Original article Siluro-Devonian miospore biostratigraphy of the Urubu River area, western Amazon Basin, northern Brazil Biostratigraphie des miospores du siluro–de ´vonien dans la re ´gion de la rivie `re Urubu, ouest du Bassin Amazonien, nord du Bre ´sil Philippe Steemans a, * , Claudia Rubinstein b , José Henrique G. de Melo c a NFSR Research Associate, Paléobotanique, Paléopalynologie, Micropaléontologie (PPM), University of Liège, Bâtiment B-18, Parking 40, allée du 6-Août, 4000 Liège-1, Belgium b CONICET, Unidad de Paleopalinología, IANIGLA, CRICYT, C.C. 131, 5500 Mendoza, Argentina c PETROBRAS/CENPES/PDEXP/BPA, Cidade Universitaria, Q.7, Ilha do Fundão, 21941-598 Rio de Janeiro, Brazil Received 5 July 2006; accepted 27 June 2007 Available online 4 March 2008 Abstract Well-preserved miospores have been observed in the Urubu River area from the western part of the Amazon Basin, northern Brazil, in rocks belonging to the uppermost Pitinga and Manacapuru Formations. A previous palynological work based on chitinozoans has dated these strata as Late Ludlow, Early Pridoli and Early Lochkovian. Although samples are relatively poor in miospores, the present study reveals the most diversified Silurian–Earliest Lochkovian palynoflora ever reported from South America, an interval where miospore information is still very rare. Sixty-four species have been identified, of which two are new (Artemopyra urubuense and Retusotriletes amazonensis). One single specimen attributed to the genus Grandispora, identical to unpublished forms identified by one of us (P.S.) in Lochkovian sections of Saudi Arabia, has been found in a Lochkovian sample from the Urubu River. These constitute the oldest geological record of Grandispora. Among the abundant acritarchs, specimens of the genus Schizocystia have been observed for the first time in pre-Devonian rocks. Although the Late Silurian–Early Lochkovian miospore biostratigraphy is still in its infancy in South America, it seems that Amazon Basin assemblages do not show significant differences in composition in comparison to coeval palynofloras from other areas of northwestern Gondwana. This is surprising since the miospore localities under consideration are widely distributed over high to low paleolatitudes, i.e. from cool temperate to dry subtropical belts. Conversely, Siluro-Devonian Gondwanan assemblages differ considerably from the contemporary palynofloras of the Old Red Sandstone Continent and adjacent areas, thus contradicting recent phytogeographic interpretations based on megafloras. # 2008 Elsevier Masson SAS. All rights reserved. Résumé Des miospores bien préservées ont été observées dans la région de la rivière Urubu, dans la partie ouest du Bassin d’Amazonie, au nord du Brésil, dans des couches appartenant à la partie supérieure de la Formation de Pitinga et à la Formation de Manacapuru. Une étude palynologique précédente, basée sur les chitinozoaires, a permis de dater les échantillons palynologiques comme étant respectivement d’âge Ludlovien supérieur, Pridoli inférieur et Lochkovien inférieur. Bien que les échantillons soient relativement pauvres en spécimens de miospores, la présente étude porte sur les assemblages les plus diversifiés de la limite siluro-dévonienne observés jusqu’à présent en Amérique du sud où les informations sur les miospores de ces niveaux stratigraphiques sont encore rares. Soixante-quatre espèces ont été identifiées, parmi lesquelles deux nouvelles espèces sont décrites (Artemopyra urubuense et Retusotriletes amazonensis). Un spécimen attribué au genre Grandispora, identique à d’autres spécimens non publiés et identifiés en Arabie Saoudite par l’un de nous (P.S.), a été découvert dans un des échantillons lochkoviens de la rivière Urubu. Ils constituent les plus anciennes observations géologiques de spécimens de Grandispora. Parmi les abondants acritarches, des spécimens du genre Schizocystia ont été observés pour la première fois dans des couches plus anciennes que le Dévonien. http://france.elsevier.com/direct/GEOBIO Disponible en ligne sur www.sciencedirect.com Geobios 41 (2008) 263–282 * Corresponding author. E-mail address: [email protected] (P. Steemans). 0016-6995/$ – see front matter # 2008 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.geobios.2007.06.003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

http://france.elsevier.com/direct/GEOBIO
Disponible en ligne sur www.sciencedirect.com
Geobios 41 (2008) 263–282
Original article
Siluro-Devonian miospore biostratigraphy of the Urubu River area,western Amazon Basin, northern Brazil
Biostratigraphie des miospores du siluro–devonien dans la region
de la riviere Urubu, ouest du Bassin Amazonien, nord du Bresil
Philippe Steemans a,*, Claudia Rubinstein b, José Henrique G. de Melo c
a NFSR Research Associate, Paléobotanique, Paléopalynologie, Micropaléontologie (PPM), University of Liège,Bâtiment B-18, Parking 40, allée du 6-Août, 4000 Liège-1, Belgium
b CONICET, Unidad de Paleopalinología, IANIGLA, CRICYT, C.C. 131, 5500 Mendoza, Argentinac PETROBRAS/CENPES/PDEXP/BPA, Cidade Universitaria, Q.7, Ilha do Fundão, 21941-598 Rio de Janeiro, Brazil
Received 5 July 2006; accepted 27 June 2007
Available online 4 March 2008
Abstract
Well-preserved miospores have been observed in the Urubu River area from the western part of the Amazon Basin, northern Brazil, in rocksbelonging to the uppermost Pitinga and Manacapuru Formations. A previous palynological work based on chitinozoans has dated these strata asLate Ludlow, Early Pridoli and Early Lochkovian. Although samples are relatively poor in miospores, the present study reveals the most diversifiedSilurian–Earliest Lochkovian palynoflora ever reported from South America, an interval where miospore information is still very rare. Sixty-fourspecies have been identified, of which two are new (Artemopyra urubuense and Retusotriletes amazonensis). One single specimen attributed to thegenus Grandispora, identical to unpublished forms identified by one of us (P.S.) in Lochkovian sections of Saudi Arabia, has been found in aLochkovian sample from the Urubu River. These constitute the oldest geological record of Grandispora. Among the abundant acritarchs,specimens of the genus Schizocystia have been observed for the first time in pre-Devonian rocks.
Although the Late Silurian–Early Lochkovian miospore biostratigraphy is still in its infancy in South America, it seems that Amazon Basinassemblages do not show significant differences in composition in comparison to coeval palynofloras from other areas of northwestern Gondwana.This is surprising since the miospore localities under consideration are widely distributed over high to low paleolatitudes, i.e. from cool temperateto dry subtropical belts. Conversely, Siluro-Devonian Gondwanan assemblages differ considerably from the contemporary palynofloras of the OldRed Sandstone Continent and adjacent areas, thus contradicting recent phytogeographic interpretations based on megafloras.# 2008 Elsevier Masson SAS. All rights reserved.
Résumé
Des miospores bien préservées ont été observées dans la région de la rivière Urubu, dans la partie ouest du Bassin d’Amazonie, au nord duBrésil, dans des couches appartenant à la partie supérieure de la Formation de Pitinga et à la Formation de Manacapuru. Une étude palynologiqueprécédente, basée sur les chitinozoaires, a permis de dater les échantillons palynologiques comme étant respectivement d’âge Ludlovien supérieur,Pridoli inférieur et Lochkovien inférieur. Bien que les échantillons soient relativement pauvres en spécimens de miospores, la présente étude portesur les assemblages les plus diversifiés de la limite siluro-dévonienne observés jusqu’à présent en Amérique du sud où les informations sur lesmiospores de ces niveaux stratigraphiques sont encore rares. Soixante-quatre espèces ont été identifiées, parmi lesquelles deux nouvelles espècessont décrites (Artemopyra urubuense et Retusotriletes amazonensis). Un spécimen attribué au genre Grandispora, identique à d’autres spécimensnon publiés et identifiés en Arabie Saoudite par l’un de nous (P.S.), a été découvert dans un des échantillons lochkoviens de la rivière Urubu. Ilsconstituent les plus anciennes observations géologiques de spécimens de Grandispora. Parmi les abondants acritarches, des spécimens du genreSchizocystia ont été observés pour la première fois dans des couches plus anciennes que le Dévonien.
* Corresponding author.E-mail address: [email protected] (P. Steemans).
0016-6995/$ – see front matter # 2008 Elsevier Masson SAS. All rights reserved.doi:10.1016/j.geobios.2007.06.003

P. Steemans et al. / Geobios 41 (2008) 263–282264
Quoique les analyses palynologiques biostratigraphiques basées sur les miospores à la limite siluro-dévonienne soient encore peudéveloppées en Amérique du Sud, il semblerait que la composition des assemblages en miospores en provenance d’Amazonie ne montreguère de différences significatives par rapport à ceux qui lui sont contemporains dans d’autres régions du nord-ouest gondwanien. Cela pourraitparaître surprenant puisque les localités contenant des miospores sont réparties depuis des régions allant de hautes à basses latitudes, depuis desclimats tempérés froids à des climats secs subtropicaux. Par ailleurs, les assemblages du Gondwana montrent de nombreuses différences avec lesassemblages du Continent des Vieux Grès Rouges et des ses régions limitrophes, contredisant certaines hypothèses phytogéographiques baséessur la mégaflore.# 2008 Elsevier Masson SAS. All rights reserved.
Resumen
Miosporas bien preservadas han sido registradas en rocas de la parte superior de la Formación Pitinga y en la Formación Manacapuru, en laregión del Río Urubu, situada en la parte oeste de la Cuenca del Amazonas, norte de Brasil. Una contribución palinológica previa, basada en elestudio de quitinozoos, permitió asignar los estratos portadores al Ludloviano tardío, Pridoliano temprano y Lochkoviano temprano. A pesar de quelas muestras son relativamente pobres en número de especímenes, el presente trabajo muestra la palinoflora del Silúrico-Lochkoviano tempranomás diversa registrada hasta el presente en América del Sur, continente donde la información sobre mioesporas de este intervalo estratigráfico esaún escasa. Se han identificado sesenta y cuatro especies entre la cuales se describen dos nuevas especies (Artemopyra urubuense y Retusotriletesamazonensis).
Un único espécimen atribuido al género Grandispora, idéntico a especimenes inéditos hallados en estratos lochkovianos de Arabia Saudita poruno de los autores (PS), ha sido encontrado en una de las muestras de edad lochkoviana del Río Urubu. Este constituye el registro geológico másantiguo de Grandispora. Entre los abundantes acritarcos presentes, especimenes del género Schizocystia se encuentran por primera vez en estratosmás antiguos que devónicos.
Si bien los estudios biostratigráficos de mioesporas del Silúrico-Lochkoviano temprano en América del Sur se hallan en sus primeras etapas, lasasociaciones de la Cuenca del Amazonas no parecen mostrar diferencias significativas de composición, respecto a asociaciones coetáneas de otrasáreas del noroeste de Gondwana. Esto puede resultar sorprendente dado que las localidades con mioesporas consideradas se encuentranampliamente distribuidas en latitudes altas a bajas, desde climas templados fríos a climas secos subtropicales. Por otra parte, las asociacionesgondwánicas siluro-devónicas difieren considerablemente de las palinofloras contemporáneas del Continente de las Viejas Areniscas Rojas y susregiones adyacentes, en contradicción con interpretaciones fitogeográficas recientes basadas en megafloras.# 2008 Elsevier Masson SAS. All rights reserved.
Keywords: Silurian; Devonian; Amazon Basin; Miospore; Biostratigraphy; Palaeogeography
Mots clés : Silurien ; Dévonien ; Basin d’Amazonie ; Miospore ; Biostratigraphie ; Paléogéographie
Palabras clave : Siluriano; Devonianos; Cuenca del Amazonas; Miosporas; Biostratigráficos; Paleogeográficas
1. Introduction
The study area is located to the northeast of Manaus City,Amazonas State, northern Brazil. The valleys of the Urubuand Uatumã Rivers were prospected in the 1950’s and 1990’sby geologists of the Brazilian state-owned oil company,Petrobras. Both rivers are left bank tributaries of the AmazonRiver (Fig. 1). Based on samples partly restudied in thepresent paper and from other parts of the Amazon Basin,Lange (1967) erected a first chitinozo-anacritarch biostrati-graphic scheme for the area. Recent chitinozoan studies inBrazil (Grahn, 1999; Grahn and Melo, 2002; Grahn et al.,2000, 2005) have allowed Lange’s biostratigraphic scheme tobe progressively modified and eventually replaced. In thelight of these newer results, the chitinozoan taxonomy andbiostratigraphy of the Urubu area was also restudied (Grahnand Melo, 2003).
The samples analyzed here for miospores have beenpreviously investigated by Grahn and Melo (2003) forchitinozoans. Although they are not rich in miospores, theassemblages are diversified, and their study is importantbecause available miospore data for the Late Silurian andSiluro-Devonian boundary is still rare in South America.
2. Previous miospores studies
Previous reports on Late Silurian–Early Lochkovianmiospores are geographically limited. In the northeastern partof Western Gondwana, the available information derivesmainly from: Spain (Cramer and Rodríguez, 1977; Crameret al., 1976; Richardson et al., 2001; Rodríguez, 1978a, 1978b,1978c, 1983), Brittany (Moreau-Benoit, 1976, 1980, 1994;Moreau-Benoit and Poncet, 1982; Moreau-Benoit andDubreuil, 1987; Rauscher and Robardet, 1975; Steemans,1989), Libya (Jardiné and Yapaudjian, 1968; Richardson et al.,1981; Rubinstein and Steemans, 2002; Streel et al., 1990),Turkey (Steemans et al., 1996) and Saudi Arabia (Steemans,1995; Steemans et al., 2001). Otherwise, most of the other dataon this stratigraphic interval concerns the Old Red SandstoneContinent and adjacent areas: UK (e.g. Burgess and Richard-son, 1995; Lavender and Wellman, 2002; Richardson, 1967;Richardson and Lister, 1969; Richardson and McGregor, 1986;Richardson et al., 1981, 1984, 1982; Wellman, 1993a, 1993b;Wellman and Richardson, 1993, 1996; Wellman et al., 1998,2000), Ardenno-Rhenan region (e.g. Breuer et al., 2005; Rocheet al., 1986; Steemans, 1981, 1982a, 1982b, 1989; Steemansand Gerrienne, 1984; Steemans et al., 2000; Streel et al., 1981,
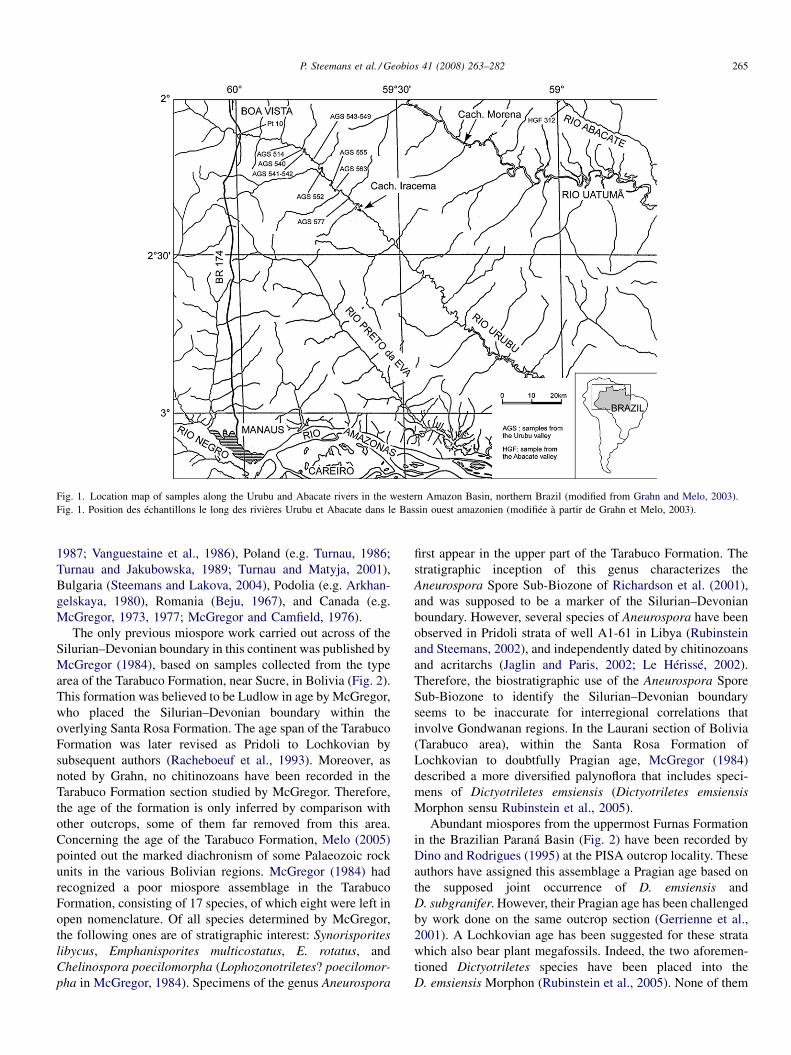
Fig. 1. Location map of samples along the Urubu and Abacate rivers in the western Amazon Basin, northern Brazil (modified from Grahn and Melo, 2003).Fig. 1. Position des échantillons le long des rivières Urubu et Abacate dans le Bassin ouest amazonien (modifiée à partir de Grahn et Melo, 2003).
P. Steemans et al. / Geobios 41 (2008) 263–282 265
1987; Vanguestaine et al., 1986), Poland (e.g. Turnau, 1986;Turnau and Jakubowska, 1989; Turnau and Matyja, 2001),Bulgaria (Steemans and Lakova, 2004), Podolia (e.g. Arkhan-gelskaya, 1980), Romania (Beju, 1967), and Canada (e.g.McGregor, 1973, 1977; McGregor and Camfield, 1976).
The only previous miospore work carried out across of theSilurian–Devonian boundary in this continent was published byMcGregor (1984), based on samples collected from the typearea of the Tarabuco Formation, near Sucre, in Bolivia (Fig. 2).This formation was believed to be Ludlow in age by McGregor,who placed the Silurian–Devonian boundary within theoverlying Santa Rosa Formation. The age span of the TarabucoFormation was later revised as Pridoli to Lochkovian bysubsequent authors (Racheboeuf et al., 1993). Moreover, asnoted by Grahn, no chitinozoans have been recorded in theTarabuco Formation section studied by McGregor. Therefore,the age of the formation is only inferred by comparison withother outcrops, some of them far removed from this area.Concerning the age of the Tarabuco Formation, Melo (2005)pointed out the marked diachronism of some Palaeozoic rockunits in the various Bolivian regions. McGregor (1984) hadrecognized a poor miospore assemblage in the TarabucoFormation, consisting of 17 species, of which eight were left inopen nomenclature. Of all species determined by McGregor,the following ones are of stratigraphic interest: Synorisporiteslibycus, Emphanisporites multicostatus, E. rotatus, andChelinospora poecilomorpha (Lophozonotriletes? poecilomor-pha in McGregor, 1984). Specimens of the genus Aneurospora
first appear in the upper part of the Tarabuco Formation. Thestratigraphic inception of this genus characterizes theAneurospora Spore Sub-Biozone of Richardson et al. (2001),and was supposed to be a marker of the Silurian–Devonianboundary. However, several species of Aneurospora have beenobserved in Pridoli strata of well A1-61 in Libya (Rubinsteinand Steemans, 2002), and independently dated by chitinozoansand acritarchs (Jaglin and Paris, 2002; Le Hérissé, 2002).Therefore, the biostratigraphic use of the Aneurospora SporeSub-Biozone to identify the Silurian–Devonian boundaryseems to be inaccurate for interregional correlations thatinvolve Gondwanan regions. In the Laurani section of Bolivia(Tarabuco area), within the Santa Rosa Formation ofLochkovian to doubtfully Pragian age, McGregor (1984)described a more diversified palynoflora that includes speci-mens of Dictyotriletes emsiensis (Dictyotriletes emsiensisMorphon sensu Rubinstein et al., 2005).
Abundant miospores from the uppermost Furnas Formationin the Brazilian Paraná Basin (Fig. 2) have been recorded byDino and Rodrigues (1995) at the PISA outcrop locality. Theseauthors have assigned this assemblage a Pragian age based onthe supposed joint occurrence of D. emsiensis andD. subgranifer. However, their Pragian age has been challengedby work done on the same outcrop section (Gerrienne et al.,2001). A Lochkovian age has been suggested for these stratawhich also bear plant megafossils. Indeed, the two aforemen-tioned Dictyotriletes species have been placed into theD. emsiensis Morphon (Rubinstein et al., 2005). None of them

Fig. 2. Early Devonian Gondwanan palaeogeographic/palaeoclimatic map modified from Scotese et al. (1999), showing the distribution of mountains (dark grey),lands (medium grey), shallow seas (pale grey) and deep seas (no shading). Location of the Southern Pole after: (A) Scotese et al. (1999), (B) Torsvik and Cocks (2004).Location of miospore records around the Siluro-Devonian boundary: (1) Amazon Basin, Brazil (Grahn and Melo, 2003; Melo and Loboziak, 2003, and present work),(2) Solimões Basin, Brazil (Rubinstein et al., 2005), (3) Cordillera Oriental, Bolivia (McGregor, 1984), (4) Paraná Basin, Brazil (Dino and Rodrigues, 1995;Gerrienne et al., 2001), (5) Precordillera, Argentina (Rubinstein et al., 2006), (6) Saudi Arabia (Steemans, 1995; Steemans et al., in preparation; Steemans et al., 2001,and unpublished data), (7) Ghadamis Basin, Libya (Rubinstein and Steemans, 2002), (8) Cantabrian Mountains, Spain (Richardson et al., 2001, and herein), (9)Brittany, France (Moreau-Benoit, 1994 and herein; Steemans, 1989 and unpublished data).Fig. 2. Carte paléogéographique/paléoclimatologiques du Gondwana au Dévonien Inférieur modifiée d’après Scotese et al. (1999) montrant la distribution desmontagnes (gris foncé), des terres émergées (gris moyen), des mers peu profondes (gris clair) et des océans profonds (blanc). Localisation du Pôle Sud d’après : (A)Scotese et al. (1999) ; (B) Torsvik et Cocks (2004). Localisation des travaux de palynologies sur les miospores réalisés près de la limite siluro-dévonienne : (1) Bassind’Amazonie, Brésil (Grahn et Melo, 2003 ; Melo et Loboziak, 2003 et travail présent) ; (2) Bassin de Solimões, Brésil (Rubinstein et al., 2005) ; (3) Cordillèreorientale, Bolivie (McGregor, 1984) ; (4) Bassin du Paraná, Brésil (Dino et Rodrigues, 1995 ; Gerrienne et al., 2001) ; (5) Précordillère, Argentine (Rubinstein et al.,2006) ; (6) Saudi Arabia (Steemans, 1995 ; Steemans et al., en préparation ; Steemans et al., 2001 et données non publiées) ; (7) Bassin de Ghadames, Libye(Rubinstein et Steemans, 2002) ; (8) Montagne Cantabrique, Espagne (Richardson et al., 2001 et inclus) ; (9) Bretagne, France (Moreau-Benoit, 1994 et inclus ;Steemans, 1989 et données non publiées).
P. Steemans et al. / Geobios 41 (2008) 263–282266
are true D. emsiensis (Allen) McGregor (1973), as they lack thecharacteristic spines at the muri junction, but instead, theyrepresent intermediate forms between D. emsiensis andD. granulatus Steemans (1989). Similarly, specimens identifiedas D. subgranifer in the Furnas Formation do not show theunevenly beaded structure along the upper edge of the muri ofthe reticulum. Excepting those species, the remainder of thepalynoflora recorded by Dino and Rodrigues has a pre-Pragian‘‘aspect’’, including such species as Synorisporites verrucatus,Emphanisporites micrornatus, Chelinospora poecilomorpha(Lophozonotriletes? poecilomorpha in Dino and Rodrigues,1995), and Amicosporites splendidus. In addition, Dibolispor-ites geikiei, a species only known in the Pridoli (Rubinstein andSteemans, 2002) and Lochkovian (Wellman and Richardson,1996), has been reported from those same strata by Gerrienneet al. (2001). This assemblage is very similar to the assemblagedescribed by McGregor (1984) in the Santa Rosa Formation, aunit which is nowadays regarded as mainly restricted to theLochkovian (Melo, 2005).
Melo and Loboziak (2003) have published on the Devonian–
Early Carboniferous miospore biostratigraphy of the AmazonBasin using numerous wells drilled in an area to the south andeast of the Urubu region. However, their Lochkovian miospore
database is relatively poor in comparison to younger intervals,because of the limited pre-Emsian sampling. In the uppermostpart of the Manacapuru Formation, samples investigated haveyielded species like Archaeozonotriletes chulus, Synorisporitestripapillatus and Cirratriradites diaphanus. The latter speciesallowed them to propose an apparent correlation with at leastthe upper part of the E. micrornatus var. sinuosus PhylogenicZone, which is within the micrornatus–newportensis OppelZone (Breuer et al., 2005; Steemans, 1989; Streel et al., 1987).The basal part of the overlying Jatapu Member of the MaecuruFormation has yielded very few identifiable miospore species,including: Archaeozonotriletes chulus, Biornatispora salopien-sis, Brochotriletes foveolatus, Chelinospora cassicula, Dibo-lisporites spp., Synorisporites spp. and Dictyotriletes emsiensis.Melo and Loboziak have correlated this assemblage with theemsiensis Interval Zone, i.e., the highest subdivision of thebreconensis–zavallatus Oppel Zone (Streel et al., 1987;Steemans, 1989) the base of which is below the lower limitof the Pragian Stage. However, as already explained, becausemost records of D. emsiensis in Gondwanan areas are actuallyassignable to the homonymous Morphon group, this species isunsuitable for accurate biostratigraphical correlations betweenthe Old Red Sandstone and Gondwana paleocontinents.

P. Steemans et al. / Geobios 41 (2008) 263–282 267
Another Lochkovian palynoflora, dated by chitinozoans(Grahn et al., 2003), comprising 53 miospore species, has beenreported from Petrobras well 1-JD-1-AM, drilled in theSolimões Basin, northwestern Brazil (Rubinstein et al.,2005). This assemblage belongs to the D. emsiensis MorphonAssemblage Zone, which is tentatively correlated with the Nthrough E Biozones of Western Europe, Lochkovian to EarlyPragian in age.
In Argentina, Silurian sediments exposed in the QuebradaAncha locality, Central Precordillera of San Juan, contain richacritarch and miospore assemblages (Rubinstein and Brussa,1999; Rubinstein et al., 2006). The Los Espejos Formation,considered as Wenlock to locally Pridoli in age, has yieldeddiverse, abundant and well-preserved marine and terrestrialpalynomorphs. The lower levels show a predominance ofacritarchs and related forms. Miospores become more frequenttowards the formation’s top, with trilete spores outnumberingthe cryptospores. Miospores include: cf. Chelinospora poeci-lomorpha, cf. Synorisporites verrucatus, cf. Clivosisporaverrucata var. convoluta, Chelinospora sp. A in Richardsonand Lister, 1969, C. sanpetrensis, Scylaspora vetusta, S. cf.scripta in Richardson et al., 2001, cf. Coronaspora cromatica,Aneurospora sp., Coronaspora sp., and Amicosporites sp. Mostof them are recorded for the first time in Argentina (Rubinsteinet al., 2006; Rubinstein and Steemans, 2007).
Fig. 3. Miospore biostratigraphic scales around the Siluro-Devonian boundary. Colbetween biozones erected by Steemans et al. (1996) and Rubinstein and Steemans (2by Richardson and McGregor (1986) and Richardson and Edwards (1989)); Colum(1989), Wellman and Richardson (1996); Column (4) Burgess and Richardson (1995)(1987); Column (7) Melo and Loboziak (2003).Fig. 3. Échelles stratigraphiques basées sur les spores à la limite siluro-dévonienne. Cles biozones sensu Steemans et al. (1996) et Rubinstein et Steemans (2002). B : bMcGregor (1986) et Richardson et Edwards (1989)) ; Colonne (2) Richardson et MRichardson (1996) ; Colonne (4) Burgess et Richardson (1995) ; Colonne (5) Richard(7) Melo et Loboziak (2003).
The first miospore zonation for the Silurian and Silurian–
Devonian boundary was erected by Richardson and McGregor(1986) in Euramerican and Libyan sections (Fig. 3). Theirscheme was later modified at the Siluro-Devonian transition byRichardson and Edwards (1989). Additional refinements havebeen introduced by Steemans et al. (1996) and Rubinstein andSteemans (Rubinstein and Steemans, 2002). An alternativeDevonian miospore biozonation for Euramerica was erected byStreel et al. (1987) and Steemans (1989). Recently, a newmiospore zonation for the Silurian–Devonian transition wasdefined in Spain (Richardson et al., 2001).
The base of the tripapillatus - spicula Assemblage Zone wasoriginally placed by Richardson and McGregor (1986) at theLudlow–Pridoli boundary (Fig. 3, column 1, B and column 2).In addition to the eponymous species, this biozone was alsocharacterised by the first appearance of patinate species of thegenera Chelinospora and Cymbosporites, and the inception oftrilete spores bearing interradial proximal papillae. However,Chelinospora has been found in the Ludlow of UK (Burgessand Richardson, 1995) and Libya (Rubinstein and Steemans,2002). Cymbosporites and tripapillate spores have also beenobserved in Ludlow sections of Libya. For this reason,Rubinstein and Steemans (2002) suggested placing the baseof the tripapillatus–spicula Biozone within the uppermostLudlow (Fig. 3, column 1, A). On the basis of observations done
umn (1) Chronostratigraphy (A: base of the Pridoli in relation to the boundary002). B: base of the Pridoli in regards of the boundary between biozones definedn (2) Richardson and McGregor (1986); Column (3) Richardson and Edwards; Column (5) Richardson et al. (2001); Column (6) Steemans (1989), Streel et al.
olonne (1) Chronostratigraphie (A : base du Pridoli par rapport aux limites entrease du Pridoli par rapport aux limites entre les biozones sensu Richardson et
cGregor (1986) ; Colonne (3) Richardson et Edwards (1989) et Wellman etson et al. (2001) ; Colonne (6) Steemans (1989) et Streel et al. (1987) ; Colonne

Fig. 4. Schematic correlation of the samples investigated. (1) Chronostratigraphic column; (2) Chitinozoan biozones; (3) Position of the samples in relation to Grahnand Melo’s (2003) biostratigraphic scheme. *: samples in stratigraphic sequence. Others are spot samples.Fig. 4. Diagramme de corrélation des échantillons étudiés. (1) Colonne chronostratigraphique ; (2) Biozones à chitinozoaires ; (3) Position des échantillons par rapportà la stratigraphie d’après Grahn et Melo (2003). * : échantillons en succession stratigraphique. Les autres sont des échantillons isolés.
P. Steemans et al. / Geobios 41 (2008) 263–282268
in the Cantabrian Mountains, Richardson et al. (2001) placedthe Ludlow–Pridoli boundary inside their new reticulata–
sanpetrensis Biozone. The Silurian–Devonian boundary wasinitially placed in the highest part of the tripapillatu–spiculaBiozone, then subsequently inside the Apiculiretusispora sp. EBiozone (Richardson and Edwards, 1989). Moreover, Richard-son et al. (2001) defined the Aneurospora spp. Sub-Biozone bythe first appearance of spores of the genus Aneurospora lackingproximal papillae, and considered the base of this biozone asclose to the Silurian–Devonian boundary. However, numerousnon-papillate specimens of Aneurospora has been observed inlayers as old as Earliest Pridoli in Libya (Rubinstein andSteemans, 2002).
Melo and Loboziak (2003) have erected two miosporebiozones in Lochkovian–Pragian sections of the AmazonBasin: the Non-spinose Zonate (NsZ) and the Dictyotriletesemsiensis Interval (Ems) Interval Zones with their basescharacterised by the first occurrences of non-spinoze zonatespores and of D. emsiensis, respectively. The boundary betweenboth zones is believed to be close to the Lochkovian–Pragianboundary by correlation with the miospore stratigraphic scaleestablished on the Old Red Sandstone Continent (Steemans,1989; Streel et al., 1987). Rubinstein et al. (2005) have sincedemonstrated that intermediate forms between ‘‘true’’D. emsiensis and other Dictyotriletes species coexisted inWestern Gondwana during the Lochkovian. Therefore, it iscurrently difficult to determine ‘‘true’’ D. emsiensis occur-rences in Gondwana. For this reason, the base of the E IntervalBiozone in the ORS Continent (characterised by the firstoccurrence of D. emsiensis in the absence of any intermediateforms) may not be strictly coeval with the base of thehomonymous biozone in Gondwana, where the variousintermediate forms of the emsiensis Morphon are present.
3. Geological setting and chitinozoan data
The samples studied herein were provided to one of us(J.H.G.M.) by PETROBRAS/CENPES geologist FernandoPellon de Miranda in the early 1990’s. They had been collectedby pioneer PETROBRAS field geologists in the 1950’s fromoutcrops along the Urubu and Abacate rivers (Fig. 1). The exact
geographic location of the sample AGS 610 is unknown. Thesamples have been investigated from the uppermost Pitinga andManacapuru Formations of the Amazon Basin. The outcropsare all situated in the northern margin of the basin (the so-calledNorth Platform — see Melo and Loboziak, 2003: Text-fig. 1).Megafossils are apparently rare in the Siluro-Devonianoutcrops of the study area, and chitinozoans are the onlymicrofossil group which has provided biostratigraphic results(Grahn and Melo, 2003) comparable to our miospore data.According to those palynomorphs, the samples collectivelyrange in age from Late Ludlow to Lochkovian. Some reworkedchitinozoans have been observed, but despite this, threechitinozoans assemblages have been defined: A (Late Ludlow),B (Early Pridoli) and C (Early Lochkovian). There is a clearseparation between Silurian and Devonian chitinozoan assem-blages, probably due to a hiatus within the Pridoli (Grahn, pers.com., 2007). Seven samples have been assigned to chitinozoanassemblage A (samples AGS 540, AGS 542, AGS 545, AGS547, AGS 549), six samples to the assemblage B (samples AGS514, AGS 548, AGS 552, AGS 555, AGS 610, HGF 312), andthree samples to assemblage C (samples Pt. 10, AGS 563 and577) (Fig. 4). According to current stratigraphic reinterpreta-tions by Yngve Grahn and one of us (J.H.G.M.), assemblages Aand possibly B would correspond to the uppermost PitingaFormation, and assemblage C to the overlying ManacapuruFormation (by contrast, assemblages B and C had beenoriginally attributed to the Manacapuru Formation, whereasassemblage A could belong to either of the two rock units). Thechitinozoan biostratigraphic results of Grahn and Melo (2003)permitted the recognition of downdip disturbances in thestratigraphic succession along the Urubu River, which may beattributed to either tectonic repetition (faulting) or unconfor-mities associated with some rock unit boundaries.
4. Miospore results (Fig. 5)
All samples investigated are rich in chitinozoans and formsof the organic-walled microphytoplankton (acritarchs, prasi-nophytes). Miospores are rare, demonstrating the marinecharacter of these strata. However, the progressive upwardincrease of the miospore/marine palynomorph ratio permits to
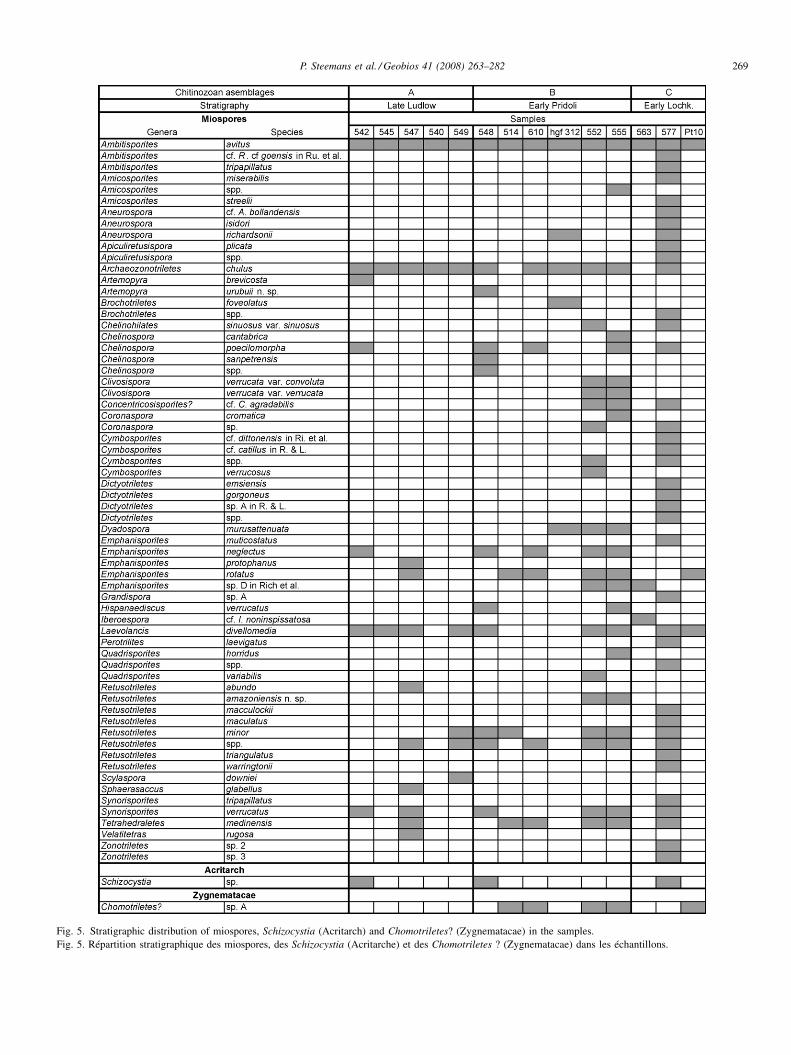
Fig. 5. Stratigraphic distribution of miospores, Schizocystia (Acritarch) and Chomotriletes? (Zygnematacae) in the samples.Fig. 5. Répartition stratigraphique des miospores, des Schizocystia (Acritarche) et des Chomotriletes ? (Zygnematacae) dans les échantillons.
P. Steemans et al. / Geobios 41 (2008) 263–282 269
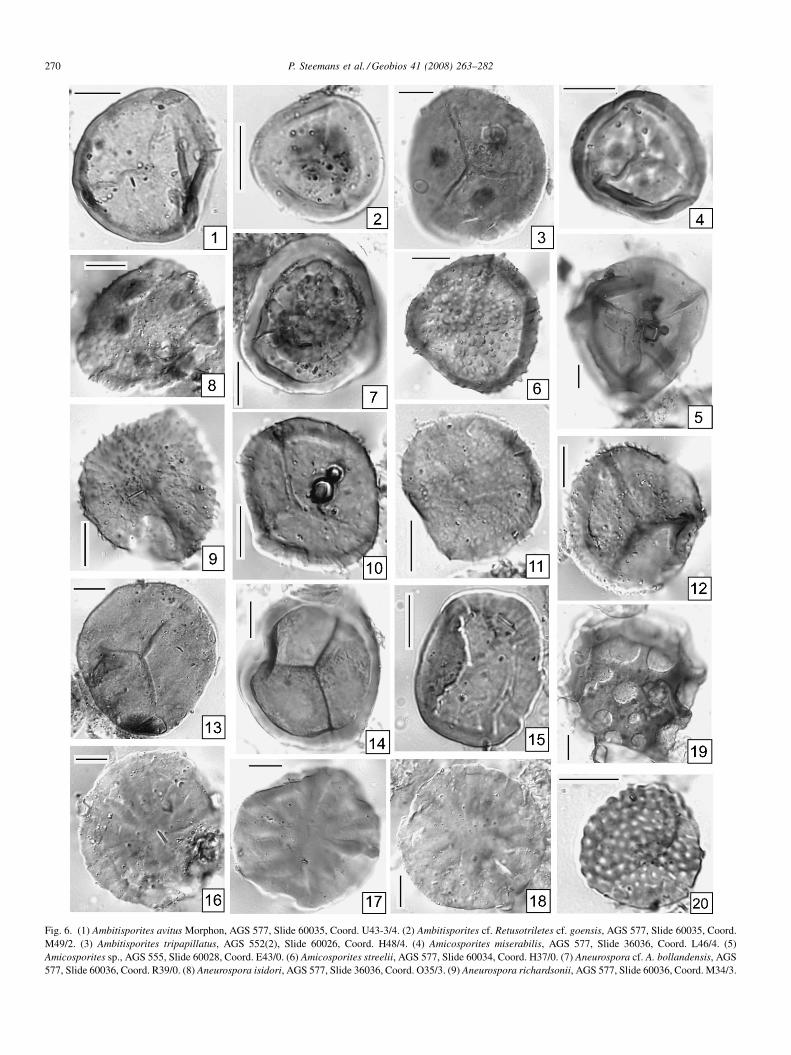
Fig. 6. (1) Ambitisporites avitus Morphon, AGS 577, Slide 60035, Coord. U43-3/4. (2) Ambitisporites cf. Retusotriletes cf. goensis, AGS 577, Slide 60035, Coord.M49/2. (3) Ambitisporites tripapillatus, AGS 552(2), Slide 60026, Coord. H48/4. (4) Amicosporites miserabilis, AGS 577, Slide 36036, Coord. L46/4. (5)Amicosporites sp., AGS 555, Slide 60028, Coord. E43/0. (6) Amicosporites streelii, AGS 577, Slide 60034, Coord. H37/0. (7) Aneurospora cf. A. bollandensis, AGS577, Slide 60036, Coord. R39/0. (8) Aneurospora isidori, AGS 577, Slide 36036, Coord. O35/3. (9) Aneurospora richardsonii, AGS 577, Slide 60036, Coord. M34/3.
P. Steemans et al. / Geobios 41 (2008) 263–282270

P. Steemans et al. / Geobios 41 (2008) 263–282 271
infer a relatively more proximal marine setting for theLochkovian sediments. Palynomorphs are usually wellpreserved, generally yellow in colour (Fig. 5).
4.1. Late Ludlow miospore assemblage
Samples AGS 543 and AGS 544 do not contain anymiospores, and the other five samples (AGS 540, AGS 542,AGS 545, AGS 547, AGS 549) are poor in miospores.Cryptospores are very rare, represented by Tetrahedraletesmedinensis, Laevolancis divellomedia, Artemopyra brevicosta(AGS 542), Sphaerasaccus glabellus and Velatitetras rugosa.The last two species have been observed in only one sample(AGS 547), and could be reworked from older sediments.Trilete spores are mainly represented by such ubiquitousspecies as Ambitisporites avitus/dilutus, Archaeozonotrileteschulus (in all five samples), Retusotriletes spp., Emphanispor-ites neglectus and E. rotatus. Spores of greater stratigraphicsignificance are the following species: Chelinospora poecilo-morpha, Emphanisporites protophanus, Scylaspora downieiand Synorisporites verrucatus. All are compatible with aLudlow age. Although S. verrucatus was formerly regarded asnot older than Pridoli (Richardson and McGregor, 1986), thisspecies was later observed in Ludlow sections of Libya(Rubinstein and Steemans, 2002).
4.2. Pridoli miospore assemblage
Pridoli samples are richer in miospores than Ludlow ones,however cryptospores remain very rare. The followingcryptospore species have been observed: Chelinohilatessinuosus var. sinuosus, Dyadospora murusdensa, Hispanae-discus verrucatus, Laevolancis divellomedia and Tetrahedra-letes medinensis. All are compatible with a Pridoli age, exceptC. sinuosus var. sinuosus which had been observed, thus far,only in the lower Lochkovian of the British Isles (Wellman andRichardson, 1996) and Saudi Arabia (Steemans et al., 2001).Currently, the total stratigraphic range of this species is stillpoorly known. Cryptospores previously identified as ‘‘Spore?with radial ribs’’ by Richardson and Ioannides (1973), anddescribed here as Artemopyra urubuense n. sp., have beenalready recorded in Libya, within the Tanezzuft Formation(Principal Shales Formation) and lower part of the AcacusFormation (Sandy/Shaly Formation). These sections corre-spond to the verrucatus–brevicosta Biozone and libycus–
poecilomorphus Biozone of Ludlow to Pridoli age. The samemiospore species has also been described as Artemopyra sp. Ain Turkey (Steemans et al., 1996), within the libycus–
poecilomorphus Biozone, independently dated Pridoli byacritarchs. Other ubiquitous miospore species already observed
(10) Aneurospora richardsonii, AGS 577, Slide 60034, Coord M43/3. (11) Aneurrichardsonii, AGS 577, Slide 36036, Coord. (13) Apiculiretusispora plicata, AGS 5760036, Coord. F37/4. (15) Artemopyra brevicosta, AGS 542, Slide 60002, Coord. K(17) Artemopyra urubuii nov. sp, Holotype, AGS 548, Slide 60012, Coord. T43/1.Brochotriletes foveolatus, HGF 312, Slide 9202737*, Coord. W23/2. (20) Brochot
in the Ludlow samples are also persistent here (A. avitus/dilutus, A. chulus, and Retusotriletes spp.). Chelinosporasanpetrensis, C. poecilomorpha, Clivosispora verrucata, andCoronaspora cromatica are known to range from the Ludlowthrough the Lochkovian. In Spain (Richardson et al., 2001),Chelinospora cantabrica (sample AGS 555) occurs in the RSBiozone (Late Ludlow to Pridoli) and H Biozone (Pridoli). InLibya (Rubinstein and Steemans, 2002), the same species hasbeen recorded as Dictyotriletes sp. in samples from thetripapillatus–spicula and Apiculiretusispora sp. E Biozones, ofLate Ludlow to Pridoli age, independently dated by chit-inozoans (Jaglin and Paris, 2002). Emphanisporites sp. D isknown in the Downton Group of UK and in Late Ludlowsections of Libya (Richardson et al., 1981). Cymbosporitesverrucosus is known in the Downtonian of UK (Richardson andLister, 1969).
The single sample from the Abacate River (HGF 312)belongs to chitinozoan assemblage B sensu Grahn and Melo(2003), and has therefore been dated Early Pridoli by theseauthors. Although poor in miospores, this sample has yieldedone reliable specimen of Brochotriletes foveolatus. This speciesis widely distributed over Euramerican and Gondwanan regionsand is thus far unknown in pre-Lochkovian sections. At leastthree possible explanations for this apparent anomaly can bedevised:
� th
osp7,43(1
ril
e chitinozoan age determination of sample HGF 312 couldbe incorrect (reworking) and in need of revision;
� th e total age span of chitinozoan assemblage B could extendinto the Early Lochkovian;
� B . foveolatus could appear earlier in South America thanelsewhere.
Therefore, the exact stratigraphic position of this sampleremains uncertain for the moment. The presence of specimensof Aneurospora richardsonii (Aneurospora sp. 2 in Rubinsteinand Steemans, 2002) is in permissive agreement with theLochkovian dating of sample HGF 312, taking into considera-tion the miospore zonation established in north Spain byRichardson et al. (2001). However, numerous Aneurosporaspecimens have already been observed in the Pridoli strata ofLibya (Rubinstein and Steemans, 2002), and thus, Aneurosporacannot be reliably used to identify the Lochkovian.
4.3. Lochkovian miospore assemblage
Of all samples studied here, AGS 577 is the richest one inmiospores. Cryptospores remain very rare. Tripapillatemiospores, typically common in the upper Silurian elsewhere,have not been observed in the Amazonian samples containing
ora richardsonii, AGS 577, Slide 60034, Coord. E34/0. (12) AneurosporaSlide 60036, Coord. S25/4. (14) Archaeozonotriletes chulus, AGS 577, Slide/0. (16) Artemopyra urubuii nov. sp., AGS 548, Slide 60011, Coord. M28/0.8) Artemopyra urubuii nov. sp., Paratype, Slide 60013, Coord. H38/3. (19)etes spp., AGS 577, Slide 60035, Coord. E29/0.
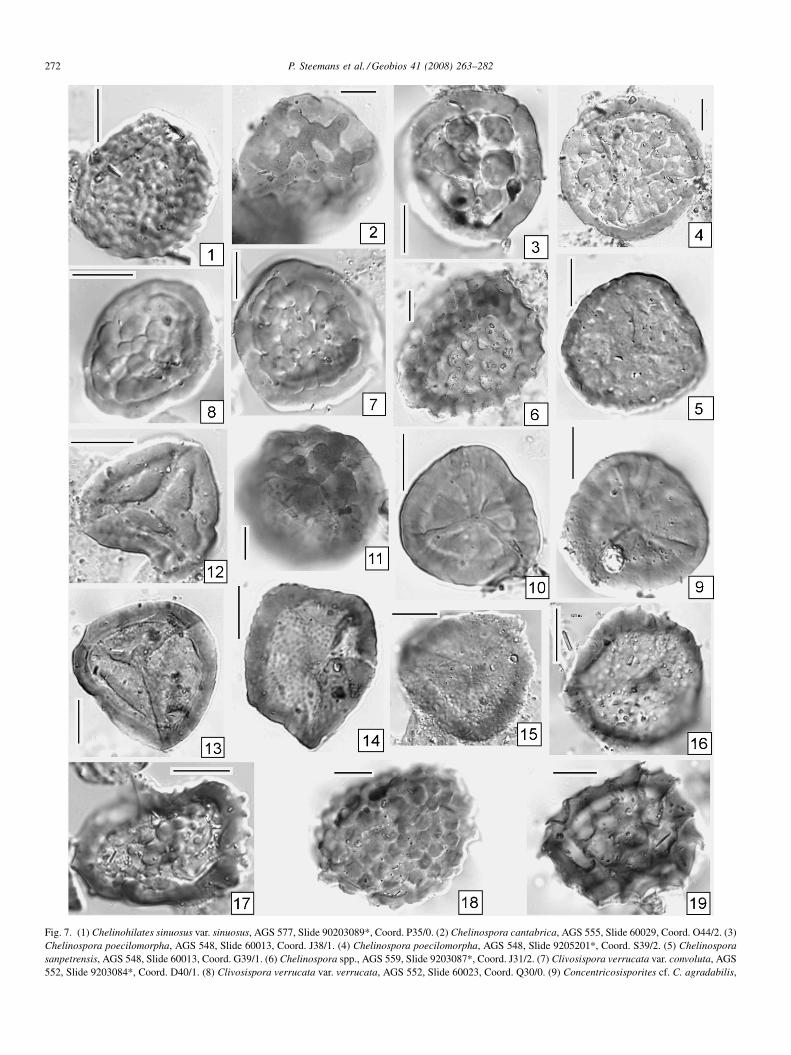
Fig. 7. (1) Chelinohilates sinuosus var. sinuosus, AGS 577, Slide 90203089*, Coord. P35/0. (2) Chelinospora cantabrica, AGS 555, Slide 60029, Coord. O44/2. (3)Chelinospora poecilomorpha, AGS 548, Slide 60013, Coord. J38/1. (4) Chelinospora poecilomorpha, AGS 548, Slide 9205201*, Coord. S39/2. (5) Chelinosporasanpetrensis, AGS 548, Slide 60013, Coord. G39/1. (6) Chelinospora spp., AGS 559, Slide 9203087*, Coord. J31/2. (7) Clivosispora verrucata var. convoluta, AGS552, Slide 9203084*, Coord. D40/1. (8) Clivosispora verrucata var. verrucata, AGS 552, Slide 60023, Coord. Q30/0. (9) Concentricosisporites cf. C. agradabilis,
P. Steemans et al. / Geobios 41 (2008) 263–282272

P. Steemans et al. / Geobios 41 (2008) 263–282 273
chitinozoan assemblage B, but are well represented in theLochkovian samples, with Ambitisporites tripapillatus, Retu-sotriletes maculatus, and Synorisporites tripapillatus. Inrespect of the other species present, some are unknown belowthe Silurian–Devonian boundary, e.g.: Amicosporites streelii,Aneurospora isidori, Perotrilites laevigatus, and particularlythe Dictyotriletes emsiensis Morphon (represented by a singlespecimen). This miospore assemblage seems to be closelyrelated to the upper Furnas Formation palynoflora of the ParanáBasin, southern Brazil (Dino and Rodrigues, 1995), which iscurrently dated Early Lochkovian (Gerrienne et al., 2001;Rubinstein et al., 2005).
Of particular interest is the presence of one single but easilyrecognizable specimen of a new species, here described asGrandispora sp. A, which has been previously observed in theLochkovian of Saudi Arabia (Steemans et al., 2001). It is theoldest species of the genus Grandispora, which so far has norecords below the upper Emsian. This finding implies that theNon-spinose Zonates Interval Zone (NsZ) of Melo andLoboziak (2003) (pre-Latest Lochkovian in age) needsprobably to be emended, in view of the presence of smallornamented zonate spores in the lower Lochkovian.
4.3.1. SchizocystiaThe samples investigated herein are usually rich in
acritarchs. Many acritarch taxa have been recognized in thecourse of the miospore study of the slides, including somespecimens of the genus Schizocystia. Because of their supposedstratigraphic interest, we have included some observations andcomments about them in this paper.
The genus Schizocystia Cookson and Eisenack, 1972 hasbeen emended and used for the first time in Palaeozoic strata byJardiné et al. (1972), in the Algerian Sahara. The single speciesS. pilosa Jardiné et al., 1972 appears in levels assigned to the‘‘Gedinnian–Siegenian’’ by these authors. Later, Jardiné et al.(1974) defined another species, S. saharica. Both S. pilosa andS. saharica first occur within Zone I of the Algerian Sahara,dated Gedinnian (Jardiné et al., 1974), with the latter persistinginto the Middle Devonian. Subsequently, S. pilosa andS. saharica have been also found in the Devonian of SouthAmerica, thus confirming their clear Gondwanan affinities.
S. pilosa and S. saharica have been regarded as markers ofthe middle part of the Zudañeziano stage, which is the lowestDevonian stage of the Cordilleran Cycle of Bolivia (Suárez-Soruco, 1989; Suárez-Soruco and Lobo-Boneta, 1983). Aprobable specimen of S. pilosa from the Santa Rosa Formation,in the Cordillera Oriental of southern Bolivia, has beenillustrated by McGregor (1984) as an ‘‘unidentified acritarch’’(Pl. 4, Fig. 16 in McGregor, 1984). Vavrdová et al. (1996)pointed out the abundance of S. pilosa in the TequejeFormation, in the Madre de Dios Basin, northern Bolivia,
AGS 577, Slide 36036, Coord. F35/3-4. (10) Concentricosisporites cf. C. agradabilisSlide 36030, Coord. O37/2. (12) Coronaspora spp., AGS 577, Slide 9203089*, CooCymbosporites cf. catillus in Richardson and Lister, AGS 577, Slide 60034, Coord. H36036, Coord. D41/0. (16) Cymbosporites spp., AGS 577, Slide 36036, Coord. MCymbosporites verrucosus, AGS 548, Slide 60013, Coord. O41/0. (19) Dictyotrile
and Limachi et al. (1996) placed the limit between the SantaRosa and Icla formations and the lower part of the IclaFormation within the Dictyotriletes Schizocystia saharica/S. piloza Zone. Although large numbers of Schizocystia spp.have been reported from Lochkovian–Pragian strata of Bolivia,Melo (2000) noted that these acritarchs are uncommon in someEarly Devonian sections investigated by PETROBRAS inBolivia.
In Brazil, Schizocystia spp. have been previously reportedfrom the uppermost Furnas Formation of the Paraná Basin, atthe PISA locality, first dated Pragian (Dino and Rodrigues,1995), then reinterpreted as Lochkovian (Gerrienne et al.,2001), and subsequently included in the D. emsiensis MorphonAssemblage Zone (Rubinstein et al., 2005). The acritarchSchizocystia is very frequent in Late Lochkovian strata of theSolimões Basin (Rubinstein et al., 2005), corresponding to theD. emsiensis Morphon Assemblage Zone, which is tentativelycorrelated with the N through the E Biozones of WesternEurope, and independently dated by means of chitinozoans. Inaddition, Schizocystia spp. (particularly S. pilosa) are alsolocally common in higher parts of the Jatapu Member of theMaecuru Formation in the Amazon Basin, within the younger(Pragian–Early Emsian) section of the Ems Biozone (J.H.G.Melo, unpublished data, 2002).
Schizocystia pilosa also occurs in the lower part of theTalacasto Formation, San Juan Precordillera of Argentina, thatranges from the upper Lochkovian to the Emsian, withindependent miospore and chitinozoan calibration (Le Hérisséet al., 1997). It is also present in the lower part of theVillavicencio Formation, of the Mendoza Precordillera,Argentina, which is dated Late Pragian to Early Emsian onthe basis of the miospores, and probably correlatable with theupper part of the Ems Biozone, in terms of the Devonianmiospore zonation of the Brazilian Amazon Basin (Rubinsteinand Steemans, in press).
In the Urubu River area, Schizocystia spp. first appearsbelow the Silurian–Devonian boundary. A few specimensassigned to Schizocystia cf. S. saharica are present in levelscorresponding to chitinozoan assemblages A and B, assigned tothe Late Ludlow and Early Pridoli respectively (Grahn andMelo, 2003), thus implying an earlier first appearance of thetaxon in the Amazon Basin (uppermost Pitinga Formation)when compared to other Western Gondwanan localities.Schizocystia pilosa is also recorded in Manacapuru Formationstrata attributed to chitinozoan assemblage C, of EarlyLochkovian age.
The first occurrence of Schizocystia spp. has previously beenconsidered as a reliable Earliest Devonian biostratigraphicalmarker in Western Gondwana. Hence, the apparent inception ofthe genus below the Silurian–Devonian boundary brings us tothe conclusions that:
, AGS 577, Slide 60036, Coord. O24/0. (11) Coronaspora cromatica, AGS 555,rd. U42/0. (13) Coronaspora spp., AGS 577, Slide 60027, Coord. O45/0. (14)30/2. (15) Cymbosporites cf. catillus in Richardson and Lister, AGS 577, Slide
45/2. (17) Cymbosporites spp., AGS 577, Slide 9203089*, Coord. F40/0. (18)tes emsiensis, AGS 577, Slide 60036, Coord. H44/3.
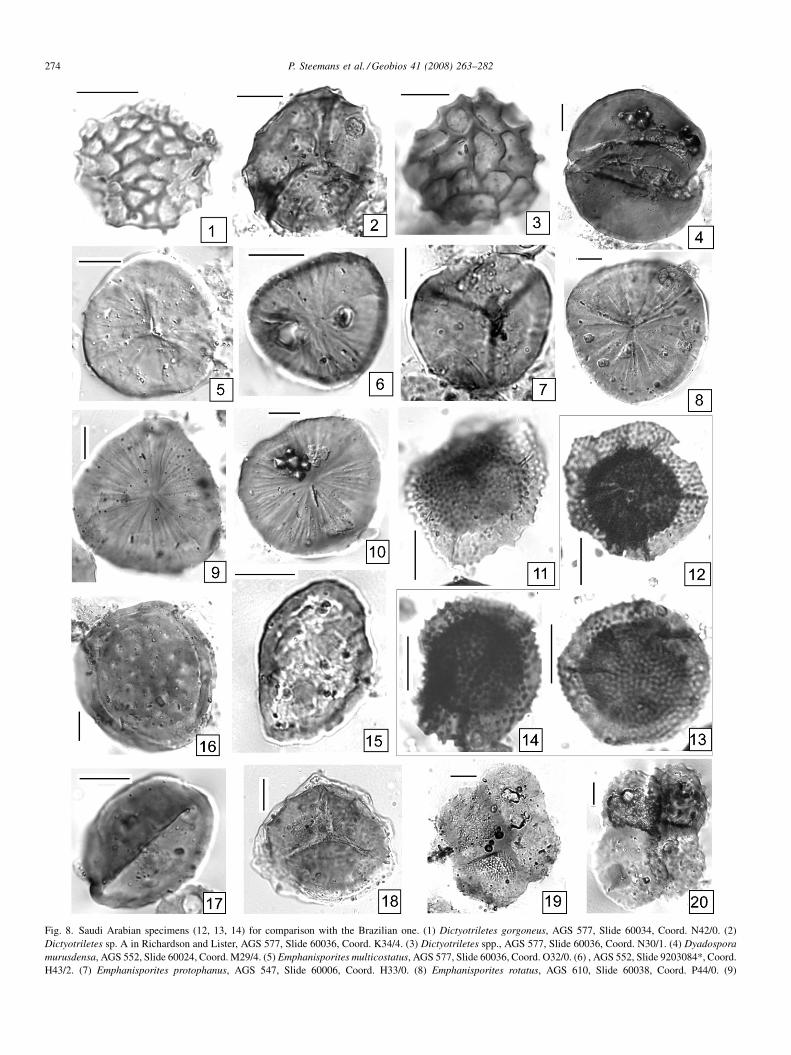
Fig. 8. Saudi Arabian specimens (12, 13, 14) for comparison with the Brazilian one. (1) Dictyotriletes gorgoneus, AGS 577, Slide 60034, Coord. N42/0. (2)Dictyotriletes sp. A in Richardson and Lister, AGS 577, Slide 60036, Coord. K34/4. (3) Dictyotriletes spp., AGS 577, Slide 60036, Coord. N30/1. (4) Dyadosporamurusdensa, AGS 552, Slide 60024, Coord. M29/4. (5) Emphanisporites multicostatus, AGS 577, Slide 60036, Coord. O32/0. (6) , AGS 552, Slide 9203084*, Coord.H43/2. (7) Emphanisporites protophanus, AGS 547, Slide 60006, Coord. H33/0. (8) Emphanisporites rotatus, AGS 610, Slide 60038, Coord. P44/0. (9)
P. Steemans et al. / Geobios 41 (2008) 263–282274

P. Steemans et al. / Geobios 41 (2008) 263–282 275
� e
Em4.SaHdi60
ither its range should be reconsidered in the light ofoccurrences in supposed Late Ludlow–Early Pridoli rocks ofthe Amazon Basin;
� th e age determination of samples AGS 542 and AGS 548 (onthe basis of chitinozoans) is inaccurate and needs biostrati-graphic re-evaluation. Miospore data from both samples isinsufficient to lend support to any of these possibilities.
5. Paleogeography
During the Siluro-Devonian interval, the South Pole wassituated close to the South American and African palaeoplatemargins. According to recent authors, the position of the SouthPole (Fig. 2) varies from latitudes close to the southernextremity of present-day Brazil (e.g. Torsvik and Cocks, 2004)to latitudes corresponding to northern Brazil (e.g. Scotese et al.,1999). Whatever the position of the South Pole, the LateSilurian and Lochkovian miospore localities in South Americaare around or above 608S paleolatitude and adjacent to theAntarctic Circle. According to location B of the South Pole(Fig. 2), the Brazilian Paraná Basin (Fig. 2, locality 4) occupieda near-polar position at that time (around 808S). The presenceof numerous miospores indicates an abundant high latitude landplant vegetation, thus confirming the existence of a warmerinterval during at least Late Silurian–Earliest Devonian times. Itcorroborates the palaeoclimatic conditions proposed by Scoteseet al. (1999), who described an extensive Cool Temperate beltthat extended from the South Pole down to mid-latitudes. It ismost likely, that no permanent ice cap existed during this timeinterval. A Dry Subtropical belt followed the perimeter ofGondwana from Spain to Australia. A Warm Temperate beltmay have lain between both areas. Surprisingly, all SouthAmerican localities, except perhaps locality 5 (Fig. 3) in SanJuan Province of Argentina (Rubinstein et al., 2006), haveyielded trilete spore assemblages remarkably similar to thosefrom lower(?) Gondwanan latitudes, with the paleoclimatevarying from cool temperate to warm temperate and drysubtropical. Conversely, Gondwanan trilete spore assemblagesshow significant differences to coeval palynofloras from the OldRed Sandstone Continent (Richardson et al., 2001; Steemanset al., in preparation). This is in apparent contradiction withresults obtained from Late Silurian megafloras (Raymond et al.,2006), which show a strong floral similarity betweenassemblages from Bolivia and Great Britain. Interestingly,almost all Gondwanan sites but locality 6 (Fig. 3) in SaudiArabia (Steemans et al., in preparation; Steemans et al., 2001)have yielded very few cryptospores. However, it should benoted that locality 6 is the only one where sediments have beendeposited in a continental basin, all others being clearly marineas indicated by numerous acritarchs and/or chitinozoans.
phanisporites sp. D in Richardson et al., AGS 555, Slide 60028, Coord. J41/0. (10)(11) Grandispora sp. A, AGS 577, Slide 60036, Coord. L41/2. (12) Grandisporaudi Arabia, Qatif, Slide 56869, Coord. Q4Emphanisporites neglectus0/0. (14)
ispanaediscus verrucatus, AGS 542, Slide 60002, Coord. P31/4. (16) Iberoesporavellomedia, AGS 548, Slide 60013, Coord. H35/0. (18) Perotrilites laevigatus, AG026, P47/0. (20) Quadrisporites variabilis, AGS 577, Slide 60036, P31/0.
6. Systematics
Examples of identified palynomorphs are provided onFigs. 6–10. The palynological slides are housed in thecollections of the PPM laboratory, University of Liège,Belgium. Each figured palynomorph is located by samplenumbers, slide numbers and England Finder coordinates. Thescale bars illustrate a length of 10 mm.
6.1. Incertae sedis
Three different genera of circular palynomorphs withconcentric ribs show close morphological affinities: Chomo-triletes Naumova, 1953, Circulisporites De Jersey emend.Norris, 1965, and Concentricystes Rossignol, 1962.
Chomotriletes comprise alete miospores with a circularoutline, and proximal and distal surfaces ornamented by closelyspaced concentric muri, which are either continuous or disjunctand may be absent on one hemisphere (Hart, 1964). Accordingto some authors, Chomotriletes can split into two hemispheres(Grenfell, 1995), whereas for others no excystment mechanismis evident (Playford and Dring, 1981). Circulisporites isornamented by continuous ribs instead of the interrupted ribs ofChomotriletes. Circulisporites is considered to be an acritarch(Norris, 1965). Concentricystes is morphologically similar toCirculisporites, and is undoubtedly of fluvial origin (Rossignol,1962). Grenfell (1995) regarded all three genera as probablezygnematacean algal spores.
Palynomorphs with concentric ribs observed in the studiedAmazonian material, mainly in samples from chitinozoanassemblage B and a single sample from assemblage C (samplePt 10), could be related to this group of zygnematacean spores.They are relatively abundant in the slides. The specimens areornamented by irregular circular ribs, which may beinterrupted, resembling fingerprints. Most of the specimenshave a proximal (?) laevigate hilum, which is equatoriallybordered by a slightly thicker crassitude. Rare specimens areassociated in dyads. In such cases, they are a little browner incolour, and the ribs of both cells seen by transparency seem tointersect. One folded specimen (Fig. 10(14)) is clearlycomposed by two loosely adpressed cells. Considering theshape of the ribs, these palynomorphs can be assigned toChomotriletes. According to our observations, the Amazonianspecimens are very similar to loosely attached cryptosporedyads which can frequently dissociate into monads. However,usually ubiquitous cryptospore taxa like Tetrahedraletes,Dyadospora, etc., are very rare in our samples, whereasrepresentatives of the genus Chomotriletes are abundant.Therefore, it is most likely that the latter are zygnemataceanalgae and not spores produced by land plants. Nevertheless,
Emphanisporites sp. D in Richardson et al., AGS 552, Slide 60022, Coord. Y47/sp. A, Saudi Arabia, Qatif, Slide 56869, Coord. G42/0. (13) Grandispora sp. A,Grandispora sp. A, Saudi Arabia, Qatif, Slide 56869, Coord. V36/4. (15.)cf. I. noninspissatosa, AGS 543, Slide 60032, Coord. K25/1. (17) LaevolancisS 577, Slide 9203089*, D28/3. (19) Quadrisporites variabilis, AGS 552, Slide
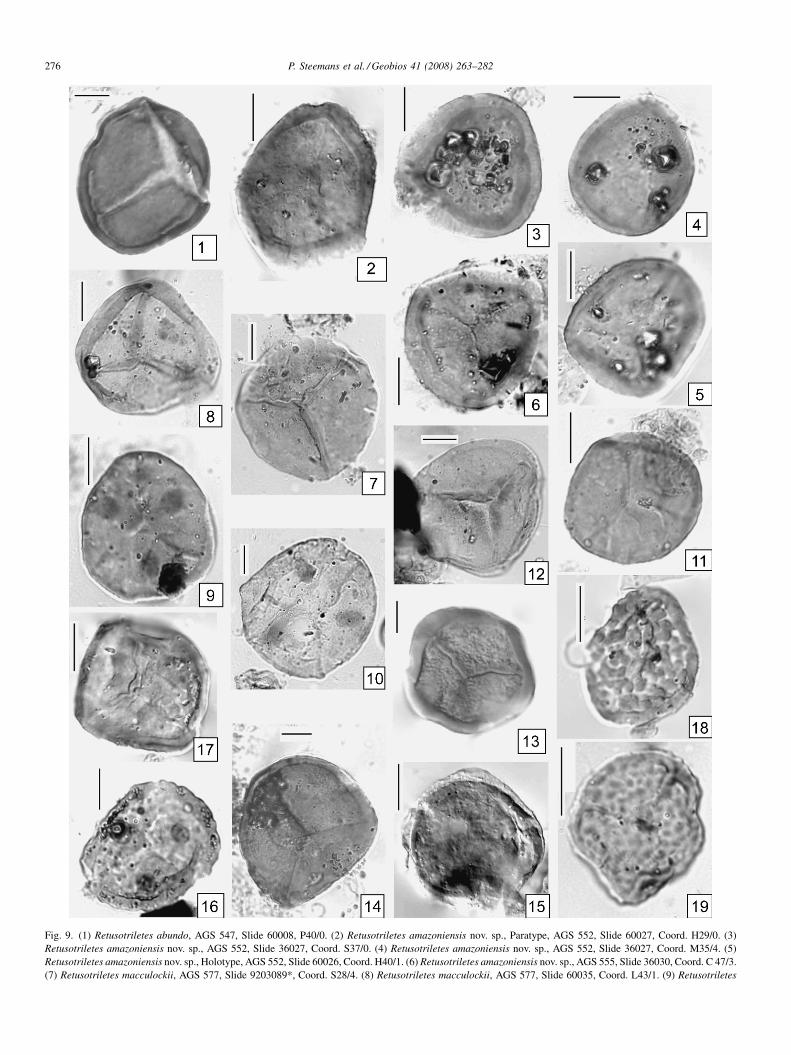
Fig. 9. (1) Retusotriletes abundo, AGS 547, Slide 60008, P40/0. (2) Retusotriletes amazoniensis nov. sp., Paratype, AGS 552, Slide 60027, Coord. H29/0. (3)Retusotriletes amazoniensis nov. sp., AGS 552, Slide 36027, Coord. S37/0. (4) Retusotriletes amazoniensis nov. sp., AGS 552, Slide 36027, Coord. M35/4. (5)Retusotriletes amazoniensis nov. sp., Holotype, AGS 552, Slide 60026, Coord. H40/1. (6) Retusotriletes amazoniensis nov. sp., AGS 555, Slide 36030, Coord. C 47/3.(7) Retusotriletes macculockii, AGS 577, Slide 9203089*, Coord. S28/4. (8) Retusotriletes macculockii, AGS 577, Slide 60035, Coord. L43/1. (9) Retusotriletes
P. Steemans et al. / Geobios 41 (2008) 263–282276

P. Steemans et al. / Geobios 41 (2008) 263–282 277
their morphological similarity to cryptospores remains intri-guing.
6.1.1. Description?Chomotriletes sp. AFig. 10(11–14).Circular to oval in shape, resembling a flat lentil usually
preserved in polar compression. Most of the specimens arepreserved as monads, and only a few (less than 10%) as dyads.Proximal exine is laevigate and thin. Exine thicker equatorially(1–2 mm) in some specimens. Distal face thin, with fineconcentric costae, 1 mm wide, somewhat irregular, showing asuccession of slight thickness variations. They are separated byfine channels, 0.5 mm wide. Ribs become irregular at the pole,forming fingerprint-like convolutions. When preserved asdyads, both cells are so perfectly superposed that it is difficultto recognise that they are dyads, except in specimens obliquelycompressed. In this case, the ribs of both cells seen bytransparency seem to intersect.
Diameters: 30 (43) 65 mm (measured on 23 specimens).Remark: if the attribution of these Amazonian specimens to
Chomotriletes is confirmed, they will constitute the oldestgeological record of this genus to our knowledge.
6.2. Miospores (Figs. 6–10)
6.2.1. New speciesArtemopyra urubuense nov. sp.Fig. 6(16–18)1973. ‘‘Spore? with radial ribs’’ in Richardson and
Ioannides, p. 288, Pl. 9, Figs. 13, 14.Derivation of name: refers to the Urubu River where the
samples have been collected.Holotype and type locality: Fig. 6(17), AGS 548, Slide
60012, Coord. T43/1.Diagnosis: a distally laevigate Artemopyra. Proximal face
with thick radial muri, becoming progressively indistincttowards the equator.
Description: circular to sub-circular amb. Presence of 14 to18 radial fragmented muri, becoming indistinct towards theequator, confined to one face, 3.5 to 4.5 mm wide. Exine thin.
Diameter: 44 (48) 51 mm (five specimens measured).Remarks: specimens observed in Brazil are very similar to
those described by Richardson and Ioannides (1973), whoconsidered them as of unknown origin because they lack atrilete mark.
Their overall morphology suggests close similarity tospores, but we prefer to include them in the cryptospore group.
Grandispora sp. ADescription: trilete camerate miospore, intexine and
exoexine amb circular. Diameter of inner body about 56 to
maculatus, AGS 577, Slide 60035, Coord. L43/3. (10) Retusotriletes maculatus, AG60034, Coord. P46/1. (12) Retusotriletes triangulatus, AGS 577, Slide 60035, CoorScylaspora downiei, AGS 540, Slide 60016, Coord. R38/2. (15) Sphaerasaccus glabAGS 577, Slide 60034, Coord. S32/1. (17) Synorisporites verrucatus, AGS 548, SlidCoord. Q38/4. (19) Synorisporites verrucatus, AGS 577, Slide 60035, Coord. F50
72% (Brazilian specimen: 67%) total spore diameter. Triletemark distinct to indistinct, sutures thin (1–2 mm in total width),that may extend to the equator of the spore. Intexine apparentlyunsculptured. Proximal surface of exoexine laevigate, distalsurface densely ornamented by round-topped or biformverrucae. Biform elements are conical surmounted by a smallhair-like spine, 0.5 (1.2) 1.8 mm (Brazilian specimen: 1.2–
1.8 mm) wide at their base, 0.5 (0.9) 1.4 mm (Brazilianspecimen: 1.10–1.2 mm) high, subcircular in plan view, and1 (1.3) 1.7 mm (Brazilian specimen: 1.2–1.5 mm) apart.
Diameter: 25 (29) 32 mm (Brazilian specimen: 32 mm)measured on eight specimens.
Comparisons: it is the smallest Grandispora ever observed.Other small Grandispora species previously described in theliterature include G. daemonii Loboziak et al., 1988, which is38–69 mm in diameter and ornamented with smaller ornaments,very densely packed, and G. inculta Allen, 1965, which is 51–
86 mm in diameter and ornamented with cones.Remarks: only one specimen has been observed in the
sample AGS 577. However, other (undescribed) specimenshave been found in the Qatif locality of Saudi Arabia (Steemanset al., in preparation; Steemans et al., 2001). Those ones areshown on Fig. 8(12–14). The description given here is based onLochkovian specimens from both localities. To our knowledge,Grandispora sp. A is the oldest Grandispora ever documented,since all previous records are Late Emsian or younger.
Retusotriletes amazonensis nov. sp.
Fig. 9(2, 3, 5, 6)Derivation of name: refers to the Amazon Basin where the
samples have been collected.Holotype and type locality: Fig 9(5), Holotype, AGS 552,
Slide 60026, Coord. H40/1.Diagnosis: a Retusotriletes with thick proximal curvaturae.Description: trilete spore with a circular to subcircular amb.
Trilete mark very thin, sometimes indistinct, extending on 4/5of the spore radius. Curvaturae forming an annulus 1 to 1.5 mmwidth with sharp edges, subparallel to the equatorial border.The distance between the annulus and the equatorial bordervaries from 1 to 4 mm on a same specimen. Distal and proximalexine laevigate, thin.
Diameters: 24 (27) 36 mm (12 specimens measured).Comparisons: no other Retusotriletes species shows a
similar proximal thick curvaturae.
6.2.2. Other miospore speciesAmbitisporites avitus Morphon Steemans et al., 1996.Ambitisporites tripapillatus Moreau-Benoit, 1976.Ambitisporites cf.? ‘‘Retusotriletes cf. goensis Rodriguez,
1983 in Rubinstein et al., 2005.Ambitisporites tripapillatus Moreau-Benoit, 1976.Amicosporites miserabilis Cramer, 1966.
S 577, Slide 60036, Coord. S25/1. (11) Retusotriletes minor, AGS 577, Slided. C43/4. (13) Scylaspora downiei, AGS 549, Slide 60021, Coord. G33/3. (14)ellus, AGS 547, Slide 60009, Coord. M28/0-2. (16) Synorisporites tripapillatus,e 60011, Coord. O31/1. (18) Synorisporites verrucatus, AGS 577, Slide 60036,/1.
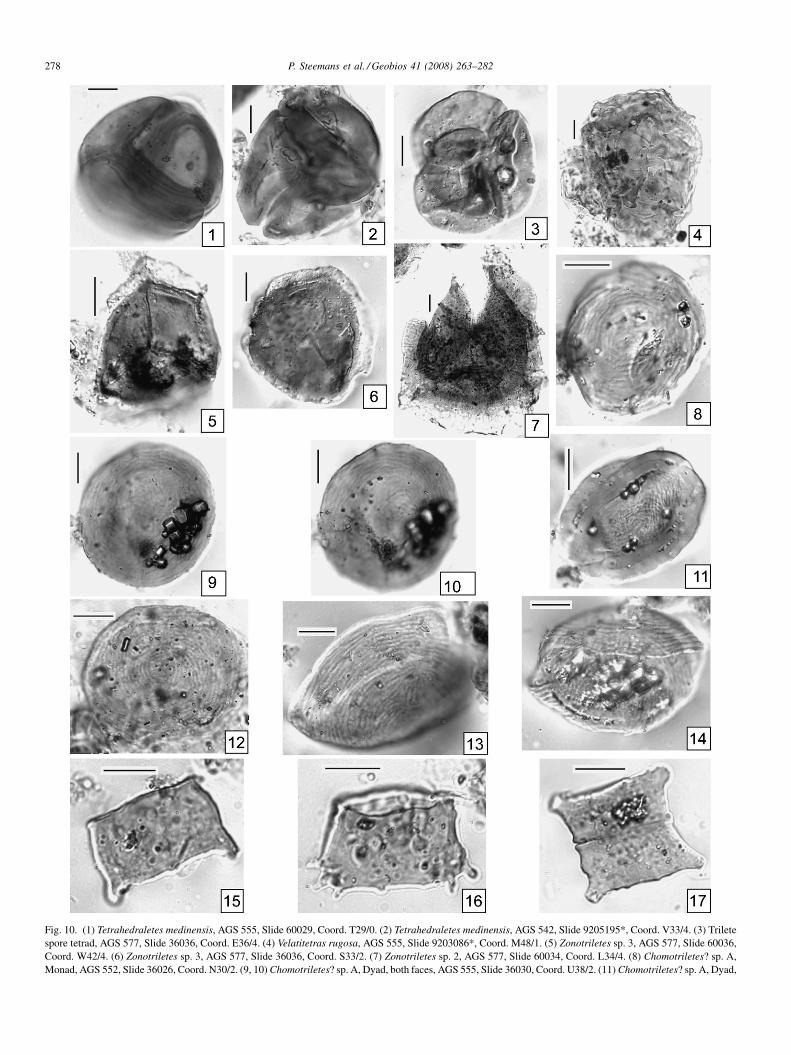
Fig. 10. (1) Tetrahedraletes medinensis, AGS 555, Slide 60029, Coord. T29/0. (2) Tetrahedraletes medinensis, AGS 542, Slide 9205195*, Coord. V33/4. (3) Triletespore tetrad, AGS 577, Slide 36036, Coord. E36/4. (4) Velatitetras rugosa, AGS 555, Slide 9203086*, Coord. M48/1. (5) Zonotriletes sp. 3, AGS 577, Slide 60036,Coord. W42/4. (6) Zonotriletes sp. 3, AGS 577, Slide 36036, Coord. S33/2. (7) Zonotriletes sp. 2, AGS 577, Slide 60034, Coord. L34/4. (8) Chomotriletes? sp. A,Monad, AGS 552, Slide 36026, Coord. N30/2. (9, 10) Chomotriletes? sp. A, Dyad, both faces, AGS 555, Slide 36030, Coord. U38/2. (11) Chomotriletes? sp. A, Dyad,
P. Steemans et al. / Geobios 41 (2008) 263–282278

P. Steemans et al. / Geobios 41 (2008) 263–282 279
Amicosporites streelii Steemans, 1989.
Aneurospora cf. Aneurospora bollandensis Steemans,1989.
Remarks: specimen similar to A. bollandensis Steemans(1989), but the three proximal inspissations that characterizethe species are not clearly visible.
Aneurospora richardsonii Richardson et al., 2001.Remarks: Aneurospora sp. 2 in Rubinstein and Steemans,
2002 is assignable to A. richardsonii.
Aneurospora isidori (Cramer and Díez) Richardson et al.,1982.
Apiculiretusispora plicata (Allen) Streel, 1967.Archaeozonotriletes chulus Morphon in Steemans et al.,
1996.Artemopyra brevicosta Burgess and Richardson, 1991.Brochotriletes foveolatus Naumova, 1953.Chelinohilates sinuosus var. sinuosus Wellman and Richard-
son, 1996.Chelinospora cantabrica Richardson et al., 2001.Chelinospora (Lophozonotriletes?) poecilomorpha
(Richardson and Ioannides) Richardson et al., 2001.Chelinospora (Brochotriletes) sanpetrensis (Rodriguez)
Richardson et al., 2001.Clivosispora verrucata var. convoluta McGregor and
Camfield, 1976.Clivosispora verrucata var. verrucata McGregor and
Camfield, 1976.
Concentricosisporites? cf. C. (Rodriguez) agradabilisRodriguez, 1983.
Remarks: the Amazonian specimens conform to theoriginal description of Rodríguez (1978b), but the proximalannular structure is not clearly visible.
Coronaspora cromatica Rodríguez emend Richardson et al.,2001.
Cymbosporites cf. dittonensis in Richardson and Lister,1969.
Cymbosporites cf. catillus in Richardson and Lister,1969.
Cymbosporites verrucosus Richardson and Lister,1969.
Dictyotriletes emsiensis Morphon Rubinstein et al., 2005.Dictyotriletes gorgoneus Cramer, 1967.Dictyotriletes sp. A in Richardson and Lister, 1969.Dyadospora murusattenuata Morphon Steemans et al.,
1995.Emphanisporites multicostatus Rodriguez, 1978b.Emphanisporites neglectus Vigran, 1964.Emphanisporites protophanus Richardson and Ioannides,
1973.Emphanisporites (McGregor) rotatus McGregor, 1973.
AGS 577, Slide 60036, Coord. V62/0. (12) Chomotriletes? sp. A, Monad, AGS 552,60024, Coord. T26/0. (14) Chomotriletes? sp. A, Dyad, lateral view, AGS 552, Slide2-4. (16) Schizocystia sp., AGS 548, Slide 60011, Coord. M35/4. (17) Schizocysti
Emphanisporites sp. D in Richardson et al., 1981.Hispanaediscus verrucatus Cramer emend Burgess and
Richardson, 1991.
Iberoespora cf. I. noninspissatosa Steemans, 1989.Remarks: the specimen illustrated here is similar to the
original species but is considerably larger in diameter(Brazilian material: 50 mm. Original description: 21 (29)36 mm).
Laevolancis divellomedia (Chibrikova) Burgess andRichardson, 1991.
Perotrilites laevigatus Steemans, 1989.Quadrisporites horridus Hennelly, 1959 ex Potonié and
Lele, 1961.Quadrisporites variabilis (Cramer) Strother, 1991.Retusotriletes abundo Rodríguez, 1978b.Retusotriletes maccullockii Wellman and Richardson, 1996.Retusotriletes maculatus McGregor and Camfield, 1976.Retusotriletes minor Kedo, 1963.Retusotriletes triangulatus (Streel) Streel, 1967.Scylaspora downiei Burgess and Richardson, 1995.Sphaerasaccus glabellus Steemans et al., 2000.Synorisporites tripapillatus Richardson and Lister, 1969.Synorisporites verrucatus Richardson and Lister, 1969.Tetrahedraletes medinensis Strother and Traverse, 1979
emend. Wellman and Richardson, 1993.Velatitetras rugosa (Strother and Traverse) Steemans et al.,
1996.Zonotriletes sp. 2 in Jardiné and Yapaudjian, 1968.Zonotriletes sp. 3 in Jardiné and Yapaudjian, 1968.
Acknowledgments
The authors wish to thank Yngve Grahn (PETROBRAS/CENPES consultant, Rio de Janeiro, Brazil) for valuableinformation on the Silurian–Earliest Devonian chitinozoanassemblages of the Amazon Basin. The author wish to thank Dr.C. Wellman (Sheffield, UK) and the anonymous reviewer fortheir comments. This paper is the result of a scientificcollaboration between the National Research Council ofArgentina (CONICET) and the National Funds for ScientificResearch of Belgium (NFSR). (PIP No. 5948; InternationalCooperation Project CONICET/FNRS 2002-2003). Theauthors publish with the permission of PETROBRAS - PetróleoBrasileiro S/A. C. Rubinstein benefited from a NFSRpostdoctoral grant at the University of Liège (2005-2006).J.H.G. Melo thanks PETROBRAS - Petróleo Brasileiro S/A forthe permission to publish this paper.
References
Arkhangelskaya, A.D., 1980. Plant spores from some Lower Devonian sectionsof the western regions of the Russian Plate. In: Byvsheva, T.V. (Ed.),
Slide 36027, Coord. R35/0. (13) Chomotriletes? sp. A, Monad, AGS 552, Slide36026, Coord. L33/2. (15) Schizocystia sp., AGS 542, Slide 60002, Coord. G41/a sp., AGS 577, Slide 60036, O24/3.

P. Steemans et al. / Geobios 41 (2008) 263–282280
Palynological research in the Proterozoic and Phanerozoic of oil and gasbearing regions of USSR. Trudy Vsesoluznogo Naucho-Issledovate skogoGeologoazvedochnogo Neftianogo Instituta, Kama Branch 217, pp. 26–46(in Russian).
Beju, D., 1967. Quelques spores, acritarches et chitinozoaires d’âge Dévonieninférieur de la plate-forme moesienne (Roumanie). Review of Palaeobotanyand Palynology 5, 39–49.
Breuer, P., Stricanne, L., Steemans, P., 2005. Morphological variation within amiospore population belonging to the genus Emphanisporites McGregor,1961, Lower Devonian, ‘‘la Gileppe’’ outcrop, Belgium. Geological Maga-zine 142, 241–253.
Burgess, N.D., Richardson, J.B., 1995. Late Wenlock to Early Pridoli cryp-tospores and miospores from south and southwest Wales, Great Britain.Palaeontographica B 236, 1–44.
Cramer, F.H., Díez, M.D.C.R., Rodríguez, R.M., Fombella, M.A., 1976. Acri-tarcos de la Formación San Pedro (Silúrico superior) de Torrestio, Provinciade León, España. Revista Española de Micropaleontología 8, 439–452.
Cramer, F.H., Rodríguez, R.M., 1977. Robledo and Arroyacas Formation(Arroyo de las Arroyacas, Province of Palencia, Spain) palynologicallydated as Late Silurian. Breviora Geologica Asturica 21, 2–4.
Dino, R., Rodrigues, M.A.C., 1995. Palinomorfos eodevonianos da FormaçãoFurnas – Bacia do Paraná. Anais da Academia Brasileira de Ciências 67,107–116.
Gerrienne, P., Bergamaschi, S., Pereira, E., Rodrigues, M.A.C., Steemans, P.,2001. An Early Devonian flora, including Cooksonia from the Paraná Basin(Brazil). In: Gerrienne, P. (Ed.), Early land plants evolution and diversifica-tion. Review of Palaeobotany and Palynology, 116. pp. 19–38.
Grahn, Y., 1999. Recent progress in Silurian and Devonian chitinozoan bios-tratigraphy of the Paraná Basin in Brazil and Paraguay, in: Rodrigues,M.A.C., Pereira, E., (Eds.), Ordovician-Devonian palynostratigraphy inWestern Gondwana: update, problems and perspectives. Univeridade doEstado do Rio de Janeiro, Rio de Janeiro, pp. 147–163.
Grahn, Y., Loboziak, S., Melo, J.H.G., 2003. Integrated correlation of LateSilurian (Pridoli s.l.) - Devonian chitinozoans and miospores in the Soli-mões Basin, northern Brazil. Acta Geologica Polonica 53, 283–300.
Grahn, Y., Melo, J.H.G., 2002. Chitinozoan biostratigraphy of the Late Devonianformations in well Caima PH-2, Tapajós River area, Amazonas Basin,northern Brazil. Review of Palaeobotany and Palynology 118, 115–139.
Grahn, Y., Melo, J.H.G., 2003. Silurian-Devonian chitinozoan biostratigraphyalong the Urubu, Uatumã and Abacate rivers in the western part of theAmazonas Basin, northern Brazil. Bulletin of Geosciences 78, 373–
391.Grahn, Y., Melo, J.H.G., Steemans, P., 2005. Contribution to the integrated
chitinozoan and miospore biostratigraphy of the Serra Grande Group(Silurian - Lower Devonian) in the Parnaíba Basin, northeast Brazil. RevistaEspañola de Micropaleontología 37, 183–204.
Grahn, Y., Pereira, E., Bergamaschi, S., 2000. Silurian and Lower Devonianchitinozoan biostratigraphy of the Paraná Basin in Brazil and Paraguay.Palynology 24, 147–176.
Grenfell, H.R., 1995. Probable fossil zygnematacean algal spore genera.Review of Palaeobotany and Palynology 84, 201–220.
Hart, G.F., 1964. Chomotriletes from the Lower Permian of South Africa.Annales of Geological Survey of South Africa 3, 149–157.
Jaglin, J.C., Paris, F., 2002. Biostratigraphy, biodiversity and palaeogeographyof Late Silurian chitinozoans from A1-61 borehole (north-western Libya).In: Steemans, P., Servais, T., Streel, M. (Eds.), Palaeozoic Palynology: aspecial issue in honour of Dr Stanislas Loboziak. Review of Palaeobotanyand Palynology 118, pp. 335–358.
Jardiné, S., Combaz, A., Magloire, L., Peniguel, G., Vachey, G., 1972. Acritar-ches du Silurien terminal et du Dévonien du Sahara Algérien. SeptièmeCongrès International de Stratigraphie et de Géologie du Carbonifère,Krefeld, 1969, C.R. 1, 295–311.
Jardiné, S., Combaz, A., Magloire, L., Peniguel, G., Vachey, G., 1974. Dis-tribution stratigraphique des acritarches dans le Paléozoïque du Saharaalgérien. Review of Palaeobotany and Palynology 18, 99–129.
Jardiné, S., Yapaudjian, L., 1968. Lithostratigraphie et palynologie du Dévo-nien-Gothlandien gréseux du Bassin de Polignac (Sahara). Revue del’Institut Français du Pétrole 23, 439–469.
Lange, F.W., 1967. Subdivisão bioestratigráfica e revisão da coluna siluro-devoniana da Bacia do baixo Amazonas. Atas do Simpósio sôbre a BiotaAmazônica 1 (Geociências) 1, 215–326.
Lavender, K., Wellman, C.H., 2002. Lower Devonian spore assemblages fromthe Arbuthnott Group at Canterland Den in the Midland Valley of Scotland.In: Steemans, P., Servais, T., Streel, M. (Eds.), Palaeozoic Palynology: aspecial issue in honour of Dr Stanislas Loboziak. Review of Palaeobotanyand Palynology 118, pp. 157–180.
Le Hérissé, A., 2002. Paleoecology, biostratigraphy and biogeography of LateSilurian to Early Devonian acritarchs and prasinophycean phycomata in wellA1-61, Western Libya, North Africa. In: Steemans, P., Servais, T., Streel, M.(Eds.), Palaeozoic Palynology: a special issue in honour of Dr StanislasLoboziak. Review of Palaeobotany and Palynology 118, pp. 359–395.
Le Hérissé, A., Rubinstein, C.V., Steemans, P., 1997. Lower Devonian paly-nomorphs from the Talacasto Formation, Cerro del Fuerte Section, San JuanPrecordillera, Argentina. Acta Universitatis Carolinae - Geologica 40(1996), 497–515.
Limachi, R., Goytia, V.H., Sarmiento, D., Arispe, O., Montecinos, R., Díaz, E.,Dalenz, A., Liachenko, N., Perez, M.E.A., 1996. Estratigrafía, geoquímica,correlaciones, ambientes sedimentarios y bioestratigrafía del Silúrico-Devónico de Bolivia. Memorias del XII Congreso Geológico de Bolivia,Tarija, 183–197.
McGregor, D.C., 1973. Lower and Middle Devonian spores of Eastern Gaspé,Canada. Palaeontographica B 142, 1–77.
McGregor, D.C., 1977. Lower and Middle Devonian spores of Eastern Gaspe,Canada. II Biostratigraphy. Palaeontographica B 163, 111–142.
McGregor, D.C., 1984. Late Silurian and Devonian spores from Bolivia.Academia Nacional de Ciencias Cordoba 69, 1–57.
McGregor, D.C., Camfield, M., 1976. Upper Silurian? to Middle Devonianspores of the Moose River Basin, Ontario. Geological Survey of Canada,Bulletin 263, 1–63.
Melo, J.H.G., 2000. Palynological evaluation and correlation of some Silurian-Devonian sections of southern Bolivia. XIV Congreso Geológico Boliviano,La Paz, 123–141.
Melo, J.H.G., 2005. Palynostratigraphy of some Paleozoic rock units of Bolivia:additional results. VI CONEXPLO – Congreso de Exploración y Desarrollode Hidrocarburos, Mar del Plata, IAPG/AAGGP CD-Rom, Trabajos Técni-cos, 20.
Melo, J.H.G., Loboziak, S., 2003. Devonian-Early Carboniferous miosporebiostratigraphy of the Amazon Basin, northern Brazil. Review of Palaeo-botany and Palynology 124, 131–202.
Moreau-Benoit, A., 1976. Les spores et débris végétaux. In: Lardeux, H. (Ed.),Les Schistes et Calcaires éodévoniens de Saint-Cénéré (Massif armoricain,France). Mémoires de la Société géologiques et minéralogique de Breta-gne19, pp. 27–58.
Moreau-Benoit, A., 1980. Les spores et débris végétaux. In: Plusquellec, Y.(Ed.), Les schistes et calcaires de ’. Mémoires de la Société géologiques etminéralogique de Bretagne 23, pp. 59–78.
Moreau-Benoit, A., 1994. Les spores des Grès de Landevennec, dans la coupede Lanvéoc, Lochkovien du Massif Armoricain, France. Revue de Micro-paléontologie 37, 75–93.
Moreau-Benoit, A., Dubreuil, M., 1987. Confirmation et découverte du Dévo-nien inférieur par la palynoplanctonologie dans les schistes et grès de laterminaison orientale du bassin d’Ancenis (sud-est du Massif armoricain).Géologie de la France 1, 37–54.
Moreau-Benoit, A., Poncet, J., 1982. Les schistes et calcaires éodévoniens deNéhou dans la coupe de Barneville (nord-est du Massif armoricain).Sédimentologie, palynologie, planctonologie, stratigraphie. Bulletin duBureau de Recherches géologiques et minières 1, 81–114.
Norris, G., 1965. Triassic and Jurassic miospores and acritarchs from theBeacon and Ferrar Groups, Victoria Land, Antarctica. New Zealand Journalof Geology and Geophysics 8, 236–277.
Playford, G., Dring, R.S., 1981. Late Devonian acritarchs from the CarnarvonBasin, Western Australia. Special Papers in Palaeontology 27, 1–78.
Racheboeuf, P.R., Le Hérissé, A., Paris, F., Babin, C., Guillocheau, F., Truyols-Massoni, M., Suárez-Soruco, R., 1993. Le Dévonien de Bolivie : bios-tratigraphie et chronostratigraphie. Comptes Rendus de l’Académie desSciences de Paris 317 (2), 602–795.

P. Steemans et al. / Geobios 41 (2008) 263–282 281
Rauscher, R., Robardet, M., 1975. Les microfossiles (acritarches, chitinozoaireset spores) des couches de passage du Silurien au Dévonien dans le Cotentin(Normandie). Annales de la Société Géologique du Nord 115, 81–92.
Raymond, A., Gensel, P.G., Stein, W.E., 2006. Late Silurian phytogeography.In: Gerrienne, P. (Ed.), Recent advances in Palaeozoic Paleobotany. Reviewof Palaeobotany and Palaeopalynology 142, pp. 165–192.
Richardson, J.B., 1967. Some British Lower Devonian assemblages and theirstratigraphic significance. Review of Palaeobotany and Palynology 1, 111–
129.Richardson, J.B., Edwards, D., 1989. Sporomorphs and plant megafossils. In:
Holland, C.H., Bassett, M.G. (Eds.), A global standard for the Siluriansystem. National Museum of Wales Geological Series, Cardiff 9, pp. 216–
226.Richardson, J.B., Ford, J.H., Parker, F., 1984. Miospores, correlation and age of
some Scottish Lower Old Red Sandstone sediments from the Strathmoreregion (Fife and Angus). Journal of Micropalaeontology 3, 109–124.
Richardson, J.B., Ioannides, N.S., 1973. Silurian palynomorphs from theTanezzuft and Acacus formations, Tripolitania, North Africa. Micropaleon-tology 19, 257–307.
Richardson, J.B., Lister, T.R., 1969. Upper Silurian and Lower Devonian sporeassemblages from the Welsh Borderland and South Wales. Palaeontology12, 201–252.
Richardson, J.B., McGregor, D.C., 1986. Silurian and Devonian spore zones ofthe Old Red Sandstone continent and adjacent regions. Geological Survey ofCanada, Bulletin 364, 1–79.
Richardson, J.B., Rasul, S.M., Al-Ameri, T., 1981. Acritarchs, miospores andcorrelation of the Ludlovian-Downtonian and Silurian-Devonian bounda-ries. Review of Palaeobotany and Palynology 34, 209–224.
Richardson, J.B., Rodríguez, R.M., Sutherland, S.J.E., 2001. Palynologicalzonation of Mid- Palaeozoic sequences from the Cantabrian Mountains NWSpain: implications for interregional and interfacies correlation of theLudford/Prídolí and Silurian/Devonian boundaries, and plant dispersalpatterns. Bulletin Natural History Museum London (Geology) 57, 115–
162.Richardson, J.B., Streel, M., Hassan, A., Steemans, P., 1982. A new spore
assemblage to correlate between the Breconian (British Isles) and theGedinnian (Belgium). Annales de la Société Géologique de Belgique105, 135–143.
Roche, M., Sabir, M., Steemans, P., Vanguestaine, M., 1986. Palynologie dela région et du sondage de Willerzie. Aarkundige Mededelingen 3,149–190.
Rodríguez, R.M., 1978a. Nuevas mioesporas de la Formación San Pedro enGeras de Gordon, Cordillera Cantabrica (Provincia de León, Noroeste deEspaña). Breviora Geologica Asturica 22, 9–16.
Rodríguez, R.M., 1978b. Mioesporas de la Formación San Pedro/Furada(Silúrico Superior-Devónico Inferior), Cordillera Cantabrica, NO deEspaña. Palinología Número Extraordinario 1, 407–433.
Rodríguez, R.M., 1978c. Miospores de la Formation San Pedro (Silurien-Dévonien) à Corniero (Province de León, Espagne). Revue de Micropa-léontologie 20, 216–221.
Rodríguez, R.M., 1983. Palinología de las Formaciones del Silúrico superior –
Devonico inferior de la Cordillera Cantabrica. Servicio de Publicaciones,Universidad de León, 1–231.
Rossignol, M., 1962. Analyse pollinique de sédiments marins Quaternaires enIsraël. II - Sédiments pléistocènes. Pollen et Spores 4, 121–148.
Rubinstein, C.V., Brussa, E.D., 1999. A palynomorph and graptolite biostrati-graphy of the Central Precordillera Silurian Basin, Argentina. In: Tongiorgi,M., Playford, G. (Eds.), Studies in Palaeozoic Palynology, Selected papersfrom the CIMP Symposium at Pisa. Bollettino della Società PaleontologicaItaliana, Modena 38, pp. 257–266.
Rubinstein, C.V., Melo, J.H.G., Steemans, P., 2005. Lochkovian (EarliestDevonian) miospores from the Solimões Basin, northern Brazil. Reviewof Palaeobotany and Palynology 133, 91–113.
Rubinstein, C.V., Steemans, P., 2002. Miospore assemblages from the Silurian-Devonian boundary, in borehole A1-61, Ghadamis Basin, Libya. In: Stee-mans, P., Servais, T., Streel, M. (Eds.), Paleozoic Palynology: a special issuein honour of Dr. Stanislas Loboziak. Review of Palaeobotany and Palyno-logy 118, pp. 397–421.
Rubinstein, C.V., Steemans, P., 2007. New palynological data from the Devo-nian Villavicencio Formation, Precordillera of Mendoza, Argentina. Ame-ghiniana 44, 3–9.
Rubinstein, C.V., Steemans, P., Astini, R.A., 2006. Silurian marine and terres-trial palynomorphs of Quebrada Ancha, Central Precordillera Basin, SanJuan Province, Argentina. XIII Simposio Argentino de Paleobotánica yPalinología, Bahia Blanca, May 22–25.
Scotese, C.R., Boucot, A.J., McKerrow, W.S., 1999. Gondwanan palaeogeo-graphy and palaeoclimatology. Journal of African Earth Sciences 28, 99–
114.Steemans, P., 1981. Étude stratigraphique des spores dans les couches de
transition « Gedinnien-Siegenien » à Nonceveux et à Spa (Belgique).Annales de la Société Géologique de Belgique 104, 41–59.
Steemans, P., 1982a. Gedinnian and Siegenian spores stratigraphy in Belgium.In: Ziegler, W., Werner, R. (Eds.), On Devonian stratigraphy and paleonto-logy of the Ardenno-Rhenish Mountains and related Devonian matters.-Courier Forschungsinstitut Senckenberg55, 165–180.
Steemans, P., 1982b. L’âge du poudingue de Fépin (base du Gedinnien) àLahonry (Belgique). Bulletin de la Société belge de Géologie (1981) 90,331–340.
Steemans, P., 1989. Palynostratigraphie de l’Eodévonien dans l’ouest del’Europe. Mémoires Explicatifs pour les Cartes Géologiques and Minéra-logiques de la Belgique, Bruxelles 27, 1–453.
Steemans, P., 1995. Silurian and Lower Emsian spores in Saudi Arabia. Reviewof Palaeobotany and Palynology 89, 91–104.
Steemans, P., Al-Ruwaili, M., Filatoff, J., Streel, M., 2001. A miospore assem-blage at the Siluro-Devonian boundary from the Tawil Formation, QatifField, Saudi Arabia. First meeting of the CIMP, Spores and Pollen Sub-commission, Cork, 2–7 September 2001, National University of Ireland, 43.
Steemans, P., Debbaut, V., Faber, A., 2000. Preliminary survey of the palyno-logical content of the Lower Devonian in the Oesling, Luxembourg. Bulletinde la Société des Naturalistes Luxembourgeois 100, 171–186.
Steemans, P., Gerrienne, P., 1984. La micro- et macroflore du Gedinnien de laGileppe, Synclinorium de la Vesdre, Belgique. Annales de la SociétéGéologique de Belgique 107, 51–71.
Steemans, P., Lakova, I., 2004. The Moesian Terrane during the Lochkovian - Anew palaeogeographic and phytogeographic hypothesis based on miosporeassemblages. Palaeogeography, Palaeoclimatology, Palaeoecology 208,225–233.
Steemans, P., Le Hérissé, A., Bozdogan, N., 1996. Ordovician and Siluriancryptospores and miospores from Southeastern Turkey. Review of Palaeo-botany and Palynology 93, 35–76.
Streel, M., Fairon-Demaret, M., Gerrienne, P., Loboziak, S., Steemans, P., 1990.Lower and Middle Devonian miospore-based stratigraphy in Libya and itsrelation to the megafloras and faunas. Review of Palaeobotany and Paly-nology 66, 229–242.
Streel, M., Fairon-Demaret, M., Otazo-Bozo, N., Steemans, P., 1981. Étudestratigraphique des spores du Dévonien inférieur au bord sud du Syncli-norium de Dinant (Belgique) et leurs applications. Annales de la SociétéGéologique de Belgique 104, 171–173.
Streel, M., Higgs, K., Loboziak, S., Riegel, W., Steemans, P., 1987. Sporestratigraphy and correlation with faunas and floras in the type marineDevonian of the Ardenno-Rhenish regions. Review of Palaeobotany andPalynology 50, 211–229.
Suárez-Soruco, R., 1989. El Ciclo Cordillerano (Silúrico-Carbonífero Inferior)en Bolivia y su relación con países limítrofes. Revista Técnica de YPFB 10,233–243.
Suárez-Soruco, R., Lobo-Boneta, J., 1983. La fase compresiva eohercínica en elsector oriental de la Cuenca Cordillerana de Bolivia. Revista Técnica deYPFB 9, 189–207.
Torsvik, T.H., Cocks, L.R.M., 2004. Earth geography from 400 to 250 Ma: apalaeomagnetic, faunal and facies review. Journal of the Geological Society161, 555–572.
Turnau, E., 1986. Lower to Middle Devonian spores from the vicinity of Pionki(Central Poland). Review of Palaeobotany and Palynology 46, 311–354.
Turnau, E., Jakubowska, L., 1989. Early Devonian miospores and age of theZwolen Formation (Old Red Sandstone facies) from Ciepielów IG-1borehole. Annales Societatis Geologorum Poloniae 59, 391–416.

P. Steemans et al. / Geobios 41 (2008) 263–282282
Turnau, E., Matyja, H., 2001. Timing of the onset of Devonian sedimentation inNorthwestern Poland: palynological evidence. Annales Societatis Geolo-gorum Poloniae 71, 67–74.
Vanguestaine, M., Steemans, P., Streel, M., 1986. Microfossiles végétaux. In:Racheboeuf, P.R. (Ed.), le Groupe de Liévin, Pridoli-Lochkovien de l’Artois(N. France). Biostratigraphie du Paléozoïque, Mémoire de la Sociétégéologique et Minière de Bretagne 3, pp. 47–55.
Vavrdová, M., Bek, J., Dufka, P., Isaacson, P.E., 1996. Palynology of theDevonian (Lochkovian to Tournaisian) sequence, Madre de Dios Basin,northern Bolivia. Vestník Ceského geologického ústavu 71, 333–349.
Wellman, C.H., 1993a. A land plant microfossil assemblage of Mid Silurian agefrom the Stonehaven Group, Scotland. Journal of Micropalaeontology 12,47–66.
Wellman, C.H., 1993b. A Lower Devonian sporomorph assemblage from theMidland Valley of Scotland. Transactions of the Royal Society of Edin-burgh. Earth Sciences 84, 117–136.
Wellman, C.H., Richardson, J.B., 1993. Terrestrial plant microfossils fromSilurian inliers of the Midland Valley of Scotland. Palaeontology 36, 155–
193.Wellman, C.H., Habgood, K., Jenkins, G., Richardson, J.B., 2000. A new
assemblage (microfossil and megafossil) from the Lower Old Red Sands-tone of the Anglo-Welsh Basin: its implications for the palaeoecology ofearly terrestrial ecosystems. Review of Palaeobotany Palynology 109, 161–
196.Wellman, C.H., Richardson, J.B., 1996. Sporomorph assemblages from the
‘‘Lower Old Red Sandstone’’ of Lorne Scotland. In: Cleal, C.J. (Ed.),Studies on early land plant spores from Britain. Special Papers in Palaeon-tology 55, pp. 41–101.
Wellman, C.H., Thomas, R.G., Edwards, D., Kenrick, P., 1998. The Coshes-ton Group (Lower Old Red Sandstone) in southwest Wales: age, correla-tion and palaeobotanical significance. Geological Magazine 135,397–412.
Related Documents











