© 2012 Landes Bioscience. Small RNAs of the Bradyrhizobium/ Rhodopseudomonas lineage and their analysis Ramakanth Madhugiri, 1 Gabriella Pessi, 2 Björn Voss, 3 Julia Hahn, 1 Cynthia M. Sharma, 4 Richard Reinhardt, 5 Jörg Vogel, 4 Wolfgang R. Hess, 3 Hans-Martin Fischer 2 and Elena Evguenieva-Hackenberg 1, * 1 Institute of Microbiology and Molecular Biology; Gießen, Germany; 2 ETH, Institute of Microbiology; Zürich, Switzerland; 3 Institute of Biology III; Freiburg, Germany; 4 Institute of Molecular Infection Biology; Research Center for Infectious Diseases; Würzburg, Germany; 5 Max Planck Institute for Plant Breeding Research; Cologne, Germany Keywords: Bradyrhizobium, Rhodopseudomonas, small RNA, non-coding RNA, nodule, symbiosis, target, prediction, microarray, dRNA-seq Abbreviations: BjrC, Bradyrhizobium japonicum small RNA candidate; dRNA-seq, differential RNA-seq; nt, nucleotides; ORF, open reading frame; sRNA, small RNA; TSS, transcriptional start site, 5'-UTR, 5' untranslated region Small RNAs (sRNAs) play a pivotal role in bacterial gene regulation. However, the sRNAs of the vast majority of bacteria with sequenced genomes still remain unknown since sRNA genes are usually difficult to recognize and thus not annotated. Here, expression of seven sRNAs (BjrC2a, BjrC2b, BjrC2c, BjrC68, BjrC80, BjrC174 and BjrC1505) predicted by genome comparison of Bradyrhizobium and Rhodopseudomonas members, was verified by RNA gel blot hybridization, microarray and deep sequencing analyses of RNA from the soybean symbiont Bradyrhizobium japonicum USDA 110. BjrC2a, BjrC2b and BjrC2c belong to the RNA family RF00519, while the other sRNAs are novel. For some of the sRNAs we observed expression differences between free-living bacteria and bacteroids in root nodules. The amount of BjrC1505 was decreased in nodules. By contrast, the amount of BjrC2a, BjrC68, BjrC80, BjrC174 and the previously described 6S RNA was increased in nodules, and accumulation of truncated forms of these sRNAs was observed. Comparative genomics and deep sequencing suggest that BjrC2a is an antisense RNA regulating the expression of inositol-monophosphatase. The analyzed sRNAs show a different degree of conservation in Rhizobiales, and expression of homologs of BjrC2, BjrC68, BjrC1505, and 6S RNA was confirmed in the free-living purple bacterium Rhodopseudomonas palustris 5D. Introduction Small RNAs (sRNAs) have been identified as essential elements of numerous regulatory networks in pro- and eukaryotic cells. Extensive searches for sRNAs in Escherichia coli and in few other bacteria demonstrated the expression of dozens to hundreds of sRNAs, most of them with still unknown function. 1-7 However, despite the availability of more than 1,000 bacterial genome sequences, the genes for sRNAs are usually difficult to recognize and, thus, are not annotated. This is mainly due to the high sequence divergence of sRNAs even in related bacteria, but also because of the difficulties to recognize promoters of genes, which lack associated open reading frames (ORFs). A particularly useful method for detection of sRNAs is their bioinformatic prediction by genome comparison of closely related bacteria, followed by experimental verification. With this approach, genes for sRNAs which are located between annotated ORFs are predicted based on sequence and/or secondary structure conservation. 8-10 We are interested in sRNAs of the Bradyrhizobium/Rhodopseudo- monas lineage of a-proteobacteria, which comprises intensively studied model organisms like Bradyrhizobium japonicum USDA 110, Bradyrhizobium sp strain BTAi1 and Rhodopseudomonas palustris. The purple non-sulfur phototrophic R. palustris is metabolically highly versatile and can perform oxidative phos- phorylation, photophosphorylation, carbon dioxide fixation and nitrogen fixation as a free-living organism. However, despite the availability of complete genomic sequences of several R. palustris strains, no sRNAs have been described for this species thus far. 11,12 The photosynthetic strain Bradyrhizobium sp BTAi1 was isolated from stem nodules of Aeschynomene indica, and its genome, as well as the genome of the phylogenetically closely related B. japonicum USDA 110, are sequenced. 13,14 B. japonicum exists either as a free-living bacterium in soil or as a nitrogen-fixing endosymbiont within the root nodules of host plants such as Glycine max (soybean). Massive physiological changes occur when free-living rhizobia convert to endosymbiotic bacteria (bacteroids). 15,16 B. japonicum USDA 110 was one of the first bacteria for which an sRNA of physiological importance was described. The sra sRNA of this organism was found to be essential for symbiotic root nodule development, and the sra RNA from Sinorhizobium meliloti could complement the symbiotic defect of a B. japonicum USDA 110 sra mutant. 17 Later, it was shown that sra RNA is a *Correspondence to: Elena Evguenieva-Hackenberg; Email: [email protected] Submitted: 03/10/11; Revised: 09/06/11; Accepted: 09/07/11 http://dx.doi.org/10.4161/rna.9.1.18008 RESEARCH PAPER RNA Biology 9:1, 47–58; January 2012; G 2012 Landes Bioscience www.landesbioscience.com RNA Biology 47

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

© 2012 Landes Bioscience.
Do not distribute.
Small RNAs of the Bradyrhizobium/Rhodopseudomonas lineage and their analysis
Ramakanth Madhugiri,1 Gabriella Pessi,2 Björn Voss,3 Julia Hahn,1 Cynthia M. Sharma,4 Richard Reinhardt,5 Jörg Vogel,4
Wolfgang R. Hess,3 Hans-Martin Fischer2 and Elena Evguenieva-Hackenberg1,*
1Institute of Microbiology and Molecular Biology; Gießen, Germany; 2ETH, Institute of Microbiology; Zürich, Switzerland; 3Institute of Biology III; Freiburg, Germany;4Institute of Molecular Infection Biology; Research Center for Infectious Diseases; Würzburg, Germany; 5Max Planck Institute for Plant Breeding Research; Cologne, Germany
Keywords: Bradyrhizobium, Rhodopseudomonas, small RNA, non-coding RNA, nodule, symbiosis, target, prediction,microarray, dRNA-seq
Abbreviations: BjrC, Bradyrhizobium japonicum small RNA candidate; dRNA-seq, differential RNA-seq; nt, nucleotides;ORF, open reading frame; sRNA, small RNA; TSS, transcriptional start site, 5'-UTR, 5' untranslated region
Small RNAs (sRNAs) play a pivotal role in bacterial gene regulation. However, the sRNAs of the vast majority of bacteria withsequenced genomes still remain unknown since sRNA genes are usually difficult to recognize and thus not annotated.Here, expression of seven sRNAs (BjrC2a, BjrC2b, BjrC2c, BjrC68, BjrC80, BjrC174 and BjrC1505) predicted by genomecomparison of Bradyrhizobium and Rhodopseudomonas members, was verified by RNA gel blot hybridization, microarrayand deep sequencing analyses of RNA from the soybean symbiont Bradyrhizobium japonicum USDA 110. BjrC2a, BjrC2band BjrC2c belong to the RNA family RF00519, while the other sRNAs are novel. For some of the sRNAs we observedexpression differences between free-living bacteria and bacteroids in root nodules. The amount of BjrC1505 wasdecreased in nodules. By contrast, the amount of BjrC2a, BjrC68, BjrC80, BjrC174 and the previously described 6S RNA wasincreased in nodules, and accumulation of truncated forms of these sRNAs was observed. Comparative genomics anddeep sequencing suggest that BjrC2a is an antisense RNA regulating the expression of inositol-monophosphatase. Theanalyzed sRNAs show a different degree of conservation in Rhizobiales, and expression of homologs of BjrC2, BjrC68,BjrC1505, and 6S RNA was confirmed in the free-living purple bacterium Rhodopseudomonas palustris 5D.
Introduction
Small RNAs (sRNAs) have been identified as essential elementsof numerous regulatory networks in pro- and eukaryotic cells.Extensive searches for sRNAs in Escherichia coli and in few otherbacteria demonstrated the expression of dozens to hundreds ofsRNAs, most of them with still unknown function.1-7 However,despite the availability of more than 1,000 bacterial genomesequences, the genes for sRNAs are usually difficult to recognizeand, thus, are not annotated. This is mainly due to the highsequence divergence of sRNAs even in related bacteria, but alsobecause of the difficulties to recognize promoters of genes, whichlack associated open reading frames (ORFs). A particularly usefulmethod for detection of sRNAs is their bioinformatic predictionby genome comparison of closely related bacteria, followed byexperimental verification. With this approach, genes for sRNAswhich are located between annotated ORFs are predicted basedon sequence and/or secondary structure conservation.8-10
We are interested in sRNAs of the Bradyrhizobium/Rhodopseudo-monas lineage of a-proteobacteria, which comprises intensivelystudied model organisms like Bradyrhizobium japonicum USDA
110, Bradyrhizobium sp strain BTAi1 and Rhodopseudomonaspalustris. The purple non-sulfur phototrophic R. palustris ismetabolically highly versatile and can perform oxidative phos-phorylation, photophosphorylation, carbon dioxide fixation andnitrogen fixation as a free-living organism. However, despite theavailability of complete genomic sequences of several R. palustrisstrains, no sRNAs have been described for this species thus far.11,12
The photosynthetic strain Bradyrhizobium sp BTAi1 was isolatedfrom stem nodules of Aeschynomene indica, and its genome, as wellas the genome of the phylogenetically closely related B. japonicumUSDA 110, are sequenced.13,14 B. japonicum exists either as afree-living bacterium in soil or as a nitrogen-fixing endosymbiontwithin the root nodules of host plants such as Glycine max(soybean). Massive physiological changes occur when free-livingrhizobia convert to endosymbiotic bacteria (bacteroids).15,16
B. japonicum USDA 110 was one of the first bacteria for whichan sRNA of physiological importance was described. The sra sRNAof this organism was found to be essential for symbiotic rootnodule development, and the sra RNA from Sinorhizobium meliloticould complement the symbiotic defect of a B. japonicumUSDA 110 sra mutant.17 Later, it was shown that sra RNA is a
*Correspondence to: Elena Evguenieva-Hackenberg; Email: [email protected]: 03/10/11; Revised: 09/06/11; Accepted: 09/07/11http://dx.doi.org/10.4161/rna.9.1.18008
RESEARCH PAPER
RNA Biology 9:1, 47–58; January 2012; G 2012 Landes Bioscience
www.landesbioscience.com RNA Biology 47

© 2012 Landes Bioscience.
Do not distribute.
discontinuous tmRNA, the gene of which is permutated inBradyrhizobium and in other a-proteobacteria.18
The tmRNA is one of the most conserved sRNAs, which,similarly to the other house-keeping sRNAs signal recognitionparticle (SRP) RNA (4.5S RNA) and RNase P RNA, can beidentified by simple BLAST analyses. Another conserved sRNA is6S RNA, the secondary structure of which resembles an openpromoter.19 In E. coli, 6S RNA is highly expressed duringtransition to stationary phase leading to sequestration of sigma70-containing RNA polymerase and therefore contributing tochanges in gene expression at late growth stage.19-21 The 6SRNA homolog of B. japonicum USDA 110 was recently identi-fied.22 So far, 6S RNA and tmRNA are the only sRNAs whoseexistence in Bradyrizobium was experimentally verified.
Over the last three years, dozens of sRNAs were experimentallydetected in the a-proteobacteria Caulobacter crescentus, Rhizobiumetli, Rhodobacter sphaeroides and Sinorhizobium meliloti.23-28
Changes in the amount of some of the sRNAs under differentstresses, in different media, and at different growth stages suggestthat they are involved in adaptation to stress, changing environ-mental conditions and in symbiosis.23-28 In the a-proteobacteriumAgrobacterium tumefaciens the sRNA AbcR1, which is homologousto the Hfq-dependent sRNA SmrC16 of S.meliloti, controls theexpression of a periplasmic substrate binding protein required foruptake of the plant-derived defense signal GABA.22,23,29
Here we describe the prediction, verification and analysis ofseven novel sRNAs in the Bradyrhizobium/Rhodopseudomonaslineage. Although the amount of the sRNAs was not significantlyinfluenced by temperature, salt or oxidative stress, changes weredetected at stationary phase and by the comparison of bacteroidsin nodules to free-living B. japonicum USDA 110 cells.
Results
Validation of predicted small RNAs in B. japonicum USDA 110.Our prediction of sRNA genes and other possible RNA elementslocated in intergenic regions (IGRs) of B. japonicum USDA 110was based on the comparison of its genome with the genome ofR. palustris strain BisB5. In addition, predictions were made bycomparing the genomes of Bradyrhizobium BTAi1 and R. palustrisBisB5. The overlap of the results of the two predictions wasinspected to exclude potential riboswitches, regulatory regionsupstream of genes for ribosomal components, and sRNAs with a
coding potential (see Material and Methods). In this way roughlyhalf of the predicted 100 elements (clusters with assignednumbers) were excluded from further analysis. Finally, 15 ofthe clusters with highest scores (Table S1) were chosen forverification in B. japonicum USDA 110. The 16th candidatesubjected to verification (BjrC1505) was predicted for the strainBTAi1 but not for USDA 110 because its sequence overlappedwith the annotated open reading frame (ORF) bsr1505 in thelatter strain. It was included here because a bsr1505 homolog ismissing in the BTAi1 and other related genomes, whereas theputative sRNA is highly conserved.
Verification of the 16 predicted sRNAs was performed by RNAgel blot analysis using total RNA from free-living B. japonicumUSDA 110 grown in PSY medium and from bacteroids insoybean nodules. The results were compared with data frommicroarray analyses.30 During the revision of the manuscript, wereceived first results of a differential RNA-seq analysis (dRNA-seq)of total RNA from free-living B. japonicum USDA 110 andbacteroids.5 The dRNA-seq analysis is based on the differentialsequencing of two cDNA libraries which allows for the annotationof transcriptional start sites. Briefly, each RNA samples wassplit into two halves: one half was treated with terminatorexonuclease (TEX) which specifically degrades processed RNAswith a 5'-mono-phosphate and thereby enriches for primarytranscripts in relative terms (Free+ and Nodule+ libraries). Thesecond half remains untreated and reflects the total RNA pool ofprocessed and primary transcripts (Free- and Nodule- libraries).Here we include the dRNA-seq data for the sRNAs identified inthis study and compare the results obtained by the threeindependent experimental approaches.
In RNA gel blot hybridizations for verification, always oneprobe for the (+) and one for the (-) strand were used, since inmost cases the direction of transcription could not be predicted.Hybridizations with these probes resulted in the detection ofspecific signals for clusters 2, 68, 80, 174, and for the sRNAcandidate overlapping bsr1505 (Table S1), but not for the other11 clusters. The detected sRNAs were named BjrC2, BjrC68,BjrC80, BjrC174 and BjrC1505, respectively (Fig. 1A, lane 1 andlanes 5 to 8). As controls and size markers, 6S RNA and 5S rRNAwere hybridized (Fig. 1A, lanes 9 and 10).
Multiple members of sRNA family RF00519. In cluster 2 twoimperfectly conserved sRNA paralogs were predicted (Table S1)which belong to the RNA family RF00519. Originally identified
Figure 1 (See opposite page). Verified sRNAs and their genomic context in B. japonicum USDA 110. (A) Verification of sRNAs belonging to the clusters 2(the sRNAs BjrC2a, BjrC2b and BjrC2c share homologous sequences leading to their simultaneous detection with probe 2f in lane 1 labeled BjrC2),68 (BjrC68), 80 (BjrC80), 174 (BjrC174) and of the sRNA overlapping with ORF bsr1505 (BjrC1505) by RNA gel blot hybridization of total RNA ofB. japonicum USDA 110 grown aerobically in PSY medium. 6S RNA (full length form of approximately 160 nt) and 5S rRNA (120 nt) were used as controlsand length standards.14,22 (B) RNA gel blot hybridizations with probes targeting different parts of BjrC80, BjrC174 and 6S RNA. For each sRNA, themembrane shown in lane 1 was stripped and its re-hybridization is shown in lane 2. The probes 80–2 (lane 1) and 80r (lane 2) were used to detect BjrC80,probes 174–2 (lane 1) and 174r (lane 2) for BjrC174, and probes 6S-2 (lane 1) and 6S-1 (lane 2) for 6S RNA. Signals representing full length sRNAs aremarked with bold arrows, the truncated forms are marked with thin arrows. (C) Schematic representation of the genomic loci of the sRNAs. Black openarrows: annotated, protein-encoding genes with their designation in the B. japonicum USDA 110 genome and genomic coordinates. Gray filled arrows:sRNA-corresponding sequences (genes or part of genes). Designations of sRNAs as defined in this study are indicated below the arrows. Black ball stickscorrespond to hairpin loops followed by polyuridine stretches which may represent rho-independent terminators. Open asterisks show the approximateposition of the sequences detected by the used hybridization probes; designation of the probes is given (see also Table S1). Distances between proteinencoding genes and sRNA sequences (indicated in nt) are based on dRNA-seq data and genome coordinates. The drawing is not to scale.
48 RNA Biology Volume 9 Issue 1

© 2012 Landes Bioscience.
Do not distribute.
as a putative suhB riboswitch, RF00519 sRNAs exist in multiplecopies in a-proteobacterial genomes, without association withspecific neighboring genes.31,32 The most conserved sequence
element of the two predicted cluster 2 paralogs, termed “2f,” wasaligned to the genome of B. japonicum USDA 110 by BLASTN,resulting in five hits in the following IGRs (in descending order of
Figure 1. For figure legend, see page 48
www.landesbioscience.com RNA Biology 49

© 2012 Landes Bioscience.
Do not distribute.
the scores): blr7730-blr7731, blr2717-bll2718, bll7804-bll7805,blr1738-bsr1739 and blr3928-bll3929. This is in agreementwith the notion that five RF00519 copies exist in IGRs of theB. japonicum USDA 110 genome.32 RNAs of the family RF00519are on average 75 nt in length and are characterized by aconserved region upstream of a hairpin followed by a stretch ofuridines which may serve as a rho-independent terminator.31,32
In the IGRs blr7730-blr7731, blr2717-bll2718 and blr1738-bsr1739, harboring the sequences of the putative sRNAs BjrC2a,BjrC2b and BjrC2c, respectively, the 2f elements are directlyfollowed by potential rho-independent terminators. In the IGRblr3928-bll3929 corresponding to a putative sRNA BjrC2d,a potential terminator is located 15 nt downstream of the 2fsequence element. No terminator was predicted downstreamof the 2f sequence element in the IGR bll7804-bll7805corresponding to a putative sRNA BjrC2e. The LocARNA andRNAalifold analyses suggest secondary structure conservation ofBjrC2a, BjrC2b, BjrC2c and BjrC2d. For the RF00519 hitBjrC2e there was no additional sequence similarity beyond the 2fsequence element (Fig. S1A).
Using probe 2f covering the highly conserved part of theRF00519 paralogs of B. japonicum USDA 110, three bands weredetected (Fig. 1A, lane 1). RNA gel blot analyses with probesspecific for the five putative RF00519 paralogs suggest that onlythe sRNAs BjrC2a, BjrC2b and BjrC2c were detected with probe2f (Fig. 1A and C; Table S1). The BjrC2a-specific probe detectedtwo small bands of approximately 65 and 70 nt (compare lanes 1and 2 in Fig. 1A). BjrC2b corresponds to the largest transcriptdetected in lane 1 (compare lanes 1 and 3 in Fig. 1A). BjrC2cwas detected as a very weak band of approximately 65 nt (Fig. 1A,lane 4). No sRNAs were detected with BjrC2d- and Bjrc2e-specific probes (not shown).
Expression of BjrC2a, BjrC2b, and BjrC2c was confirmedby microarray and dRNA-seq (Figs. 2, S2A–C). Microarrayand dRNA-seq data support expression of the BjrC2d region(Fig. S2D), whereas dRNA-seq showed no expression of theBjrC2e region (not covered by the microarray analysis). Insummary, we confirmed the expression of three RF00519 paralogsin B. japonicum USDA 110: BjrC2a, 2b and 2c. Signals corres-ponding to at least two of them, BjrC2a and BjrC2c, may overlapin RNA gel blot hybridizations with probe 2f.
The sRNAs BjrC68, BjrC80, BjrC174, BjrC1505 and 6S RNA.More than one signal was detected by RNA gel blot hybridi-zation also for BjrC80 and BjrC174 (Fig. 1A, lanes 6 and 7), and,in agreement with previous results, for 6S RNA (Fig. 1A,lane 9).22 Additional hybridizations with probes directed todifferent regions of those sRNAs strongly suggested that theshorter transcripts arise from degradation of the longer forms,which can be considered as the full-length sRNAs (Fig. 1Band C).
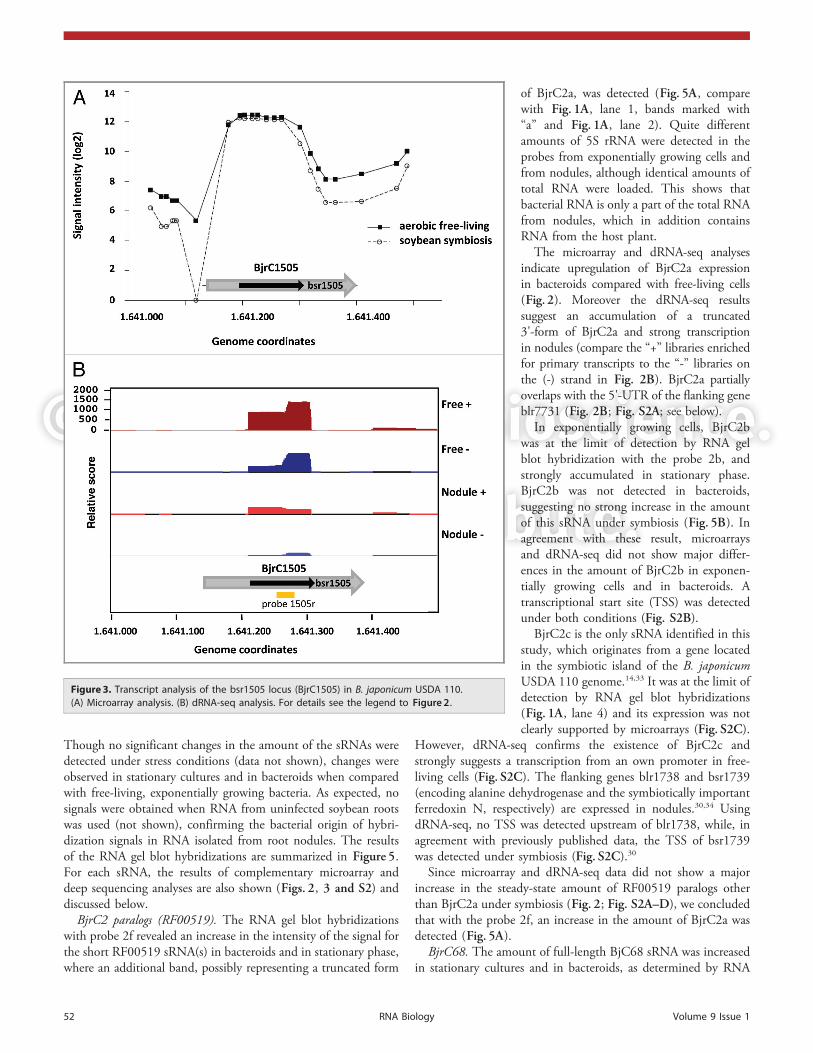
Microarray and dRNA-seq data supported the expressionof BjrC68, BjrC80, BjrC174 (Fig. S2E–G), BjrC1505 (Fig. 3;Fig. S2H) and 6S RNA (Fig. S2I). The short BjrC1505 transcriptdetected by RNA gel blot hybridization overlaps in sense withbsr1505 (Fig. 1A and C). Data from microarray and dRNA-seqrevealed high accumulation of transcripts corresponding to the
central part of the putative bsr1505 mRNA (Fig. 3), stronglysupporting the existence of an sRNA in this region.
Conservation of sRNAs in related bacteria and detection inR. palustris 5D. To analyze the conservation of the verifiedsRNAs in other bacteria, the corresponding B. japonicum USDA110 genomic regions (Table S1) were used in BLASTN searchesagainst the microbial section of NCBI. Sequence hits of lengthscomparable to the full-length or the truncated forms of thesRNAs (Fig. 1A) were further aligned and consensus secondarystructures were created with LocARNA and RNAalifold(Fig. S1B–F). BjrC68-like sequences were detected only ingenomes of Bradyrhizobium and Rhodopseudomonas species, whileBjrC80- and BjrC174-like sequences were also found in speciesof the genera Oligotropha and Nitrobacter, which both belongto the Bradyrhizobiaceae family. Sequences corresponding toBjrC1505 and to the BjrC2 paralogs (RF00519) are conservedin numerous genera of Rhizobiales (Table 1). The high degree ofconservation, both at the sequence and secondary structure level,among BjrC1505 orthologs in many genera of the Rhizobialesstrongly supports the existence of an sRNA at the bsr1505 locusin B. japonicum USDA 110 (Fig. S1F). In contrast, databasesearches failed to detect homologs of the Bsr1505 protein in otherorganisms.
Multiple BjrC2 paralogs are present in many representatives ofRhizobiales (Table 1). We noticed that the genomic context ofBjrC2a is highly conserved. When more than one copy of BjrC2-like sequences is present in a given genome, one of the copies isalways located 200 to 400 nt upstream of a gene encoding aninositol monophosphatase family protein (blr7731 in B. japoni-cum USDA 110) and is transcribed in the opposite direction(Figs. 1C and 2B). The genomic context of the other sRNAs isalso conserved. Orthologs of BjrC80 are located between a genefor a serine protease precursor and a gene for a two-componentresponse regulator (blr3130 and blr3131 in B. japonicum USDA110). The genes for BjrC174 orthologs are located betweengenes for conserved hypothetical proteins (bll0464 and bll0465in B. japonicum USDA 110, with domains DUF2794 andDUF1223, respectively), while those for BjrC68 orthologs arefound downstream of a gene encoding an ammonium transporter(blr0613 in B. japonicum USDA 110). The genes for BjrC1505orthologs are found between genes for two hypothetical proteins(bsr1504 and blr1506 in B. japonicum USDA 110) comprisingconserved domains of unknown function (DUF1192 andDUF1465; Fig. 1C).
Given the high conservation of the sRNA sequences inBradyrhizobium and Rhodopseudomonas genomes, we tried tovalidate homologous transcripts in total RNA from the non-sequenced strain R. palustris 5D. Though we failed to findBjrC174- and BjrC80-like sRNAs using probes 174r, 174–3 and80r (Table S1), the ocurrence of sRNAs similar to BjrC2a,BjrC68, BjrC1505 and 6S RNA was detected in RNA gel blotswith R. palustris 5D RNA (Fig. 4). Except for BjrC68, the sRNAsin R. palustris 5D are slightly larger (6S RNA) or shorter(the BjrC2 and BjrC1505 homologs) than the correspondingB. japonicum USDA 110 sRNAs, as judged from the main signalsin Figure 4. Using the 5S rRNA signal as reference it seems that
50 RNA Biology Volume 9 Issue 1

© 2012 Landes Bioscience.
Do not distribute.
the R. palustris homologs of BjrC2, BjrC68, and 6S RNA arehigher expressed in in R. palustris 5D than in B. japonicum USDA110 under the applied growth conditions.
Differential expression of sRNAs in free-living bacteria and insoybean root nodules. To address possible physiological roles ofthe seven candidate sRNAs and of 6S RNA, their expression in
bacteroids and in free-living B. japonicum USDA 110 cellsharvested at different growth phases or after exposure to heat,cold, salt or oxidative stress during exponential growth was com-pared by RNA gel blot analyses (see material and methods). Sincevery weak signals were obtained with the hybridization probes 2aand 2c, the short BjrC2 sRNAs were monitored with probe 2f.
Figure 2. Transcript analysis of the blr7730-7731 locus (BjrC2a) in B. japonicum USDA 110. (A) Microarray analysis. Graphical representation ofhybridization signal intensities derived from individual probe pairs of this region in microarray experiments.30 The expression profile in wild-type cellsgrown aerobically in complex PSY media and in soybean nodules is shown. Solid lines marked with “a” and “f” show the position of the probes 2a and 2fused in RNA gel blot hybridizations. (B) dRNA-seq analysis.5 Total RNA from free-living cells (Free) or from soybean root nodules (Nodule) was analyzed.Compared are cDNA libraries from untreated, total RNA (Free-, Nodule -, blue graphs), and from RNA treated with terminator exonuclease (Free+, Nodule+, red graphs) to enrich primary, transcripts with a 5’-triphosphate. cDNAs were mapped to the (+) or (–) strand of the corresponding region of the B.japonicum USDA 110 chromosome. Y-axes scales give a relative score (‰ mapped reads per genome position) and are consistent for all libraries of thesame strand (indicated for only one of the libraries). Annotated genes and the BjrC2a gene are indicated by gray and black horizontal arrows,respectively, adjacent to the genome coordinate axis.14 TSS are marked with flexed arrows; dashed arrows mark questionable TSS.
www.landesbioscience.com RNA Biology 51
© 2012 Landes Bioscience.
Do not distribute.
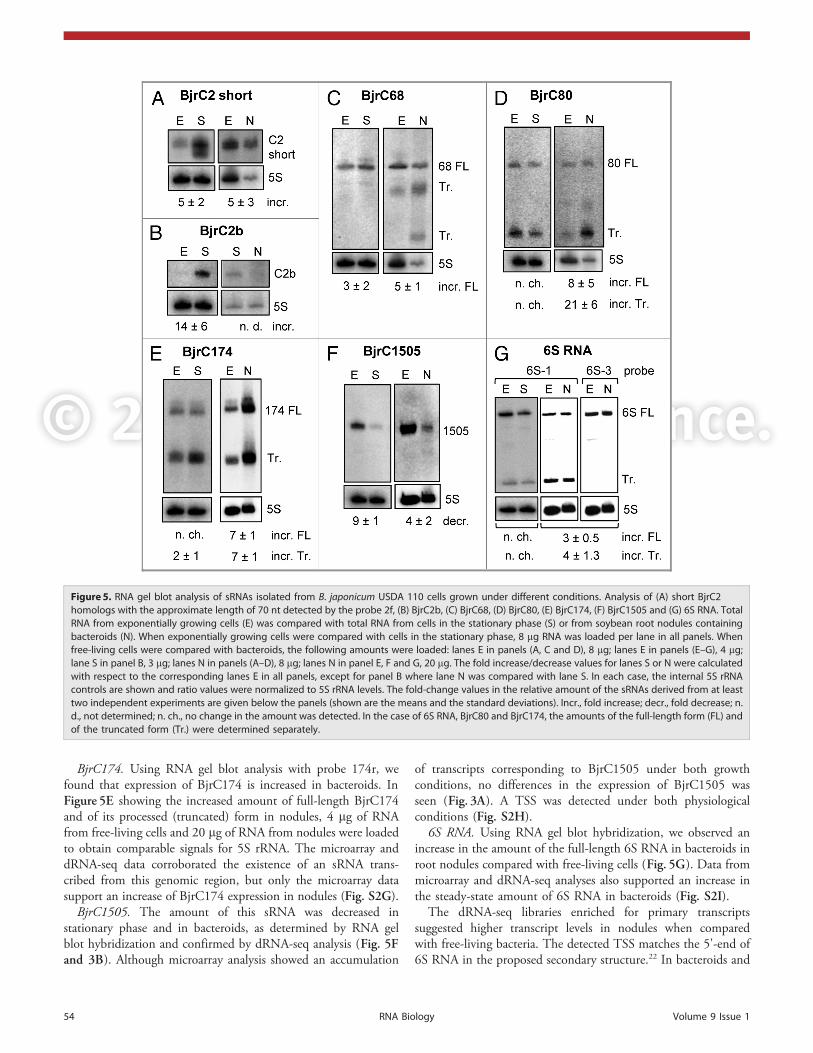
Though no significant changes in the amount of the sRNAs weredetected under stress conditions (data not shown), changes wereobserved in stationary cultures and in bacteroids when comparedwith free-living, exponentially growing bacteria. As expected, nosignals were obtained when RNA from uninfected soybean rootswas used (not shown), confirming the bacterial origin of hybri-dization signals in RNA isolated from root nodules. The resultsof the RNA gel blot hybridizations are summarized in Figure 5.For each sRNA, the results of complementary microarray anddeep sequencing analyses are also shown (Figs. 2, 3 and S2) anddiscussed below.
BjrC2 paralogs (RF00519). The RNA gel blot hybridizationswith probe 2f revealed an increase in the intensity of the signal forthe short RF00519 sRNA(s) in bacteroids and in stationary phase,where an additional band, possibly representing a truncated form
of BjrC2a, was detected (Fig. 5A, comparewith Fig. 1A, lane 1, bands marked with“a” and Fig. 1A, lane 2). Quite differentamounts of 5S rRNA were detected in theprobes from exponentially growing cells andfrom nodules, although identical amounts oftotal RNA were loaded. This shows thatbacterial RNA is only a part of the total RNAfrom nodules, which in addition containsRNA from the host plant.
The microarray and dRNA-seq analysesindicate upregulation of BjrC2a expressionin bacteroids compared with free-living cells(Fig. 2). Moreover the dRNA-seq resultssuggest an accumulation of a truncated3'-form of BjrC2a and strong transcriptionin nodules (compare the “+” libraries enrichedfor primary transcripts to the “-” libraries onthe (-) strand in Fig. 2B). BjrC2a partiallyoverlaps with the 5'-UTR of the flanking geneblr7731 (Fig. 2B; Fig. S2A; see below).
In exponentially growing cells, BjrC2bwas at the limit of detection by RNA gelblot hybridization with the probe 2b, andstrongly accumulated in stationary phase.BjrC2b was not detected in bacteroids,suggesting no strong increase in the amountof this sRNA under symbiosis (Fig. 5B). Inagreement with these result, microarraysand dRNA-seq did not show major differ-ences in the amount of BjrC2b in exponen-tially growing cells and in bacteroids. Atranscriptional start site (TSS) was detectedunder both conditions (Fig. S2B).
BjrC2c is the only sRNA identified in thisstudy, which originates from a gene locatedin the symbiotic island of the B. japonicumUSDA 110 genome.14,33 It was at the limit ofdetection by RNA gel blot hybridizations(Fig. 1A, lane 4) and its expression was notclearly supported by microarrays (Fig. S2C).
However, dRNA-seq confirms the existence of BjrC2c andstrongly suggests a transcription from an own promoter in free-living cells (Fig. S2C). The flanking genes blr1738 and bsr1739(encoding alanine dehydrogenase and the symbiotically importantferredoxin N, respectively) are expressed in nodules.30,34 UsingdRNA-seq, no TSS was detected upstream of blr1738, while, inagreement with previously published data, the TSS of bsr1739was detected under symbiosis (Fig. S2C).30
Since microarray and dRNA-seq data did not show a majorincrease in the steady-state amount of RF00519 paralogs otherthan BjrC2a under symbiosis (Fig. 2; Fig. S2A–D), we concludedthat with the probe 2f, an increase in the amount of BjrC2a wasdetected (Fig. 5A).
BjrC68. The amount of full-length BjC68 sRNA was increasedin stationary cultures and in bacteroids, as determined by RNA
Figure 3. Transcript analysis of the bsr1505 locus (BjrC1505) in B. japonicum USDA 110.(A) Microarray analysis. (B) dRNA-seq analysis. For details see the legend to Figure 2 .
52 RNA Biology Volume 9 Issue 1

© 2012 Landes Bioscience.
Do not distribute.gel blot hybridization, microarrays and dRNA-seq (Fig. 5C;Fig. S2E). Interestingly, shorter RNA products of similarabundance were detected in bacteroids by RNA gel blothybridizations, pointing to degradation or turnover of BjrC68during symbiosis (Fig. 5C). Like the full-length form, these
additional products were not detected in RNA from roots,excluding the possibility of unspecific hybridization of the probeto RNAs of the plant (data not shown).
The microarray data suggest cotranscription of BjrC68with blr0613 encoding an ammonium transporter, which isalso upregulated during symbiosis (Fig. S2E) and detected asa protein in bacteroids.34 The dRNA-seq data also suggestcotranscription, since no TSS was detected upstream of BjrC68under symbiosis, while the TSS of blr0613 was detected(Fig. S2E). However, we were not able to confirm cotrans-cription by RT-PCR using RNA from free-living cells and fromnodules. Hybridization of total RNA separated in agaroseformaldehyde gels with the probe 68r also did not detect a largetranscript containing blr0613 mRNA and BjrC68 (not shown).
BjrC80. Using RNA gel blot hybridization, we found anincrease in the amount of both the full-length and thedegraded or processed product of BjrC80 in nodules whencompared with exponential cultures, while no change wasmeasured in stationary cultures (Fig. 5D). Microarrays alsoshowed a higher expression of BjrC80 in bacteroids andsuggested cotranscription of BjrC80 with the flanking genesblr3130 and blr3131 (Fig. S2F). dRNA-seq confirmedslightly higher expression of BjrC80 in bacteroids,accumulation of shorter, processed form(s), and lack of anown TSS, consistent with cotranscription with the upstreamgene. RT-PCR analysis confirmed the cotranscription ofblr3130 and blr3131, implying that BjrC80 is processedfrom a polycistronic transcript (Fig. S2F).
Table 1. Distribution of sequences with homology to the B. japonicum USDA 110 sRNAs in genera of the order Rhizobiales
Genus BjrC2 BjrC68 BjrC80 BjrC174 BjrC1505
Bradyrhizobium + (at least 3 x, scattered inB. japonicum USDA 110)
+ + + +
Rhodopseudomonas + + + + +
Oligotropha + + + +
Nitrobacter + + + +
Methylobacterium + (several hits in IGRs, scattered) +
Mesorhizobium + +
Parvibaculum +
Brucella +
Ochrobactrum +
Xanthobacter + (2 x in an IGR)
Azorhizobium + (2 x in an IGR)
Bejerinckia + (5 x in an IGR)
Methylocella + (2 x in an IGR)
Agrobacterium + (RF00519)32
Rhizobium +
Sinorhizobium + (Sm8)25
Plus indicates that homologous sequences with the approximate length of the B. japonicum USDA 110 sRNAs (as estimated by RNA gel blot analysis, Fig. 1Aand Table S1), were detected by BLAST searches in the strains specified below. An exception is the detection of BjrC2-like sequences in Agrobacterium,Rhizobium and Sinorhizobium: in these cases, the 30 nt sequence CCCTCCTTGGGTGTTTCCTCCCTAGACTTG is present 200 to 400 nt upstream of the geneencoding an inositol monophosphatase family protein. In several genomes multiple BjrC2-like sequences were found. Our analysis does not exclude that ingenomes of Rhodopseudomonas, Oligotropha, Nitrobacter and in Rhizobiaceae, several imperfect copies of BjrC2-like sequences are present. IGR, intergenicregion.
Figure 4. Detection of sRNAs in R. palustris 5D. RNA gel blots documenting theexpression of sRNAs in R. palustris 5D (R. pal.) which are homologous toBjrC1505, BjrC2, BjrC68 and 6S RNA of B. japonicum USDA 110 (B. jap.). Thetype of the detected sRNA and the species from which the analyzed total RNAoriginates are indicated on the right side and on top of the panels,respectively.
www.landesbioscience.com RNA Biology 53
© 2012 Landes Bioscience.
Do not distribute.
BjrC174. Using RNA gel blot analysis with probe 174r, wefound that expression of BjrC174 is increased in bacteroids. InFigure 5E showing the increased amount of full-length BjrC174and of its processed (truncated) form in nodules, 4 mg of RNAfrom free-living cells and 20 mg of RNA from nodules were loadedto obtain comparable signals for 5S rRNA. The microarray anddRNA-seq data corroborated the existence of an sRNA trans-cribed from this genomic region, but only the microarray datasupport an increase of BjrC174 expression in nodules (Fig. S2G).
BjrC1505. The amount of this sRNA was decreased instationary phase and in bacteroids, as determined by RNA gelblot hybridization and confirmed by dRNA-seq analysis (Fig. 5Fand 3B). Although microarray analysis showed an accumulation
of transcripts corresponding to BjrC1505 under both growthconditions, no differences in the expression of BjrC1505 wasseen (Fig. 3A). A TSS was detected under both physiologicalconditions (Fig. S2H).
6S RNA. Using RNA gel blot hybridization, we observed anincrease in the amount of the full-length 6S RNA in bacteroids inroot nodules compared with free-living cells (Fig. 5G). Data frommicroarray and dRNA-seq analyses also supported an increase inthe steady-state amount of 6S RNA in bacteroids (Fig. S2I).
The dRNA-seq libraries enriched for primary transcriptssuggested higher transcript levels in nodules when comparedwith free-living bacteria. The detected TSS matches the 5'-end of6S RNA in the proposed secondary structure.22 In bacteroids and
Figure 5. RNA gel blot analysis of sRNAs isolated from B. japonicum USDA 110 cells grown under different conditions. Analysis of (A) short BjrC2homologs with the approximate length of 70 nt detected by the probe 2f, (B) BjrC2b, (C) BjrC68, (D) BjrC80, (E) BjrC174, (F) BjrC1505 and (G) 6S RNA. TotalRNA from exponentially growing cells (E) was compared with total RNA from cells in the stationary phase (S) or from soybean root nodules containingbacteroids (N). When exponentially growing cells were compared with cells in the stationary phase, 8 mg RNA was loaded per lane in all panels. Whenfree-living cells were compared with bacteroids, the following amounts were loaded: lanes E in panels (A, C and D), 8 mg; lanes E in panels (E–G), 4 mg;lane S in panel B, 3 mg; lanes N in panels (A–D), 8 mg; lanes N in panel E, F and G, 20 mg. The fold increase/decrease values for lanes S or N were calculatedwith respect to the corresponding lanes E in all panels, except for panel B where lane N was compared with lane S. In each case, the internal 5S rRNAcontrols are shown and ratio values were normalized to 5S rRNA levels. The fold-change values in the relative amount of the sRNAs derived from at leasttwo independent experiments are given below the panels (shown are the means and the standard deviations). Incr., fold increase; decr., fold decrease; n.d., not determined; n. ch., no change in the amount was detected. In the case of 6S RNA, BjrC80 and BjrC174, the amounts of the full-length form (FL) andof the truncated form (Tr.) were determined separately.
54 RNA Biology Volume 9 Issue 1

© 2012 Landes Bioscience.
Do not distribute.
in free-living bacteria, short primary pRNA transcripts (10 to 17nt in length) were detected, which are probably synthesized byRNA polymerase during the release of 6S RNA (Fig. S2I).19,20
This indicates an active turnover of 6S RNA in free-living cellsand in bacteroids. Turnover is also supported by the detection of amajor degradation or processing product (truncated form) of 6SRNA by RNA gel blot hybridization. A specific truncated formaccumulates since it was detected with the probe 6S-1 targetingthe 3'-part of the central bulge of 6S RNA, but not with the probe6S-2 directed against the 5'-part of the central bulge or withthe probe 6S-3, which targets the 5'-end of 6S RNA (Figs. 5Gand 1B; Fig. S2I).22
Potential targets of the sRNAs BjrC1505 and BjrC2a. Mostof the bacterial regulatory sRNAs which have been functionallycharacterized to date act as antisense RNAs by imperfect base-pairing interactions with target mRNAs. However, for somesRNAs it has been shown that the regulatory function can also beexerted by binding of specific proteins.19,35 Here, possible mRNAtargets of the B. japonicum USDA 110 sRNAs were predictedusing TargetRNA and IntaRNA.36,37 Due to the high GC-contentof B. japonicum USDA 110 many potential targets with question-able significance were predicted.38 Therefore we considered onlypotential targets predicted by both algorithms. Such predictionswere obtained for potential targets of BjrC1505 and BjrC2a. As apossible target of BjrC1505, the translational start of blr4023(encoding acetolactate synthase) was predicted. Furthermore, apotential base-pairing of the highly conserved BjrC2a part (andthus of the RF00519 paralogs) with the 5'-UTR of bll7804(encoding 3-hydroxyacyl-CoA dehydrogenase type II) wassuggested. The pairing potential between the sRNAs and thepredicted targets is shown in Figure S3.
The highly conserved genomic context of BjrC2a raised thequestion whether this sRNA may act as an antisense RNA toregulate the expression of the inositol monophosphatase geneblr7731 (Fig. 1C). In such a case the blr7731 mRNA shouldharbor a large 5'-UTR. Indeed, dRNA-seq strongly supports theexistence of a TSS at position 8474751, which would lead to ablr7731 transcript partially overlapping with the antisense BjrC2a(Fig. 2B; Fig. S2A). Enrichment of primary blr7731 transcriptsstarting at this position was detected only in free-living cells butnot in bacteroids when the expression of BjrC2a is increased,supporting the view that BjrC2a downregulates blr7731.
Discussion
In this work, a computational approach was used to identifysRNAs in the Bradyrhizobium/Rhodopseudomonas lineage. Ourmethod generally identifies sRNAs genes in IGRs, and thussRNAs overlapping with annotated ORFs like BjrC1505 could beoverlooked. The following observations suggests the existence ofan sRNA encoded at the bsr1505 locus in B. japonicum USDA110: (1) the high degree of conservation among BjrC1505orthologs in Rhizobiales, (2) the failure to detect homologs of theBsr1505 protein in other organisms, (3) the failure to detect theBsr1505 protein by proteomic analyses of B. japonicum USDA110 during symbiosis and in free-living conditions (G. Pessi,
unpublished data), and (4) the detection of sRNA BjrC1505under both conditions (Figs. 3 and 5F).34,39 These facts suggestbsr1505 to be mis-annotated and that it should be replaced by agene encoding BjrC1505.
Taken together, the in silico analysis predicted approximately100 clusters of putative regulatory RNA elements, out of whichapproximately 50 were assumed to represent sRNAs. We analyzed16 clusters in more detail (15 clusters for putative sRNAspredicted with highest scoring for B. japonicum USDA 110 andthe cluster corresponding to BjrC1505, which was predicted forBradyrhizobium sp. BTAi1). From these, five were confirmed tocorrespond to sRNAs. This yield is comparable to that reported insimilar studies on the computational prediction of bacterialsRNAs.40 The reliability of the method is demonstrated by theprediction of 6S RNA and of members of the RNA familyRF00519 in cluster 2 with strong support (Table S1). The othervalidated clusters correspond to novel sRNAs.
We observed accumulation of high amounts of truncated formsof BjrC68, BjrC80, BjrC174 and 6S RNA in B. japonicum USDA110, particularly during symbiosis (Fig. 5). Likewise, truncatedforms of BjrC1505, BjrC68 and 6S RNA were detected inR. palustris (Fig. 4). Truncated forms could represent degradationproducts, and their accumulation could reflect a fast turnover ofthe sRNAs and/or slower decay of the truncated forms undercertain conditions. Alternatively, the truncated derivatives couldcorrespond to functional, processed forms of primary sRNAs,similarly to the abundant processed form of AcrZ in Salmonella.41
The quantitative analysis of the sRNAs by RNA gel blothybridization was compared with the microarray analysis, whichonly allows for a semi-quantitative comparison of RNAs, and tothe dRNA-seq data, which semi-quantitatively reflect differencesin RNA levels.5,16,30,42 Generally, the three methods displayedsimilar tendencies, and decreases or increases in the amounts weresupported by at least two of the methods for all sRNAs.
The sizes in the RNA gel blots are smaller than the predictedsizes, with exception of BjrC2b, which is larger in the RNA gelblot (Table S1). Probably the predictions include conservedregulatory sequences, and/or the RNA gel blots detect alreadyprocessed forms. In the case of BjrC2b we see an example of asRNA with a unique 5'-part and highly conserved 3'-partbelonging to RF00519. When compared with the dRNA-seqdata, the sizes of BjrC2b, BjrC2c, BjrC68 and BjrC1505 wereslightly overestimated in the RNA gel blots, while the sizes of thefull length BjrC80 and BjrC2a were underestimated (Table S1).Persistent secondary structures of the GC-rich sRNAs mayaccount for the observed differences, as previously discussed forS. meliloti.7 Solexa enables sequencing of approximately 100 nt atthe 5'-end of an RNA. Obviously, length estimation of 180 nt forBjrC80 and of 163 nt for 6S RNA by dRNA-seq results fromsequencing of 3'-fragments in addition to the corresponding full-length forms and/or 5'-fragments (Table S1; Fig. S2F and S2I).
The predicted interactions between sRNAs and their targetmRNAs by two independent algorithms suggest that BjrC1505and BjrC2a (or RF00519 paralogs) may regulate gene expressionat the post-transcriptional level as described for enterobacterialsRNAs.2,43,44 Notably, the expression pattern of these sRNAs in
www.landesbioscience.com RNA Biology 55

© 2012 Landes Bioscience.
Do not distribute.
bacteroids fits with that of the predicted targets. Consistent withthe increase in the amount of BjrC2a in bacteroids, the expressionof bll7804 (3-hydroxyacyl-CoA dehydrogenase type II) was notdetected in a transcriptomics and proteomics study, in whichexpression of 43% of the predicted B. japonicum USDA 110 genesand/or proteins was demonstrated in bacteroids.34 Likewise, thedecrease in the amount of BjrC1505 during symbiosis is in linewith the expression of its putative target, acetolactate synthase(blr4023) in bacteroids.34,45 Furthermore, the suggestion thatBjrC2a paralogs act as antisense RNAs regulating inositol mono-phosphatase in Rhizobiales fits well to the reported need for a finetuning the synthesis of this protein in Rhizobium leguminosarumfor successful symbiosis with plants.46,47
Since genomes of non-symbiotic Rhodopseudomonas andsymbiotic Bradyrhizobium species were used for the predictionof sRNAs, many sRNAs with increased expression in bacteroidsare probably not exclusively involved in the interaction betweenB. japonicum and its host plant, but may contribute to other,more general aspects of bacteroid physiology. This view is furthersupported by the finding that the amount of some of these sRNAsis also increased in stationary phase during free-living growth. Inconclusion, our work led to the identification of several novelsRNAs as a first step in the analysis of the still unexplored pool ofsRNAs in the Bradyrhizobium/Rhodopseudomonas lineage.
Material and Methods
Computational methods. Predictions of sRNA genes and otherRNA elements located within intergenic regions were made aspreviously described.10 In brief, all intergenic regions longer than50 nt were compared among the three investigated genomes usingBLASTN. Genome sequences were obtained from the finishedmicrobial genomes website at GenBank (www.ncbi.nlm.nih.gov/genomes/lproks.cgi) with the following accession numbers:B. japonicum USDA 110 (NC_004463), Bradyrhizobium BTAi1(NC_009485) and R. palustris BisB5 (NC_007958). Based on theBLASTN results, homologous sequence regions were clustered,aligned using ClustalW, and analyzed for structural significancewith RNAz.48,49 Alignments were postprocessed using the toolrnazSelectSeqs.pl with default parameters from the RNAzpackage. RNAz was applied in a sliding window approach (astep size of 10 nt and different window sizes were used, namely80,100,120,140 and 160 nt) of which the window with highestprobability was selected. Elements were considered high-scoringwhen they achieved an RNAz probability of $0.5 or their Z-scorewas #-2.0.
The Rfam database was searched for known RNA families.31
For secondary structure prediction of individual sequences Mfoldwas used, for predictions based on multiple sequence alignmentsthe program RNAalifold.50,51 To analyze the coding potential ofthe predicted sRNAs the program RNAcode was used, whichdifferentiates between non-coding and possibly coding regionsbased on nucleotide substitution patterns in an alignment of atleast three homologous sequences.52 This approach lead to theidentification of a maximum of three possible candidates forpeptide-coding or dual function sRNAs with reasonable p values
of less than 0.05. Potential mRNA targets were predicted byTargetRNA and IntaRNA.36,37 The sequence of each validatedsRNA was used to search each annotated mRNA and its flankingregions in the genome of B. japonicum USDA 110 for basepairingpotential. The p value threshold was set to 0.01 when TargetRNAwas used, and a perfectly matching seed of 7 nt was applied whenIntaRNA was used.
Bacterial strains and growth conditions. The strains used inthis work were R. palustris 5D and B. japonicum 110spc4, whichis a spectinomycin-resistant derivate of B. japonicum USDA 110(US Department of Agriculture).53,54
Free-living B. japonicum 110spc4 cells were grown at 28–30°Cin PSY (complex peptone-salt-yeast extract medium) supple-mented with spectinomycin (100 mg ml21).55 Cells were harvestedin exponential phase (OD600 = 0.4 to 0.5), or in stationary phase(OD600 = 0.9 to 1.1). The following stress conditions wereapplied to exponentially growing cultures: heat stress, 30 min at39°C; cold stress, 30 min at 20°C; oxidative stress, 5 min exposureto 2 mM H2O2; salt stress, 30 min at 50 mM NaCl. Controlcultures were incubated for the same time without applying stressconditions. For symbiotic conditions, soybean seedlings (Glycinemax L. Merr., cv Williams; kindly provided by D. N. Navarro andF. Temprano, CIFA-Las Torres Tomejil, Spain) were inoculatedwith B. japonicum 110spc4 and root nodules were harvested 21 dpost infection.16,56 R. palustris 5D was grown at 32°C in minimalmalate salt medium.57
RNA isolation and RNA gel blot analysis. Isolation of totalRNA from free-living cells of B. japonicum 100spc4 andR. palustris 5D by a modified TRIzol method, and RNA gelblot hybridization were performed as previously described.7,58
RNA from soybean nodules and from uninfected soybean rootswas isolated by the modified hot-phenol method.16 RNA samples(7.5 mg RNA from free-living bacteria or 15 mg RNA fromnodules or uninfected roots for initial screening; amounts usedin Figure 5 are specified in the legend) were separated on 10%polyacrylamide-urea gels, transferred onto a nylon membraneand hybridized with oligonucleotides, which were radioactivelylabeled at the 5'-end using c-32P-ATP and polynucleotide kinase.After overnight hybridization at 56°C in a solution containingthe labeled probe (2 to 5 � 106 cpm), 6 � SSC, 0.5% SDSand salmon sperm DNA (50 mg/ml), the membranes werewashed twice in 0.01% SDS, 5 � SSC at room temperature.Membranes were stripped for 20 min at 96°C in 0.1% SDS andre-hybridized.
The probes used for RNA gel blot analysis are given inTable S1. Signals were detected and analyzed using a BioRadmolecular imager and the QuantityOne (BioRad) software. Theintensity of the sRNA bands was normalized to the intensity ofthe 5S rRNA.
Microarray analysis. The genomic region around each sRNAgene was specifically analyzed using global expression data froma microarray of B. japonicum 110spc4 cells grown aerobically inPSY medium supplemented with 0.1% L-arabinose and fromB. japonicum 110spc4 soybean bacteroids harvested 21 d postinfection as described previously.16,30 For the tiling analyses,signals from each probe pairs were analyzed using GTRANS
56 RNA Biology Volume 9 Issue 1

© 2012 Landes Bioscience.
Do not distribute.
(Affymetrix) with a bandwith of 70 and visualized using Prism.Transcriptomics experiments were done using a custom-madeAffymetrix GeneChip representing 8,266 annotated B. japonicumUSDA 110 genes plus intergenic regions.30
RT-PCR. Reverse transcription (RT)-PCR was performed withthe One-Step-RT-PCR Kit (Qiagen, Hilden, Germany) using50 ng/ml DNaseI-treated RNA. Reaction products wereseparated on agarose gels and stained with ethidium bromide.Following primers were used: blr3130f 5'-GGCAAGAGCGT-GAGCACC-3', blr3131r 5'-GAGGTCACCTTCTCTAAAC-ATG-3', bjrC68r_short 5'-CTCAACGGTCGGGGCATTG-3',blr0613f 5'-GCATCGCAGGTCTCGTCGG-3'.
dRNA-seq analysis. Differential RNA sequencing (dRNA-seq)of total RNA from nodules and from cultures at OD600 = 0.5 wasperformed as previously described.5 Depletion of processed RNAswas performed as previously described.5 Briefly, total RNA fromfree-living bacteria and nodules was treated with TerminatorTM5'-phosphate-dependent exonuclease (TEX) (Epicentre#TER51020) or in buffer alone followed by TAP (tobacco acidpyrophosphatase) treatment. cDNA libraries for Solexa sequenc-ing were prepared at Vertis Biotechnology AG (Germany) similarlyas previously described.59 Solexa sequencing was performed on
Illumina Genome Analyzer IIx machine. Mapping of cDNAlibraries to the Bradyrhizobium japonicum USDA 110 genome(NC_004463) using the segemehl software was done as previouslydescribed.5,60 For each library (Free-/+ TEX and Nodule -/+ TEX)graphs representing the number of mapped reads per nucleotidewere calculated and visualized using the Integrated GenomeBrowser software from Affymetrix as previously described.59
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Angelika Balzer and Sebastian Thalmann (University ofGiessen) for help in some experiments, and Konrad U. Förstner(University of Würzburg) for help with the computational analysisof deep sequencing data. This work was supported by DeutscheForschungsgemeinschaft, SPP1258, grant Ev42/4–1 to E.E.-H.and He2544/4–2 to W.R.H.
Supplemental Material
Supplemental materials can be found at:www.landesbioscience.com/journals/rnabiology/article/18008
References1. Altuvia S. Identification of bacterial small non-coding
RNAs: experimental approaches. Curr Opin Microbiol2007; 10:257-61; PMID:17553733; http://dx.doi.org/10.1016/j.mib.2007.05.003
2. Storz G, Altuvia S, Wassarman KM. An abundance ofRNA regulators. Annu Rev Biochem 2005; 74:199-217;PMID:15952886; http://dx.doi.org/10.1146/annurev.biochem.74.082803.133136
3. Sharma CM, Vogel J. Experimental approaches forthe discovery and characterization of regulatory smallRNA. Curr Opin Microbiol 2009; 12:536-46; PMID:19758836; http://dx.doi.org/10.1016/j.mib.2009.07.006
4. Georg J, Voss B, Scholz I, Mitschke J, Wilde A, HessWR. Evidence for a major role of antisense RNAs incyanobacterial gene regulation. Mol Syst Biol 2009;5:305; PMID:19756044; http://dx.doi.org/10.1038/msb.2009.63
5. Sharma CM, Hoffmann S, Darfeuille F, Reignier J,Findeiss S, Sittka A, et al. The primary transcriptome ofthe major human pathogen Helicobacter pylori. Nature2010; 464:250-5; PMID:20164839; http://dx.doi.org/10.1038/nature08756
6. Liu JM, Camilli A. A broadening world of bacterial smallRNAs. Curr Opin Microbiol 2010; 13:18-23; PMID:20022798; http://dx.doi.org/10.1016/j.mib.2009.11.004
7. Schlüter JP, Reinkensmeier J, Daschkey S, Evguenieva-Hackenberg E, Janssen S, Jänicke S, et al. A genome-wide survey of sRNAs in the symbiotic nitrogen-fixingalpha-proteobacterium Sinorhizobium meliloti. BMCGenomics 2010; 11:245; PMID:20398411; http://dx.doi.org/10.1186/1471-2164-11-245
8. Axmann IM, Kensche P, Vogel J, Kohl S, Herzel H,Hess WR. Identification of cyanobacterial non-codingRNAs by comparative genome analysis. Genome Biol2005; 6:R73; PMID:16168080; http://dx.doi.org/10.1186/gb-2005-6-9-r73
9. Livny J, Teonadi H, Livny M, Waldor MK. High-throughput, kingdom-wide prediction and annotation ofbacterial non-coding RNAs. [Erratum in: PLoS One;3][11]. PLoS ONE 2008; 3:e3197; PMID:18787707;http://dx.doi.org/10.1371/journal.pone.0003197
10. Voss B, Georg J, Schon V, Ude S, Hess WR. Bio-computational prediction of non-coding RNAs in modelcyanobacteria. BMC Genomics 2009; 10:123; PMID:19309518; http://dx.doi.org/10.1186/1471-2164-10-123
11. Larimer FW, Chain P, Hauser L, Lamerdin J, MalfattiS, Do L, et al. Complete genome sequence of themetabolically versatile photosynthetic bacterium Rhodo-pseudomonas palustris. Nat Biotechnol 2004; 22:55-61;PMID:14704707; http://dx.doi.org/10.1038/nbt923
12. Oda Y, Larimer FW, Chain PS, Malfatti S, Shin MV,Vergez LM, et al. Multiple genome sequences revealadaptations of a phototrophic bacterium to sedimentmicroenvironments. Proc Natl Acad Sci USA 2008;105:18543-8; PMID:19020098; http://dx.doi.org/10.1073/pnas.0809160105
13. Giraud E, Moulin L, Vallenet D, Barbe V, Cytryn E,Avarre JC, et al. Legumes symbioses: absence of Nodgenes in photosynthetic bradyrhizobia. Science 2007;316:1307-12; PMID:17540897; http://dx.doi.org/10.1126/science.1139548
14. Kaneko T, Nakamura Y, Sato S, Minamisawa K,Uchiumi T, Sasamoto S, et al. Complete genomicsequence of nitrogen-fixing symbiotic bacteriumBradyrhizobium japonicum USDA 110. DNA Res2002; 9:189-97; PMID:12597275; http://dx.doi.org/10.1093/dnares/9.6.189
15. Becker A, Bergès H, Krol E, Bruand C, Rüberg S,Capela D, et al. Global changes in gene expression inSinorhizobium meliloti 1021 under microoxic andsymbiotic conditions. Mol Plant Microbe Interact2004; 17:292-303; PMID:15000396; http://dx.doi.org/10.1094/MPMI.2004.17.3.292
16. Pessi G, Ahrens CH, Rehrauer H, Lindemann A,Hauser F, Fischer HM, et al. Genome-wide transcriptanalysis of Bradyrhizobium japonicum bacteroids insoybean root nodules. Mol Plant Microbe Interact2007; 20:1353-63; PMID:17977147; http://dx.doi.org/10.1094/MPMI-20-11-1353
17. Ebeling S, Kündig C, Hennecke H. Discovery of arhizobial RNA that is essential for symbiotic rootnodule development. J Bacteriol 1991; 173:6373-82;PMID:1717438
18. Keiler KC, Shapiro L, Williams KP. tmRNAs thatencode proteolysis-inducing tags are found in all knownbacterial genomes: A two-piece tmRNA functions inCaulobacter. Proc Natl Acad Sci USA 2000; 97:7778-83; PMID:10884408; http://dx.doi.org/10.1073/pnas.97.14.7778
19. Wassarman KM. 6S RNA: a regulator of transcription.Mol Microbiol 2007; 65:1425-31; PMID:17714443;http://dx.doi.org/10.1111/j.1365-2958.2007.05894.x
20. Wassarman KM, Storz G. 6S RNA regulates E. coliRNA polymerase activity. Cell 2000; 101:613-23;PMID:10892648; http://dx.doi.org/10.1016/S0092-8674(00)80873-9
21. Barrick JE, Sudarsan N, Weinberg Z, Ruzzo WL,Breaker RR. 6S RNA is a widespread regulator ofeubacterial RNA polymerase that resembles an openpromoter. RNA 2005; 11:774-84; PMID:15811922;http://dx.doi.org/10.1261/rna.7286705
22. Voss B, Hölscher M, Baumgarth B, Kalbfleisch A, KayaC, Hess WR, et al. Expression of small RNAs inRhizobiales and protection of a small RNA and itsdegradation products by Hfq in Sinorhizobium meliloti.Biochem Biophys Res Commun 2009; 390:331-6;PMID:19800865; http://dx.doi.org/10.1016/j.bbrc.2009.09.125
23. del Val C, Rivas E, Torres-Quesada O, Toro N, Jimenez-Zurdo JI. Identification of differentially expressedsmall non-coding RNAs in the legume endosymbiontSinorhizobium meliloti by comparative genomics. MolMicrobiol 2007; 66:1080-91; PMID:17971083; http://dx.doi.org/10.1111/j.1365-2958.2007.05978.x
24. Ulvé VM, Sevin EW, Chéron A, Barloy-Hubler F.Identification of chromosomal alpha-proteobacterialsmall RNAs by comparative genome analysis anddetection in Sinorhizobium meliloti strain 1021. BMCGenomics 2007; 8:467; PMID:18093320; http://dx.doi.org/10.1186/1471-2164-8-467
25. Valverde C, Livny J, Schluter JP, Reinkensmeier J,Becker A, Parisi G. Prediction of Sinorhizobium melilotisRNA genes and experimental detection in strain 2011.BMC Genomics 2008; 9:416; PMID:18793445;http://dx.doi.org/10.1186/1471-2164-9-416
www.landesbioscience.com RNA Biology 57

© 2012 Landes Bioscience.
Do not distribute.
26. Landt SG, Abeliuk E, McGrath PT, Lesley JA,McAdams HH, Shapiro L. Small non-coding RNAsin Caulobacter crescentus. Mol Microbiol 2008; 68:600-14; PMID:18373523; http://dx.doi.org/10.1111/j.1365-2958.2008.06172.x
27. Berghoff BA, Glaeser J, Sharma CM, Vogel J, Klug G.Photooxidative stress-induced and abundant small RNAsin Rhodobacter sphaeroides. Mol Microbiol 2009; 74:1497-512; PMID:19906181; http://dx.doi.org/10.1111/j.1365-2958.2009.06949.x
28. Vercruysse M, Fauvart M, Cloots L, Engelen K, ThijsIM, Marchal K, et al. Genome-wide detection ofpredicted non-coding RNAs in Rhizobium etli expressedduring free-living and host-associated growth using ahigh-resolution tiling array. BMC Genomics 2010;11:53; PMID:20089193; http://dx.doi.org/10.1186/1471-2164-11-53
29. Wilms I, Voss B, Hess WR, Leichert IL, Narberhaus F.Small RNA-mediated control of the Agrobacteriumtumefaciens GABA receptor. Mol Microbiol 2011;80:492-506; PMID:21320185; http://dx.doi.org/10.1111/j.1365-2958.2011.07589.x
30. Hauser F, Pessi G, Friberg M, Weber C, Rusca N,Lindemann A, et al. Dissection of the Bradyrhizobiumjaponicum NifA+σ54 regulon, and identification of aferredoxin gene (fdxN) for symbiotic nitrogen fixation.MolGenet Genomics 2007; 278:255-71; PMID:17569992;http://dx.doi.org/10.1007/s00438-007-0246-9
31. Gardner PP, Daub J, Tate JG, Nawrocki EP, Kolbe DL,Lindgreen S, et al. Rfam: updates to the RNA familiesdatabase. Nucleic Acids Res 2009; 37:D136-40; PMID:18953034; http://dx.doi.org/10.1093/nar/gkn766
32. Corbino KA, Barrick JE, Lim J, Welz R, Tucker BJ,Puskarz I, et al. Evidence for a second class ofS-adenosylmethionine riboswitches and other regula-tory RNA motifs in alpha-proteobacteria. Genome Biol2005; 6:R70; PMID:16086852; http://dx.doi.org/10.1186/gb-2005-6-8-r70
33. Kündig C, Hennecke H, Göttfert M. Correlated physicaland genetic map of the Bradyrhizobium japonicum 110genome. J Bacteriol 1993; 175:613-22; PMID:8423135
34. Delmotte N, Ahrens CH, Knief C, Qeli E, Koch M,Fischer HM, et al. An integrated proteomics andtranscriptomics reference data set provides new insightsinto the Bradyrhizobium japonicum bacteroid meta-bolism in soybean root nodules. Proteomics 2010;10:1391-400; PMID:20104621; http://dx.doi.org/10.1002/pmic.200900710
35. Romeo T. Global regulation by the small RNA-bindingprotein CsrA and the non-coding RNA molecule CsrB.Mol Microbiol 1998; 29:1321-30; PMID:9781871;http://dx.doi.org/10.1046/j.1365-2958.1998.01021.x
36. Tjaden B. TargetRNA: a tool for predicting targets ofsmall RNA action in bacteria. Nucleic Acids Res 2008;36:W109-113; PMID:18477632; http://dx.doi.org/10.1093/nar/gkn264
37. Smith C, Heyne S, Richter AS, Will S, Backofen R.Freiburg RNA Tools: a web server integratingINTARNA, EXPARNA and LOCARNA. NucleicAcids Res 2010; 38:W373-377; PMID:20444875;http://dx.doi.org/10.1093/nar/gkq316
38. Vockenhuber MP, Sharma CM, Statt MG, Schmidt D,Xu Z, Dietrich S, et al. Deep sequencing-based identifica-tion of small non-coding RNAs in Streptomyces coelicolor.RNA Biol 2011; 8:468-77; PMID:21521948; http://dx.doi.org/10.4161/rna.8.3.14421
39. Koch M, Delmotte N, Rehrauer H, Vorholt JA, PessiG, Hennecke H. Rhizobial adaptation to hosts, a newfacet in the legume root-nodule symbiosis. Mol PlantMicrobe Interact 2010; 23:784-90; PMID:20459317;http://dx.doi.org/10.1094/MPMI-23-6-0784
40. Backofen R, Hess WR. Computational prediction ofsRNAs and their targets in bacteria. RNA Biol 2010;7:33-42; PMID:20061798; http://dx.doi.org/10.4161/rna.7.1.10655
41. Papenfort K, Said N, Welsink T, Lucchini S, HintonJC, Vogel J. Specific and pleiotropic patterns of mRNAregulation by ArcZ, a conserved, Hfq-dependentsmall RNA. Mol Microbiol 2009; 74:139-58; PMID:19732340; http://dx.doi.org/10.1111/j.1365-2958.2009.06857.x
42. Croucher NJ, Thomson NR. Studying bacterialtranscriptomes using RNA-seq. Curr Opin Microbiol2010; 13:619-24; PMID:20888288; http://dx.doi.org/10.1016/j.mib.2010.09.009
43. Altuvia S, Zhang A, Argaman L, Tiwari A, Storz G. TheEscherichia coli OxyS regulatory RNA represses fhlAtranslation by blocking ribosome binding. EMBO J1998; 17:6069-75; PMID:9774350; http://dx.doi.org/10.1093/emboj/17.20.6069
44. Majdalani N, Cunning C, Sledjeski D, Elliott T,Gottesman S. DsrA RNA regulates translation of RpoSmessage by an anti-antisense mechanism, independentof its action as an antisilencer of transcription. ProcNatl Acad Sci USA 1998; 95:12462-7; PMID:9770508; http://dx.doi.org/10.1073/pnas.95.21.12462
45. Zawoznik MS, Tomaro ML. Effect of chlorimuron-ethyl on Bradyrhizobium japonicum and its symbiosiswith soybean. Pest Manag Sci 2005; 61:1003-8;PMID:15920784; http://dx.doi.org/10.1002/ps.1077
46. Janczarek M, Skorupska A. The Rhizobium legumino-sarum bv. trifolii pssB gene product is an inositolmonophosphatase that influences exopolysaccharidesynthesis. Arch Microbiol 2001; 175:143-51; PMID:11285742; http://dx.doi.org/10.1007/s002030000250
47. Janczarek M, Skorupska A. Regulation of pssA and pssBgene expression in Rhizobium leguminosarum bv. trifoliiin response to environmental factors. Antonie van Leeu-wenhoek 2004; 85:217-27; PMID:15031650; http://dx.doi.org/10.1023/B:ANTO.0000020342.39867.fb
48. Larkin MA, Blackshields G, Brown NP, Chenna R,McGettigan PA, McWilliam H, et al. Clustal Wand Clustal X version 2.0. Bioinformatics 2007; 23:2947-8; PMID:17846036; http://dx.doi.org/10.1093/bioinformatics/btm404
49. Gruber AR, Findeiß S, Washietl S, Hofacker IL, StadlerPF. RNAZ 2.0: improved noncoding RNA detection.Pac Symp Biocomput 2010; 15:69-79; PMID:19908359
50. Zuker M. Mfold web server for nucleic acid folding andhybridization prediction. Nucleic Acids Res 2003;31:3406-15; PMID:12824337; http://dx.doi.org/10.1093/nar/gkg595
51. Bernhart SH, Hofacker IL, Will S, Gruber AR, StadlerPF. RNAalifold: improved consensus structure predic-tion for RNA alignments. BMC Bioinformatics 2008;9:474; PMID:19014431; http://dx.doi.org/10.1186/1471-2105-9-474
52. Washietl S, Findeiß S, Müller SA, Kalkhof S, von BergenM, Hofacker IL, et al. RNAcode: Robust discrimina-tion of coding and noncoding regions in compara-tive sequence data. RNA 2011; 17:578-94; PMID:21357752; http://dx.doi.org/10.1261/rna.2536111
53. Regensburger B, Hennecke H. RNA polymerase fromRhizobium japonicum. Arch Microbiol 1983; 135:103-9;PMID:6639271; http://dx.doi.org/10.1007/BF00408017
54. Evguenieva-Hackenberg E, Klug G. RNase III proces-sing of intervening sequences found in helix 9 of23S rRNA in the alpha subclass of Proteobacteria.J Bacteriol 2000; 182:4719-29; PMID:10940010;http://dx.doi.org/10.1128/JB.182.17.4719-4729.2000
55. Mesa S, Hauser F, Friberg M, Malaguti E, FischerHM, Hennecke H. Comprehensive assessment of theregulons controlled by the FixLJ-FixK2-FixK1 cascade inBradyrhizobium japonicum. J Bacteriol 2008; 190:6568-79; PMID:18689489; http://dx.doi.org/10.1128/JB.00748-08
56. Göttfert M, Hitz S, Hennecke H. Identification of nodSand nodU, two inducible genes inserted between theBradyrhizobium japonicum nodYABC and nodIJ genes.Mol Plant Microbe Interact 1990; 3:308-16; PMID:2134855; http://dx.doi.org/10.1094/MPMI-3-308
57. Drews G. Mikrobiologisches Praktikum, 3rd ed.Springer-Verlag, Berlin, Germany, 1976.
58. Madhugiri R, Evguenieva-Hackenberg E. RNase J isinvolved in the 5'-end maturation of 16S rRNA and23S rRNA in Sinorhizobium meliloti. FEBS Lett 2009;583:2339-42; PMID:19540834; http://dx.doi.org/10.1016/j.febslet.2009.06.026
59. Sittka A, Lucchini S, Papenfort K, Sharma CM, RolleK, Binnewies TT, et al. Deep sequencing analysis ofsmall noncoding RNA and mRNA targets of the globalpost-transcriptional regulator, Hfq. PLoS Genet 2008;4:e1000163; PMID:18725932; http://dx.doi.org/10.1371/journal.pgen.1000163
60. Hoffmann S, Otto C, Kurtz S, Sharma CM, KhaitovichP, Vogel J, et al. Fast mapping of short sequenceswith mismatches, insertions and deletions using indexstructures. PLOS Comput Biol 2009; 5:e1000502;PMID:19750212; http://dx.doi.org/10.1371/journal.pcbi.1000502
58 RNA Biology Volume 9 Issue 1
Related Documents











