C-C
O2.
kg1 dr
y so
il.d
1
HighMediumLow
1 3 7 14 21 28 49 70 91 127
010
2030
4050
60
Time (days)
HighMediumLow
1 3 7 14 21 28 49 70 91 127
010
2030
4050
60
HighMediumLow
Fig. 2 Cumulative and daily fructose mineralisation for high(dashed lines), medium (dotted lines) and low (solid lines) diver-sity treatments in undisturbed (left), sieved (middle) and dispersed
(right) samples. Values are expressed as mg C-CO2 kg!1 dry soilequivalent and mg C-CO2 kg!1day!1 dry soil equivalent, respec-tively. Error bars represent standard error (n=3)
Biol Fertil Soils
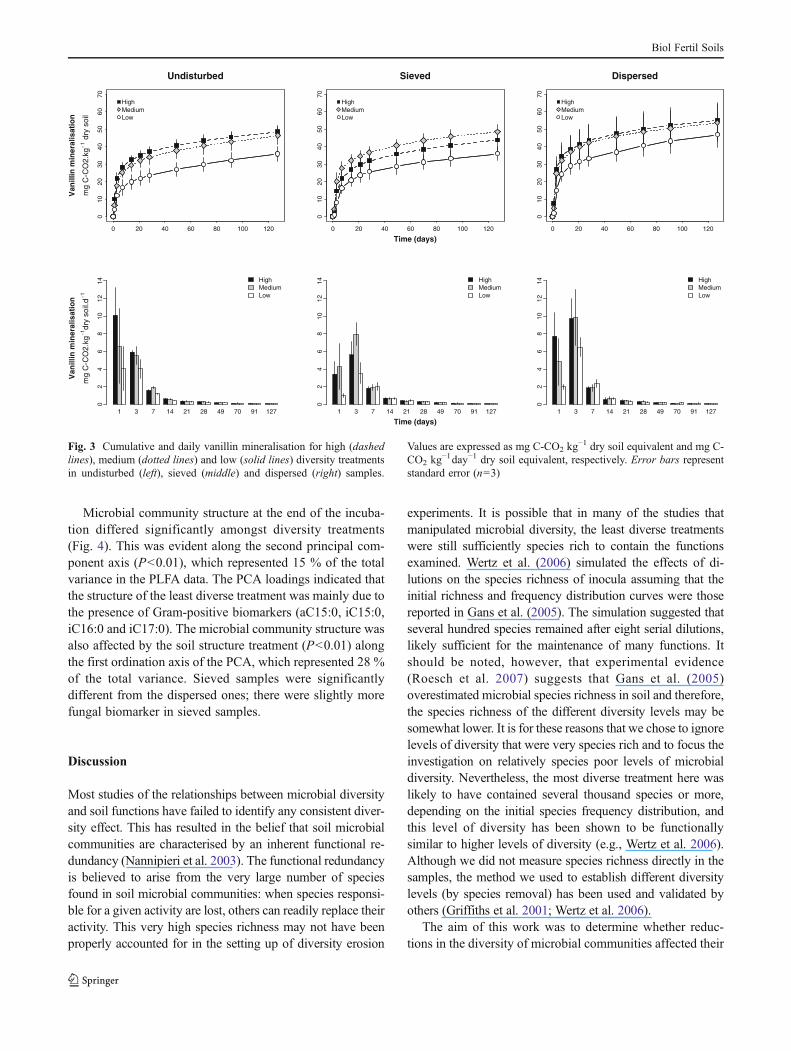
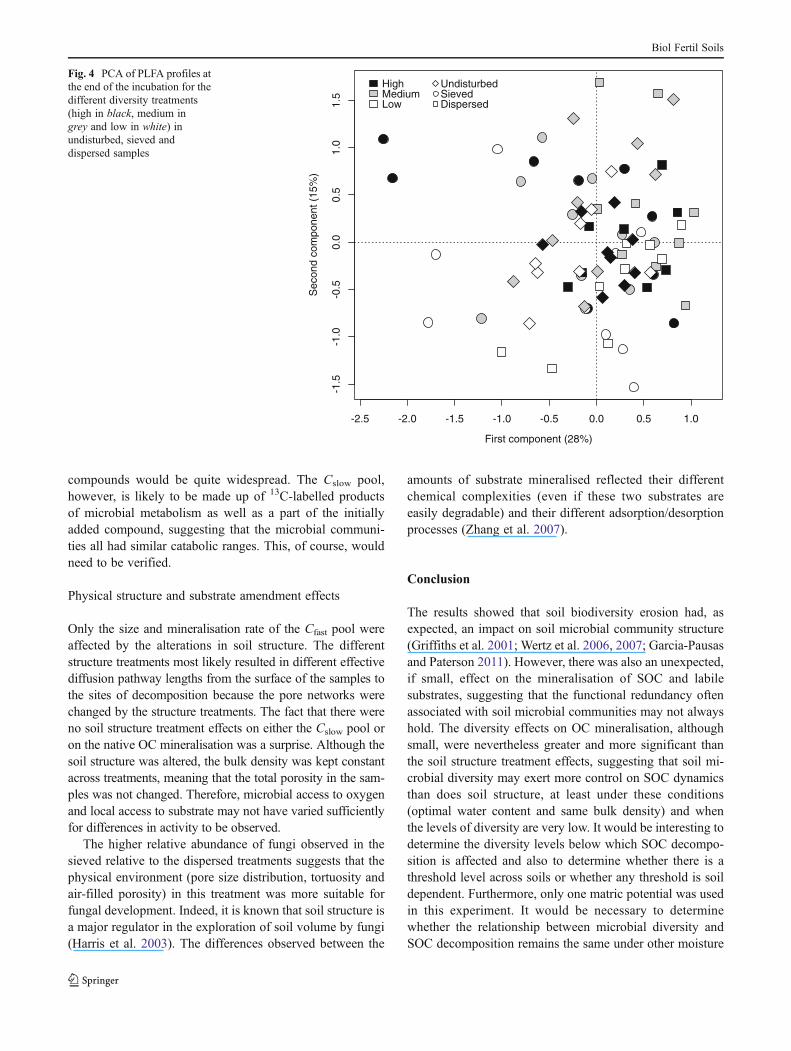
Microbial community structure at the end of the incuba-tion differed significantly amongst diversity treatments(Fig. 4). This was evident along the second principal com-ponent axis (P<0.01), which represented 15 % of the totalvariance in the PLFA data. The PCA loadings indicated thatthe structure of the least diverse treatment was mainly due tothe presence of Gram-positive biomarkers (aC15:0, iC15:0,iC16:0 and iC17:0). The microbial community structure wasalso affected by the soil structure treatment (P<0.01) alongthe first ordination axis of the PCA, which represented 28 %of the total variance. Sieved samples were significantlydifferent from the dispersed ones; there were slightly morefungal biomarker in sieved samples.
Discussion
Most studies of the relationships between microbial diversityand soil functions have failed to identify any consistent diver-sity effect. This has resulted in the belief that soil microbialcommunities are characterised by an inherent functional re-dundancy (Nannipieri et al. 2003). The functional redundancyis believed to arise from the very large number of speciesfound in soil microbial communities: when species responsi-ble for a given activity are lost, others can readily replace theiractivity. This very high species richness may not have beenproperly accounted for in the setting up of diversity erosion
experiments. It is possible that in many of the studies thatmanipulated microbial diversity, the least diverse treatmentswere still sufficiently species rich to contain the functionsexamined. Wertz et al. (2006) simulated the effects of di-lutions on the species richness of inocula assuming that theinitial richness and frequency distribution curves were thosereported in Gans et al. (2005). The simulation suggested thatseveral hundred species remained after eight serial dilutions,likely sufficient for the maintenance of many functions. Itshould be noted, however, that experimental evidence(Roesch et al. 2007) suggests that Gans et al. (2005)overestimated microbial species richness in soil and therefore,the species richness of the different diversity levels may besomewhat lower. It is for these reasons that we chose to ignorelevels of diversity that were very species rich and to focus theinvestigation on relatively species poor levels of microbialdiversity. Nevertheless, the most diverse treatment here waslikely to have contained several thousand species or more,depending on the initial species frequency distribution, andthis level of diversity has been shown to be functionallysimilar to higher levels of diversity (e.g., Wertz et al. 2006).Although we did not measure species richness directly in thesamples, the method we used to establish different diversitylevels (by species removal) has been used and validated byothers (Griffiths et al. 2001; Wertz et al. 2006).
The aim of this work was to determine whether reduc-tions in the diversity of microbial communities affected their
0 20 40 60 80 100 120
010
2030
4050
6070
Van
illin
min
eral
isat
ion
mg
C-C
O2.
kg1 d
ry s
oil
Undisturbed
HighMediumLow
0 20 40 60 80 100 120
010
2030
4050
6070
Time (days)
Sieved
HighMediumLow
0 20 40 60 80 100 120
010
2030
4050
6070
Dispersed
HighMediumLow
1 3 7 14 21 28 49 70 91 127
Van
illin
min
eral
isat
ion
02
46
810
1214
mg
C-C
O2.
kg1 dr
y so
il.d
1
HighMediumLow
1 3 7 14 21 28 49 70 91 127
02
46
810
1214
Time (days)
HighMediumLow
1 3 7 14 21 28 49 70 91 127
02
46
810
1214 High
MediumLow
Fig. 3 Cumulative and daily vanillin mineralisation for high (dashedlines), medium (dotted lines) and low (solid lines) diversity treatmentsin undisturbed (left), sieved (middle) and dispersed (right) samples.
Values are expressed as mg C-CO2 kg!1 dry soil equivalent and mg C-
CO2 kg!1day!1 dry soil equivalent, respectively. Error bars represent
standard error (n=3)
Biol Fertil Soils
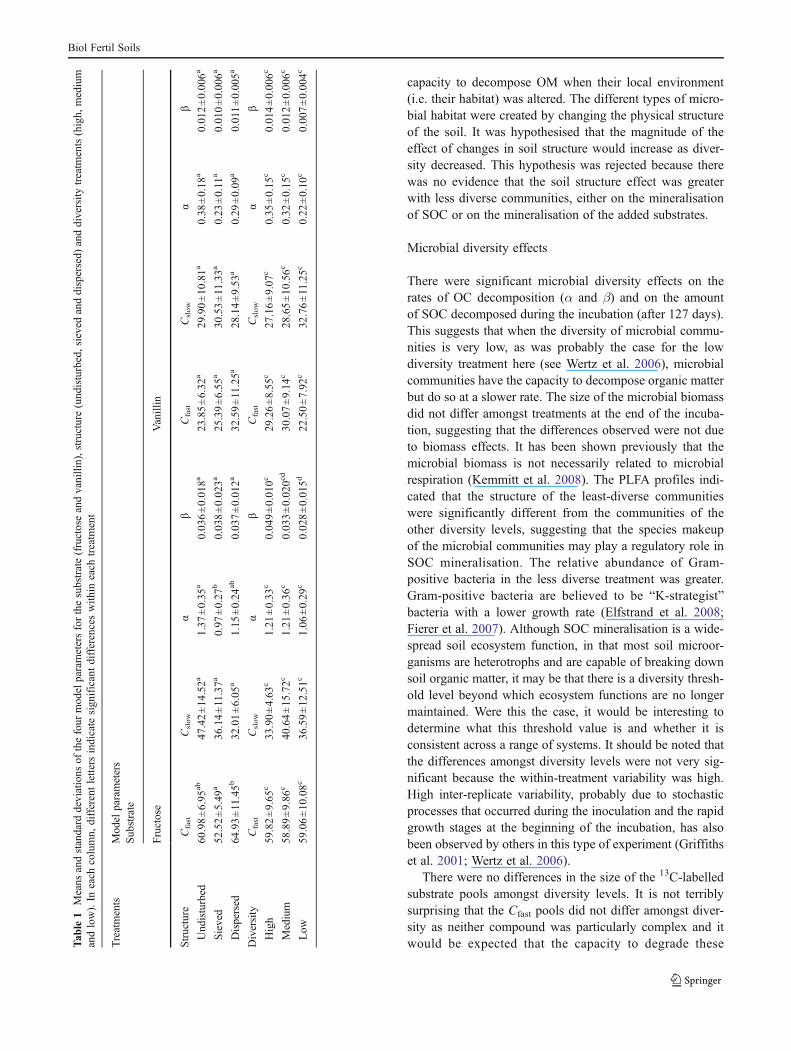
capacity to decompose OM when their local environment(i.e. their habitat) was altered. The different types of micro-bial habitat were created by changing the physical structureof the soil. It was hypothesised that the magnitude of theeffect of changes in soil structure would increase as diver-sity decreased. This hypothesis was rejected because therewas no evidence that the soil structure effect was greaterwith less diverse communities, either on the mineralisationof SOC or on the mineralisation of the added substrates.
Microbial diversity effects
There were significant microbial diversity effects on therates of OC decomposition (! and ") and on the amountof SOC decomposed during the incubation (after 127 days).This suggests that when the diversity of microbial commu-nities is very low, as was probably the case for the lowdiversity treatment here (see Wertz et al. 2006), microbialcommunities have the capacity to decompose organic matterbut do so at a slower rate. The size of the microbial biomassdid not differ amongst treatments at the end of the incuba-tion, suggesting that the differences observed were not dueto biomass effects. It has been shown previously that themicrobial biomass is not necessarily related to microbialrespiration (Kemmitt et al. 2008). The PLFA profiles indi-cated that the structure of the least-diverse communitieswere significantly different from the communities of theother diversity levels, suggesting that the species makeupof the microbial communities may play a regulatory role inSOC mineralisation. The relative abundance of Gram-positive bacteria in the less diverse treatment was greater.Gram-positive bacteria are believed to be “K-strategist”bacteria with a lower growth rate (Elfstrand et al. 2008;Fierer et al. 2007). Although SOC mineralisation is a wide-spread soil ecosystem function, in that most soil microor-ganisms are heterotrophs and are capable of breaking downsoil organic matter, it may be that there is a diversity thresh-old level beyond which ecosystem functions are no longermaintained. Were this the case, it would be interesting todetermine what this threshold value is and whether it isconsistent across a range of systems. It should be noted thatthe differences amongst diversity levels were not very sig-nificant because the within-treatment variability was high.High inter-replicate variability, probably due to stochasticprocesses that occurred during the inoculation and the rapidgrowth stages at the beginning of the incubation, has alsobeen observed by others in this type of experiment (Griffithset al. 2001; Wertz et al. 2006).
There were no differences in the size of the 13C-labelledsubstrate pools amongst diversity levels. It is not terriblysurprising that the Cfast pools did not differ amongst diver-sity as neither compound was particularly complex and itwould be expected that the capacity to degrade theseT
able1
Means
andstandard
deviations
ofthefour
mod
elparametersforthesubstrate(fructoseandvanillin),structure
(und
isturbed,sievedanddispersed)
anddiversity
treatm
ents(high,
medium
andlow).In
each
column,
differentletters
indicate
sign
ificantdifferenceswith
ineach
treatm
ent
Treatments
Mod
elparameters
Substrate
Fructose
Vanillin
Structure
Cfast
Cslow
!"
Cfast
Cslow
!"
Und
isturbed
60.98±6.95
ab47
.42±14
.52a
1.37
±0.35
a0.03
6±0.01
8a23
.85±6.32
a29
.90±10
.81a
0.38
±0.18
a0.01
2±0.00
6a
Sieved
52.52±5.49
a36
.14±11.37a
0.97
±0.27
b0.03
8±0.02
3a25
.39±6.55
a30
.53±11.33a
0.23
±0.11
a0.01
0±0.00
6a
Dispersed
64.93±11.45b
32.01±6.05
a1.15
±0.24
ab0.03
7±0.01
2a32
.59±11.25a
28.14±9.53
a0.29
±0.09
a0.011±0.00
5a
Diversity
Cfast
Cslow
!"
Cfast
Cslow
!"
High
59.82±9.65
c33
.90±4.63
c1.21
±0.33
c0.04
9±0.01
0c29
.26±8.55
c27
.16±9.07
c0.35
±0.15
c0.01
4±0.00
6c
Medium
58.89±9.86
c40
.64±15
.72c
1.21
±0.36
c0.03
3±0.02
0cd
30.07±9.14
c28
.65±10
.56c
0.32
±0.15
c0.01
2±0.00
6c
Low
59.06±10
.08c
36.59±12
.51c
1.06
±0.29
c0.02
8±0.01
5d22
.50±7.92
c32
.76±11.25c
0.22
±0.10
c0.00
7±0.00
4c
Biol Fertil Soils
compounds would be quite widespread. The Cslow pool,however, is likely to be made up of 13C-labelled productsof microbial metabolism as well as a part of the initiallyadded compound, suggesting that the microbial communi-ties all had similar catabolic ranges. This, of course, wouldneed to be verified.
Physical structure and substrate amendment effects
Only the size and mineralisation rate of the Cfast pool wereaffected by the alterations in soil structure. The differentstructure treatments most likely resulted in different effectivediffusion pathway lengths from the surface of the samples tothe sites of decomposition because the pore networks werechanged by the structure treatments. The fact that there wereno soil structure treatment effects on either the Cslow pool oron the native OC mineralisation was a surprise. Although thesoil structure was altered, the bulk density was kept constantacross treatments, meaning that the total porosity in the sam-ples was not changed. Therefore, microbial access to oxygenand local access to substrate may not have varied sufficientlyfor differences in activity to be observed.
The higher relative abundance of fungi observed in thesieved relative to the dispersed treatments suggests that thephysical environment (pore size distribution, tortuosity andair-filled porosity) in this treatment was more suitable forfungal development. Indeed, it is known that soil structure isa major regulator in the exploration of soil volume by fungi(Harris et al. 2003). The differences observed between the
amounts of substrate mineralised reflected their differentchemical complexities (even if these two substrates areeasily degradable) and their different adsorption/desorptionprocesses (Zhang et al. 2007).
Conclusion
The results showed that soil biodiversity erosion had, asexpected, an impact on soil microbial community structure(Griffiths et al. 2001; Wertz et al. 2006, 2007; Garcia-Pausasand Paterson 2011). However, there was also an unexpected,if small, effect on the mineralisation of SOC and labilesubstrates, suggesting that the functional redundancy oftenassociated with soil microbial communities may not alwayshold. The diversity effects on OC mineralisation, althoughsmall, were nevertheless greater and more significant thanthe soil structure treatment effects, suggesting that soil mi-crobial diversity may exert more control on SOC dynamicsthan does soil structure, at least under these conditions(optimal water content and same bulk density) and whenthe levels of diversity are very low. It would be interesting todetermine the diversity levels below which SOC decompo-sition is affected and also to determine whether there is athreshold level across soils or whether any threshold is soildependent. Furthermore, only one matric potential was usedin this experiment. It would be necessary to determinewhether the relationship between microbial diversity andSOC decomposition remains the same under other moisture
-2.5 -2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
First component (28%)
Sec
ond
com
pone
nt (
15%
)
HighMediumLow
UndisturbedSievedDispersed
Fig. 4 PCA of PLFA profiles atthe end of the incubation for thedifferent diversity treatments(high in black, medium ingrey and low in white) inundisturbed, sieved anddispersed samples
Biol Fertil Soils

conditions in order to better predict how decomposition willbe affected by future climate change.
Acknowledgments This work was funded by the Agence Nationalede la Recherche (ANR) under the “SYSCOMM (Systèmes complexeset modélisation mathématique)” Program (MEPSOM project) and bythe Région Ile-de-France (DIM Astréa). The authors would like tothank Daniel Billiou for help with the elemental analysis and GérardBardoux for the isotopic measurements. They would also like to thankMichel Bertrand and the Unité expérimentale for the access and formaintaining the “La Cage” long-term field trial.
References
Allison SD (2005) Cheaters, diffusion and nutrients constrain decompo-sition by microbial enzymes in spatially structured environments.Ecol Lett 8:626–635. doi:10.1111/j.1461-0248.2005.00756.x
Attard E, Recous S, Chabbi A, De Berranger C, Guillaumaud N,Labreuche J, Philippot L, Schmid B, Le Roux X (2011) Soilenvironmental conditions rather than denitrifier abundance anddiversity drive potential denitrification after changes in land uses.Glob Chang Biol 17:1975–1989. doi:10.1111/j.1365-2486.2010.02340.x
Balesdent J, Besnard E, Arrouays D, Chenu C (1998) The dynamics ofcarbon in particle-size fractions of soil in a forest-cultivationsequence. Plant Soil 201:49–57. doi:10.1023/A:1004337314970
Baumann K, Dignac MF, Rumpel C, Bardoux G, Sarr A, Steffens M,Maron PA (2013) Soil microbial diversity affects soil organicmatter decomposition in a silty grassland soil. Biogeochemistry.doi: 10.1007/s10533-012-9800-6
Berns AE, Philipp H, Narres HD, Burauel P, Vereecken H, Tappe W(2008) Effect of gamma-sterilization and autoclaving on soilorganic matter structure as studied by solid state NMR, UV andfluorescence spectroscopy. Eur J Soil Sci 59:540–550.doi:10.1111/j.1365-2389.2008.01016.x
Cardinale BJ, Srivastava DS, Duffy JE, Wright JP, Downing AL,Sankaran M, Jouseau C (2006) Effects of biodiversity on thefunctioning of trophic groups and ecosystems. Nature 443:989–992. doi:10.1038/nature05202
Chaparro JM, Sheflin AM,Manter DK, Vivanco JM (2012) Manipulatingthe soil microbiome to increase soil health and plant fertility. BiolFertil Soils 48:489–499. doi:10.1007/s00374-012-0691-4
Chapin FS III, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM,Reynolds HL, Hooper DU, Lavorel S, Sala OE, Hobbie SE, MackMC, Díaz S (2000) Consequences of changing biodiversity.Nature 405:234–242. doi:10.1038/35012241
Elfstrand S, Lagerlöf J, Hedlund K, Mårtensson A (2008) Carbonroutes from decomposing plant residues and living roots into soilfood webs assessed with 13C labelling. Soil Biol Biochem40:2530–2539. doi:10.1016/j.soilbio.2008.06.013
Fierer N, Bradford M, Jackson R (2007) Toward an ecological classi-fication of soil bacteria. Ecology 88:1354–1364
Frostegård A, Bååth E (1996) The use of phospholipid fatty acidanalysis to estimate bacterial and fungal biomass in soil. BiolFertil Soils 22:59–65
Frostegård A, Bååth E, Tunlid A (1993) Shifts in the structure of soilmicrobial communities in limed forests as revealed by phospho-lipid fatty-acid analysis. Soil Biol Biochem 25:723–730
Gans J, Wolinsky M, Dunbar J (2005) Computational improvementsreveal great bacterial diversity and high metal toxicity in soil.Science 309:1387–1390. doi:10.1126/science.1112665
Garcia-Pausas J, Paterson E (2011)Microbial community abundance andstructure are determinants of soil organic matter mineralisation in the
presence of labile carbon. Soil Biol Biochem 43:1705–1713.doi:10.1016/j.soilbio.2011.04.016
Griffiths BS, Ritz K, Bardgett RD, Cook R, Christensen S, Ekelund F,Sørensen SJ, Bååth E, Bloem J, de Ruiter PC, Dolfing J,Nicolardot B (2000) Ecosystem response of pasture soil commu-nities to fumigation-induced microbial diversity reductions: anexamination of the biodiversity–ecosystem function relationship.Oikos 90:279–294. doi:10.1034/j.1600-0706.2000.900208.x
Griffiths BS, Ritz K, Wheatley R, Kuan HL, Boag B, Christensen S,Ekelund F, Sorensen SJ, Muller S, Bloem J (2001) An examina-tion of the biodiversity–ecosystem function relationship in arablesoil microbial communities. Soil Biol Biochem 33:1713–1722
Harris K, Young IM, Gilligan CA, Otten W, Ritz K (2003) Effect ofbulk density on the spatial organisation of the fungus Rhizoctoniasolani in soil. FEMS Microbiol Ecol 44:45–56. doi:10.1111/j.1574-6941.2003.tb01089.x
Hassink J (1992) Effects of soil texture and structure on carbon andnitrogen mineralization in grassland soils. Biol Fertil Soils14:126–134. doi:10.1007/BF00336262
Hector A, Schmid B, Beierkuhnlein C, Caldeira MC, Diemer M,Dimitrakopoulos PG, Finn JA, Freitas H, Giller PS, Good J,Harris R, Högberg P, Huss-Danell K, Joshi J, Jumpponen A,Körner C, Leadley PW, Loreau M, Minns A, Mulder CPH,O'Donovan G, Otway SJ, Pereira JS, Prinz A, Read DJ, Scherer-Lorenzen M, Schulze E-D, Siamantziouras A-SD, Spehn EM,Terry AC, Troumbis AY, Woodward FI, Yachi S, Lawton JH(1999) Plant diversity and productivity experiments inEuropean grasslands. Science 286:1123–1127. doi:10.1126/science.286.5442.1123
Kemmitt SJ, Lanyon CV, Waite IS, Wen Q, Addiscott TM, Bird NRA,O'Donnell AG, Brookes PC (2008) Mineralization of native soilorganic matter is not regulated by the size, activity or compositionof the soil microbial biomass—a new perspective. Soil BiolBiochem 40:61–73. doi:10.1016/j.soilbio.2007.06.021
Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime JP, Hector A,Hooper DU, Huston MA, Raffaelli D, Schmid B, Tilman D,Wardle DA (2001) Biodiversity and ecosystem functioning: cur-rent knowledge and future challenges. Science 294:804–808.doi:10.1126/science.1064088
Loreau M, Mouquet N, Gonzalez A (2003) Biodiversity as spatialinsurance in heterogeneous landscapes. PNAS 100:12765–12770. doi:10.1073/pnas.2235465100
Martin SL, Mooney SJ, Dickinson MJ, West HM (2012) Soil structuralresponses to alterations in soil microbiota induced by the dilutionmethod and mycorrhizal fungal inoculation. Pedobiologia55:271–281. doi:10.1016/j.pedobi.2012.06.001
McNamara NP, Black HIJ, Beresford NA, Parekh NR (2003) Effects ofacute gamma irradiation on chemical, physical and biologicalproperties of soils. Appl Soil Ecol 24:117–132. doi:10.1016/S0929-1393(03)00073-8
Nannipieri P, Ascher J, Ceccherini MT, Landi L, Pietramellara G,Renella G (2003) Microbial diversity and soil functions. Eur JSoil Sci 54:655–670. doi:10.1046/j.1351-0754.2003.0556.x
Nielsen UN, Ayres E, Wall DH, Bardgett RD (2011) Soil biodiversityand carbon cycling: a review and synthesis of studies examiningdiversity–function relationships. Eur J Soil Sci 62:105–116.doi:10.1111/j.1365-2389.2010.01314.x
Roesch LF, Fulthorpe RR, Riva A, Casella G, Hadwin AKM, KentAD, Daroubs SH, Camargo FAO, Farmerie WG, Triplett EW(2007) Pyrosequencing enumerates and contrasts soil microbialdiversity. ISME J 1:283–290. doi:10.1038/ismej.2007.53
Ruamps LS, Nunan N, Chenu C (2011) Microbial biogeography at thesoil pore scale. Soil Biol Biochem 43:280–286. doi:10.1016/j.soilbio.2010.10.010
Sleutel S, Bouckaert L, Buchan D, Van Loo D, Cornelis WM, SangaHG (2012) Manipulation of the soil pore and microbial
Biol Fertil Soils

community structure in soil mesocosm incubation studies. SoilBiol Biochem 45:40–48. doi:10.1016/j.soilbio.2011.09.016
Strong DT, Wever HD, Merckx R, Recous S (2004) Spatial location ofcarbon decomposition in the soil pore system. Eur J Soil Sci55:739–750. doi:10.1111/j.1365-2389.2004.00639.x
Tilman D, Reich PB, Isbell F (2012) Biodiversity impacts ecosystemproductivity as much as resources, disturbance, or herbivory.PNAS 109:10394–10397. doi:10.1073/pnas.1208240109
Wertz S, Degrange V, Prosser JI, Poly F, Commeaux C, Freitag T,Guillaumaud N, Le Roux X (2006) Maintenance of soil function-ing following erosion of microbial diversity. Environ Microbiol8:2162–2169. doi:10.1111/j.1462-2920.2006.01098.x
Wertz S, Degrange V, Prosser JI, Poly F, Commeaux C,Guillaumaud N, Le Roux X (2007) Decline of soil microbial
diversity does not influence the resistance and resilience ofkey soil microbial functional groups following a model dis-turbance. Environ Microbiol 9:2211–2219. doi:10.1111/j.1462-2920.2007.01335.x
Zelles L (1997) Phospholipid fatty acid profiles in selected members ofsoil microbial communities. Chemosphere 35:275–294.doi:10.1016/S0045-6535(97)00155-0
Zelles L (1999) Fatty acid patterns of phospholipids and lipopoly-saccharides in the characterisation of microbial communitiesin soil: a review. Biol Fertil Soils 29:111–129. doi:10.1007/s003740050533
Zhang Q-F, Jiang Z-T, Gao H-J, Li R (2007) Recovery of vanillin fromaqueous solutions using macroporous adsorption resins. Eur FoodRes Technol 226:377–383. doi:10.1007/s00217-006-0548-x
Biol Fertil Soils
Related Documents











