RESEARCH REPORT Short-Term Limb Immobilization Affects Cognitive Motor Processes Lucette Toussaint and Aurore Meugnot Universite ´ de Poitiers We examined the effects of a brief period of limb immobilization on the cognitive level of action control. A splint placed on the participants’ left hand was used as a means of immobilization. We used a hand mental rotation task to investigate the immobilization-induced effects on motor imagery performance (Experiments 1 and 2) and a number mental rotation task to investigate whether immobilization-induced effects are also found when visual imagery is involved (Experiment 2). We also examined whether the effects of immobilization vary as a function of individuals’ vividness of motor imagery (Experiment 2). The immobilized participants performed the mental rotation tasks before and immediately after the splint removal. The control group did not undergo the immobilization procedure. For hand stimuli, response time analysis showed a lack of task-repetition benefit following immobilization (Experiments 1 and 2) except when the visual imagery task was performed first (Experiment 2). Following immobilization, a flattening in the response time profile for left hand stimuli was observed as a function of stimuli rotation (Experiments 1 and 2), especially for participants with less vivid motor imagery (Experiment 2). We did not find an immobilization-induced effect on number stimuli. These findings revealed that the cognitive representation of hand movements is modified by immobilization and that sensorimotor deprivation specifically affects motor simulation of the immobilized hand. We discuss the possibility that immobi- lization affects the sensorimotor system due to the reduced processing of proprioceptive feedback, which lead some participants to switch from a motor to a visual imagery strategy. Keywords: motor imagery, visual imagery, upper limb immobilization, effector dependence, sensorimo- tor representation Experience plays a fundamental role in human brain develop- ment and subsequent behavior. Neurophysiological and functional imaging studies have shown that motor training results in an enlarged cortical sensorimotor representation (Hlustik, Solodkin, Noll, & Small, 2004; Pascual-Leone & Torres, 1993), whereas a lack of physical activity leads to a reduced representation (Liepert, Tegenthoff, & Malin, 1995; Zanette et al., 1997). The changes in sensorimotor cortex organization lead to well-known motor per- formance modifications with regard to motor skill acquisition, but little is known about the behavioral consequences of an imposed low level of physical activity. The present study examines the effects of 2 days of upper limb sensorimotor deprivation by means of a splint on cognitive motor processes. We used motor imagery (i.e., internal movement simulation) to investigate the cognitive aspect of action control following immobilization because it avoids confounding related to motor execution. Motor imagery is considered to be the cognitive level of action processing (Jean- nerod, 2001). For this purpose, the hand laterality task (Parsons, 1987, 1994), in which subjects are requested to identify the later- ality of hand images presented in various orientations, is particu- larly appropriate to study factors affecting cognitive aspects of action. This task implicitly triggers motor imagery processes, as evidenced in behavioral and neuroimaging studies (de Lange, Helmich, & Toni, 2006; Kosslyn, DiGirolamo, Thompson, & Alpert, 1998; Parsons, 1994; Parsons, Gabrieli, Phelps, & Gazza- niga, 1998; Sekiyama, 1982). The embodied nature of high-level cognitive processes has been shown in research areas such as language understanding (Glen- berg, Sato, & Cattaneo, 2008), processing of action-related words (Tomasino, Weiss, & Fink, 2010), and number magnitude process- ing (Badets & Pesenti, 2010). With regard to motor imagery, the embodied nature of this process remains controversial. Some stud- ies have reported that motor imagery performance is influenced by peripheral changes, such as the current posture of the subject’s own hands (Ionta, Fourkas, Fiorio, & Aglioti, 2007; Sirigu & Duhamel, 2001), the amputation of an upper limb (Nico, Daprati, Rigal, Parsons, & Sirigu, 2004), and focal hand dystonia, or writer’s cramp (Fiorio, Tinazzi, & Aglioti, 2006). In these studies, left- and right-handedness judgments were strongly affected by the changes in the actual body states or by the absence of the real effector. Similar results have been observed in patients suffering from chronic arm pain (Schwoebel, Friedman, Duda, & Coslett, 2001); these patients exhibited slower response times, especially when judging hand laterality of hand images that required large- This article was published Online First June 11, 2012. Lucette Toussaint and Aurore Meugnot, Centre de Recherches sur la Cognition et l’Apprentissage, Universite ´ de Poitiers, Poitiers, France. We thank Yves Alme ´cija (Centre National de la Recherche Scientifique/ Unite ´ Mixte de Recherche 7295) for technical support. Correspondence concerning this article should be addressed to Lucette Toussaint, Centre de Recherches sur la Cognition et l’Apprentissage, CNRS/UMR 7295, Universite ´ de Poitiers, 5 Rue The ´odore Lefebvre, 86000 Poitiers, France. E-mail: [email protected] Journal of Experimental Psychology: © 2012 American Psychological Association Learning, Memory, and Cognition 2013, Vol. 39, No. 2, 623– 632 0278-7393/13/$12.00 DOI: 10.1037/a0028942 623

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH REPORT
Short-Term Limb Immobilization Affects Cognitive Motor Processes
Lucette Toussaint and Aurore MeugnotUniversite de Poitiers
We examined the effects of a brief period of limb immobilization on the cognitive level of action control.A splint placed on the participants’ left hand was used as a means of immobilization. We used a handmental rotation task to investigate the immobilization-induced effects on motor imagery performance(Experiments 1 and 2) and a number mental rotation task to investigate whether immobilization-inducedeffects are also found when visual imagery is involved (Experiment 2). We also examined whether theeffects of immobilization vary as a function of individuals’ vividness of motor imagery (Experiment 2).The immobilized participants performed the mental rotation tasks before and immediately after the splintremoval. The control group did not undergo the immobilization procedure. For hand stimuli, responsetime analysis showed a lack of task-repetition benefit following immobilization (Experiments 1 and 2)except when the visual imagery task was performed first (Experiment 2). Following immobilization, aflattening in the response time profile for left hand stimuli was observed as a function of stimuli rotation(Experiments 1 and 2), especially for participants with less vivid motor imagery (Experiment 2). We didnot find an immobilization-induced effect on number stimuli. These findings revealed that the cognitiverepresentation of hand movements is modified by immobilization and that sensorimotor deprivationspecifically affects motor simulation of the immobilized hand. We discuss the possibility that immobi-lization affects the sensorimotor system due to the reduced processing of proprioceptive feedback, whichlead some participants to switch from a motor to a visual imagery strategy.
Keywords: motor imagery, visual imagery, upper limb immobilization, effector dependence, sensorimo-tor representation
Experience plays a fundamental role in human brain develop-ment and subsequent behavior. Neurophysiological and functionalimaging studies have shown that motor training results in anenlarged cortical sensorimotor representation (Hlustik, Solodkin,Noll, & Small, 2004; Pascual-Leone & Torres, 1993), whereas alack of physical activity leads to a reduced representation (Liepert,Tegenthoff, & Malin, 1995; Zanette et al., 1997). The changes insensorimotor cortex organization lead to well-known motor per-formance modifications with regard to motor skill acquisition, butlittle is known about the behavioral consequences of an imposedlow level of physical activity. The present study examines theeffects of 2 days of upper limb sensorimotor deprivation by meansof a splint on cognitive motor processes. We used motor imagery(i.e., internal movement simulation) to investigate the cognitiveaspect of action control following immobilization because itavoids confounding related to motor execution. Motor imagery isconsidered to be the cognitive level of action processing (Jean-
nerod, 2001). For this purpose, the hand laterality task (Parsons,1987, 1994), in which subjects are requested to identify the later-ality of hand images presented in various orientations, is particu-larly appropriate to study factors affecting cognitive aspects ofaction. This task implicitly triggers motor imagery processes, asevidenced in behavioral and neuroimaging studies (de Lange,Helmich, & Toni, 2006; Kosslyn, DiGirolamo, Thompson, &Alpert, 1998; Parsons, 1994; Parsons, Gabrieli, Phelps, & Gazza-niga, 1998; Sekiyama, 1982).
The embodied nature of high-level cognitive processes has beenshown in research areas such as language understanding (Glen-berg, Sato, & Cattaneo, 2008), processing of action-related words(Tomasino, Weiss, & Fink, 2010), and number magnitude process-ing (Badets & Pesenti, 2010). With regard to motor imagery, theembodied nature of this process remains controversial. Some stud-ies have reported that motor imagery performance is influenced byperipheral changes, such as the current posture of the subject’sown hands (Ionta, Fourkas, Fiorio, & Aglioti, 2007; Sirigu &Duhamel, 2001), the amputation of an upper limb (Nico, Daprati,Rigal, Parsons, & Sirigu, 2004), and focal hand dystonia, orwriter’s cramp (Fiorio, Tinazzi, & Aglioti, 2006). In these studies,left- and right-handedness judgments were strongly affected by thechanges in the actual body states or by the absence of the realeffector. Similar results have been observed in patients sufferingfrom chronic arm pain (Schwoebel, Friedman, Duda, & Coslett,2001); these patients exhibited slower response times, especiallywhen judging hand laterality of hand images that required large-
This article was published Online First June 11, 2012.Lucette Toussaint and Aurore Meugnot, Centre de Recherches sur la
Cognition et l’Apprentissage, Universite de Poitiers, Poitiers, France.We thank Yves Almecija (Centre National de la Recherche Scientifique/
Unite Mixte de Recherche 7295) for technical support.Correspondence concerning this article should be addressed to Lucette
Toussaint, Centre de Recherches sur la Cognition et l’Apprentissage,CNRS/UMR 7295, Universite de Poitiers, 5 Rue Theodore Lefebvre,86000 Poitiers, France. E-mail: [email protected]
Journal of Experimental Psychology: © 2012 American Psychological AssociationLearning, Memory, and Cognition2013, Vol. 39, No. 2, 623–632
0278-7393/13/$12.00 DOI: 10.1037/a0028942
623

amplitude-imagined movements. The embodied nature of move-ment simulation has been recently confirmed and extended by deLange et al.’s (2006) work with healthy participants. These authorsshowed that the current posture of the left and right hand affectsmotor imagery performance in an effector-specific manner (seealso Ionta & Blanke, 2009; Shenton, Schwoebel, & Coslett, 2004),suggesting a close link between the mentally represented limb andits physical counterpart.
By contrast, some studies revealed a dissociation between theability to simulate an action and the ability to execute it. Johnsonand collaborators (Johnson, 2000; Johnson, Sprehn, & Saykin,2002) showed that movement simulation can be successfully car-ried out in both acute and chronic hemiplegic patients, regardlessof whether the limb is paralyzed. These results suggest that theinternal limb representation has not been recalibrated to reflectperipheral modifications and that sensorimotor representation isnot activity dependent. Evidence for the robustness of sensorimo-tor representation comes from another study with lower limbamputees (Curtze, Otten, & Postema, 2010). Despite the anatom-ical changes in the body periphery, no difference was observed inthe mental rotation of feet for judgments based on the affected(absent) versus unaffected limb. Overall, these results are contraryto those reported on the continuous updating of body representa-tion with regard to peripheral factors (de Lange et al., 2006; Fiorioet al., 2006; Ionta et al., 2007; Nico et al., 2001; Schwoebel et al.,2001). These inconsistencies in pathological populations led us toexamine the robustness of sensorimotor representation in healthyparticipants. Of primary interest in the present study is whethershort-term, upper limb immobilization affects the ability to repre-sent hand movement.
Recent studies have shown that a splint used to immobilizeupper or lower limbs for many weeks results in changes in both theperipheral musculature (Seki, Taniguchi, & Narusawa, 2001) andthe central nervous system (Huber et al., 2006; Kaneko, Mu-rakami, Onari, Kurumadani, & Kawaguchi, 2003; Liepert et al.,1995; Zanette et al., 1997). Cortical plastic changes in normalpeople may also appear following a brief period of limb immobi-lization. Facchini, Romani, Tinazzi, and Aglioti (2002) reported adecrease of cortical excitability after 3–4 days of two-hand fingerimmobilization. In the same vein, Huber et al. (2006) showed thatarm immobilization for only 12 hr induced a decrease of bothsomatosensory and motor-evoked potential over the contralateralsensorimotor cortex. For these authors, the central reorganizationof the sensorimotor areas following immobilization could be theorigin of motor performance deteriorations, as revealed by theincreased variability in the hand path areas of reaching move-ments. The immobilization-induced motor alterations were alsoreported by Moisello et al. (2008). They observed changes inmovement control that were similar to those reported for deaffer-ented patients (Sainburg, Ghilardi, Poizner, & Ghez, 1995). Asproposed by Moisello et al., the alteration of hand and jointtrajectories following short-term limb immobilization could resultin changes to sensorimotor memories as a result of the reducedprocessing of proprioceptive signals. The effects of sensorimotordeprivation on subsequent motor performance are thus more likelyto be related to plastic changes in the sensorimotor areas becausethese changes occur too early to be ascribed to changes in musclestructure (Facchini et al., 2002). However, one cannot exclude thepossibility that some difficulties in executing real hand movements
occur after immobilization due to peripheral changes, such asankylosis of the hand joints. The extent to which the less effectivemovement outcome after immobilization is specifically due to aninadequate sensorimotor representation remains to be clarified.
In the present study, we investigated whether participants withshort-term hand immobilization have difficulty mentally simulat-ing hand movements and examined whether this impairment isspecific to the immobilized hand or whether it is also observedwhen the nonimmobilized hand is mentally rotated. For this pur-pose, we used a left- and right-hand judgment task (Experiments 1and 2), which implicitly triggers motor imagery processes (Par-sons, 1994). To determine whether hand pictures presented atdifferent orientations correspond to left or right hand rotation,normal participants mentally rotated their own hands from theircurrent position to the orientation of the hand picture (i.e., asubject-centered strategy). In this task, it has been shown thatmental rotation processes are subject to the same biomechanicalconstraints as actual movements. Some authors reported that re-sponse time increases with the length of hand trajectories and formovements that reach the limits of what is biomechanically pos-sible (i.e., for unnatural hand orientation; Nı Choisdealbha, Brady,& Maguinness, 2011; Nico et al., 2004; Parsons, 1994; Sekiyama,1982). As a result, we assumed that the central origin of short-termupper limb immobilization should lead to impaired motor imageryperformance, especially for the most constraining postures. Fur-thermore, if the absence of physical exercise causes an alteration inthe representation of specific body parts, then an atypical pattern ofmotor imagery performance should be observed for the immobi-lized left hand (compared with the nonimmobilized right hand)after 2 days of sensorimotor deprivation.
In Experiment 2, we also examined whether upper limb short-term immobilization only affects motor imagery performance orwhether performance affects are also found when visual imagery isinvolved. To this aim, we used a supplementary nonbody mentalrotation task (i.e., with number stimuli), which did not elicit amotor strategy (Dalecki, Hoffmann, & Bock, 2012; Ganis, Keenan,Kosslyn, & Pascual-Leone, 2000; Jola & Mast, 2005; Kosslyn etal., 1998; Tomasino, Rumiati, & Umilta, 2003) when participantswere not instructed to relate the objects to their hands (Kosslyn etal., 1998; Kosslyn, Thompson, Wraga, & Alpert, 2001; Tomasino& Rumiati, 2004). Finally, at least two mechanisms can be usedwhen solving mental rotation task: Visual imagery processes (i.e.,object-centered strategy) engage many visuospatial cortical areas,whereas motor imagery processes (i.e., subject-centered strategy)engage the sensory and motor cortex areas. We expectedimmobilization-induced effects only for the motor imagery task.
Experiment 1
Method
Participants. Twenty-two right-handed university students(18–30 years of age) took part in the experiment. They weredivided into two groups (five men and six women in each group):a control group (mean age � 23.2 years) and an immobilizedgroup (mean age � 21.5 years). The participants were healthy, hadnormal or corrected-to-normal vision, and had no history of motoror neurological disorders. The study protocol was approved by thelocal ethics committee. All participants gave their written informed
624 TOUSSAINT AND MEUGNOT

consent prior to their inclusion in this study and received €20 fortheir participation. Before testing, all participants were naive to theaims of the experiment.
Material and task. Participants were seated in front of acomputer screen. Their left hand was placed near the keyboard,and their right hand was placed on the keyboard, with index andmiddle fingers on two marked keys. The index and middle fingerswere on the left and right marked keys, respectively. Participantshad to judge the laterality hand stimuli presented individually onthe computer screen by pressing the left (for left hand) or right (forright hand) keys as accurately and as quickly as possible. TheE-Prime 2.0 software package was used to present hand stimuliand to record the participants’ responses (accuracy and responsetimes). The stimuli were interspersed with a black fixation crossdisplayed randomly for 500, 1,000, or 1,500 ms. Each stimulusremained visible until the participant’s response was given.
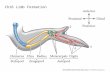
Stimuli consisted of hand figures created with Poser 6.0 soft-ware. Left and right hands (13 cm in height, 7 cm in width) weremirror images of each other (see Figure 1). They were presented indifferent orientations in the picture plane (40°, 80°, 120°, and 160°in a clockwise or a counterclockwise direction). Note that a clock-wise direction corresponds to a medial rotation for the left handand a lateral rotation for the right hand, whereas the reverse is truefor counterclockwise directions. Figure 1 illustrates the 16 stimuli(2 hands � 4 rotations � 2 directions) used in the present exper-iment.
Procedure. Before the start of the experiment, participantswere divided into two groups (control vs. immobilized). We im-mobilized the left hand of one group for 48 hr; the other groupserved as controls. We used a rigid splint (a padded wrist supportwith adjustable aluminum palmar stay, model DonJoy “ComfortDigit,” DJO, Guildford, United Kingdom) to immobilize the wristand three fingers (the index, middle, and ring fingers). Eachparticipant’s left arm was placed in a sling to ensure that theparticipant kept the left hand at rest during the 2 days of immo-bilization.
The mental rotation task was performed during two experimen-tal sessions 48 hr apart for both groups (pre- and posttests). Thetask was performed before immobilization (pretest) and immedi-ately after splint removal (posttest) for the immobilized group.
For all participants, the mental rotation task was divided intotwo phases. The first training phase was designed to familiarizeparticipants with the task. They were shown 16 trials (2 hands �4 rotations � 2 directions) in a random order. Participants had tojudge the laterality of the hands by pressing on the appropriatemarked key without time constraints. All participants correctlyunderstood the task instructions.
The second experimental phase was similar to the trainingphase, except that participants were asked to respond as accuratelyand as quickly as possible. The experimental phase was composedof four blocks of 16 randomly presented trials (2 hands � 4rotations � 2 directions). Sixty-four trials were performed by eachparticipant. No specific imagery instructions were given from thebeginning to the end of the experiment.
Data analysis. Only data from correct responses were used tosubsequently analyze response times. Moreover, response timeoutliers (�2.5 standard deviations) computed for each angularrotation were excluded from the analyses (less than 2%). Nosignificant Pearson correlation was found between accuracy scoresand response times in both groups for both sessions, indicating nospeed–accuracy trade-off. Analyses of variance (ANOVAs) wereperformed on both mean accuracy (%) and response times (ms)with group (control vs. immobilized) as a between-subjects factorand session (pretest vs. posttest), hand (left vs. right), rotation (40°,80°, 120°, 160°), and direction (medial vs. lateral) as within-subjects factors. Post hoc comparisons were carried out by meansof a Newman–Keuls test. Alpha was set at .05 for all analyses.
Results
Accuracy. The ANOVA performed on the percentage ofcorrect responses yielded only a mean effect of rotation, F(3,60) � 17.95, p � .001, �p
2 � .47. Post hoc comparisons revealedthat correct responses were significantly more frequent for the 40°(M � 98%, SD � 6%), 80° (M � 98%, SD � 6%), and 120° (M �95%, SD � 7%) hand rotations than for the 160° (M � 89%, SD �11%; ps � .001), regardless of the group and the session.
Response times. The ANOVA yielded main effects of ses-sion, F(1, 20) � 20.6, p � .001, �p
2 � .51, and rotation, F(3, 60) �120.2, p � .001, �p
2 � .86. We also found two significant inter-actions: Group � Session, F(1, 20) � 5.6, p � .01, �p
2 � .17, andGroup � Session � Hand � Rotation, F(3, 60) � 2.8, p � .04,�p
2 � .12. As illustrated in Figure 2 and confirmed by post hoccomparisons, response times decreased from pretest to posttest forall the stimuli (i.e., from 40° to 160° for both right and left hands;ps � .01) for controls only. A reduced response time from pretestto posttest appeared for the 160° left hand stimuli in the immobi-lized group (p � .001), whereas performance did not vary for theother stimuli (ps � .25). Subsequent polynomial analyses revealedsignificant quadratic increases in response time with increasedrotation angles for both hands, tests, and groups, Fs(1, 20) � 5.52,ps � .02, except for posttest data of the left hand stimuli in theimmobilized group, F(1, 20) � 2.76, p � .11. The latter results areconsistent with lower response times observed for 160° left handstimuli following 48 hr of left hand immobilization.
Discussion
The general purpose of the present experiment was to explorethe influence of short-term upper limb immobilization on cognitive
Figure 1. Right and left back hand stimuli shown in different orientations(40°, 80°, 120°, and 160° following medial or lateral rotations).
625SHORT-TERM LIMB IMMOBILIZATION
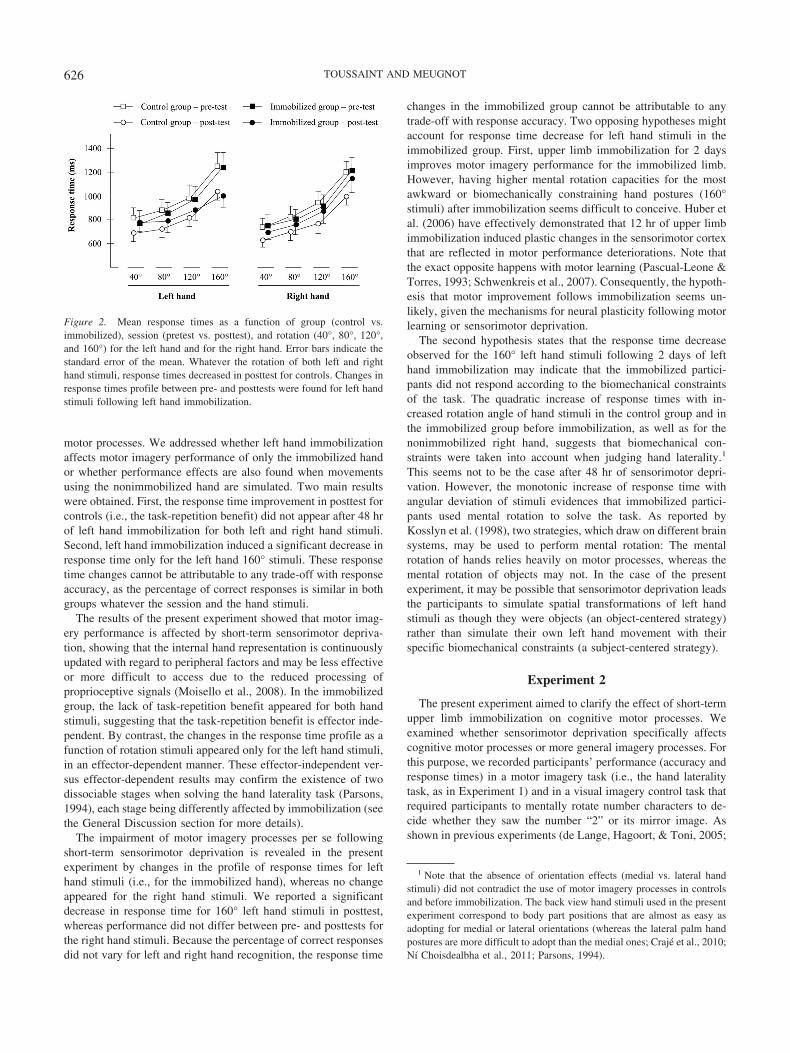
motor processes. We addressed whether left hand immobilizationaffects motor imagery performance of only the immobilized handor whether performance effects are also found when movementsusing the nonimmobilized hand are simulated. Two main resultswere obtained. First, the response time improvement in posttest forcontrols (i.e., the task-repetition benefit) did not appear after 48 hrof left hand immobilization for both left and right hand stimuli.Second, left hand immobilization induced a significant decrease inresponse time only for the left hand 160° stimuli. These responsetime changes cannot be attributable to any trade-off with responseaccuracy, as the percentage of correct responses is similar in bothgroups whatever the session and the hand stimuli.
The results of the present experiment showed that motor imag-ery performance is affected by short-term sensorimotor depriva-tion, showing that the internal hand representation is continuouslyupdated with regard to peripheral factors and may be less effectiveor more difficult to access due to the reduced processing ofproprioceptive signals (Moisello et al., 2008). In the immobilizedgroup, the lack of task-repetition benefit appeared for both handstimuli, suggesting that the task-repetition benefit is effector inde-pendent. By contrast, the changes in the response time profile as afunction of rotation stimuli appeared only for the left hand stimuli,in an effector-dependent manner. These effector-independent ver-sus effector-dependent results may confirm the existence of twodissociable stages when solving the hand laterality task (Parsons,1994), each stage being differently affected by immobilization (seethe General Discussion section for more details).
The impairment of motor imagery processes per se followingshort-term sensorimotor deprivation is revealed in the presentexperiment by changes in the profile of response times for lefthand stimuli (i.e., for the immobilized hand), whereas no changeappeared for the right hand stimuli. We reported a significantdecrease in response time for 160° left hand stimuli in posttest,whereas performance did not differ between pre- and posttests forthe right hand stimuli. Because the percentage of correct responsesdid not vary for left and right hand recognition, the response time
changes in the immobilized group cannot be attributable to anytrade-off with response accuracy. Two opposing hypotheses mightaccount for response time decrease for left hand stimuli in theimmobilized group. First, upper limb immobilization for 2 daysimproves motor imagery performance for the immobilized limb.However, having higher mental rotation capacities for the mostawkward or biomechanically constraining hand postures (160°stimuli) after immobilization seems difficult to conceive. Huber etal. (2006) have effectively demonstrated that 12 hr of upper limbimmobilization induced plastic changes in the sensorimotor cortexthat are reflected in motor performance deteriorations. Note thatthe exact opposite happens with motor learning (Pascual-Leone &Torres, 1993; Schwenkreis et al., 2007). Consequently, the hypoth-esis that motor improvement follows immobilization seems un-likely, given the mechanisms for neural plasticity following motorlearning or sensorimotor deprivation.
The second hypothesis states that the response time decreaseobserved for the 160° left hand stimuli following 2 days of lefthand immobilization may indicate that the immobilized partici-pants did not respond according to the biomechanical constraintsof the task. The quadratic increase of response times with in-creased rotation angle of hand stimuli in the control group and inthe immobilized group before immobilization, as well as for thenonimmobilized right hand, suggests that biomechanical con-straints were taken into account when judging hand laterality.1
This seems not to be the case after 48 hr of sensorimotor depri-vation. However, the monotonic increase of response time withangular deviation of stimuli evidences that immobilized partici-pants used mental rotation to solve the task. As reported byKosslyn et al. (1998), two strategies, which draw on different brainsystems, may be used to perform mental rotation: The mentalrotation of hands relies heavily on motor processes, whereas themental rotation of objects may not. In the case of the presentexperiment, it may be possible that sensorimotor deprivation leadsthe participants to simulate spatial transformations of left handstimuli as though they were objects (an object-centered strategy)rather than simulate their own left hand movement with theirspecific biomechanical constraints (a subject-centered strategy).
Experiment 2
The present experiment aimed to clarify the effect of short-termupper limb immobilization on cognitive motor processes. Weexamined whether sensorimotor deprivation specifically affectscognitive motor processes or more general imagery processes. Forthis purpose, we recorded participants’ performance (accuracy andresponse times) in a motor imagery task (i.e., the hand lateralitytask, as in Experiment 1) and in a visual imagery control task thatrequired participants to mentally rotate number characters to de-cide whether they saw the number “2” or its mirror image. Asshown in previous experiments (de Lange, Hagoort, & Toni, 2005;
1 Note that the absence of orientation effects (medial vs. lateral handstimuli) did not contradict the use of motor imagery processes in controlsand before immobilization. The back view hand stimuli used in the presentexperiment correspond to body part positions that are almost as easy asadopting for medial or lateral orientations (whereas the lateral palm handpostures are more difficult to adopt than the medial ones; Craje et al., 2010;Nı Choisdealbha et al., 2011; Parsons, 1994).
Figure 2. Mean response times as a function of group (control vs.immobilized), session (pretest vs. posttest), and rotation (40°, 80°, 120°,and 160°) for the left hand and for the right hand. Error bars indicate thestandard error of the mean. Whatever the rotation of both left and righthand stimuli, response times decreased in posttest for controls. Changes inresponse times profile between pre- and posttests were found for left handstimuli following left hand immobilization.
626 TOUSSAINT AND MEUGNOT

Kosslyn et al., 1998, 2001), mental rotation of nonbody images didnot elicit a motor strategy when participants were not asked toimagine themselves physically manipulating the objects. Thesestudies, as well as results obtained in Experiment 1, led us toassume evidence for immobilization-induced perturbations in themotor imagery task, but not in the visual imagery task.
A second aim of Experiment 2 was to examine whether thechanges in response time profile for left hand stimuli following lefthand immobilization can be replicated and whether these changesare correlated with participants’ imagery capacities. In the presentexperiment, the Vividness of Movement Imagery Questionnaire(VMIQ; Isaac, Marks, & Russell, 1986) was used as a measure ofvisual imagery (i.e., an object-centered strategy) and motor imag-ery (i.e., a subject-centered strategy) capacities. In the hand later-ality task, if the immobilization-induced effects result in a switchfrom a motor imagery to a visual imagery strategy, as suggested inExperiment 1, it may be possible that this switch could be easierfor participants with good vividness of visual imagery and/or forparticipants with poor vividness of motor imagery.
Method
Participants. Thirty-two right-handed university students(18–30 years of age) took part in the experiment. They weredivided into two groups: a control group (mean age � 19.9 years;eleven men, five women) and an immobilized group (mean age �19.0 years; nine men, seven women). None of the participantsparticipated in Experiment 1. They were healthy, had normal orcorrected-to-normal vision, and had no history of motor or neuro-logical disorders. The study protocol was approved by the localethics committee. All participants gave their written informedconsent prior to their inclusion in this study and received €20 fortheir participation. Before testing, all participants were naive aboutthe aims of the experiment.
Material and tasks. Participants performed two tasks: a handmental rotation task (i.e., the hand laterality task) and a nonbodymental rotation task. Only the images displayed on the computerscreen varied between the two tasks. The hand mental rotation taskwas similar to that used in Experiment 1 (see Figure 1). In thenonbody mental rotation task, participants had to report whetherthe number “2” image (sized 4.5 � 3 cm) was presented in itscanonical form or its mirror image (see Figure 3), regardless of its
angular rotation (40°, 80°, 120°, and 160° in a clockwise or acounterclockwise direction). In both tasks, participants respondedas accurately and as quickly as possible by pressing the appropriateleft or right marked keys with the index or the middle finger oftheir right nonimmobilized hand. As in Experiment 1, we used theE-Prime 2.0 software package to present stimuli and to record theparticipants’ responses (accuracy and response times).
Procedure. Before the start of the experiment, participantswere divided into two groups (control vs. immobilized). Theimmobilization procedure was identical to the one used in Exper-iment 1. We immobilized the participants’ left hand for 48 hr bymeans of a rigid splint. Each participant’s left arm was placed ina sling. In Experiment 2, to ensure that the participants kept theirleft hands at rest as much as possible during the 2 days ofimmobilization, we placed an actimeter (pounds/min) on theirimmobilized and nonimmobilized hands. Participants were askednever to remove the splint and the actimeter during the immobi-lization period. During the 48 hr immobilization period, on aver-age 258 � 81 pounds/min were recorded for the immobilized handand 822 � 212 pounds/min for the nonimmobilized hand. Fourparticipants were not retained in the present experiment; theiractimeter values for the left immobilized hand were more than 450pounds/min, indicating that they did not strictly follow the immo-bilization procedure. Four new participants took part in the immo-bilized group in order to maintain a balanced experimental design(these participants were included in the Participants sectionabove).
The motor imagery task (with hand stimuli) and the visualimagery task (with the number “2” stimuli) were performed duringtwo experimental sessions 48 hr apart (pre- and posttests; seeExperiment 1 for more details). The order of presentation of thetwo imagery tasks was counterbalanced across participants (hand–number tasks vs. number–hand tasks). For all participants, themotor imagery task and the visual imagery task were divided intotwo phases. During the first familiarization phase, the participantswere shown 16 randomly presented trials (2 hands or numbers �4 rotations � 2 directions). No time constraints were imposedduring the familiarization phase. During the second experimentalphase, the participants were shown six blocks of 16 trials (i.e., 96trials per participant) presented in a random order. The participantshad to respond as accurately and as quickly as possible.
At the beginning of the first experimental session, participantscompleted the VMIQ (Isaac et al., 1986). The VMIQ consists of 24items relevant to both the visual imagery of movement (i.e.,movement is simulated from an external, third-person perspective)and the imagery of kinesthetic sensations (i.e., movement is sim-ulated from an internal, first-person perspective). For each imageryperspective, measures of imagery vividness are assessed on a5-point Likert-type scale. High scores represent low vividness ofmovement imagery (for visual and for kinesthetic imagery), withthe reverse being true for low scores. Mean VMIQ scores areillustrated in Table 1.
Data analysis. Only data from correct responses were usedsubsequently to analyze response times. Moreover, response timeoutliers (�2.5 standard deviations) computed for each angularrotation were excluded from the analyses (less than 2.5% in eachimagery task). For the hand mental rotation task, ANOVAs wereperformed on mean accuracy (%) and response times (ms) withgroup (control vs. immobilized) and task order (hand–number vs.
Figure 3. Number “2” and its mirror image shown in different orienta-tions (40°, 80°, 120°, and 160° following clockwise or counterclockwiserotations).
627SHORT-TERM LIMB IMMOBILIZATION
number–hand) as between-subjects factors and session (pretest vs.posttest), hand (left vs. right), rotation (40°, 80°, 120°, 160°) anddirection (medial vs. lateral) as within-subjects factors. For thenumber mental rotation task, ANOVAs were performed withgroup and task order (hand–number vs. number– hand) asbetween-subjects factors and session, image (number “2” vs. itsmirror image), rotation, and direction (clockwise vs. counterclock-wise) as within-subjects factors. Post hoc comparisons were car-ried out by means of a Newman–Keuls test. Alpha was set at .05for all analyses.
Results
The hand mental rotation task. This task aimed to examinewhether left hand immobilization affects motor imagery processesas observed in Experiment 1. We expected immobilization toaffect the task-repetition benefit and induce changes in the re-sponse time profile for left hand stimuli. Moreover, in this section,we also examine whether the changes in response time profile forleft hand stimuli are correlated with participants’ imagery capac-ities (i.e., VMIQ scores from an external versus an internal per-spective).
Accuracy. The ANOVA performed on the percentage ofcorrect responses yielded only a mean effect of rotation, F(3,84) � 26.58, p � .001, �p
2 � .49. Post hoc comparisons revealedthat correct responses were significantly more frequent for the 40°(M � 95%, SD � 9%) and 80° (M � 95%, SD � 8%) handrotations than for the 160° (M � 87%, SD � 16%; ps � .001), aswell as for the 120° (M � 93%, SD � 11%) than for the 160°stimuli (p � .03), regardless of the group and the session.
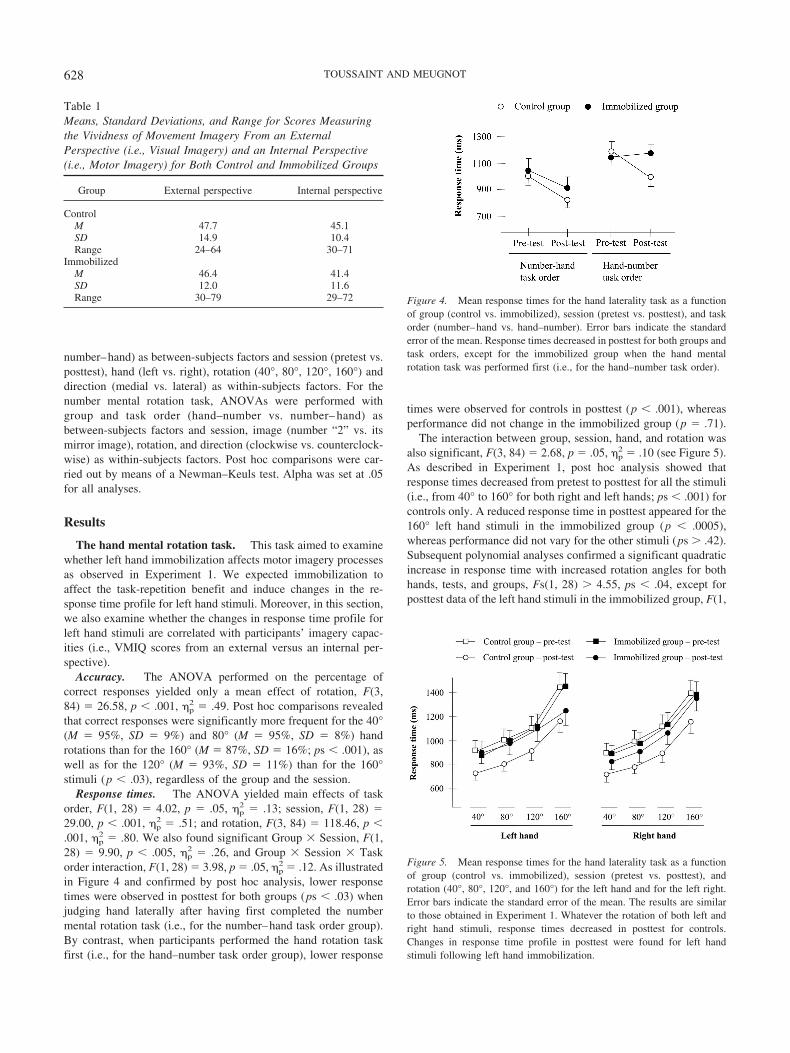
Response times. The ANOVA yielded main effects of taskorder, F(1, 28) � 4.02, p � .05, �p
2 � .13; session, F(1, 28) �29.00, p � .001, �p
2 � .51; and rotation, F(3, 84) � 118.46, p �.001, �p
2 � .80. We also found significant Group � Session, F(1,28) � 9.90, p � .005, �p
2 � .26, and Group � Session � Taskorder interaction, F(1, 28) � 3.98, p � .05, �p
2 � .12. As illustratedin Figure 4 and confirmed by post hoc analysis, lower responsetimes were observed in posttest for both groups (ps � .03) whenjudging hand laterally after having first completed the numbermental rotation task (i.e., for the number–hand task order group).By contrast, when participants performed the hand rotation taskfirst (i.e., for the hand–number task order group), lower response
times were observed for controls in posttest (p � .001), whereasperformance did not change in the immobilized group (p � .71).
The interaction between group, session, hand, and rotation wasalso significant, F(3, 84) � 2.68, p � .05, �p
2 � .10 (see Figure 5).As described in Experiment 1, post hoc analysis showed thatresponse times decreased from pretest to posttest for all the stimuli(i.e., from 40° to 160° for both right and left hands; ps � .001) forcontrols only. A reduced response time in posttest appeared for the160° left hand stimuli in the immobilized group (p � .0005),whereas performance did not vary for the other stimuli (ps � .42).Subsequent polynomial analyses confirmed a significant quadraticincrease in response time with increased rotation angles for bothhands, tests, and groups, Fs(1, 28) � 4.55, ps � .04, except forposttest data of the left hand stimuli in the immobilized group, F(1,
Table 1Means, Standard Deviations, and Range for Scores Measuringthe Vividness of Movement Imagery From an ExternalPerspective (i.e., Visual Imagery) and an Internal Perspective(i.e., Motor Imagery) for Both Control and Immobilized Groups
Group External perspective Internal perspective
ControlM 47.7 45.1SD 14.9 10.4Range 24–64 30–71
ImmobilizedM 46.4 41.4SD 12.0 11.6Range 30–79 29–72 Figure 4. Mean response times for the hand laterality task as a function
of group (control vs. immobilized), session (pretest vs. posttest), and taskorder (number–hand vs. hand–number). Error bars indicate the standarderror of the mean. Response times decreased in posttest for both groups andtask orders, except for the immobilized group when the hand mentalrotation task was performed first (i.e., for the hand–number task order).
Figure 5. Mean response times for the hand laterality task as a functionof group (control vs. immobilized), session (pretest vs. posttest), androtation (40°, 80°, 120°, and 160°) for the left hand and for the left right.Error bars indicate the standard error of the mean. The results are similarto those obtained in Experiment 1. Whatever the rotation of both left andright hand stimuli, response times decreased in posttest for controls.Changes in response time profile in posttest were found for left handstimuli following left hand immobilization.
628 TOUSSAINT AND MEUGNOT
28) � 1.93, p � .19, which is consistent with lower response timesobserved for 160° left hand stimuli following left hand immobili-zation.
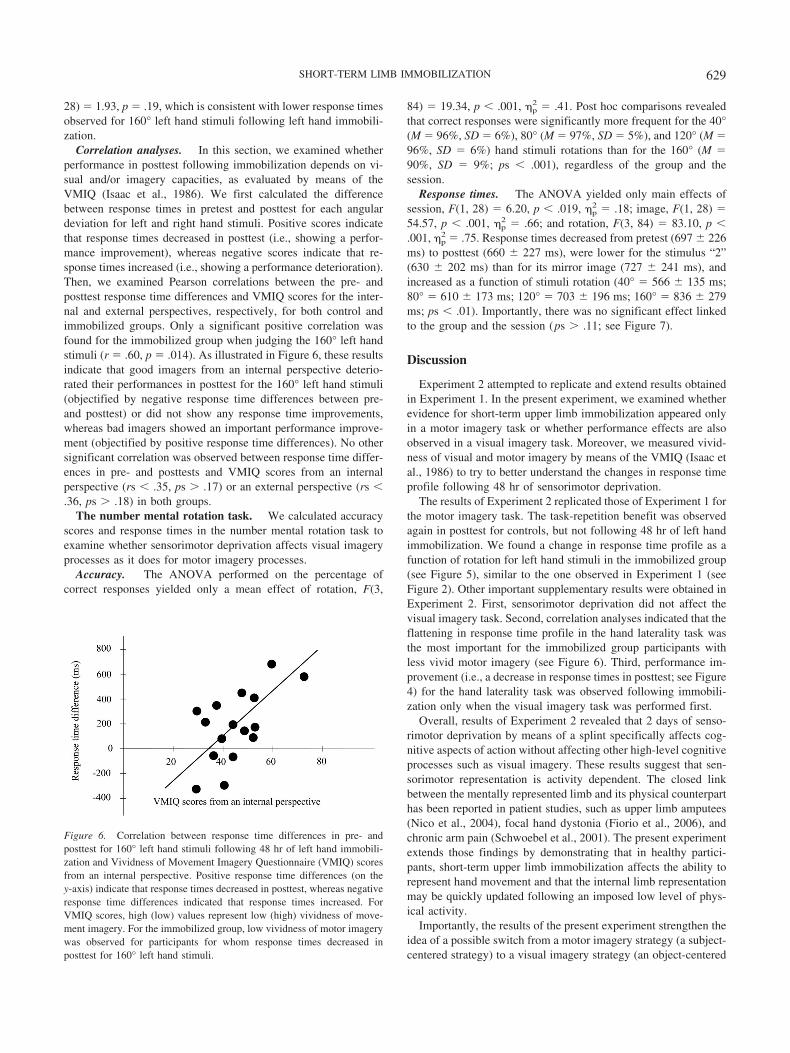
Correlation analyses. In this section, we examined whetherperformance in posttest following immobilization depends on vi-sual and/or imagery capacities, as evaluated by means of theVMIQ (Isaac et al., 1986). We first calculated the differencebetween response times in pretest and posttest for each angulardeviation for left and right hand stimuli. Positive scores indicatethat response times decreased in posttest (i.e., showing a perfor-mance improvement), whereas negative scores indicate that re-sponse times increased (i.e., showing a performance deterioration).Then, we examined Pearson correlations between the pre- andposttest response time differences and VMIQ scores for the inter-nal and external perspectives, respectively, for both control andimmobilized groups. Only a significant positive correlation wasfound for the immobilized group when judging the 160° left handstimuli (r � .60, p � .014). As illustrated in Figure 6, these resultsindicate that good imagers from an internal perspective deterio-rated their performances in posttest for the 160° left hand stimuli(objectified by negative response time differences between pre-and posttest) or did not show any response time improvements,whereas bad imagers showed an important performance improve-ment (objectified by positive response time differences). No othersignificant correlation was observed between response time differ-ences in pre- and posttests and VMIQ scores from an internalperspective (rs � .35, ps � .17) or an external perspective (rs �.36, ps � .18) in both groups.
The number mental rotation task. We calculated accuracyscores and response times in the number mental rotation task toexamine whether sensorimotor deprivation affects visual imageryprocesses as it does for motor imagery processes.
Accuracy. The ANOVA performed on the percentage ofcorrect responses yielded only a mean effect of rotation, F(3,
84) � 19.34, p � .001, �p2 � .41. Post hoc comparisons revealed
that correct responses were significantly more frequent for the 40°(M � 96%, SD � 6%), 80° (M � 97%, SD � 5%), and 120° (M �96%, SD � 6%) hand stimuli rotations than for the 160° (M �90%, SD � 9%; ps � .001), regardless of the group and thesession.
Response times. The ANOVA yielded only main effects ofsession, F(1, 28) � 6.20, p � .019, �p
2 � .18; image, F(1, 28) �54.57, p � .001, �p
2 � .66; and rotation, F(3, 84) � 83.10, p �.001, �p
2 � .75. Response times decreased from pretest (697 � 226ms) to posttest (660 � 227 ms), were lower for the stimulus “2”(630 � 202 ms) than for its mirror image (727 � 241 ms), andincreased as a function of stimuli rotation (40° � 566 � 135 ms;80° � 610 � 173 ms; 120° � 703 � 196 ms; 160° � 836 � 279ms; ps � .01). Importantly, there was no significant effect linkedto the group and the session (ps � .11; see Figure 7).
Discussion
Experiment 2 attempted to replicate and extend results obtainedin Experiment 1. In the present experiment, we examined whetherevidence for short-term upper limb immobilization appeared onlyin a motor imagery task or whether performance effects are alsoobserved in a visual imagery task. Moreover, we measured vivid-ness of visual and motor imagery by means of the VMIQ (Isaac etal., 1986) to try to better understand the changes in response timeprofile following 48 hr of sensorimotor deprivation.
The results of Experiment 2 replicated those of Experiment 1 forthe motor imagery task. The task-repetition benefit was observedagain in posttest for controls, but not following 48 hr of left handimmobilization. We found a change in response time profile as afunction of rotation for left hand stimuli in the immobilized group(see Figure 5), similar to the one observed in Experiment 1 (seeFigure 2). Other important supplementary results were obtained inExperiment 2. First, sensorimotor deprivation did not affect thevisual imagery task. Second, correlation analyses indicated that theflattening in response time profile in the hand laterality task wasthe most important for the immobilized group participants withless vivid motor imagery (see Figure 6). Third, performance im-provement (i.e., a decrease in response times in posttest; see Figure4) for the hand laterality task was observed following immobili-zation only when the visual imagery task was performed first.
Overall, results of Experiment 2 revealed that 2 days of senso-rimotor deprivation by means of a splint specifically affects cog-nitive aspects of action without affecting other high-level cognitiveprocesses such as visual imagery. These results suggest that sen-sorimotor representation is activity dependent. The closed linkbetween the mentally represented limb and its physical counterparthas been reported in patient studies, such as upper limb amputees(Nico et al., 2004), focal hand dystonia (Fiorio et al., 2006), andchronic arm pain (Schwoebel et al., 2001). The present experimentextends those findings by demonstrating that in healthy partici-pants, short-term upper limb immobilization affects the ability torepresent hand movement and that the internal limb representationmay be quickly updated following an imposed low level of phys-ical activity.
Importantly, the results of the present experiment strengthen theidea of a possible switch from a motor imagery strategy (a subject-centered strategy) to a visual imagery strategy (an object-centered
Figure 6. Correlation between response time differences in pre- andposttest for 160° left hand stimuli following 48 hr of left hand immobili-zation and Vividness of Movement Imagery Questionnaire (VMIQ) scoresfrom an internal perspective. Positive response time differences (on they-axis) indicate that response times decreased in posttest, whereas negativeresponse time differences indicated that response times increased. ForVMIQ scores, high (low) values represent low (high) vividness of move-ment imagery. For the immobilized group, low vividness of motor imagerywas observed for participants for whom response times decreased inposttest for 160° left hand stimuli.
629SHORT-TERM LIMB IMMOBILIZATION

strategy) following sensorimotor deprivation. First, evidence forimmobilization-induced motor imagery perturbations (i.e., for thehand laterality task) that varied with task order might corroboratethis idea. The task-repetition benefit when judging hand lateralitywas found following immobilization only when participants per-formed the visual imagery task first (for the number–hand taskorder group). The present result suggests that the ability for par-ticipants to disengage from less efficient sensorimotor processes(or subject-centered strategy) due to immobilization could befacilitated by the previous use of a visual, object-centered imagerystrategy. Second, correlation analyses revealed that participantswith good vividness of motor imagery had more difficulties thanparticipants with poor vividness of motor imagery to mentallysimulate the largest amplitude movements (160° rotation stimuli)corresponding to the immobilized hand: The difference calculatedbetween their response times in pre- and posttest (with eithernegative values or values close to zero; see Figure 6) confirmstheir difficulties. By contrast, poor imagers from an internal per-spective (subject-centered strategy) had lower response times fol-lowing immobilization (posttest vs. pretest), as revealed by posi-tive response time differences. Consequently, when participantshad fewer capacities to mentally simulate a movement from asubject-centered strategy, they more easily switched from asubject-centered to an object-centered strategy when the sensori-motor system was affected. We are conscious of the speculativenature of our comments and of the necessity to lead other research-ers to investigate in more depth the possible flexibility in mentalrotation strategies according to the constraints of the situation aswell as individual characteristics.
General Discussion
The current study shows for the first time that short-term upperlimb immobilization specifically affects the cognitive motor pro-cesses without affecting other higher cognitive processes, such asvisual imagery. In healthy subjects, the internal hand representa-tion can be quickly updated following sensorimotor deprivation.Importantly, the embodied nature of the movement simulationprocesses is confirmed by changes in response time profile as a
function of stimuli rotation in the immobilized group, whereas thefirst stage devoted to the identification–recognition processes (Par-sons, 1994) of the visually presented hand stimuli appeared effec-tor independent. The two-stage recognition processes have beennoted by Parsons (1994): an implicit identification–recognitionprocess based on a whole body representation, followed by aconfirmatory process requiring the mental rotation of either the leftor right hand. The temporal evolution of the identification–recognition and mental rotation processes (i.e., recognition priorrotation) has been confirmed by event-related potential recordings(Overney, Michel, Harris, & Pegna, 2005). The lack of task-repetition benefit observed in the present study (Experiments 1 and2) after left hand immobilization for both left and right handstimuli suggests that the initial analyses of the hand stimuli (whichis based on a stored whole body schema representation) may betime-consuming due to difficulties accessing sensorimotor repre-sentations. This suggestion is consistent with immobilization-neural changes reported in a transcranial magnetic stimulationstudy (Facchini et al., 2002). Sensorimotor restriction induced thereduction of cortical excitability or the synaptic depression of therestricted limb, but also on homotopic regions of the cortex. Inthe same vein as results reported in our present experiment, im-pairments of limb representation objectified by delayed responsetimes in the hand laterality task have been reported in patients withunilateral phantom upper limb pain compared with healthy sub-jects (Reinersmann et al., 2010). Importantly, the response timesfor hand stimuli corresponding to the patients’ affected limbs wereequally delayed as that of hand stimuli of the contralateral, unaf-fected limbs. However, because information on the response timeprofile as a function of angular rotation of hand stimuli was notreported in Reinersmann et al.’s study (2010), we cannot knowwhether the difficulties encountered by their patients resulted fromdeficits associated with the first recognition stage or the lattermental rotation stage.
In the present study, the effector-dependent changes induced byonly 48 hr of left hand immobilization on movement simulationmay be an indicator of the continuously updated sensorimotormemories in the contralateral sensorimotor area due to the reducedprocessing of proprioceptive feedback relative to the immobilizedhand. Moreover, as suggested at the end of Experiment 1, it maybe possible that the sensorimotor deprivation causes the sensori-motor system solicited by motor imagery to become less efficientand thus led the immobilized participants to favor nonmotor im-agery processes (i.e., to use an object-centered strategy rather thana subject-centered strategy) to solve the hand laterality task. Asimilar explanation has been proposed by Wilson et al. (2004),who reported a small trade-off (similar to the one observed in ourexperiments) between response times and hand rotation stimuli forchildren with a developmental coordination disorder comparedwith control children. In the same vein, de Lange et al. (2005)reported that response times increased as a function of stimulirotation more in the hand mental rotation task than in the lettermental rotation task. Consequently, the nonsignificant quadraticincrease of response times with increase rotation angle of handstimuli after immobilization leads us to suggest that the reducedprocessing of sensorimotor information due to immobilization ledparticipants to switch from a less efficient motor to a more effi-cient visual imagery strategy to solve the hand recognition task.
Figure 7. Mean response times as a function of group (control vs.immobilized), session (pretest vs. posttest), and rotation (40°, 80°, 120°,and 160°) for the stimulus “2” and for its mirror image. Error bars indicatethe standard error of the mean. Analysis of variance yielded only maineffects of test, stimulus, and rotation.
630 TOUSSAINT AND MEUGNOT

In conclusion, the sensorimotor system quickly adapts to animposed low level of physical activity, as it does following motortraining (Pascual-Leone & Torres, 1993; Schwenkreis et al., 2007),except that the opposite happens. Even a brief period of sensori-motor deprivation can induce some deficits in sensorimotor rep-resentations and be the origin of motor performance deteriorations.This means that measures should be taken to try to counteract thedeleterious effect of immobilization on the functioning of thecentral nervous system. Explicit motor imagery instructions arewidely used in sports (see Holmes & Calmels, 2008, for a review)as well as by nonathletes (Gentili, Han, Schweighofer, & Papax-anthis, 2010; Jackson, Lafleur, Malouin, Richards, & Doyon,2003; Toussaint & Blandin, 2010; Toussaint, Robin, & Blandin,2010) to improve motor performance. Does this mean that motorimagery training can be used to prevent the immobilized-inducedsensorimotor alterations as soon as a splint is applied? Futureresearch is necessary to investigate this topic.
References
Badets, A., & Pesenti, M. (2010). Creating number semantics throughfinger movement perception. Cognition, 115, 46 –53. doi:10.1016/j.cognition.2009.11.007
Craje, C., van Elk, M., Beeren, M., van Schie, T. T., Bekkering, H., &Steenbergen, B. (2010). Compromised motor planning and motor imag-ery in right hemiparetic cerebral palsy. Research in DevelopmentalDisabilities, 31, 1313–1322. doi:10.1016/j.ridd.2010.07.010
Curtze, C., Otten, B., & Postema, K. (2010). Effects of lower limbamputation on the mental rotation of feet. Experimental Brain Research,201, 527–534. doi:10.1007/s00221-009-2067-z
Dalecki, M., Hoffmann, U., & Bock, O. (2012). Mental rotation of letters,body parts and complex scenes: Separate or common mechanisms?Human Movement Science. Advance online publication. doi:10.1016/j.humov.2011.12.001
de Lange, F. P., Hagoort, P., & Toni, I. (2005). Neural topography andcontent of movement representations. Journal of Cognitive Neurosci-ence, 17, 97–112. doi:10.1162/0898929052880039
de Lange, F. P., Helmich, R. C., & Toni, I. (2006). Posture influencesmotor imagery: An fMRI study. NeuroImage, 33, 609 – 617. doi:10.1016/j.neuroimage.2006.07.017
Facchini, S., Romani, M., Tinazzi, M., & Aglioti, S. M. (2002). Time-related changes of excitability of the human motor system contingentupon immobilization of the ring and little fingers. Clinical Neurophys-iology, 113, 367–375. doi:10.1016/S1388-2457(02)00009-3
Fiorio, M., Tinazzi, M., & Aglioti, S. M. (2006). Selective impairment ofhand mental rotation in patients with focal hand dystonia. Brain, 129,47–54. doi:10.1093/brain/awh630
Ganis, G., Keenan, J. P., Kosslyn, S. M., & Pascual-Leone, A. P. (2000).Transcranial magnetic stimulation of primary motor cortex affects men-tal rotation. Cerebral Cortex, 10, 175–180. doi:10.1093/cercor/10.2.175
Gentili, R., Han, C. E., Schweighofer, N., & Papaxanthis, C. (2010). Motorlearning without doing: Trial-by-trial improvement in motor perfor-mance during mental training. Journal of Neurophysiology, 104, 774–783. doi:10.1152/jn.00257.2010
Glenberg, A. M., Sato, M., & Cattaneo, L. (2008). Use-induced motorplasticity affects the processing of abstract and concrete language. Cur-rent Biology, 18, R290–R291. doi:10.1016/j.cub.2008.02.036
Hlustik, P., Solodkin, A., Noll, D. C., & Small, S. L. (2004). Corticalplasticity during three-week motor skill learning. Journal of ClinicalNeurophysiology, 21, 180 –191. doi:10.1097/00004691-200405000-00006
Holmes, P., & Calmels, C. (2008). A neuroscientific review of imagery and
observation use in sport. Journal of Motor Behavior, 40, 433–445.doi:10.3200/JMBR.40.5.433-445
Huber, R., Ghilardi, M. F., Massimini, M., Ferrarelli, F., Riedner, B. A.,Peterson, M. J., & Tononi, G. (2006). Arm immobilization causescortical plastic changes and locally decreases sleep slow wave activity.Nature Neuroscience, 9, 1169–1176. doi:10.1038./nn1758
Ionta, S., & Blanke, O. (2009). Differential influence of hands posture onmental rotation of hands and feet in left and right handers. ExperimentalBrain Research, 195, 207–217. doi:10.1007/s00221-009-1770-0
Ionta, S., Fourkas, A. D., Fiorio, M., & Aglioti, S. M. (2007). The influenceof hands posture on mental rotation of hands and feet. ExperimentalBrain Research, 183, 1–7. doi:10.1007/s00221-007-1020-2
Isaac, A., Marks, D. F., & Russell, D. G. (1986). An instrument forassessing imagery of movement: The Vividness of Movement ImageryQuestionnaire (VMIQ). Journal of Mental Imagery, 10, 23–30.
Jackson, P. L., Lafleur, M. F., Malouin, F., Richards, C. L., & Doyon, J.(2003). Functional cerebral reorganization following motor sequencelearning through mental practice with motor imagery. NeuroImage, 20,1171–1180. doi:10.1016/S1053-8119(03)00369-0
Jeannerod, M. (2001). Neural simulation of action: A unifying mechanismfor motor cognition. NeuroImage, 14, 429 – 439. doi:10.1006/nimg.2001.0832
Johnson, S. H. (2000). Imagining the impossible: Intact motor representa-tions in hemiplegics. NeuroReport, 11, 729 –732. doi:10.1097/00001756-200003200-00015
Johnson, S. H., Sprehn, G., & Saykin, A. J. (2002). Intact motor imageryin chronic upper limb hemiplegics: Evidence for activity-independentaction representations. Journal of Cognitive Neuroscience, 14, 841–852.doi:10.1162/089892902760191072
Jola, C., & Mast, F. W. (2005). Mental object rotation and egocentric bodytransformation: Two dissociable processes? Spatial Cognition and Com-putation, 5, 217–237.
Kaneko, F., Murakami, T., Onari, K., Kurumadani, H. & Kawaguchi, K.(2003). Decreased cortical excitability during motor imagery after disuseof an upper limb in humans. Clinical Neurophysiology, 114, 2397–2403.doi:10.1016/S1388-2457(03)00245-1
Kosslyn, S. M., DiGirolamo, G. J., Thompson, W. L., & Alpert, N. M.(1998). Mental rotation of objects versus hands: Neural mechanismsrevealed by positron emission tomography. Psychophysiology, 35, 151–161. doi:10.1111/1469-8986.3520151
Kosslyn, S. M., Thompson, W. L., Wraga, M., & Alpert, N. M. (2001).Imagining rotation by endogenous versus exogenous forces: Distinctneural mechanisms. NeuroReport, 12, 2519 –2525. doi:10.1097/00001756-200108080-00046
Liepert, J., Tegenthoff, M., & Malin, J. P. (1995). Changes of corticalmotor area size during immobilization. Electroencephalography andClinical Neurophysiology, 97, 382–386. doi:10.1016/0924-980X(95)00194-P
Moisello, C., Bove, M., Huber, R., Giovanni, A., Battaglia, F., Tononi, G.,& Ghilardi, M. F. (2008). Short-term limb immobilization affects motorperformance. Journal of Motor Behavior, 40, 165–176. doi:10.3200/JMBR.40.2.165-176
N´i Choisdealbha, A. N., Brady, N., & Maguinness, C. (2011). Differingroles for the dominant and non-dominant hands in the hand lateralitytask. Experimental Brain Research, 211, 73–85. doi:10.1007/s00221-011-2652-9
Nico, D., Daprati, E., Rigal, F., Parsons, L. M., & Sirigu, A. (2004). Leftand right hand recognition in upper limb amputees. Brain, 127, 120–132. doi:10.1093/brain/awh006
Overney, L. S., Michel, C. M., Harris, I. M., & Pegna, A. J. (2005).Cerebral processes in mental transformations of body parts: Recognitionprior to rotation. Cognitive Brain Research, 25, 722–734. doi:10.1016/j.cogbrainres.2005.09.024
Parsons, L. M. (1987). Imagined spatial transformations of one’s hand and
631SHORT-TERM LIMB IMMOBILIZATION

feet. Cognitive Psychology, 19, 178 –241. doi:10.1016/0010-0285(87)90011-9
Parsons, L. M. (1994). Temporal and kinematic properties of motor be-havioral reflected in mentally simulated action. Journal of ExperimentalPsychology: Human Perception and Performance, 20, 709–730. doi:10.1037/0096-1523.20.4.709
Parsons, L. M., Gabrieli, J. D., Phelps, E. A., & Gazzaniga, M. S. (1998).Cerebrally lateralized mental representations of hand shape and move-ment. Journal of Neuroscience, 18, 6539–6548.
Pascual-Leone, A., & Torres, F. (1993). Plasticity of the sensorimotorcortex representation of the reading finger in Braille readers. Brain, 116,39–52. doi:10.1093/brain/116.1.39
Reinersmann, A., Haarmeyer, G. S., Blankenburg, M., Frettlöh, J., Kru-mova, E. K., Ocklenburg, S., & Maier, C. (2010). Left is where the L isright. Significantly delayed reaction time in limb laterality recognition inboth CRPS and phantom limb pain patients. Neuroscience Letters, 486,240–245. doi:10.1016/j.neulet.2010.09.062
Sainburg, R. L., Ghilardi, M. F., Poizner, H., & Ghez, C. (1995). Controlof limb dynamics in normal subjects and patients without propriocep-tion. Journal of Neurophysiology, 73, 820–835.
Schwenkreis, P., El Tom, S., Ragert, P., Pleger, B., Tegenthoff, M., &Dinse, H. R. (2007). Assessment of sensorimotor cortical representationasymmetries and motor skills in violin players. European Journal ofNeuroscience, 26, 3291–3302. doi:10.1111/j.1460-9568.2007.05894.x
Schwoebel, J., Friedman, R., Duda, N., & Coslett, H. B. (2001). Pain andthe body schema: Evidence for peripheral effects on mental representa-tions of movement. Brain, 124, 2098 –2104. doi:10.1093/brain/124.10.2098
Seki, K., Taniguchi, Y., & Narusawa, M. (2001). Alterations in contractileproperties of human skeletal muscle induced by joint immobilization.Journal of Physiology, 530, 521–532. doi:10.1111/j.1469-7793.2001.0521k.x
Sekiyama, K. (1982). Kinesthetic aspects of mental representations in theidentification of left and right hands. Perception & Psychophysics, 32,89–95. doi:10.3758/BF03204268
Shenton, J. T., Schwoebel, J., & Coslett, H. B. (2004). Mental motor
imagery and the body schema: Evidence for proprioceptive dominance.Neuroscience Letters, 370, 19–24. doi:10.1016/j.neulet.2004.07.053
Sirigu, A., & Duhamel, J. R. (2001). Motor and visual imagery as twocomplementary but neutrally dissociable mental processes. Journal of Cog-nitive Neuroscience, 13, 910–919. doi:10.1162/089892901753165827
Tomasino, B., & Rumiati, R. I. (2004). Effects of strategies on mentalrotation and hemispheric lateralization: Neuropsychological evidence.Journal of Cognitive Neuroscience, 16, 878 – 888. doi:10.1162/089892904970753
Tomasino, B., Rumiati, R. I., & Umilta, C. A. (2003). Selective deficit ofmotor imagery as tapped by a left–right decision of visually presentedhands. Brain and Cognition, 53, 376–380. doi:10.1016/S0278-2626(03)00147-7
Tomasino, B., Weiss, P. H., & Fink, G. R. (2010). To move or not to move:Imperatives modulate action-related verb processing in the motor sys-tem. Neuroscience, 169, 246 –258. doi:10.1016/j.neuroscience.2010.04.039
Toussaint, L., & Blandin, Y. (2010). On the role of imagery modalities onmotor learning. Journal of Sports Sciences, 28, 497–504. doi:10.1080/02640410903555855
Toussaint, L., Robin, N., & Blandin, Y. (2010). On the content of senso-rimotor representations after actual and motor imagery practice. MotorControl, 14, 159–175.
Wilson, P. H., Maruff, P., Butson, M., Williams, J., Lum, J., & Thomas,P. R. (2004). Internal representation of movement in children withdevelopmental coordination disorder: A mental rotation task. Develop-mental Medicine & Child Neurology, 46, 754–759. doi:10.1111/j.1469-8749.2004.tb00995.x
Zanette, G., Tinazzi, M., Bonato, C., di Summa, A., Manganotti, P., Polo,A., & Fiaschi, A. (1997). Reversible changes of motor cortical outputsfollowing immobilization of the upper limb. Electroencephalographyand Clinical Neurophysiology, 105, 269 –279. doi:10.1016/S0924-980X(97)00024-6
Received July 21, 2011Revision received April 19, 2012
Accepted April 23, 2012 �
632 TOUSSAINT AND MEUGNOT
Related Documents