Psychopharmacology ( . ):3–19 DOI 10.1007/s00213-004-1920-9 REVIEW Xavier Guitart . Xavier Codony . Xavier Monroy Sigma receptors: biology and therapeutic potential Received: 24 July 2003 / Accepted: 14 April 2004 # Springer-Verlag Abstract More than 20 years after the identification of the sigma receptors as a unique binding site in the brain and in the peripheral organs, several questions regarding this receptor are still open. Only one of the subtypes of the receptor has been cloned to date, but the endogenous ligand still remains unknown, and the possible association of the receptor with a conventional second messenger system is controversial. From the very beginning, the sigma receptors were associated with various central nervous system disorders such as schizophrenia or move- ment disorders. Today, after hundreds of papers dealing with the importance of sigma receptors in brain function, it is widely accepted that sigma receptors represent a new and different avenue in the possible pharmacological treatment of several brain-related disorders. In this review, what is known about the biology of the sigma receptor regarding its putative structure and its distribution in the central nervous system is summarized first. The role of sigma receptors regulating cellular functions and other neurotransmitter systems is also addressed, as well as a short overview of the possible endogenous ligands. Finally, although no specific sigma ligand has reached the market, different pharmacological approaches to the alleviation and treatment of several central nervous system disorders and deficits, including schizophrenia, pain, memory deficits, etc., are discussed, with an overview of different compounds and their potential therapeutic use. Keywords Sigma receptors . Schizophrenia . Drug abuse . Clinical potential . Brain disorder Introduction The process of understanding sigma receptors has been a long, complicated and somewhat mysterious one, and the story is still far from complete. In an early work, Haertzen (1970) described the psychomimetic effects of cyclazocine and nalorphine in humans. Later on, Martin and colleagues (1976), working in morphine-dependent and non-depen- dent chronic spinal dogs, reported the psychomimetic effects of N-allyl-normetazocine (SKF 10,047). These effects could not be explained by the actions on μ-opioid or κ-opioid receptors, and contributed to the proposal of a σ-opioid receptor. Nevertheless, the enantiomeric selectivity of the sigma receptor for the (+)-isoforms of the benzomorphans rather than for their (−)-isoforms (Brady et al. 1982), and the fact that the effects of sigma ligands neither in vivo nor in vitro were blocked by classical opioid antagonists (Iwamoto 1981; Vaupel 1983) led to some confusion regarding the true nature of sigma binding sites. Moreover, the sigma receptor was also identified as the binding site for phencyclidine (PCP) within the N-methyl-D-aspartate (NMDA) glutamate receptor (Zukin et al. 1984; Mendel- sohn et al. 1985). However, further research in more recent years conclusively defined the nature of sigma receptors, and they were finally reclassified as a non-opioid and non- PCP unique site (Quirion et al. 1992). Since sigma receptors were first identified in the central nervous system, extensive work has been performed in various laboratories in an attempt to elucidate their physiological functions and, despite the initial confusion, significant advances have been made in delineating the functions of sigma receptors. These advances have been summarized in several review articles describing their known physiological roles, as well as potential clinical uses of sigma receptors and their ligands (Walker et al. 1990; Debonnel 1993; Su 1993; Novakova 1998; Bowen 2000; Maurice et al. 2001; Matsumoto et al. 2003). This review examines the known functions of the sigma receptors regarding the different types that have been described, the cellular signals coupled to the stimulation- X. Guitart (*) . X. Codony . X. Monroy Biological Discovery and CNS Research Department, Research Center, Esteve, S.A., Mare de Deu de Montserrat 221, 08041 Barcelona, Spain e-mail: [email protected] Tel.: +34-93-4466063 Fax: +34-93-4466220 08/06/04 - PROOF PAGEMAKEUP 213_1920

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Psychopharmacology ( . ) : 3–19DOI 10.1007/s00213-004-1920-9
REVIEW
Xavier Guitart . Xavier Codony . Xavier Monroy
Sigma receptors: biology and therapeutic potential
Received: 24 July 2003 / Accepted: 14 April 2004# Springer-Verlag
Abstract More than 20 years after the identification ofthe sigma receptors as a unique binding site in the brainand in the peripheral organs, several questions regardingthis receptor are still open. Only one of the subtypes of thereceptor has been cloned to date, but the endogenousligand still remains unknown, and the possible associationof the receptor with a conventional second messengersystem is controversial. From the very beginning, thesigma receptors were associated with various centralnervous system disorders such as schizophrenia or move-ment disorders. Today, after hundreds of papers dealingwith the importance of sigma receptors in brain function, itis widely accepted that sigma receptors represent a newand different avenue in the possible pharmacologicaltreatment of several brain-related disorders. In this review,what is known about the biology of the sigma receptorregarding its putative structure and its distribution in thecentral nervous system is summarized first. The role ofsigma receptors regulating cellular functions and otherneurotransmitter systems is also addressed, as well as ashort overview of the possible endogenous ligands.Finally, although no specific sigma ligand has reachedthe market, different pharmacological approaches to thealleviation and treatment of several central nervous systemdisorders and deficits, including schizophrenia, pain,memory deficits, etc., are discussed, with an overview ofdifferent compounds and their potential therapeutic use.
Keywords Sigma receptors . Schizophrenia . Drug abuse .Clinical potential . Brain disorder
Introduction
The process of understanding sigma receptors has been along, complicated and somewhat mysterious one, and thestory is still far from complete. In an early work, Haertzen(1970) described the psychomimetic effects of cyclazocineand nalorphine in humans. Later on, Martin and colleagues(1976), working in morphine-dependent and non-depen-dent chronic spinal dogs, reported the psychomimeticeffects of N-allyl-normetazocine (SKF 10,047). Theseeffects could not be explained by the actions on μ-opioidor κ-opioid receptors, and contributed to the proposal of aσ-opioid receptor.
Nevertheless, the enantiomeric selectivity of the sigmareceptor for the (+)-isoforms of the benzomorphans ratherthan for their (−)-isoforms (Brady et al. 1982), and the factthat the effects of sigma ligands neither in vivo nor in vitrowere blocked by classical opioid antagonists (Iwamoto1981; Vaupel 1983) led to some confusion regarding thetrue nature of sigma binding sites. Moreover, the sigmareceptor was also identified as the binding site forphencyclidine (PCP) within the N-methyl-D-aspartate(NMDA) glutamate receptor (Zukin et al. 1984; Mendel-sohn et al. 1985). However, further research in more recentyears conclusively defined the nature of sigma receptors,and they were finally reclassified as a non-opioid and non-PCP unique site (Quirion et al. 1992).
Since sigma receptors were first identified in the centralnervous system, extensive work has been performed invarious laboratories in an attempt to elucidate theirphysiological functions and, despite the initial confusion,significant advances have been made in delineating thefunctions of sigma receptors. These advances have beensummarized in several review articles describing theirknown physiological roles, as well as potential clinicaluses of sigma receptors and their ligands (Walker et al.1990; Debonnel 1993; Su 1993; Novakova 1998; Bowen2000; Maurice et al. 2001; Matsumoto et al. 2003).
This review examines the known functions of the sigmareceptors regarding the different types that have beendescribed, the cellular signals coupled to the stimulation-
X. Guitart (*) . X. Codony . X. MonroyBiological Discovery and CNS Research Department, ResearchCenter,Esteve, S.A., Mare de Deu de Montserrat 221,08041 Barcelona, Spaine-mail: [email protected].: +34-93-4466063Fax: +34-93-4466220
08/06/04 - PROOF PAGEMAKEUP 213_1920

blockade of the sigma receptors, and also focuses onexamining specific aspects of sigma receptor ligandsrelated to mental and neurological alterations, and theirputative therapeutic applications. This article will try tosummarize the current understanding in the study of sigmareceptors, but does not pretend to be extensive and not allthe work and references can be cited.
Classification of sigma receptors
Sigma receptors have been extensively characterized inbinding studies. These receptors are distinct from anyother known neurotransmitter receptors and have beendefined as non-opioid binding sites with high affinity for1,3-di (2-tolyl) guanidine (DTG), (+)-3-(3-hydroxyphe-nyl)-N-(1-propyl)piperidine [(+)-3-PPP], or haloperidol.
Biochemical and pharmacological studies stronglysuggest that there are multiple subtypes of sigmareceptors. The best characterized binding sites aredesignated as sigma-1 and sigma-2 receptor types(Hellewell and Bowen 1990; Quirion et al. 1992; Bowen2000). These receptor subtypes differ primarily in theirdrug selectivity. Sigma-1 sites exhibit high affinity andstereoselectivity for the (+)-isomers of pentazocine,cyclazocine, and SKF 10,047, whereas sigma-2 sitestend to prefer the (−)-stereoisomers (Bowen et al. 1993;Hellewell et al. 1994). Sigma-1 and sigma-2 receptors alsodiffer in their tissue distribution patterns, subcellularlocalizations, and apparent molecular weight (Itzhak1994; Torrence-Campbell and Bowen 1996).
The sigma-1 receptor has been recently cloned fromvarious sources, including guinea-pig liver (Hanner et al.1996), human placental choriocarcinoma cells (Kekuda etal. 1996), human brain (Prasad et al. 1998), rat brain (Sethet al. 1998; Mei and Pasternak 2001), and mouse brain(Pan et al. 1998). Based on the predicted 223 amino acid
sequence of the receptor, it shares no structural homologywith opioid receptors or other neurotransmitter receptors,although sigma receptors expressed in the central nervoussystem are highly similar to those expressed in peripheralorgans. The locus coding for the sigma-1 receptor islocated in the human chromosome 9 and in the mousechromosome 4. The gene is ∼7 kbp long and contains fourexons and three introns (Prasad et al. 1998). The aminoacid sequence of the cloned sigma-1 receptor shows nohomology with other mammalian proteins, but sharesapproximately 30% identity with the yeast gene thatencodes a sterol C8–C7 isomerase that is necessary forcholesterol synthesis (Moebius et al. 1997a,b). However, amammalian C8–C7 has been recently cloned (Hanner et al.1995), but this protein shows no similarity to either theyeast isomerase or the sigma receptor. In contrast, sigma-1receptors do not possess sterol isomerase activity (Hanneret al. 1996).
Based on hydropathy analysis of the amino acidsequence of the sigma-1 receptor, it was suggested thatthis receptor includes a single putative transmembranedomain (Mei and Pasternak 2001). Other recent studiescompared the homology between this putative transmem-brane domain and segments of other known transmem-brane domains; these studies suggested that the sigma-1receptor has two transmembrane segments, resulting in anextracellular loop of approximately 50 amino acids and anintracellular C-terminus of approximately 125 amino acids(Aydar et al. 2002). According to this model, the N-terminus is very short and also localized intracellularly.Figure 1 illustrates the two structural models that havebeen proposed for the sigma-1 receptor. The amino acidsequence is not consistent with that of a classical G-protein-coupled receptor, which makes it especiallydifficult to predict the effects of agonists or antagonistsof this receptor, at least in the traditional way, by directlinkage to stimulation or inhibition of intracellular
Fig. 1 Predicted amino acidsequence of the rat brain sigmareceptor. Different models havebeen proposed (see text fordetails). Receptor models with asingle putative transmembranedomain (a) and two putativetransmembrane domains (b)
2 08/06/04 - PROOF PAGEMAKEUP 213_1920
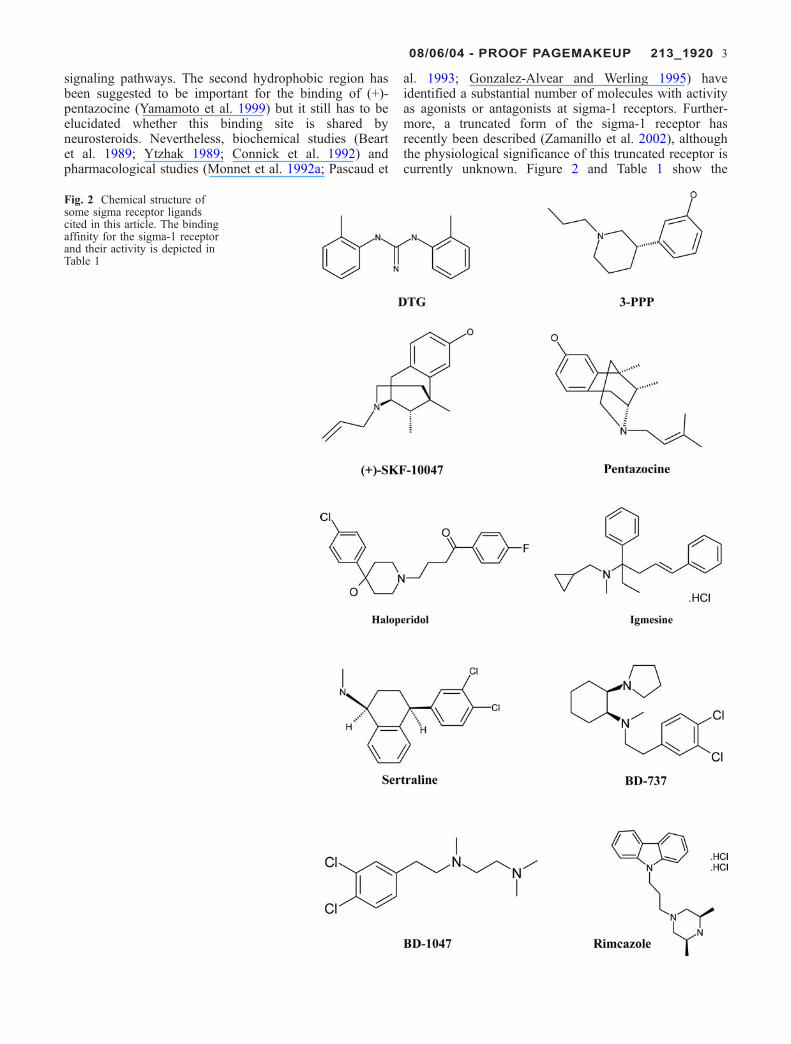
signaling pathways. The second hydrophobic region hasbeen suggested to be important for the binding of (+)-pentazocine (Yamamoto et al. 1999) but it still has to beelucidated whether this binding site is shared byneurosteroids. Nevertheless, biochemical studies (Beartet al. 1989; Ytzhak 1989; Connick et al. 1992) andpharmacological studies (Monnet et al. 1992a; Pascaud et
al. 1993; Gonzalez-Alvear and Werling 1995) haveidentified a substantial number of molecules with activityas agonists or antagonists at sigma-1 receptors. Further-more, a truncated form of the sigma-1 receptor hasrecently been described (Zamanillo et al. 2002), althoughthe physiological significance of this truncated receptor iscurrently unknown. Figure 2 and Table 1 show the
Fig. 2 Chemical structure ofsome sigma receptor ligandscited in this article. The bindingaffinity for the sigma-1 receptorand their activity is depicted inTable 1
08/06/04 - PROOF PAGEMAKEUP 213_1920 3
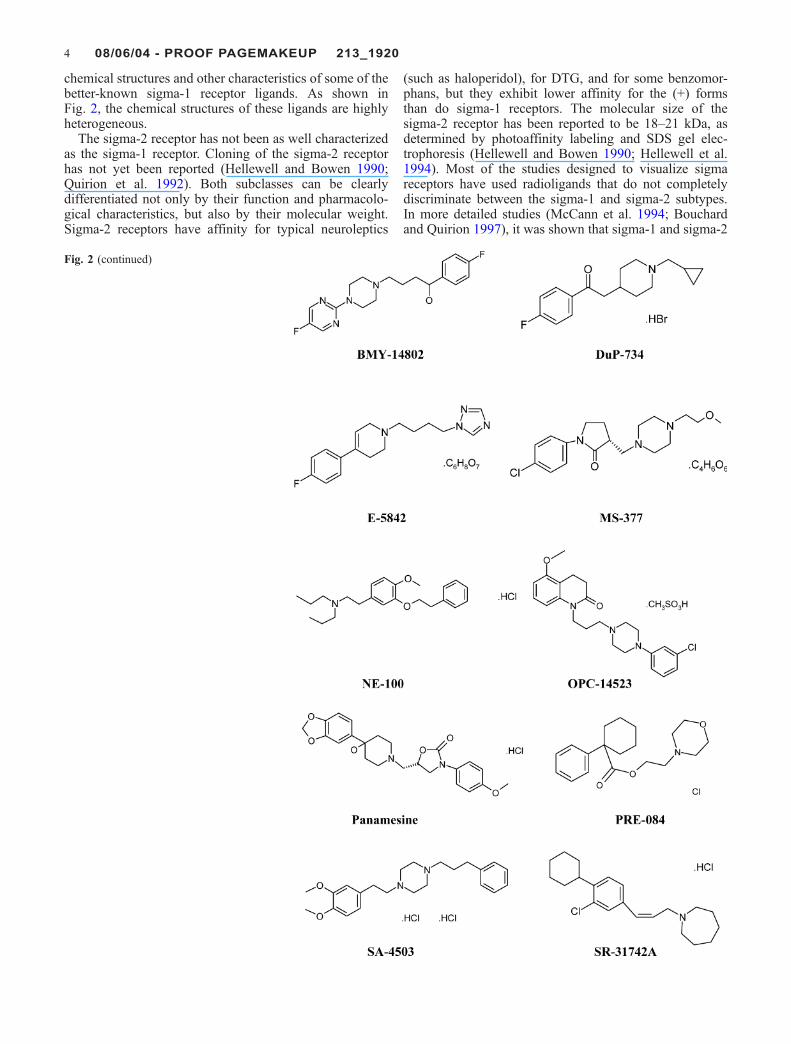
chemical structures and other characteristics of some of thebetter-known sigma-1 receptor ligands. As shown inFig. 2, the chemical structures of these ligands are highlyheterogeneous.
The sigma-2 receptor has not been as well characterizedas the sigma-1 receptor. Cloning of the sigma-2 receptorhas not yet been reported (Hellewell and Bowen 1990;Quirion et al. 1992). Both subclasses can be clearlydifferentiated not only by their function and pharmacolo-gical characteristics, but also by their molecular weight.Sigma-2 receptors have affinity for typical neuroleptics
(such as haloperidol), for DTG, and for some benzomor-phans, but they exhibit lower affinity for the (+) formsthan do sigma-1 receptors. The molecular size of thesigma-2 receptor has been reported to be 18–21 kDa, asdetermined by photoaffinity labeling and SDS gel elec-trophoresis (Hellewell and Bowen 1990; Hellewell et al.1994). Most of the studies designed to visualize sigmareceptors have used radioligands that do not completelydiscriminate between the sigma-1 and sigma-2 subtypes.In more detailed studies (McCann et al. 1994; Bouchardand Quirion 1997), it was shown that sigma-1 and sigma-2
Fig. 2 (continued)
4 08/06/04 - PROOF PAGEMAKEUP 213_1920

receptors are generally co-localized, although they may bepresent in different ratios (Leitner et al. 1994).
Sigma-2 receptors have been implicated in the regula-tion of cell proliferation and cell viability. Cytotoxicity ofsigma ligands has been observed both in vivo and in vitro(Vilner and Bowen 1993; Vilner et al. 1995). These effectsare mediated by sigma receptors; ligands with affinity forother receptors or ion channels did not exhibit cytotoxicactivity, and the pharmacological profile of these com-pounds clearly implicates sigma-2 receptors (Bowen et al.1995a). Selective sigma-1 ligands did not induce anycytotoxic effects on cells.
Endogenous ligands of the sigma receptors
Despite considerable advances in our understanding ofsigma receptors over the past several years, an endogenousligand has not yet been found. Identification of anendogenous sigma ligand would improve our under-standing of the role of sigma receptors in physiologicaland pathophysiological brain functions. A number ofknown peptides and other neurotransmitters have beenclearly shown to be ineffective in displacing selectivesigma ligands from sigma receptors. Neurotransmittersthat do not interact with the sigma receptor includeserotonin, norepinephrine, dopamine, and histamine(Weber et al. 1986; DeHaven-Hudkins and Fleissner1992); as well as several amino acids, such as glutamate,glycine, aspartate, and cysteine (Craviso and Musacchio1983; Klein and Musacchio 1989); and various peptides,such as the endorphins, dynorphins, enkephalins, andsubstance P (Samovilova et al. 1988; DeHaven-Hudkinsand Fleissner 1992).
Neuropeptide Y (NPY) deserves particular attention. Ithas been repeatedly suggested that NPY is an endogenousligand for certain sigma receptors. Roman et al. (1989a)showed that two peptides, NPY and YY, which had highaffinity for sigma receptor, both were able to displace the[3H](+)-SFK 10,047 binding. They found that IC50 valuesof these two peptides were approximately 10 nM, althoughthese results could not be repeated later by another group(Tam and Mitchell 1991).
However, in vivo experiments showed that NPY andsigma ligands can modulate NMDA-induced neuronalactivity in the hippocampus (Monnet et al. 1992c,d; Aultand Werling 1997) and NMDA-stimulated [3H]dopaminerelease from rat cortical and striatal slices (Ault andWerling 1998; Hong and Werling 2001). In a more recentstudy, autoradiography was used to show that the presenceof micromolar concentrations of sigma-1 receptor agonistsor antagonists did not alter [35S]GTPγS binding inducedby nanomolar concentrations of NPY in the rat brain.Thus, it appears that NPY does not act as an endogenousligand of sigma-1 receptors.
Steroids have also been considered as putative ligandsof sigma-1 receptors, and the interaction between steroidsand the sigma-1 receptor was first suggested from in vitrobinding experiments in guinea-pig brain and spleen. Suand collaborators (1988) showed an inhibition of [3H](+)-SKF 10,047 binding in the guinea-pig brain and of [3H]-haloperidol in the spleen by progesterone with Ki values inthe nanomolar range. Other steroids, such as testosterone,pregnenolone sulfate, or deoxycorticosterone, exhibit Kivalues in the micromolar range (McCann and Su 1991;Maurice et al. 1996). Confirming these studies, othergroups have shown that progesterone was indeed theneurosteroid which most potently inhibited the binding of
Table 1 Activity of somesigma-1 receptor ligand com-pounds
Compound Sigma-1 activity Sigma-1 affinity (IC50) References
DTG Agonist 45 de Costa and He (1994)(+)-3-PPP Agonist 48 de Costa and He (1994)(+)-SKF-10,047 Agonist 97 de Costa and He (1994)(+)-Pentazocine Agonist 5.8 Chaki et al. (1994)Haloperidol Antagonist 2.3 Chaki et al. (1994)Igmesine Agonist 39 Roman et al. (1990)Sertraline Agonist 6.8 Schmidt et al. (1989)BD-737 Agonist 2 (Ki) Bowen et al. (1992)BD-1047 Antagonist 36 Matsumoto et al. (1995)Rimcazole Antagonist 520 Schmidt et al. (1989)BMY-14,802 Antagonist 66 Koe et al. (1989)DuP734 Antagonist 10 (Ki) Tam et al. (1992)E-5842 Antagonist 5 Guitart et al. (1998)MS-377 Antagonist 73 (Ki) Takahashi et al. (1999)NE-100 Antagonist 1.6 Chaki et al. (1994)OPC-14523 Agonist 47 Oshiro et al. (2000)Panamesine Antagonist 6 Grunder et al. (1999)PRE-084 Agonist 44 Su et al. (1991)SA4503 Agonist 17 Matsuno et al. (1996a,b)SR31742A Antagonist 5.3 Poncelet et al. (1993)
08/06/04 - PROOF PAGEMAKEUP 213_1920 5

several sigma-1 ligands in different preparations, such asthe rat brain, splenocytes plasma membrane, and livermicrosomes (Ross 1991; Klein and Musacchio 1994;Yamada et al. 1994). However, many sigma-1 ligands(haloperidol, carbapentane, DTG, (+)-3-PPP, and rimca-zole) inhibit the [3H]-progesterone binding in rat liverpreparations or porcine liver solubilized fractions (Yamadaet al. 1994; Meyer et al. 1998).
Exogenous administration of neurosteroids leads to adose-dependent inhibition of [3H](+)-SKF 10,047 tosigma-1 receptors (Maurice et al. 1996). In theseexperiments, progesterone was shown to be the mostpotent inhibitor relative to pregnenolone sulphate andDHEA sulphate, although the inhibition of these last twocompounds seems to be unrelated to their in vitro affinity.Modulation of the endogenous steroid levels (adrenalec-tomy—castration) has been shown to enhance the in vivo[3H](+)-SKF 10,047 binding, while the use of pharmaco-logical agents, which augment progesterone concentration,induces a decrease of the in vivo [3H](+)-SKF 10,047binding (Phan et al. 1999). Thus, it seems clear thatneuroactive steroids may clearly interact with the sigma-1receptor being the Ki value of progesterone for the sigmareceptor compatible with an in vivo interaction (forcomprehensive and complete reviews of this area, seeMaurice et al. 1999, 2001).
The interaction of neurosteroids with the sigma receptormost likely occurs through a similar mechanism by whichneurosteroids interact with the NMDA receptor (Bowlby1993; Gore 2001; Lambert et al. 2001) or the γ-aminobutyric acid type-A (GABAA) receptor (Gundlachet al. 1986).
Sigma receptors and signal transduction
Many neurotransmitter receptors are coupled to effectorsystems through various G-proteins, and these G-proteinsplay a critical role in mediating signal transductionmechanisms. Classical examples of regulation of secondmessenger systems by the interaction of a receptor with itseffector include the G-protein coupling of cell surfacereceptors to adenylyl cyclase or phospholipase C (PLC).However, several neurotransmitter receptors are known toregulate some ion channel functions in the absence ofdirect interactions with G-proteins.
The downstream signal transduction pathways regulatedby sigma receptors have not yet been elucidated. Thus, itremains controversial whether or not sigma receptors areassociated with G-proteins. A number of studies haveinvestigated the role of G-proteins in sigma receptorfunction, as outlined below. Early radioligand bindingstudies showed that the binding parameters of severalsigma ligands, such as (+)-SKF 10,047 and (+)-3-PPP,were altered by guanosine triphosphate (GTP) and itsstable analogs such as Gpp(NH)p (Beart et al. 1989;Ytzhak 1989; Connick et al. 1992), suggesting that thesigma receptor was coupled to a G-protein signalingmechanism. More recently, it has been reported that some
sigma receptor ligands stimulate G-protein activity insynaptic membranes from mouse prefrontal cortex (To-kuyama et al. 1997, 1999) and that the binding of [35S]GTPγS in guinea-pig spleen is stimulated by (+)-penta-zocine (Maruo et al. 2000). In addition, several studieshave shown that the binding affinity of [3H](+)-3-PPP forthe sigma receptor was clearly diminished after mem-branes were treated with pertussis toxin or cholera toxin(Ytzhak 1989; Basile et al. 1992). Moreover, some cellulareffects of sigma-1 ligands may be inhibited by pretreat-ment with cholera toxin or pertussis toxin (Monnet et al.1994; Soriani et al. 1999). These findings suggested thatG-proteins are involved in, at least, some of the effects ofsigma-1 receptors.
Despite the considerable evidence suggesting that sigmasites are coupled to G-proteins, other findings imply thatthere is no such coupling, or that it is of a peculiar andcharacteristic nature. For example, the binding parametersof sigma-1 agonists were not altered in response to GTPγStreatment in membrane preparations from rat and guinea-pig cerebellum, as determined with two different radioli-gands for the sigma-1 receptor ([3H]haloperidol and [3H](+)-pentazocine) (Hong and Werling 2000). This is similarto previous findings that used guanine nucleotides andguinea-pig membrane preparations (DeHaven-Hudkins etal. 1992). In order to avoid a possible disruption of thechain effector-receptor-G-protein that could occur in abiochemical approach, autoradiographic studies were usedto show that [35S]GTPγS binding is not stimulated by thepresence of sigma-1 agonists (Hong and Werling 2000).Altogether, these data clearly suggest that a consensus onthis point is still far from being reached.
In electrophysiological studies, the effect of (+)-penta-zocine on N-methyl-D-aspartate (NMDA) responses inhippocampal neurons was not affected by pertussis toxin,whereas cholera toxin altered the effect of (+)-pentazocineeffect on potassium A-current in frog pituitary melano-trope cells (Monnet et al. 1994; Soriani et al. 1999). Incontrast, (+)-pentazocine (as high as 10 μM) has beenreported to have no effect on either basal or forskolin-stimulated adenylyl cyclase in human neuroblastoma BE(2)-C cells (Ryan-Moro et al. 1996). E-5842, a highlyselective sigma-1 receptor ligand and putative atypicalantipsychotic (Guitart and Farré 1998; Guitart et al. 1998),was shown to decrease forskolin-stimulated adenylylcyclase activity (Monroy et al. 2001). Furthermore,when rats were chronically treated with E-5842, increasesin adenylyl cyclase type-I immunoreactivity were ob-served in the brain, whereas acute treatment with E-5842had no effect on adenylyl cyclase activity (Monroy et al.2001).
Phosphoinositide turnover and inositol phosphates havealso been studied in relation to their possible regulation bysigma receptor ligands. Bowen and co-workers and otherlaboratories have shown that sigma ligands may modulatethe function of some receptors coupled to stimulation ofphosphoinositide turnover (Bowen et al. 1988, 1992;Bowen 1994; Novakova et al. 1998). In rat cardiacmyocytes, sigma ligands have been shown to affect cell
6 08/06/04 - PROOF PAGEMAKEUP 213_1920

contraction and calcium movement (Ela et al. 1996). Inthis way, a sigma-1 agonist such as BD737 causes a rapidincrease in inositol triphosphate (IP3) production, whereasBD1047, a putative sigma-1 antagonist, causes a moresustained increase in IP3 levels (Novakova et al. 1998).This effect has been explained by suggesting that BD1047may not act as a true antagonist in cardiac myocytes. Inprimary cultures of rat cortical neurons, sigma ligands(either agonists or antagonists) attenuate NMDA-inducedincreases in calcium (Hayashi et al. 1995; Klette et al.1997). However, in neuron-like cultured cell lines and inbrain synaptosomes, sigma-1 receptor ligands (includingpurported agonists and antagonists) failed to increase IP3levels (Cutts et al. 1993).
It has recently been shown that the sigma-2 receptorregulates Ca2+ release from endoplasmic reticulum stores,and that this effect was reversed by sigma receptorantagonists (Bowen et al 1996; Vilner and Bowen 2000).This study also showed that two different types ofincreases in Ca2+ occur in response to sigma-2 ligands;one is immediate, transient, and is likely derived from theendoplasmic reticulum, whereas the other is sustained andhas been suggested to be derived from mitochondrialstores (Vilner and Bowen 2000). It has also been proposedthat the activation of sigma-2 receptors initiates a Ca2+-dependent process in PC12 cells, which may regulateamphetamine-stimulated dopamine release via Ca2+/cal-modulin kinase II (Weatherspoon and Werling 1999). Incontrast to sigma-1 receptors, it has been proposed thatsigma-2 receptors are lipid raft proteins that would affectcalcium signaling through sphingolipid products (Gebre-selassie and Bowen 2002; Crawford et al. 2002).
Based on the findings outlined above, it has beenhypothesized that different cell types respond to sigma-1receptor activation in different ways (Novakova et al.1995, 1998). More recently, activation of sigma-1receptors by (+)-pentazocine has been shown to decreasePLC-mediated hypoglossal activity in isolated guinea-pigbrainstem preparations and to induce the translocation ofprotein kinase C to the plasma membrane via a novelmechanism of action of intracellular receptors (Morin-Surun et al. 1999). In a recent study, the changes in PLCactivity in different brain regions (including the striatumand the frontal cortex) have been described in response toeither single or repeated doses of the sigma-1 ligand E-5842 (Romero et al. 2000). In addition to changes in PLCactivity, levels of PLCβ and the associated G-proteinGq/11α were also upregulated in the frontal cortex of ratschronically treated with E-5842. Sigma receptors havealso been repeatedly linked to the modulation of proteinkinases, especially protein kinase C (Morin-Surun et al.1999; Derbez et al. 2002; Nuwayhid and Werling 2003)
Taken together, the data clearly suggest that classicalsecond messenger systems play a role in the action ofsigma ligands, although this probably occurs via amechanism different from the classical and well-describedones. Are there different types of sigma-1 receptors indifferent cell types? Is the sigma-1 binding proteinrecently cloned merely a component of multi-subunit
receptors? Is the sigma receptor only an auxiliary subunitof concrete ion channels? Further studies are required toanswer these and other important questions.
Sigma receptors as modulators
Cooperation between sigma-1 receptors and ion channelsmust be considered when characterizing the signal trans-duction pathways and cellular mechanisms by which thesereceptors function. Sigma receptors have been shown tomodulate various ion channels in several cell types (Morioet al. 1994; Soriani et al. 1999; Wilke et al. 1999a;Lupardus et al. 2000; Aydar et al. 2002; Karasawa et al.2002; Zhang and Cuevas 2002). Sigma receptors inhibitvoltage-gated potassium channels (Wilke et al. 1999b),and pentazocine and SKF 10,047 can modulate potassiumchannels in peptidergic nerve terminals despite intraterm-inal perfusion with GTPγS or GDPβS, suggesting that G-proteins do not play a role in these responses (Lupardus etal. 2000). Based on experiments performed in cell-attached patches, it has been reported that the sigmareceptor and ion channels must be in close vicinity tocooperate (Lupardus et al. 2000), which would argueagainst the existence of a classical mechanism by whichchannels are modulated through G-proteins and proteinphosphorylation (Wickman and Clapham 1995; Jonas andKaczmarek 1996). A recent report (Aydar et al. 2002) hasshown that sigma receptors associate with the voltage-gated potassium channel Kv1.4, which suggests thatprotein–protein interactions may play a role in sigmareceptor signal transduction.
In recent years, Su’s laboratory has presented elegantresults showing the molecular mechanism that governs theregulation of intracellular calcium signaling at the endo-plasmic reticulum through the participation of sigma-1receptors. It has been suggested that ternary complexesoccur as a result of interactions between the sigma receptorand various other proteins, including IP3 receptors andankyrin (Hayashi and Su 2001). According to this model,under the stimulation of the sigma-1 receptors by receptoragonists [cocaine or (+)-pentazocine or neurosteroids], thesigma-1 receptor/ankyrin complex would dissociate fromthe IP3 receptors and would be translocated to the plasmaor nuclear membranes. In the presence of a sigma-1receptor antagonist, this effect would be avoided andsigma-1 receptors would dissociate from ankyrin and IP3receptors which would stay on the endoplasmic reticulum.Following with this model, and based on the studiesperformed in NG-108 cells (Hayashi et al. 2000; Hayashiand Su 2001), in resting conditions, the dissociation of thesigma-1 receptor/ankyrin complex elicited by sigma-1agonists produces no detectable influence on calciumefflux from the endoplasmic reticulum and the cells needto be stimulated in the sense that appropriate levels of IP3are necessary. Under these conditions the dissociation ofthe sigma-1 receptor/ankyrin complex from IP3 receptorswould facilitate and indirectly promote the calcium effluxfrom these receptors (see a recent paper of Su and Hayashi
08/06/04 - PROOF PAGEMAKEUP 213_1920 7

2003 for a comprehensive review). Although this is a veryinteresting hypothesis, it still has to explain how theagonists and the antagonists of the sigma-1 receptors reachthe intracellular sites where the sigma-1 receptors/ankyrin/IP3 receptors are. This model implies that sigma-1 ligands,such as cocaine, (+)-pentazocine, or NE-100, are able toeasily cross the plasma membrane to reach the intracellularstores of sigma-1 receptors, but does not explain how.
Other experimental evidences also suggest that sigmareceptors play a role in modulating intracellular calciumlevels. In this sense, it was shown that calcium channelmodulators have differential effects on the binding of [3H]-DTG to sigma receptor sites (Rothman et al. 1991).Another study showed that sigma ligands can regulateintracellular calcium levels via modulation of the nicotinicacetylcholine receptor in chromaffin cells (Paul et al.1993). Catecholamine release induced by nicotine receptorstimulation was inhibited by (+)-pentazocine, haloperidol,and (−)-SKF 10,047.
Regulation of high-voltage-activated calcium channelsby sigma receptors has also been described in culturedhippocampal pyramidal neuron preparations, where in-creases in intracellular free calcium were blocked by sigmaligands (Church and Fletcher 1995). A similar result wasobserved in neuroprotection experiments using sigmaligands in rat primary cortical neurons, where sigma-mediated neuroprotection was shown to be mediatedthrough the modulation of intracellular calcium levels andvoltage-gated calcium channels (Klette et al. 1995). It hasbeen recently shown that sigma receptor agonists rapidlydepress peak calcium-channel currents, using the perfo-rated-patch configuration of the whole cell patch-clamprecording technique in neurons from the superior cervicalganglia (Zhang and Cuevas 2002).
It has also been suggested that sigma receptors canregulate glutamatergic transmission by functionally mod-ulating the NMDA receptor complex (Monnet et al. 1990,1992b; Ohno and Watanabe 1995; Bouchard et al. 1997;Maurice and Lockhart 1997; Gronier and Debonnel 1999).Iontophoretic or intravenous administration of DTG, asigma ligand, potentiates NMDA-induced firing of CA3pyramidal neurons (Monnet et al. 1992b). A similarpotentiation has been described using other sigma ligands(Debonnel and deMontigny 1996; Bergeron and Debonnel1997), generally considered to be agonists of the receptor.The effect was not observed in response to sigma-1antagonists, such as haloperidol, NE-100, or BMY 14802.Interestingly, most sigma agonists produce bell-shapeddose–response curves on the NMDA-induced effect(Couture and Debonnel 1998), and this has been suggestedto be due to the recruitment of different subtypes of sigmareceptors at higher doses (Bergeron and Debonnel 1997).Administration of the new selective sigma-2 ligand Lu 28-179 (Moltzen et al. 1995) dose dependently potentiated theNMDA response in the CA3 region of rat hippocampus,whereas ibogaine, a sigma-2 ligand with moderate affinityfor the receptor (Bowen et al. 1995b), slightly increasedthe NMDA response (Couture and Debonnel 1998). Toexplain such differences, it has been proposed that several
sigma-2 receptor subtypes may exist (Couture andDebonnel 1998). However, it is important to note thatthese effects are not exclusive to sigma-1 ligands or mixedsigma-1/sigma-2 ligands. Taken together, these electro-physiological studies suggest that one function of sigmareceptors is modulation of NMDA responses.
Anatomical distribution of sigma receptors
Early studies on the subcellular distribution of sigmareceptors indicated that [3H] (+)-SKF 10,047 binding siteswere enriched in the microsomal fraction of rat brain(McCann and Su 1990) and in non-synaptic areas of theplasma membrane (Roman et al. 1989b). Subcellularlocalization studies have shown that the sigma-1 receptoris primarily associated with neuronal perikarya anddendrites, and localized to the plasma membrane, themitochondrial membrane, and the endoplasmic reticulum(McLean and Weber 1988). Given this unusual distribu-tion, it has been proposed that, on activation, the sigma-1receptor is translocated from the endoplasmic reticulum tothe plasma membrane or to the nuclear membrane (Morin-Surun et al. 1999; Hayashi and Su 2001).
Different studies showed the particular distribution ofsigma-2 receptors inside the cell. These receptors havebeen proposed to be intracellular receptors (Bowen 2000)located in the membranes of the endoplasmic reticulumand mitochondria, and may, therefore, release calciumfrom these intracellular stores.
The anatomical distribution of sigma-1 receptors in thecentral nervous system has been well characterized. Sigmareceptors were first visualized using different radioligands,such as [3H] (+)-3-PPP, [3H]-SKF 10,047, [3H]-DTG, or[3H]NE-100 (Samovilova et al. 1988; Largent et al. 1986;Jansen et al. 1991; Okuyama et al. 1995a), and bindingsites were reported to be concentrated in limbic structures(including different areas of the hippocampus as thepyramidal and non-pyramidal layers, and the granularlayer of the gyrus dentatus), areas of the forebrainincluding the septum, the paraventricular nucleus of thehypothalamus, the anterodorsal thalamic nucleus, and indiscrete regions of the midbrain and hindbrain such as thedorsal raphe, the substantia nigra and the locus coeruleus,and in the cerebellum.
Recently, in situ hybridization and immunohistochem-ical techniques have been used to study sigma receptorlocalization (McLean and Weber 1988; Alonso et al. 2000;Zamanillo et al. 2000). Sigma receptors were found to bewidely distributed, although they seem to be concentratedin brain areas involved in motor functions, in limbic areas,sensory areas, and areas associated with endocrinefunctions (McCann et al. 1994).
A more detailed study was able to discriminate betweenthe two subtypes of sigma receptors and showed thatsigma-1 and sigma-2 receptors are differentially distrib-uted in the brain (Bouchard and Quirion 1997). By meansof autoradiographic studies using [3H]-DTG (with thepresence of (+)-pentazocine), Bouchard and Quirion
8 08/06/04 - PROOF PAGEMAKEUP 213_1920

(1997) showed the sigma-2 distribution in the rat brain.Only a few areas in the brain are particularly enriched withsigma-2 sites, these areas being implicated in the controlof posture and movement (Walker et al. 1990, 1994).
Using immunohistochemistry with a specific antibodyagainst the sigma-1 receptor, the distribution of thisreceptor in the adult rat central nervous system has beendescribed (Alonso et al. 2000). High levels of sigma-1immunostaining were associated with neurons in specificbrain regions, including the olfactory bulb, several hypo-thalamic nuclei, the septum, the central gray, certain motornuclei of the hindbrain, and the dorsal horn of the spinalcord. Intensely immunostained cells were observed in thehippocampus (especially in the dentate gyrus). Interest-ingly, in the dorso-lateral striatum only a small number ofweakly stained cells could be observed, whereas moder-ately immunostained cells were observed in the nucleusaccumbens.
Figure 3 shows the regional distribution of the sigma-1receptor as determined by means of Western blot using adifferent antibody (Zamanillo et al. 2002) than that used inthe immunohistochemical studies. The distribution of thesigma-1 receptor protein is compared with its mRNAdistribution.
In addition to the central nervous system, the sigma-1receptor is also widely distributed in peripheral organs. Inthe digestive tract (Samovilova and Vinogradov 1992),binding assays and autoradiographic studies showed thatsigma-1 receptors are enriched in the mucosal andsubmucosal regions, with less labeling in the muscularregions. Sigma-1 receptors are also present in the liver(Dumont and Lemaire 1991; Hellewell et al. 1994;Maurice et al. 1996), kidney (Hellewell et al. 1994),heart (Jansen et al. 1992; Ela et al. 1994), and sexualorgans (Jansen et al. 1992). In a similar way to the sigma-1receptor, sigma-2 receptor has also been found in differentorgans, including liver and kidney in high densities(Hellewell et al. 1994).
Potential therapeutic applications of sigma receptor ligands
Although further experiments are clearly needed todelineate the precise mechanisms of action and theendogenous ligand(s) of sigma receptors, a large numberof pharmacological, biochemical, and even therapeuticfunctions have been proposed for these receptors (Su andJunien 1994). Given the widespread distribution of sigmareceptors in the central nervous system, sigma ligands maybe useful in the treatment of different central disorders.These may eventually include modulation of mooddisorders, amnesic and cognitive deficits, the effectsproduced by drugs of abuse, tardive dyskinesia, potenti-ation of analgesia, and others. Moreover, in view of theirdistribution in peripheral organs, sigma receptors couldalso be implicated in gastrointestinal movements underconditions of stress, duodenal bicarbonate secretion,contraction of vas deferens, regulation of plasma levelsof corticosterone, prolactin, adrenocorticotropin, contrac-tility of cardiac cells, and many others.
While this article was being prepared, our laboratory,together with another group, produced a sigma-1 receptorknockout mouse (Langa et al. 2003). The study of theseanimals should help to clarify the role of sigma-1 receptorsin different physiological and pathophysiological pro-cesses where these receptors have been implicated.However, as these mice still express the sigma-2 receptor,they may also help to dissociate the involvement of sigma-1 and sigma-2 receptors in the manifestation of differentpathologies. The generation of these animals shouldundoubtedly start a new era of research in the field ofsigma receptors; but, although some phenotypical aspectshave already been disclosed (Langa et al. 2003), others arestill being studied (unpublished results from our labora-tory), and possible compensatory mechanisms to the lackof the sigma-1 receptor should not be discarded.
Sigma ligands and schizophrenia and movementdisorders
The first suggestion that sigma receptors might beinvolved in the pathophysiology of schizophrenia arosefrom the finding that synthetic ligands such as (+)-SKF10,047 exhibited psychotomimetic effects and from thefact that many neuroleptics exhibit high affinity for sigmareceptors. These findings led to speculation that sigmareceptors could play a role in the antipsychotic actions ofthese drugs (Gillgan and Tam 1994).
In addition to the role played by the central dopami-nergic system in schizophrenia, it is also important toconsider the glutamate system and the intricate relation-ship between dopamine, glutamate, and sigma receptors inthis disease. The role of NMDA in the pathogenesis ofschizophrenia has generated considerable interest becausePCP, an NMDA receptor channel-blocking agent, caninduce a schizophrenia-like psychosis in humans. Severalpost-mortem studies have shown abnormalities in gluta-mate concentrations and glutamate receptor densities in
Fig. 3 Regional distribution of the sigma-1 receptor in the rat brain.a Typical Western-blot image using an antibody specific for thesigma-1 receptor. Note the different relative distribution of thesigma-1 receptor protein. b Graph showing the distribution ofsigma-1 receptor mRNA in several brain areas as studied usingNorthern blot. FC frontal cortex, ST dorso-lateral striatum, CBcerebellum, PC parietal cortex, NA nucleus accumbens, HPhippocampus, LC locus coeruleus, DR dorsal raphe, VT ventraltegmental area, SN substantia nigra
08/06/04 - PROOF PAGEMAKEUP 213_1920 9

schizophrenic brains, and some biochemical and beha-vioral studies have suggested that antipsychotic drugs mayalter NMDA receptor functions.
However, there is no correlation between the affinity ofneuroleptic drugs for sigma receptors and their therapeuticefficacy. Thus, it has been suggested that sigma receptorsmay instead mediate the undesirable motor side effects ofantipsychotic drugs (Walker et al. 1990; Walker et al.1994). Moreover, the relationship between the ability ofneuroleptics to interact with sigma-1 and sigma-2receptors and their tendency to induce dystonic reactionsin humans has recently been studied (Matsumoto andPouw 2000). It has been suggested that the motor sideeffects induced by neuroleptics could be mediated throughboth sigma-1 and sigma-2 receptors.
Sigma receptors have been detected in cortical andlimbic structures in human post-mortem brain (Weissmanet al. 1998), and a selective loss of sigma sites inschizophrenia has been reported (Weissman et al. 1991).However, studies on polymorphisms of the sigma-1receptor gene and schizophrenia have yielded contra-dictory results (Ishiguro et al. 1998; Ohmori et al. 2000;Uchida et al. 2003). No selective sigma ligand has yetbeen marketed as an antipsychotic drug, but manydescriptions of novel putative antipsychotic compoundswith high affinity for the sigma receptor have beenreported (Chaki et al. 1994; Bartoszky et al. 1996;Takahashi et al. 1999).
Among these, [α-(4-fluorophenyl)-4-(5-fluoro-2-pyri-midinyl)-1-piperazine-butanol hydrochloride] (BMY14802) is a compound with high affinity for sigma,serotonin-1A, and serotonin-2 receptors, and a weakaffinity for dopamine and other neurotransmitter receptors(Taylor and Dekleva 1987). BMY 14802 was firstidentified as an antipsychotic candidate based on classicalneuropharmacological tests. BMY 14802 inhibits apomor-phine-induced climbing in mice (Taylor et al. 1993) andinhibits apomorphine-induced stereotypy in rats with lesspotency than that of haloperidol—a prototypical com-pound—but similar potency to that of clozapine. BMY14802 is also able to block sensitization to methamphet-amine (Ujike et al. 1992), and the development ofsensitization to cocaine (Ujike et al. 1996), which is apharmacological model of schizophrenia.
BMY 14802 exhibited a favorable pre-clinical profilesuggesting that it held promise as a drug that couldmediate antipsychotic effects in humans without produ-cing the untoward extrapyramidal side effects associatedwith standard neuroleptics. However, results of clinicaltrials showed no significant improvement in psychiatricsymptoms in response to BMY 14802 treatment (Gewirtzet al. 1994).
4-(4-Fluorophenyl)-1,2,3,6-tetrahydro-1-[4-(1,2,4-tria-zol-1-il)butyl] pyridine citrate (E-5842) is a compoundwith remarkable affinity for the sigma-1 receptor, and withlow-to-moderate affinity for other central nervous systemreceptors, including several dopamine, serotonin, andglutamate receptors (Guitart et al. 1998). E-5842 hasbeen widely tested as an antipsychotic at the pre-clinical
level. E-5842 inhibits apomorphine-induced climbing inmice and inhibits the conditioned avoidance response inrats with ED50 values different from those of dopamine D2
antagonists such as haloperidol (also a sigma receptorligand) and risperidone (Guitart et al. 1998). E-5842 alsodose dependently inhibits amphetamine-induced locomo-tor activity in rats, with no apparent induction of catalepsy.Acute administration of E-5842 also increases the expres-sion of Fos protein in selected regions of the rat forebrain(Guitart and Farré 1998), similar to what has been shownwith panamesine (EMD 57455), a selective sigma receptorligand that has been tested as a putative antipsychotic inclinical trials. In the case of E-5842, Fos levels wereincreased in the prefrontal cortex and the nucleusaccumbens, with little effect in the dorso-lateral striatum,clearly suggesting the possibility that this sigma com-pound may exert antipsychotic activity.
Sigma receptors may also interact with or modulate thefunction of glutamatergic receptors. Repeated administra-tion of E-5842 alters immunoreactivity levels of varioussubunits of the ionotropic glutamate receptors (Guitart etal. 2000), an effect that had been previously described forother antipsychotic drugs (Fitzgerald et al. 1995; Tasceddaet al. 1999), but not for any known sigma-1 receptorligands.
N,N-dipropyl-2-[4-methoxy-3-(2-phenylethoxy)-phe-nyl]-ethylamine monohydrochloride (NE-100) is anotherputative antipsychotic compound with high affinity for thesigma-1 receptor, and lower affinity for the sigma-2,dopamine, serotonin, and PCP receptors (Okuyama et al.1993; Chaki et al. 1994). NE-100 antagonizes thebehavioral effects of PCP and does not induce catalepsyin rats (Okuyama et al. 1993, 1994, 1995b). NE-100 andother sigma-1 antagonists have also been shown toregulate dopamine release through the NMDA receptor(Chaki et al. 1998) or to modulate NMDA-inducedcurrents in dopaminergic neurons. Together with the datafrom studies of E-5842 (Guitart et al. 1998), experimentalevidence clearly suggests that sigma compounds canmodulate both dopaminergic and glutamatergic neuro-transmission.
Remoxipride has considerable binding affinity for thesigma receptor and has been shown to exhibit atypicalantipsychotic activity in humans. However, subchronicadministration of remoxipride did not affect either thedensity or the affinity of sigma-1 receptors in the rat brain(Ericson and Ross 1992), as determined by [3H] (+)-3-PPPradioligand binding. This fact is quite remarkable, giventhat haloperidol, which is also a sigma-1 receptor antag-onist, has been reported to decrease sigma-1 bindingwithout affecting sigma-1 mRNA levels in rat and guinea-pig brain (Inoue et al. 2000). Furthermore, repeatedadministration of E-5842 has been shown to increasesigma-1 receptor mRNA levels in several brain areas(Zamanillo et al. 2000).
Over the last decade, other compounds with highaffinity for the sigma receptor have been evaluated forpossible use in the treatment of schizophrenia. NPC16377, MS-377, SR-31742A, and some other compounds
10 08/06/04 - PROOF PAGEMAKEUP 213_1920

mentioned above have been described as potent andselective sigma ligands with antipsychotic potential. Someof these, like remoxipride, BMY 14802, or SR-31742A,have been discontinued, whereas panamesine and SL82.0715 exhibited favorable efficacy in open clinicalstudies (Frieboes et al. 1997).
Along with the putative beneficial effects of certainsigma ligands as antipsychotic drugs, it has beenhypothesized that sigma ligands could also contribute tothe various undesirable motor side effects observed afterlong-term treatment with antipsychotic drugs. There isanatomical evidence supporting a role for sigma receptorsin the motor side effects of certain neuroleptics (Walkerand Martin 1994). Brainstem nuclei that control musclesthat mediate the motor side effects of neuroleptics areenriched in sigma receptors (Bouchard and Quirion 1997),and as mentioned previously, substantial levels of sigmareceptors are found in the basal ganglia (Gundlach et al.1986). Furthermore, brain regions such as the cerebellumand the red nucleus, which are clearly related to dystonicreactions, are also enriched in sigma receptors (McLeanand Weber 1988; Bouchard and Quirion 1997). In earlystudies, it was shown that selective sigma receptoragonists may elicit dystonic reactions when directlyinjected into the red nucleus (Matsumoto et al. 1990),whereas injection of non-sigma ligands failed to inducesuch reactions (Matsumoto et al. 1990, 1995). The abilityof some sigma receptor antagonists to attenuate orofacialdyskinesias and dystonic reactions in animals suggests thatthese drugs may hold promise as potential therapeuticapproaches to prevent such undesired movements (Matsu-moto et al. 1995; Tran et al. 1998). The sigma-2 receptorseems to play the most important role in controlling motorfunction (Walker et al. 1993). Conversely, the sigma-1receptor was thought to be less important (McCraken et al.1999). Nevertheless, a subsequent study comparing neu-roleptic binding to the sigma-1 and sigma-2 receptors andthe induction of dystonic reactions (Matsumoto and Pouw2000) suggested that sigma-1 receptors also play a role inmotor functions. Other studies seem to confirm theimportance of both receptors in the regulation of dystonicreactions (Okumura et al. 1996; Ghelardini et al. 2000;Yoshida et al. 2000).
Sigma ligands and drugs of abuse
Drug addiction is among the most serious problems facingsociety today, and it is clear that the psychological aspectsof addiction are mediated via neurobiological mechanismsin the brain (Nestler 2001). Unfortunately, there arecurrently no effective pharmacological treatments for theabuse of cocaine, D-amphetamine, and other abused drugs.
As it has been described below, the central dopaminer-gic neurotransmission systems that are likely involved inthe pathophysiology of schizophrenia may be modulatedby sigma receptors. Central dopaminergic systems alsorepresent, at least in part, the neuroanatomical substrate ofdrug addiction. Sigma-1 receptors have been hypothesized
to be involved in the effects of cocaine, given the affinityof cocaine for the receptor (Sharkey et al. 1988; Ritz andGeorge 1993; Matsumoto et al. 2001; Su and Hayashi2001; Maurice et al. 2002). Selective sigma-1 receptorantagonists such as BMY 14802, BD1047, and BD1063(Menkel et al. 1991; McCraken et al. 1999) decrease thelocomotor effects of cocaine, which has also beenobserved when mice are treated with antisense oligodeox-ynucleotides corresponding to the sequence of sigma-1receptors (Matsumoto et al. 2001, 2002). In another set ofexperiments, it has been shown that sigma-1 antagonistsmay attenuate the behavioral sensitization induced bycocaine (Ujike et al. 1996). It has also been shown thatintraperitoneal administration of sigma-1 antagonists, suchas NE-100 and BD1047, significantly attenuated orblocked the cocaine-induced conditioned place preference,a paradigm used to evaluate the rewarding properties ofthis drug and other drugs of abuse (Romieu et al. 2000).Changes in gene expression induced by cocaine have alsobeen reported in the brains of mice. The upregulation bycocaine of three genes identified as the fos-related antigen-2, the G-protein coupled receptor 27 and the ataxiatelangiectasia murine homolog was prevented by the useof the sigma-1 antagonist BD1063, suggesting that someprotective actions of the sigma receptor antagonistsinvolve transcriptional mechanisms (Matsumoto et al.2003).
When taken together, these data support the idea thatactivation of sigma-1 receptors by cocaine can beimportant for both the short-term and long-term effectsof the drug. The data also suggest that sigma-1 receptorsrepresent a useful target for the development of futuretherapeutic strategies (see Maurice et al. 2002; Matsumotoet al. 2003 for comprehensive reviews). However, a directrole of sigma-1 receptors in other aspects of cocaineaddiction has not been established.
A role for sigma-1 receptors in the actions of otherpsychostimulant drugs has also been proposed. Repeatedadministration of methamphetamine increased [3H] (+)-pentazocine binding in rat brain (Itzhak 1993). Sigmaligands also block the development of behavioral sensi-tization induced by repeated administration of metham-phetamine (Ujike et al. 1992; Takahashi et al. 2000). Othersigma-1 receptor antagonists, such as NPC 16377 and E-5842, have also been reported to partially block D-amphetamine-stimulated locomotor activity (Clissold et al.1993; Guitart et al. 1998).
The role of sigma-2 receptors in the effects of drugs ofabuse is less clear, and the investigation of their roleopposing the actions of cocaine is complicated by the lackof an amino acid sequence of the receptor that can be usedto construct useful antisense oligodeoxynucleotides. How-ever, the sigma-2 receptor system has been shown tomodulate amphetamine-stimulated dopamine release invitro (Izenwasser et al. 1998; Weatherspoon and Werling1999). More recently, it has been shown that SM-21, anovel sigma-2 antagonist (Ghelardini et al. 2000), canattenuate the locomotor-stimulating effects in cocaine inmice (Matsumoto and Mack 2001), although the mecha-
08/06/04 - PROOF PAGEMAKEUP 213_1920 11

nism of action of SM-21 has yet not been fullycharacterized
Memory and attention deficits
It has been proposed that sigma agonists have beneficialeffects in several models of amnesia. Some sigma-1ligands have been shown to prevent or reverse scopol-amine-induced amnesia in rats and mice (Earley et al.1991; Matsuno et al. 1994). Receptor agonists increaseacetylcholine release in both the hippocampus and thefrontal cortex (Matsuno et al. 1993, 1995) and maypotentiate several NMDA-evoked responses in selectedregions of the hippocampus (Hong and Werling 2000).SA4503 is a highly selective sigma-1 receptor agonist thathas been shown to reverse scopolamine-induced amnesiaand to attenuate learning impairments in animals withcortical cholinergic dysfunctions (Matsuno et al. 1997;Senda et al. 1998a). However, SA4503 protects retinalcells against glutamate-induced neurotoxicity (Senda et al.1998b). SA4503 has also been shown to attenuate deficitsin spatial working and reference memory induced bydizocilpine in the delay-interposed radial maze task (Zouet al. 2000). In a similar way, PRE-084, a selective sigma-1 receptor agonist, reduces MK-801-induced learningimpairments in mice (Maurice et al. 1994).
Neuroactive steroids may also play a role in themodulation of memory by sigma receptors. Neuroactivesteroids may modulate GABAA receptor function andvarious NMDA-evoked responses, and can also interactwith sigma receptors (Maurice et al. 1999, 2001).Pregnenolone, dehydroepiandrosterone (DHEA), and pro-gesterone exhibit significant affinity for the sigma-1receptor. Pregnenolone, DHEA, and their sulfate estershave also been shown to enhance memory retentionproperties in learning-impaired mice (Flood et al. 1992;Maurice et al. 1997). In another set of experiments, it wasshown that sigma-1 agonists, such as PRE-084, SA4503,DHEA, pregnenolone, and their sulfate esters, attenuatedthe β25–35-amyloid peptide-induced learning impairmentin mice by interacting with sigma-1 receptors (Maurice etal. 1997, 1998). In these experiments, progesteronebehaved as an antagonist, blocking the effects ofneuroactive steroids, and some sigma-1 agonists.
Depression and anxiety
Recent studies have identified novel putative antidepres-sant compounds that have little, if any, effect on endog-enous biogenic amine levels. Sigma receptor agonists arebeing considered as such putative antidepressants (Matsu-no et al. 1996a; Ukai et al. 1998; Sanchez and Papp 2000).Selective sigma-1 receptor agonists such as (+)-pentazo-cine, (+)-SKF 10,047, or SA4503 decrease the duration ofimmobility in the forced swimming test and in the tailsuspension test (Matsuno et al. 1996b; Ukai et al. 1998;Urani et al. 2001), two experimental models widely used
to assess the antidepressant activity of new compounds.The sigma receptor agonist OPC-14523 (Oshiro et al.2000; Tottori et al. 2001) and the selective sigma-1receptor agonist igmesine (JO 1784) also show antide-pressant-like activity in pre-clinical animal models(Roman et al. 1990; Akunne et al. 2001). In a recentstudy (Urani et al. 2002), it has been shown that theantidepressant-like activity of igmesine requires modula-tion of intracellular calcium mobilization. In line withwhat was referred in the previous section, neurosteroidshave also been suggested to have a beneficial effect ondepressive states through a sigma-1 receptor-mediatedeffect (Maurice et al. 1996; see also Maurice et al. 2001;Van Broekhoven and Verkes 2003, for reviews).
The selective sigma-2 receptor ligand Lu 28-179(siramesine) exhibits antidepressant activity in the chronicmild stress model of depression (Sanchez and Papp 2000).In another study, the effects of Lu 28-179 were assessed invarious animal models of anxiety in rodents. Lu 28-179facilitated the exploratory behavior of mice and rats in theblack and white two-compartment box. Repeated treat-ment with Lu 28-179 significantly increased the durationof social interaction compared with controls, and noanxiogenic-like effects were observed on withdrawal.Furthermore, Lu 28-179 reversed shock-induced suppres-sion of drinking in rats (Sanchez et al. 1997). Lu 28-179 iscurrently under development in clinical trials as a potentialtreatment for anxiety (Heading 2001). It was recentlyreported that (+)-pentazocine and the antidepressantsimipramine and fluvoxamine, which have activity atsigma-1 receptors, may enhance the nerve growth factor-induced neurite sprouting in PC12 cells (Takebayashi et al.2002). This is of interest in that it has been proposed thatthe therapeutic actions of antidepressants may involveneurite sprouting.
Pain
A possible role for sigma receptors in antinociception wassuggested from studies that showed a relationship betweensigma systems and opioid analgesia (Chien and Pasternak1993, 1994). In this sense, an antiopioid sigma-1 systemhas been proposed to exist in mice and rats, where sigma-1receptor antagonists would potentiate opioid analgesia(Chien and Pasternak 1993, 1995). Using the tail-flickassay, it has been shown that sigma-1 antagonists, such ashaloperidol, have no effect on tail-flick latencies whengiven alone, but significantly increase the analgesicresponse to morphine. Haloperidol also increases tail-flick latencies in response to treatment with selectivekappa receptor agonists with low intrinsic analgesicactivity, whereas the sigma-1 agonist (+)-pentazocinediminishes this tail-flick latency.
In a series of elegant experiments, the specificity ofmodulation of opioid analgesia by sigma-1 receptors wasassessed using antisense oligodeoxynucleotides to down-regulate sigma-1 receptors (King et al. 1997; Mei andPasternak 2002). Mice that received antisense treatment
12 08/06/04 - PROOF PAGEMAKEUP 213_1920

showed a markedly enhanced response to both kappareceptor drugs and morphine, similar to the effects ofhaloperidol.
Conclusions and final remarks
Sigma receptors have been described for more than25 years and, in some ways, they remain controversial.Although considerable effort has been directed towardsdefining the intracellular pathways that mediate sigmareceptor signaling, and some second messenger system hasbeen associated with sigma receptors (calcium, IP3, PKC),the possible intracellular pathways have not been fullyworked out. Neither has any endogenous sigma receptorligand yet been discovered, despite considerable effort inthis area. However, the landscape is becoming clearer asmore information on the biology, pharmacology, andfunction of these receptors becomes available. An increas-ing body of evidence suggests that the sigma receptor maybe unique, in that it may modify synaptic transmissionthrough the regulation of other neurotransmitter receptors.Thus, sigma receptor ligands may modulate glutamate anddopamine release. Several years ago, when the hypothe-tical roles of sigma receptors were first being identified,the possible therapeutic implications of sigma ligandswere merely speculative. Today, it seems that sigmareceptors may be involved in many neuronal processes, aswell as in the pathophysiology of certain psychiatricdisorders, including depression, schizophrenia, motordisturbances, pain, drug addiction, attention deficitdisorders, and probably cancer (an issue that has notbeen addressed in this review). Identification of theputative endogenous ligand(s) for sigma receptors shouldgreatly help to clarify the picture. Despite this gap in ourknowledge, many significant advances have taken placeover the past several years in the field of sigma receptorresearch. The generation of knockout mice for the sigma-1receptor has recently been reported, and these animalsshould undoubtedly start a new era of research in the fieldof sigma receptors. Altogether, these advances mayeventually lead to the therapeutic use of sigma receptorligands for treatment and/or prevention of some of thesedevastating human psychiatric disorders.
References
Akunne HC, Zoski KT, Whetzel SZ, Cordon JJ, Brandon RM,Roman F, Pugsley TA (2001) Neuropharmacological profile ofa selective sigma ligand igmesine: a potential antidepressant.Neuropharmacology 41:138–149
Alonso G, Phan V-L, Guillemain I, Saunier M, Legrand A, AnoalM, Maurice T (2000) Immunocytochemical localization of thesigma1 receptor in the adult rat central nervous system.Neuroscience 97:155–170
Ault DT, Werling LL (1997) Differential modulation of NMDA-stimulated [3H]dopamine release from rat striatum by neuro-peptide Y and sigma receptor ligands. Brain Res 760:210–217
Ault DT, Werling LL (1998) Neuropeptide Y-mediated enhancementof NMDA-stimulated [3H]dopamine release from rat prefrontalcortex is reversed by sigma1 receptor antagonists. SchizophrRes 31:27–36
Aydar E, Palmer CP, Klyachko VA, Jackson MB (2002) The sigmareceptor as a ligand-regulated auxiliary potassium channelsubunit. Neuron 34:399–410
Bartoszky GD, Bender HM, Hellman J, Schnorr C, Seyfred CA(1996) EMD 57445: a selective sigma ligand with the profile ofan atypical neuroleptic. CNS Drug Rev 2:175–194
Basile AS, Paul I, Mirchevich A, Kuijpers G, de Costa B (1992)Modulation of (+)-[3H]pentazocine binding to guinea pigcerebellum by divalent ions. Mol Pharmacol 42:882–889
Beart PM, O’Shea RD, Manallack DT (1989) Regulation of sigma-receptors: high- and low-affinity agonist states GTP shifts andup-regulation by rimcazole and 13-Di(2-tolyl) guanidine. JNeurochem 53:779–788
Bergeron R, Debonnel G (1997) Effects of low and high doses ofselective sigma ligands: further evidence suggesting theexistence of different subtypes of sigma receptors. Psycho-pharmacology 129:215–224
Bouchard P, Quirion R (1997) [3H]1,3-di(2-tolyl)guanidine and [3H](+)pentazocine binding sites in the rat brain: autoradiographicvisualization of the putative sigma 1 and sigma 2 receptorsubtypes. Neuroscience 76:467–477
Bouchard P, Maurice T, St-Pierre S, Privat A, Quirion R (1997)Neuropeptide Y and the calcitonin gene-related peptide atten-uate learningimpairments induced by MK-801 via a sigmareceptor-related mechanism. EurJ Neurosci 9:2142–2152
Bowen WD (1994) Interaction of sigma receptors with signaltransduction pathways and effects on second messengers. In:Itzhak Y (ed) Sigma receptors. Academic, San Diego, pp 139–170
Bowen WD (2000) Sigma receptors: recent advances and newclinical potentials. Pharm Acta Helv 74:211–218
Bowen WD, Kirschner BN, Newman AH, Rice KC (1988) Sigmareceptors negatively modulate agonist-stimulated phosphoino-sitide metabolism in rat brain. Eur J Pharmacol 177:111–118
Bowen WD, Walker JM, de Costa BR, Wu R, Tolentino PJ, Finn D,Rothman RB, Rice KC (1992) Characterization of theenantiomers of cis-N-[2-(34-dichlorophenyl)ethyl]-N-methyl-2-(1-pyrrolidinyl)cyclohexylamine (BD737 and BD738):novel compounds with high affinity selectivity and biologicalefficacy at sigma receptors. J Pharmacol Exp Ther 262:32–40
Bowen WD, de Costa BR, Hellewell SB, Walker JM, Rice KC(1993) [3H](+)-Pentazocine: a potent and highly selectivebenzomorphan-based probe for sigma-1 receptors. Mol Neuro-pharmacol 3:117–126
Bowen WD, Bertha CM, Vilner BJ, Rice KC (1995a) CB-64D andCB-184: ligands with high sigma-2 receptor affinity andsubtype selectivity. Eur J Pharmacol 278:257–260
Bowen WD, Vilner BJ, Williams W, Bertha CM, Kuehne ME,Jacobson AE (1995b) Ibogaine and its congeners are sigma 2receptor-selective ligands with moderate affinity. Eur J Phar-macol 279:R1–R3
Bowen WD, Vilner BJ, Bandarage UK, Kuehne ME (1996)Ibogaine and ibogamine modulate intracellular calcium levelsvia interaction with sigma-2 receptors. Soc Neurosci Abstr 22(7875):2006
Bowlby MR (1993) Pregnenolone sulphate potentiation of N-methyl-D-aspartate receptor channels in hippocampal neurons.Mol Pharmacol 43:813–819
Brady KT, Balster RL, May EL (1982) Stereoisomers of N-allylnormetazocine: phencyclidine-like behavioral effects insquirrel monkeys and rats. Science 215:178–180
Chaki S, Tanaka M, Muramatsu M, Otomo S (1994) NE-100, anovel potent sigma ligand preferentially binds to sigma 1binding sites in guinea pig brain. Eur J Pharmacol 251:R1–R2
Chaki S, Okuyama S, Ogawa S, Tomisawa K (1998) Regulation ofNMDA-induced [3H]dopamine release from rat hippocampalslices through sigma-1 binding sites. Neurochem Int 33:29–34
08/06/04 - PROOF PAGEMAKEUP 213_1920 13

Chien C-C, Pasternak GW (1993) Functional antagonism ofmorphine analgesia by (+)-pentazocine: evidence for an anti-opioid σ1 system. Eur J Pharmacol 250:R7–R8
Chien C-C, Pasternak GW (1994) Selective antagonism of opioidanalgesia by a sigma system. J Pharmacol Exp Ther 271:1583–1590
Chien C-C, Pasternak GW (1995) Sigma antagonists potentiateopioid analgesia in rats. Neurosci Lett 190:137–139
Church J, Fletcher EJ (1995) Blockade by sigma site ligands of highvoltage-activated Ca2+ channels in rat and mouse culturedhippocampal pyramidal neurons. Br J Pharmacol 116:2801–2810
Clissold DB, Pontecorvo MJ, Jones BE, Abreu ME, Karbon EW,Erikson RH, Natalie KJ Jr, Borosky S, Hartman T, MansbachRS, Balster RL, Ferkany JW, Enna SJ (1993) NPC 16377: apotent and selective σ-ligand. II. Behavioral and neuroprotec-tive profile. J Pharmacol Exp Ther 265:876–886
Connick JH, Hanlon G, Roberts J, France L, Fox PK, Nicholson CD(1992) Multiple sigma binding sites in guinea-pig and rat brainmembranes: G-protein interactions. Br J Pharmacol 107:726–731
de Costa BR, He X-S (1994) Structure–activity relationships andevolution of sigma receptor ligands (1976-present). In: Itzhak Y(ed) Sigma receptors. Academic, San Diego, pp 45–111
Couture S, Debonnel G (1998) Modulation of the neuronal responseto N-methyl-D-aspartate by selective sigma 2 ligands. Synapse29:62–71
Craviso GL, Musacchio JM (1983) High affinity dextromethorphanbinding sites in guinea pig. II. Competition experiments. MolPharmacol 23:629–640
Crawford KW, Coop A, Bowen WD (2002) σ2 Receptors regulatechanges in sphingolipid levels in breast tumor cells. Eur JPharmacol 443:207–209
Cutts JM, de Costa BR, Bowen WD (1993) Sigma ligands havereduced ability to inhibit the muscarinic phosphoinositideresponse in cells deficient in sigma-1 receptors. In: Harris LS(ed) Problems of Drug Dependence 1992. Proceedings of the54th annual scientific meeting, National Institute of DrugAbuse Research Monograph 132, US Government PrintingOffice, p 403
Debonnel G (1993) Current hypotheses on sigma receptors and theirphysiological role: possible implications in psychiatry. JPsychiatry Neurosci 18:157–172
Debonnel G, deMontigny C (1996) Modulation of NMDA anddopaminergic neurotransmission by sigma ligands: possibleimplications for the treatment of psychiatric disorders. Life Sci58:721–734
DeHaven-Hudkins DL, Fleissner LC (1992) Competitive interac-tions at [3H]-13-(2-tolyl)guanidine (DTG)-defined sigma rec-ognition sites in guinea pig brain. Life Sci 50:PL65–PL70
DeHaven-Hudkins DL, Fleissner LC, Ford-Rice FY (1992)Characterization of the binding of [3H](+)-pentazocine tosigma recognition sites in guinea pig brain. Eur J Pharmacol227:371–378
Derbez AE, Mode RM, Werling LL (2002) σ2-Receptor regulationof dopamine transporter via activation of protein kinase C. JPharmacol Exp Ther 301:306–314
Dumont M, Lemaire S (1991) Interaction of 1,3-di(2-[5-3H]tolyl)guanidine with σ-2 binding sites in rat heart membranepreparations. Eur J Pharmacol 209:245–248
Earley B, Burke M, Leonard BE, Gouret CJ, Junien JL (1991)Evidence for an anti-amnesic effect of JO 1784 in the rat: apotent and selective ligand for the sigma receptor. Brain Res546:282–286
Ela C, Barg J, Vogel Z, Hasin Y, Eliam Y (1994) Sigma receptorligands modulate contractility Ca++ influx and beating rate incultured cardiac myocytes. J Pharmacol Exp Ther 269:1300–1309
Ela C, Hasin Y, Eilam Y (1996) Apparent desensitization of a sigmareceptor sub-population in neonatal rat cardiac myocytes bypre-treatment with sigma receptor ligands. Eur J Pharmacol295:275–280
Ericson H, Ross SB (1992) Subchronic treatment of rats withremoxipride fails to modify σ binding sites in the brain. Eur JPharmacol 226:157–161
Fitzgerald LW, Deutch AY, Gasic G, Heinemann SF, Nestler EJ(1995) Regulation of cortical and subcortical glutamate recep-tor subunit expression by antipsychotic drugs. J Neurosci15:2453–2461
Flood JF, Morley JE, Roberts E (1992) Memory-enhancing effectsin male mice of pregnenolone and steroids metabolicallyderived from it. Proc Natl Acad Sci U S A 89:1567–1571
Frieboes RM, Murck H, Wiedemann K, Holsboer F, Steiger A(1997) Open clinical trial of the sigma ligand panamesine inpatients with schizophrenia. Psychopharmacology 132:82–88
Gebreselassie D, Bowen WD (2002) σ2 Receptors are componentsof sphingolipid/cholesterol-rich membrane rafts. Soc NeurosciAbstr 235.20
Gewirtz GR, Gorman JM, Volavka J, Macaluso J, Gribkoff G,Taylor DP, Borison R (1994) BMY 14802, a sigma receptorligand for the treatment of schizophrenia. Neuropsychophar-macology 10:37–40
Ghelardini C, Galeotti N, Bartolini A (2000) Pharmacologicalidentification of SM-21, the novel sigma(2) antagonist. Phar-macol Biochem Behav 67:659–662
Gillgan PJ, Tam SW (1994) Sigma receptor ligands: potential drugsfor the treatment of CNS disorders? Drug News Perspect 7:13–18
Gonzalez-Alvear GM, Werling LL (1995) Sigma1 receptors in ratstriatum regulate NMDA-stimulated [3H]dopamine release viaa presynaptic mechanism. Eur J Pharmacol 294:713–719
Gore AC (2001) Gonadotropin-releasing hormone neurons NMDAreceptors and their regulation by steroid hormones across thereproductive life cycle. Brain Res Rev 37:235–248
Gronier B, Debonnel G (1999) Involvement of sigma receptors inthe modulation of the glutamatergic/NMDA neurotransmissionin the dopaminergic systems. Eur J Pharmacol 368:183–196
Grunder G, Muller MJ, Andreas J, Heydari N, Wetzel H, SchlosserR, Schlegel S, Nickel O, Eissner D, Benkert O (1999)Occupancy of striatal D(2)-like dopamine receptors aftertreatment with the sigma ligand EMD 57445: a putativeatypical antipsychotic. Psychopharmacology 146:81–86
Guitart X, Farré AJ (1998) The effect of E-5842, a σ receptor ligandand potential atypical antipsychotic on Fos expression in ratforebrain. Eur J Pharmacol 363:127–130
Guitart X, Codony X, Ballarín M, Dordal A, Farré AJ (1998) E-5842: a new potent and preferential sigma ligand: preclinicalpharmacological profile. CNS Drug Rev 4:201–224
Guitart X, Méndez R, Ovalle S, Andreu F, Carceller A, Farré AJ,Zamanillo D (2000) Regulation of ionotropic glutamatereceptor subunits in different rat brain areas by a preferentialsigma1 receptor ligand and potential atypical antipsychotic.Neuropsychopharmacology 23:539–546
Gundlach AL, Largent BL, Snyder SH (1986) Autoradiographiclocalization of sigma receptor binding sites in guinea pig andrat central nervous system with (+)3H-3-(3-hydroxyphenyl)-N-(1-propyl)-piperidine. J Neurosci 6:642–647
Haertzen CA (1970) Subjective effects of narcotic antagonistscyclazocine and nalorphine on the addiction research centerinventory (ARCI). Psychopharmacologia 18:366–377
Hanner M, Moebius FF, Weber F, Grabner M, Striessnig J,Glossmann (1995) Phenylalkylamine Ca2+ antagonist bindingprotein. Molecular cloning, tissue distribution and heterologousexpression. J Biol Chem 271:7551–7557
Hanner M, Moebius FF, Flandorfer A, Knaus HG, Striessnig J,Kempner E, Glossmann H (1996) Purification molecularcloning and expression of the mammalian sigma1-bindingsite. Proc Natl Acad Sci U S A 93:8072–8077
Hayashi T, Su T-P (2001) Regulating ankyrin dynamics: roles ofsigma-1 receptors. Proc Natl Acad Sci U S A 98:491–496
14 08/06/04 - PROOF PAGEMAKEUP 213_1920

Hayashi T, Kagaya A, Takebayashi M, Shimizu M, Uchitomi Y,Motohashi N, Yamawaki S (1995) Modulation by sigmaligands of intracellular free Ca++ mobilization by N-methyl-D-aspartate in primary culture of rat frontal cortical neurons. JPharmacol Exp Ther 275:207–214
Hayashi T, Maurice T, Su TP (2000) Ca (2+) signaling via sigma(1)-receptors: novel regulatory mechanism affecting intracellularCa (2+)concentration. J Pharmacol Exp Ther 293:788–798
Heading C (2001) Siramesine. Curr Opin Investig Drugs 2:266–270Hellewell SB, Bowen WD (1990) A sigma-like binding site in rat
pheochromocytoma (PC12) cells: decreased affinity for (+)-benzomorphans and lower molecular weight suggest a differentsigma receptor form from that in guinea pig brain. Brain Res527:244–253
Hellewell SB, Bruce A, Feinstein G, Orringer J, Williams W, BowenWD (1994) Rat liver and kidney contain high densities ofsigma-1 and sigma-2 receptors: characterization by ligandbinding and photoaffinity labeling. Eur J Pharmacol MolPharmacol Sect 268:9–18
Hong W, Werling LL (2000) Evidence that the sigma(1) receptor isnot directly coupled to G proteins. Eur J Pharmacol 408:117–125
Hong W, Werling LL (2001) Lack of effects by s ligands onneuropeptide Y-induced G-protein activation in rat hippocam-pus and cerebellum. Brain Res 901:208–218
Inoue A, Sugita S, Shoji H, Ichimoto H, Hide I, Nakata Y (2000)Repeated haloperidol treatment decreases σ1 receptor bindingbut does not affect its mRNA levels in the guinea pig or ratbrain. Eur J Pharmacol 401:307–316
Ishiguro H, Ohtsuki T, Toru M, Itokawa M, Aoki J, Shibuya H,Kurumaji A, Okubo Y, Iwawaki A, Ota K, Shimizu H,Hamaguchi H, Arinami T (1998) Association between poly-morphisms in the type 1 sigma receptor gene and schizophre-nia. Neurosci Lett 257:45–48
Itzhak Y (1993) Repeated methamphetamine-treatment alters brainsigma receptors. Eur J Pharmacol 230:243–244
Itzhak Y (1994) Multiple sigma binding sites in the brain. In: ItzhakY (ed) Sigma receptors. Academic, San Diego, pp 113–137
Iwamoto ET (1981) Locomotor activity and antinociception afterputative mu, kappa and sigma opioid receptor agonists in therat: influence of dopaminergic agonists and antagonists. JPharmacol Exp Ther 217:451–460
Izenwasser S, Thompson-Montgomery D, Deben SE, ChowdhuryIN, Werling LL (1998) Modulation of amphetamine-stimulated(transporter-mediated) dopamine release in vitro by σ2 receptoragonists and antagonists. Eur J Pharmacol 346:189–196
Jansen KLR, Faull RLM, Dragunow M, Leslie RA (1991)Autoradiographic distribution of sigma receptors in humanneocortex hippocampus basal ganglia cerebellum pineal andpituitary glands. Brain Res 559:172–177
Jansen KL, Elliot M, Leslie RA (1992) Sigma receptors in rat brainand testes show similar reductions in response to chronichaloperidol. Eur J Pharmacol 214:281–283
Jonas EA, Kaczmarek LK (1996) Regulation of potassium channelsby protein kinases. Curr Opin Neurobiol 6:318–323
Karasawa J, Yamamoto H, Yamamoto T, Sagi N, Horikomi K, Sora I(2002) MS-377, a selective sigma receptor ligand indirectlyblocks the action of PCP in the N-methyl-D-aspartate receptorion-channel complex in primary cultured rat neuronal cells. LifeSci 70:1631–1642
Kekuda R, Prasad PD, Fei YJ, Leibach FH, Ganapathy V (1996)Cloning and functional expression of the human type 1 sigmareceptor (hSigmaR 1). Biochem Biophys Res Commun229:553–558
King MA, Pan Y-X, Mei J, Chang A, Xu J, Pasternak GW (1997)Enhanced kappa opioid analgesia by antisense targeting the σ1receptor. Eur J Pharmacol 331:R5–R7
Klein MD, Musacchio JM (1989) High affinity dextromethorphanbinding sites in guinea pig brain. Effect of sigma ligands andother agents. J Pharmacol Exp Ther 251:207–215
Klein MD, Musacchio JM (1994) Effects of cytochrome P450ligands on the binding of [3H]dextrometorphan and sigmaligands to guinea-pig brain. In: Itzhak Y (ed) Sigma receptors.Academic, San Diego, pp 243–262
Klette KL, DeCoster MA, Moreton JE, Tortella FC (1995) Role ofcalcium in sigma-mediated neuroprotection in rat primarycortical neurons. Brain Res 704:31–41
Klette KL, Lin Y, Clapp LE, DeCoster MA, Moreton JE, Tortella FC(1997) Neuroprotective sigma ligands attenuate NMDA andtrans-ACPD-induced calcium signaling in rat primary neurons.Brain Res 756:231–240
Koe BK, Burkhart CA, Lebel LA (1989) (+)-[3H]3-(3-hydroxyphe-nyl)-N-(1-propyl)-piperidine binding to σ receptors in mousebrain in vivo. Eur J Pharmacol 161:263–266
Lambert JJ, Harney SC, Belelli D, Peters JA (2001) Neurosteroidmodulation of recombinant and synaptic GABAA receptors. IntRev Neurobiol 46:177–205
Langa F, Codony X, Tovar V, Lavado A, Gimenez E, Cozar P,Cantero M, Dordal A, Hernández E, Pérez R, Monroy X,Zamanillo D, Guitart X, Montoliu L (2003) Generation andphenotypic analysis of sigma receptor type I (σ1) knockoutmice. Eur J Neurosci 18:2188–2196
Largent BL, Gundlach AL, Snyder SH (1986) Pharmacological andautoradiographic discrimination of sigma and phencyclidinereceptor binding sites in brain with (+)-[3H]SKF 10,047 (+)-[3H]-3-[3-hydroxyphenyl]-N-(1-propyl)piperidine and [3H]-1-[1-(2-thienyl)cyclohexyl]-piperidine. J Pharmacol Exp Ther238:739–748
Leitner ML, Hohmann AG, Patrick SL, Walker JM (1994) Regionalvariation in the ratio of sigma 1 to sigma 2 binding in rat brain.Eur J Pharmacol 259:65–69
Lupardus PJ, Wilke RA, Aydar E, Palmer CP, Chen Y, Ruoho AE,Jackson MB (2000) Membrane-delimited coupling betweensigma receptors and K+ channels in rat neurohypophysialterminals requires neither G-protein nor ATP. J Physiol526:527–539
Martin WR, Eades CE, Thompson JA, Huppler RE (1976) Theeffects of morphine and nalorphine-like drugs in the non-dependent and morphine-dependent chronic spinal dog. JPharmacol Exp Ther 197:517–532
Maruo J, Yoshida A, Shimohira I, Matsuno K, Mita S, Ueda H(2000) Binding of [35S]GTPgammaS stimulated by (+)-penta-zocine sigma receptor agonist is abundant in the guinea pigspleen. Life Sci 67:599–603
Matsumoto RR, Mack AL (2001) (±)-SM 21 attenuates theconvulsive and locomotor stimulatory effects of cocaine inmice. Eur J Pharmacol 417:R1–R2
Matsumoto RR, Pouw B (2000) Correlation between neurolepticbinding to sigma(1) and sigma(2) receptors and acute dystonicreactions. Eur J Pharmacol 401:155–160
Matsumoto RR, Hemstreet MK, Lai NL, Thurkauf A, de Costa BR,Rice KC, Hellewell SB, Bowen WB, Walker JM (1990) Drugspecificity of pharmacological dystonia. Pharmacol BiochemBehav 36:151–155
Matsumoto RR, Bowen WD, Tom MA, Vo VN, Truong DD, deCosta BR (1995) Characterization of two novel σ receptorligands: antidystonic effects in rats suggests s receptor antag-onism. Eur J Pharmacol 365:35–38
Matsumoto RR, McCraken KA, Friedman MJ, Pouw B, de CostaBR, Bowen WD (2001) Conformationally restricted analogs ofBD1008 and an antisense oligodeoxynucleotide targeting sigma1 receptors produce anti-cocaine effects in mice. Eur JPharmacol 419:163–174
Matsumoto RR, McCracken KA, Pouw B, Zhang Y, Bowen WD(2002) Involvement of sigma receptors in the behavioral effectsof cocaine: evidence from novel ligands and antisenseoligodeoxynucleotides. Neuropharmacology 42:1043–1055
Matsumoto RR, Liu Y, Lerner M, Howard EW, Brackett DJ (2003)Sigma receptors: potential medications development target foranti-cocaine agents. Eur J Pharmacol 469:1–12
08/06/04 - PROOF PAGEMAKEUP 213_1920 15

Matsuno K, Matsunaga K, Senda T, Mita S (1993) Increase inextracellular acetylcholine level by sigma ligands in rat frontalcortex. J Pharmacol Exp Ther 265:851–859
Matsuno K, Senda T, Matsunaga K, Mita S (1994) Amelioratingeffects of sigma receptor ligands on the impairment of passiveavoidance tasks in mice: involvement in the central acetylcho-linergic system. Eur J Pharmacol 261:43–51
Matsuno K, Senda T, Kobayashi T, Mita S (1995) Involvement ofsigma 1 receptor in (+)-N-allylnormetazocine-stimulated hip-pocampal cholinergic functions in rats. Brain Res 690:200–206
Matsuno K, Kobayashi T, Tanaka MK, Mita S (1996a) Sigma 1receptor subtype is involved in the relief of behavioral despairin the mouse forced swimming test. Eur J Pharmacol 286:213–217
Matsuno K, Nakazawa M, Okamoto K, Kawashima Y, Mita S(1996b) Binding properties of SA4503: a novel and selectivesigma 1 receptor agonist. Eur J Pharmacol 306:271–279
Matsuno K, Senda T, Kobayashi T, Okamoto K, Nakata K, Mita S(1997) SA4503: a novel cognitive enhancer with sigma1receptor agonistic properties. Behav Brain Res 83:221–224
Maurice T, Lockhart BP (1997) Neuroprotective and anti-amnesicpotentials of sigma receptor ligands. Prog Neuropsychophar-macol Biol Psychiatry 21:69–102
Maurice T, Su TP, Parish DW, Nabeshima T, Privat A (1994) PRE-084: a sigma selective PCP derivative attenuates MK-801-induced impairment of learning in mice. Pharmacol BiochemBehav 49:859–869
Maurice T, Roman FJ, Privat A (1996) Modulation by neurosteroidsof the in vivo (+)-[3H]SKF 10,047 binding to sigma1 receptorsin the mouse forebrain. J Neurosci Res 46:734–743
Maurice T, Junien JL, Privat A (1997) Dehydroepiandrosteronesulphate attenuates dizocilpine-induced learning impairment inmice via σ1-receptors. Behav Brain Res 83:159–164
Maurice T, Su TP, Privat A (1998) Sigma1 (σ1) receptor agonistsand neurosteroids attenuate β25–35-amyloid peptide-inducedamnesia in mice through a common mechanism. Neuroscience83:413–428
Maurice T, Phan VL, Urani A, Kamei H, Noda Y, Nameshima T(1999) Neuroactive steroids as endogenous effectors for thesigma1 (σ1) receptor: pharmacological evidence and therapeu-tic opportunities. Jpn J Pharmacol 81:125–155
Maurice T, Urani A, Phan V-L, Romieu P (2001) The interactionbetween neuroactive steroids and the σ1 receptor function:behavioral consequences and therapeutic opportunities. BrainRes Rev 37:116–132
Maurice T, Martin-Fardon R, Romieu P, Matsumoto RR (2002)Sigma1(σ1) receptor antagonists represent a new strategyagainst cocaine addiction and toxicity. Neurosci Biobehav Rev26:499–527
McCann DJ, Su TP (1990) Haloperidol-sensitive (+)[3H]SKF10,047 binding sites (sigma sites) exhibit a unique distributionin rat brain subcellular fractions. Eur J Pharmacol 188:211–218
McCann DJ, Su T-P (1991) Solubilization and characterization ofhaloperidol-sensitive (+)-[3H]SKF 10,047 binding sites (sigmasites) from rat liver membranes. J Pharmacol Exp Ther257:547–554
McCann DJ, Weissman AD, Su TP (1994) Sigma-1 and sigma-2sites in rat brain: comparison of regional ontogenic andsubcellular patterns. Synapse 17:182–189
McCraken KA, Bowen WD, Matsumoto RR (1999) Novel σreceptor ligands attenuate the locomotor stimulatory effects ofcocaine. Eur J Pharmacol 365:35–38
McLean S, Weber E (1988) Autoradiographic visualization ofhaloperidol-sensitive sigma receptors in guinea-pig brain.Neuroscience 25:259–269
Mei J, Pasternak GW (2001) Molecular cloning and pharmacolo-gical characterization of the rat sigma1 receptor. BiochemPharmacol 62:349–355
Mei J, Pasternak GW (2002) σ1 Receptor modulation of opioidanalgesia in the mouse. J Pharmacol Exp Ther 300:1070–1074
Mendelsohn LG, Kalra V, Johnson BG, Kerchner GA (1985) Sigmaopioid receptor: characterization and co-identity with thephencyclidine receptor. J Pharmacol Exp Ther 233:597–602
Menkel M, Terry P, Pontecorvo M, Katz JL, Witkin JM (1991)Selective sigma ligands block stimulant effects of cocaine. EurJ Pharmacol 201:251–252
Meyer C, Schmieding K, Falkenstein E, Wehling M (1998) Arehigh-affinity progesterone binding sites(s) from porcine livermicrosomes members of the σ receptor family? Eur JPharmacol 347:293–299
Moebius FF, Striessnig J, Glossmann H (1997a) The mysteries ofsigma receptors: new family members reveal a role incholesterol synthesis. Trends Pharmacol Sci 18:67–70
Moebius FF, Reiter RJ, Hanner M, Glossmann H (1997b) Highaffinity of sigma 1-binding sites for sterol isomerizationinhibitors: evidence for a pharmacological relationship withthe yeast sterol C8–C7 isomerase. Br J Pharmacol 121:1–6
Moltzen EK, Perregaard J, Meier E (1995) Sigma ligands withsubnanomolar affinity and preference for the sigma 2 bindingsite. 2. Spiro-joined benzofuran, isobenzofuran, and benzopy-ran piperidines. J Med Chem 38:2009–2017
Monnet FP, Debonnel G, Junien JL, deMontigny C (1990) N-methyl-D-aspartate-induced neuronal activity is selectivelymodulated by sigma receptors. Eur J Pharmacol 179:441–445
Monnet FP, Blier P, Debonnel G, de Montigny C (1992a)Modulation by sigma ligands of N-methyl-D-aspartate-induced[3H]noradrenaline release. Naunyn Schmiedebergs Arch Phar-macol 346:32–39
Monnet FP, Debonnel G, de Montigny C (1992b) In vivoelectrophysiological evidence for a selective modulation of N-methyl-D-aspartate-induced neuronal activation in rat CA3dorsal hippocampus by sigma ligands. J Pharmacol Exp Ther261:123–130
Monnet FP, Debonnel G, Fournier A, de Montigny C (1992c)Neuropeptide Y potentiates selectively the N-methyl-D-aspar-tate response in the CA3 dorsal hippocampus. II. Involvementof a subtype of sigma receptor. J Pharmacol Exp Ther263:1219–1225
Monnet FP, Fournier A, Debonnel G, de Montigny C (1992d)Neuropeptide Y potentiates selectively the N-methyl-D-aspar-tate response in the CA3 dorsal hippocampus. I. Involvement ofan atypical neuropeptide Y receptor. J Pharmacol Exp Ther263:1212–1218
Monnet FP, Debonnel G, Bergeron R, Gronier B, de Montigny C(1994) The effects of sigma ligands and of neuropeptide Y onN-methyl-D-aspartate-induced neuronal activation of CA3 dor-sal hippocampus neurones are differentially affected bypertussis toxin. Br J Pharmacol 112:709–715
Monroy X, Romero G, Pérez MP, Farré AJ, Guitart X (2001)Decrease of adenylyl cyclase activity and expression by asigma1 receptor ligand and putative atypical antipsychotic.Neuroreport 12:1989–1992
Morin-Surun MP, Collin T, Denavit-Saubié M, Baulieu E-E, MonnetFP (1999) Intracellular σ1 receptor modulates phospholipase Cand protein kinase C activities in the brainstem. Proc Natl AcadSci U S A 96:8196–8199
Morio Y, Tanimoto H, Yakushiji T, Morimoto Y (1994) Character-ization of the currents induced by sigma ligands in NCB20neuroblastoma cells. Brain Res 637:190–196
Nestler EJ (2001) Molecular neurobiology of addiction. Am JAddict 10:201–217
Novakova M (1998) Sigma receptors: with special reference tocardiac muscle. Exp Clin Cardiol 3:113–120
Novakova M, Ela C, Barg J, Vogel Z, Hasin Y, Eilam Y (1995)Inotropic action of sigma receptor ligands in isolated cardiacmyocytes from adult rats. Eur J Pharmacol 286:19–30
Novakova M, Ela C, Bowen WD, Hasin Y, Eilam Y (1998) Highlyselective sigma receptor ligands elevate inositol 1,4,5-trispho-sphate production in rat cardiac myocytes. Eur J Pharmacol353:315–327
16 08/06/04 - PROOF PAGEMAKEUP 213_1920

Nuwayhid SJ, Werling LL (2003) σ1-Receptor agonist-mediatedregulation of N-methyl-D-aspartate-stimulated [3H]dopaminerelease is dependent upon protein kinase C. J Pharmacol ExpTher 304:364–369
Ohmori O, Shinkai T, Suzuki T, Okano C, Kojima H, Terao T,Nakamura J (2000) Polymorphisms of the sigma(1) receptorgene in schizophrenia: an association study. Am J Med Genet96:118–122
Ohno M, Watanabe S (1995) Intrahippocampal administration of(+)-SKF 10,047: a sigma ligand reverses MK-801-inducedimpairment of working memory in rats. Brain Res 684:237–242
Okumura K, Ujike H, Akiyama K, Kuroda S (1996) BMY-14802reversed the sigma receptor agonist-induced neck dystonia inrats. J Neural Transm 103:1153–1161
Okuyama S, Imagawa Y, Ogawa S, Araki H, Ajima A, Tanaka M,Muramatsu M, Nakazato A, Yamaguchi K, Yoshida M, OtomoS (1993) NE-100, a novel sigma receptor ligand: in vivo tests.Life Sci 53:285–290
Okuyama S, Imagawa Y, Sakagawa T, Nakazato A, Yamaguchi K,Katoh M, Yamada S, Araki H, Otomo S (1994) NE-100, anovel sigma receptor ligand: effect on phencyclidine-inducedbehaviors in rats dogs and monkeys. Life Sci 55:PL133–PL138
Okuyama S, Chaki S, Yae T, Nakazato A, Muramatsu M (1995a)Autoradiographic characterization of binding sites for [3H]NE-100 in guinea pig brain. Life Sci 57:PL333–PL337
Okuyama S, Ogawa S, Nakazato A, Tomizawa K (1995b) Effect ofNE-100, a novel sigma receptor ligand on phencyclidine-induced delayed cognitive dysfunction in rats. Neurosci Lett189:60–62
Oshiro Y, Sakurai Y, Sato S, Kurahashi N, Tanaka T, Kikuchi T,Tottori K, Uwahodo Y, Miwa T, Nishi T (2000) 3,4-Dihydro-2(1H))-quinolinone as a novel antidepressant drug: synthesis andpharmacology of 1-[3-[4-(3-chlorophenyl)-1-piperazinyl]pro-pyl]-3,4-dihydro-5-methoxy-2(1H)-quinolinone and its deriva-tives. J Med Chem 43:177–189
Pan YX, Mei JF, Xu J, Wan BL, Zuckerman A, Pasternak GW(1998) Cloning and characterization of a σ1 receptor. JNeurochem 70:2279–2285
Pascaud XB, Chovet M, Roze C, Junien JL (1993) Neuropeptide Yand sigma receptor agonists act through a common pathway tostimulate duodenal alkaline secretion in rats. Eur J Pharmacol231:389–394
Paul IA, Basile AS, Rojas E, Youdim MB, De Costa B, Skolnick P,Pollard HB, Kuijpers GA (1993) Sigma receptors modulatenicotinic receptor function in adrenal chromaffin cells. FASEBJ 7:1171–1178
Phan VL, Su TP, Privat A, Maurice T (1999) Modulation ofsteroidal levels by adrenalectomy/castration and inhibition ofneurosteroid synthesis enzymes affect sigma1 receptor-mediated behaviour in mice. Eur J Neurosci 11:2385–2396
Poncelet M, Santucci V, Paul R, Gueudet C, Lavastre S, Guitard J,Steinberg R, Terranova JP, Breliere JC, Soubrie P, Le Fur G(1993) Neuropharmacological profile of a novel and selectiveligand of the sigma site: SR 31742A. Neuropharmacology32:605–615
Prasad PD, Li HW, Fei YJ, Ganapathy ME, Fujita T, Plumley LH,Yang-Feng TL, Leibach FH, Ganapathy V (1998) Exon–intronstructure analysis of promoter region and chromosomal local-ization of the human type 1 sigma receptor gene. J Neurochem70:443–451
Quirion R, Bowen WD, Itzhak Y, Junien JL, Musacchio JM,Rothman RB, Su T-P, Tam SW, Taylor DP (1992) A proposalfor the classification of sigma binding sites. Trends PharmacolSci 13:85–86
Ritz MC, George FR (1993) Cocaine-induced seizures and lethalityappear to be associated with distinct central nervous systembinding sites. J Pharmacol Exp Ther 264:1333–1343
Roman F, Pascaud X, Duffy O, Vauche D, Martin B, Junien JL(1989a) Neuropeptide Y and peptide YY interact with rat brainsigma and PCP binding sites. Eur J Pharmacol 174:301–302
Roman F, Pascaud X, Chomette G, Bueno L, Junien JL (1989b)Autoradiographic localization of sigma opioid receptors in thegastrointestinal tract of the guinea pig. Gastroenterology 97:76–82
Roman FJ, Pascaud X, Martin B, Vauche D, Junien JL (1990) JO1784: a potent and selective ligand for rat and mouse brain σ-sites. Eur J Pharmacol 42:439–440
Romero G, Pérez MP, Carceller A, Monroy X, Farré AJ, Guitart X(2000) Changes in phosphoinositide signalling activity andlevels of the alpha subunit of Gq/11 protein in rat brain inducedby E-5842, a sigma1 receptor ligand and potential atypicalantipsychotic. Neurosci Lett 290:189–192
Romieu P, MartinFardon R, Maurice T (2000) Involvement of thesigma 1 receptor in the cocaine-induced conditioned placepreference. Neuroreport 11:2885–2888
Ross SB (1991) Heterogeneous binding of σ radioligands in the ratbrain and liver: possible relationship to subforms of cytochromeP450. Pharmacol Toxicol 68:293–301
Rothman RB, Reid A, Mahboubi A, Kim CH, De Costa BR,Jacobson AE, Rice KC (1991) Labeling by [3H]13-di(2-tolyl)guanidine of two high affinity binding sites in guinea pig brain:evidence for allosteric regulation by calcium channel antago-nists and pseudoallosteric modulation by sigma ligands. MolPharmacol 39:222–232
Ryan-Moro J, Chien CC, Standifer KM, Pasternak GW (1996)Sigma binding in a human neuroblastoma cell line. NeurochemRes 21:1309–1314
Samovilova NN, Vinogradov VA (1992) Subcellular distribution of(+)-[3H]SKF 10047 binding sites in rat liver. Eur J PharmacolMol Pharmacol 225:69–74
Samovilova NN, Nagornaya LV, Vinogradov VA (1988) (+)-[3H]SKF 10,047 binding sites in rat liver. Eur J Pharmacol147:259–264
Sanchez C, Papp M (2000) The selective sigma(2) ligand Lu 28-179has an antidepressant-like profile in the rat chronic mild stressmodel of depression. Behav Pharmacol 11:117–124
Sanchez C, Arnt J, Costall B, Kelly ME, Meier E, Naylor RJ,Perregaard J (1997) The selective sigma2-ligand Lu 28-179 haspotent anxiolytic-like effects in rodents. J Pharmacol Exp Ther283:1323–1332
Schmidt A, Lebel L, Koe BK, Seeger T, Heym J (1989) Sertralinepotently displaces (+)-[3H]3-PPP binding to sigma sites in ratbrain. Eur J Pharmacol 165:335–336
Senda T, Matsuno K, Kobayashi T, Nakazawa M, Nakata K, Mita S(1998a) Ameliorative effect of SA4503, a novel cognitiveenhancer on the basal forebrain lesion-induced impairment ofthe spatial learning performance in rats. Pharmacol BiochemBehav 59:129–134
Senda T, Mita S, Kaneda K, Kikuchi M, Akaike A (1998b) Effect ofSA4503, a novel sigma1 receptor agonist against glutamateneurotoxicity in cultured rat retinal neurons. Eur J Pharmacol42:105–111
Seth P, Fei YJ, Huang W, Leibach FH, Ganapathy V (1998) Cloningand functional characterization of a sigma receptor from ratbrain. J Neurochem 70:922–931
Sharkey J, Glen KA, Wolfe S, Kuhar MJ (1988) Cocaine binding atsigma receptors. Eur J Pharmacol 149:171–174
Soriani O, Foll FL, Roman F, Monnet FP, Vaudry H, Cazin L (1999)A-current down-modulated by sigma receptor in frog pituitarymelanotrope cells through a G protein-dependent pathway. JPharmacol Exp Ther 289:321–328
Su TP (1993) Delineating biochemical and functional properties ofsigma receptors: emerging concepts. Crit Rev Neurobiol 7:187–203
Su T-P, Junien JL (1994) Sigma receptors in the central nervoussystem and the periphery. In: Itzhak Y (ed) Sigma receptors.Academic, San Diego, pp 21–44
Su TP, Hayashi T (2001) Cocaine affects the dynamics ofcytoskeletal proteins via sigma(1) receptors. Trends PharmacolSci 22:456–458
08/06/04 - PROOF PAGEMAKEUP 213_1920 17

Su TP, Hayashi T (2003) Understanding the molecular mechanismof sigma-1 receptors: towards a hypothesis that sigma-1receptors are intracellular amplifiers for signal transduction.Curr Med Chem 10:2073–2080
Su T-P, London ED, Jaffe JH (1988) Steroid binding at sigmareceptor suggests a link between endocrine nervous andimmune systems. Science 240:219–221
Su TP, Wu XZ, Cone EJ, Shukla K, Gund TM, Dodge AL, ParishDW (1991) Sigma compounds derived from phencyclidine:identification of PRE-084, a new selective sigma ligand. JPharmacol Exp Ther 259:543–550
Takahashi S, Sonehara K, Takagi K, Miwa T, Horikomi K, Mita N,Nagase H, Iizuka K, Sakai K (1999) Pharmacological profile ofMS-377, a novel selective antipsychotic agent with selectiveaffinity for sigma receptors. Psychopharmacology 145:295–302
Takahashi S, Miwa T, Horikomi K (2000) Involvement of sigma 1receptors in methamphetamine-induced behavioural sensitiza-tion in rats. Neurosci Lett 289:21–24
Takebayashi M, Hayashi T, Su TP (2002) Nerve growth factor-induced neurite sprouting in PC12 cells involves sigma-1receptors: implications for antidepressants. J Pharmacol ExpTher 303:1227–1237
Tam SW, Mitchell KN (1991) Neuropeptide Y and peptide YY donot bind to brain sigma and phencyclidine binding sites. Eur JPharmacol 193:121–122
Tam SW, Steinfels GF, Gilligan PJ, Schimdt WK, Cook L (1992)DuP 724 [1-(cyclopropylmethyl)-4-(2′(4″-fluorophenyl)-2′-ox-oethyl)-piperidine HBr]: a sigma- and 5-hydroxytryptamine2receptor antagonist; receptor-binding electrophysiological andneuropharmacological profiles. J Pharmacol Exp Ther263:1167–1174
Tascedda F, Lovati E, Blom JMC, Muzzioli P, Brunello N, RacagniG, Riva MA (1999) Regulation of ionotropic glutamatereceptors in the rat brain in response to the atypical antipsy-chotic seroquel (quetiapine fumarate). Neuropsychopharmacol-ogy 21:211–217
Taylor DP, Dekleva J (1987) The potential antipsychotic BMY14802 selectively binds to sigma sites. Fed Proc 46:1304
Taylor DP, Eison MS, Moon SL, Schlemmer RF Jr, Shukla UA,VanderMaelen CP, Yocca FD, Gallant DJ, Behling SH,Boissard CG, Braselton JP, Davis HH Jr, Duquette MN,Lamy RC, Libera JM, Ryan E, Wright RN (1993) A role for σbinding in the antipsychotic profile of BMY 14802? In: SouzaEB, Clouet D, London E (eds) Sigma PCP and NMDAreceptors. NIDA Research Monograph Series, pp 125–157
Tokuyama S, Hirata K, Ide A, Ueda H (1997) Sigma ligandsstimulate GTPase activity in mouse prefrontal membranes—evidence for the existence of metabotropic sigma receptor.Neurosci Lett 233:141–144
Tokuyama S, Hirata K, Yoshida A, Maruo J, Matsuno K, Mita S,Ueda H (1999) Selective coupling of mouse brain metabotropicsigma (σ) receptor with recombinant Gi1. Neurosci Lett268:85–88
Torrence-Campbell C, Bowen WD (1996) Differential solubilizationof rat liver sigma 1 and sigma 2 receptors: retention of sigma 2sites in particulate fraction. Eur J Pharmacol 304:201–210
Tottori K, Miwa T, Uwahodo Y, Yamada S, Nakai M, Oshiro Y,Kikuchi T, Altar CA (2001) Antidepressant-like responses tothe combined sigma and 5-HT1A receptor agonist OPC-14523.Neuropharmacology 41:976–988
Tran TT, de Costa BR, Matsumoto RR (1998) Microinjection ofsigma ligands into cranial nerve nuclei produces vacuouschewing in rats. Psychopharmacology 137:191–200
Uchida N, Ujike H, Nakata K, Takaki M, Nomura A, Katsu T,Tanaka Y, Imamura T, Sakai A, Kuroda S (2003) Noassociation between the sigma receptor type 1 gene andschizophrenia: results of analysis and meta-analysis of case-control studies. BMC Psychiatry 3:13
Ujike H, Kanzaki A, Okumura K, Akiyama K, Otsuki S (1992)Sigma antagonist BMY 14802 prevents methamphetamine-induced sensitization. Life Sci 50:PL129–PL134
Ujike H, Kuroda S, Otsuki S (1996) Sigma receptor antagonistsblock the development of sensitization to cocaine. Eur JPharmacol 296:123–128
Ukai M, Maeda H, Nanya Y, Kameyama T, Matsuno K (1998)Beneficial effects of acute and repeated administrations ofsigma receptor agonists on behavioral despair in mice exposedto tail suspension. Pharmacol Biochem Behav 61:247–252
Urani A, Roman FJ, Phan V-L, Su T-P, Maurice T (2001) Theantidepressant-like effect induced by σ1-receptor agonists andneuroactive steroids in mice submitted to the forced swimmingtest. J Pharmacol Exp Ther 298:1269–1279
Urani A, Romieu P, Portales-Casamar E, Roman FJ, Maurice T(2002) The antidepressant-like effect induced by the sigma1(σ1) receptor agonist igmesine involves modulation of intra-cellular calcium mobilization. Psychopharmacology 163:26–35
Van Broekhoven F, Verkes RJ (2003) Neurosteroids in depression: areview. Psychopharmacology 165:97–110
Vaupel DB (1983) Naltrexone fails to antagonize the sigma effectsof PCP and SKF 10,047 in the dog. Eur J Pharmacol 92:269–274
Vilner BJ, Bowen WD (1993) Sigma receptor-active neurolepticsare cytotoxic to C6 glioma cells in culture. Eur J PharmacolMol Pharmacol Sect 244:199–201
Vilner BJ, Bowen WD (2000) Modulation of cellular calcium bysigma-2 receptors. Release from intracellular stores in humanSK-N-SH neuroblastoma cells. J Pharmacol Exp Ther 292:900–911
Vilner BJ, de Costa BR, Bowen WD (1995) Cytotoxic effects ofsigma ligands: sigma receptor-mediated alterations in cellularmorphology and viability. J Neurosci 15:117–134
Walker JM, Bowen WD, Walker FO, Matsumoto RR, De Costa B,Rice KC (1990) Sigma receptors: biology and function.Pharmacol Rev 42:355–402
Walker JM, Bowen WD, Patrick SL, Williams WE, Mascarella SW,Bai X, Carroll FI (1993) A comparison of (−)-deoxybenzo-morphans devoid of opiate activity with their dextrorotatoryphenolic counterparts suggests role of σ2 receptors in motorfunction. Eur J Pharmacol 231:61–68
Walker JM, Martin WJ, Hohmann AG, Hemstreet MK, Roth JS,Leitner ML, Weiser SD, Patrick SL, Patrick RL, Matsumoto RR(1994) Role of sigma receptors in brain mechanisms ofmovement. In: Itzhak Y (ed) Sigma receptors. Academic, SanDiego, pp 205–224
Weatherspoon JK, Werling LL (1999) Modulation of amphetamine-stimulated [3H]dopamine release from rat pheochromocytoma(PC12) cells by σ type 2 receptors. J Pharmacol Exp Ther289:278–284
Weber E, Sonders M, Quarum M, McLean S, Pou S, Keana JFW(1986) 1,3-Di(2-[5-3H]tolyl)guanidine: a selective ligand thatlabels sigma-type receptors for psychotomimetic opiates andantipsychotic drugs. Proc Natl Acad Sci U S A 83:8784–8788
Weissman AD, Casanova MF, Kleinman JE, London ED, De SouzaEB (1991) Selective loss of cerebral cortical sigma but notPCP-binding sites in schizophrenia. Biol Psychiatry 29:41–54
Weissman AD, Su TP, Hedreen JC, London DE (1998) Sigmareceptors in post-mortem human brains. J Pharmacol Exp Ther247:29–33
Wickman KD, Clapham DE (1995) G-protein regulation of ionchannels. Curr Opin Neurobiol 5:278–285
Wilke RA, Mehta RP, Lupardus PJ, Chen Y, Ruoho AE, JacksonMB (1999a) Sigma receptor photolabeling and sigma receptor-mediated modulation of potassium channels in tumor cells. JBiol Chem 274:18387–18392
Wilke RA, Lupardus PJ, Grandy DK, Rubinstein M, Low MJ,Jackson MB (1999b) K+ channel modulation in rodentneurohypophysial nerve terminals by sigma receptors and notby dopamine receptors. J Physiol 517:391–406
Yamada M, Nishigami T, Nakasho K, Nishimoto Y, Miyaji H (1994)Relationship between σ-like site and progesterone-binding siteof adult male rat liver microsomes. Hepatology 20:1271–1280
18 08/06/04 - PROOF PAGEMAKEUP 213_1920

Yamamoto H, Miura R, Yamamoto T, Shinohara K, Watanabe M,Okuyama S, Nakazato A, Nukada T (1999) Amino acidresidues in the transmembrane domain of the type 1 sigmareceptor critical for ligand binding. FEBS Lett 445:19–22
Yoshida K, Takahashi H, Sato K, Higuchi H, Shimizu T (2000)Biperiden hydrochlorate ameliorates dystonia of rats producedby microinjection of sigma ligands into the red nucleus.Pharmacol Biochem Behav 67:497–500
Ytzhak Y (1989) Multiple affinity binding states of the sigmareceptor: effect of GTP-binding protein-modifying agents. MolPharmacol 36:512–517
Zamanillo D, Andreu F, Ovalle S, Pérez MP, Romero G, Farré AJ,Guitart X (2000) Up-regulation of sigma1 receptor mRNA inrat brain by a putative atypical antipsychotic and sigma receptorligand. Neurosci Lett 282:169–172
Zamanillo D, Romero G, Dordal A, Pérez P, Vincent L, Méndez R,Andreu F, Hernández E, Pérez R, Monroy X, Ovalle S, GuitartX (2002) Increase of forskolin-stimulated adenylyl cyclase andAP-1 activities by sigma1 receptor expression. FENS AbstrA04617
Zhang H, Cuevas J (2002) Sigma receptors inhibit high-voltage-activated calcium channels in rat sympathetic and parasympa-thetic neurons. J Neurophysiol 87:2867–2879
Zou LB, Yamada K, Sasa M, Nakata Y, Nabeshima T (2000) Effectsof sigma1 receptor agonist SA4503 and neuroactive steroids onperformance in a radial arm maze task in rats. Neuropharma-cology 39:1617–1627
Zukin SR, Brady KT, Slifer BL, Balster RL (1984) Behavioral andbiochemical stereoselectivity of sigma opiate/PCP receptors.Brain Res 294:174–177
08/06/04 - PROOF PAGEMAKEUP 213_1920 19
Related Documents











