Small RNAs containing the pathogenic determinant of a chloroplast-replicating viroid guide the degradation of a host mRNA as predicted by RNA silencing Beatriz Navarro 1 , Andreas Gisel 2 , Maria Elena Rodio 1 , Sonia Delgado 3 , Ricardo Flores 3,* and Francesco Di Serio 1,* 1 Istituto di Virologia Vegetale (CNR), Unita ` Organizzativa di Bari, Via Amendola 165/A, 70126 Bari, Italy, 2 Istituto di Tecnologie Biomediche (CNR), Unita ` Organizzativa di Bari, Via Amendola 122/D, 70126 Bari, Italy, and 3 Instituto de Biologı´a Molecular y Celular de Plantas (UPV-CSIC), Campus Universidad Polite ´ cnica, Avenida de los Naranjos, 46022 Valencia, Spain Received 27 January 2012; accepted 9 February 2012; published online 2 April 2012. *For correspondence (e-mail [email protected]; rfl[email protected]). GenBank accession numbers for the nucleotide sequences reported: JN377825–JN377892. SUMMARY How viroids, tiny non-protein-coding RNAs (250–400 nt), incite disease is unclear. One hypothesis is that viroid-derived small RNAs (vd-sRNAs; 21–24 nt) resulting from the host defensive response, via RNA silencing, may target for cleavage cell mRNAs and trigger a signal cascade, eventually leading to symptoms. Peach latent mosaic viroid (PLMVd), a chloroplast-replicating viroid, is particularly appropriate to tackle this question because it induces an albinism (peach calico, PC) strictly associated with variants containing a specific 12–14-nt hairpin insertion. By dissecting albino and green leaf sectors of Prunus persica (peach) seedlings inoculated with PLMVd natural and artificial variants, and cloning their progeny, we have established that the hairpin insertion sequence is involved in PC. Furthermore, using deep sequencing, semi-quantitative RT-PCR and RNA ligase-mediated rapid amplification of cDNA ends (RACE), we have determined that two PLMVd-sRNAs containing the PC-associated insertion (PC-sRNA8a and PC-sRNA8b) target for cleavage the mRNA encoding the chloroplastic heat-shock protein 90 (cHSP90), thus implicating RNA silencing in the modulation of host gene expression by a viroid. Chloroplast malformations previously reported in PC-expressing tissues are consistent with the downregulation of cHSP90, which participates in chloroplast biogenesis and plastid-to- nucleus signal transduction in Arabidopsis. Besides PC-sRNA8a and PC-sRNA8b, both deriving from the less- abundant PLMVd ()) strand, we have identified other PLMVd-sRNAs potentially targeting peach mRNAs. These results also suggest that sRNAs derived from other PLMVd regions may downregulate additional peach genes, ultimately resulting in other symptoms or in a more favorable host environment for viroid infection. Keywords: chloroplast development, heat-shock protein 90, non-coding RNAs, PLMVd, RNA silencing, viroid pathogenesis. INTRODUCTION Viroids are minimal RNAs that infect and often cause severe diseases in plants (Flores et al., 2005; Tsagris et al., 2008; Ding, 2009). Based on their properties, among which the ability to replicate through specific pathways of a rolling- circle mechanism in certain subcellular compartments is key, viroid species are classified into two families. The family Pospiviroidae, type species Potato spindle tuber viroid (PSTVd) (Diener, 1972; Gross et al., 1978), clusters viroids replicating and accumulating in the nucleus, whereas the family Avsunviroidae, type species Avocado sunblotch viroid (ASBVd) (Symons, 1981; Hutchins et al., 1986), includes viroids with hammerhead ribozymes in both polarity strands that mediate self-cleavage of their replicative intermediates generated in plastids (mostly chloroplasts) (Flores et al., 2000). Despite being composed of just a small (246–401 nt), circular, non-protein-coding RNA, viroids can usurp and redirect the host machinery for completing their infectious cycle. Therefore, viroids largely differ from viruses, the replication, movement and pathogenesis of which partly rely on proteins encoded in their own genome. However, similarly to changes incited by viruses, viroid infections cause profound changes in their host homeostasis (Itaya ª 2012 The Authors 991 The Plant Journal ª 2012 Blackwell Publishing Ltd The Plant Journal (2012) 70, 991–1003 doi: 10.1111/j.1365-313X.2012.04940.x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Small RNAs containing the pathogenic determinant of achloroplast-replicating viroid guide the degradation of a hostmRNA as predicted by RNA silencing
Beatriz Navarro1, Andreas Gisel2, Maria Elena Rodio1, Sonia Delgado3, Ricardo Flores3,* and Francesco Di Serio1,*
1Istituto di Virologia Vegetale (CNR), Unita Organizzativa di Bari, Via Amendola 165/A, 70126 Bari, Italy,2Istituto di Tecnologie Biomediche (CNR), Unita Organizzativa di Bari, Via Amendola 122/D, 70126 Bari, Italy, and3Instituto de Biologıa Molecular y Celular de Plantas (UPV-CSIC), Campus Universidad Politecnica, Avenida de los Naranjos,
46022 Valencia, Spain
Received 27 January 2012; accepted 9 February 2012; published online 2 April 2012.
*For correspondence (e-mail [email protected]; [email protected]).
GenBank accession numbers for the nucleotide sequences reported: JN377825–JN377892.
SUMMARY
How viroids, tiny non-protein-coding RNAs (�250–400 nt), incite disease is unclear. One hypothesis is that
viroid-derived small RNAs (vd-sRNAs; 21–24 nt) resulting from the host defensive response, via RNA silencing,
may target for cleavage cell mRNAs and trigger a signal cascade, eventually leading to symptoms. Peach latent
mosaic viroid (PLMVd), a chloroplast-replicating viroid, is particularly appropriate to tackle this question
because it induces an albinism (peach calico, PC) strictly associated with variants containing a specific 12–14-nt
hairpin insertion. By dissecting albino and green leaf sectors of Prunus persica (peach) seedlings inoculated
with PLMVd natural and artificial variants, and cloning their progeny, we have established that the hairpin
insertion sequence is involved in PC. Furthermore, using deep sequencing, semi-quantitative RT-PCR and RNA
ligase-mediated rapid amplification of cDNA ends (RACE), we have determined that two PLMVd-sRNAs
containing the PC-associated insertion (PC-sRNA8a and PC-sRNA8b) target for cleavage the mRNA encoding
the chloroplastic heat-shock protein 90 (cHSP90), thus implicating RNA silencing in the modulation of host
gene expression by a viroid. Chloroplast malformations previously reported in PC-expressing tissues are
consistent with the downregulation of cHSP90, which participates in chloroplast biogenesis and plastid-to-
nucleus signal transduction in Arabidopsis. Besides PC-sRNA8a and PC-sRNA8b, both deriving from the less-
abundant PLMVd ()) strand, we have identified other PLMVd-sRNAs potentially targeting peach mRNAs.
These results also suggest that sRNAs derived from other PLMVd regions may downregulate additional peach
genes, ultimately resulting in other symptoms or in a more favorable host environment for viroid infection.
Keywords: chloroplast development, heat-shock protein 90, non-coding RNAs, PLMVd, RNA silencing, viroid
pathogenesis.
INTRODUCTION
Viroids are minimal RNAs that infect and often cause severe
diseases in plants (Flores et al., 2005; Tsagris et al., 2008;
Ding, 2009). Based on their properties, among which the
ability to replicate through specific pathways of a rolling-
circle mechanism in certain subcellular compartments is
key, viroid species are classified into two families. The family
Pospiviroidae, type species Potato spindle tuber viroid
(PSTVd) (Diener, 1972; Gross et al., 1978), clusters viroids
replicating and accumulating in the nucleus, whereas the
family Avsunviroidae, type species Avocado sunblotch
viroid (ASBVd) (Symons, 1981; Hutchins et al., 1986), includes
viroids with hammerhead ribozymes in both polarity strands
that mediate self-cleavage of their replicative intermediates
generated in plastids (mostly chloroplasts) (Flores et al.,
2000). Despite being composed of just a small (246–401 nt),
circular, non-protein-coding RNA, viroids can usurp and
redirect the host machinery for completing their infectious
cycle. Therefore, viroids largely differ from viruses, the
replication, movement and pathogenesis of which partly
rely on proteins encoded in their own genome. However,
similarly to changes incited by viruses, viroid infections
cause profound changes in their host homeostasis (Itaya
ª 2012 The Authors 991The Plant Journal ª 2012 Blackwell Publishing Ltd
The Plant Journal (2012) 70, 991–1003 doi: 10.1111/j.1365-313X.2012.04940.x

et al., 2002; Tessitori et al., 2007; Wang et al., 2011), which
ultimately result in the onset of visible symptoms.
RNA silencing, a regulatory network that modulates host
gene expression and protects plants and most other eukar-
yotes against invading nucleic acids, such as transposons,
viruses and transgenes, is triggered by double-stranded
RNAs (dsRNAs) and highly-structured single-stranded RNAs
(ssRNAs) that are processed by Dicer-like (DCL) RNases into
small RNAs (sRNAs) (Carthew and Sontheimer, 2009; Chen,
2009). The two major classes of sRNAs are microRNAs
(miRNAs) and small interfering RNAs (siRNAs), with host
RNA-dependent RNA polymerases (RDRs) mediating an
amplification circuit resulting into secondary siRNAs able
to promote RNA silencing in a non-cell-autonomous manner
(Voinnet, 2008; Dunoyer and Voinnet, 2009). The sRNAs are
loaded into AGO proteins and guide the RNA-inducing
silencing complex (RISC) to target their cognate RNAs or
DNAs (Mallory and Vaucheret, 2010).
To defend themselves, plant viruses encode in their
genomes proteins suppressing the RNA-silencing response
that they trigger in their hosts (Csorba et al., 2009; Ding,
2010). As a side effect, these RNA-silencing suppressors
(RSSs) may impair host developmental pathways regulated
by RNA silencing, explaining at least in part the phenotypic
alterations accompanying most virus infections (Kasschau
et al., 2003; Jay et al., 2011). However, in the absence of
viroid-encoded RSSs, this model cannot account for symp-
toms induced by viroids. In addition to the conventional
model regarding a direct interaction of the genomic viroid
RNA with host proteins and/or RNAs as the primary event of
viroid pathogenesis (reviewed by Flores et al., 2005), an
intriguing alternative model was first proposed when viroid-
derived sRNAs (vd-sRNAs), structurally similar to sRNAs,
were detected in PSTVd-infected tissues (Papaefthimiou
et al., 2001). Accordingly, symptoms would result from
vd-sRNAs loading RISC and targeting specific host mRNAs
for inactivation. This hypothesis has been extended to
explain pathogenesis of satellite RNAs of plant viruses
(Wang et al., 2004) and to involve RDR6-derived secondary
siRNAs in viroid pathogenesis (Gomez et al., 2008).
Recently, direct experimental evidence has been supplied
showing that the yellow symptoms induced in tobacco by
the concurrent infection of Cucumber mosaic virus (CMV)
and its Y-satellite RNA (Y-sat) result from silencing the
chlorophyll biosynthetic gene CHLI by a 22-nt Ysat-derived
siRNA (Shimura et al., 2011; Smith et al., 2011). However, in
spite of multiple reports on the identification and character-
ization of vd-sRNAs accumulating in tissue infected by
nucleus- and chloroplast-replicating viroids (Papaefthimiou
et al., 2001; Martınez de Alba et al., 2002; Itaya et al., 2007;
Di Serio et al., 2009, 2010; Navarro et al., 2009; St-Pierre
et al., 2009; Bolduc et al., 2010; Martınez et al., 2010),
attempts to validate this hypothesis for viroids have failed
so far, with the evidence being circumstantial and restricted
to correlating symptom severity with vd-sRNA accumulation
(Markarian et al., 2004; Wang et al., 2004, 2011; Matousek
et al., 2007; Gomez et al., 2008; Diermann et al., 2010).
Moreover, in most instances a similar correlation also exists
with the genomic viroid RNA, making it difficult to draw
reliable inferences (Itaya et al., 2001; Carbonell et al., 2008;
Di Serio et al., 2010).
To address the issue of whether vd-sRNAs have a role in
pathogenesis, Peach latent mosaic viroid (PLMVd) (Hernan-
dez and Flores, 1992) seems particularly convenient. PLMVd,
a member of the family Avsunviroidae that replicates in the
chloroplast (Bussiere et al., 1999; Rodio et al., 2007), may
induce in its natural host (Prunus persica, peach) a broad
variety of symptoms that includes a severe albinism (peach
calico, PC) in leaves, stems and fruits. PC is associated with
variants of 348–351 nt containing a specific insertion of
12–14 nt (Malfitano et al., 2003) that is absent in latent and
mosaic-inducing variants of 335–338 nt (Ambros et al., 1998,
1999; Flores et al., 2006).
Interestingly, the pathogenic determinant for PC has been
mapped at this insertion, which folds into a hairpin capped
by a UUUU loop (Malfitano et al., 2003; Rodio et al., 2006).
Further dissection of the molecular pathway underlying PC
has shown that PLMVd variants inducing this syndrome
interfere with the maturation of plastid rRNAs, thus impair-
ing plastid translation and chloroplast biogenesis (Rodio
et al., 2007). The recent release of the complete sequence of
the peach genome (peach v1.0; International Peach Genome
Initiative, http://www.rosaceae.org/peach/genome), pro-
vides a critical resource for genome-wide analyses of
PLMVd-induced alterations in this host. Here, we report that
the sequence of the pathogenic determinant for PC is crucial
for eliciting this symptomatology, and supply direct evi-
dence supporting that vd-sRNAs containing the PC deter-
minant program RISC for cleaving a host mRNA, a finding
with potential implications on viroid pathogenesis.
RESULTS
Molecular dissection of the pathogenic determinant for PC
In a previous work we showed that, in addition to the cap-
ping UUUU loop, the stem of the hairpin insertion seems to
play a role in PC (Figure 1a) (Rodio et al., 2006). To further
investigate this role, we selected two natural PLMVd variants
with the same capping UUUU loop but differing stems: the
reference PC-inducing variant C40 with a 4-bp stem (Malfit-
ano et al., 2003) (Figure 1b, left), and variant P1.148 with a
3-bp stem that is unable to elicit this syndrome (Rodio et al.,
2006) (Figure 1c, left). Next, the C and A at position 349 in
variants C40 and P1.148, respectively, were changed into A
and C to generate the artificial variants C40(A349) and
P1.148(C349), in which the predicted stem of the hairpin
insertion was shorter and longer than in their respective
wild-type counterparts (Figure 1b–e, left panels). Slash
992 Beatriz Navarro et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

inoculation of dimeric transcripts from the natural and
mutated C40 and P1.148 variants, and dot-blot hybridization
of leaf RNA preparations, showed that the four variants were
highly infectious (the eight GF-305 peach seedlings of each
block became infected) and generated similar viroid titers
(data not shown). These plants were observed for symptom
expression and, to establish a relationship between the
structure and pathogenicity of the infecting variants, their
progeny were cloned and sequenced 6 months post inocu-
lation (6 mpi). To better define this relationship, the albino
sectors of symptomatic leaves were carefully separated
from the adjacent green tissues before extraction, as in a
previous study (Rodio et al., 2006). In addition, for getting a
more complete view of variant distribution within the
PC-expressing seedlings, leaves from an asymptomatic
(green) branch were also analyzed.
As anticipated, variant C40 elicited severe PC symptoms in
the eight inoculated seedlings, whereas variant P1.148 did
not cause any leaf symptom (Figure 1b,c). In line with earlier
results (Malfitano et al., 2003; Rodio et al., 2006), most C40
progeny variants accumulating in albino tissues presented a
minor modification (a G fi A substitution, not disrupting the
hairpin stem) (Figure 1b), whereas in two of the eight
sequenced variants additional minor modifications (includ-
ing a covariation and a substitution) enlarged the stem up to
5 bp (Figure 1b). Intriguingly, all PLMVd variants recovered
(a)
(b)
(c)
(d)
(e)
Figure 1. Peach phenotype and progeny of nat-
ural and artificial PLMVd variants with specific
hairpin insertions.
(a) Partial and complete albinism induced by
variant C40 (left), and schematic representation
of the secondary structure of PLMVd, with the
hairpin insertion represented with a broken line
(right).
(b–e) Results of bioassays with the natural vari-
ants C40, P1.148 (b, c) and their mutant deriva-
tives C40(A349) and P1.148(C349) (d, e), the
hairpin insertion of which (with light-blue back-
ground) plus the flanking 5¢ and 3¢ nucleotides
are depicted on the left. Nucleotides in blue (d, e,
left) denote changes introduced by site-directed
mutagenesis at position 349 in variants C40 and
P1.148 (b, c, left), respectively. Nucleotides in red
denote recurring substitutions emerging natu-
rally with respect to the parental hairpin inser-
tions, with additional, less frequent substitutions
indicated with circles, together with the variants
in which they were found. The ratio of symp-
tomatic to infected seedlings is reported on the
top of each panel in brackets. The sequence of
the hairpin insertions and the name of the
progeny variants recovered from albino (a) and/
or green (g) tissues of one representative inoc-
ulated seedling are also indicated (b–e, middle
and/or left).
Viroid-induced RNA silencing of host mRNA 993
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

from the asymptomatic branch of the same plant contained
a similar extended 5-bp hairpin stem (Figure 1b, right panel).
On the other hand, half of P1.148 progeny variants preserved
the hairpin folding of the parental insertion, but the other
half showed longer or shorter stems (Figure 1c).
Unexpectedly, all seedlings inoculated with the mutant
variant C40(A349), and six of eight seedlings inoculated with
the mutant variant P1.148(C349), also displayed PC (Fig-
ure 1d,e). However, the two parental variants were not
recovered in their respective progeny. Instead, in both cases,
the hairpin insertions of variants accumulating in albino and
green tissues were similar, respectively, to those of variants
from albino and green tissues of seedlings inoculated with
their corresponding natural variants (Figure 1, compare
panels d and e with b and c).
Altogether, results from these bioassays show that the
typical hairpin insertion (composed of a UUUU loop and a
4-bp stem previously associated with the pathogenic deter-
minant for PC) was prevalent in PLMVd progeny variants
from albino tissues, irrespective of the specific hairpin
insertion of the inoculated natural or mutant variant. In
contrast, and also irrespective of the specific hairpin inser-
tion of the inoculated variant, PLMVd progeny from green
tissues mostly presented a hairpin insertion with a UUUU
loop but with a 5-bp stem resulting from an additional A:U
pair. These data strongly suggested that the latter variants
did not incite PC. To provide direct support for this hypoth-
esis, we inoculated eight GF-305 peach seedlings with a
dimeric transcript of variant C40(A349)-g12, recovered from
the progeny of variant C40(A349) in the previous experi-
ment, and with an additional A:U pair in the 5-bp stem
(Figure 1d, right panel). All inoculated seedlings remained
asymptomatic and most progeny variants presented the
same hairpin insertion (Figure S1), thereby showing that
variants of this kind cannot elicit PC.
Collectively, these data support the view that the struc-
tural requirements conferring PC depend not only on the
capping loop (UUUU) of the hairpin insertion, but also on
the stem composition (Rodio et al., 2006). However, at this
stage we could not clearly discern whether the sequence or
the specific morphology of the hairpin insertion were
required for inducing PC.
The primary rather than the secondary structure of the
pathogenic determinant plays a major role in PC
We showed previously that the insertion found in some
PLMVd variants always adopts a hairpin folding (Malfitano
et al., 2003; Rodio et al., 2006). To examine whether the
sequence of this hairpin also plays a role in PC, we generated
the artificial variant C40(stem), which differs from its
parental C40 in having the eight nucleotides forming the
stem interchanged (Figure 2). Therefore, the sequence of the
12-nt hairpin insertion of variants C40(stem) and C40 are
largely different, whereas their secondary structure is pre-
served.
Then, dimeric transcripts of these two variants were slash-
inoculated in two blocks of eight peach seedlings each. Dot-
blot hybridization showed that all inoculated plants were
infected at 6 mpi, and that their viroid titer was similar (data
not shown). Remarkably, plants inoculated with variant
C40(stem) remained asymptomatic, with all progeny vari-
ants preserving the stem stability and some displaying
mutations in the capping loop (Figure 2). In contrast, control
seedlings inoculated with variant C40 expressed typical PC
symptoms. Therefore, these data show that preservation of
the secondary structure (folding into a hairpin with a
capping UUUU loop) is not sufficient for conferring PC
pathogenicity to this structural element, and strongly sup-
port that its primary structure, particularly the 12-nt
sequence GA(A/G)CUUUUGUUC (hereafter named PC-asso-
ciated insertion) characteristic of variants from albino
tissues, is critical in this respect.
PLMVd-sRNAs derived from the pathogenic determinant for
PC accumulate during infection
In view of the apparent role played by the sequence of the
PC-associated insertion, we examined whether pathogenesis
could operate by the silencing-based model proposed pre-
viously for viroids and plant satellite RNAs (Wang et al.,
2004). Adapting this model to our system implicitly entails
that (+) or ()) PLMVd-sRNAs involved in PC should contain,
at least partially, the PC-associated insertion.
To explore this possibility we subjected the sRNAs
accumulating in PC-expressing seedlings to deep sequencing.
Inoculated Progeny in green tissue
C40(stem)-g03, C40(stem)-g09, C40(stem)-g11
C40(stem)
U U U
U G C .
A U .
G C .
A C
G . U
C40(stem) (symptomatic/infected: 0/8)
U C U
U G C .
A U .
G C .
U C
G . C
Ag11 A g11
U g09 U
U U U
G C .
A U .
G C .
A C
A . U
A g07 g08
U C U A
G C .
A U .
A U .
A C
G . C
C40(stem)-g06, C40(stem)-g07, C40(stem)-g08
U U U
U G C .
A U .
A U .
A C
G . C
C40(stem)-g02 C40(stem)-g12
A-
Figure 2. Peach phenotype and progeny of the artificial PLMVd variant C40(stem).
Nucleotides in blue (left) denote changes introduced by site-directed mutagenesis in the hairpin insertion of natural variant C40 to interchange the nucleotides of the
hairpin stem, preserving its stability. Other details are as described in the legend to Figure 1.
994 Beatriz Navarro et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

This approach has already been adopted in a study aimed at
comparing the vd-sRNAs accumulating in leaves of GF-305
peach seedlings infected with the PC-inducing variant C40
and a mosaic-inducing PLMVd variant (GDS6) (Di Serio
et al., 2009). This previous C40 sRNAs library was prepared
from symptomatic leaves that also contained green sectors.
Considering the role of the PC-associated insertion in
pathogenesis and the uneven distribution of variants with
this structural element in symptomatic plants (see above),
we generated a C40 sRNA library from albino leaf tissues
carefully separated from the surrounding green sectors in
order to enrich it with vd-sRNAs from variants containing the
PC-associated insertion. In parallel, we also generated an
sRNAs library from leaves of a mock-inoculated peach
seedling as a negative control. Gel-purified sRNAs were
linked to bar-coded adaptors for deep sequencing of both
libraries in the same channel.
Of approximately 20 815 300 reads, 96.2% were attribu-
table to the two bar-coded sRNA preparations (53.2 and
46.8% for the C40 and the mock-inoculated sample, respec-
tively). Reads for sRNAs between 18 and 26 nt (9 014 628
and 7 557 838 for the C40 and the mock-inoculated sample,
respectively), adopted a profile with two prominent 21- and
24-nt peaks in both cases (Figure S2). When PLMVd-sRNAs
matching perfectly the (+) and ()) sequences of variant C40
and its progeny (Figure 1b) were searched, 1 925 916 reads
(21.3% of the total generated by sRNAs between 18 and
26 nt) were retrieved from the C40 sample, corresponding to
8015 non-redundant sequences (Appendix S1), with minimal
PLMVd-related sequences detected in the mock-inoculated
sample (3961 reads representing 0.05% of the total, 611 of
which were non-redundant); the latter reads most likely
result from a minor contamination, because none of these
PLMVd-related sequences matched perfectly with the peach
genome. In line with results of the previous deep-sequenc-
ing experiment (Di Serio et al., 2009), (+) PLMVd-sRNAs
(57.6%) were slightly more abundant than their ()) counter-
parts (42.4%), with a similar polarity distribution being
observed after segregating PLMVd-sRNAs in size classes
(Figure S3). Moreover, also in line with previous results (Di
Serio et al., 2009), size classes of 21 and 22 nt were largely
prevalent (64.5 and 23.8%, respectively) within the PLMVd-
sRNAs population, whereas a minor fraction of 20-, 23- and
24-nt vd-sRNAs (around 10.2% altogether) was retrieved
(Figure S3). Mapping the PLMVds-RNA reads along the
genomic (+) and ()) RNAs revealed specific hot-spot profiles
(Figure S4). A parallel mapping of the non-redundant
PLMVd-sRNAs also revealed profiles with hot spots corre-
sponding, as expected, to genomic regions with high
nucleotide variability in the progeny variants (Figure S5).
A search for PLMVd-sRNAs spanning totally or partially (in
at least one nucleotide) the sequence of the PC-associated
insertion from C40 and its progeny variants revealed 111
non-redundant (+) and ()) 21-nt RNAs (6993 reads). Addi-
tionally, 100 non-redundant (+) and ()) vd-sRNAs of 22 nt
(6799 reads) were mapped at the same structural element.
These data show that the PC-associated insertion is indeed a
source of vd-sRNAs (hereafter referred to as PC-sRNAs),
although at this stage we did not know whether, like genuine
miRNAs, they might target host mRNAs for inactivation.
Host mRNAs are potentially targeted by PC-sRNAs
To search transcripts that could be targeted by PLMVd-
sRNAs for RISC-mediated degradation, we took advantage
of the complete sequence of the peach genome released
recently (peach v1.0; International Peach Genome Initiative,
http://www.rosaceae.org/peach/genome), and applied the
RNAhybrid program (Rehmsmeier et al., 2004). The search
was restricted to sRNAs of 21 nt because this is the prevalent
size of PLMVd-sRNAs and plant miRNAs (Di Serio et al.,
2009; St-Pierre et al., 2009; Figure S3). Moreover, we took
into consideration that miRNA-directed mRNA cleavage in
plants requires a high degree of base pairing, and that
mismatches in bona fide miRNA:target duplexes are mostly
located in the first position and after position 13 relative to
the 5¢ terminus of the miRNA (Mallory et al., 2004). There-
fore, a position-dependent scoring matrix for predicting
plant miRNA targets (Fahlgren and Carrington, 2010)
was applied to all possible duplexes formed by 21-nt
PLMVd-sRNAs and predicted peach mRNAs identified as
potential targets. A maximum score cut-off of 2.5, which is
below the cut-off of 3.5 established for reliable sensitivity
and specificity in Arabidopsis thaliana (Fahlgren and
Carrington, 2010), was adopted. This cut-off of 2.5 seemed
appropriate for our search because the duplexes between
miRNAs identified in our libraries conserved in other species
(including miR156, miR159, miR160, miR162, miR164,
miR166, miR168, miR171, miR172, miR393, miR394, miR395,
miR396 and miR397) and their potential targets in the com-
plete peach genome showed scores from 0 to 3. The search
resulted in 65 predicted peach mRNA targets, most encoding
proteins with functional annotation (P. persica – JGI v1.0;
http://www.phytozome.net/cgi-bin/gbrowse/peach; Table
S1), and 30 of them having been detected as peach tran-
scripts (peach assembled expressed sequence tags, ESTs,
from peachv1pasa_assemblies).
Considering that the sequence of the PC-associated
insertion is critical for eliciting PC, and that this insertion is
a source of vd-sRNAs (PC-sRNAs) in albino tissues (see
above), we next concentrated on the duplexes formed by
PC-sRNAs and peach mRNAs detected as transcripts
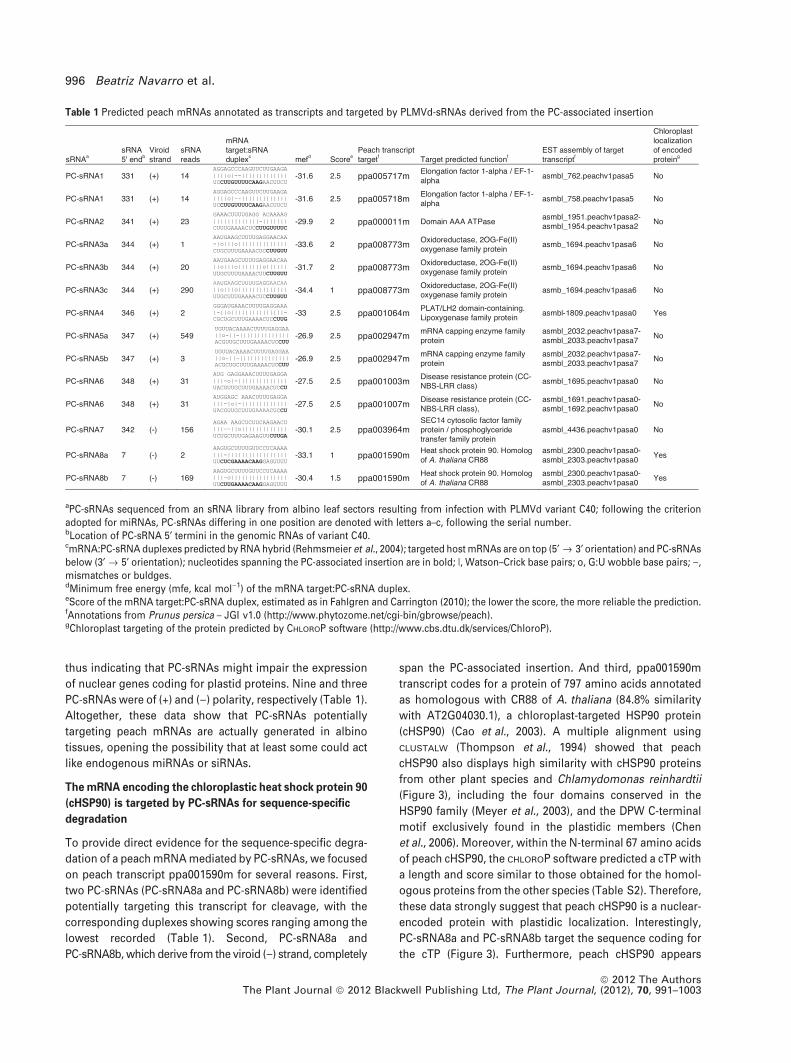
(Table 1). Overall, 12 PC-sRNAs were found potentially
targeting 10 predicted peach mRNAs because PC-sRNA1
and PC-sRNA6 could target two different mRNAs (Table 1).
This table also shows that two of the ten encoded proteins
contain a predicted chloroplastic transit peptide (cTP) (Li and
Chiu, 2010), according to CHLOROP software (http://www.
cbs.dtu.dk/services/ChloroP) (Emanuelsson et al., 1999),
Viroid-induced RNA silencing of host mRNA 995
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003
thus indicating that PC-sRNAs might impair the expression
of nuclear genes coding for plastid proteins. Nine and three
PC-sRNAs were of (+) and ()) polarity, respectively (Table 1).
Altogether, these data show that PC-sRNAs potentially
targeting peach mRNAs are actually generated in albino
tissues, opening the possibility that at least some could act
like endogenous miRNAs or siRNAs.
The mRNA encoding the chloroplastic heat shock protein 90
(cHSP90) is targeted by PC-sRNAs for sequence-specific
degradation
To provide direct evidence for the sequence-specific degra-
dation of a peach mRNA mediated by PC-sRNAs, we focused
on peach transcript ppa001590m for several reasons. First,
two PC-sRNAs (PC-sRNA8a and PC-sRNA8b) were identified
potentially targeting this transcript for cleavage, with the
corresponding duplexes showing scores ranging among the
lowest recorded (Table 1). Second, PC-sRNA8a and
PC-sRNA8b, which derive from the viroid ()) strand, completely
span the PC-associated insertion. And third, ppa001590m
transcript codes for a protein of 797 amino acids annotated
as homologous with CR88 of A. thaliana (84.8% similarity
with AT2G04030.1), a chloroplast-targeted HSP90 protein
(cHSP90) (Cao et al., 2003). A multiple alignment using
CLUSTALW (Thompson et al., 1994) showed that peach
cHSP90 also displays high similarity with cHSP90 proteins
from other plant species and Chlamydomonas reinhardtii
(Figure 3), including the four domains conserved in the
HSP90 family (Meyer et al., 2003), and the DPW C-terminal
motif exclusively found in the plastidic members (Chen
et al., 2006). Moreover, within the N-terminal 67 amino acids
of peach cHSP90, the CHLOROP software predicted a cTP with
a length and score similar to those obtained for the homol-
ogous proteins from the other species (Table S2). Therefore,
these data strongly suggest that peach cHSP90 is a nuclear-
encoded protein with plastidic localization. Interestingly,
PC-sRNA8a and PC-sRNA8b target the sequence coding for
the cTP (Figure 3). Furthermore, peach cHSP90 appears
Table 1 Predicted peach mRNAs annotated as transcripts and targeted by PLMVd-sRNAs derived from the PC-associated insertion
aPC-sRNAs sequenced from an sRNA library from albino leaf sectors resulting from infection with PLMVd variant C40; following the criterionadopted for miRNAs, PC-sRNAs differing in one position are denoted with letters a–c, following the serial number.bLocation of PC-sRNA 5¢ termini in the genomic RNAs of variant C40.cmRNA:PC-sRNA duplexes predicted by RNA hybrid (Rehmsmeier et al., 2004); targeted host mRNAs are on top (5¢ fi 3¢ orientation) and PC-sRNAsbelow (3¢ fi 5¢ orientation); nucleotides spanning the PC-associated insertion are in bold; |, Watson–Crick base pairs; o, G:U wobble base pairs; ),mismatches or buldges.dMinimum free energy (mfe, kcal mol)1) of the mRNA target:PC-sRNA duplex.eScore of the mRNA target:PC-sRNA duplex, estimated as in Fahlgren and Carrington (2010); the lower the score, the more reliable the prediction.fAnnotations from Prunus persica – JGI v1.0 (http://www.phytozome.net/cgi-bin/gbrowse/peach).gChloroplast targeting of the protein predicted by CHLOROP software (http://www.cbs.dtu.dk/services/ChloroP).
996 Beatriz Navarro et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

particularly appealing in the context of PC pathogenesis
because its homologous CR88 is required for chloroplast
biogenesis in A. thaliana (Cao et al., 2003); this protein,
perhaps interacting with HSP70, may be needed for assem-
bling the core of a multichaperone complex involved in the
biogenesis/maintenance of thylakoid membranes (Schroda
and Muhlhaus, 2009), which is the developmental pathway
specifically altered in PC-expressing tissues (Rodio et al., 2007).
We next proceeded comparing the steady-state level of
cHSP90 mRNA in mock-inoculated and PLMVd-infected
peach seedlings expressing PC. In line with data from the
peach EST database, RT-PCR amplification of a 405-bp cDNA
corresponding to the 5¢ terminus of the peach mRNA for
cHSP90, followed by cloning and sequencing, showed that
the gene cHSP90 is indeed transcribed in the mock-inocu-
lated control. However, RT-PCR estimates using the same
primers and serial cDNA dilutions showed a substantially
lower steady-state level of the cHSP90 transcript in albino
leaf sectors than in green tissues [from the same
C40-inoculated seedling, or from a mock-inoculated seed-
ling or a seedling infected by the latent variant C40(A349)-
g12]. As additional controls we also tested the levels of the
transcripts from genes psbA and rpoB, which were lower
and higher in albino than in green tissues, respectively
(Figure 4), in agreement with a previous report showing that
mRNAs from genes transcribed by the plastid-encoded
polymerase (like psbA) are negatively affected in albino
tissues, whereas those transcribed by the nuclear-encoded
polymerase (like rpoB) over-accumulate (Rodio et al., 2007).
The lower accumulation of the cHSP90 transcript in albino
tissues is consistent with its targeting by PC-sRNA8a and
PC-sRNA8b for RISC-mediated degradation (Figure 5). This
hypothesis was validated by RNA ligase-mediated rapid
amplification of cDNA ends (RLM-RACE) experiments with
RNA preparations from albino tissues: four out of five cDNA
clones were from fragments of the cHSP90 transcript with
5¢ termini identical to the cleavage site predicted by
PC-sRNA8a and PC-sRNA8b (Figure 5), between positions
10 and 11 from their 5¢ termini, with the cleavage site
inferred from the other clone mapping at one nucleotide
| Nucleotide binding domainA. thaliana -MAPALSRSLYTSPLTSVPITP-----VSSRLSHLRSSFLPHGGALRTGVSCS---WNLEK--RCNRFAVKCDAAVAEK--ETTEEGSGEKFEYQAEVSRLLDLIVHSLYSHKEVFLREL 107P. persica -MAPVLSRSLATASLASLPSSSPFTLRNPSKALSLRSAFVPQNG-LRKGFSCGGLKWKLES--KNRGISIRCDAAVAEK--EATDT-PGEKFEYQAEVTRLMDLIVHSLYSHKEVFLREL 113S. cereale -MAPALSRTLGPSSVAALRPSP-------SRGLPLAALLPQGKRSS-SARGVR---WEAG---RGRLVGARCASAVAEKTAGEEEEAAGEKFEYQAEVSRLMDLIVHSLYSHKEVFLREL 105O. sativa -MAPSLSRSLGASSVAALRPCA-------GRVRAPGAGAARGSGSARCGRGVR---WEAGSGSRGRLVRVRCDAAVAEK---AEETAEEEKFEYQAEVSRLMDLIVHSLYSHKEVFLREL 106C. reinhardtii MMLRGLSSGMRQARAQSSASAAS-----AARPLPLLARGNGVSSALLSGASPC----AAVTAAASLRPLPAGRGPVLMRAAATEAASGSETFTYQAEVDRLMDMIVNSLYSNREVFLREL 111
ATP binding sitenucleotide binding domain
A. thaliana VSNASDALDKLRFLSVTEPSLLGDGGDLEIRIKPDPDNGTITITDTGIGMTKEELIDCLGTIAQSGTSKFLKALKENKDLGADNGLIGQFGVGFYSAFLVAEKVVVSTKSPKSDKQYVWE 227P. persica VSNASDALDKLRFLSVTEPSLLGDAGELQIRIKPDPDNGTITITDTGIGMTKEELIDCLGTIAQSGTSKFLKALKENKDLGADNGLIGQFGVGFYSAFLVAEKVVVSTKSPRSDKQYVWE 233S. cereale VSNASDALDKLRFLSVTDSSVLADGGELEIRIKPDPDAGTITITDSGIGMTKDELKDCLGTIAQSGTSKFLKALKENKELGADNGLIGQFGVGFYSAFLVAEKVVVSTKSPKTDKQYIWE 225O. sativa VSNASDALDKLRFLGVTDSSLLADGGELEIRIKPDPDAGTITITDTGIGMTKDELKDCLGTIAQSGTSKFLKALKENKDLGADNGLIGQFGVGFYSAFLVAEKVVVSTKSPKSDKQYVWE 226C. reinhardtii ISNASDALDKARFLSLTDPSVLAGREELDIRISADKEKGTLVIEDSGIGMSREQLLSNLGTIARSGTRKFMEAMAAKG----DTNLIGQFGVGFYSAFLVADRVMVQSKSPEEAKHWVWE 227
Nucleotide binding domain | Charged linke | Middle domain A. thaliana SVADSSSYLIREETDPDNILRRGTQITLYLREDDKYEFAESTRIKNLVKNYSQFVGFPIYTWQ-----------EKSRTIEVEEDEPVKEGEE-GEPK--KKKTTKTEKYWDWELANETK 333P. persica AAADSSSYVIREETDPENLIRRGTQITLYLRPDDKYEFSEPARIQGLVKNYSQFVSFPIYTWQ-----------EKSRTVEVEEEEEPKEGEE-PKPEGEKKKKTKTEKYWDWELANETK 341S. cereale AEANSSSYVIREETDPEKMLTRGTQITLFLREDDKYEFADPARIQGLVKNYSQFVSFPIFTWQ-----------EKSRTVEVE-EEESKEGEETAEGEKEEKKKTITEKYWDWELANETK 333O. sativa GVADSSSYVIKEETDPEKMLTRGTQITLVLRPDDKFEFADPGRIQGLVKNYSQFVSFPIYTWQ-----------EKSRTVEVEEDEEAKEGEEAKEGE-QKKKKTITEKYWDWELANETK 334C. reinhardtii AKAGSHQYSIREDEAKD--LVRGTRITLYLK-EDAAEMADTVKITQLIKQYSQFIAFPIKVYAPKKEPRKVVDEEATKKKQAAADAKAKEAGEEAAKPVEPVMKTEYDEVWDWRLENENK 344
Middle domainA. thaliana PLWMRNSKEVEKGEYNEFYKKAFNEFLDPLAHTHFTTEGEVEFRSILYIPGMGPLNNEDVTNPKTKNIRLYVKRVFISDDFDGELFPRYLSFVKGVVDSDDLPLNVSREILQESRIVRIM 453P. persica PIWMRNPKEVEKDEYHEFYKKTFSEFLDPVAYTHFTTEGEVEFRSVLYIPGMGPLNNEDVVNAKTKNIRLYVKRIFISDDFDGELFPRYLSFVKGVVDSNDLPLNVSREILQESRIVRIM 461S. cereale PIWMRNPKEVEETEYNEFYKKAFNEFLDPLAHAHFTTEGEVEFRSVLYIPGMAPLSNEEIMNPKTKNIRLYVKRVFISDDFDGELFPRYLSFVKGVVDSNDLPLNVSREILQESRIVRIM 453O. sativa PIWMRSPKEIEKTEYNEFYKKAFNEFLDPLAYTHFTTEGEVEFRSVLYIPGMAPLSNEEIMNPKTKNIRLYVKRVFISDDFDGELFPRYLSFVKGVVDSNDLPLNVSREILQESRIVRIM 454C. reinhardtii PIWTRSPKDVSETAYNDFFKTTFGEFLDPLAHVHFNVEGTIEFSSILYIPGMAPFEQQNMQ-QRSKSIKLYVKRVFISDEFDEDLMPRYLAFVKGVVDSSDLPLNVSREILQESRIVRVI 463
Middle domainA. thaliana RKRLIRKTFDMIQEISESENKEDYKKFWENFGRFLKLGCIEDTGNHKRITPLLRFFSSKNEEELTSLDDYIENMGENQKAIYYLATDSLKSAKSAPFLEKLIQKDIEVLYLVEPIDEVAI 573P. persica RKRLVRKTFDMIQEISESENKEDYKKLWENFGRFLKLGCIEDSGNHKRLTPLLRFYSSKSEEELISLDDYVENMPENQKAIYYLAADSLKSAKSAPFLEKLVQKDIEVLYLVEPIDEVAI 581S. cereale RKRLVRKTFDMIQDIADKDNKEDYKKFGESFGKFMKLGCIEDSGNQKRLAPLLRFYSSKNETDLISLDQYVENMPETQKAIYYIATDSLQSAKTAPFLEKLLQKDIEVLYLIEPIDEVAI 573O. sativa RKRLVRKTFDMIEEISEKDDKEDYKKFWESFGKFIKLGCIEDTGNHKRLAPLLRFHTSKNEGDLISLDQYVENMPENQKAIYYIATDSLQSAKTAPFLEKLVQKDIEVLYLIEPIDEVAI 574C. reinhardtii RKQLVRRSIEMLEELAGKEGGEDYKTFWEAFGRNIKYGVIEDTENRERLSKLLRFSSSKAEDSLTSLDEYVGRMGANQKTIYYMAADSVAAARAAPFMEAMVAKGIEVLYLTEPIDEACV 583
Middle domain | Dimerization/co-chaperoneA. thaliana QNLQTYKEKK------FVDISKE--DLELGDEDEVKDREAKQEFNLLCDWIKQQLGDKVAKVQVSNRLSSSPCVLVSGKFGWSANMERLMKAQALGDTSSLEFMRGRRILEINPDHPIIK 685P. persica QNLQTYKEKK------FVDISKE--DLELGDEDEVKERETEQEYNLLCDWIKQQLGDKVAKVQVSKRLSSSPCVLVSGKFGWSANMERLMKAQALGDTASLEFMRGRRILEVNPDHPIVK 693S. cereale QNLQTYKEKK------FVDISKE--DLELGDEDEDKE-ETKQEYTLLCDWIKQQLGDKVAKVQISKRLSSSPCVLVSGKFGWSANMERLMKAQTLGDTSSLEFMRGRRIFEINPDHPIVK 684O. sativa QNLQTYKEKK------FVDISKE--DLELGDEDEDNK-ESKQEYTLLCDWVKQQLGDKVAKVQISKRLSLSPCVLVSGKFGWSANMERLMKAQTLGDTSSLEFMRGRRIFEINPDHPIVK 685C. reinhardtii TNLGKYGPDKNGPQYELVDVSKEGVSLDEGEDEKKKAEEVAKDMAPVVDFLKKALGERVEKVTVSNRLLDSPCALVTSKFGWSANMERIMRSQALGDARAMEYMKGRKIMEINPNHDIIA 703
interaction Dimerization/co-chaperone interaction |A. thaliana DLNAACKNAPESTEATRVVDLLYDTAIISSGFTPDSPAELGNKIYEMMAMAVGGRWGRVEEEEESSTVNEGDDKSG---------ETEVVEPSEVRAES---DPWQD-- 780P. persica DLKAACKNAPDSTDAKRAVDLLYDTALISSGFTPDSPAELGNKIYEMMAMALGGRWGRSEDEEAETEVADGDAGESSDASAGEVVEAQVIEPSDVRAES---DPWSD-- 797S. cereale DLSAACKNEPDSTEAKRAVELLYETALISSGYTPESPAELGGKIYEMMTIALGGRWGRS-----GAEEAETNVDGDSS----EGVVPEVIEPSEVRTENE-NDPWRD-- 781O. sativa DLNAACKNEPESTEAKRAVELLYETALISSGYTPDSPAELGGKIYEMMTIALGGRWGRSDTETEAATTGDASTETGSS----EATVTEVIEPSEVRPES---DPWRD-- 785C. reinhardtii GIKTLLK-EKDEDRARDLSELLYETALITSGFQVDSPKDYASKVFTLMKIALG-YDILSEAEEQAAAAAPQAAEAAAAPKAAEAAAVPKVEATPVDAEVVSDDPWKKSA 810
Figure 3. Comparison of chloroplast-targeted cHSP90 from different species.
Alignment of the amino acid sequences of cHSP90 from Prunus persica, Arabidopsis thaliana (NP_178487) (Cao et al., 2003), Secale cereale (CAA82945) (Schmitz
et al., 1996), Oryza sativa (NP_001062103) (Chen et al., 2006) and Chlamydomonas reinhardtii (XP_001702984) (Willmund and Schroda, 2005). Residues with a black
background are conserved in all sequences, whereas those in grey are conserved in four of the five sequences. Amino acids with similar chemical properties are
denoted as N/Q, D/E, R/K, S/T, F/Y, A/G and V/I/L/M. Domains in cHSP90s are according to Meyer et al. (2003). The C-terminal amino acid of the chloroplast transit
peptide predicted by the CHLOROP program (Emanuelsson et al., 1999) is in red. The C-terminal DPW motif conserved in all chloroplastic HSP90 species is in green
(boxed). Amino acids in blue denote those encoded by the fragment of the peach cHSP90 mRNA predicted to be targeted by PC-sRNA8a and PC-sRNA8b.
Viroid-induced RNA silencing of host mRNA 997
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

downstream. These data conclusively showed that
PC-sRNA8a and PC-sRNA8b downregulate a peach mRNA,
as predicted by an RNA silencing mechanism. Conversely,
the lack of the same RLM-RACE products when using RNA
preparations from green tissues [from the C40-inoculated
seedling, or from a mock-inoculated seedling or a seedling
infected by the latent variant C40(A349)-g12], together with
the similar cHSP90 mRNA levels detected in the same
tissues (Figure 4), indicate that this peach transcript is not
targeted for RISC-mediated degradation in the asymptom-
atic tissues. Finally, these findings are also consistent
with the biased distribution of PLMVd variants observed
in symptomatic seedlings (Figure 1): PCsRNA8a and
PC-sRNA8b are exclusively generated from variants con-
taining the PC-associated insertion, which only accumulate
in albino leaf sectors. To give further credence to this view,
two additional peach sRNA libraries were generated and
sequenced from green leaves of PC-expressing and non-
symptomatic seedlings inoculated with variants C40 and
P1.148, respectively (Figure S6). Although PLMVd-sRNAs
deriving from the hairpin insertion were sequenced in both
libraries, none contained the PC-associated insertion. In
particular, PC-sRNA8a and PC-sRNA8b were not found, and
the PLMVd-sRNAs sequenced that could form duplexes with
cHSP90 mRNA were derived from insertions of variants
characteristically accumulating in green tissues. However,
the corresponding scores were high (from 5 to 7.5; see
Figure S7), further supporting the hypothesis that the sRNAs
resulting from PLMVd variants without the PC-associated
insertion do not prime RISC for cHSP90 mRNA cleavage.
DISCUSSION
In the first part of this study we furthered the characterization
of the PLMVd determinant for PC by dissecting albino and
green sectors of GF-305 peach seedlings inoculated with
natural and artificial variants of this viroid, and subsequently
cloning the resulting progeny. This approach has revealed
the 12-nt sequence GA(A/G)CUUUUGUUC as the insertion
conferring PC to infecting PLMVd variants. In agreement
with previous data (Malfitano et al., 2003; Rodio et al., 2006),
this insertion folds into a hairpin capped by a UUUU loop,
and differs in the stem composition from other hairpin
insertions found in PLMVd variants recovered from green
tissues. Therefore, the primary rather than secondary
structure of the pathogenic determinant is involved in elic-
iting PC, as clearly illustrated by the artificial variant
C40(stem). The infecting ability of this variant probably
results from the eight mutations introduced not affecting the
stem stability of the hairpin insertion (Figure 2), in line with
cDNA dilution 1:2 1:20 1:200
cHSP90
Healthy
C40(349)-g12(green)
C40 (albino)
C40 (green)
18S psbA rpoB
1:2 1:20 1:200 1:2 1:20 1:200 1:2 1:20 1:200
Figure 4. Transcript accumulation in GF-305 leaves.
RT-PCR semiquantitative estimation of two nucleus-encoded transcripts (the
18S rRNA and the mRNA of the chloroplast-targeted cHSP90) and two plastid-
encoded transcripts (the mRNAs of the rpoB subunit and of the psbA
component of photosystem II). The cDNAs, generated by random-primed
reverse transcription of equalized RNA preparations, were serially diluted
before PCR amplification with gene-specific primer pairs. The amplified
products were analyzed by agarose gel electrophoresis and ethidium bromide
staining. Only gel sections containing the amplified cDNAs are presented.
Results are representative of three replicates conducted on two biologically
independent samples.
1/5 4/5
5’-..AGCCUGAGAAGUGCUUUUGUUCCUCAAAAUGGGCUCAGGAAGG..-3’||| |||||||||||||||||
3’-UUCUCGAAAACAAGGAGUUUU-5’PC-sRNA8a
cHSP90 mRNA
336 7
AAAA
AA
G
G U UU
U
UU C G A
UCC
C. . . .
7
336
scaffold_1: 28257983..28265359
28258k 28259k 28260k 28261k 28262k 28263k 28264k
Transcriptppa001590m
Figure 5. Specific cleavage of peach cHSP90 mRNA
mediated by PC-sRNAs.
Upper panel, predicted cHSP90 transcript and loca-
tion of the gene into the peach genome v1.0 (http://
www.phytozome.org). Middle panel, cHSP90
mRNA:PC-sRNA8a duplex (in bold), with the
sequence of PC-sRNA8a corresponding to the
PC-associated insertion underlined (note that they
are of complementary polarity). Arrows mark the
predicted and validated cleavage sites by RLM-
RACE from five independent clones, with fractions
indicating the number of clones producing the same
results. Lower panel, schematic representation of
the secondary structure predicted for PLMVd variant
C40 with nucleotides complementary to PC-sRNA8a
highlighted, and those forming part of the PC-induc-
ing insertion on a black background.
998 Beatriz Navarro et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

previous observations with another member of the same
genus, Chrysanthemum chlorotic mottle viroid (CChMVd;
Navarro and Flores, 1997), which also remains viable as long
as the introduced mutations preserve elements of its sec-
ondary and tertiary structure (De la Pena et al., 1999; Gago
et al., 2005).
Altogether, these findings are consistent with the silenc-
ing-based model of pathogenesis proposed for viroid and/or
satellite RNAs (Papaefthimiou et al., 2001; Wang et al.,
2004), recently validated experimentally for the yellow
symptoms induced by a specific satellite RNA (Y-sat) of
CMV (Shimura et al., 2011; Smith et al., 2011). However,
regarding viroids, previous attempts to obtain support for
this model have met with limited success. Although some
reports highlight the positive correlation between symptom
severity and the accumulation of vd-sRNAs in different
plant–viroid combinations (Itaya et al., 2001; Markarian
et al., 2004; Matousek et al., 2007), including transgenic
Solanum lycopersicum (tomato) lines expressing a non-
infectious PSTVd hairpin RNA (Wang et al., 2004), others
have failed to find a similar correlation in some transgenic
lines expressing the same non-infectious PSTVd hairpin
RNA, and apparently accumulating similar vd-sRNA titers
(Schwind et al., 2009). Moreover, although RDR6, an enzyme
mediating synthesis of secondary siRNAs, has been
involved in symptom induction by Hop stunt viroid in
Nicotiana benthamiana (Gomez et al., 2008), symptoms
elicited by PSTVd in the same plant species are RDR6-
independent (Di Serio et al., 2010). Setting aside these
contradictory results, none of the previous studies have
provided direct experimental support for the vd-sRNAs
actually mediating cleavage of host mRNAs; at best, a
correlation between the downregulation of tomato mRNAs
potentially targeted by PSTVd sRNAs and symptom expres-
sion has been found (Wang et al., 2011). Finally, even if
vd-sRNAs may target and downregulate, in a sequence-
specific manner, the overexpression of a reporter gene (Vogt
et al., 2004; Itaya et al., 2007) or the accumulation of viroid
RNA in infected plants (Carbonell et al., 2008), these results
do not prove that a similar mechanism operates against host
genes in natural infections.
In the second part of this study we supply solid experi-
mental evidence showing that the accumulation level of the
mRNA coding for the chloroplast-targeted cHSP90 from
peach is reduced in the albino sectors of PC-expressing
leaves. Our data, obtained by a combination of deep
sequencing, semi-quantitative RT-PCR and RLM-RACE, are
consistent with the sequence-specific cleavage of this mRNA
guided by PLMVd-sRNAs (PC-sRNA8a and PC-sRNA8b)
generated from the PC-associated insertion of some PLMVd
variants, thus illustrating that at least this viroid can indeed
modify its host gene expression through RNA silencing.
Chloroplast developmental defects and malfunctioning of
plastid-to-nucleus signaling reported in PC-expressing
tissues (Rodio et al., 2007) support a role for the down-
regulation of cHSP90 in eliciting the albino phenotype,
because this chloroplast-targeted protein has been involved
in chloroplast biogenesis and plastid-to-nucleus signal
transduction in Arabidopsis and Chlamydomonas (Cao
et al., 2003; Willmund and Schroda, 2005; Willmund et al.,
2008). Additional support for this view comes from the
yellow phenotype observed in a mutant line of A. thaliana
with a single amino acid change in cHSP90 (Cao et al., 2003),
and from two cHSP90 T-DNA null-mutant lines (EMB 1956-1
and EMB 1956-2) of A. thaliana that are embryo defective,
and display white embryos and seeds (Meinke et al., 2008).
Although it has recently been shown that the yellow
symptoms induced in Nicotiana tabacum by the Y-sat of
CMV result from silencing the chlorophyll biosynthetic gene
CHLI with a 22-nt siRNA derived from this satellite RNA
(Shimura et al., 2011; Smith et al., 2011), another more
complex RNA silencing-based model has been offered for
explaining the attenuation of the yellowing symptomatol-
ogy of N. benthamiana induced by a different satellite RNA
of the same virus (Hou et al., 2011), indicating that the
pathogenic mechanism proposed for Y-sat may not be
general. Whether the silencing of gene cHSP90 guided by
PC-sRNA8a and PC-sRNA8b is sufficient for inciting PC is not
known because, at this stage, we cannot rule out the
involvement of other factors. However, PLMVd-sRNAs other
than PC-sRNAs can be excluded as direct players in this
respect, because they lack nucleotides derived from the
insertion strictly associated with PC.
Nine and three PC-sRNAs potentially targeting peach
mRNAs have (+) and ()) polarity, respectively (Table 1), but
both PC-sRNA8a and PC-sRNA8b derive from the ()) polarity
strand. These results indicate that the activity of vd-sRNAs in
promoting host mRNA cleavage may not depend on their
relative abundance. In fact, the PLMVd region encompassing
the PC determinant does not map at a hot spot in the profile
of vd-sRNA reads (Figure S4); neither does the Y-sat region
from which the 22-nt (+) species silencing CHLI mRNA
derives (Shimura et al., 2011; Smith et al., 2011). This finding
is in agreement with results of a previous study in which
viroid dsRNAs, probably produced in the cytoplasm by host
RDR(s), were proposed to serve as substrates for generating
PLMVd-sRNAs (Di Serio et al., 2009). Consistent with this
dsRNA-based origin of most PLMVd-sRNAs, the passenger
strand of PC-sRNA8b was also identified in the sRNA library
(Appendix S1, sequence ID: MAC-18-1596391_21_3_0), but
with a number of reads (only three) significantly lower than
that of its guide counterpart (169 reads; Table 1) mediating
cleavage of the cHSP90 mRNA. Two other points are worthy
of note: (i) PC-sRNA1 and PC-sRNA6 may form duplexes
with more than one peach mRNA, suggesting that a small
viroid region has the potential for silencing multiple host
targets; and (ii) the U prevalence in the four 5¢ terminal
positions of PC-sRNA8a and PC-sRNA8b is remarkable
Viroid-induced RNA silencing of host mRNA 999
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

because this trait is also found in the 22-nt siRNA from Y-sat
of CMV silencing CHLI mRNA.
As PLMVd accumulates genetic heterogeneity very rap-
idly (Ambros et al., 1999; Rodio et al., 2006; this work), in line
with the observation that the related CChMVd has the
highest mutation rate reported for any biological entity
(Gago et al., 2009), new PLMVd-sRNAs may be generated
with time, targeting other peach genes and eventually
resulting in the fluctuating phenotype characteristic of most
PLMVd infections (Flores et al., 2006). Consistent with this
view, most PLMVd-sRNAs with potential peach targets
derive from viroid regions displaying high sequence vari-
ability (Figure S5). As previously proposed for viruses
(Moissiard and Voinnet, 2006; Llave, 2010), changes in host
gene expression induced by PLMVd-sRNAs could generate a
more favorable environment for viroid infection. Recipro-
cally, viroid modulation of host gene expression via RNA
silencing might play a role in the molecular evolution of
these minimal infectious agents.
EXPERIMENTAL PROCEDURES
Plant material and growing conditions
Leaf tissues were from GF-305 peach [Prunus persica (L.) Batsch]seedlings slash-inoculated with buffer or dimeric head-to-tail tran-scripts generated in vitro from natural and artificial PLMVd variantsinducing different foliar symptoms or no symptoms (see below). Sixweeks after inoculation, symptom expression was stimulated bychilling the seedlings at 4�C in the darkness for 6–8 weeks, and thentransferring them back to the glasshouse to favor the emergence ofnew flushes.
RNA extraction and dot-blot hybridization
Total nucleic acid preparations were extracted from leaves (60 mg)with phenol-chloroform, recovered by ethanol precipitation andresuspended in water (50 ll) (Rodio et al., 2007). When indicated,albino sectors from PC-expressing leaves were excised from thesurrounding green tissues with a razor blade. PLMVd accumulationin infected tissues was quantified by spotting aliquots (5 ll) of 1/5, 1/25 and 1/125 dilutions of the nucleic acid preparations ontopositively charged nylon membranes (Roche Diagnostics GmbH,http://www.roche.com) that were hybridized with a PLMVd-specificdigoxigenin-labeled riboprobe (Rodio et al., 2006).
Amplification and sequencing of PLMVd progeny variants
For cloning PLMVd-cDNAs from progeny variants, nucleic acidspreparations (150 ll) were obtained from leaf tissues (60 mg) by amodified silica-gel capture system (Foissac et al., 2001); whendealing with symptomatic leaves, albino and green sectors weredissected beforehand. Aliquots (5 ll) were used for synthesizingfirst-strand cDNA with random hexamers and the High CapacityReverse Transcription kit (Applied Biosystem, http://www.appliedbiosystems.com). The resulting cDNAs were PCR-amplified with theprimer pair FPLMV-57 (5¢-CACACCCCCCTCGGAACCAACCG-3¢) andFPLMV-58 (5¢-ATCCAGGTACCGCCGTAGAAAC-3¢), complementaryand identical to positions 202–180 and 203–224 of the referencePLMVd variant (Hernandez and Flores, 1992; Ambros et al., 1998),respectively, and the Expand High Fidelity PCR system (RocheDiagnostics GmbH). Amplicons of the expected size were cloned
in p-GEM-T Easy plasmid (Promega, http://www.promega.com)and sequenced (MWG-Biotech, http://www.mwg-biotech.com).The GenBank IDs for PLMVd progeny variants are: JN377825–JN377849; JN377851–JN377862; JN377864–JN377874; andJN377876–JN377891.
PLMVd infectious clones, site-directed mutagenesis and
inoculation of in vitro transcripts
Plasmids containing head-to-tail PLMVd-cDNA dimeric inserts fromnatural variants C40 (AJ550912) and P1.148 (DQ222050) have beendescribed previously (Malfitano et al., 2003; Rodio et al., 2006); fordescriptions of plasmids of variants C40(A349)-g12 (JN377855),P1.148(C349) (JN377863), C40(A349) (JN377850) and C40(stem)(JN377875), see Appendix S2. Recombinant plasmids were linearizedwith appropriate restriction enzymes and transcribed with T7 or SP6RNA polymerases. The resulting products were analyzed by elec-trophoresis in 5% polyacrylamide gels containing 1X TBE buffer and8 M urea, eluted and slash-inoculated into GF-305 peach seedlings.
Deep sequencing and bioinformatics analyses
The protocol for purifying the sRNAs, adaptor ligation, RT-PCRamplification, library purification and high-throughput DNAsequencing on the Illumina Genome Analyzer (Fasteris SA, http://www.fasteris.com), has been reported (Di Serio et al., 2010). Fourlibraries were sequenced. Two bar-coded leaf samples, from mock-inoculated and C40-infected GF-305 seedlings, were analyzed in asingle channel in the Illumina EAS269 GAII; the sRNA library fromthe C40-infected GF-305 seedling was generated from albino sectorsdissected from adjacent green tissues. The two additional bar-co-ded libraries from green leaf tissues of C40- and P1.148-infectedGF305 seedlings were sequenced in a single channel in the IlluminaGenome Analyzer HiSeq 2000. Raw data from the Illumina platformwere fed into an in-house pipeline, which removed barcodes andadaptors and split the clean sequences by size. Sequence sizesbetween 18 and 26 nt were blasted (BLASTN; Altschul et al., 1997)against a selected set of PLMVd variants, and a mapping profileagainst the consensus of the multiple alignment (CLUSTALW;Thompson et al., 1994) was generated. To find out potential targetsof PLMVd-sRNAs, sequences perfectly matching genomic RNAs ofC40 and its progeny were used for RNAhybrid (Rehmsmeier et al.,2004) searching on exon sequence fragments of the peach genome(Peach v1.0; International Peach Genome Initiative, http://www.rosaceae.org/peach/genome). The quality of the duplex pair-ing was estimated as proposed previously (Fahlgren and Carring-ton, 2010). Peach mRNAs detected as transcripts and formingpotential duplexes with PLMVd-sRNAs spanning totally or partially(in at least one nucleotide) the PC-associated insertion (PC-sRNAs)were further analysed by CHLOROP (http://www.cbs.dtu.dk/services/ChloroP/; Emanuelsson et al., 1999) to predict the presence ofchloroplastic transit peptides (Li and Chiu, 2010) in the encodedproteins.
Transcript analysis
Transcript levels were estimated by RT-PCR. Total RNAs (200 ng),obtained by treating total nucleic acid preparations with RQ1DNase I (Promega), were reverse transcribed with random hexa-mers. Aliquots (2 ll) of serial dilutions (1:2, 1:20 and 1:200) of theresulting cDNAs were added to amplification reactions (25 ll)catalyzed with Go-Taq DNA polymerase (Promega). The cyclingprogram, consisting in an initial denaturation at 94�C for 3 min and30 cycles (94�C for 30 s, 50�C for 30 s for 18S, psbA and RpoB, and55�C for cHSP90, and 72�C for 30 s), with a final extension at 72�C for7 min, was adopted according to preliminary experiments showing
1000 Beatriz Navarro et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

that cDNA amplification was in the logarithmic phase. A cDNAfragment (405 nt) of cHSP90 mRNA was RT-PCR amplified withprimers cHSP90-Fw (5¢-CAATGGCTCCAGTTCTAAGCA-3¢) andcHSP90-Rv (5¢-GCAGAGAGGGCTCAGTCACACTCAA-3¢), identicaland complementary, respectively, to positions 84–104 and 464–488of this transcript. Sequencing of the resulting product confirmed theexpected cDNA (JN377892). Primer pairs for cDNA amplification ofthe 18S rRNA and psbA and RpoB transcripts have been describedpreviously (Rodio et al., 2007).
5¢ RNA ligase-mediated rapid amplification of cDNA ends
(RLM-RACE)
RQ1 DNase I-treated total RNAs (1.2 lg) were incubated at 20�C for6 h with an RNA adaptor (5¢-GUUCAGAGUUCUACAGUCCGACG-AUC-3¢) and 0.5 U of T4 RNA ligase (Promega). Ligated RNAs werereverse transcribed with primer cHSP90-Rv as described before.First PCR was then performed using the forward primer P2 (5¢-AATGATACGGCGACCACCGACAGGTTCAGAGTTCTACAGTCCGA-3¢),with the 3¢ moiety (in bold) homologous to the RNA adaptor, and thereverse primer cHSP90-Rv reported above, whose 5¢ end maps288 nt downstream the cleavage site of peach cHSP90 mRNApredicted by PC-sRNA8a and PC-sRNA8b. The resulting product wasamplified with the same primer P2 and the nested reverse primercHSP90-nes-Rv (5¢-CCTTGTGGCTGTATAGACTATG-3¢), comple-mentary to positions 383–404 of peach cHSP90 mRNA. Followingelectrophoresis in a 1.2% agarose gel, the nested PCR product(248 bp) was excised, cloned into the pGEM-T easy vector (Pro-mega) and sequenced.
ACKNOWLEDGEMENTS
This work was supported by a dedicated grant from the ItalianMinistry of Economy and Finance to the CNR (Legge n. 191/2009),the Dipartimento Agroalimentare of the CNR of Italy (A. Leone andD. Mariotti 2008 award for advanced research in agriculture to FDS)and by the Ministerio de Ciencia e Innovacion of Spain (grantsBFU2008-03154 and BFU2011-28443 to RF).
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. Peach phenotype and progeny of the natural PLMVdvariant C40(A349)-g12.Figure S2. Relative distribution by size classes of total sRNAs fromGF-305 peach leaves infected by variant C40 (blue) or mockinoculated (red).Figure S3. Distribution by size classes, polarity and abundance ofPLMVd-sRNAs in the C40-infected sample.Figure S4. Location and frequency of the 5¢ termini of PLMVd-sRNAsalong the genomic RNAs.Figure S5. Number and location of the 5¢ termini of 21-nt non-redundant (nr) PLMVd-sRNAs along the genomic RNAs (a), andnucleotide variability detected in variant C40 and its progeny (b).Figure S6. Distribution by size classes, polarity and abundance ofPLMVd-sRNAs in the green tissue of C40- and P1.148-infectedsamples.Figure S7. Duplexes potentially formed by the cHSP90 mRNA andPLMVd-sRNAs with hairpin insertions obtained by deep sequencingof sRNA libraries from green leaves of C40- and P1.148-infectedpeach seedlings (a and b, respectively).Table S1. Predicted peach mRNAs targeted by PLMVd-sRNAsmatching perfectly the genomic sequence of variant C40 and itsprogeny.
Table S2. Chloroplast transit peptides and their length in cHSP90 offive species predicted by the CHLOROP program.Appendix S1. PLMVd-sRNAs (18–26 nt) from albino leaf sectorsinfected with PLMVd variant C40.Appendix S2. Experimental procedures.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W.
and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation
of protein database search programs. Nucleic Acids Res. 25, 3389–3402.
Ambros, S., Hernandez, C., Desvignes, J.C. and Flores, R. (1998) Genomic
structure of three phenotypically different isolates of peach latent mosaic
viroid: implications of the existence of constraints limiting the heteroge-
neity of viroid quasi-species. J. Virol. 72, 7397–7406.
Ambros, S., Hernandez, C. and Flores, R. (1999) Rapid generation of genetic
heterogeneity in progenies from individual cDNA clones of peach latent
mosaic viroid in its natural host. J. Gen. Virol. 80, 2239–2252.
Bolduc, F., Hoareau, C., St-Pierre, P. and Perreault, J.P. (2010) In-depth
sequencing of the siRNAs associated with peach latent mosaic viroid
infection. BMC Mol. Biol. 11, 16.
Bussiere, F., Lehoux, J., Thompson, D.A., Skrzeczkowski, L.J. and Perreault,
J. (1999) Subcellular localization and rolling circle replication of
peach latent mosaic viroid: hallmarks of group A viroids. J. Virol. 73,
6353–6360.
Cao, D., Froehlich, J.E., Zhang, H. and Cheng, C.L. (2003) The chlorate-resis-
tant and photomorphogenesis-defective mutant cr88 encodes a chloro-
plast-targeted HSP90. Plant J. 33, 107–118.
Carbonell, A., Martınez de Alba, A.E., Flores, R. and Gago, S. (2008)
Double-stranded RNA interferes in a sequence-specific manner with
infection of representative members of the two viroid families. Virology,
371, 44–53.
Carthew, R.W. and Sontheimer, E.J. (2009) Origins and Mechanisms of
miRNAs and siRNAs. Cell, 136, 642–655.
Chen, X. (2009) Small RNAs and their roles in plant development. Annu. Rev.
Cell Dev. Biol. 25, 21–44.
Chen, B., Zhong, D. and Monteiro, A. (2006) Comparative genomics and
evolution of the HSP90 family of genes across all kingdoms of organisms.
BMC Genomics, 7, 156.
Csorba, T., Pantaleo, V. and Burgyan, J. (2009) RNA silencing: an antiviral
mechanism. Adv. Virus Res. 75, 35–71.
De la Pena, M., Navarro, B. and Flores, R. (1999) Mapping the molecular
determinant of pathogenicity in a hammerhead viroid: a tetraloop within
the in vivo branched RNA conformation. Proc. Natl Acad. Sci. USA, 96,
9960–9965.
Di Serio, F., Gisel, A., Navarro, B., Delgado, S., Martınez de Alba, A.E.,
Donvito, G. and Flores, R. (2009) Deep sequencing of the small RNAs de-
rived from two symptomatic variants of a chloroplastic viroid: implications
for their genesis and for pathogenesis. PLoS ONE, 4, e7539.
Di Serio, F., Martınez de Alba, A.E., Navarro, B., Gisel, A. and Flores, R.
(2010) RNA-dependent RNA polymerase 6 delays accumulation and
precludes meristem invasion of a nuclear-replicating viroid. J. Virol. 84,
2477–2489.
Diener, T.O. (1972) Potato spindle tuber viroid. VIII. Correlation of infectivity
with a UV-absorbing component and thermal denaturation properties of
the RNA. Virology, 50, 606–609.
Diermann, N., Matousek, J., Junge, M., Riesner, D. and Steger, G. (2010)
Characterization of plant miRNAs and small RNAs derived from potato
spindle tuber viroid (PSTVd) in infected tomato. Biol. Chem. 391, 1379–
1390.
Ding, B. (2009) The biology of viroid-host interactions. Annu. Rev. Phytopa-
thol. 47, 105–131.
Ding, S.W. (2010) RNA-based antiviral immunity. Nat. Rev. Immunol. 10, 632–
644.
Viroid-induced RNA silencing of host mRNA 1001
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

Dunoyer, P. and Voinnet, O. (2009) Movement of RNA silencing between
plant cells: is the question now behind us? Trends Plant Sci. 14, 643–
644.
Emanuelsson, O., Nielsen, H. and von Heijne, G. (1999) ChloroP, a neural
network-based method for predicting chloroplast transit peptides and their
cleavage sites. Protein Sci. 8, 978–984.
Fahlgren, N. and Carrington, J.C. (2010) miRNA target prediction in plants.
Methods Mol. Biol. 592, 51–57.
Flores, R., Daros., J.A. and Hernandez, C. (2000) The Avsunviroidae family:
viroids with hammerhead ribozymes. Adv. Virus Res. 55, 271–323.
Flores, R., Hernandez, C., Martınez de Alba, A.E., Daros., J.A. and Di Serio, F.
(2005) Viroids and viroid-host interactions. Annu. Rev. Phytopathol. 43,
117–139.
Flores, R., Delgado, S., Rodio, M.E., Ambros, S., Hernandez, C. and Di Serio, F.
(2006) Peach latent mosaic viroid: not so latent. Mol. Plant Pathol. 7, 209–
221.
Foissac, X., Svanella-Dumas, L., Dulucq, M.J., Gentit, P. and Candresse, T.
(2001) Polyvalent detection of fruit tree tricho, capillo and foveaviruses by
nested RT-PCR using degenerated and inosine containing primers (PDO
RT-PCR). Acta Hortic. 550, 37–43.
Gago, S., De la Pena, M. and Flores, R. (2005) A kissing-loop interaction in a
hammerhead viroid RNA critical for its in vitro folding and in vivo viability.
RNA, 11, 1073–1083.
Gago, S., Elena, S.F., Flores, R. and Sanjuan, R. (2009) Extremely high muta-
tion rate of a hammerhead viroid. Science, 323, 1308.
Gomez, G., Martınez, G. and Pallas, V. (2008) Viroid-induced symptoms in
Nicotiana benthamiana plants are dependent on RDR6 activity. Plant
Physiol. 148, 414–423.
Gross, H.J., Domdey, H., Lossow, C., Jank, P., Raba, M., Alberty, H. and
Sanger, H.L. (1978) Nucleotide sequence and secondary structure of potato
spindle tuber viroid. Nature, 273, 203–208.
Hernandez, C. and Flores, R. (1992) Plus and minus RNAs of peach latent
mosaic viroid self-cleave in vitro through hammerhead structures. Proc.
Natl Acad. Sci. USA, 89, 3711–3715.
Hou, W.N., Duan, C.G., Fang, R.X., Zhou, X.Y. and Guo, H.S. (2011) Satellite
RNA reduces expression of the 2b suppressor protein resulting in the
attenuation of symptoms caused by cucumber mosaic virus infection. Mol.
Plant Pathol. 12, 595–605.
Hutchins, C., Rathjen, P.D., Forster, A.C. and Symons, R.H. (1986) Self-cleav-
age of plus and minus RNA transcripts of avocado sunblotch viroid. Nucleic
Acids Res. 14, 3627–3640.
Itaya, A., Folimonov, A., Matsuda, Y., Nelson, R.S. and Ding, B. (2001) Potato
spindle tuber viroid as inducer of RNA silencing in infected tomato. Mol.
Plant-Microbe Interact. 14, 1332–1334.
Itaya, A., Matsuda, Y., Gonzales, R.A., Nelson, R.S. and Ding, B. (2002) Potato
spindle tuber viroid strains of different pathogenicity induces and sup-
presses expression of common and unique genes in infected tomato. Mol.
Plant-Microbe Interact. 15, 990–999.
Itaya, A., Zhong, X., Bundschuh, R., Qi, Y., Wang, Y., Takeda, R., Harris, A.R.,
Molina, C., Nelson, R.S. and Ding, B. (2007) A structured viroid RNA is
substrate for Dicer-like cleavage to produce biologically active small RNAs
but is resistant to RISC-mediated degradation. J. Virol. 81, 2980–2994.
Jay, F., Wang, Y., Yu, A., Taconnat, L., Pelletier, S., Colot, V., Renou, J.P. and
Voinnet, O. (2011) Misregulation of AUXIN RESPONSE FACTOR 8 underlies
the developmental abnormalities caused by three distinct viral silencing
suppressors in Arabidopsis. PLoS Pathog. 7, e1002035.
Kasschau, K.D., Xie, Z., Allen, E., Llave, C., Chapman, E.J., Krizan, K.A. and
Carrington, J.C. (2003) P1/HC-Pro, a viral suppressor of RNA silencing,
interferes with Arabidopsis development and miRNA unction. Dev. Cell, 4,
205–217.
Li, H.M. and Chiu, C.C. (2010) Protein transport into chloroplasts. Annu. Rev.
Plant Biol. 61, 157–180.
Llave, C. (2010) Virus-derived small interfering RNAs at the core of plant-virus
interactions. Trends Plant Sci. 15, 701–707.
Malfitano, M., Di Serio, F., Covelli, L., Ragozzino, A., Hernandez, C. and Flores,
R. (2003) Peach latent mosaic viroid variants inducing peach calico contain
a characteristic insertion that is responsible for this symptomatology.
Virology, 313, 492–501.
Mallory, A. and Vaucheret, H. (2010) Form, function, and regulation of ARG-
ONAUTE proteins. Plant Cell, 22, 3879–3889.
Mallory, A.C., Reinhart, B.J., Jones-Rhoades, M.W., Tang, G., Zamore, P.D.,
Barton, M.K. and Bartel, D.P. (2004) MicroRNA control of PHABULOSA in
leaf development: importance of pairing to the microRNA 5¢ region. EMBO
J. 23, 3356–3364.
Markarian, N., Li, H.W., Ding, S.W. and Semancik, J.S. (2004) RNA
silencing as related to viroid induced symptom expression. Arch. Virol.
149, 397–406.
Martınez de Alba, A.E., Flores, R. and Hernandez, C. (2002) Two chloroplastic
viroids induce the accumulation of the small RNAs associated with post-
transcriptional gene silencing. J. Virol. 76, 13094–13096.
Martınez, G., Donaire, L., Llave, C., Pallas, V. and Gomez, G. (2010) High-
throughput sequencing of Hop stunt viroid-derived small RNAs from
cucumber leaves and phloem. Mol. Plant Pathol. 11, 347–359.
Matousek, J., Kozlova, P., Orctova, L., Schmitz, A., Pesina, K., Bannach, O.,
Diermann, N., Steger, G. and Riesner, D. (2007) Accumulation of viroid-
specific small RNAs and increase of nucleolytic activities linked to viroid-
caused pathogenesis. Biol. Chem. 388, 1–13.
Meinke, D., Muralla, R., Sweeney, C. and Dickerman, A. (2008) Identifying
essential genes in Arabidopsis thaliana. Trends Plant Sci. 13, 483–491.
Meyer, P., Prodromou, C., Hu, B., Vaughan, C., Roe, S.M., Panaretou, B., Piper,
P.W. and Pearl, L.H. (2003) Structural and functional analysis of the middle
segment of hsp90: implications for ATP hydrolysis and client protein and
cochaperone interactions. Mol. Cell, 11, 647–658.
Moissiard, G. and Voinnet, O. (2006) RNA silencing of host transcripts by cau-
liflower mosaic virus requires coordinated action of the four Arabidopsis
Dicer-like proteins. Proc. Natl Acad. Sci. USA, 103, 19593–19598.
Navarro, B. and Flores, R. (1997) Chrysanthemum chlorotic mottle
viroid: unusual structural properties of a subgroup of self-cleaving viroids
with hammerhead ribozymes. Proc. Natl Acad. Sci. USA, 94, 11262–11267.
Navarro, B., Pantaleo, V., Gisel, A., Moxon, S., Dalmay, T., Bisztray, G.,
Di Serio, F. and Burgyan, J. (2009) Deep sequencing of viroid-derived small
RNAs from grapevine provides new insights on the role of RNA silencing in
plant-viroid interaction. PLoS ONE, 4, e7686.
Papaefthimiou, I., Hamilton, A.J., Denti, M.A., Baulcombe, D.C., Tsagris, M.
and Tabler, M. (2001) Replicating potato spindle tuber viroid RNA is
accompanied by short RNA fragments that are characteristic of post-tran-
scriptional gene silencing. Nucleic Acids Res. 29, 2395–2400.
Rehmsmeier, M., Steffen, P., Hochsmann, M. and Giegerich, R. (2004) Fast
and effective prediction of microRNA/target duplexes. RNA, 10, 1507–1517.
Rodio, M.E., Delgado, S., Flores, R. and Di Serio, F. (2006) Variants of peach
latent mosaic viroid inducing peach calico: uneven distribution in infected
plants and requirements of the insertion containing the pathogenicity
determinant. J. Gen. Virol. 87, 231–240.
Rodio, M.E., Delgado, S., De Stradis, A.E., Gomez, M.D., Flores, R. and
Di Serio, F. (2007) A viroid RNA with a specific structural motif inhibits
chloroplast development. Plant Cell, 19, 3610–3626.
Schmitz, G., Schmidt, M. and Feierabend, J. (1996) Characterization of a
plastid-specific HSP90 homologue: identification of a cDNA sequence,
phylogenetic descendence and analysis of its mRNA and protein expres-
sion. Plant Mol. Biol. 30, 479–492.
Schroda, M. and Muhlhaus, T. (2009) A ‘foldosome’ in the chloroplast? Plant
Signal Behav. 4, 301–303.
Schwind, N., Zwiebel, M., Itaya, A., Ding, B., Wang, M.B., Krczal, G. and
Wassenegger, M. (2009) RNAi-mediated resistance to potato spindle tuber
viroid in transgenic tomato expressing a viroid hairpin RNA construct. Mol.
Plant Pathol. 10, 459–469.
Shimura, H., Pantaleo, V., Ishihara, T., Myojo, N., Inaba, J., Sueda, K., Burg-
yan, J. and Masuta, C. (2011) A viral satellite RNA induces yellow symp-
toms on tobacco by targeting a gene involved in chlorophyll biosynthesis
using the RNA silencing machinery. PLoS Pathog. 7, e1002021.
Smith, N.A., Eamens, A.L. and Wang, M.B. (2011) Viral small interfering RNAs
target host genes to mediate disease symptoms in plants. PLoS Pathog. 7,
e1002022.
St-Pierre, P., Hassen, F., Thompson, D. and Perreault, J.P. (2009) Character-
ization of the siRNAs associated with peach latent mosaic viroid infection.
Virology, 383, 178–182.
Symons, R.H. (1981) Avocado sunblotch viroid: primary sequence and pro-
posed secondary structure. Nucleic Acids Res. 9, 6527–6537.
Tessitori, M., Maria, G., Capasso, C., Catara, G., Rizza, S., De Luca, V., Catara,
A., Capasso, A. and Carginale, V. (2007) Differential display analysis of gene
1002 Beatriz Navarro et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003

expression in Etrog citron leaves infected by citrus viroid III. Biochim.
Biophys. Acta, 1769, 228–235.
Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUSTAL W: improv-
ing the sensitivity of progressive multiple sequence alignment through
sequence weighting, position-specific gap penalties and weight matrix
choice. Nucleic Acids Res. 22, 4673–4680.
Tsagris, E.M., Martınez de Alba, A.E., Gozmanova, M. and Kalantidis, K. (2008)
Viroids. Cell. Microbiol. 10, 2168–2179.
Vogt, U., Pelissier, T., Putz, A., Razvi, F., Fischer, R. and Wassenegger, M.
(2004) Viroid-induced RNA silencing of GFP-viroid fusion transgenes does
not induce extensive spreading of methylation or transitive silencing. Plant
J. 1, 107–118.
Voinnet, O. (2008) Use, tolerance and avoidance of amplified RNA silencing
by plants. Trends Plant Sci. 13, 317–328.
Wang, M.B., Bian, X.Y., Wu, L.M. et al. (2004) On the role of RNA silencing in
the pathogenicity and evolution of viroids and viral satellites. Proc. Natl
Acad. Sci. USA, 101, 3275–3280.
Wang, Y., Shibuya, M., Taneda, A., Kurauchi, T., Senda, M., Owens, R.A. and
Sano, T. (2011) Accumulation of potato spindle tuber viroid-specific small
RNAs is accompanied by specific changes in gene expression in two
tomato cultivars. Virology, 413, 72–83.
Willmund, F. and Schroda, M. (2005) HEAT SHOCK PROTEIN 90C is a bona
fide Hsp90 that interacts with plastidic HSP70B in Chlamydomonas rein-
hardtii. Plant Physiol. 138, 2310–2322.
Willmund, F., Dorn, K.V., Schulz-Raffelt, M. and Schroda, M. (2008) The
chloroplast DnaJ homolog CDJ1 of Chlamydomonas reinhardtii is part of a
multichaperone complex containing HSP70B, CGE1, and HSP90C. Plant
Physiol. 148, 2070–2082.
Viroid-induced RNA silencing of host mRNA 1003
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 70, 991–1003
Related Documents











