Sites of Interaction of a Precursor Polypeptide on the Export Chaperone SecB Mapped by Site-directed Spin Labeling Jennine M. Crane 1 , Yuying Suo 1 , Angela A. Lilly 1 , Chunfeng Mao 1 Wayne L. Hubbell 2 and Linda L. Randall 1 ⁎ 1 Department of Biochemistry, University of Missouri, Columbia, MO 65211, USA 2 Jules Stein Eye Institute and Department of Chemistry and Biochemistry, University of California, Los Angeles, CA 90095-7008, USA Export of protein into the periplasm of Escherichia coli via the general secretory system requires that the transported polypeptides be devoid of stably folded tertiary structure. Capture of the precursor polypeptides before they fold is achieved by the promiscuous binding to the chaperone SecB. SecB delivers its ligand to export sites through its specific binding to SecA, a peripheral component of the membrane translocon. At the translocon the ligand is passed from SecB to SecA and subsequently through the SecYEG channel. We have previously used site-directed spin labeling and electron paramagnetic resonance spectroscopy to establish a docking model between SecB and SecA. Here we report use of the same strategy to map the pathway of a physiologic ligand, the unfolded form of precursor galactose-binding protein, on SecB. Our set of SecB variants each containing a single cysteine, which was used in the previous study, has been expanded to 48 residues, which cover 49% of the surface of SecB. The residues on SecB involved in contacts were identified as those that, upon addition of the unfolded polypeptide ligand, showed changes in spectral line shape consistent with restricted motion of the nitroxide. We conclude that the bound precursor makes contact with a large portion of the surface of the small chaperone. The sites on SecB that interact with the ligand are compared with the previously identified sites that interact with SecA and a model for transfer of the ligand is discussed. © 2006 Elsevier Ltd. All rights reserved. *Corresponding author Keywords: precursor polypeptide; SecB; export; site-directed spin labeling; chaperone Introduction Export of polypeptides into the periplasmic space through the cytoplasmic membrane in the Gram negative bacterium Escherichia coli is mediated by two systems. One, the Tat system, handles proteins that fold into stable structures in the cytosol before they are translocated. 1 The other, the general secretory, or Sec, system cannot handle folded proteins at all but must transfer polypeptides before they acquire their final folded structure. 2 In addition to a pathway through the membrane, provided by the translocon comprising a SecYEG core and the accessory proteins SecDF, YajC, the general secretory system includes soluble chaperones that allow early capture of precursor polypeptides. SecB is such a chaperone. It binds to its ligands with high affinity, selecting them by virtue of their non-native state. The complex undergoes a rapid equilibrium between the bound and free states. The rate of folding of the polypeptide relative to the rate of binding SecB determines a kinetic partitioning between the irre- versible non-productive pathway of folding in the cytosol and the pathway of export. 3 SecB and its bound ligand forms a complex with SecA which itself has affinity for SecY. Thus, the ternary complex of SecA:SecB:precursor engages the translocon. SecA, itself an ATPase, undergoes a cycle of ATP binding and hydrolysis to achieve transfer of the ligand from SecB to SecA and subsequently through the Abbreviations used: EPR, electron paramagnetic resonance; MALDI, matrix-assisted laser desorption ionization. E-mail address of the corresponding author: [email protected] doi:10.1016/j.jmb.2006.07.021 J. Mol. Biol. (2006) 363, 63–74 0022-2836/$ - see front matter © 2006 Elsevier Ltd. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1016/j.jmb.2006.07.021 J. Mol. Biol. (2006) 363, 63–74
Sites of Interaction of a Precursor Polypeptide on theExport Chaperone SecB Mapped by Site-directed SpinLabeling
Jennine M. Crane1, Yuying Suo1, Angela A. Lilly1, Chunfeng Mao1
Wayne L. Hubbell2 and Linda L. Randall1⁎
1Department of Biochemistry,University of Missouri,Columbia, MO 65211, USA2Jules Stein Eye Institute andDepartment of Chemistry andBiochemistry, University ofCalifornia, Los Angeles,CA 90095-7008, USAAbbreviations used: EPR, electronresonance; MALDI, matrix-assistedionization.E-mail address of the correspondi
0022-2836/$ - see front matter © 2006 E
Export of protein into the periplasm of Escherichia coli via the generalsecretory system requires that the transported polypeptides be devoid ofstably folded tertiary structure. Capture of the precursor polypeptidesbefore they fold is achieved by the promiscuous binding to the chaperoneSecB. SecB delivers its ligand to export sites through its specific binding toSecA, a peripheral component of the membrane translocon. At thetranslocon the ligand is passed from SecB to SecA and subsequentlythrough the SecYEG channel. We have previously used site-directed spinlabeling and electron paramagnetic resonance spectroscopy to establish adocking model between SecB and SecA. Here we report use of the samestrategy to map the pathway of a physiologic ligand, the unfolded form ofprecursor galactose-binding protein, on SecB. Our set of SecB variants eachcontaining a single cysteine, which was used in the previous study, has beenexpanded to 48 residues, which cover 49% of the surface of SecB. Theresidues on SecB involved in contacts were identified as those that, uponaddition of the unfolded polypeptide ligand, showed changes in spectralline shape consistent with restricted motion of the nitroxide. We concludethat the bound precursor makes contact with a large portion of the surfaceof the small chaperone. The sites on SecB that interact with the ligand arecompared with the previously identified sites that interact with SecA and amodel for transfer of the ligand is discussed.
© 2006 Elsevier Ltd. All rights reserved.
Keywords: precursor polypeptide; SecB; export; site-directed spin labeling;chaperone
*Corresponding authorIntroduction
Export of polypeptides into the periplasmic spacethrough the cytoplasmic membrane in the Gramnegative bacterium Escherichia coli is mediated bytwo systems. One, the Tat system, handles proteinsthat fold into stable structures in the cytosol beforethey are translocated.1 The other, the generalsecretory, or Sec, system cannot handle foldedproteins at all but must transfer polypeptides beforethey acquire their final folded structure.2 In addition
paramagneticlaser desorption
ng author:
lsevier Ltd. All rights reserve
to a pathway through the membrane, provided bythe translocon comprising a SecYEG core and theaccessory proteins SecDF, YajC, the general secretorysystem includes soluble chaperones that allow earlycapture of precursor polypeptides. SecB is such achaperone. It binds to its ligands with high affinity,selecting themby virtue of their non-native state. Thecomplex undergoes a rapid equilibrium between thebound and free states. The rate of folding of thepolypeptide relative to the rate of binding SecBdetermines a kinetic partitioning between the irre-versible non-productive pathway of folding in thecytosol and the pathway of export.3 SecB and itsbound ligand forms a complexwith SecAwhich itselfhas affinity for SecY. Thus, the ternary complex ofSecA:SecB:precursor engages the translocon. SecA,itself an ATPase, undergoes a cycle of ATP bindingand hydrolysis to achieve transfer of the ligand fromSecB to SecA and subsequently through the
d.

Figure 1. Measures of mobility. The peak to peakwidth of the central resonance line (ΔHpp) is measured asindicated and is equivalent to the peak width at half-height of an absorbance spectrum. The spectral breadth(2A′zz) is the distance between the outermost hyperfineextrema. The spectrum used to illustrate these parametersis that of L67R1.
64 Ligand Binding on Chaperone Mapped by SDSL and EPR
membrane. Elucidation of the mechanism of transferrequires a detailed description of the contactsbetween the members of the ternary complex.We have previously determined the orientation of
the docking of SecA onto SecB4 and now weinvestigate the binding of ligand. When physiologicligands, precursors of polypeptides of 30 – 50 kDa ina non-native state, that are destined for export to theperiplasmic space bind to SecB, the portion of thepolypeptide that makes contact with the chaperonespans a minimal stretch of 150 amino acid resi-dues.5–7 SecB is a small, homotetrameric chaperoneof 69 kDa organized as a dimer of dimers,8,9 and thusto accommodate such a length of polypeptide theligand must wrap around the surface. Examinationof the X-ray crystal structure of SecB reveals a 70 Ålong groove at the interface of the dimers on eachside of the tetramer.8 It was proposed that theligand binds in this groove. Our study describedhere indicates that in addition to residues in theputative binding groove there are many otherregions of contact between SecB and its ligands.We have defined the binding interface by usingsite-directed spin labeling to examine variants ofSecB each containing a nitroxide spin label at aunique position. Comparison of the spectral lineshapes for SecB free in solution and in complexwith precursor galactose-binding protein allowedidentification of sites of contact between theligand and the chaperone.
Results
We have mapped the surface of contact betweenan unfolded polypeptide ligand that wraps aroundthe chaperone by exploiting the fact that the lineshape of an electron paramagnetic resonance (EPR)spectrum of spin-labeled SecB contains informationabout the mobility of the nitroxide side-chain on thenanosecond timescale (for a review see Schneiderand Freed10 and Fajer11). If in a complex of ligandand chaperone the ligand either makes a directcontact with the nitroxide or is held in close vicinityone would expect the motion of the nitroxide to beconstrained. As a nitroxide goes from highly mobileto constrained or immobile two features of thespectrum change. First, the central line widthbroadens (Figure 1, ΔHpp) which is also seen as adecrease in the intensity of the central line since allspectra are normalized. Second, the overall spectralbreadth increases, that is, the total intensity is spreadover a wider range of the magnetic field. For spectrareflecting slow motional states the hyperfineextrema are well resolved and the separation ofthe extrema can be used as a measure of the spectralbreadth (Figure 1, 2A′zz). Here we use these simpleindicators of mobility, which are apparent by visualinspection, to determine which areas on the surfaceof SecB interact with the ligand. It is a formalpossibility that constraints on the nitroxide resultnot from a direct contact but from a conformationalchange. Nevertheless, determination of the sites that
show constrained mobility should allow us todetermine the surface of SecB that interacts withthe precursor polypeptide ligand.A total of 41 of the 48 residues studied here were
previously examined in our study of docking ofSecA onto spin-labeled SecB.4 The reader is referredto that publication for a detailed description ofwhether the line shapes of those spectra reflectisotropic motion and whether resolved spectralcomponents are due to multiple dynamic states ofthe nitroxide in slow exchange or are due toanisotropic motion of a nitroxide in one dynamicstate.The crystal structure of SecB from E. coli9 was used
as a guide in selection of side-chains for labeling thatare at solvent-exposed sites to avoid disruption ofthe protein structure and to maximize the prob-ability of observing contacts. Cysteine was substi-tuted for the native amino acid at 67 positions.Nineteen of the variant proteins aggregated duringpurification and were not pursued further. Theremaining 48 variants were reacted with a sulfhy-dryl specific methanethiosulfonate reagent to gen-erate a nitroxide side-chain as shown in Figure 2.The spin-labeled proteins were shown to be prop-erly folded and were tested for activity in bindingunfolded precursor galactose-binding protein, anatural ligand of SecB, by size-exclusion chromato-graphy, (data not shown, see Activity assays inMaterials and Methods for details).SecB forms a complex with its natural ligands
only if the polypeptides are presented in a non-native state. The binding is of high affinity (Kd forunfolded precursor galactose-binding protein, theligand used here, is approximately 35 nM12 as
Figure 2. The methanethiosul-fonate spin label and the side-chain(R1) that it generates.
Figure 3. Reversal of complex formation betweenprecursor galactose-binding protein and SecB. Black tracesare free SecB, green traces are SecB and galactose-bindingprotein, blue traces are SecB and galactose-binding proteinafter addition of 3 mM CaCl2 and incubation at roomtemperature for 30 min before acquiring the spectrum at6 °C.
65Ligand Binding on Chaperone Mapped by SDSL and EPR
determined by titration calorimetry in GnHCl atboth 0.1 M and 0.2 M), but the system is in a rapiddynamic equilibrium. Binding to SecB is readilyreversible and each time the ligand is released itundergoes a kinetic partitioning between folding toits native state and rebinding to SecB.13,14 Precursorgalactose-binding protein is an ideal ligand forthese studies because the rate of folding can bemodulated by temperature, as is true for thefolding of all proteins, and also by calcium. Thenative structure of galactose-binding protein isstabilized by a calcium ion and the rate of foldingof the polypeptide is drastically decreased in theabsence of calcium.14 In the experimental designused here the precursor, unfolded in 1 M GnHCl,was added to the SecB by rapid dilution into asolution held on ice so that the final concentrationof the GnHCl was 0.17 M and calcium was chelatedby 1 mM EGTA. The spectra were collected at 6 °C.Under these conditions the rate of folding of theligand is sufficiently slow so that the partitioning ispoised to favor complex formation. Addition ofCaCl2 to 3 mM to the mixture results in a shift ofthe partitioning to favor folding. Thus, the changesin spectral line shape observed upon addition ofprecursor galactose-binding protein to spin-labeledSecB can be shown to be specific for binding of anunfolded ligand by adding CaCl2 to reverse thechange. Figure 3 shows two examples of such areversal of the line shape (compare black traces,free SecB; green traces, SecB and precursor galac-tose-binding protein; blue traces, SecB and pre-cursor galactose-binding protein with CaCl2, notethat the black and blue traces overlay).The spectra of the spin-labeled variants of SecB
free in solution and those of SecB in complex withprecursor were compared to identify areas at whichmotion of the nitroxide side-chain was constrained,indicating contact with the ligand. We have groupedthe sites examined into three classes: those with ahigh degree of visible change in the spectra, thosewith significant changes that are less readily seenand those showing very small change, which weconsider insignificant. It should be noted that thegroupings are qualitative and although constraint ofmotion of a nitroxide does indicate that the residuelies on or close to the pathway taken by the ligand,we cannot assign contributions to the energy ofstabilization of the complex to any particularcontacts. The goal of this study and of our previousstudy of spin-labeled SecB is to map the bindinginterface between SecB and each of its bindingpartners, the unfolded ligand and SecA, so that wemay gain insight into the role of the ternary complexin export. To facilitate comparison of the surface ofcontact with the precursor to that with SecAwe have
included some data from our previous work. Thecombined results from both studies indicate that thebinding sites for precursor and SecA show consider-able overlap.The group of residues that show the highest
degree of constraint when precursor makes acomplex with SecB are shown in Figure 4 (comparethe spectral lines shown in black, free SecB withthose shown in green, SecB in complex withprecursor). The constraints on the motion of thenitroxides when the unfolded ligand binds can bereadily seen as an increase in spectral breadth that isobvious both from the outward movement of thehyperfine extrema and the decreases in amplitude ofthe center lines. The nitroxides at four of thesepositions, K41R1, D43R1, D45R1 and L126R1 showthe same degree of constraint when SecB is incomplex with either the ligand or with SecA. Theother seven positions show either very little contact(A87R1 and P130R1) or no contact (Q14R1, Q33R1,L42R1, S85R1 and N127R1) when SecA binds(Figure 4, compare the black traces, free SecB withthe red traces, SecB in complex with SecA).
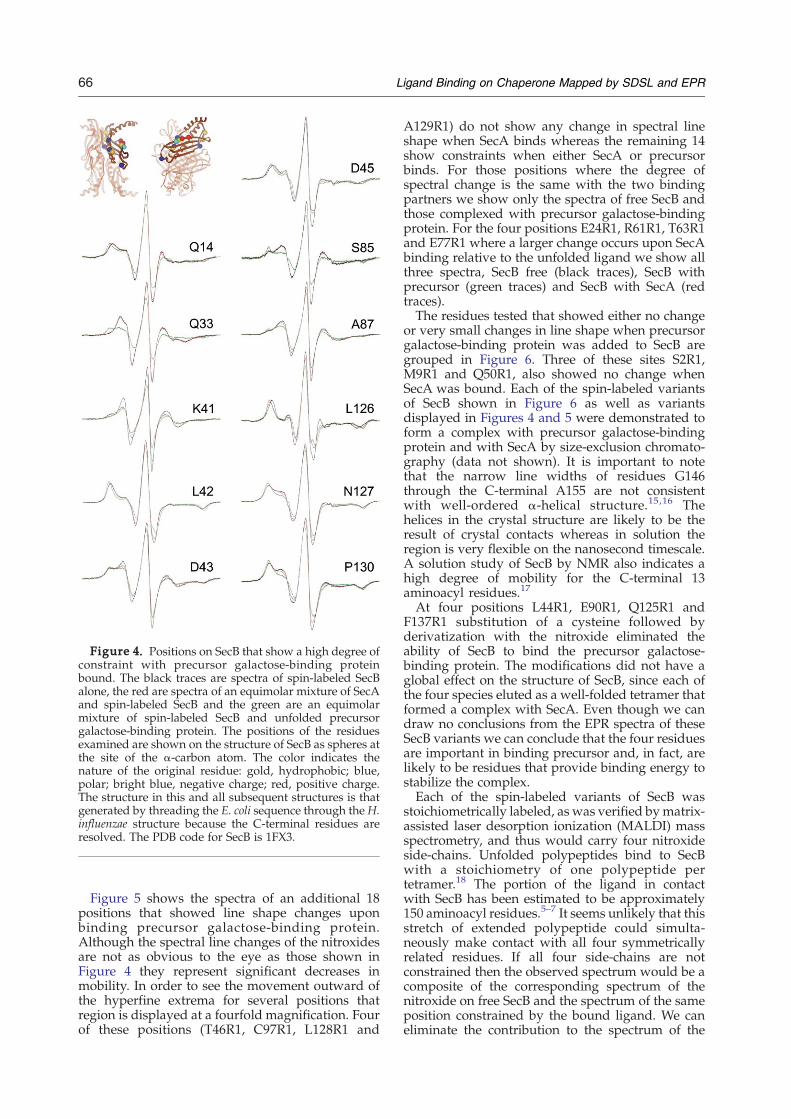
Figure 4. Positions on SecB that show a high degree ofconstraint with precursor galactose-binding proteinbound. The black traces are spectra of spin-labeled SecBalone, the red are spectra of an equimolar mixture of SecAand spin-labeled SecB and the green are an equimolarmixture of spin-labeled SecB and unfolded precursorgalactose-binding protein. The positions of the residuesexamined are shown on the structure of SecB as spheres atthe site of the α-carbon atom. The color indicates thenature of the original residue: gold, hydrophobic; blue,polar; bright blue, negative charge; red, positive charge.The structure in this and all subsequent structures is thatgenerated by threading the E. coli sequence through the H.influenzae structure because the C-terminal residues areresolved. The PDB code for SecB is 1FX3.
66 Ligand Binding on Chaperone Mapped by SDSL and EPR
Figure 5 shows the spectra of an additional 18positions that showed line shape changes uponbinding precursor galactose-binding protein.Although the spectral line changes of the nitroxidesare not as obvious to the eye as those shown inFigure 4 they represent significant decreases inmobility. In order to see the movement outward ofthe hyperfine extrema for several positions thatregion is displayed at a fourfold magnification. Fourof these positions (T46R1, C97R1, L128R1 and
A129R1) do not show any change in spectral lineshape when SecA binds whereas the remaining 14show constraints when either SecA or precursorbinds. For those positions where the degree ofspectral change is the same with the two bindingpartners we show only the spectra of free SecB andthose complexed with precursor galactose-bindingprotein. For the four positions E24R1, R61R1, T63R1and E77R1 where a larger change occurs upon SecAbinding relative to the unfolded ligand we show allthree spectra, SecB free (black traces), SecB withprecursor (green traces) and SecB with SecA (redtraces).The residues tested that showed either no change
or very small changes in line shape when precursorgalactose-binding protein was added to SecB aregrouped in Figure 6. Three of these sites S2R1,M9R1 and Q50R1, also showed no change whenSecA was bound. Each of the spin-labeled variantsof SecB shown in Figure 6 as well as variantsdisplayed in Figures 4 and 5 were demonstrated toform a complex with precursor galactose-bindingprotein and with SecA by size-exclusion chromato-graphy (data not shown). It is important to notethat the narrow line widths of residues G146through the C-terminal A155 are not consistentwith well-ordered α-helical structure.15,16 Thehelices in the crystal structure are likely to be theresult of crystal contacts whereas in solution theregion is very flexible on the nanosecond timescale.A solution study of SecB by NMR also indicates ahigh degree of mobility for the C-terminal 13aminoacyl residues.17
At four positions L44R1, E90R1, Q125R1 andF137R1 substitution of a cysteine followed byderivatization with the nitroxide eliminated theability of SecB to bind the precursor galactose-binding protein. The modifications did not have aglobal effect on the structure of SecB, since each ofthe four species eluted as a well-folded tetramer thatformed a complex with SecA. Even though we candraw no conclusions from the EPR spectra of theseSecB variants we can conclude that the four residuesare important in binding precursor and, in fact, arelikely to be residues that provide binding energy tostabilize the complex.Each of the spin-labeled variants of SecB was
stoichiometrically labeled, as was verified bymatrix-assisted laser desorption ionization (MALDI) massspectrometry, and thus would carry four nitroxideside-chains. Unfolded polypeptides bind to SecBwith a stoichiometry of one polypeptide pertetramer.18 The portion of the ligand in contactwith SecB has been estimated to be approximately150 aminoacyl residues.5–7 It seems unlikely that thisstretch of extended polypeptide could simulta-neously make contact with all four symmetricallyrelated residues. If all four side-chains are notconstrained then the observed spectrum would be acomposite of the corresponding spectrum of thenitroxide on free SecB and the spectrum of the sameposition constrained by the bound ligand. We caneliminate the contribution to the spectrum of the
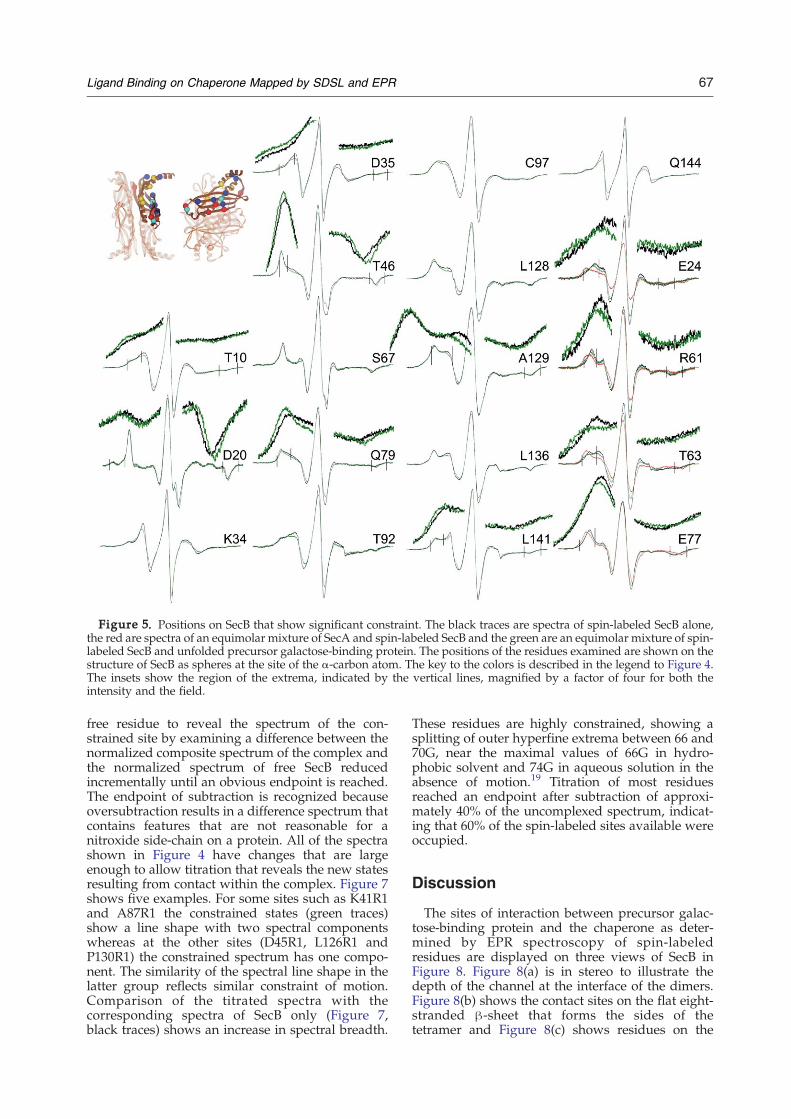
Figure 5. Positions on SecB that show significant constraint. The black traces are spectra of spin-labeled SecB alone,the red are spectra of an equimolar mixture of SecA and spin-labeled SecB and the green are an equimolar mixture of spin-labeled SecB and unfolded precursor galactose-binding protein. The positions of the residues examined are shown on thestructure of SecB as spheres at the site of the α-carbon atom. The key to the colors is described in the legend to Figure 4.The insets show the region of the extrema, indicated by the vertical lines, magnified by a factor of four for both theintensity and the field.
67Ligand Binding on Chaperone Mapped by SDSL and EPR
free residue to reveal the spectrum of the con-strained site by examining a difference between thenormalized composite spectrum of the complex andthe normalized spectrum of free SecB reducedincrementally until an obvious endpoint is reached.The endpoint of subtraction is recognized becauseoversubtraction results in a difference spectrum thatcontains features that are not reasonable for anitroxide side-chain on a protein. All of the spectrashown in Figure 4 have changes that are largeenough to allow titration that reveals the new statesresulting from contact within the complex. Figure 7shows five examples. For some sites such as K41R1and A87R1 the constrained states (green traces)show a line shape with two spectral componentswhereas at the other sites (D45R1, L126R1 andP130R1) the constrained spectrum has one compo-nent. The similarity of the spectral line shape in thelatter group reflects similar constraint of motion.Comparison of the titrated spectra with thecorresponding spectra of SecB only (Figure 7,black traces) shows an increase in spectral breadth.
These residues are highly constrained, showing asplitting of outer hyperfine extrema between 66 and70G, near the maximal values of 66G in hydro-phobic solvent and 74G in aqueous solution in theabsence of motion.19 Titration of most residuesreached an endpoint after subtraction of approxi-mately 40% of the uncomplexed spectrum, indicat-ing that 60% of the spin-labeled sites available wereoccupied.
Discussion
The sites of interaction between precursor galac-tose-binding protein and the chaperone as deter-mined by EPR spectroscopy of spin-labeledresidues are displayed on three views of SecB inFigure 8. Figure 8(a) is in stereo to illustrate thedepth of the channel at the interface of the dimers.Figure 8(b) shows the contact sites on the flat eight-stranded β-sheet that forms the sides of thetetramer and Figure 8(c) shows residues on the
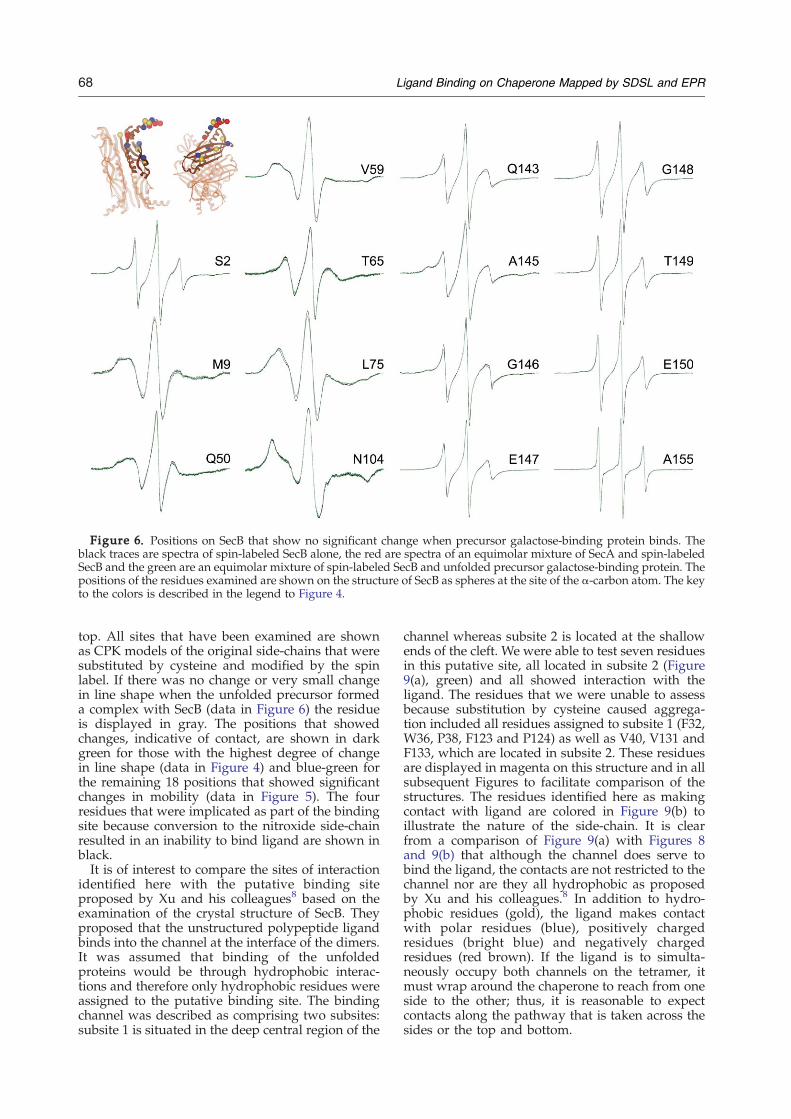
Figure 6. Positions on SecB that show no significant change when precursor galactose-binding protein binds. Theblack traces are spectra of spin-labeled SecB alone, the red are spectra of an equimolar mixture of SecA and spin-labeledSecB and the green are an equimolar mixture of spin-labeled SecB and unfolded precursor galactose-binding protein. Thepositions of the residues examined are shown on the structure of SecB as spheres at the site of the α-carbon atom. The keyto the colors is described in the legend to Figure 4.
68 Ligand Binding on Chaperone Mapped by SDSL and EPR
top. All sites that have been examined are shownas CPK models of the original side-chains that weresubstituted by cysteine and modified by the spinlabel. If there was no change or very small changein line shape when the unfolded precursor formeda complex with SecB (data in Figure 6) the residueis displayed in gray. The positions that showedchanges, indicative of contact, are shown in darkgreen for those with the highest degree of changein line shape (data in Figure 4) and blue-green forthe remaining 18 positions that showed significantchanges in mobility (data in Figure 5). The fourresidues that were implicated as part of the bindingsite because conversion to the nitroxide side-chainresulted in an inability to bind ligand are shown inblack.It is of interest to compare the sites of interaction
identified here with the putative binding siteproposed by Xu and his colleagues8 based on theexamination of the crystal structure of SecB. Theyproposed that the unstructured polypeptide ligandbinds into the channel at the interface of the dimers.It was assumed that binding of the unfoldedproteins would be through hydrophobic interac-tions and therefore only hydrophobic residues wereassigned to the putative binding site. The bindingchannel was described as comprising two subsites:subsite 1 is situated in the deep central region of the
channel whereas subsite 2 is located at the shallowends of the cleft. We were able to test seven residuesin this putative site, all located in subsite 2 (Figure9(a), green) and all showed interaction with theligand. The residues that we were unable to assessbecause substitution by cysteine caused aggrega-tion included all residues assigned to subsite 1 (F32,W36, P38, F123 and P124) as well as V40, V131 andF133, which are located in subsite 2. These residuesare displayed in magenta on this structure and in allsubsequent Figures to facilitate comparison of thestructures. The residues identified here as makingcontact with ligand are colored in Figure 9(b) toillustrate the nature of the side-chain. It is clearfrom a comparison of Figure 9(a) with Figures 8and 9(b) that although the channel does serve tobind the ligand, the contacts are not restricted to thechannel nor are they all hydrophobic as proposedby Xu and his colleagues.8 In addition to hydro-phobic residues (gold), the ligand makes contactwith polar residues (blue), positively chargedresidues (bright blue) and negatively chargedresidues (red brown). If the ligand is to simulta-neously occupy both channels on the tetramer, itmust wrap around the chaperone to reach from oneside to the other; thus, it is reasonable to expectcontacts along the pathway that is taken across thesides or the top and bottom.
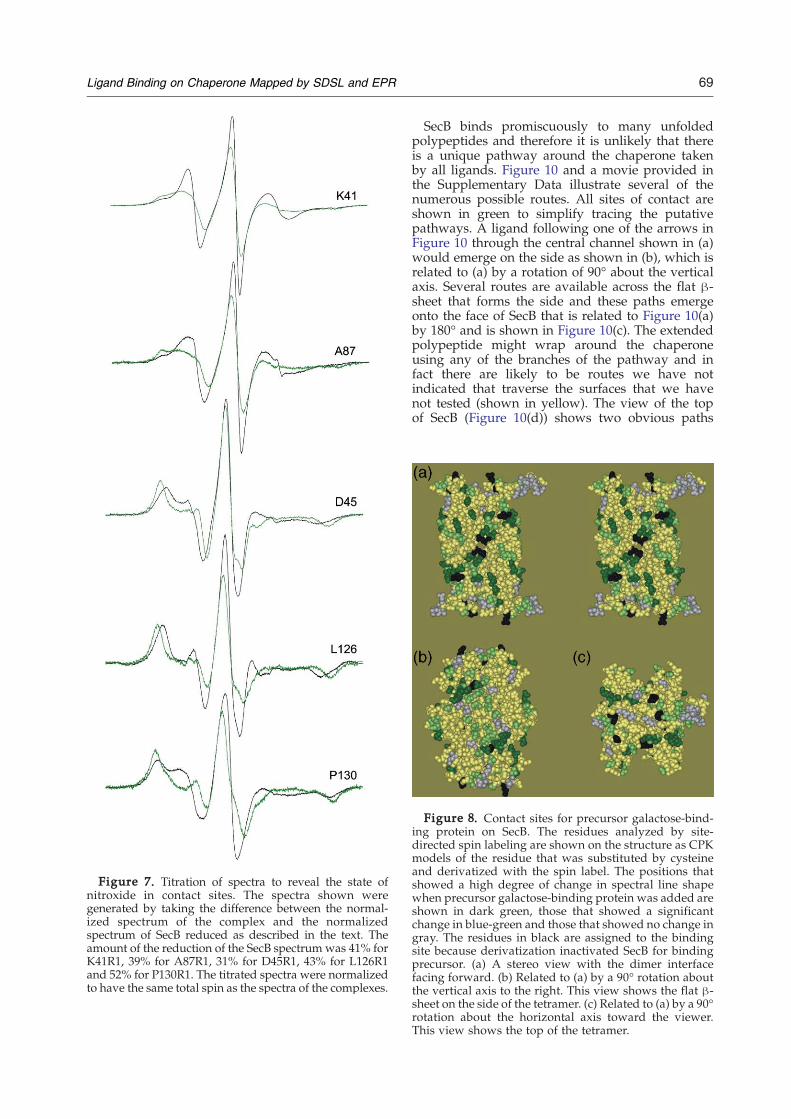
Figure 7. Titration of spectra to reveal the state ofnitroxide in contact sites. The spectra shown weregenerated by taking the difference between the normal-ized spectrum of the complex and the normalizedspectrum of SecB reduced as described in the text. Theamount of the reduction of the SecB spectrum was 41% forK41R1, 39% for A87R1, 31% for D45R1, 43% for L126R1and 52% for P130R1. The titrated spectra were normalizedto have the same total spin as the spectra of the complexes.
69Ligand Binding on Chaperone Mapped by SDSL and EPR
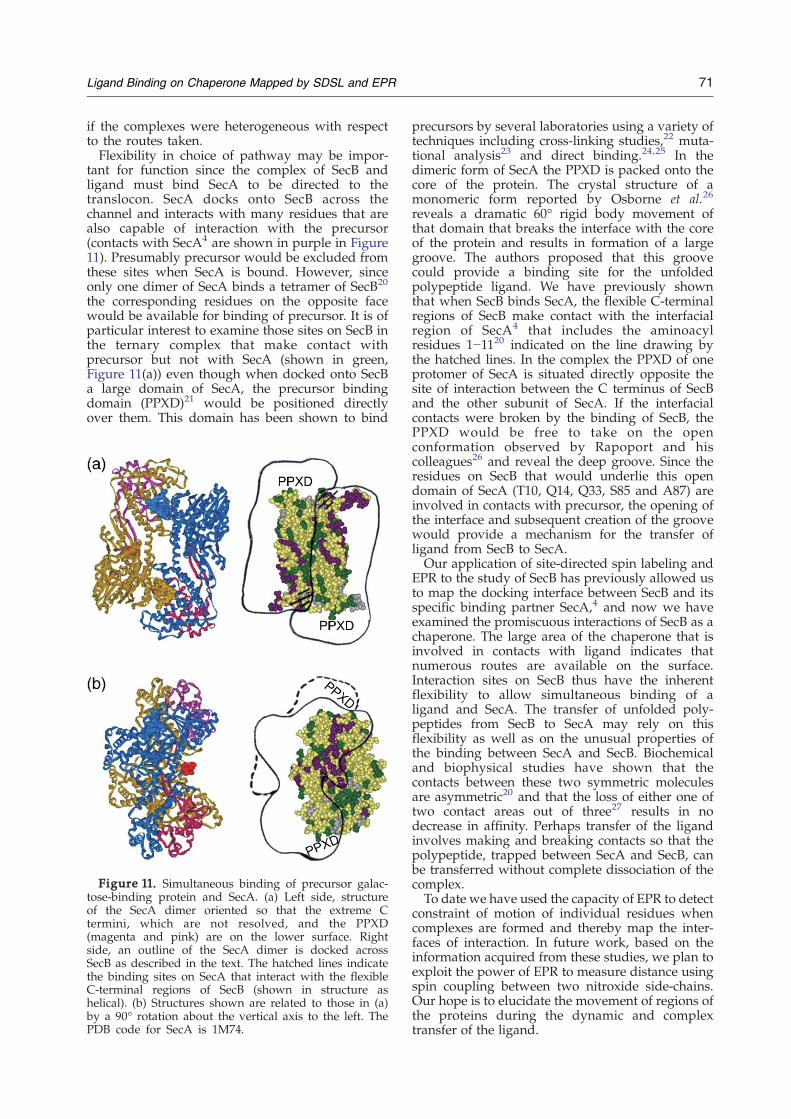
SecB binds promiscuously to many unfoldedpolypeptides and therefore it is unlikely that thereis a unique pathway around the chaperone takenby all ligands. Figure 10 and a movie provided inthe Supplementary Data illustrate several of thenumerous possible routes. All sites of contact areshown in green to simplify tracing the putativepathways. A ligand following one of the arrows inFigure 10 through the central channel shown in (a)would emerge on the side as shown in (b), which isrelated to (a) by a rotation of 90° about the verticalaxis. Several routes are available across the flat β-sheet that forms the side and these paths emergeonto the face of SecB that is related to Figure 10(a)by 180° and is shown in Figure 10(c). The extendedpolypeptide might wrap around the chaperoneusing any of the branches of the pathway and infact there are likely to be routes we have notindicated that traverse the surfaces that we havenot tested (shown in yellow). The view of the topof SecB (Figure 10(d)) shows two obvious paths
Figure 8. Contact sites for precursor galactose-bind-ing protein on SecB. The residues analyzed by site-directed spin labeling are shown on the structure as CPKmodels of the residue that was substituted by cysteineand derivatized with the spin label. The positions thatshowed a high degree of change in spectral line shapewhen precursor galactose-binding protein was added areshown in dark green, those that showed a significantchange in blue-green and those that showed no change ingray. The residues in black are assigned to the bindingsite because derivatization inactivated SecB for bindingprecursor. (a) A stereo view with the dimer interfacefacing forward. (b) Related to (a) by a 90° rotation aboutthe vertical axis to the right. This view shows the flat β-sheet on the side of the tetramer. (c) Related to (a) by a 90°rotation about the horizontal axis toward the viewer.This view shows the top of the tetramer.
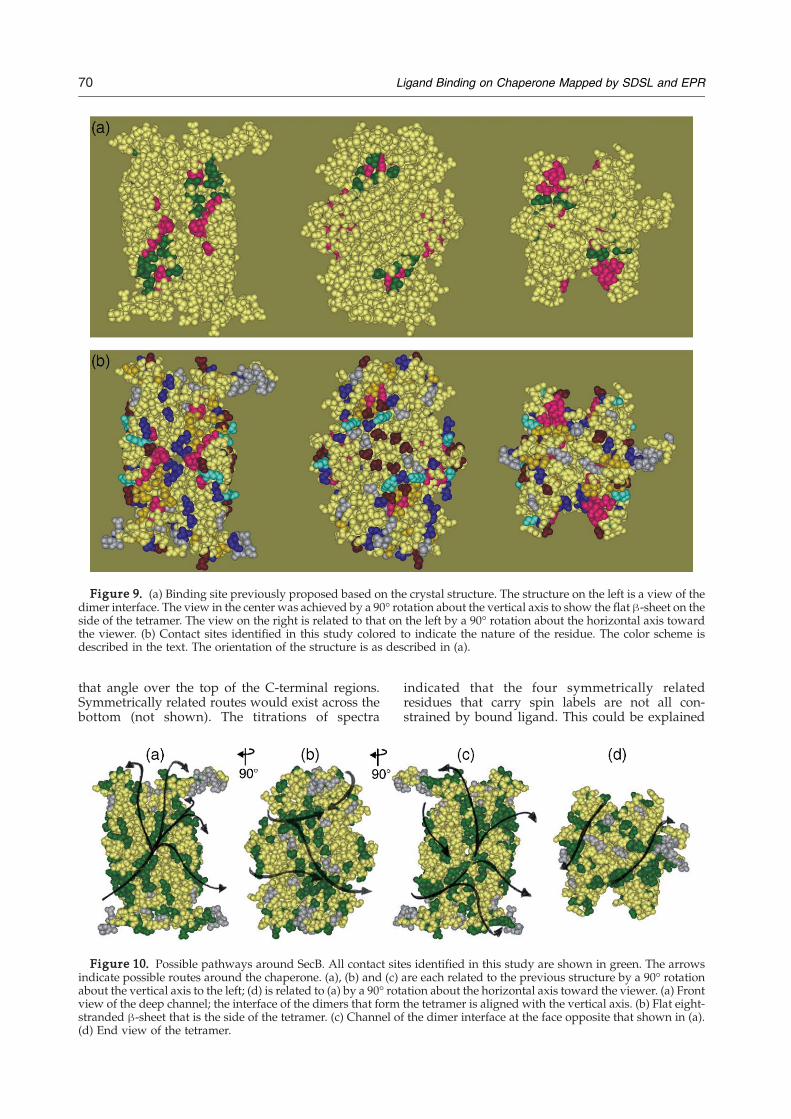
Figure 9. (a) Binding site previously proposed based on the crystal structure. The structure on the left is a view of thedimer interface. The view in the center was achieved by a 90° rotation about the vertical axis to show the flat β-sheet on theside of the tetramer. The view on the right is related to that on the left by a 90° rotation about the horizontal axis towardthe viewer. (b) Contact sites identified in this study colored to indicate the nature of the residue. The color scheme isdescribed in the text. The orientation of the structure is as described in (a).
70 Ligand Binding on Chaperone Mapped by SDSL and EPR
that angle over the top of the C-terminal regions.Symmetrically related routes would exist across thebottom (not shown). The titrations of spectra
Figure 10. Possible pathways around SecB. All contact sitindicate possible routes around the chaperone. (a), (b) and (c)about the vertical axis to the left; (d) is related to (a) by a 90° rotview of the deep channel; the interface of the dimers that formstranded β-sheet that is the side of the tetramer. (c) Channel of(d) End view of the tetramer.
indicated that the four symmetrically relatedresidues that carry spin labels are not all con-strained by bound ligand. This could be explained
es identified in this study are shown in green. The arrowsare each related to the previous structure by a 90° rotationation about the horizontal axis toward the viewer. (a) Frontthe tetramer is aligned with the vertical axis. (b) Flat eight-the dimer interface at the face opposite that shown in (a).
71Ligand Binding on Chaperone Mapped by SDSL and EPR
if the complexes were heterogeneous with respectto the routes taken.Flexibility in choice of pathway may be impor-
tant for function since the complex of SecB andligand must bind SecA to be directed to thetranslocon. SecA docks onto SecB across thechannel and interacts with many residues that arealso capable of interaction with the precursor(contacts with SecA4 are shown in purple in Figure11). Presumably precursor would be excluded fromthese sites when SecA is bound. However, sinceonly one dimer of SecA binds a tetramer of SecB20
the corresponding residues on the opposite facewould be available for binding of precursor. It is ofparticular interest to examine those sites on SecB inthe ternary complex that make contact withprecursor but not with SecA (shown in green,Figure 11(a)) even though when docked onto SecBa large domain of SecA, the precursor bindingdomain (PPXD)21 would be positioned directlyover them. This domain has been shown to bind
Figure 11. Simultaneous binding of precursor galac-tose-binding protein and SecA. (a) Left side, structureof the SecA dimer oriented so that the extreme Ctermini, which are not resolved, and the PPXD(magenta and pink) are on the lower surface. Rightside, an outline of the SecA dimer is docked acrossSecB as described in the text. The hatched lines indicatethe binding sites on SecA that interact with the flexibleC-terminal regions of SecB (shown in structure ashelical). (b) Structures shown are related to those in (a)by a 90° rotation about the vertical axis to the left. ThePDB code for SecA is 1M74.
precursors by several laboratories using a variety oftechniques including cross-linking studies,22 muta-tional analysis23 and direct binding.24,25 In thedimeric form of SecA the PPXD is packed onto thecore of the protein. The crystal structure of amonomeric form reported by Osborne et al.26
reveals a dramatic 60° rigid body movement ofthat domain that breaks the interface with the coreof the protein and results in formation of a largegroove. The authors proposed that this groovecould provide a binding site for the unfoldedpolypeptide ligand. We have previously shownthat when SecB binds SecA, the flexible C-terminalregions of SecB make contact with the interfacialregion of SecA4 that includes the aminoacylresidues 1−1120 indicated on the line drawing bythe hatched lines. In the complex the PPXD of oneprotomer of SecA is situated directly opposite thesite of interaction between the C terminus of SecBand the other subunit of SecA. If the interfacialcontacts were broken by the binding of SecB, thePPXD would be free to take on the openconformation observed by Rapoport and hiscolleagues26 and reveal the deep groove. Since theresidues on SecB that would underlie this opendomain of SecA (T10, Q14, Q33, S85 and A87) areinvolved in contacts with precursor, the opening ofthe interface and subsequent creation of the groovewould provide a mechanism for the transfer ofligand from SecB to SecA.Our application of site-directed spin labeling and
EPR to the study of SecB has previously allowed usto map the docking interface between SecB and itsspecific binding partner SecA,4 and now we haveexamined the promiscuous interactions of SecB as achaperone. The large area of the chaperone that isinvolved in contacts with ligand indicates thatnumerous routes are available on the surface.Interaction sites on SecB thus have the inherentflexibility to allow simultaneous binding of aligand and SecA. The transfer of unfolded poly-peptides from SecB to SecA may rely on thisflexibility as well as on the unusual properties ofthe binding between SecA and SecB. Biochemicaland biophysical studies have shown that thecontacts between these two symmetric moleculesare asymmetric20 and that the loss of either one oftwo contact areas out of three27 results in nodecrease in affinity. Perhaps transfer of the ligandinvolves making and breaking contacts so that thepolypeptide, trapped between SecA and SecB, canbe transferred without complete dissociation of thecomplex.To date we have used the capacity of EPR to detect
constraint of motion of individual residues whencomplexes are formed and thereby map the inter-faces of interaction. In future work, based on theinformation acquired from these studies, we plan toexploit the power of EPR to measure distance usingspin coupling between two nitroxide side-chains.Our hope is to elucidate the movement of regions ofthe proteins during the dynamic and complextransfer of the ligand.

72 Ligand Binding on Chaperone Mapped by SDSL and EPR
Materials and Methods
Mutagenesis and protein purification
The strategy of site-directed spin labeling requires thatonly the cysteine at the site of interest be modified by thenitroxide reagent via sulfhydryl chemistry. Ideally allnative cysteine residues in the protein of interest arereplaced and a unique cysteine is introduced at thedesired site. SecB contains four native cysteine residues(C76, C97, C102 and C113). We were unable to replaceC76 or C113 without disrupting the structure of SecB.Fortunately only C97 showed reactivity in the nativeprotein. Therefore to create our base protein for thisstudy we substituted C97 with serine and C102 withleucine. This base protein forms a wild-type complexwith precursor galactose-binding protein as assessed bysize exclusion chromatography (see Size exclusionchromatography below), but does not react with thespin label as shown by MALDI mass spectrometry. Theplasmid pAL204 carrying the gene for the base proteinunder the T7 promoter was modified by standardrecombinant DNA techniques (Quickchange, Strategene)to generate the cysteine mutants used here. Each mutantwas subjected to DNA sequencing to confirm the change.The plasmids were transformed into BL21(DE3) (Nova-gen) or BL21(DE3) secB∷Tn10 srl∷Tn10 recA (a secB nullstrain obtained from Carol Kumamoto). The proteinswere purified from the appropriate strains as describedfor wild-type SecB.20 Precursor galactose-binding proteinwas purified as described.14 The protein concentrationswere determined spectrophotometrically at 280 nm usingextinction coefficients of 47,600 M−1 cm−1 for SecBtetramer and 37,410 M−1 cm−1 for precursor galactose-binding protein.
Spin-labeling of SecB variants
Immediately before labeling with the nitroxide reagent(1-oxyl-2,2,5,5-tetramethylpyrroline-3-methyl)-metha-nethiosulfonate (Toronto Research Chemicals Inc.), thereducing agent, Tris-(2-carboxyethyl) phosphine hydro-chloride (TCEP, Molecular Probes) was removed byexchange from the usual storage buffer (10 mM Hepes-KOH, 300 mM KOAc, 2 mM TCEP (pH 7.6)) into 10 mMHepes-HAc, 300 mM KOAc (pH 6.7) using a Nap10column (Amersham). The proteins were concentratedusing a Centricon 30 (Millipore). Typically 0.2 ml of eachSecB variant, at a concentration of approximately 1.3 mMmonomer, was incubated with an equimolar amount ofthe spin labeling reagent. To generate C97R1 we used thenative, wild-type protein. In the case of residues that wereclose enough to show spin−spin interaction (D20, E24,E77and Q79) the labeling was carried out with a mixtureof the spin label and an analog (a generous gift fromKálmán Hideg), which has no free electron, at a ratio of1:1.5. D35 shows spin−spin interaction across the interfaceof the dimer of dimers. This interaction was reduced bymixing labeled SecBD35R1 with an equal amount ofunlabeled SecB and incubating to allow the spontaneousexchange of dimers. The spin label reagent and the analogwere each prepared as a stock of 100 mM in acetonitrileand stored in the dark at −80 °C. The reaction was allowedto proceed on ice in the dark for 2 – 3 h. To remove excessspin label and to separate the spin-labeled species thatwere tetrameric from aggregated species the entirereaction mixture was applied to a BioSep-SEC-S 3000
column (600 mm×7.8 mm, Phenomenex), equilibrated in10 mM Hepes-HAc, 300 mM KOAc, 5 mMMg(OAc)2 (pH6.7). The proteins that eluted in the position of tetramerwere concentrated using a Nanosep filter concentrator,10,000 MW cut off (Pall Life Sciences). The concentrationof each spin-labeled protein was determined by absor-bance at 280 nm. Each spin-labeled variant was analyzedby MALDI mass spectrometry and in all cases the labelingwas near stoichiometric. The proteins were stored at−80 °C. The spin-labeled variants are designated by theamino acid and residue number that has been substitutedby cysteine and modified by the nitroxide side-chain, R1.Thus, K41R1 indicates that lysine at position 41 wasconverted to cysteine and reacted with the spin labelreagent.
Activity assays
To ascertain that introduction of the nitroxide did notdisrupt structure all spin labeled variants were subjectedto analysis by HPLC using a size-exclusion column (TSKG3000SW, TosoHaas). At four positions (N104, L128,A129, and F137) derivatization of the cysteine by thespin label resulted in considerable aggregation (∼50% insome cases). However, the protein that eluted as atetramer could be isolated by size-exclusion chromato-graphy and if used immediately did not repopulate theaggregate. All spin-labeled variants included in this EPRstudy were shown to be tetrameric and to form complexeswith precursor galactose-binding protein as well as withSecA by size-exclusion chromatography as described(data not shown).20
EPR measurements
EPR spectroscopy was performed on an X-band spectro-meter, a Bruker Elexsys E500 or Bruker EMX, with a highsensitivity resonator. Protein samples of 5 μl (spin labelwas either 80 μM or 160 μM) were loaded into syntheticsilica capillaries (0.6 mm i.d.×0.84 mm o.d.) sealed at oneend. All spectra were acquired using incident microwavepower at 20 mW, and a 100 kHz field modulation of 1 to 4gauss as appropriate. All spectra were recorded at 6 °Cwith a scan width of 100 gauss centered at 3356 gauss.Between 15 and 25 scans were averaged for each spectralline. All spectra were normalized and further analyzedusing the Labview programs written by Christian Alten-bach (UCLA).Precursor galactose-binding protein was chosen as the
natural ligand used in this study because its rate of foldingcan be modulated by temperature and the absence ofcalcium. For each experiment equimolar precursor galac-tose-binding protein, unfolded in 10 mM Hepes-HAc,300 mM KOAc, 5 mM Mg(OAc)2, 1 mM EGTA, 1 MGnHCl (pH 7.6), was added to the spin-labeled SecBvariant (20 or 40 μM) in 10 mM Hepes-Hac, 300 mMKOAc, 5 mM Mg(OAc)2, 1 mM EGTA (pH 6.7) by rapiddilution into a solution held on ice so that the finalconcentration of the GnHCl was 0.17 M. The spectra werecollected at 6 °C in the presence of 1 mM EGTA to chelatethe calcium in order to slow the rate of folding ofprecursor. As a control for the effect of GnHCl bufferalone was added to spin-labeled SecB in place of precursorgalactose-binding protein so that the final concentration ofGnHCl was 0.17MGnHCl. The Kd for the complex formedbetween SecB and precursor galactose-binding protein isapproximately 35 nM.12

73Ligand Binding on Chaperone Mapped by SDSL and EPR
Calculation of timescale of tumbling
EPR is sensitive to motion of the nitroxide side-chain inthe timescale of 0.1 ns to 20 ns. The molecular tumbling ofSecB (τc=60 ns) is too slow to be averaged into the spectrareported here and thus the spectra are interpreted in termsof local fluctuations of the backbone and internal motionof the nitroxide side-chain. The timescale of tumbling (τc)for SecB was calculated from the rotational diffusionconstant (Dr) of the protein as follows:
H c ¼ 16 Dr
; Dr ¼ kT8kDa3
;
where k is Boltzmann's constant, T is temperature, η is theviscosity of the solvent, and a is the radius of hydration.We have determined the radius of hydration of SecB to be3.3 nm by quasi-elastic light scatter using an in-linedetector (Wyatt QELS, Wyatt Technology Corporation)following chromatography on a BioSep-SEC-S4000 sizeexclusion column (7.8 mm internal diameter × 30 cm,Phenomenex) in 10 mM Hepes-KOH, 300 mM KOAc,5 mM Mg(OAc)2 (pH 7.6) at 7 °C. The ASTRA softwareprovided with the instrument was used to calculate thehydrodynamic radius from the diffusion coefficientmeasured for material at the apex of the chromatographicpeak. Using 3.3 nm as the radius, τc for SecB is calculatedto be 60 ns.
Calculation of surface area
The Hemophilus influenzae structure has more residuesresolved than does the E. coli structure. Therefore, forsurface area calculations a homology model was createdby threading the sequence of E. coli SecB through thestructure of the H. influenzae SecB. The accessible surfacearea of individual residues of SecB was calculated usingCalc-surface Program28 (available at the NIH Computa-tional Molecular Biology web site†) with a probe size of1.4 Å. Because of limitations in the electron density of thecrystal structure, it was possible to obtain surface areas foronly 176 of the 192 mutated residues in the tetramer (48mutations per protomer). The surface area of missingresidues was reported as the average of the surface areafor that residue on the other protomers.
Generation of movie provided in Supplementary Data
Still images were generated using the PyMol moleculargraphics system v0.99. The Connolly solvent surface wasgenerated using a probe with radius 1.4 Å.
Acknowledgements
We thank Gseping Liu for purifying the SecA usedin the activity assays in this study, Beverly B. DaGueof The Proteomics Center at the University ofMissouri for MALDI mass spectrometry, ChristianAltenbach of UCLA for providing the software usedfor data analysis, Hilary C. Vasel for technical
†http://molbio.info.nih.gov/molbio/
assistance and Kálmán Hideg, University of Pécs,Hungary, for the generous gift of the analog of thespin label. We are grateful to Dylan B. Cooper forgeneration of themovie in SupplementaryData. Thiswork was supported in part by NIH research grantsGM29798 (L.L.R.) and EY05216 (W.L.H.), an endow-ment from the Hugo Wurdack Trust at the Uni-versity of Missouri (L.L.R.), the Jules Stein ProfessorEndowment (W.L.H.), and the Bruce Ford Bundyand Anne Smith Bundy Foundation (W.L.H.).
Supplementary Data
Supplementary data associated with this articlecan be found, in the online version, at doi:10.1016/j.jmb.2006.07.021
References
1. Berks, B. C., Sargent, F. & Palmer, T. (2000). The Tatprotein export pathway. Mol. Microbiol. 35, 260–274.
2. Economou, A. (2002). Bacterial secretome: the assem-bly manual and operating instructions. Mol. Membr.Biol. 19, 159–169.
3. Randall, L. L. & Hardy, S. J. S. (1995). High selectivitywith low specificity: how SecB has solved theparadox of chaperone binding. Trends Biochem. Sci.20, 65–69.
4. Crane, J. M., Mao, C., Lilly, A. A., Smith, V. F., Suo, Y.,Hubbell, W. L. & Randall, L. L. (2005). Mapping of thedocking of SecA onto the chaperone SecB by site-directed spin labeling: insight into the mechanism ofligand transfer during protein export. J. Mol. Biol. 353,295–307.
5. Topping, T. B. & Randall, L. L. (1994). Determinationof the binding frame within a physiological ligand forthe chaperone SecB. Protein Sci. 3, 730–736.
6. Khisty, V. J., Munske, G. R. & Randall, L. L. (1995).Mapping of the binding frame for the chaperoneSecB within a natural ligand, galactose-bindingprotein. J. Biol. Chem. 270, 25920–25927.
7. Smith, V. F., Hardy, S. J. S. & Randall, L. L. (1997).Determination of the binding frame of the chaperoneSecB within the physiological ligand oligopeptide-binding protein. Protein Sci. 6, 1746–1755.
8. Xu, Z., Knafels, J. D. & Yoshino, K. (2000). Crystalstructure of the bacterial protein export chaperoneSecB. Nature Struct. Biol. 7, 1172–1177.
9. Dekker, C., de Kruijff, B. & Gros, P. (2003). Crystalstructure of SecB from Escherichia coli. J. Struct. Biol.144, 313–319.
10. Schneider, D. & Freed, J. (1989). Calculating slowmotional resonance spectra: a user's guide. In SpinLabeling: Theory and Application, Biological MagneticResonance (Berliner, L.&Reuben, J., eds), vol. 8, pp. 1–76.
11. Fajer, P. G. (2000). Electron spin resonance spectro-scopy labeling in peptide and protein analysis. InEncyclopedia of Analytical Chemistry (Meyers, R. A.,ed.), pp. 5725–5761, John Wiley & Sons Ltd,London.
12. Randall, L. L., Topping, T. B., Suciu, D. &Hardy, S. J. S.(1998). Calorimetric analyses of the interactionbetween SecB and its ligands. Protein Sci. 7, 1195–1200.
13. Hardy, S. J. S. & Randall, L. L. (1991). A kineticpartitioning model of selective binding of nonnative

74 Ligand Binding on Chaperone Mapped by SDSL and EPR
proteins by the bacterial chaperone SecB. Science, 251,439–443.
14. Topping, T. B. & Randall, L. L. (1997). Chaperone SecBfrom Escherichia colimediates kinetic partitioning via adynamic equilibrium with its ligands. J. Biol. Chem.272, 19314–19318.
15. Isas, J. M., Langen, R., Haigler, H. T. & Hubbell, W. L.(2002). Structure and dynamics of a helical hairpinand loop region in annexin 12: a site-directed spinlabeling study. Biochemistry, 41, 1464–1473.
16. Columbus, L., Kalai, T., Jeko, J., Hideg, K. &Hubbell, W. L. (2001). Molecular motion of spinlabeled side chains in alpha-helices: analysis byvariation of side chain structure. Biochemistry, 40,3828–3846.
17. Volkert, T. L., Baleja, J. D. & Kumamoto, C. A. (1999).A highly mobile C-terminal tail of the Escherichia coliprotein export chaperone SecB. Biochem. Biophys. Res.Commun. 264, 949–954.
18. Bruce, J. E., Smith, V. F., Liu, C., Randall, L. L. & Smith,R. D. (1998). The observation of chaperone-ligandnoncovalent complexes with electrospray ionizationmass spectrometry. Protein Sci. 7, 1180–1185.
19. Gross, A., Columbus, L., Hideg, K., Altenbach, C. &Hubbell, W. L. (1999). Structure of the KcsA potassiumchannel from Streptomyces lividans: a site-directed spinlabeling study of the second transmembrane segment.Biochemistry, 38, 10324–10335.
20. Randall, L. L., Crane, J. M., Lilly, A. A., Liu, G., Mao,C., Patel, C. N. & Hardy, S. J. S. (2005). Asymmetricbinding between SecA and SecB two symmetricproteins: implications for function in export. J. Mol.Biol. 348, 479–489.
21. Hunt, J. F., Weinkauf, S., Henry, L., Fak, J. J.,McNicholas, P., Oliver, D. B. & Deisenhofer, J.(2002). Nucleotide control of interdomain interactionsin the conformational reaction cycle of SecA. Science,297, 2018–2026.
22. Kimura, E., Akita, M., Matsuyama, S. &Mizushima, S.(1991). Determination of a region in SecA thatinteracts with presecretory proteins in Escherichiacoli. J. Biol. Chem. 266, 6600–6606.
23. Kourtz, L. & Oliver, D. (2000). Tyr-326 plays a criticalrole in controlling SecA-preprotein interaction. Mol.Microbiol. 37, 1342–1356.
24. Baud, C., Karamanou, S., Sianidis, G., Vrontou, E.,Politou, A. S. & Economou, A. (2002). Allostericcommunication between signal peptides and theSecA protein DEAD motor ATPase domain. J. Biol.Chem. 277, 13724–13731.
25. Papanikou, E., Karamanou, S., Baud, C., Frank, M.,Sianidis, G., Keramisanou, D. et al. (2005). Identifica-tion of the preprotein binding domain of SecA. J. Biol.Chem. 280, 43209–43217.
26. Osborne, A. R., Clemons, W. M., Jr & Rapoport, T. A.(2004). A large conformational change of the translo-cation ATPase SecA. Proc. Natl Acad. Sci. USA, 101,10937–10942.
27. Patel, C. N., Smith, V. F. & Randall, L. L. (2006).Characterization of three areas of interactions stabiliz-ing complexes between SecA and SecB, two proteinsinvolved in protein export. Protein Sci. 15, 1379–1386.
28. Gerstein, M. (1992). A resolution-sensitive procedurefor comparing protein surfaces and its application tothe comparison of antigen-combining sites. ActaCrystallog. sect. A, 48, 271–276.
Edited by F. Schmid
(Received 8 May 2006; received in revised form 10 July 2006; accepted 11 July 2006)Available online 15 July 2006
Related Documents











