Single-Step Nitrification Models Erroneously Describe Batch Ammonia Oxidation Profiles when Nitrite Oxidation Becomes Rate Limiting Kartik Chandran, 1 Barth F. Smets 1,2 1 Environmental Engineering Program, University of Connecticut, Storrs, Connecticut 2 Department of Civil & Environmental Engineering, University of Connecticut, Storrs, Connecticut 06269, telephone: (860) 486-2270; e-mail: [email protected]. Received 24 June 1999; accepted 30 November 1999 Abstract: Nitrification involves the sequential biological oxidation of reduced nitrogen species such as ammo- nium-nitrogen (NH 4 + -N) to nitrite-nitrogen (NO 2 - -N) and nitrate-nitrogen (NO 3 - -N). The adequacy of modeling NH 4 + -N to NO 3 - -N oxidation as one composite biochemi- cal reaction was examined at different relative dynamics of NH 4 + -N to NO 2 - -N and NO 2 - -N to NO 3 - -N oxidation. NH 4 + -N to NO 2 - -N oxidation and NO 2 - -N to NO 3 - -N oxi- dation by a mixed nitrifying consortium were uncoupled using selective inhibitors allylthiourea and sodium azide. The kinetic parameters of NH 4 + -N to NO 2 - -N oxidation (q max,ns and K S,ns ) and NO 2 - -N to NO 3 - -N oxidation (q max,nb and K S,nb ) were determined by a rapid extant respirometric technique. The stoichiometric coefficients relating nitrogen removal, oxygen uptake and biomass synthesis were derived from an electron balanced equa- tion. NH 4 + -N to NO 2 - -N oxidation was not affected by NO 2 - -N concentrations up to 100 mg NO 2 - -N L -1 . NO 2 - -N to NO 3 - -N oxidation was noncompetitively inhibited by NH 4 + -N but was not inhibited by NO 3 - -N concentrations up to 250 mg NO 3 - -N L -1 . When NH 4 + -N to NO 2 - -N oxi- dation was the sole rate-limiting step, complete NH 4 + -N to NO 3 - -N oxidation was adequately modeled as one composite process. However, when NH 4 + -N to NO 2 - -N oxidation and NO 2 - -N to NO 3 - -N oxidation were both rate limiting, the estimated lumped kinetic parameter es- timates describing NH 4 + -N to NO 3 - -N oxidation were un- realistically high and correlated. These findings indicate that the use of single-step models to describe batch NH 4 + oxidation yields erroneous kinetic parameters when NH 4 + -to-NO 2 - oxidation is not the sole rate-limiting pro- cess throughout the assay. Under such circumstances, it is necessary to quantify NH 4 + -N to NO 2 - -N oxidation and NO 2 - -N to NO 3 - -N oxidation, independently. © 2000 John Wiley & Sons, Inc. Biotechnol Bioeng 68: 396–406, 2000. Keywords: nitrification; biokinetics; extant respirometry; single-step modeling; dual rate-limitation INTRODUCTION Nitrification involves the conversion of ammonium- nitrogen (NH 4 + -N) to nitrite-nitrogen (NO 2 - -N) and nitrate- nitrogen (NO 3 - -N). Traditionally, nitrification has been mathematically treated as one composite biochemical pro- cess, with the assumption that NH 4 + -N to NO 2 - -N oxidation limits the overall transformation of NH 4 + -N to NO 3 - -N (Grady et al., 1999; Henze et al., 1987). However, when NH 4 + -N and NO 2 - -N oxidation both limit overall nitrifica- tion at different stages of the process, the single-step rep- resentation is inadequate. Under such circumstances, the individual characterization of both NH 4 + -N and NO 2 - -N oxidation is necessary. An accurate knowledge of the kinet- ics of each step is required to formulate appropriate design and control strategies for achieving desired NH 4 + -N and NO 2 - -N removal. Numerous biokinetic studies have been performed using pure nitrifying cultures, nitrifying enrich- ment consortia and activated sludge (Painter, 1970; Sharma and Ahlert, 1977). However, significant variation exists in literature with respect to the reported kinetic estimates due to the wide array of biokinetic estimation techniques em- ployed. Additionally, the initial conditions imposed on some biokinetic assays render the resulting kinetic param- eters inappropriate for extrapolation to continuous reactor configurations (Volskay and Grady, 1988). Because batch biokinetic assays are often used to estimate kinetic param- eters for use in continuous reactor modeling, we considered it imperative to evaluate whether single-step nitrification models are appropriate to describe NH 4 + -N oxidation pro- files obtained from batch assays. The primary objectives of this study were as follows: 1. To quantify, the individual steps of NH 4 + -N and NO 2 - -N oxidation by a nitrifying enrichment culture using an extant respirometic technique. 2. To examine the effect of principal nitrogen species on each oxidation step. 3. To determine the kinetics of complete NH 4 + -N to NO 3 - -N oxidation using a single-step nitrification model. 4. To evaluate the adequacy of the single-step nitrification model in depicting complete NH 4 + -N oxidation by com- Correspondence to: B. F. Smets © 2000 John Wiley & Sons, Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Single-Step Nitrification ModelsErroneously Describe Batch AmmoniaOxidation Profiles when Nitrite OxidationBecomes Rate Limiting
Kartik Chandran,1 Barth F. Smets1,2
1Environmental Engineering Program, University of Connecticut,Storrs, Connecticut2Department of Civil & Environmental Engineering, University ofConnecticut, Storrs, Connecticut 06269, telephone: (860) 486-2270;e-mail: [email protected].
Received 24 June 1999; accepted 30 November 1999
Abstract: Nitrification involves the sequential biologicaloxidation of reduced nitrogen species such as ammo-nium-nitrogen (NH4
+-N) to nitrite-nitrogen (NO2−-N) and
nitrate-nitrogen (NO3−-N). The adequacy of modeling
NH4+-N to NO3
−-N oxidation as one composite biochemi-cal reaction was examined at different relative dynamicsof NH4
+-N to NO2−-N and NO2
−-N to NO3−-N oxidation.
NH4+-N to NO2
−-N oxidation and NO2−-N to NO3
−-N oxi-dation by a mixed nitrifying consortium were uncoupledusing selective inhibitors allylthiourea and sodium azide.The kinetic parameters of NH4
+-N to NO2−-N oxidation
(qmax,ns and KS,ns) and NO2−-N to NO3
−-N oxidation(qmax,nb and KS,nb) were determined by a rapid extantrespirometric technique. The stoichiometric coefficientsrelating nitrogen removal, oxygen uptake and biomasssynthesis were derived from an electron balanced equa-tion. NH4
+-N to NO2−-N oxidation was not affected by
NO2−-N concentrations up to 100 mg NO2
−-N L−1. NO2−-N
to NO3−-N oxidation was noncompetitively inhibited by
NH4+-N but was not inhibited by NO3
−-N concentrationsup to 250 mg NO3
−-N L−1. When NH4+-N to NO2
−-N oxi-dation was the sole rate-limiting step, complete NH4
+-Nto NO3
−-N oxidation was adequately modeled as onecomposite process. However, when NH4
+-N to NO2−-N
oxidation and NO2−-N to NO3
−-N oxidation were bothrate limiting, the estimated lumped kinetic parameter es-timates describing NH4
+-N to NO3−-N oxidation were un-
realistically high and correlated. These findings indicatethat the use of single-step models to describe batch NH4
+
oxidation yields erroneous kinetic parameters whenNH4
+-to-NO2− oxidation is not the sole rate-limiting pro-
cess throughout the assay. Under such circumstances, itis necessary to quantify NH4
+-N to NO2−-N oxidation and
NO2−-N to NO3
−-N oxidation, independently. © 2000 JohnWiley & Sons, Inc. Biotechnol Bioeng 68: 396–406, 2000.Keywords: nitrification; biokinetics; extant respirometry;single-step modeling; dual rate-limitation
INTRODUCTION
Nitrification involves the conversion of ammonium-nitrogen (NH4
+-N) to nitrite-nitrogen (NO2−-N) and nitrate-
nitrogen (NO3−-N). Traditionally, nitrification has been
mathematically treated as one composite biochemical pro-cess, with the assumption that NH4
+-N to NO2−-N oxidation
limits the overall transformation of NH4+-N to NO3
−-N(Grady et al., 1999; Henze et al., 1987). However, whenNH4
+-N and NO2−-N oxidation both limit overall nitrifica-
tion at different stages of the process, the single-step rep-resentation is inadequate. Under such circumstances, theindividual characterization of both NH4
+-N and NO2−-N
oxidation is necessary. An accurate knowledge of the kinet-ics of each step is required to formulate appropriate designand control strategies for achieving desired NH4
+-N andNO2
−-N removal. Numerous biokinetic studies have beenperformed using pure nitrifying cultures, nitrifying enrich-ment consortia and activated sludge (Painter, 1970; Sharmaand Ahlert, 1977). However, significant variation exists inliterature with respect to the reported kinetic estimates dueto the wide array of biokinetic estimation techniques em-ployed. Additionally, the initial conditions imposed onsome biokinetic assays render the resulting kinetic param-eters inappropriate for extrapolation to continuous reactorconfigurations (Volskay and Grady, 1988). Because batchbiokinetic assays are often used to estimate kinetic param-eters for use in continuous reactor modeling, we consideredit imperative to evaluate whether single-step nitrificationmodels are appropriate to describe NH4
+-N oxidation pro-files obtained from batch assays.
The primary objectives of this study were as follows:
1. To quantify, the individual steps of NH4+-N and NO2
−-Noxidation by a nitrifying enrichment culture using anextant respirometic technique.
2. To examine the effect of principal nitrogen species oneach oxidation step.
3. To determine the kinetics of complete NH4+-N to
NO3−-N oxidation using a single-step nitrification
model.4. To evaluate the adequacy of the single-step nitrification
model in depicting complete NH4+-N oxidation by com-Correspondence to:B. F. Smets
© 2000 John Wiley & Sons, Inc.

paring single-step kinetic estimates of NH4+-N to
NO3−-N oxidation with estimates for NH4
+-N to NO2−-N
oxidation.
MATERIALS AND METHODS
Cultivation of a Nitrifying Enrichment Culture
A nitrifying enrichment culture was grown in a 2-L labo-ratory scale sequencing batch reactor (BIOFLO III, NewBrunswick Scientific Co., Edison, NJ). The inoculum forthe nitrifying culture was kindly provided by Dr. Robert M.Cowan (Rutgers University, New Brunswick, NJ). To en-rich for a nitrifying population, an inorganic medium devoidof organic carbon was fed to the SBR. Ammonium sulfate(500 mg-N L−1) was provided as the sole energy source ina buffered solution containing the requisite macro- and mi-cronutrients (Hockenbury and Grady, 1977). Periodic addi-tion of a 1M sodium carbonate (Na2CO3) solution fulfilledboth CO2 fixation and alkalinity requirements. The reactorcontents were thoroughly agitated to ensure adequate mix-ing. The dissolved oxygen (DO) concentration in the reactorwas continuously monitored and maintained in excess of 1mg/L during the fill and react periods with filtered labora-tory air. The reactor was operated in a pH range of 7.5 ± 0.2.A solids retention time (SRT) of 20 days was maintained bydirect wastage of solids from the reactor. The reactor wasoperated at four diurnal cycles of 6 h. Reactor performancewas monitored by periodically measuring reactor and efflu-ent suspended solids and influent and effluent NH4
+-N,NO2
−-N, and NO3−-N concentrations. Upon attainment of
steady state, mixed liquor was periodically withdrawn fromthe SBR and used in respirometric tests.
Extant Respirometric Assay for MonitoringNitrification Activity
Extant assays were performed on mixed liquor sampleswithdrawn from the parent nitrifying SBR towards the endof the react cycle. The biomass samples were harvested bycentrifugation at 4000g for 10 minutes and resuspended innitrogen free mineral medium at pH 7.5. The employedextant respirometric assay was conducted essentially as de-scribed earlier (Ellis et al., 1996). The assay was performedin a 100-mL jacketed glass vessel, maintained at 25°C. Therespirometric vessel was completely filled with mixed li-quor withdrawn from the parent SBR and sealed with theinsertion of a Clark-type polarographic DO electrode (YSIModel 5331, Yellow Springs, OH). The contents of therespirometric vessel were thoroughly mixed using a mag-netic stir-bar. A decrease in the DO level in the vessel dueto substrate oxidation was measured by the DO probe andcontinuously acquired by a personal computer interfaced toa DO monitor (YSI Model 5300, Yellow Springs, OH) by amulti-channel data acquisition device (LabPC+, NationalInstruments, Austin, TX). DO profiles were acquired at a
user-defined frequency (typically 4 Hz) from two respiro-metric vessels operated in tandem.
Reagent Solutions
Substrate stock solutions were freshly prepared prior to eachset of respirometric assays using laboratory grade ammo-nium sulfate ((NH4)2SO4, Fisher Scientific Co., NJ) andsodium nitrite (NaNO2, Sigma Chemical Co., MO). Al-lylthiourea (C4H8N2S, Acros, NJ) and sodium azide (NaN3,Fisher Scientific Co.) were used to selectively inhibitNH4
+-N to NO2−-N oxidation and NO2
−-N to NO3−-N oxi-
dation, respectively.
Nitrogen Analysis
NH4+-N was analyzed using an ammonia gas-sensing elec-
trode (HNU Systems, MA) with a range of operation be-tween 7 × 10−3 to 1.4 × 104 mg NH4
+-N L−1. NO2−-N was
assayed using a modification of the sulfanilic acid-N-naphthylethylenediamine addition method (Eaton et al.,1995) with operational range between 5 × 10−3 to 1 mgNO2
−-N L−1 (Eaton et al., 1995). NO3−-N was estimated
using a nitrate ion-specific electrode (Corning, NY) with anoperational range between 1.4 × 10−2 and 1.4 × 104 mgNO3
−-N L−1.
Convention for Expressing State Variables
To facilitate tracking of all state variables during respiro-metric assays, we introduce an approach to express concen-trations of nitrogen containing species in terms of the ni-trogenous oxygen demand (NOD). The oxygen demand of areduced nitrogen species in a given molecule is computedwith respect to an appropriate oxidized reference nitrogenspecies. When NH4
+-N to NO2−-N oxidation is considered,
NO2−-N is the reference species; consequently NH4
+-N hasa NOD of 3.43 mg/mg N, and NO2
−-N has a NOD of 0mg/mg N. When oxidation to NO3
−-N is considered,NO3
−-N is the reference species; consequently NH4+-N has
an NOD of 4.57 mg/mg N, NO2−-N has an NOD of 1.14
mg/mg N, and NO3−-N has an NOD of 0 mg/mg N.
Determination of Biomass Concentrations
Biomass concentrations were measured as chemical oxygendemand (COD) using commercially available reagents(Hach Chemical Co., Loveland CO). Since the influent wasdevoid of soluble COD, the total COD (tCOD) concentra-tion was taken as a measure of biomass concentration. Thefractions of NH4
+-N and NO2−-N oxidizing biomass were
assumed to be directly proportional to their respective bio-mass yields (see Appendix):
Xns~COD! = tCODfS,ns
fS,ns+fS,nb
3
(1)
CHANDRAN AND SMETS: SINGLE-STEP NITRIFICATION MODELING UNDER DUAL RATE-LIMITATION 397

Xnb~COD! = tCOD
fS,nb
3
fS,ns+fS,nb
3
(2)
whereXns is the concentration of NH4+-N oxidizing biomass
(mg COD L−1), Xnb is the concentration of NO2−-N oxidiz-
ing biomass (mg COD L−1), fS,nsis the biomass yield coef-ficient for NH4
+-N to NO2−-N oxidation (mgXns COD/mg
NH4+-N oxidized), andfS,nbis the biomass yield coefficient
for NO2−-N to NO3
−-N oxidation (mgXnb COD/mg NO2−-
NOD oxidized). The (1⁄3) term in Eqs. (1) and (2) derivesfrom the fact that 1 mg NH4
+-NOD is oxidized to1⁄3 mgNO2
−-NOD.
Selective Inhibition of NH4+-N Oxidation by
Allylthiourea (ATU)
Pre-incubations were performed with ATU (Hooper andTerry, 1973) to determine the minimum concentration atwhich NH4
+-N oxidation was completely inhibited in thepresence and absence of NH4
+-N. Nitrogen specific mea-surements were carried out before and after the pre-incubation to examine NH4
+-N accumulation and NO2−-N
production.
Selective Inhibition of NO2−-N Oxidatioin by
Sodium Azide
Biomass samples were incubated with different NaN3 con-centrations in the presence and absence of NO2
−-N, to de-termine the optimum conditions for selective inhibition ofNO2
−-N oxidation (Aleem and Sewell, 1981; Ginestet et al.,1998). The samples were analyzed before and after incuba-tion to examine NO2
−-N accumulation and NO3−-N produc-
tion.
Data Analysis and Parameter Estimation
The dissolved oxygen (DO) profiles obtained from respiro-metric assays were time-averaged prior to analysis and cor-rected for endogenous oxygen uptake occurring before sub-strate injection and after substrate exhaustion. Three differ-ential equations linking biomass growth, substrateconsumption, and oxygen uptake were formulated and si-multaneously solved using a fourth-order Runge–Kuttamethod. The biokinetic parameters describing the appropri-ate nitrogen oxidation step,mmax andKS, were estimated byminimizing the sum of the squared errors (SSE) with respectto the experimental oxygen uptake profiles using theSOLVERt utility in MS Excelt.
Biokinetic Models for NH4+-N and
NO2−-N Oxidation
The kinetics of NH4+-N to NO2
−-N oxidation were esti-mated by fitting the numerical solutions of Eqs. (3)–(5) to
oxygen uptake profiles obtained in response to NH4+-N
pulses, when NO2−-N oxidation was selectively inhibited.
These experiments are termed isolated NH4+-N oxidation
assays.
dSnh
dt=
~1 + ~0.3fS,ns!!
fS,nsmmax,ns
XnsSnh
KS,ns+ Snh(3)
dOUns
dt=
~1 − fS,ns!
fS,nsmmax,ns
XnsSnh
KS,ns+ Snh(4)
dXns
dt= mmax,ns
XnsSnh
KS,ns+ Snh(5)
where Snh is the NH4+-N concentration (mg NOD L−1),
OUns is the cumulative oxygen uptake due to NH4+-N to
NO2−-N oxidation (mg O2 L−1), mmax,ns is the maximum
specific growth rate of NH4+-N oxidizing biomass (h−1),
andKS,ns is the half-saturation coefficient of NH4+-N oxi-
dizing biomass (mg NOD L−1). (See Fig. A-1 for derivationof stoichiometric coefficients).
The parameters for NO2−-N to NO3
−-N oxidation wereobtained by fitting the numerical solution of Eqs. (6)–(8) toDO responses to NO2
−-N pulses. These experiments aretermed isolated NO2
−-N oxidation assays. In formulatingthe stoichiometric relationship between NO2
−-N consump-tion and oxygen uptake, it was assumed that all the NO2
−-Nprocessed by the nitrite oxidizing bacteria is oxidized toNO3
−-N and that assimilative reduction of NO2−-N does not
occur. The basis for this assumption is that NH4+-N is the
preferred nitrogen source for NO2−-N oxidizing microor-
ganisms.
dSno2
dt= −
mmax,nb
fS,nb
XnbSno2
KS,nb+ Sno2(6)
dOUnb
dt=
~1 − fS,nb!
fS,nbmmax,nb
XnbSno2
KS,nb+ Sno2(7)
dXnb
dt= mmax,nb
XnbSno2
KS,nb+ Sno2(8)
where Sno2 is the NO2−-N concentration (mg NOD L−1),
OUnb is the cumulative oxygen uptake due to NO2−-N to
NO3−-N oxidation (mg O2 L−1), mmax,nb is the maximum
specific growth rate of NO2−-N oxidizing biomass (h−1),
andKS,nb is the half-saturation coefficient of NO2−-N oxi-
dizing biomass (mg NOD L−1). (See Fig. A-2 for derivationof stoichiometric coefficients).
The lumped kinetics of NH4+-N to NO3
−-N oxidationwere obtained by fitting the numerical solution of thesingle-step nitrification model (Eqs. (9)–(11) to DO re-sponses to NH4
+-N pulses in the absence of NO2−-N oxi-
dation inhibition. These experiments are termed completeNH4
+-N oxidation assays.
dSnh
dt= −
~1 + ~0.4fS,L!!
fS,Lmmax,L
XnhSnh
KS,L + Snh(9)
398 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 68, NO. 4, MAY 20, 2000

dOUnh
dt=
~1 − fS,L!
fS,Lmmax,L
XnhSnh
KS,L + Snh(10)
dXnh
dt= mmax,L
XnhSnh
KS,L + Snh(11)
whereXnh is the total nitrifying biomass concentration (mgCOD L−1), OUnh is the cumulative oxygen uptake due tocomplete NH4
+-N oxidation (mg L−1), mmax,L is the lumpedmaximum specific growth rate (h−1), KS,L is the lumpedhalf-saturation coefficient (mg NOD L−1), and fS,L is thelumped biomass yield coefficient (mgXnh NOD/mg NH4
+-NOD oxidized to NO3
−-N).The biomass yield coefficientsfS,ns and fS,L were esti-
mated from the difference between the total cumulativeoxygen uptake during a respirometric assay and the injectedNH4
+-N concentration. It can easily be shown that
fS,ns=~Snho − OUns,f!
~Snho + ~0.3OUns,f!!(12)
fS,L =~Snho − OUnh,f!
~Snho + ~0.4OUns,f!!(13)
where the second term in the denominator is a correctionfactor for assimilated NH4
+-N that does not change oxida-tion state, andSnho is the injected NH4
+-N concentration(mg NOD L−1), OUns,f is the total cumulative oxygen up-take due to NH4
+-N to NO2−-N oxidation (mg O2 L−1), and
OUnh,f is the total cumulative oxygen uptake due to NH4+-N
to NO3−-N oxidation (mg O2 L−1).
From the entire pool of experiments conducted, an aver-agefS,nsvalue of 0.084 mgXnsCOD/mg NH4
+-NOD and anaveragefS,L value of 0.079 mgXnh COD/mg NH4
+-NODwere calculated. Minor adjustments to theSnho values weremade in individual experiments based onOUf andfS,ns,avgorfS,L,avg (Eqs. (14) and (15)):
S8nho = OUns,f
~1 + ~0.3fS,ns,avg!!
~1 − fS,ns,avg!(14)
S8nho = OUnh,f
~1 + ~0.4fS,L,avg!!
~1 − fS,L,avg!(15)
whereS8nho is the adjusted initial NH4+-N concentration (mg
NOD L−1), fS,ns,avgis the averagefS,ns (mg Xns COD/mgNH4
+-NOD oxidized to NO2−-N), andfS,L,avgis the average
fL,ns (mg Xnh COD/mg NH4+-NOD oxidized to NO3
−-N).The biomass yield associated with NO2
−-N to NO3−-N
oxidation is given by
fS,nb=Sno2o− OUnb,f
Sno2o(16)
whereSno2ois the injected NO2−-N concentration (mg NOD
L−1) andOUnb,f is the total cumulative oxygen uptake due toNO2
−-N to NO3−-N oxidation (mg O2 L−1).
fS,nb could not be evaluated from the NO2−-N oxidation
respirograms because the differences between the injected
NO2−-N concentrations and the total cumulative oxygen up-
take were too small to estimate reliably. Therefore, a litera-ture fS,nb value of 0.1 mgXnb COD/NO2
−-NOD oxidizedwas used (Sharma and Ahlert, 1977). Minor corrections toSno2o values were made in individual experiments (Eq.(17)):
S8no2o=OUnb,f
~1 − fS,nb,avg!(17)
where S8no2o is the adjusted initial NO2−-N concentration
(mg NOD L−1) and fS,nb,avg is the averagefS,nb (mg Xnb
COD/mg NO2−-NOD oxidized to NO3
−-N).The maximum specific NH4
+-N and NO2−-N oxidation
activities were expressed as maximum specific substrateconsumption rate (Eqs. (18)–(20).
qmax,ns= mmax,ns
~1 + ~0.3fS,ns!!
fS,ns(18)
qmax,L = mmax,L
~1 + ~0.4fS,L!!
fS,L(19)
qmax,nb=mmax,nb
fS,nb(20)
whereqmax,ns is the maximum specific NH4+-N consump-
tion rate during NH4+-N to NO2
−-N oxidation (mg NOD mgCOD−1 h−1), qmax,L is the maximum specific NH4
+-N con-sumption rate during NH4
+-N to NO3−-N oxidation (mg
NOD mg COD−1 h−1), andqmax,nbis the maximum specificNO2
−-N consumption rate (mg NOD mg COD−1 h−1).Individual parameters were obtained from several months
of experimental testing. Variability in the parameter poolwas minimized by calculating the inner and outer bound-aries for each parameter estimate and discarding the ex-treme outliers from the overall parameter set (Ott, 1993).
Effect of NO2−-N on NH4
+-N to NO2−-N Oxidation
NO2−-N oxidation was completely inhibited by addition of
24 mM NaN3. Subsequently, increasing NO2−-N concentra-
tions in the reactor were attained by repeated NO2−-N in-
jections. The kinetic parameter estimates of NH4+-N to
NO2−-N oxidation obtained at different NO2
−-N concentra-tions were fit to a linear reversible enzymatic inhibitionmodel (Hartmann and Laubenberger, 1968) to determine theextent and nature of inhibition caused by NO2
−-N.
Effect of NH4+-N on NO2
−-N to NO3−-N Oxidation
NH4+-N oxidation was completely inhibited by pre-
incubation with 2 mg ATU L−1 and 2 mg NH4+-N L−1. The
effect of NH4+-N concentrations on NO2
−-N to NO3−-N
oxidation was quantified by correlatingmmax,nbandKS,nbatvarious NH4
+-N concentrations to a linear reversible enzy-matic inhibition model.
CHANDRAN AND SMETS: SINGLE-STEP NITRIFICATION MODELING UNDER DUAL RATE-LIMITATION 399
Dual Limitation of Complete NH4+-N Oxidation
To evaluate the adequacy of the single-step nitrificationmodel under artificially enhanced dual limitation of NH4
+-Nto NO3
−-N oxidation by both NH4+-N to NO2
−-N andNO2
−-N to NO3−-N oxidation, NO2
−-N to NO3−-N oxida-
tion was selectively and partially inhibited by exposing bio-mass samples to 0.24 and 0.48mM NaN3. Injections ofNH4
+-N and NO2−-N performed in duplicate NaN3 supple-
mented reactors allowed the concurrent estimation of ki-netic parameters for NH4
+-N to NO3−-N oxidation and
NO2−-N to NO3
−-N oxidation.
RESULTS
Steady-State Nitrifying SBR Performance
Except for occasional instabilities, the nitrifying SBR con-sistently operated at a target solids retention time (SRT) of20 days and a hydraulic retention time (HRT) of 1.25 days.The steady state reactor and effluent biomass concentrationswere 1003 ± 262 and 51 ± 28 mg COD L−1, respectively (n4 64). The effluent NH4
+-N, NO2−-N, and NO3
−-N con-centrations were 1.3 ± 2.4, 0.14 ± 0.58, and 537 ± 182 mgN L−1, respectively (n 4 90).
Selective Inhibition of NH4+-N Oxidation
by Allylthiourea
The severity of ATU induced inhibition depended on bothNH4
+-N and ATU pre-incubation concentrations. Success-ful inhibition of NH4
+-N oxidation was achieved by incu-bation with 2 mg ATU L−1 and 2 mg NH4
+-N L−1 for 60min. Inhibition was confirmed by the accumulation ofNH4
+-N but not of NO2−-N during the incubation period
and further corroborated by the lack of oxygen uptake inresponse to a subsequent NH4
+-N injection. At 2 mg L−1,ATU did not effect NO2
−-N oxidation (data not shown).
Selective Inhibition of NO2−-N Oxidation by
Sodium Azide
Sodium azide instantaneously inhibited NO2−-N oxidation,
obviating the need for preincubations. NO2−-N to NO3
−-Noxidation was only partially inhibited at 12mM NaN3, whileat higher NaN3 concentrations (48 and 96mM) bothNH4
+-N and NO2−-N oxidation were inhibited (data not
shown). An optimum NaN3 concentration of 24mM enabledselective inhibition of NO2
−-N oxidation, without affectingNH4
+-N oxidation. This optimum concentration is consis-tent with a recent report (Ginestet et al., 1998).
Biokinetics of Nitrification
The kinetics of both NH4+-N to NO2
−-N oxidation andNO2
−-N to NO3−-N oxidation changed gradually over the
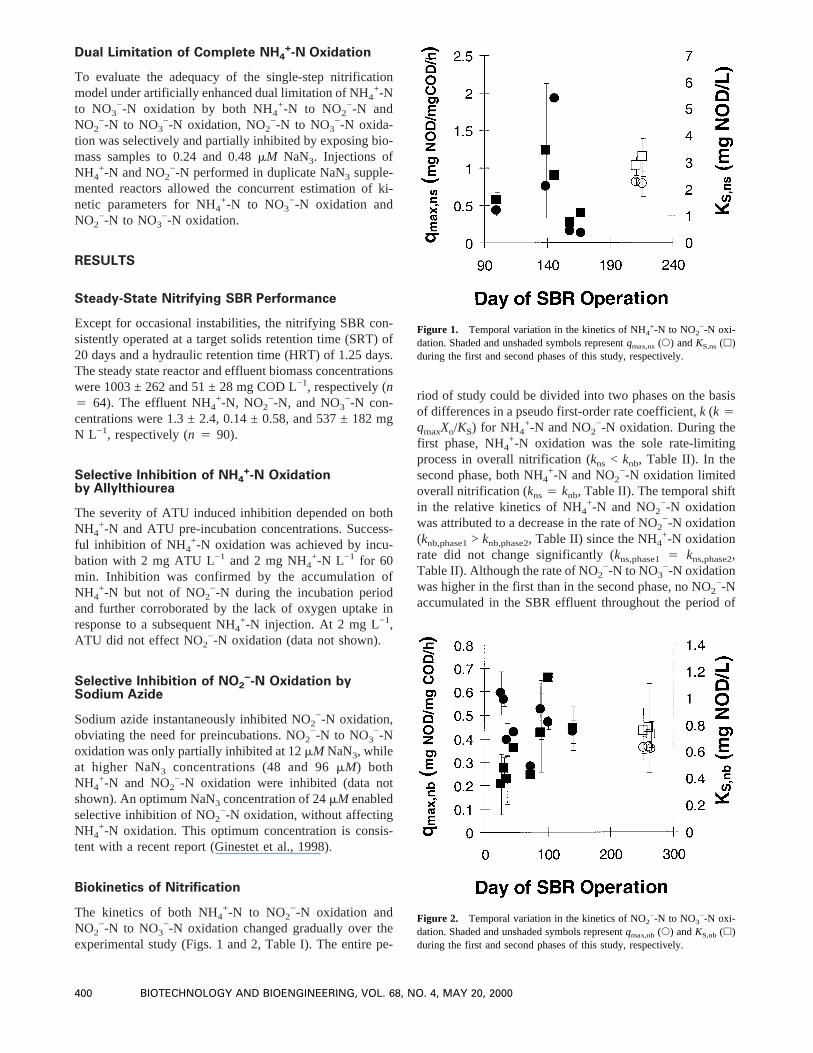
experimental study (Figs. 1 and 2, Table I). The entire pe-
riod of study could be divided into two phases on the basisof differences in a pseudo first-order rate coefficient,k (k 4qmaxXo/KS) for NH4
+-N and NO2−-N oxidation. During the
first phase, NH4+-N oxidation was the sole rate-limiting
process in overall nitrification (kns < knb, Table II). In thesecond phase, both NH4
+-N and NO2−-N oxidation limited
overall nitrification (kns4 knb, Table II). The temporal shiftin the relative kinetics of NH4
+-N and NO2−-N oxidation
was attributed to a decrease in the rate of NO2−-N oxidation
(knb,phase1> knb,phase2, Table II) since the NH4+-N oxidation
rate did not change significantly (kns,phase14 kns,phase2,Table II). Although the rate of NO2
−-N to NO3−-N oxidation
was higher in the first than in the second phase, no NO2−-N
accumulated in the SBR effluent throughout the period of
Figure 1. Temporal variation in the kinetics of NH4+-N to NO2
−-N oxi-dation. Shaded and unshaded symbols representqmax,ns(s) andKS,ns(h)during the first and second phases of this study, respectively.
Figure 2. Temporal variation in the kinetics of NO2−-N to NO3
−-N oxi-dation. Shaded and unshaded symbols representqmax,nb(s) andKS,nb(h)during the first and second phases of this study, respectively.
400 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 68, NO. 4, MAY 20, 2000

this study. Thus, the lower NO2−-N oxidation rate in the
second phase was mainly caused by a change in the kineticparameters and not by a decrease in the NO2
−-N oxidizingbiomass concentration.
The maximum initial substrate concentrations used toquantify NH4
+-N to NO2−-N oxidation and NO2
−-N toNO3
−-N oxidation (∼4 mg NH4+-N L−1 and∼3 mg NO2
−-NL−1 respectively) were much lower than the concentrationsat which selfinhibition of NH4
+-N and NO2−-N oxidation is
significant in activated sludge (Anthonisen, 1974) and bothNH4
+-N and NO2−-N oxidation kinetics were modeled us-
ing the Monod equation for non-inhibitory substrates.
Effect of Dominant Nitrogen Species on NH4+-N
to NO2−-N Oxidation
At NO2−-N concentrations from approximately 2 to 100 mg
NO2−-N L−1 there was no monotonic effect on eitherqmax,ns
or KS,ns(Fig. 3). The lack of inhibition of NH4+-N oxidation
is consistent with the high threshold inhibitory levels (1,200mg NO2
−-N L−1) reported in the literature (Prakasam andLoehr, 1972). The effect of NO3
−-N on NH4+-N to NO2
−-N
oxidation was not determined, assuming acclimation of thenitrifying enrichment culture to high NO3
−-N concentra-tions (537 ± 182 mg NO3
−-N L−1) observed in the SBR.
Effect of Dominant Nitrogen Species on NO2−-N
to NO3−-N Oxidation
Based on a single-coefficient linear inhibition model, theeffect of NH4
+-N on NO2−-N to NO3
−-N oxidation wasdescribed by a noncompetitive inhibition coefficient ofKi,nh
4 62 mg NH4+-N L−1 (Fig. 4). NO2
−-N to NO3−-N oxida-
tion was not inhibited by NO3−-N concentrations up to 250
mg N L−1 (Fig. 5). The absence of any effect of the NO3−-N
concentrations tested on the kinetics of NO2−-N oxidation
was expected due to the high NO3−-N concentrations (537 ±
182 mg NO3−-N L−1) observed in the parent SBR.
Lumped Kinetics of NH4+-N to NO3
−-N Oxidation
During the first phase, there was no statistical differencebetween the pseudo first-order rate coefficient obtainedfrom complete NH4
+-N oxidation assays and that obtained
Table II. Statistical inferences made on kinetic data pool.a
Reaction considered inparameter estimation Assay conducted
Hypothesis onb
qmax (df, a, b) KS (df, a, b) k (df, a, b)
NH4+-N → NO3
−-N fromvs
Complete NH4+-N oxidation (Phase 1) qmax,L 4 qmax,ns
(8, 0.05, 0.13)KS,L > KS,ns
(27, 0.05, −)kL 4 kns
(18, 0.05, 0.43)NH4
+-N → NO2−-N from Isolated NH4
+-N oxidation (Phase 1)NH4
+-N → NO2−-N from
vsIsolated NH4
+-N oxidation (Phase 1) qmax,ns4 qmax,nb
(8, 0.05, 0.1)KS,ns> KS,nb
(9, 0.05, −)kns < knb
(36, 0.05, −)NO2
−-N → NO3−-N from Isolated NO2
−-N oxidation (Phase 1)NH4
+-N → NO2−-N from
vsIsolated NH4
+-N oxidation (Phase 2) qmax,ns> qmax,nb
(4, 0.05, −)KS,ns> KS,nb
(4, 0.05, −)kns 4 knb
(7, 0.05, 0.25)NO2
−-N → NO3−-N from Isolated NO2
−-N oxidation (Phase 2)NH4
+-N → NO2−-N from
vsIsolated NH4
+-N oxidation (Phase 1) qmax,ns,14 qmax,ns,2
(9, 0.05, 0.22)KS,ns,1< KS,ns,2
(8, 0.05, −)kns,1 4 kns,2
(11, 0.05, 0.03)NH4
+-N → NO2−-N from Isolated NH4
+-N oxidation (Phase 2)NO2
−-N → NO3−-N from
vsIsolated NO2
−-N oxidation (Phase 1) qmax,nb,m1> qmax,nb,2
(40, 0.05, −)KS,nb,1< KS,nb,2
(31, 0.05, −)Knb,1 > knb,2
(32, 0.05, −)NO2
−-N → NO3−-N from Isolated NO2
−-N oxidation (Phase 2)
aParameters (p1 and p2) were compared at a confidence level of 95% (a 4 0.05) with the null hypothesis: p14 p2.bk, pseudo first-order rate coefficient4 (qmaxX/KS)df, degrees of freedom;a, probability of incorrectly rejecting the null hypothesis when it is true;b, probability of incorrectly accepting the null hypothesis
when it is false; only applicable when the null hypothesis is not rejected and the parameters being compared are not significantly different. Subscript L refersto lumped kinetic parameters describing NH4
+-N to NO3−-N oxidation. Subscripts 1 and 2 refer to phase 1 and 2 of the study, respectively.
Table I. Biokinetics of nitrification measured throughout the experimental study.
Reaction consideredin parameter estimation Phase Assay
qmax mg NOD(mg COD h)−1
[avg ± SD]
Ks
(mg NOD L−1)[avg ± SD] n
NH4+-N → NO2
−-N 1 Isolated 0.6 ± 0.54 1.63 ± 0.54 9NH4
+-N → NO2−-N 2 Isolated 0.82 ± 0.11 3.05 ± 0.56 5
NO2−-N → NO3
−-N 1 Isolated 0.47 ± 0.11 0.53 ± 0.29 31NO2
−-N → NO3−-N 2 Isolated 0.36 ± 0.04 0.75 ± 0.19 12
NH4+-N → NO3
−-N 1 Complete 0.44 ± 0.15 2.41 ± 0.11 32
CHANDRAN AND SMETS: SINGLE-STEP NITRIFICATION MODELING UNDER DUAL RATE-LIMITATION 401
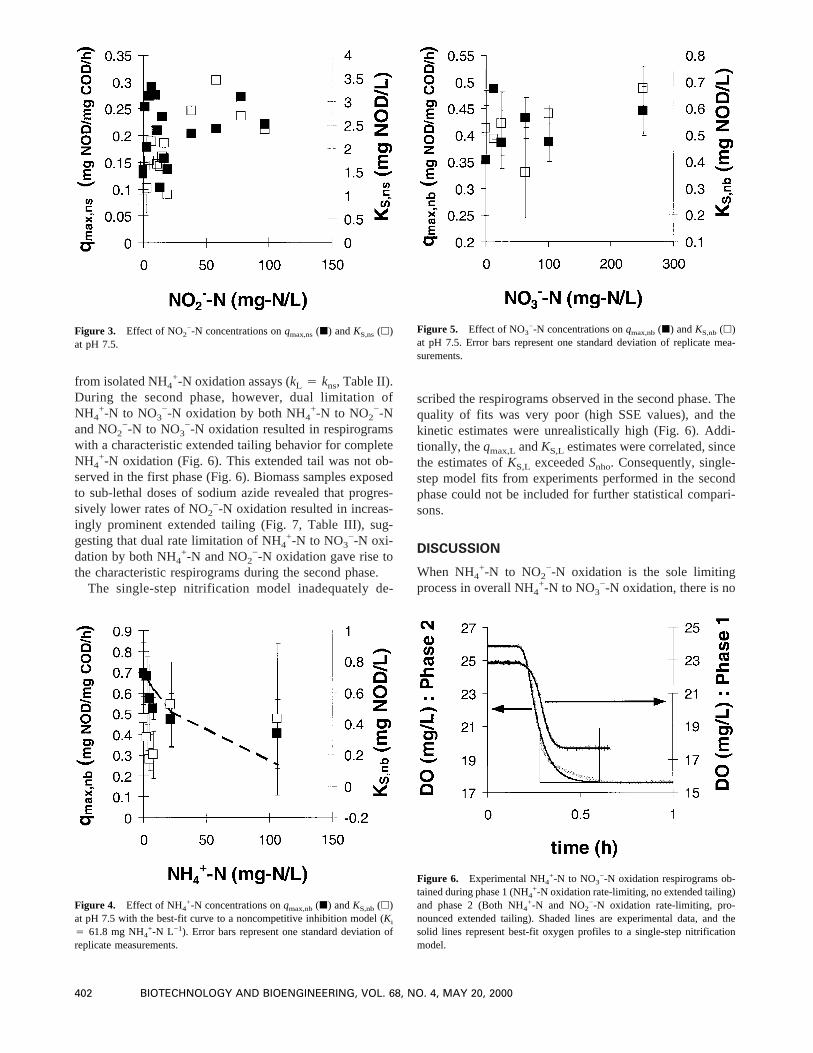
from isolated NH4+-N oxidation assays (kL 4 kns, Table II).
During the second phase, however, dual limitation ofNH4
+-N to NO3−-N oxidation by both NH4
+-N to NO2−-N
and NO2−-N to NO3
−-N oxidation resulted in respirogramswith a characteristic extended tailing behavior for completeNH4
+-N oxidation (Fig. 6). This extended tail was not ob-served in the first phase (Fig. 6). Biomass samples exposedto sub-lethal doses of sodium azide revealed that progres-sively lower rates of NO2
−-N oxidation resulted in increas-ingly prominent extended tailing (Fig. 7, Table III), sug-gesting that dual rate limitation of NH4
+-N to NO3−-N oxi-
dation by both NH4+-N and NO2
−-N oxidation gave rise tothe characteristic respirograms during the second phase.
The single-step nitrification model inadequately de-
scribed the respirograms observed in the second phase. Thequality of fits was very poor (high SSE values), and thekinetic estimates were unrealistically high (Fig. 6). Addi-tionally, theqmax,L andKS,L estimates were correlated, sincethe estimates ofKS,L exceededSnho. Consequently, single-step model fits from experiments performed in the secondphase could not be included for further statistical compari-sons.
DISCUSSION
When NH4+-N to NO2
−-N oxidation is the sole limitingprocess in overall NH4
+-N to NO3−-N oxidation, there is no
Figure 3. Effect of NO2−-N concentrations onqmax,ns(j) andKS,ns(h)
at pH 7.5.
Figure 4. Effect of NH4+-N concentrations onqmax,nb(j) andKS,nb(h)
at pH 7.5 with the best-fit curve to a noncompetitive inhibition model (Ki
4 61.8 mg NH4+-N L−1). Error bars represent one standard deviation of
replicate measurements.
Figure 5. Effect of NO3−-N concentrations onqmax,nb(j) andKS,nb(h)
at pH 7.5. Error bars represent one standard deviation of replicate mea-surements.
Figure 6. Experimental NH4+-N to NO3
−-N oxidation respirograms ob-tained during phase 1 (NH4
+-N oxidation rate-limiting, no extended tailing)and phase 2 (Both NH4
+-N and NO2−-N oxidation rate-limiting, pro-
nounced extended tailing). Shaded lines are experimental data, and thesolid lines represent best-fit oxygen profiles to a single-step nitrificationmodel.
402 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 68, NO. 4, MAY 20, 2000

transient build-up of intermediates and NH4+-N to NO3
−-Noxidation can be described adequately using a single-stepnitrification model (Table II). However, when NH4
+-N andNO2
−-N oxidation both limit the overall oxidation ofNH4
+-N to NO3−-N, a single-step model is not applicable.
When the rate of NO2−-N to NO3
−-N oxidation is slowerthan the rate of NH4
+-N to NO2−-N oxidation, NO2
−-N per-sists even after NH4
+-N is exhausted. Since the single-stepmodel is also a single-factor model, the oxidation of thetransiently accumulated NO2
−-N is erroneously modeled asNH4
+-N oxidation, leading to incorrect lumped kinetic pa-rameter estimates. Therefore, if single-step models are to beused for the interpretation and parameter estimation fromexperimental data, it is important to confirm that NH4
+-N toNO2
−-N oxidation is truly the sole rate-limiting process. Inthe scenario that multiple biochemical processes limit over-all nitrification, alternative models must be considered.
Although NO2−-N accumulation during nitrification has
been recognized in natural habitats, full-scale engineeredsystems, and laboratory-scale reactors (Anthonisen et al.,1976; Knowles et al., 1965), overall nitrification is usually
limited by NH4+-N to NO2
−-N oxidation (Benefield andRandall, 1980; Poduska and Andrews, 1975). Transient ac-cumulation of NO2
−-N and the extended tailing pattern has,however, been observed in batch biokinetic estimation as-says (Gee et al., 1990b; Mauret et al., 1996; Ossenbruggenet al., 1996). Generally, pronounced extended tailing incomplete NH4
+-N oxidation respirograms is associated witha high initial NH4
+-N concentration. For instance, Mauret etal. (1996) observed the extended tail in batch assays with anSnho of 90 mg-N L−1 but not with anSnho of 35 mg-N L−1
(Mauret et al., 1996). This observation suggested possibleinhibition of NO2
−-N oxidation by high initial NH4+-N con-
centrations. Extended tailing pattern has, however, alsobeen observed withSnho as low as 6.5 mg-N L−1 (Ossen-bruggen et al., 1996) which is toward the lower end ofinhibitory NH4
+-N concentrations reported in literature (An-thonisen, 1974). In our study, inhibition of NO2
−-N oxida-tion by NH4
+-N can be ruled out due to the lowSnhoapplied(Snho < 5 mg NH4
+-N L−1) and the experimentally deter-mined inhibition coefficient (KI,nh 4 62 mg NH4
+-N L−1).As a result, the dual limitation and the extended tailingpattern were attributed to slow NO2
−-N oxidation kinetics,which was consistent with the kinetic parameters estimatedfrom the isolated assays. By explicitly measuring the kinet-ics of the individual steps as well as the overall process ofnitrification, we were able to demonstrate unambiguouslythat NO2
−-N to NO3−-N oxidation may be rate limiting dur-
ing a part of the batch nitrification assay. Under such con-ditions, we showed that the mathematical treatment of com-plete batch NH4
+-N oxidation as a single step is inappro-priate.
Several studies have been devoted to the explicit deter-mination of NH4
+-N and NO2−-N oxidation kinetics using
mixed nitrifying cultures. These studies involve either directsubstrate measurements (Gee et al., 1990b; Knowles et al.,1965; Mauret et al., 1996), the measurement of a surrogateanalyte such as oxygen (Brouwer et al., 1998), or a combi-nation of both (Ossenbruggen et al., 1996; Ossenbruggen etal., 1991). An explicit respirometric determination ofNH4
+-N and NO2−-N oxidation kinetics by a mixed nitrify-
ing culture requires the uncoupling of the NH4+-N oxidizing
and NO2−-N oxidizing biomass fractions. Both kinetic and
metabolic uncoupling is possible. In kinetic uncoupling,NH4
+-N to NO2−-N oxidation is quantified at high back-
ground NO2−-N oxidation rates (Chudoba et al., 1985); in
metabolic uncoupling, one of the two nitrification steps isselectively inhibited (Ginestet et al., 1998; Surmacz-Gorskaet al., 1996). Respirometry and metabolic uncoupling wererecently used to measure rates of NH4
+-N to NO2−-N oxi-
dation by activated sludge, although a direct estimation ofmaximum specific activity and affinity coefficient was notattempted (Ginestet et al., 1998; Surmacz-Gorska, et al.,1996). In our protocol, the acquisition of complete respiro-grams allowed the estimation of the kinetic parameters(qmax andKS) for both NH4
+-N and NO2−-N oxidation. In
sum, we have presented an improved and simple respirom-
Table III. Kinetics of NO2−-N to NO3
−-N oxidation at various NaN3concentrations.
NaN3 (mM)
qmax,nb
mg NOD(mg COD h)−1
avg ± SDKS,nb (mg NOD L−1)
avg ± SD n
0 0.36 ± 0.01 0.73 ± 0.1 30.24 0.25 ± 0.02 1.31 ± 0.08 20.48 0.15 ± 0.00 0.67 ± 0.08 2
Figure 7. NH4+-N to NO3
−-N oxidation profiles obtained during thesecond phase of this study by exposing biomass samples to NaN3: 0 mM(A), 0.24 mM (B), and 0.48mM NaN3 (C). Kinetics of NO2
−-N oxidationwere obtained from NO2
−-N injections performed concurrently (Table III).The rectangles depict the extended tailing in the respirograms due to in-creasingly lower NO2
−-N oxidation rates.
CHANDRAN AND SMETS: SINGLE-STEP NITRIFICATION MODELING UNDER DUAL RATE-LIMITATION 403

etry-based assay that permits the complete kinetic charac-terization of each nitrification step.
Interaction Between Nitrogen Species Involvedin Nitrification
Each nitrification step is reportedly more adversely affectedby the substrate of the other step than by its own substrate(Anthonisen et al., 1976; Poduska and Andrews, 1975). Inthis study, NH4
+-N was observed to be more inhibitory toNO2
−-N to NO3−-N oxidation than NO2
−-N was to NH4+-N
to NO2−-N oxidation, consistent with literature reports (An-
thonisen, 1974; Prakasam and Loehr, 1972). NO2−-N to
NO3−-N oxidation was not affected by NO3
−-N up to themaximum concentration tested (250 mg NO3
−-N L−1), inagreement with a reported high tolerance to NO3
−-N (Boonand Laudelot, 1962).
Interactions Between NH4+-N and NO2
−-NOxidizing Microorganisms
The implicit assumption in quantifying each nitrificationstep via metabolic uncoupling, is that no direct interactionexists between the NH4
+-N and NO2−-N oxidizing popula-
tions. Previously, however, it was reported that NO2−-N
oxidation is greatly enhanced in the presence of a robustNH4
+-N oxidizing population while NH4+-N oxidation is
independent of the NO2−-N oxidizing population (Gee et al.,
1990a). Our results do not support this claim. The isolatedkinetics of NO2
−-N oxidation obtained with and withoutcomplete inhibition of NH4
+-N oxidizing activity were notsignificantly different (qmax,nb,ATU+4 qmax,nb,ATU−, n 4 4,a 4 0.05,b 4 0.12;KS,nb,ATU+4 KS,nb,ATU−, n 4 4, a 40.05,b 4 0.08).
CONCLUSIONS
Batch respirometry is an excellent tool for the rapid estima-tion of biokinetic parameters with minimal analytical effort,provided appropriate mathematical models exist to interpretthe obtained respirograms. We derived mathematical ex-pressions that couple the sequential oxidation of NH4
+-N toNO2
−-N and NO2−-N to NO3
−-N to oxygen uptake in batchassays. This mathematical description was facilitated by ex-pressing the concentrations of nitrogen species in terms ofoxygen demand equivalents. These models were success-fully used to estimate kinetics of individual nitrificationsteps by selective metabolic inhibition. A conventionalsingle-step, single-factor model that considers completeNH4
+-N to NO3−-N oxidation as one biochemical process
was also derived in terms of oxygen uptake. Such nitrifica-tion models are appropriate only as long as NH4
+-N toNO2
−-N oxidation is the sole rate-limiting step in NH4+-N
to NO3−-N oxidation. During a long-term study with a ni-
trifying enrichment consortium, we observed shifts in therelative dynamics of NH4
+-N to NO2−-N oxidation and
NO2−-N to NO3
−-N oxidation. These shifts resulted in dual
rate limitation by both processes in batch NH4+-N oxidation
assays. Under such conditions, quantifying NH4+-N to
NO3−-N oxidation using a single-step model resulted in er-
roneous kinetic estimates. We also observed that the kinet-ics of NH4
+-N to NO2−-N oxidation and NO2
−-N to NO3−-N
oxidation were minimally impacted by the presence of ni-trogen species that did not constitute the direct energy sub-strate. Additionally, there was no evidence of any interac-tion between the NH4
+-N and NO2−-N oxidizing microor-
ganisms.Given that respirograms obtained during NH4
+-N toNO3
−-N oxidation contain information pertaining to bothNH4
+-N and NO2−-N oxidation, it may be possible to de-
termine kinetic parameter estimates describing each nitrifi-cation step from this respirogram. In a subsequent manu-script, we will present an appropriate experimental designstrategy and a mathematical model that facilitate the quan-tification of both NH4
+-N and NO2−-N oxidation from com-
plete NH4+-N to NO3
−-N oxidation profiles.
APPENDIX
Of the electrons arising from the oxidation of NH4+-N and
NO2−-N a fraction is channeled via a terminal cytochrome
oxidase to oxygen. The remaining electrons travel via elec-tron carriers such as NAD(P)H to effect CO2 fixation (Figs.A-1 and A-2). The fraction of the total electrons availablefrom reduced nitrogen species that is used for CO2 fixationis termedfs, and the fraction that is used for oxygen reduc-tion and energy generation is termedfe (Grady et al., 1999).The sum offs andfe equals 1 (Grady et al., 1999).fs is alsoequal to the biomass yield coefficient expressed asX CODsynthesized/NOD oxidized.
NH4+-N is the preferred assimilative nitrogen source for
both NH4+-N and NO2
−-N oxidizing bacteria. Therefore inthe case of NH4
+-N oxidation, only a fraction of the total
Figure A-1. Stoichiometric links between nitrogen removal, oxygen con-sumption, and biomass growth during NH4
+-N to NO2−-N oxidation.
404 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 68, NO. 4, MAY 20, 2000

NH4+-N is oxidized and the remainder is incorporated into
biomass without any change in oxidation state. Consideringone mg NH4
+-NOD processed, the fraction of NH4+-N oxi-
dized is:
frSnh,oxid =
1
fS,ns
1
fS,ns+ iNOD/COD
, (A-1)
where frSnh,oxid is the fraction of total NH4+-N processed
that is oxidized,fS,nsis the fraction of total electrons avail-able from NH4
+-N to NO2−-N oxidation that is used for CO2
fixation, andiNOD/COD is nitrogen content of biomass (mgNOD/X COD).
For NH4+-N to NO2
−-N oxidation, iNOD/COD equals 0.3mg NOD/mgXns COD and Eq. (A-1) can be simplified toyield
frSnh,oxid =1
1 + 0.3fS,ns. (A-2)
For NH4+-N to NO3
−-N oxidation, iNOD/COD equals 0.4mg NOD/mgXnh COD andfrSnh,oxid is given by
frSnh,oxid =1
1 + 0.4fS,L, (A-3)
where fS,L is the fraction of total electrons available fromNH4
+-N to NO3−-N oxidation that is used for CO2 fixation.
In NO2−-N oxidation, all the NO2
−-N processed is oxi-dized since NH4
+-N acts as the nitrogen source. Therefore,frSno2oxid 4 1.
Calculation of NH4+-N and NO2
−-N OxidizingBiomass Concentrations
To derive expressions that relate the concentrations of bio-mass fractions to their yield coefficients, a basis of 1 mg
NH4+-NOD oxidized was considered. The biomass synthe-
sized as a result of this oxidation is
DXns 4 fS,nsSnh,oxid 4 fnh,oxid 4 fS,ns. (A-4)
The oxidation of 1 mg NH4+-N results in the formation of
1 mg NO2−-N. Because electrons are lost during this oxi-
dation, 3.43 mg NH4+-NOD is oxidized to 1.14 mg NO2
−-NOD. Thus the oxidation of 1 mg NH4
+-NOD results in (1⁄3)mg NO2
−-NOD, all of which is oxidized to NO3−-N.
DXnb 4 fS,nbSno2,oxid 4 1⁄3 fS,nb, (A-5)
wherefS,nb is the fraction of total electrons available fromNO2
−-N to NO3−-N oxidation that is used for CO2 fixation.
Finally, in a mixed nitrifying consortium, the NH4+-N
and NO2−-N oxidizing biomass fractions are given by the
following equations:
frXns=
DXns
DXns + DXnb=
fS,ns
fS,ns+fS,nb
3
, (A-6)
frXnb=
DXnb
DXns + DXnb=
fS,nb
3
fS,ns+fS,nb
3
. (A-7)
NOMENCLATURE
Snh NH4+-N concentration (mg NOD L−1)
Xns NH4+-N oxidizing biomass concentration (mg COD L−1)
mmax,ns maximum specific growth rate associated with NH4+-N to
NO2−-N oxidation (h−1)
KS,ns half-saturation coefficient for NH4+-N to NO2
−-N oxidation(mg NOD L−1)
qmax,ns maximum specific NH4+-N consumption rate (mg NOD mg
COD−1 h−1)fS,ns biomass yield coefficient associated with NH4
+-N toNO2
−-N oxidation (mgXns COD produced/mg NH4+-NOD
oxidized)OUns oxygen uptake accompanying NH4
+-N to NO2−-N oxidation
(mg O2 L−1)Sno2 NO2
−-N concentration (mg NOD L−1)Xnb NO2
−-N oxidizing biomass concentration (mg COD L−1)mmax,nb maximum specific growth rate associated with NO2
−-N toNO3
−-N oxidation (h−1)KS,nb half-saturation coefficient for NO2
−-N to NO3−-N oxidation
(mg NOD L−1)qmax,nb maximum specific NO2
−-N consumption rate (mg NOD mgCOD−1 h−1)
fS,nb biomass yield coefficient associated with NO2−-N to
NO3−-N oxidation (mgXnb NOD produced/mg NO2
−-NODoxidized)
OUnb oxygen uptake accompanying NO2−-N to NO3
−-N oxidation(mg O2 L−1)
Xnh total nitrifying biomass concentration (mg COD L−1)mmax,L lumped maximum specific growth rate (h−1)KS,L lumped half-saturatioin coefficient (mg NOD L−1)qmax,L lumped maximum specific NH4
+-N consumption rate (mgNOD mg COD−1 h−1)
fS,L lumped biomass yield coefficient associated with NH4+-N to
NO3−-N oxidation (mgXnh NOD produced/mg NH4
+-NODoxidized)
Figure A-2. Stoichiometric links between nitrogen removal, oxygen con-sumption, and biomass growth during NO2
−-N to NO3−-N oxidation.
CHANDRAN AND SMETS: SINGLE-STEP NITRIFICATION MODELING UNDER DUAL RATE-LIMITATION 405

OUnh oxygen uptake accompanying oxidation of NH4+-N to
NO3−-N (mg O2 L−1)
iNOD/COD nitrogen content of biomass (mg NOD/mgX COD)Ki inhibition coefficient (mg L−1)k pseudo first-order rate coefficient (h−1)SSE sum of squared errors (mg2 L−2)a probability of incorrectly rejecting the null hypothesis in
statistical comparison tests when it is trueb probability of incorrectly accepting the null hypothesis in
statistical comparison tests when it is false, only applicablewhen the null hypothesis is not rejected and the parametersbeing compared are not significantly different
References
Aleem MIH, Sewell DL. 1981. Mechanism of nitrite oxidation and oxido-reductase systems inNitrobacter agilis.Curr Microbiol 5:267–272.
Anthonisen AC. 1974. The effects of free ammonia and free nitrous acid onthe nitrification process. Ph.D. (Eng.) Thesis. Cornell University,Ithaca, NY.
Anthonisen AC, Loehr RC, Prakasam TBS, Srinath EG. 1976. Inhibition ofnitrification by ammonia and nitrous acid. J Water Pollut Cont Fed48:835–852.
Benefield LD, Randall CW. 1980. Activated sludge and its process modi-fication. Englewood Cliffs, NJ: Prentice-Hall Inc.
Boon B, Laudelot H. 1962. Kinetics of nitrite oxidation byNitrobacterwinogradsky.Biochem J 85:440–447.
Brouwer H, Klapwijk A, Keesman KJ. 1998. Identification of activatedsludge and wastewater characteristics using respirometric batch ex-periments. Water Res 32:1240–1254.
Chudoba J, Cech JS, Chudoba P. 1985. The effects of aeration tank con-figuration on nitrification kinetics. J Water Pollut Cont Fed 57:1078–1083.
Eaton AD, Clesceri LS, Greenberg AE, editors. 1995. Standard Methodsfor the Examination of Water and Wastewater. Washington, D.C.:APHA, AWWA, and WEF.
Ellis TG, Barbeau DS, Smets BF, Grady CPL Jr. 1996. Respirometrictechnique for determination of extant kinetic parameters describingbiodegradation. Water Environ Res 68:917–926.
Gee CS, Pfeffer JT, Suidan MT. 1990a.Nitrosomonasand Nitrobacterinteractions in biological nitrification. J Environ Eng 116:4–17.
Gee CS, Suidan MT, Pfeffer JT. 1990b. Modeling of nitrification undersubstrate-inhibiting conditions. J Environ Eng 116:18–31.
Ginestet P, Audic JM, Urbain V, Block JC. 1998. Estimation of nitrifyingbacterial activities by measuring oxygen uptake in the presence of themetabolic inhibitors allylthiourea and azide. Appl Environ Microbiol64:2266–2268.
Grady CPL Jr, Daigger GT, Lim HC. 1999. Biological wastewater treat-ment. New York, Marcel Dekker.
Hartmann L, Laubenberger G. 1968. Toxicity measurements in activatedsludge. J San Eng Proc ASCE 94:247.
Henze M, Grady CPL Jr, Gujer W, Marais GVR, Matsuo T. 1987. Ageneral model for single-sludge wastewater treatment systems. WaterRes 5:505–515.
Hockenbury MR, Grady CPL Jr. 1977. Inhibition of nitrification: Effects ofselected organic compounds. J Water Pollut Cont Fed 49:768–777.
Hooper AB, Terry KR. 1973. Specific inhibitors of ammonia oxidation inNitrosomonas.J Bacteriol 115:480–485.
Knowles G, Downing AL, Barrett MJ. 1965. Determination of kineticconstants for nitrifying bacteria in mixed culture, with the aid of anelectronic computer. J Gen Microbiol 38:263–278.
Mauret M, Paul E, Puech-Costes E, Maurette MT, Baptiste P. 1996. Ap-plication of experimental research methodology to the study of nitri-fication in mixed culture. Water Sci Technol 34:245–252.
Ossenbruggen PJ, Spanjers HA, Aspegren H, Klapwijk A. 1991. Designingexperiments for model identification of the nitrification process. WaterSci Technol 24:9–16.
Ossenbruggen PJ, Spanjers H, Klapwijk K. 1996. Assessment of a two-stepnitrification model for activated sludge. Water Res 30:939–953.
Ott RL. 1993. An introduction to statistical methods and data analysis.Belmont, CA: Duxbury Press.
Painter HA. 1970. A review of literature on inorganic nitrogen metabolismin microorganisms. Water Res 4:393–450.
Poduska RA, Andrews JF. 1975. Dynamics of nitrification in the activatedsludge process. J Water Pollut Cont Fed 47:2599–2619.
Prakasam TBS, Loehr RC. 1972. Microbial nitrification and denitrificationin concentrated wastes. Water Res 6:497–509.
Sharma B, Ahlert RC. 1977. Nitirification and nitrogen removal. WaterRes 11:897–925.
Surmacz-Gorska J, Gernaey K, Demuynck C, Vanrolleghem P, VerstraeteW. 1996. Nitrification monitoring in activated sludge by oxygen up-take rate (OUR) measurements. Water Res 30:1228–1236.
Volskay VT Jr, Grady CPL Jr. 1988. Toxicity of selected RCRA com-pounds to activated sludge microorganisms. J Water Pollut Cont Fed60:1850–1856.
406 BIOTECHNOLOGY AND BIOENGINEERING, VOL. 68, NO. 4, MAY 20, 2000
Related Documents











