Significantly skewed memory CD8 + T cell subsets in HIV-1 infected infants during the first year of life 1 Nazma Mansoor 2,a , Brian Abel 2,a , Thomas J. Scriba 2 , Jane Hughes 2 , Marwou de Kock 2 , Michele Tameris 2 , Sylvia Mlenjeni 2 , Lea Denation 2 , Francesca Little 3 , Sebastian Gelderbloem 2 , Anthony Hawkridge 2 , W. Henry Boom 4 , Gilla Kaplan 5 , Gregory D. Hussey 2 , and Willem A. Hanekom 2 2 South African Tuberculosis Vaccine Initiative, Institute of Infectious Diseases and Molecular Medicine and School of Child and Adolescent Health, University of Cape Town, Anzio Road, Observatory, 7925, South Africa 3 Department of Statistical Sciences, University of Cape Town, Rondebosch, South Africa 4 Tuberculosis Research Unit, Case Western Reserve University and University Hospitals Case Medical Center, Cleveland, Ohio, USA 5 Public Health Research Institute, University of Medicine and Dentistry of New Jersey, Newark, New Jersey, USA Abstract HIV-1 infection causes a severe T cell compromise; however, little is known about changes in naïve, memory, effector and senescent T cell subsets during the first year of life. T cell subsets were studied over the first year of life in blood from 3 infant cohorts: untreated HIV-infected, HIV-exposed but uninfected, and HIV-unexposed. In HIV-infected infants, the frequency of CCR7 + CD45RA + naïve CD8 + T cells was significantly decreased, whilst the frequency of CCR7 − CD45RA − effector memory CD8 + T cells was increased, compared with the control cohorts. A larger population of CD8 + T cells in HIV-infected infants displayed a phenotype consistent with senescence. Differences in CD4 + T cell subset frequencies were less pronounced, and no significant differences were observed between exposed and unexposed HIV-uninfected infants. We concluded that the proportion of naïve, memory, effector and senescent CD8 + T cells during the first year of life is significantly altered by HIV-1 infection. Keywords CD4; CD8; memory; HIV-1; infants 1 Competing interests: The authors declare no conflicts of interest. Correspondence should be addressed to Willem A. Hanekom, University of Cape Town Health Sciences, Anzio Road, Observatory, 7925, South Africa, Tel. +27 21 406 6080, Fax. +27 21 406 6081, email: [email protected]. a Nazma Mansoor and Brian Abel contributed equally to this work. This work has been presented, in part, at the 13 th International Congress of Immunology in Rio de Janeiro, Brazil, 21–25 August 2007. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript Clin Immunol. Author manuscript; available in PMC 2010 March 1. Published in final edited form as: Clin Immunol. 2009 March ; 130(3): 280–289. doi:10.1016/j.clim.2008.09.006. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Significantly skewed memory CD8+ T cell subsets in HIV-1 infectedinfants during the first year of life1
Nazma Mansoor2,a, Brian Abel2,a, Thomas J. Scriba2, Jane Hughes2, Marwou de Kock2,Michele Tameris2, Sylvia Mlenjeni2, Lea Denation2, Francesca Little3, SebastianGelderbloem2, Anthony Hawkridge2, W. Henry Boom4, Gilla Kaplan5, Gregory D. Hussey2,and Willem A. Hanekom22 South African Tuberculosis Vaccine Initiative, Institute of Infectious Diseases and MolecularMedicine and School of Child and Adolescent Health, University of Cape Town, Anzio Road,Observatory, 7925, South Africa3 Department of Statistical Sciences, University of Cape Town, Rondebosch, South Africa4 Tuberculosis Research Unit, Case Western Reserve University and University Hospitals CaseMedical Center, Cleveland, Ohio, USA5 Public Health Research Institute, University of Medicine and Dentistry of New Jersey, Newark,New Jersey, USA
AbstractHIV-1 infection causes a severe T cell compromise; however, little is known about changes in naïve,memory, effector and senescent T cell subsets during the first year of life. T cell subsets were studiedover the first year of life in blood from 3 infant cohorts: untreated HIV-infected, HIV-exposed butuninfected, and HIV-unexposed. In HIV-infected infants, the frequency of CCR7+CD45RA+ naïveCD8+ T cells was significantly decreased, whilst the frequency of CCR7−CD45RA− effector memoryCD8+ T cells was increased, compared with the control cohorts. A larger population of CD8+ T cellsin HIV-infected infants displayed a phenotype consistent with senescence. Differences in CD4+ Tcell subset frequencies were less pronounced, and no significant differences were observed betweenexposed and unexposed HIV-uninfected infants. We concluded that the proportion of naïve, memory,effector and senescent CD8+ T cells during the first year of life is significantly altered by HIV-1infection.
KeywordsCD4; CD8; memory; HIV-1; infants
1Competing interests: The authors declare no conflicts of interest.Correspondence should be addressed to Willem A. Hanekom, University of Cape Town Health Sciences, Anzio Road, Observatory,7925, South Africa, Tel. +27 21 406 6080, Fax. +27 21 406 6081, email: [email protected] Mansoor and Brian Abel contributed equally to this work.This work has been presented, in part, at the 13th International Congress of Immunology in Rio de Janeiro, Brazil, 21–25 August 2007.Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptClin Immunol. Author manuscript; available in PMC 2010 March 1.
Published in final edited form as:Clin Immunol. 2009 March ; 130(3): 280–289. doi:10.1016/j.clim.2008.09.006.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

BackgroundAcquired cell-mediated immunity (CMI) is critical for control and clearance of many humaninfections [1]. The main mediators of CMI, CD4+ and CD8+ T cells, exist in the peripheralcirculation either as naïve or as antigen-experienced cells. The latter consist of heterogenouspopulations that may be classified according to immediate or delayed effector function,expression of homing receptors, cytokine secretion profile, proliferation potential andlongevity or survival characteristics [2;3]. Effector memory T cells (TEM) are characterised bythe surface phenotype CD45RA−CCR7−CD62Llo. Following antigenic challenge, these cellsmigrate to inflamed tissues and demonstrate potent effector function. They proliferate poorlyand are short-lived. TEM that re-express CD45RA are postulated to be terminally differentiated,and termed TEMRA [2;3]. By contrast, central memory T cells (TCM), characterised by thesurface phenotype CD45RA−CCR7+CD62Lhi, are long-lived in the host, home to lymph nodes,and proliferate efficiently to differentiate into effector cells [4;5;6;7]. T cells may be furtherclassified by expression of CD28, a costimulatory molecule which is associated with betterfunctional capacity of the cells [8], and CD57, which is thought to be a marker of replicativesenescence [9].
HIV infection results in severe disruption and depletion of the mucosal, lymphatic andperipheral T lymphocyte compartments. Systemic changes in the entire CD4+ and CD8+ T cellpopulations have been reported in HIV-1 infected adults [10;11;12], children[13;14;15], andinfants[13;15;16]. An increased activation state of these T cells occurs during HIV infection[17;18] and may be directly linked to T cell differentiation [19]. Adult HIV-1 infection ischaracterised by a loss of naïve CD4+ and CD8+ T cells and an expansion of CD27− and/orCD28− T cells, and similar observations have been reported for HIV-infected infants andchildren[14;20;21]. Further, a skewing in T cell phenotype from TCM to TEM (loss of CCR7and CD62L expression) has been reported in HIV infected adults and children [22], whileCCR7 expression has been shown to correlate inversely with HIV-1 viral load [23;24].
Limited knowledge exists about longitudinal phenotypic changes during the first year of lifein peripheral blood T cells from antiretroviral treatment (ART)-naïve infants perinatallyinfected with HIV-1, and from HIV-uninfected infants born to infected mothers. HIV-infectedinfants have a ten-fold increase in mortality, compared with HIV-unexposed infants. This ispartly because infection prior to the full development of the immune system results indevastating destruction [25;26], and opportunistic infections may be lethal. Importantly, infantsexposed to HIV-1 but not infected have mortality rates up to three times greater than thoseborn to HIV-uninfected mothers [27]. The severity of maternal HIV disease has been linkedto increased mortality and morbidity in uninfected infants. Although environmental factorsmay contribute significantly to this increased mortality, immunological factors may alsocontribute, as immunological differences between exposed, uninfected infants and unexposedinfants have been shown: high level T cell activation has been reported in uninfected neonatesborn to infected mothers [13;28;29;30].
Our aim was to compare the phenotype and differentiation status of naïve and memory T cellpopulations longitudinally during the first year of life in infants infected by HIV, exposed toHIV but not infected, and in unexposed infants. We show that CD8+ T cells derived from HIV-infected infants were markedly skewed towards an effector phenotype, rather than thepredominant naïve phenotype observed in uninfected infants. Moreover, a greater frequencyof CD8+ T cells from HIV-infected infants displayed a senescent phenotype, compared withCD8+ T cells from uninfected infants.
Mansoor et al. Page 2
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

MethodsStudy population
Infants born to HIV-infected and non-infected women from the Worcester region of theWestern Cape, South Africa, were enrolled before the age of 3 months. HIV-1 infection statuswas determined in infants by PCR, and each was classified into 1 of 3 groups: non-HIV-infectedborn to a non-HIV-infected mother (HIV−), non-HIV-infected born to an HIV-infected mother(Exposed HIV−), or HIV-infected (HIV+). During the study period, roll-out of antiretroviraltherapy (ART) as part of the mother-to-child HIV transmission prevention programmecommenced. Toward the end of the project, roll-out of ART for treatment pediatric HIV-1infection was also initiated. Infants who received ART were excluded from the analysis.
The protocol was approved by the Research Ethics Committee of the University of Cape Town,and by the Institutional Review Board of the University of Medicine and Dentistry of NewJersey. Good clinical practice and ethical guidelines of the US Department of Health andHuman Services and the South African Medical Research Council were followed in the conductof the project; this included written informed parental consent.
Blood collectionEDTA-anticoagulated peripheral blood was collected from the infants at 3, 6, 9, and 12 monthsof age. In addition to the immunological analysis reported here, a complete blood count andHIV-1 viral load (EasyQ HIV1, version1.2 bioMérieux NucliSens®) were performed at eachtimepoint.
Flow cytometric and hematology analysisWhole blood was transported at room temperature to the laboratory and processed within 4hours of collection. Fifty microlitres of whole blood were incubated with fluorescence-conjugated antibodies against CD3-PE, CD4-allophycocyanin, CD8-PerCP, CD45-FITC,CD45RA-FITC, CCR7-PE, CD62L-PE, CD28-PE, and CD57-PE (all obtained from BDBiosciences), in different combinations, at room temperature for 30 minutes. Red cells werethen lysed with FACS lysing solution (BD Biosciences), and white cells fixed in 1% para-formaldehyde. All cells were acquired on a FACSCalibur flow cytometer (BD Biosciences),equipped for four colour detection, using Cellquest (BD Biosciences). Analysis was completedusing Flowjo (Treestar) software. A complete blood count was performed on all infants,allowing calculation of absolute numbers of CD4+ and CD8+ T cell subsets, as CD45 was usedin flow cytometric panels to delineate total lymphocyte populations. A rigorous gating strategywas employed which was consistently applied to all participants. All antibodies in the panelswere optimally titrated before use in the study.
Statistical considerationsThe nonparametric Kruskal-Wallis test, with Dunn’s Multiple Comparison Test, were used toevaluate differences between study groups, and these statistical tests were performed usingPrism 4.03 (GraphPad). A mixed effects linear regression model was used to compare the timeprofiles of the CD8 T cell populations (Supplementary Figure 5). Maximum likelihoodestimation was used so that the models coped with missing data arising from infant drop-outbecause observations at each time point influences estimates of treatment or other effects atevery other time point due to the specification of the covariance pattern[31]. Where data wasnot normally distributed, values were log transformed.
Mansoor et al. Page 3
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ResultsParticipant characteristics
Among 298 infants born to HIV+ mothers enrolled, 20 were ultimately identified as HIV+ bya positive viral amplification test. The number of patients in the HIV+ group decreasedgradually over the 12 month period (Table I): 8 infants died early during the study, conductedduring a period when ART was not routinely available in South Africa, and 4 infants were lostto follow-up. Three infants received ART, resulting in exclusion from analysis. Twenty fiveof the 278 non-infected infants born to HIV-infected mothers were included in the “ExposedHIV−“ group. Eleven of these infants were lost to follow-up over their first year or life. Twentythree infants born to HIV-negative mothers (termed “HIV−“) were also enrolled. Of these, 4were ultimately lost to follow-up. At 3 months of age, blood of only 16 of 20 HIV+ infants wasexamined.
Total, CD4 and CD8 lymphocyte counts and viral loadThere were no differences in total lymphocyte counts between the groups, at all time points(Table I). The frequencies of CD4+ lymphocytes in the HIV+ group were lower than in theother groups (Figure IA), while absolute numbers differed between the HIV+ and the HIV−
groups at 6 months of age (Table I). The frequencies of CD8+ T cells were significantly higherin the HIV+ group, compared with the other 2 groups (Figure IB), and absolute numbers werehigher in HIV+ infants than in the other 2 groups at 3 months. As expected, the ratios ofCD4+ to CD8+ T cell frequencies were significantly reduced in the HIV+ group, comparedwith the other groups (Figure IC).
Participants from the HIV+ group had a detectable plasma HIV-1 viral load, which showed acharacteristic gradual decline over the first year of life (Table I).
Memory phenotype of CD8+ T cellsTo investigate the memory phenotype of CD8+ T cells, we determined the expression ofCD45RA and CCR7, or of CD45RA and CD62L, by flow cytometry (Figure II). The frequencyof naïve CD8+ T cells (CD8+ TNaïve; CD45RA+CCR7+) was approximately 3-fold lower inHIV+ infants at 3 months of age, compared with infants from the other 2 groups (Figure IIC;Supplementary Figure IA). This frequency decreased in all groups over the next 9 months, butHIV+ infants had consistently lower frequencies throughout (Figure IIC, Supplementary FigureIA). Differences between groups were not as prominent when the absolute number of CD8+
TNaïve cells was evaluated (Figure IIE; Supplementary Figure IIA); however, the HIV− grouphad an increased CD8+ TNaïve subset, compared with the Exposed HIV− group, at 6 and 12months. Very similar patterns were observed when CD62L was substituted for CCR7 in theanalysis (data not shown).
The frequency of effector memory CD8+ T cells (CD8 TEM; CD45RA−CCR7−) wassignificantly higher in the HIV+ group at all time points, compared with the other 2 groups,and increased over time in all groups (Figure IID, left panel, Supplementary Figure IB). Theabsolute numbers of CD8+ TEM cells followed a similar pattern (Figure IIF left panel;Supplementary Figure IIB). A mixed effects linear regression modelling of this data alsodemonstrated significant differences in the naïve and effector CD8 T cell subsets between theHIV-infected group and the control groups (Supplementary Figure 5). At 3 months of age,HIV+ infants also had a significantly higher frequency of CD8+ TEM cells that re-expressedCD45RA (TEM45RA), compared with the Exposed HIV− group (Figure IID middle panel,Supplementary Figure IC). At later time points there were no differences in the frequency ofthese cells for the 3 groups, but absolute numbers were higher in HIV+ infants throughout the
Mansoor et al. Page 4
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

first year of life (Figure IIF middle panel; Supplementary Figure IIC). Again, when CD62Lwas substituted for CCR7, similar patterns were observed (data not shown).
The frequencies of central memory CD8+ T cells (TCM) in the 3 groups, were extremely lowwhen CCR7 was used to define this subset (CD45RA−CCR7+) (Figure IID, right panel,Supplementary Figures ID). When CD62L was used instead of CCR7 to define TCM, increasedfrequencies and absolute numbers of CD8 TCM cells could be detected in the HIV+group,compared with the other 2 groups, over the first year of life (Figure IID, right panel;Supplementary Figures ID and IID).
Memory phenotype of CD4+ T cellsPhenotypic differences in CD4+ T cells were far less pronounced than in the CD8+ T cells,with only occasional differences that were significant between HIV+ and the HIV− infants(Figure III; Supplementary Figures III and IV). Interestingly, CCR7 appeared to be a bettermarker to differentiate subsets, compared with CD62L. There were no dramatic differencesbetween Exposed HIV− infants and HIV− infants in any of the CD4+ T cell subsets.
Expression of CD28 and CD57The absolute number of CD8+ T cells expressing the senescence marker CD57 was higher inthe HIV+ group at most time points, compared with the HIV− Exposed group. Surprisingly,expression of this marker tended to be higher in HIV− infants, compared with Exposed HIV−
infants (Figure IVA).
The absolute number of CD8+ T cells not expressing the costimulatory molecule CD28 wasmarkedly higher in HIV+ infants, compared with the others, at most time points (Figure IVB).CD57 and CD28 expression on CD4+ T cells was not different between the groups (data notshown).
DiscussionThis is a comprehensive report of changes in memory CD4+ and CD8+ T cell subsets inuntreated HIV-infected infants (HIV+), which were also compared with HIV-exposed butuninfected infants (Exposed HIV−), and with HIV-unexposed infants (HIV−). The data showskewing of predominantly CD8+ T cell populations in HIV-infected infants, compared withthe other infant groups.
As early as 3 months after birth, the frequency of naïve CD8+ T cells was markedly reducedin HIV+ infants. In HIV-infected adults, loss of naïve T cells is the result of increased immunedifferentiation, driven by immune activation, which is characteristic of HIV-1 infection [32;33;34;35]. In adults, the naïve CD4+ T cell compartment is more severely depleted than theCD8 compartment [11]. We found that loss of naïve CD8+ T cells was more prominent ininfants, whereas Mc Closkey et al. reported a significant loss of naïve CD4 and CD8 T cellsin the first year of life [14], and Gallagher et al. demonstrated a significant decrease in naïveCD8 T cells already at 6 weeks of age [36]. Naive T cell loss perturbs T cell homeostasis andconsequently, thymic output [11]. Clinical studies have shown that thymic dysfunction isassociated with rapid progression of disease in perinatally infected infants [25;37;38].
The numbers and proportions of effector memory CD8+ T cells (TEM) were significantlyincreased at all time points in HIV-infected infants. Such maturational skewing is welldescribed in HIV-infected adults [22;39]. The clonal expansion of antigen-specific effector Tcells is important for immunity against primary infections, whereas the development andmaintenance of memory T cell populations is critical for protective immunity againstreinfections. HIV-driven, non-specific maturation may thus be particularly detrimental to the
Mansoor et al. Page 5
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

development of immunological memory in infants. Our results contrast with findings of Jordan,et al., who failed to show changes in the distribution of memory and effector T cell populationsin children aged 2–14 [22]; however, the latter study investigated very small cohorts of 10HIV-infected and 4 uninfected children, and all HIV-infected children had received ART.
The CD8+ TEMRA population, effector memory cells that have re-expressed CD45RA, wasalso increased in HIV+ infants, compared with Exposed HIV− infants. This increase has alsobeen reported in adult HIV-1 infection [22], and may reflect chronic immune activation thatdrives cells into a state of terminal differentiation.
Expression of the costimulatory molecule CD28 and of the senescence marker CD57 were alsodifferent between our infant groups. CD8+ T cells of HIV-infected infants displayed aphenotype consistent with senescence, with low CD28 and elevated CD57 expression. HIV-specific CD8+ T cells detected very early after birth, in utero-infected infants [40;41;42], maytherefore be prone to senescence, and be less functional, especially in the absence of specificCD4+ T cell help [40;43;44]. These phenotypes have been shown to be prevalent in HIV−
infected adults with high viremia, where the senescent T cells have also shown reducedproliferative capacity [9;45]. Similarly, adults [46;47;48;49;50] and children [14;21] with highviral replication characteristically have CD8+ T cell downregulation of CD28, which leads todysregulation of normal T cell function. Interestingly, we found that HIV-exposed butuninfected infants expressed significantly lower levels of CD57 than HIV-1 unexposed infants.This result was surprising, since increased immune activation is expected in exposed HIV-uninfected infants [13;28;29;30], which should result in greater T cell senescence, comparedwith unexposed infants. Further study would be necessary to clarify this finding.
CCR7 and CD62L expression are often used interchangeably with CD45RA to define memorysubsets. We showed that in infants, significant differences between the expression of these 2markers do exist. For example, populations of central memory CD8+ T cells were detectableonly when CD62L was used, and CCR7 was a better marker to define effector memoryCD4+ T cells. We do not know why these markers were not equivalently useful to delineate Tcell subsets in infants, but this warrants further inspection, and these differences should betaken into account when measuring these subsets in infant studies.
The significant drop-out rates, primarily due to death of HIV-infected infants later in the firstyear of life, constituted a limitation of our study. These death rates were consistent with thoseobserved in other cohorts of African infants not on antiretroviral therapy [26]. We thereforealso assessed our results with maximum likelihood mixed effects linear regression modelling,which copes with loss to follow-up. These models showed results that were similar to thosewithout applying the models. In turn, sensitivity of the models were tested by applying threedifferent methods of imputation of missing data and comparing this with analysis of just thosepatients who had complete data: there was excellent agreement between the results of all fouranalyses (data not shown). Invariably, results from HIV-uninfected infants remained differentfrom HIV-infected infants. Additionally, we did not prospectively address covariates that couldhave had an effect on infant immunity, such as maternal viral load and immune status, maternalthereapy, infant nutritional status, and additional infections in infants. Regardless, the T cellphenotype in HIV-infected infants during the first year of life remained strikingly different tothe control groups. We therefore believe this study’s results are unique and completely novel,particularly as the current routine availability of antiretroviral therapy precludes this kind ofstudy from ever being repeated.
To conclude, the changes in T cell phenotype reported here suggest a gross dysfunction in Tcell development and maturation in HIV-infected infants. A reduction of CD4+ T cells andincrease in CD8+ T cells, with lower CD4/CD8 ratios, in HIV-infected infants is well
Mansoor et al. Page 6
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

documented [25]; however, we now demonstrate, in addition, that subsets of these cells arealso significantly altered. Furthermore, the observation that total lymphocyte counts betweenthe 3 groups are similar indicates that there is a re-distribution of the existing T cells from theCD4 to the CD8 compartment, and from naive T cells to effector T cells. In contrast, very littledifference between HIV exposed but uninfected and HIV-1 unexposed infants was observed.The findings from this study indicate that T cell memory subsets are severely disrupted in HIV-infected infants, and this may have dire implications on the efficacy of childhood vaccination,and may contribute to the increased morbidity and mortality of infected infants. Our novelresults also underscore the positive impact that early antiretroviral therapy may have inreversing the abnormal immune profile. This hypothesis is substantiated by recent findings ofmarkedly improved clinical outcome of infants who have started antiretroviral therapy at 6weeks of age, or at the earliest time point after HIV diagnosis, compared with infants who havestarted therapy at later ages [51].
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsThis work was supported by the Elizabeth Glaser Pediatric AIDS Foundation and the TBRU of the NIH (NO1-AI70022).
References1. Klenerman P, Hill A. T cells and viral persistence: lessons from diverse infections. Nat Immunol
2005;6:873–9. [PubMed: 16116467]2. Sallusto F, Geginat J, Lanzavecchia A. Central memory and effector memory T cell subsets: function,
generation, and maintenance. Annu Rev Immunol 2004;22:745–63. [PubMed: 15032595]3. Seder RA, Ahmed R. Similarities and differences in CD4+ and CD8+ effector and memory T cell
generation. Nat Immunol 2003;4:835–42. [PubMed: 12942084]4. Marshall DR, Turner SJ, Belz GT, Wingo S, Andreansky S, Sangster MY, Riberdy JM, Liu T, Tan M,
Doherty PC. Measuring the diaspora for virus-specific CD8+ T cells. Proc Natl Acad Sci U S A2001;98:6313–8. [PubMed: 11344265]
5. Masopust D, Vezys V, Marzo AL, Lefrancois L. Preferential localization of effector memory cells innonlymphoid tissue. Science 2001;291:2413–7. [PubMed: 11264538]
6. Reinhardt RL, Khoruts A, Merica R, Zell T, Jenkins MK. Visualizing the generation of memory CD4T cells in the whole body. Nature 2001;410:101–5. [PubMed: 11242050]
7. Wherry EJ, Teichgraber V, Becker TC, Masopust D, Kaech SM, Antia R, von Andrian UH, AhmedR. Lineage relationship and protective immunity of memory CD8 T cell subsets. Nat Immunol2003;4:225–34. [PubMed: 12563257]
8. Lloyd TE, Yang L, Tang DN, Bennett T, Schober W, Lewis DE. Regulation of CD28 costimulationin human CD8+ T cells. J Immunol 1997;158:1551–8. [PubMed: 9029089]
9. Brenchley JM, Karandikar NJ, Betts MR, Ambrozak DR, Hill BJ, Crotty LE, Casazza JP, Kuruppu J,Migueles SA, Connors M, Roederer M, Douek DC, Koup RA. Expression of CD57 defines replicativesenescence and antigen-induced apoptotic death of CD8+ T cells. Blood 2003;101:2711–20. [PubMed:12433688]
10. Douek DC. Disrupting T-cell homeostasis: how HIV-1 infection causes disease. AIDS Rev2003;5:172–7. [PubMed: 14598566]
11. Douek DC, Picker LJ, Koup RA. T cell dynamics in HIV-1 infection. Annu Rev Immunol2003;21:265–304. [PubMed: 12524385]
12. Hazenberg MD, Hamann D, Schuitemaker H, Miedema F. T cell depletion in HIV-1 infection: howCD4+ T cells go out of stock. Nat Immunol 2000;1:285–9. [PubMed: 11017098]
Mansoor et al. Page 7
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

13. Clerici M, Saresella M, Colombo F, Fossati S, Sala N, Bricalli D, Villa ML, Ferrante P, Dally L,Vigano A. T-lymphocyte maturation abnormalities in uninfected newborns and children with verticalexposure to HIV. Blood 2000;96:3866–71. [PubMed: 11090071]
14. McCloskey TW, Kohn N, Lesser M, Bakshi S, Pahwa S. Immunophenotypic analysis of HIV-infectedchildren: alterations within the first year of life, changes with disease progression, and longitudinalanalyses of lymphocyte subsets. Cytometry 2001;46:157–65. [PubMed: 11449406]
15. Shearer WT, Easley KA, Goldfarb J, Rosenblatt HM, Jenson HB, Kovacs A, McIntosh K. Prospective5-year study of peripheral blood CD4, CD8, and CD19/CD20 lymphocytes and serum Igs in childrenborn to HIV-1 women. The P(2)C(2) HIV Study Group. J Allergy Clin Immunol 2000;106:559–66.[PubMed: 10984378]
16. Kharbanda M, McCloskey TW, Pahwa R, Sun M, Pahwa S. Alterations in T-cell receptor Vbetarepertoire of CD4 and CD8 T lymphocytes in human immunodeficiency virus-infected children. ClinDiagn Lab Immunol 2003;10:53–8. [PubMed: 12522039]
17. Deeks SG, Kitchen CM, Liu L, Guo H, Gascon R, Narvaez AB, Hunt P, Martin JN, Kahn JO, LevyJ, McGrath MS, Hecht FM. Immune activation set point during early HIV infection predictssubsequent CD4+ T-cell changes independent of viral load. Blood 2004;104:942–7. [PubMed:15117761]
18. Hazenberg MD, Otto SA, van Benthem BH, Roos MT, Coutinho RA, Lange JM, Hamann D, PrinsM, Miedema F. Persistent immune activation in HIV-1 infection is associated with progression toAIDS. Aids 2003;17:1881–8. [PubMed: 12960820]
19. Aandahl EM, Quigley MF, Moretto WJ, Moll M, Gonzalez VD, Sonnerborg A, Lindback S, HechtFM, Deeks SG, Rosenberg MG, Nixon DF, Sandberg JK. Expansion of CD7(low) and CD7(negative)CD8 T-cell effector subsets in HIV-1 infection: correlation with antigenic load and reversion byantiretroviral treatment. Blood 2004;104:3672–8. [PubMed: 15308569]
20. Rabin RL, Roederer M, Maldonado Y, Petru A, Herzenberg LA, Herzenberg LA. Alteredrepresentation of naive and memory CD8 T cell subsets in HIV-infected children. J Clin Invest1995;95:2054–60. [PubMed: 7738172]
21. Resino S, Correa R, Bellon JM, Munoz-Fernandez MA. Preserved immune system in long-termasymptomatic vertically HIV-1 infected children. Clin Exp Immunol 2003;132:105–12. [PubMed:12653844]
22. Jordan KA, Furlan SN, Gonzalez VD, Karlsson AC, Quigley MF, Deeks SG, Rosenberg MG, NixonDF, Sandberg JK. CD8 T cell effector maturation in HIV-1-infected children. Virology2006;347:117–26. [PubMed: 16406047]
23. Champagne P, Ogg GS, King AS, Knabenhans C, Ellefsen K, Nobile M, Appay V, Rizzardi GP,Fleury S, Lipp M, Forster R, Rowland-Jones S, Sekaly RP, McMichael AJ, Pantaleo G. Skewedmaturation of memory HIV-specific CD8 T lymphocytes. Nature 2001;410:106–11. [PubMed:11242051]
24. Harari A, Vallelian F, Pantaleo G. Phenotypic heterogeneity of antigen-specific CD4 T cells underdifferent conditions of antigen persistence and antigen load. Eur J Immunol 2004;34:3525–33.[PubMed: 15484193]
25. Kourtis AP, Ibegbu C, Nahmias AJ, Lee FK, Clark WS, Sawyer MK, Nesheim S. Early progressionof disease in HIV-infected infants with thymus dysfunction. N Engl J Med 1996;335:1431–6.[PubMed: 8875920]
26. Newell ML, Coovadia H, Cortina-Borja M, Rollins N, Gaillard P, Dabis F. Mortality of infected anduninfected infants born to HIV-infected mothers in Africa: a pooled analysis. Lancet 2004;364:1236–43. [PubMed: 15464184]
27. Newell ML, Brahmbhatt H, Ghys PD. Child mortality and HIV infection in Africa: a review. Aids2004;18(Suppl 2):S27–34. [PubMed: 15319741]
28. Rich KC, Siegel JN, Jennings C, Rydman RJ, Landay AL. Function and phenotype of immature CD4+ lymphocytes in healthy infants and early lymphocyte activation in uninfected infants of humanimmunodeficiency virus-infected mothers. Clin Diagn Lab Immunol 1997;4:358–61. [PubMed:9144377]
Mansoor et al. Page 8
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

29. Kuhn L, Meddows-Taylor S, Gray G, Tiemessen C. Human immunodeficiency virus (HIV)-specificcellular immune responses in newborns exposed to HIV in utero. Clin Infect Dis 2002;34:267–76.[PubMed: 11740717]
30. Ssewanyana I, Elrefaei M, Dorsey G, Ruel T, Jones NG, Gasasira A, Kamya M, Nakiwala J, AchanJ, Charlebois E, Havlir D, Cao H. Profile of T cell immune responses in HIV-infected children fromUganda. J Infect Dis 2007;196:1667–70. [PubMed: 18008251]
31. Brown, H.; Prescott, R. Applied Mixed Models in Medicine. Wiley; 2004.32. Brugnoni D, Prati E, Malacarne F, Gorla R, Airo P, Cattaneo R. The primary response to HIV infection
is characterized by an expansion of activated CD8+ CD28-cells. Aids 1996;10:104–6. [PubMed:8924239]
33. Gruters RA, Terpstra FG, De Goede RE, Mulder JW, De Wolf F, Schellekens PT, Van Lier RA,Tersmette M, Miedema F. Immunological and virological markers in individuals progressing fromseroconversion to AIDS. Aids 1991;5:837–44. [PubMed: 1892588]
34. Papagno L, Spina CA, Marchant A, Salio M, Rufer N, Little S, Dong T, Chesney G, Waters A,Easterbrook P, Dunbar PR, Shepherd D, Cerundolo V, Emery V, Griffiths P, Conlon C, McMichaelAJ, Richman DD, Rowland-Jones SL, Appay V. Immune activation and CD8+ T-cell differentiationtowards senescence in HIV-1 infection. PLoS Biol 2004;2:E20. [PubMed: 14966528]
35. Roederer M, Dubs JG, Anderson MT, Raju PA, Herzenberg LA, Herzenberg LA. CD8 naive T cellcounts decrease progressively in HIV-infected adults. J Clin Invest 1995;95:2061–6. [PubMed:7738173]
36. Gallagher K, Gorre M, Harawa N, Dillon M, Wafer D, Stiehm ER, Bryson Y, Song D, Dickover R,Plaeger S. Timing of lymphocyte activation in neonates infected with human immunodeficiencyvirus. Clin Diagn Lab Immunol 1997;4:742–7. [PubMed: 9384301]
37. Meyers A, Shah A, Cleveland RH, Cranley WR, Wood B, Sunkle S, Husak S, Cooper ER. Thymicsize on chest radiograph and rapid disease progression in human immunodeficiency virus 1-infectedchildren. Pediatr Infect Dis J 2001;20:1112–8. [PubMed: 11740315]
38. Nahmias AJ, Clark WS, Kourtis AP, Lee FK, Cotsonis G, Ibegbu C, Thea D, Palumbo P, Vink P,Simonds RJ, Nesheim SR. Thymic dysfunction and time of infection predict mortality in humanimmunodeficiency virus-infected infants. CDC Perinatal AIDS Collaborative Transmission StudyGroup. J Infect Dis 1998;178:680–5. [PubMed: 9728535]
39. Harari A, Petitpierre S, Vallelian F, Pantaleo G. Skewed representation of functionally distinctpopulations of virus-specific CD4 T cells in HIV-1-infected subjects with progressive disease:changes after antiretroviral therapy. Blood 2004;103:966–72. [PubMed: 12958069]
40. Buseyne F, Burgard M, Teglas JP, Bui E, Rouzioux C, Mayaux MJ, Blanche S, Riviere Y. Early HIV-specific cytotoxic T lymphocytes and disease progression in children born to HIV-infected mothers.AIDS Res Hum Retroviruses 1998;14:1435–44. [PubMed: 9824321]
41. Luzuriaga K, Holmes D, Hereema A, Wong J, Panicali DL, Sullivan JL. HIV-1-specific cytotoxic Tlymphocyte responses in the first year of life. J Immunol 1995;154:433–43. [PubMed: 7995957]
42. Thobakgale CF, Ramduth D, Reddy S, Mkhwanazi N, de Pierres C, Moodley E, Mphatswe W,Blanckenberg N, Cengimbo A, Prendergast A, Tudor-Williams G, Dong K, Jeena P, Kindra G, BobatR, Coovadia H, Kiepiela P, Walker BD, Goulder PJ. Human Immunodeficiency Virus-Specific CD8+ T-Cell Activity Is Detectable from Birth in the Majority of In Utero-Infected Infants. J Virol2007;81:12775–84. [PubMed: 17881456]
43. Lohman BL, Slyker JA, Richardson BA, Farquhar C, Mabuka JM, Crudder C, Dong T, Obimbo E,Mbori-Ngacha D, Overbaugh J, Rowland-Jones S, John-Stewart G. Longitudinal assessment ofhuman immunodeficiency virus type 1 (HIV-1)-specific gamma interferon responses during the firstyear of life in HIV-1-infected infants. J Virol 2005;79:8121–30. [PubMed: 15956557]
44. Riviere Y, Buseyne F. Cytotoxic T lymphocytes generation capacity in early life with particularreference to HIV. Vaccine 1998;16:1420–2. [PubMed: 9711782]
45. Palmer BE, Blyveis N, Fontenot AP, Wilson CC. Functional and phenotypic characterization of CD57+CD4+ T cells and their association with HIV-1-induced T cell dysfunction. J Immunol2005;175:8415–23. [PubMed: 16339584]
Mansoor et al. Page 9
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

46. Choremi-Papadopoulou H, Viglis V, Gargalianos P, Kordossis T, Iniotaki-Theodoraki A, KosmidisJ. Downregulation of CD28 surface antigen on CD4+ and CD8+ T lymphocytes during HIV-1infection. J Acquir Immune Defic Syndr 1994;7:245–53. [PubMed: 7906302]
47. Haffar OK, Smithgall MD, Wong JG, Bradshaw J, Linsley PS. Human immunodeficiency virus type1 infection of CD4+ T cells down-regulates the expression of CD28: effect on T cell activation andcytokine production. Clin Immunol Immunopathol 1995;77:262–70. [PubMed: 7586736]
48. Swigut T, Shohdy N, Skowronski J. Mechanism for down-regulation of CD28 by Nef. Embo J2001;20:1593–604. [PubMed: 11285224]
49. Borthwick NJ, Bofill M, Gombert WM, Akbar AN, Medina E, Sagawa K, Lipman MC, Johnson MA,Janossy G. Lymphocyte activation in HIV-1 infection. II. Functional defects of CD28- T cells. Aids1994;8:431–41. [PubMed: 8011246]
50. Vingerhoets JH, Vanham GL, Kestens LL, Penne GG, Colebunders RL, Vandenbruaene MJ, GoemanJ, Gigase PL, De Boer M, Ceuppens JL. Increased cytolytic T lymphocyte activity and decreased B7responsiveness are associated with CD28 down-regulation on CD8+ T cells from HIV-infectedsubjects. Clin Exp Immunol 1995;100:425–33. [PubMed: 7539725]
51. Rabie, H.; Violari, A.; Madhi, S.; Gibb, DM.; Steyn, J.; van Niekerk, R.; Josipovic, D.; Innes, S.;Cotton, MF. CROI. Boston, USA: 2008. Complications of BCG Vaccination in HIV-infected and -uninfected Children: evidence from the Children with HIV Early Antiretroviral Therapy (CHER)Study.
Mansoor et al. Page 10
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure I.Frequencies of total CD4+ T cells (A) and CD8+ T cells (B), among total T cells and CD4/CD8ratios (C), in the peripheral blood of infants from the HIV+, HIV− Exposed, or HIV− groups,over the first year of life, measured by flow cytometry. Boxes in boxplots represent mediansand interquartile ranges, with the total range as the whiskers. Differences: *p < 0.05; **p <0.01; ***p< 0.001, Kruskal-Wallis test, with Dunn’s Multiple Comparison Test.
Mansoor et al. Page 11
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
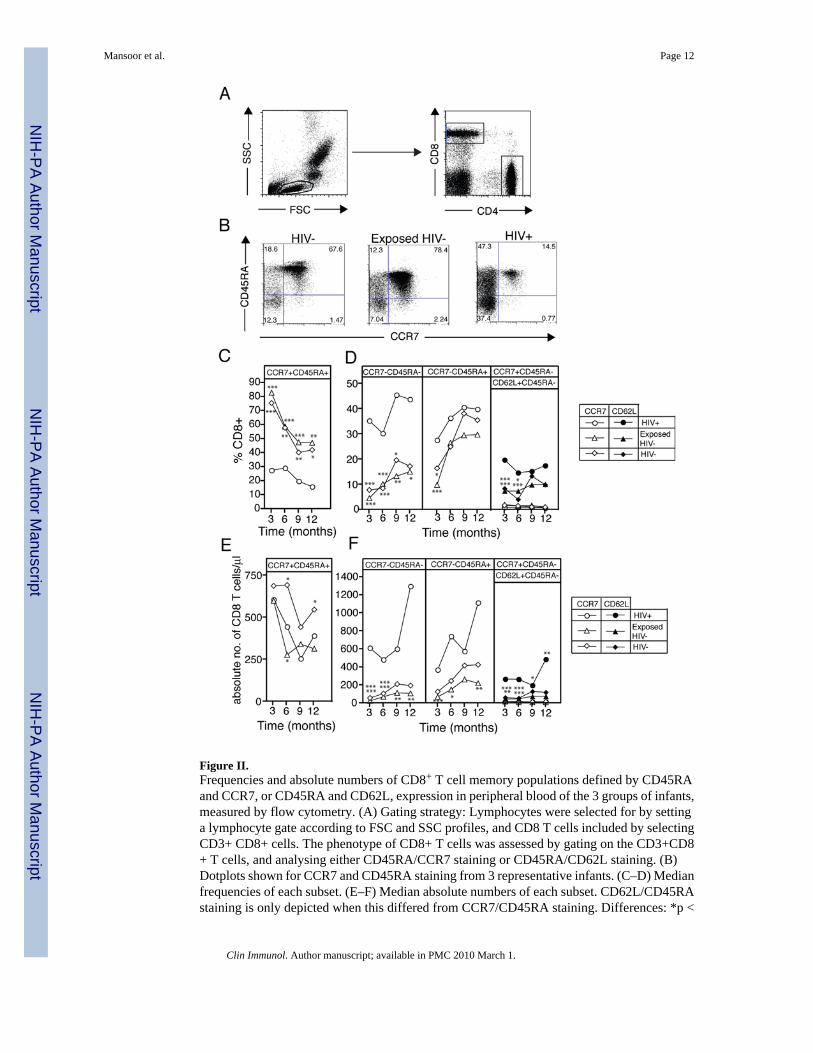
Figure II.Frequencies and absolute numbers of CD8+ T cell memory populations defined by CD45RAand CCR7, or CD45RA and CD62L, expression in peripheral blood of the 3 groups of infants,measured by flow cytometry. (A) Gating strategy: Lymphocytes were selected for by settinga lymphocyte gate according to FSC and SSC profiles, and CD8 T cells included by selectingCD3+ CD8+ cells. The phenotype of CD8+ T cells was assessed by gating on the CD3+CD8+ T cells, and analysing either CD45RA/CCR7 staining or CD45RA/CD62L staining. (B)Dotplots shown for CCR7 and CD45RA staining from 3 representative infants. (C–D) Medianfrequencies of each subset. (E–F) Median absolute numbers of each subset. CD62L/CD45RAstaining is only depicted when this differed from CCR7/CD45RA staining. Differences: *p <
Mansoor et al. Page 12
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

0.05; **p < 0.01; ***p< 0.001, Kruskal-Wallis test, with Dunn’s Multiple Comparison Test.The distributions are shown in Supplementary Figures I and II.
Mansoor et al. Page 13
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
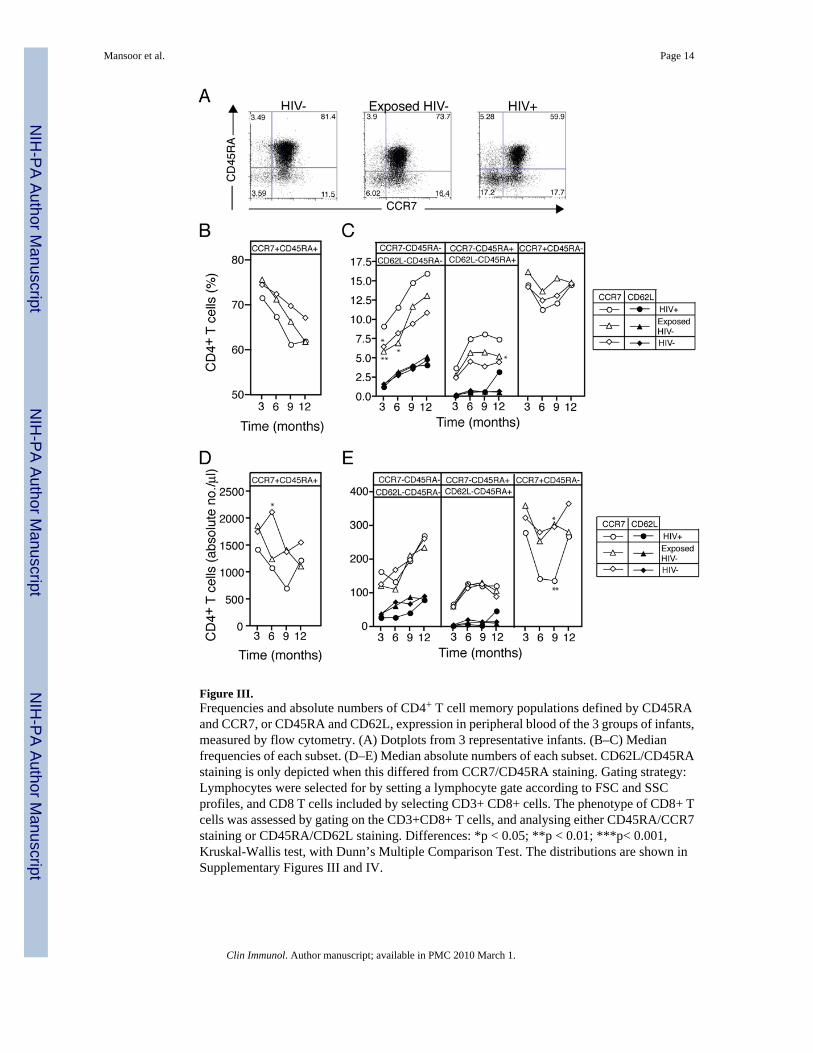
Figure III.Frequencies and absolute numbers of CD4+ T cell memory populations defined by CD45RAand CCR7, or CD45RA and CD62L, expression in peripheral blood of the 3 groups of infants,measured by flow cytometry. (A) Dotplots from 3 representative infants. (B–C) Medianfrequencies of each subset. (D–E) Median absolute numbers of each subset. CD62L/CD45RAstaining is only depicted when this differed from CCR7/CD45RA staining. Gating strategy:Lymphocytes were selected for by setting a lymphocyte gate according to FSC and SSCprofiles, and CD8 T cells included by selecting CD3+ CD8+ cells. The phenotype of CD8+ Tcells was assessed by gating on the CD3+CD8+ T cells, and analysing either CD45RA/CCR7staining or CD45RA/CD62L staining. Differences: *p < 0.05; **p < 0.01; ***p< 0.001,Kruskal-Wallis test, with Dunn’s Multiple Comparison Test. The distributions are shown inSupplementary Figures III and IV.
Mansoor et al. Page 14
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
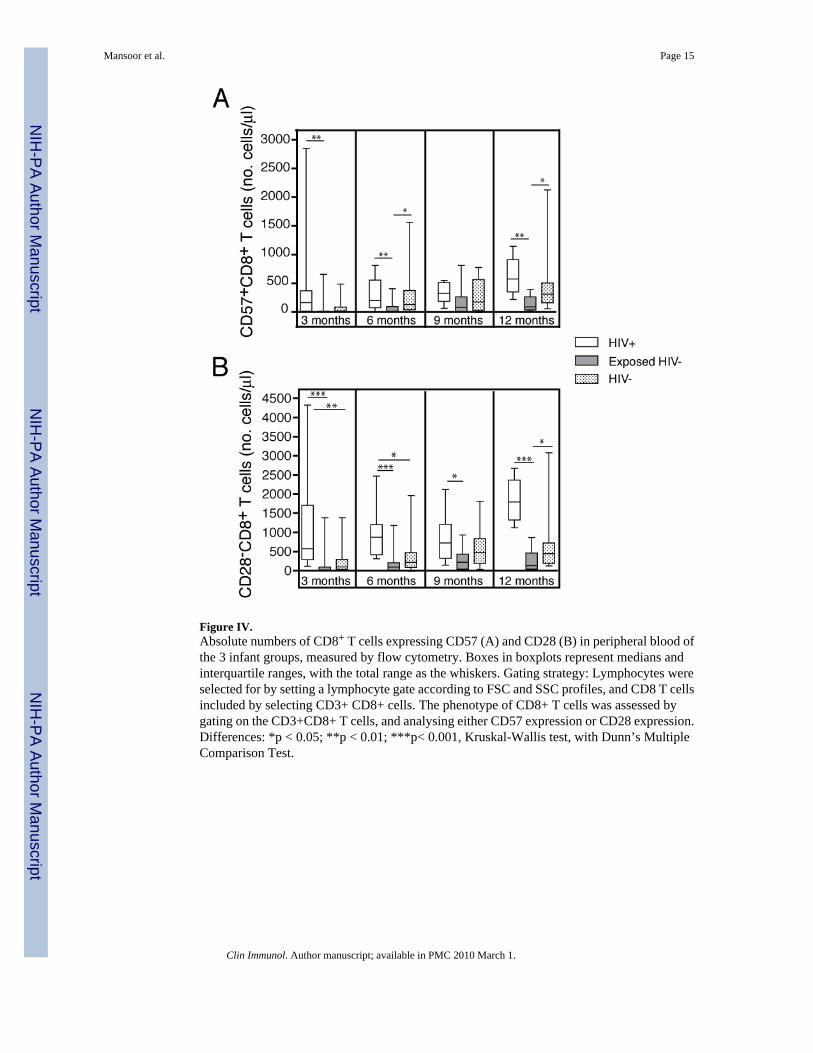
Figure IV.Absolute numbers of CD8+ T cells expressing CD57 (A) and CD28 (B) in peripheral blood ofthe 3 infant groups, measured by flow cytometry. Boxes in boxplots represent medians andinterquartile ranges, with the total range as the whiskers. Gating strategy: Lymphocytes wereselected for by setting a lymphocyte gate according to FSC and SSC profiles, and CD8 T cellsincluded by selecting CD3+ CD8+ cells. The phenotype of CD8+ T cells was assessed bygating on the CD3+CD8+ T cells, and analysing either CD57 expression or CD28 expression.Differences: *p < 0.05; **p < 0.01; ***p< 0.001, Kruskal-Wallis test, with Dunn’s MultipleComparison Test.
Mansoor et al. Page 15
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Mansoor et al. Page 16Ta
ble
1
Age
3 m
onth
s6
mon
ths
9 m
onth
s12
mon
ths
HIV
stat
usH
IV+
Exp
.H
IV−
HIV−
HIV
+E
xp.
HIV−
HIV−
HIV
+E
xp.
HIV−
HIV−
HIV
+E
xp.
HIV−
HIV−
Sam
ple
size
, n16
2523
1118
219
1722
514
19
Lym
phoc
ytes
(×10
9 /lite
r), m
edia
n(I
QR
)
8.1
(5.4
–10.
7)6.
4(5
.1–8
.2)
6.4
(5.2
–8.5
)5.
8(3
.6–9
.8)
5.6
(4.2
–7.4
)7.
6(5
.2–9
.0)
5.7
(4.2
–6.5
)6.
5(4
.2–8
.1)
6.1
(4.5
–8.6
)7.
7(5
.9–9
.7)
5.1
(3.4
–7.1
)6.
5(4
.5–9
.3)
CD
4(ly
mph
ocyt
es/u
l) m
edia
n(I
QR
)
2024
(120
4–28
40)
2519
(194
0–28
74)
2465
(202
0–30
32)
1469
(614
–256
2)18
24(1
383–
2124
)27
63(1
603–
3359
)a12
66(7
60–2
038)
2211
(143
0–23
19)
2061
(161
1–26
05)
1872
(112
1–26
59)
1833
(139
4–20
84)
2190
(158
1–28
77)
CD
8(ly
mph
ocyt
es/u
l) m
edia
n(I
QR
)
1569
(118
6–29
38)
732
(565
–127
0)b
1019
(778
–125
0)a
1460
(879
–305
7)64
0(4
62–9
80)c,
d12
93(8
92–1
567)
1464
(127
0–24
78)
766
(461
–174
8)11
39(9
06–1
734)
2819
(212
7–36
53)
653
(515
–977
)b,e
1162
(869
–215
1)
Vira
l loa
d Lo
g 10 R
NA
copi
es/m
l med
ian
(IQ
R)
5.7
(4.9
–6.2
)N
/AN
/A5.
6(4
.8–5
.9)
N/A
N/A
4.9
(3.8
–5.5
)N
/AN
/A4.
9(2
.4–5
.7)
N/A
N/A
a HIV
+ an
d H
IV−
(p<0
.05)
b HIV
+ an
d Ex
p. H
IV−
(p<0
.001
)
c HIV
+ an
d Ex
p. H
IV−
(p<0
.01)
d Exp.
HIV−
and
HIV−
(p<0
.01)
e Exp.
HIV−
and
HIV−
(p<0
.05)
Clin Immunol. Author manuscript; available in PMC 2010 March 1.
Related Documents











