Neuromusc ular Jun ctio n Lectu re Notes These notes supplement the lectures and cover background material that will help place the experiments in context. © University of Minnesota Version: July, 2008

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 1/35
Neuromuscular JunctionLecture Notes
These notes supplement the lectures and cover background material that will help placethe experiments in context.
© University of MinnesotaVersion: July, 2008

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 2/35

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 3/35

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 4/35

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 5/35
1 Muscle Contraction: cardiac,smooth and skeletal
1. Types On the basis of structure, contractile properties and control mechanisms, threetypes of muscle can be identified: 1) skeletal muscle, 2) smooth muscle and 3) cardiacmuscle. Although there are significant differences between these muscle types, the force-generating mechanisms are similar.
Skeletal Muscle: most skeletal
muscle is attached to bone and itscontraction is responsible for supporting and moving the skeleton.The contraction of these muscles isinitiated by action potential propagating down motoneurons to themuscle and can be under voluntarycontrol.
Smooth Muscle: sheets of smoothmuscle surround various hollow
organs and tubes (e.g., stomach,intestines, urinary bladder, uterus, blood vessels and airways).Contraction of these cells may propelthe luminal contents through theorgan or regulate internal flows bychanging tube diameters. Single andgroups of smooth muscle cells arealso found distributed throughoutorgans and perform various other functions: e.g., iris of the eye and
attachment of hair. Smooth musclecontraction can be spontaneous or controlled by: the autonomic nervoussystem, hormones and other chemicalsignals.
Cardiac Muscle: The muscle of theheart surrounds four pumping
NMJ Lecture Notes 3

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 6/35
chambers. Contraction of cardiac muscle provides the impetus for the movement of blood through the pulmonary and systemic circulatory systems. Spontaneous cyclingof an intrinsic pacemaker triggers each heartbeat (or contraction). However theautonomic nervous system and circulating hormones modulate the frequency of thisactivation.
2. Structure and Function of Skeletal Muscle
If one sections through a skeletal muscle, one can observed that it is organized into bundles of fibers call fascicles. The individual muscle fibers, multinucleated cells,contain long slender structures called myofibrils. These are made of myofilaments,which are organized into sarcomeres, the functional unit of contractions.
Both skeletal and cardiac muscle have a striated appearance under a light microscope,due to the organization of the myofilaments.
Each myofibril is composed of thick and thin filaments arranged in a repeating patternalong their length. thick filaments are composed primarily of the protein myosin and thethin filaments are made up the three proteins, troponin, tropomyosin and actin. It is thecyclic binding between myosin heads of the thick filament and actin of the thin filaments,crossbridge formation, that allows of force production or muscle shortening. It should benoted, that there exist other proteins within sarcomere which have recently been shown tohave a role in contractile function, e.g., the elastic protein titan (also known asconnectin).
3. The Motor Unit A single motor unit consistsof one motor neuron and all of the muscle fibers itinnervates. The cell bodies of motor neurons arelocated within the brainstem or spinal cord. Theaxons of these neurons are myelinated and large indiameter, and thus are able to propagate action potentials at high velocities. Once an alpha motor neuron is activated to produce an action potential,all of the fibers innervated by this neuron areactivated and contract simultaneously. Eachmotor unit is made up of one type of musclefibers: i.e., slow twitch, fast-twitch fatigable or fast-twitch fatigue resistant.
NMJ Lecture Notes 4

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 7/35
4. Excitation-Contraction Coupling This refers to the sequence of events by which anaction potential in the plasma membrane of the muscle fiber leads to force production viaan increase in intracellular calcium and crossbridge formation and turn-over. Excitation begins at the neuromuscular junction and then the action potential spreads over thesurface membrane and inward into the fiber via the transverse tubule system
(invaginations of the sarcolemma). This inward excitation activates calcium release fromthe sarcoplasmic reticulum. The calcium then binds to the thin filament and crossbridgeformation occurs.
NMJ Lecture Notes 5

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 8/35
4. The Neuromuscular Junction
Each branch of a motoneuronforms a single junction with amuscle fiber. The myelin sheath
surrounding the motor axon endsnear the surface of the musclefiber and the axon divides into anumber of short processes thatlie embedded in grooves on themuscle-fiber surface. Thisregion of the sarcolemma(muscle membrane) is known asthe motor end plate.Acetylcholine is theneurotransmitter in these
synapses. End-plate potentials(EPPs) can be recorded at themotor end plate when the presynaptic membrane isactivated to release vesicles containing the acetylcholine.
Steps in neuromuscular transmission:
1) nerve action potential.2) calcium entry into the presynaptic terminus.3) release of Ach quanta.4) diffusion of Ach across cleft.5) combination of Ach with post-synaptic receptors and Ach breakdown viaesterase.6) opening of Na+/K + channels (cation channels).7) postsynaptic membrane depolarization (EPP).8) muscle action potential.
NMJ Lecture Notes 6
7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 9/35
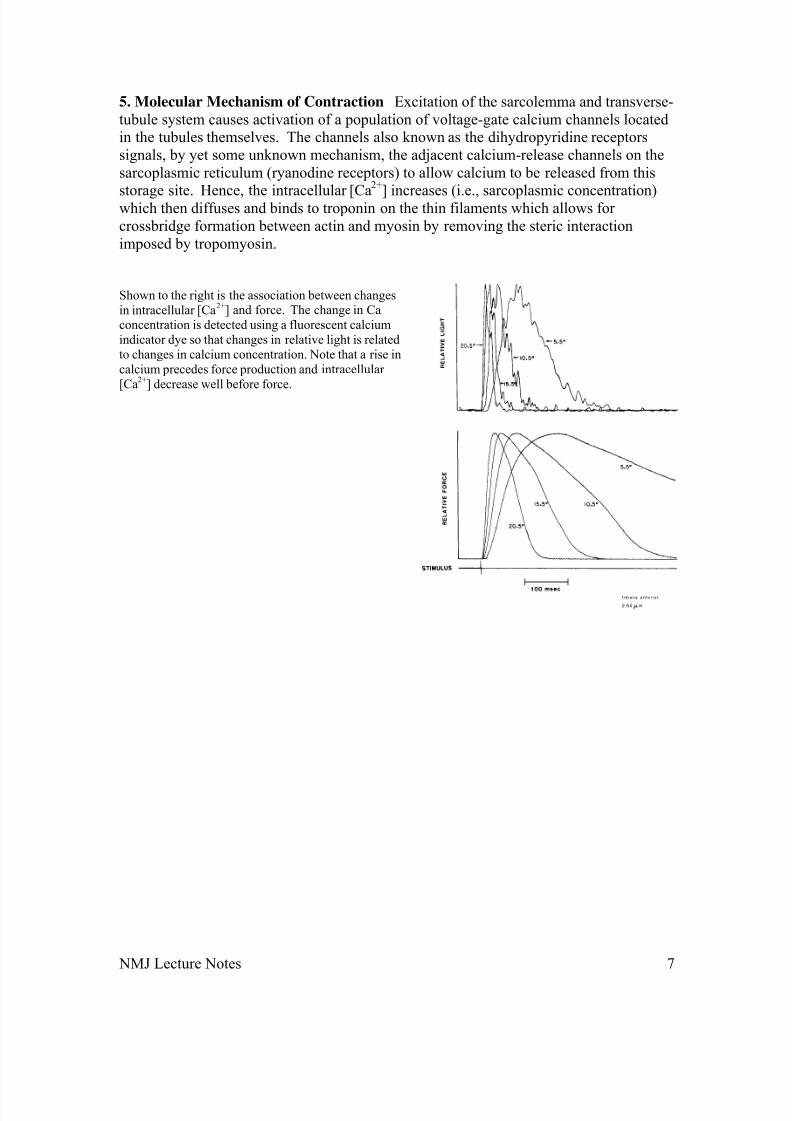
5. Molecular Mechanism of Contraction Excitation of the sarcolemma and transverse-tubule system causes activation of a population of voltage-gate calcium channels locatedin the tubules themselves. The channels also known as the dihydropyridine receptorssignals, by yet some unknown mechanism, the adjacent calcium-release channels on thesarcoplasmic reticulum (ryanodine receptors) to allow calcium to be released from this
storage site. Hence, the intracellular [Ca
2+
] increases (i.e., sarcoplasmic concentration)which then diffuses and binds to troponin on the thin filaments which allows for crossbridge formation between actin and myosin by removing the steric interactionimposed by tropomyosin.
Shown to the right is the association between changesin intracellular [Ca2+] and force. The change in Caconcentration is detected using a fluorescent calciumindicator dye so that changes in relative light is relatedto changes in calcium concentration. Note that a rise incalcium precedes force production and intracellular [Ca2+] decrease well before force.
NMJ Lecture Notes 7

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 10/35
Functional Overview
A neuromuscular junctionsends excitatory signals fromthe CNS via theneurotransmitter,
acetylcholine which binds tonicotinic receptors on the post-synaptic membrane.The binding causes a localchange in the voltage of thesarcolemma affectingneighboring channels (Na+ toenter and eventually K+ toflow out). This ionmovement produces theaction potential which
propagates along thesarcolemma and inward via
the transverse tubule system.This rapid voltage changeinitiates the gating of dihydropyridine receptorswhich in turn causes therelease of calcium from thesarcoplasmic reticulum viathe ryanodine receptors. Thereleased calcium binds totroponin inducing aconformation change intropomyosin, also acomponent of the contractileapparatus, which in turn
allows crossbridge formation between actin and myosin (an energy dependent process). The crossbridgeformation leads to muscle fiber shortening and the generation of force. Crossbridge cycling will proceeduntil calcium dissociates from troponin and the inhibitory influence of tropomyosin is reestablished. Thedissociation occurs because calcium release stops and its active uptake (requiring ATP) into thesarcoplasmic reticulum causes a reduction.
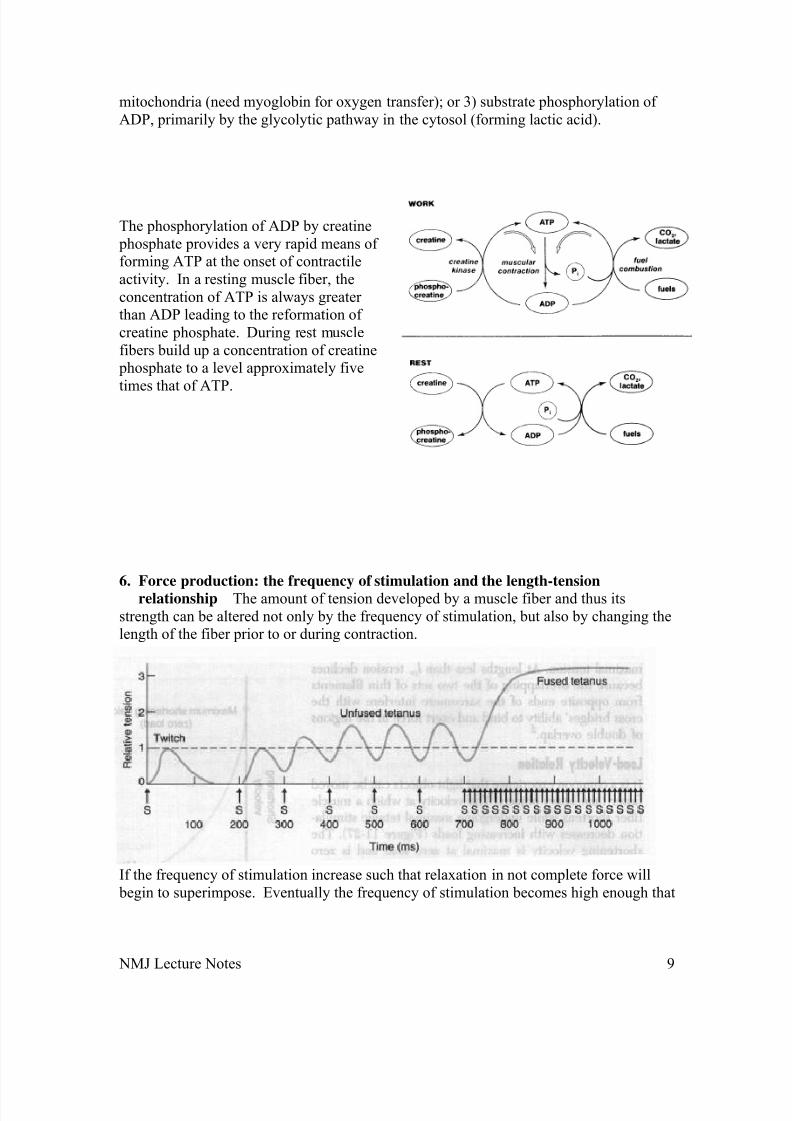
7. Metabolic pathways producing the ATP utilized during muscle contraction
There are three primary ways amuscle fiber canform ATP duringcontractile activity:1) phosphorylationof ADP by creatine phosphate; 2)oxidative phosphorylation of ADP in
NMJ Lecture Notes 8
7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 11/35
mitochondria (need myoglobin for oxygen transfer); or 3) substrate phosphorylation of ADP, primarily by the glycolytic pathway in the cytosol (forming lactic acid).
The phosphorylation of ADP by creatine phosphate provides a very rapid means of forming ATP at the onset of contractileactivity. In a resting muscle fiber, theconcentration of ATP is always greater than ADP leading to the reformation of creatine phosphate. During rest musclefibers build up a concentration of creatine phosphate to a level approximately five
times that of ATP.
6. Force production: the frequency of stimulation and the length-tension
relationship The amount of tension developed by a muscle fiber and thus itsstrength can be altered not only by the frequency of stimulation, but also by changing thelength of the fiber prior to or during contraction.
If the frequency of stimulation increase such that relaxation in not complete force will begin to superimpose. Eventually the frequency of stimulation becomes high enough that
NMJ Lecture Notes 9

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 12/35
the force becomes fused. Further increase in the frequency will cause more force to be produced until eventually a maximum is reached.
If one stretches skeletal or cardiac muscle the magnitude of subsequent contractions will be altered. If the muscle is unloaded, i.e., the sarcomere spacing compressed, there is
little force or shortening that can occur. Skeletal muscle has an optimal length (l0) atwhich force is maximal due to the greatest possible numbers of crossbridges can beformed. Most muscles in the human body are attached so to have near their optimallength at rest.
Because skeletal muscle can shorten allowances need to be made for the sarcolemma toalso conform to this changes without being damaged. Structural proteins are presentwhich link the myofilaments to the surface membrane and extracellular matrix. One of these proteins is dystrophin which is lacking is patients with Muscular Dystrophy.
7. Contraction in Smooth Muscle
This type of muscle lacks cross-striated banding patterns and the nerves which caninnervate it arise from the autonomic nervous system. Nevertheless, smooth muscle alsouses cross-bridge movements between actin and myosin molecules to produce force.
NMJ Lecture Notes 10

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 13/35
Each smooth-muscle fiber is a spindle-shaped cell with a diameter ranging between 2 and10 µm. Smooth muscle cells have only a single nucleus and can continue to divide. Twotypes of filaments are present in the cytoplasm: thick filaments containing myosin andthin composed of actin. The actin filaments are anchored either to the plasma membraneor to cytoplasmic structures known as dense bodies, the smooth muscle equivalent to z-
lines.Tension produced by smoothmuscle alsovaries withlength, but therange of lengthand amount of shortening thatsmooth musclecan achieve is
greater thanskeletal muscle.
The pathwaysleading to anincrease incytoplasmic[Ca2+] and toforce generationdifferssignificantly between smoothand skeletalmuscle.Crossbridges insmooth musclecan form oncemyosin is phosphorylated by a calcium dependent process (enzyme). There are twosources of calcium which leads to an increase in cytoplasmic concentration prior tocontractions: 1) the sarcoplasmic reticulum , and 2) extracellular calcium.
NMJ Lecture Notes 11

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 14/35
Some smooth muscle will generate action potentials, but they differ from skeletal musclein that Ca2+ is the primary ion responsible for the membrane depolarization. Some cellsspontaneously produce action potential and have pacemaker like properties. Other types
of smooth muscle are innervated by nerves. The released neurotransmitter can bereleased onto varicosities along the fibers or in spaces between fibers which then requiresgreater diffusion of the neurotransmitter to the receptor sited. In addition in certainmultiunit smooth muscle arrangements, excitation may be initiated on some cells viainnervation and then transmitted to additional cells via gap junctions between cells.
8. Cardiac Muscle The cardiac-muscle cells of the myocardium are arranged in layersthat are tightly bound together and completely encircle the blood-filled chambers.
Cardiac muscle combines the properties of both skeletal muscle and smooth muscle. The
cells are striated as the result of an arrangement of thick myosin and thin actin filaments.However, cardiac cells are considerably shorter than skeletal muscle fibers and haveseveral branching processes. Adjacent cells are joined end to end at structures calledintercalated disks, within which are desmosomes that hold the cells together and to whichthe myofibrils are attached.
Approximately 1 percent of cardiac-muscle tissue have specialized features that form theconducting system of the heart. They send information to the contractile cells via gap junctions. The conducting system initiates the heartbeat and helps spread the impulserapidly through the heart.
The heart receives a rich supply of sympathetic (norepinephrine) and parasympathetic(acetylcholine) innervation contained in the vagus nerve.
On-Line Muscle References
The excellent chapter on muscle from Vander, Sherman & Luciano's Human Physiologytext, one of the most widely used physiology texts in the world. The chapter from the2004 edition is available online atwww.me.umn.edu/labs/hmd/lab/docs/widmaier_samplech9.pdf
The Dept of Radiology at the University of Washington has an excellent upper and lower extremity Muscle Atlas with images suitable for presentations. The site has a simple andfree copyright form to use the images for academic purposes.www.rad.washington.edu/atlas2/
Medline Plus has an excellent section on muscle disorders.www.nlm.nih.gov/medlineplus/muscledisorders.html
NMJ Lecture Notes 12

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 15/35
2 The Neuromuscular Junction
Each branch of a motoneuron forms a single junction with a muscle fiber. The myelinsheath surrounding the motor axon ends near the surface of the muscle fiber and the axondivides into a number of short processes that lie embedded in grooves on the muscle-fiber surface. This region of the sarcolemma (muscle cell membrane) is known as the motor end plate. Acetylcholine is the neurotransmitter in these synapses. End-plate potentials(EPPs) can be recorded at the motor end plate when the presynaptic membrane isactivated to release vesicles containing the acetylcholine.
1) Nerve and muscle have separate, intact, plasmalemmas, which are separated by a 50nm gap (500Å) known as the synaptic cleft.
NMJ Lecture Notes 13

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 16/35
2) An unmyelinated motoneuron terminal (i.e. presynaptic end of the axon) sits in aspecialized groove of the skeletal muscle fiber to form the neuromuscular end plate.
a) There is only one presynaptic nerve per muscle fiber.
b) Each motoneuron has several ending; each innervates only one muscle fiber.
c) All of the muscle fibers in a given motor unit contract in unison when their motoneuron fires an action potential.
d) All muscle fibers in a motor unit are of the same fiber type (either all-slow or fasttwitch).
3) The junction or end plate region of the skeletal muscle fiber is specialized and
different from the rest of the plasmalemma.a) Synaptic infoldings of the plasmalemma in the cleft greatly increase the
membrane surface area.
b) Receptors (protein molecules) for acetylcholine (ACHR) are located near the cleftedge of the infoldings. In denervated muscle fibers, the ACHRs spread of theentire muscle plasmalemma(sarcolemma).
i) The skeletalneuromuscular junction ACHR isnicotinic sensitivereceptor.
ii) The nicotinicACHR structure iswell characterized(i.e., cloned andsequenced)
iii) In its proteinmoiety, the ACHR contains:(1) Binding sites for
acetylcholine(ACH) and likemolecules (agonists and antagonists).
(2) Ligand-gated cation channel(3) Several types of modulator sites
NMJ Lecture Notes 14

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 17/35
c) The are 5 subunits of the ACHR: 2 alpha, beta, gamma and epsilon.
d) The channel and the ACH binding sites are on the alpha subunits.
e) The ACHR is a non-specific cation channel which opens and closes in response toACH binding and unbinding (insensitive to TTX or TEA)
i) In the presence of an elevated [ACH] in the cleft ACH binds to theextracellular side of the receptor and the channel opens.
FastReaction 2[A] + [R] < -- > [AR] + [A] < -- > A2R < -- > A2R*Channel closed closed closed openStates:
↑ ↓
AR A2R Desensitized ← desensitized
ii) The binding of the 2 ACH molecules to a single receptor elicits positivecooperativity.
iii) When the ACHR channel opens at a normal muscle fiber restingmembrane potential, Em =90 mV, the net current through the channel isinward and depolarizing.
iv) ACH unbinds from the ACHR after the channel closes when the [ACH] in
the endplate decreases due to diffusion from the cleft and is broken down byan acetylcholinesterase (ACHE). Prolonged ACH (> 100 msec) stimulationleads to inactivation of the channels through a change to the desensitized state.
v) Then the end plate channel opens, net current (I) from all cations ininward: this positive charge (q+) movement through the myoplasm to thesurrounding sarcolemma cause a capacitate change (depolarization) whichthen affects the gating of the voltage-sensitive Na+ and K+ channels.
vi) ACHRs can be irreversibly blocked by the snake poison
vii) Curare block binding of ACH to its receptors. Curare is a non-activatingor non-depolarizing block.
viii) Nicotine acts at the NMJ and binds to the ACHR.
ix) There are substances, which mimic ACH, but are not readily broken down by ACHE, thus that cause and initial opening of the channel and then
NMJ Lecture Notes 15

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 18/35
inactivation through desensitization. Succinylcholine is one of thesedepolarizing muscle relaxants.
4) The nerve terminal has vesicles (50 nm in diameter) containing ACH which fuse with
the plasmalemma and release ACH into the cleft after the nerve AP depolarizes themembrane and Ca2+ enters through channels in the nerve terminal.
a) Formation of ACH in nerve terminal.
Acetyl TransferaseAcetylCoA + Choline ------------------------ > ACH + CoA
b) ACH is stored in vesicles in the nerve terminal.
c) Quantum: Smallest amount of ACH released. Probably the amount of ACH in a“standard” presynaptic vesicle is: Quantum = 2,000 to 10,000 ACH molecules.
5) The NMJ cleft is filled with extracellular fluid and ground substance, which alsocontains the enzyme acetylcholinesterase (ACHE).
ACHEACH -------------- > Acetate + Choline
6) ACHE acts only on unbound ACH. Acetate and choline are transported back into thenerve terminal. Acetate is converted to acetylCoA (in mitochondria) and thencombines with choline to reform ACH.
a) Organophosphates inhibit ACHE and thus prolong ACH lifetime. Inhibitors of ACHE are: physostigmine and neostigmine, which are used clinically to reverseneuromuscular blockage.
Characteristics of ACH release:
1) As a result of AP depolarization, Ca2+ enters the nerve terminal through a voltage-gated channel.
a) 4 Ca2+ act cooperatively to release one quantum.
b) Reducing extracellular Ca2+ reduces ACH release.
NMJ Lecture Notes 16

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 19/35
c) Mg2+ competes with Ca2+ and does not active ACH release; increasingextracellular Mg2+ decreases ACH release.
2) One nerve AP causes the release of approximately 300 quanta (vesicles)
1 AP ------ > 300 quanta released ----------- > 1,500,000 ACh molecules
(assuming 5,000 ACH/quantum) Some of this ACH diffuses out from the cleft andsome is broken down by ACHE: approximately 200,000 molecules bind to ACHR toopen channels in the endplates.
3) Factors which alter or block nerve APs will alter ACH release:
a) Local anesthetics (e.g., procaine) inhibit voltage-gated Na+ channels and interfere
with AP transmission in the nerve. Some local anesthetics also act on the ACHR by promoting desensitization and/or by blocking the channel. b) An increase [K]o causes prolonged depolarization of the nerve and thus partial
inactivation of the voltage-gated Na+ channels, thus alters AP transmission.
Hemicholinium inhibits uptake of choline into the nerve terminus and thus decreasesACH production and storage. The result is decreases ACH/quantum.Botulinum toxins block the release of ACH from the nerve terminals (i.e., paralysis from bad tuns).
Characteristics of the End Plate Potential (EPP):
1) Opening of the end plate channels and the subsequent net inward current sets up atransient depolarization of the sarcolemma adjacent to the end plate.
a) The end plate potential (EPP) spreads electrotonically and thus decrements inamplitude with distance from the end plate region. The EPP itself in not propagated but serves as the stimulus to drive the Em to threshold for an AP to beinitiated. The EPP can be seen in isolation of an AP by treating the muscle fiberswith tetrodotoxin (TTX) which blocks the voltage-gated Na+ channels.
b) The EPP brings the adjacent sarcolemma to and beyond threshold: the voltage-gated channels in the non-endplate region of the membrane are then responsiblefor the propagation of the AP throughout the length of the muscle fiber.
c) EPPs last approximately 5-10 msec.
d) Normally: 1 nerve AP -- > 1 EPP -- > 1 muscle AP -- > a single twitch .
NMJ Lecture Notes 17

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 20/35
e) The typical EPP amplitude is –30 mV, which represents current throughapproximately 100,000 open ACH endplate channels.
NMJ Lecture Notes 18
7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 21/35
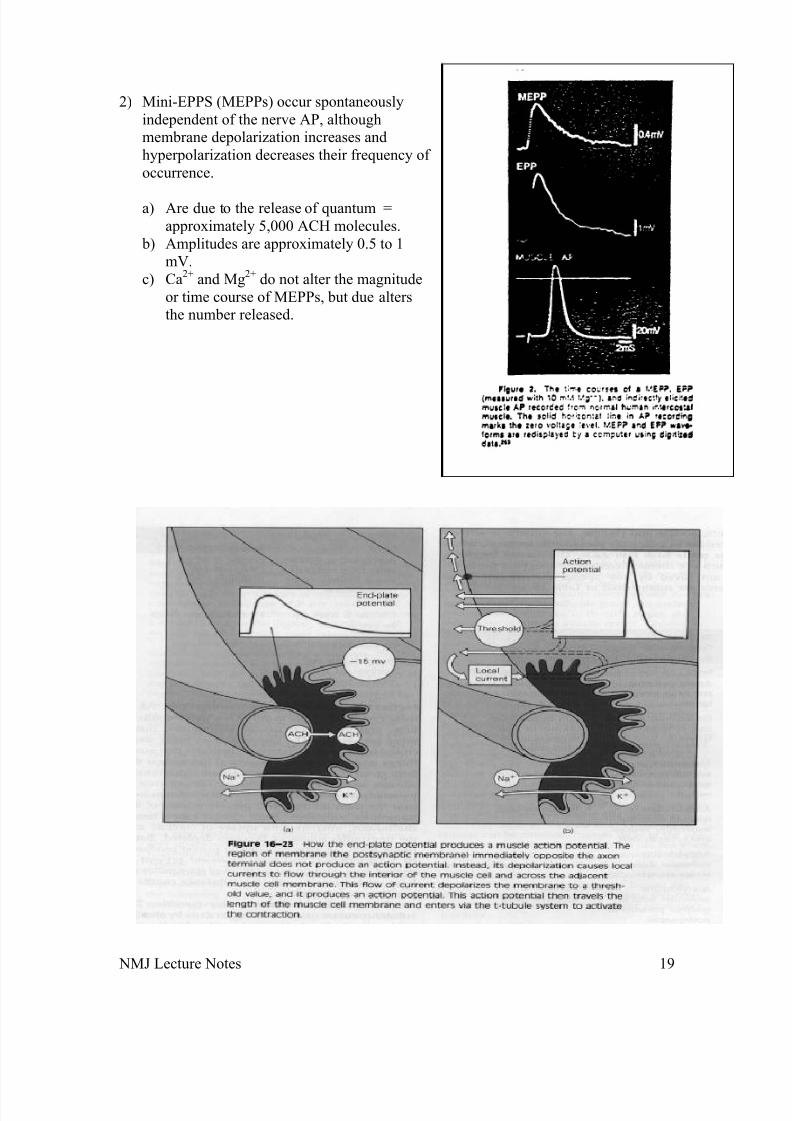
2) Mini-EPPS (MEPPs) occur spontaneouslyindependent of the nerve AP, althoughmembrane depolarization increases andhyperpolarization decreases their frequency of occurrence.
a) Are due to the release of quantum =approximately 5,000 ACH molecules.
b) Amplitudes are approximately 0.5 to 1mV.
c) Ca2+ and Mg2+ do not alter the magnitudeor time course of MEPPs, but due altersthe number released.
NMJ Lecture Notes 19

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 22/35
Review: Steps in neuromuscular transmission:
1) nerve action potential.2) calcium entry into the presynaptic terminus.3) release of Ach quanta.
4) diffusion of Ach across cleft.5) combination of Ach with post-synaptic receptors and Ach breakdown viaesterase.6) opening of Na+/K + channels (cation channels).7) postsynaptic membrane depolarization (EPP).8) muscle action potential.
NMJ Lecture Notes 20

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 23/35
3 Membrane Resting Potential
The nervous system uses electrical signals to communicate over relatively enormous'biological distances'. It does so with speed and accuracy, and deals in a vast traffic of signals distributed to millions of cells. Present-day emphasis on the nerve membraneand the resting potential is far more than a political dogma to bedevil the student. For inunderstanding these processes, we glimpse the basic strategy used by neurons to carry-ontheir mission of information processing. This strategy--sometimes loosely called theionic hypothesis - appears to be common to all nerve cells, and to be a specialization of the general phenomenon of irritability present in all cells from the dawn of life.
Our knowledge of the, subject is by no means complete, but sufficient elements have
been assembled to allow the conclusion that this strategy itself is a remarkably simpleone, but also elegant in that there is a capacity for many individual variations on acommon theme. The necessary elements appear to be: 1) the presence of an ion-selective, semi-permeable membrane; 2) the ability of the cell to concentrate differentamounts of sodium, potassium and chloride across the membrane, 3) passive physicalforces of diffusion and electrical gradients; and 4) an active process (principally operatingon sodium and potassium ions), that aids in maintaining the various ionic concentration profiles.
1) The Nerve Membrane. The nerve membrane has been extolled to you in biochemistry, biology and physiology, so we will not dwell in detail on it here. Suffice it
to say that there exists a membrane composed basically of a bi-layer of lipid leafletsembedded in which at various distances are proteins. Many of these proteins have water permeable pores or channels in them which allow one or more ion species through. Someof these channel-bearing proteins are "gated" (can open and close as the result of something that forces a change in their structure), some are theorized to be passive andalways open. These latter channels are what are called “resting channels", and we willconcentrate on these, as it is thought they are responsible for the resting permeability of the membrane and therefore for the resting potential. The "gated" channels operate to produce changes in the membrane potential and therefore mediate informationexchanges.1
In the absence of any active -information processing, the nerve membrane maintains a potential difference between the inside of the cell and outside of some 70 to 90 millivolts,with the inside negative (1 mV = 10-3 volts), and this voltage difference is called the
1 No one has ever "seen" a resting channel, and there are various alternative explanations that could accountfor the passive, resting permeability of the membrane. For instance, gated channels that flicker open andclosed on a probabilistic basis could account for much of the resting permeability However, for presentdidactic reasons, it is easier to just theorize that resting channels exist.
NMJ Lecture Notes 21
7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 24/35
resting potential. 70 millivolts may seem like a very small electrical force compared tothe 120 volts of city power you may have painfully sampled on occasion, but this 70 mVis acting across a correspondingly small distance. To obtain a "feel" for the actual forceone has to look at the ratio of the resting potential to the thickness of the membrane. 70mV across an approximately 100 membrane corresponds to a force of 70,000
volts/centimeter. This is more than adequate to make proteins with charged groupsimbedded in the membrane stand-up and salute!2 Indeed, such a powerful electrical fieldcan and does regulate the three- dimensional configuration of some of the gatedmembrane protein channels ("voltage gated channels") causing them to open and/or closedepending on voltage changes that occur across the membrane.
2) Ionic Profiles. The typical mammalian extracellular solution contains about 120 mM(millimoles) of sodium, 4 mM of potassium and 124 mM of chloride ions. Inside thecell, the relative concentrations are almost reversed. We find about 12 mm of sodium,110 mM of potassium and 9 mM of chloride ions. There are sufficient, large,impermeable anions (bicarbonate, glutamate, aspartate and organo-phosphates and -COO-
groups of cellular proteins) to bring the sum total of negative charges inside about equalto the number of negative ions outside. Thus from a macroscopic view, the inside andoutside contain an almost equal number of both positive and negative ions. The table below sums up the ion concentrations:
Inside Outside Ratio of outside/inside
Sodium 12 mM 120 mM 10Potassium 110 mM 4mM 0.0363Chloride 9 mM 124 mM 13.78"A” ~(113 mM) -- --
From a macroscopic view, then, the inside and outside contain an almost equal number of both positive and negative ions. There must exist, however, a small imbalance of chargeimmediately across the membrane or there would be no resting potential. It should beemphasized though, that the required amount of excess negative charge inside, separatedfrom the outside by less than 100 Angstroms, is almost immeasurably small. In fact, theability of small amounts of charge to produce significant voltage changes across thenerve membrane is one of the great utilities of the system: small amounts of charge
migrating the small distance across the membrane introduce very rapid, precise
changes in voltage
3) Passive Physical Forces of Diffusion and Voltage: The Nernst Fquation.Given the ionic profiles, we are faced with explaining how they lead to the resting potential. As a first step, consider what the situation would be if the membrane were permeable only to one ion species (had only one kind of protein channel that was permeable to only one kind of ion). Taking potassium (K +) first, the relatively large
2 1 Ả = 10-8 cm., 100 Ả = 10-6 cm. To scale the membrane and voltage to recognizable dimensions,
multiply both the membrane thickness and the voltage by 106. This gives I cm. for the membrane now, and70 x 10-3 xIO6 =7o x 103 volts = 70,000 volts for the scaled electric field.
NMJ Lecture Notes 22

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 25/35
concentration of the K+ ions on the inside should cause K + ions to diffuse outwardtoward the more dilute solution of K + ions outside. Every ion of K + that diffuses acrossthe membrane will carry a positive charge with it, leaving behind an excess of negativecharge. It is just this separation of charges which will produce an electric field, or potential. Moreover, increasing the number of charges which are separated, will tend to
impede subsequent, positively charged K+ ions from diffusing outward. Indeed, it can beshown that an equilibrium will soon be reached such that the electrical field will exactlyoppose the force of diffusion! In this state, as many potassium ions are attracted acrossthe membrane from outside to inside by excess negative charges, as are shoved frominside to outside by the force of diffusion. The quantitative expression of this balance isthe Nernst Equation for the single ion species.
⎟⎟ ⎠
⎞⎜⎜⎝
⎛ =⎟⎟
⎠
⎞⎜⎜⎝
⎛ =
i
o
i
o
C
C
zF
RT
C
C
zF
RT E log3.2ln
where Co = outside concentration; Ci = inside concentration; z = the charge(valence) of the ion; and 2.3 RT/F = 61.5 mV, at 37°C.
Substituting in the concentration values for potassium, we get:
( ) mV xK
K E
i
oK 9044.15.61)04.0log(5.61
110
4log5.61log5.61 =−=+=⎟
⎠
⎞⎜⎝
⎛ +=⎟⎟
⎠
⎞⎜⎜⎝
⎛ =
EK = -90 mV if the membrane were permeable only to potassium ions. In other words, avoltage difference of -90 mV would exactly oppose the diffusion force for the listed ionicconcentrations of potassium across the nerve cell membrane. This is called the
equilibrium potential for potassium. Looking at chloride, we obtain3
mV Cl
Cl
Cl
Cl E
o
i
i
oCl 70log5.61log5.61 −=⎟⎟
⎠
⎞⎜⎜⎝
⎛ +=⎟⎟
⎠
⎞⎜⎜⎝
⎛ −=
This would be the value of the membrane potential if the membrane were permeable onlyto chloride ions. For sodium, the equilibrium potential is
mV Na
Na E
i
o Na 5.61)10log(5.61
12
120log5.61log5.61 +=+=⎟
⎠
⎞⎜⎝
⎛ +=⎟⎟
⎠
⎞⎜⎜⎝
⎛ =
That is, the membrane potential would be 61.5 millivolts, inside positive, if themembrane were permeable only to sodium ions.
3 Note that multiplying a log ratio by –1 simply reverses the position of the numerator and denominator.
This is an algebraic trick employed to keep minus signs out of textbook equations. Its end result here isthat the equilibrium expression for chloride has the inside and outside concentrations reversed whencompared with K + and Na+.
NMJ Lecture Notes 23

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 26/35
Looking at the values obtained we can draw a number of conclusions about the actualstate of affairs. First, the actual resting potential of -70 mV agrees well with the chlorideequilibrium potential. This implies that chloride is distributed passively across themembrane. If we artificially pass current into the cell to change the resting potential,
chloride concentrations will also change (over a period of time) such that they will balance out the right hand side of the Nernst equation to equal the artificially imposedresting potential. Nothing is acting on chloride other than the forces of diffusion andvoltage. Second, neither potassium nor sodium appear to be in equilibrium. We would predict that with a resting potential of -70 mV and the given concentrations of Na and K,a small amount of potassium should tend to diffuse out of the cell and a much larger amount of Na should diffuse into the cell if both ions were equally permeable to themembrane. In fact, potassium permeates the resting cell membrane about 50 times morereadily than sodium, and about an equal amount of potassium tends to diffuse out assodium goes in. Our third, conclusion is that given the figures on sodium and potassium,some force other than the passive forces of diffusion and electrical field is necessary to
maintain the measured quantity of potassium so high (and that of sodium so low) insidethe cell. This is where the role of the sodium-potassium 'pump' (sometimes just called the'sodium pump") figures in. Active transport in terms of the ATP-dependent sodium-
potassium 'pump' is constantly at work to maintain the K+ and Na+ concentrations
at their stated values. This is homeostasis in action.
The Na-K active transport may be considered as a background process in that the pumpcranks along, using cellular energy to maintain the K+ and Na+ gradients constant. If something happens to transiently change the leakage rates of these two ions, it is knownthat the pump will speed up a little bit or slow down a little bit to catch things back-upwithin a few seconds or minutes, thereby maintaining the desired gradients. Indeed, theATP-ase activity of the sodium-potassium coupled transport is one of the principlehomeostatic mechanisms of most cell membranes studied. Present studies are movingcloser to physically characterizing the structure and exact function of this system.One other thing to note: by exchanging one Na+ for one K+ ion, the pump is "electricallyneutral"; i.e., it does not change the charge concentration across the membrane, andtherefore it does not contribute directly to either the resting potential or to changes in theresting potential. It works indirectly by maintaining the differential ion concentrations,which in turn exert their passive forces as expressed in the Nernst equation above and inthe Goldman- Hodgkin-Katz equation explained below.4
4
There are actually a number of different, generic Na-K pumps and for most of them, the exchangeratios are not exactly 1:1. In these cases, the pumps are considered "electrogenic" in that they do contributedirectly to the resting potential via unequal pumping ratios, but they still continue to contribute indirectly aswell by maintaining ionic gradients. The electrogenic effects of all such pumps studied are actually minor ones, however, and they contribute only a few mV of potential. Much is made of them in many recenttexts, unfortunately, as they are a more recent and "hot" research discovery topic, and authors of textbookstend to overplay new things. Students should be as astute as we are, however, and realize that this aspect,while exciting to study in the research laboratory, should not detract from the basic concept and utility of the over-all homeostatic process, which is to maintain the constant ionic concentration gradients across themembrane.
NMJ Lecture Notes 24

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 27/35
The Goldman-Hodgkin-Katz Equation. We are still left with the question, why 70millivolts for the resting potential? A clue to the answer lies in realizing that the cellmembrane is not permeable to just one ion species but is instead permeable to all threemajor ions, Na, K and Cl. We must quantitatively account for the relative permeabilitiesof these ions in order to understand why -70 mV, and to do that we have to go beyond the
Nernst equation. If one were to stir a small quantity of oranges and a large quantity of grapefruit on a platform with many holes the size of oranges and only a very few largeenough for grapefruit, the net result would be that more oranges than grapefruit wouldfall through. The same principle holds for relative permeability of ions. The relative permeability of the membrane to the various charged ions will determine which ionspecies is most important in carrying charges across the membrane, and therefore whichof the ion species will have the greatest influence on 'setting' the resting membrane potential.
The mathematics of the diffusion process gets complicated in so doing, but the three mennamed above were able, over about 10 years time of independent and of collaborative
effort, to mold a relatively simple expression which considers both concentrations and permeabilities. This is the GHK Equation, expressed as
⎥⎦
⎤⎢⎣
⎡
++
++=
oCli Naik
iClo Naok
ClP NaPK P
ClP NaPK P
F
RT E ln
where:
PK = 1 x 10-6 (membrane permeability of potassium compared to free diffusion)PCl = 1 x 10-6 (membrane permeability of chloride compared to free diffusion)P Na = 2 x 10-8 (membrane permeability of sodium compared to free diffusion)
Note that the Goldman-Hodgkin-Katz equation gives a potential for the steady-statecondition where as many plus charges are flowing out across the membrane as areflowing in. The same is true for minus charges. Only chloride is in thermodynamicequilibrium, and the resting potential is a steady-state potential defined as no net flow of charge. It is assumed, of course, that the concentration profiles remain constant due tothe active transport of Na and K by the sodium- potassium pump. In fact, membrane permeabilities of both K and Na are so low that a nerve axon can be metabolically poisoned to block active transport, and the concentration profiles will change measurablyonly after many minutes or hours (depending on the diameter and volume of the fiber).
While not dwelling excessively on mathematics, it is instructive to play a few algebraicgames with the G-H-K equation to illustrate the relative importance of the terms withinthe brackets. Divide all terms upstairs and down by PK . (This does not change theequation, as PK /PK = 1, but it allows for rearrangement of terms).
NMJ Lecture Notes 25

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 28/35
⎥⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢⎢
⎣
⎡
⎟⎟ ⎠
⎞⎜⎜⎝
⎛ +⎟⎟
⎠
⎞⎜⎜⎝
⎛ +
⎟⎟ ⎠
⎞⎜⎜⎝
⎛ +⎟⎟
⎠
⎞⎜⎜⎝
⎛ +
=
o
K
Cl
i
K
Na
i
i
K
Cl
o
K
Na
o
ClP
P Na
P
PK
ClP
P Na
P
PK
F
RT E ln
Now, disregard the terms with chloride, since we know that chloride will passivelydistribute its concentration to agree with whatever K and Na determine. (You might trycalculating E with and without deleting the chloride term to convince yourself of this).Finally, if you get tired of writing PNa/PK, let this quantity be b, substitute it in, and weobtain:
⎥⎦
⎤⎢⎣
⎡
+
+=⎥
⎦
⎤⎢⎣
⎡
+
+=
ii
oo
ii
oo
NaK
NaK
NaK
NaK
F
RT E
b
b
b
blog5.61ln
Note that in the resting state, b is small because P Na is about 50 times smaller than PK which divides it in the term. If you substitute in the concentrations, you will see that the potassium concentration dominates (because Na terms are multiplied by about .02). Weexpect, and we do find, that changing extracellular K concentration will greatly affect theresting membrane potential while changing extracellular Na causes little change.
The Strategy.One last feature remains to be considered, and herein lies the insight into the basicstrategy employed by the neuron in affecting voltage changes as its information signalingmechanism. Since Na and K are, in fact, not in thermodynamic equilibrium, and becausethe cell will maintain the ionic profiles cited above, a small change in the membrane's
permeability to either K or Na will cause the trans-membrane potential to change. Theresting state is, then, a true source of potential energy. Increase sodium permeability for a fraction of a second, and the cell will depolarize (become less negative inside or less polarized) towards the sodium equilibrium potential. Increase PK and the membrane willhyperpolarize (become more negative or more polarized) towards the potassiumequilibrium potential. Few ions need to flow (and these will generally be handled over the long haul by active transport). The concentrations remain effectively the same, butthe membrane potential will fluctuate rapidly towards the equilibrium potential of whatever ion appears to dominate in the equation. Figure 1 at the end of this essaysummarizes the situation.
The change in permeability may be brought about by chemicals acting on chemicallygated channels in the membrane (as it is in synaptic transmission), or it may be thatforced changes in the electrical field across the membrane may trigger a voltage gatedchannel (as is the case for the action potential). The permeabilities of both Na and K mayeither increase (by opening gated channels) or decrease (by blocking resting channels),they may do so simultaneously or in sequence, as there are a large number of different,chemically gated and voltage gated channels. Other ions (chloride for instance, calciumin cases such as in cardiac muscle) may also figure into the permeability changes, but
NMJ Lecture Notes 26

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 29/35
they do so in the same manner as sodium and potassium. In other words, the equationcan be extended within the brackets to incorporate other ion species, and while it makesthe equation more complicated to look at, the strategy by which membrane potential ischanged remains the same.
What isn't known is all the physical details of the different types of channels. We doknow that the major ion species all have membrane proteins which are pretty muchselective to each of them, and this makes common sense in considering how exquisiteand important is the control over permeabilities. Moreover, there are some channels thatare equally permeable to both Na and K (try putting b = 1 into the GHK equation and predicting what direction the membrane potential will go when this type of gated channelopens up). We also know that the permeability sensitivity of neuronal membranes tovarious chemicals or to changes in electrical potential varies from cell to cell (i.e.,different cells have different kinds of gated channels) and indeed, varies across thedifferent parts of each neuron. Thus different drugs may affect different neurons or may block permeability changes on only a specific part of the neuron because of the drug's
effect on a specific channel.Modern research now accepts the overall concepts given here, and the exciting newresearch emphasis is exactly concentrating on understanding the proteins that make upthe gated channels. Considerable progress has been made on a few of these: the sodiumion channel that is voltage gated and responsible for action potentials in nerve axons has been isolated, cloned and sequenced; the acetylcholine receptor of the neuromuscular junction, which contains a large channel that lets both Na and K through simultaneously,has been isolated, purified, and sequenced; in addition numerous antibodies that bind tospecific parts of this protein have been made and used to help us understand its three-dimensional structure. Other channels have been sufficiently isolated to allow for their placement into "artificial" membranes in such a way as to make study of them easier.Understanding these channels better should lead to a more complete understanding of membrane. As pathologies of some channels lead to specific disease syndromes,understanding their structure should eventually lead to better clinical control of thediseases. As an example, auto immune attack on the above mentioned acetylcholinereceptor leads to the disease called myasthenia gravis, and you will hear more about thislater in the course. As the molecular biology of this receptor gets better understood,means for curing this disease becomes increasingly probable. That may be as it 'is, andone can get very excited about the research. What is just as exciting, however, is that wenow understand the basic functional concepts underlying the membrane potential andhow it is controlled. As explained above, it really isn't all that complicated, but it is anelegant system in its purity, efficiency and in the number of combinations and permutations that can lead from it. Indeed, understanding the membrane potential andthe natural strategy for controlling it through permeability changes really reduces muchthe rest of basic neurophysiology to practical examples, and that should make any studentof this subject happy!
NMJ Lecture Notes 27

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 30/35
-100
-90
-80
-70-60
-50
-40
-30
-20
-10
0
10
20
30
40
50
60
70 61.5 mV
EK
ENa
Resting (PNa/PK ) = 0.02
Hyperpolarization < 0.02
Depolarization > 0.02
Potential (mV)
Figure 1. The membrane potential depends on its relative permeability to ions.
Permeability, and thus the membrane voltage, changes as voltage-dependentor chemically-dependent gated channels open and close. At the resting potential, P Na/PK = 0.02. When the ratio becomes larger than 0.02 (i.e.increases or decreases), depolarization results. When the ratio becomes lessthan 0.02 ( decreases or increases), hyperpolarization results. The twoequilibrium potentials, E = 61.5 mV and E = -90 mV, set the upper andlower limits of the possible potential changes.
NMJ Lecture Notes 28

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 31/35
4 NMJ References
If you want to read more about neuromuscular physiology, try these references.
Vander A, Sherman J, Luciano D (2001). Human Physiology: The Mechanisms of Body
Function, 8th ed. McGraw-Hill.
An excellent overview of human physiology. Covers everything, but has nice sections on nerveand muscle. Used in the core undergrad intro to physiology courses (PHYSL 3051, 6051) at theUniversity of Minnesota.
Kandel E, Schwartz J, Jessell T (2000). Principles of Neural Science, 4th ed. McGraw-Hill.
The bible of neurosciences. Excellent chapters on nerve and the NMJ. Every student of theneurosciences should own this book.
Hodgkin A (1992). Chance and Design. Cambridge University Press.
Short autobiography of one author of the Hodgkin-Huxley equations. A wonderful book about hiswork, starting as a student, which elucidated the ionic basis of neuronal and muscle excitability.
Koch, C (1999). Biophysics of Computation. Oxford University Press.
Reference for biophysics of neurons. Chapters 1-4, 6 and 8 of particular interest for this week.
Hille, B (2001) Ion Channels of Excitable Membranes. 3rd Ed. Sinauer, Sunderland ,Mass.
Standard textbook and reference on ion channels.
Oakley B, Schafer R (1978) Experimental Neurobiology: A Laboratory Manual,University of Michigan Press.
One of the only step-by-step guides on methods of basic neuroscience experiments. Out of print. If you get lucky, you might find one in a used book stores.
Loeb G, Gans C (1986). Electromyography for Experimentalists. University of ChicagoPress.
Excellent coverage of experimental equipment for neurosciences, including how to build your own. The book is a little old, but most of the information is still valid.
Adrian RH (1956) The effect of internal and external potassium concentration on themembrane potential of frog muscle. J Physiol 133:631-658.
NMJ Lecture Notes 29

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 32/35
The classic paper on this topic. Included in the readings section of these lecture notes.
Magleby KI (1984) Neuromuscular transmission. In: The Anatomy, Physiology, and
Biochemistry of Muscle. Chapter 13, pp. 393-418.
Coverage of the NMJ, classic experiments on recording EPPS and MEPPs. Included in the
readings section of these lecture notes.
Matthews, G.G. (1998). Cellular Physiology of Nerve and Muscle. 3rd ed. BlackwellScience.
Excellent overview of the principles at work in excitable cells. Textbook used for Univ of Minn physiology courses.
Aidley, D. (1998). The Physiology of Excitable Cells. 4th ed. Cambridge Univ. Press
Excellent overview of the principles at work in nerve and muscle cells.
NMJ Lecture Notes 30

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 33/35
5 Readings
This section contains primary source material that should be read prior to the course.
Contents
Adrian RH (1956) The effect of internal and external potassium concentration on themembrane potential of frog muscle. J Physiol 133:631-658.
A classic paper on how potassium concentration changes change the resting potential of amembrane. You will be doing a similar experiment in this course.
Magleby KL (1984) Neuromuscular transmission. In: The Anatomy, Physiology, and
Biochemistry of Muscle. Chapter 13, pp. 393-418.
Coverage of the NMJ, classic experiments on recording EPPS and MEPPs. Relevant to allmicroelectrode recording experiments you will do this week. Included in the readings section of these lecture notes.
Engle AE (1994). Congenital myasthenic syndromes. In Neurologic Clinics of North
America, 12(2):401-437.
Overview of NMJ disorders. You only need to skim this one.
Durfee, W.K. and P.A. Iaizzo. Rehabilitation and muscle testing. In: Encyclopedia of Medical Devices and Instrumentation, 2nd ed . J.G. Webster, ed., Vol 6, pp 62-71,Hoboken, John Wiley & Sons, 2006.
Review of clinical human muscle force testing. (available on-line atwww.me.umn.edu/~wkdurfee/publications/wiley-chap-2006.pdf
NMJ Lecture Notes 31

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 34/35

7/28/2019 Sodium Pump
http://slidepdf.com/reader/full/sodium-pump 35/35
**Note to handout assembler: Replace this page
with the following articles:
Adrian RH (1956) The effect of internal and external potassium concentration onthe membrane potential of frog muscle. J Physiol 133:631-658.
Magleby KL (1984) Neuromuscular transmission. In: The Anatomy, Physiology,and Biochemistry of Muscle. Chapter 13, pp. 393-418.
Engle AE (1994). Congenital myasthenic syndromes. In Neurologic Clinics of North America, 12(2):401-437.
Durfee, W.K. and P.A. Iaizzo. Rehabilitation and muscle testing. In: Encyclopediaof Medical Devices and Instrumentation, 2nd ed . J .G. Webster, ed., Vol 6,pp 62-71, Hoboken, J ohn Wiley & Sons, 2006.
(available on-line atwww.me.umn.edu/~wkdurfee/publications/wiley-chap-2006.pdf )
Related Documents











