RESEARCH PAPER Short-term changes of fish assemblages observed in the near-pristine reefs of the Phoenix Islands Sangeeta Mangubhai • Ayron M. Strauch • David O. Obura • Gregory Stone • Randi D. Rotjan Received: 14 August 2012 / Accepted: 20 September 2013 / Published online: 24 November 2013 Ó Springer Science+Business Media Dordrecht 2013 Abstract Climate change-related disturbances are increasingly recognized as critical threats to biodiver- sity and species abundance. On coral reefs, climate disturbances have known consequences for reef fishes, but it is often difficult to isolate the effect of coral bleaching from preceding or simultaneous distur- bances such as fishing, pollution, and habitat loss. In this study, pre-bleaching surveys of fish family assemblages in the remote Phoenix Islands in 2002 are compared to post-bleaching in 2005, following severe thermal stress. Post-bleaching, total coral cover decreased substantially, as did the combined abun- dance of all fish families. Yet, changes in abundance for specific fish families were not uniform, and varied greatly from site to site. Of the 13 fish families examined, 3 exhibited significant changes in abun- dance from 2002 to 2005, regardless of site (Carang- idae, Chaetodontidae, and serranid subfamily Epinephelinae). For these families, we explored whether changes in abundance were related to island type (island vs atoll) and/or declining coral cover (percent change). Carangidae on islands experienced larger changes in abundance than those on atolls, though declines in abundance over time were not associated with changes in live coral cover. In contrast, for Chaetodontidae, declines in abundance over time were most dramatic on atolls, and were also associated with changes in live coral cover. The remoteness of the Phoenix Islands excludes many typical local anthropogenic stressors as drivers of short-term changes; observed changes are instead more likely attributed to natural variation in fish populations, or associated with coral loss following the 2002–2003 major thermal stress event. Keywords Climate change Á Disturbance Á Coral reef Á Kiribati Á Chaetodontidae Introduction Disturbance plays an important role in determining species diversity and community structure (Petraitis et al. 1989; Pickett and White 1986; Sousa 1984). Climate change-related disturbances are increasingly recognized as critical threats to biodiversity (Thomas S. Mangubhai IUCN Oceania Regional Office, 5 Ma’afu Street, Suva, Fiji S. Mangubhai Á A. M. Strauch Á D. O. Obura Á G. Stone Á R. D. Rotjan (&) John H. Prescott Marine Laboratory, New England Aquarium, One Central Wharf, Boston, MA 02110, USA e-mail: [email protected] D. O. Obura CORDIO East Africa, P.O. Box 10135, Bamburi, Mombasa 80101, Kenya G. Stone Global Marine Division, Conservation International, 2011 Crystal Drive, Arlington, VA 22202, USA 123 Rev Fish Biol Fisheries (2014) 24:505–518 DOI 10.1007/s11160-013-9327-5

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH PAPER
Short-term changes of fish assemblages observedin the near-pristine reefs of the Phoenix Islands
Sangeeta Mangubhai • Ayron M. Strauch •
David O. Obura • Gregory Stone •
Randi D. Rotjan
Received: 14 August 2012 / Accepted: 20 September 2013 / Published online: 24 November 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Climate change-related disturbances are
increasingly recognized as critical threats to biodiver-
sity and species abundance. On coral reefs, climate
disturbances have known consequences for reef fishes,
but it is often difficult to isolate the effect of coral
bleaching from preceding or simultaneous distur-
bances such as fishing, pollution, and habitat loss. In
this study, pre-bleaching surveys of fish family
assemblages in the remote Phoenix Islands in 2002
are compared to post-bleaching in 2005, following
severe thermal stress. Post-bleaching, total coral cover
decreased substantially, as did the combined abun-
dance of all fish families. Yet, changes in abundance
for specific fish families were not uniform, and varied
greatly from site to site. Of the 13 fish families
examined, 3 exhibited significant changes in abun-
dance from 2002 to 2005, regardless of site (Carang-
idae, Chaetodontidae, and serranid subfamily
Epinephelinae). For these families, we explored
whether changes in abundance were related to island
type (island vs atoll) and/or declining coral cover
(percent change). Carangidae on islands experienced
larger changes in abundance than those on atolls,
though declines in abundance over time were not
associated with changes in live coral cover. In
contrast, for Chaetodontidae, declines in abundance
over time were most dramatic on atolls, and were also
associated with changes in live coral cover. The
remoteness of the Phoenix Islands excludes many
typical local anthropogenic stressors as drivers of
short-term changes; observed changes are instead
more likely attributed to natural variation in fish
populations, or associated with coral loss following
the 2002–2003 major thermal stress event.
Keywords Climate change � Disturbance �Coral reef � Kiribati � Chaetodontidae
Introduction
Disturbance plays an important role in determining
species diversity and community structure (Petraitis
et al. 1989; Pickett and White 1986; Sousa 1984).
Climate change-related disturbances are increasingly
recognized as critical threats to biodiversity (Thomas
S. Mangubhai
IUCN Oceania Regional Office, 5 Ma’afu Street,
Suva, Fiji
S. Mangubhai � A. M. Strauch � D. O. Obura �G. Stone � R. D. Rotjan (&)
John H. Prescott Marine Laboratory, New England
Aquarium, One Central Wharf, Boston, MA 02110, USA
e-mail: [email protected]
D. O. Obura
CORDIO East Africa, P.O. Box 10135, Bamburi,
Mombasa 80101, Kenya
G. Stone
Global Marine Division, Conservation International,
2011 Crystal Drive, Arlington, VA 22202, USA
123
Rev Fish Biol Fisheries (2014) 24:505–518
DOI 10.1007/s11160-013-9327-5

et al. 2004; Balmford et al. 2005; Smale and Wernberg
2013); such disturbances are wide-ranging and include
acute as well as chronic events with varying frequency
and intensity (e.g. Hoegh-Guldberg et al. 2007).
Predicting the more subtle impacts of disturbance
is difficult, especially in highly diverse ecosystems
that are changing prior to the measurement of
historical baseline states. Such is the case for coral
reefs, which are among the most diverse and complex
ecosystems on the planet and for which little historical
data of pre-anthropogenic disturbance conditions are
available (though see Jackson 2001; Pandolfi et al.
2003).
Tropical reefs, created by coral animals, support
diverse communities of reef fishes, with over 4,000
tropical fish species worldwide (Allen 2007; Lieske
and Myers 1994). Declines in reef fish abundance have
been well documented on nearshore reefs, largely due
to direct causes such as overfishing, habitat loss, and
pollution (reviewed by Jones and Syms 1998; Wilson
et al. 2006). Similarly, an increasing number of studies
have shown that rising temperatures, increased storm
frequency and intensity, and habitat loss due to climate
change have substantial impacts on reef fishes (Mun-
day et al. 2008; Pratchett et al. 2008, 2011; Chong-
Seng et al. 2012). However, on most reefs, multiple
disturbances may occur simultaneously, thus con-
founding the influence of any one disturbance. Climate
change disturbance events are generally coupled with
local anthropogenic impacts (acute and/or chronic).
As a result, the majority of climate-related disturbance
reef research has inevitably occurred against a back-
drop of concurrent local anthropogenic stressors,
revealing the combined impacts of these stressors
instead of solely isolating the impacts of climatolog-
ical disturbances (but see Christensen et al. 1996;
Sandin et al. 2008).
Recent reviews and workshops have assessed
current knowledge gaps and highlighted the immedi-
ate need to explore reef fish responses to climate
change disturbances, with heavy emphasis on under-
standing how fish community dynamics and diversity
change as a result of habitat loss or changing habitat
composition (Munday et al. 2008; Wilson et al.
2010a). For example, fish diversity and abundance
decline with coral loss (Wilson et al. 2006; Graham
et al. 2006; Jones et al. 2004). Examining the
taxonomic and functional group responses to bleach-
ing is an active topic of research, and fishes do not
have a universal response to bleaching and/or coral
loss. For example, some groups, such as herbivores
that benefit from conversion of coral to algal cover in
the short term, have in some cases been shown to
increase following coral loss (Lindahl et al. 2001)
while fishes with a known coral-dependency cannot
tolerate coral mortality (e.g. Bonin et al. 2009). A
recent review also highlighted the need for tropical
field observations of response to climate change
events to complement the majority of studies that
have thus far focused on lab manipulations (Wernberg
et al. 2012a, b).
The Phoenix Islands, located just south of the
equator in the Central Pacific in the Republic of
Kiribati, are ideally located for examining the natural
variation of fish assemblages over time, as well as the
potential impacts of coral bleaching. Phoenix Islands
reefs are protected from local anthropogenic stressors
such as fishing, coastal development, and pollution.
These islands are largely uninhabited, and are among
the most remote reefs on earth (3 days by sea from the
closest port). Kanton is the only inhabited atoll, and
between 2000 and 2012 has hosted a caretaker
population of fewer than 50 residents. Orona was
settled briefly from 2001 to 2003 under a Kiribati
government trial resettlement scheme, while the
remaining atolls have been uninhabited since the
1960s.
Expeditions to the Phoenix Islands in 2000 and
2002 found the reefs to be in excellent condition with
coral cover averaging 45.1 and 58.1 %, respectively,
with a maximum cover of 100 % live coral (Obura
et al. 2011; Obura and Stone 2002) and 516 species of
fishes (Allen and Bailey 2011). However, during the
expedition in 2002 the early stages of coral bleaching
were observed on one of the atolls in the Phoenix
Islands, and over the following 10 months a sea
surface temperature (SST) ‘hotspot’ developed and
remained over the central Pacific from June 2002 to
March 2003 (Alling et al. 2007). Data from in situ
temperature loggers showed the highest maximum
SST was recorded in November 2002, being about
0.5–1 �C warmer than the following two years. The
hotspot did not fully dissipate and remained for an
extended period of 21 Degree Heating Weeks (DHW).
Post-bleaching surveys in 2005 documented live coral
cover, which was significantly reduced to 12.1 %
overall in the Phoenix Islands (Obura and Mangubhai
2011a). This level of coral loss is consistent with other
506 Rev Fish Biol Fisheries (2014) 24:505–518
123

studies where bleaching has had substantial negative
impact on the abundance and composition of coral
communities (e.g. Vargas-Angel et al. 2011). Given
their remoteness and protection from local anthropo-
genic stressors, the reefs are considered relatively
pristine, with highly abundant fish populations (Obura
et al. 2011; Obura and Stone 2002), which provides an
opportunity to investigate the impacts of bleaching,
decoupled from other concurrent stressors.
The remarkable abundance of fishes across all
trophic groups that are largely overfished elsewhere
makes the Phoenix Islands an ideal study site
compared to more developed and/or exploited Central
Pacific reefs (Allen and Bailey 2011). The lack of local
anthropogenic impacts in the Phoenix Islands also
allows for examination of how natural topographic
features contribute to relative changes in benthic and
fish assemblages. Studies examining how coral cover
differs from site to site often note high between-site
variability, but it is often difficult to decouple the
influence of anthropogenic versus natural features of
each site. For example, remote and undeveloped
lagoonal islands may be differently sensitive to
thermal stress compared to non-lagoonal islands
(Obura and Mangubhai 2011), given their differences
in water quality and nutrient characteristics. However,
different fishes might be predicted to respond differ-
ently to thermal stress and resulting loss of coral
habitat, depending on their trophic mode and flexibil-
ity, mobility, lifespan, and other natural history
characteristics (e.g. Wilson et al. 2006; Pratchett
et al. 2008). The extreme remoteness of the archipel-
ago, coupled with the severity of this thermal event,
make the Phoenix Islands a relatively unique location
for exploring reef fish response to climate change
disturbance in the field.
In this study, we examined the short-term
(*3 year) impacts of severe thermal stress on fish
assemblages in the Phoenix Islands. We compared fish
family abundances pre- and post-bleaching by site,
and explored several hypotheses to explain why the
abundances of some families differed, including site
location, island type, and hard coral cover. We tested
the hypothesis that fish decline differed between
lagoonal atolls and non-lagoonal islands, and that fish
decline was correlated to hard coral cover decline. For
fish abundance data, the family level is the highest
taxonomic resolution available for 2002 and 2005.
Methods
Study site
The Phoenix Islands in the Republic of Kiribati are
comprised of eight islands (Nikumaroro, Kanton,
Orona, Enderbury, Manra, Rawaki, McKean and
Birnie) and two shallow submerged reefs (Carondelet
and Winslow), all located within the equatorial region
of the central Pacific Ocean (2.5–5�S, 174.8–170.1�W)
(Fig. 1). The three largest islands (Nikumaroro, Kan-
ton, and Orona) are atolls with lagoons that connect to
the sea. The remaining islands have no interior lagoon
that interacts with the sea (Fig. 1).
In 2002, a total of 451 fish species were recorded in
the Phoenix Islands, including 212 new records for the
island group, for a total of 516 total known species in
217 genera and 67 families (Allen and Bailey 2011). In
2002, nine permanent monitoring sites were estab-
lished around the Phoenix Islands in an effort to
standardize sampling locations over time (Fig. 1).
Eight of nine sites were chosen on the leeward sides of
islands and atolls, for both consistency and accessi-
bility. These sites were visited in 2002 (pre-bleach-
ing), and again in 2005 (post-bleaching).
Hard coral cover analysis
The reef platform of the Phoenix Islands is typically
\100 m wide and within a similar distance from the
shoreline. Full descriptions of each island are available
from Obura et al. (2011). Study sites were marked by
GPS position and navigated using distinct coastal and
underwater features. Within sites, coral communities
were surveyed by haphazardly-chosen sampling loca-
tions. Benthic cover was sampled using photo quadrats
collected at a single depth (12–15 m) on the forereef,
along the reef edge. High resolution digital images
(5–8 megapixels) were collected with a digital camera
held *60 cm above the substratum with the image
plane parallel to the surface. To analyze images, 25
points were chosen from an even 5 9 5 grid and coral
cover was determined at each point. Four images were
aggregated (100 points) as a single replicate (Obura
and Mangubhai 2011), reflecting a compromise
between the size of a sample unit (4 m2) and grain/
resolution of photoquadrats (25 pts per image). In
2002, five replicates per site were used, while in 2005,
Rev Fish Biol Fisheries (2014) 24:505–518 507
123

ten replicates were used per site. Hard coral cover
surveys were analyzed between years and sites with a
two-way analysis of variance (ANOVA). More
detailed analyses of benthic change can be found in
Obura and Mangubhai (2011). Both coral and fish
surveys were done at the same sites shown in Fig. 1.
Family-level fish surveys
At each site, 6–12 circular point counts were
conducted with a 7 m radius (area totaling
*150 m2 each) to measure fish abundance (as in
Wilson et al. 2010b). Data were gathered mainly at
the fish family level, with some notes subfamilies
(genus and species were not quantified in point
counts), by the same observer in both 2002 and
2005. Family-level resolution was chosen for long-
term monitoring given the needs of Kiribati at the
time, consistent with guidance under the Global
Coral Reef Monitoring Network (Salvat 2002;
English et al. 1997). Point counts were separated
by at least 15–20 m underwater. In each replicate,
the abundances of fishes in 13 key fish families were
Fig. 1 Map of the Kiribati
Phoenix Islands showing the
location of the permanent
monitoring study sites
examined in this study. The
small black square
represents the boundary of
the Phoenix Islands
Protected Area, created in
2006. Black circles on each
island map denote sampling
sites. Note the changing
scale bars on each island
graph. Maps created by
K. Lagueux
508 Rev Fish Biol Fisheries (2014) 24:505–518
123
recorded—Acanthuridae, Balistidae, Carangidae, Chae-
todontidae, serranid subfamily Epinephelinae, Haemu-
lidae, Labridae, Lethrinidae, Lutjanidae, Pomacanthi-
dae, Scaridae and Sphyraenidae; and elasmobranchs.
One family was counted at a time, focusing on the
faster moving and shyer species first (e.g. Carangidae,
Scaridae), and then counting the more sedentary
fishes (e.g. serranid subfamily Epinephelinae) that
tended to hide in the reef matrix. Other serranids
were not included in surveys. Faster swimming,
highly mobile fish that traversed each transect were
also included. The time taken to complete a count
varied depending on the number of fish present, but
ranged from 3 to 10 min. To reduce positive skew
due to either rare occurrences or dense schools of
mobile and transient fishes, abundances were
log(x ? 1) transformed. In order to estimate the
within-site variation, we considered point counts as
the statistical unit of replication.
A two-way ANOVA with site as a random factor
was used to examine year and site effects within fish
families on log(x ? 1) transformed data. Changes
across all fish families across sites were also explored
using a two-way ANOVA, looking at the mean
abundance of all fishes per site from 2002 and 2005.
We conducted post hoc power analyses assuming an
effect size of 0.8, an alpha of 0.05, and inputted the
calculated standard deviation for each taxonomic
group. For all between-year comparisons, there was
enough power to detect a difference ([0.8) given our
sample size.
Exploring hypotheses for changes in fish
abundance
To explore hypotheses that might explain temporal
changes in fish abundance, we examined the effects of
island type (lagoonal atoll versus non-lagoonal island)
and fish family abundance for the 3 families that
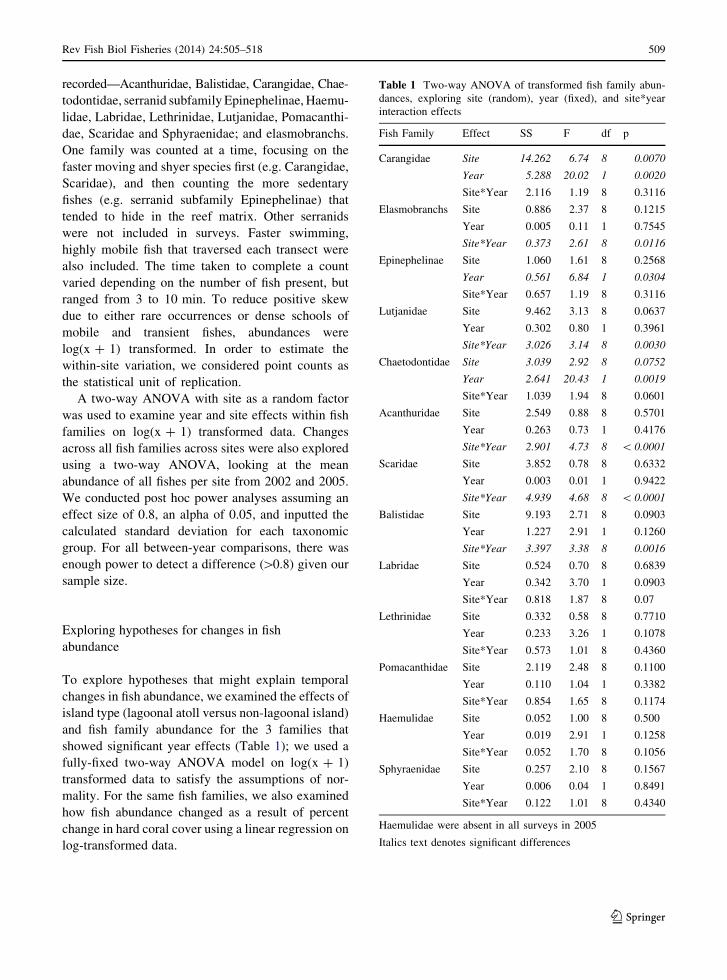
showed significant year effects (Table 1); we used a
fully-fixed two-way ANOVA model on log(x ? 1)
transformed data to satisfy the assumptions of nor-
mality. For the same fish families, we also examined
how fish abundance changed as a result of percent
change in hard coral cover using a linear regression on
log-transformed data.
Table 1 Two-way ANOVA of transformed fish family abun-
dances, exploring site (random), year (fixed), and site*year
interaction effects
Fish Family Effect SS F df p
Carangidae Site 14.262 6.74 8 0.0070
Year 5.288 20.02 1 0.0020
Site*Year 2.116 1.19 8 0.3116
Elasmobranchs Site 0.886 2.37 8 0.1215
Year 0.005 0.11 1 0.7545
Site*Year 0.373 2.61 8 0.0116
Epinephelinae Site 1.060 1.61 8 0.2568
Year 0.561 6.84 1 0.0304
Site*Year 0.657 1.19 8 0.3116
Lutjanidae Site 9.462 3.13 8 0.0637
Year 0.302 0.80 1 0.3961
Site*Year 3.026 3.14 8 0.0030
Chaetodontidae Site 3.039 2.92 8 0.0752
Year 2.641 20.43 1 0.0019
Site*Year 1.039 1.94 8 0.0601
Acanthuridae Site 2.549 0.88 8 0.5701
Year 0.263 0.73 1 0.4176
Site*Year 2.901 4.73 8 \ 0.0001
Scaridae Site 3.852 0.78 8 0.6332
Year 0.003 0.01 1 0.9422
Site*Year 4.939 4.68 8 \ 0.0001
Balistidae Site 9.193 2.71 8 0.0903
Year 1.227 2.91 1 0.1260
Site*Year 3.397 3.38 8 0.0016
Labridae Site 0.524 0.70 8 0.6839
Year 0.342 3.70 1 0.0903
Site*Year 0.818 1.87 8 0.07
Lethrinidae Site 0.332 0.58 8 0.7710
Year 0.233 3.26 1 0.1078
Site*Year 0.573 1.01 8 0.4360
Pomacanthidae Site 2.119 2.48 8 0.1100
Year 0.110 1.04 1 0.3382
Site*Year 0.854 1.65 8 0.1174
Haemulidae Site 0.052 1.00 8 0.500
Year 0.019 2.91 1 0.1258
Site*Year 0.052 1.70 8 0.1056
Sphyraenidae Site 0.257 2.10 8 0.1567
Year 0.006 0.04 1 0.8491
Site*Year 0.122 1.01 8 0.4340
Haemulidae were absent in all surveys in 2005
Italics text denotes significant differences
Rev Fish Biol Fisheries (2014) 24:505–518 509
123

510 Rev Fish Biol Fisheries (2014) 24:505–518
123

Results
Benthic response to thermal stress
In 2002, the highest mean coral cover was found at
Weird Eddie (83 %), Lone Palm (77 %), and Satellite
Beach (71 %), while Algae Corner had the lowest mean
coral cover (10 %), as reported in Obura and Mangubhai
(2011). Following substantial coral mortality in 2005,
the highest mean coral cover was recorded at Lone Palm
(43 %) and Deepwater (38 %), representing almost half
of their 2002 means. The greatest coral cover losses
were recorded on the three atolls: on Kanton, Weird
Eddie and Satellite Beach lost 97 and 87 % cover,
respectively; on Nikumaroro cover declined by 80 % at
Amelia’s Lost Causeway and 97 % on Windward Wing,
and on Orona, Dolphin Ledge lost 92 % live coral cover.
There was a significant year effect (F = 444.9,
df = 1,1, p \ 0.001), site effect (F = 20.07, df = 8,8,
p \ 0.001) and year*site interaction (F = 11.47,
df = 8,8, p \ 0.001) comparing hard coral cover over
time (two-way ANOVA, F = 44.06, df = 17, 149,
p \ 0.001), which explained most of the variation in the
data (R2 = 0.82). At all sample sites, there was a
significant decrease in hard coral cover (Fig. 2 insets).
Differences in fish family abundance between sites
We found significant site effects for Carangidae, and
Chaetodontidae (Table 1) and significant year effects for
Carangidae, Chaetodontidae, and Epinephelinae
(Table 1). Five families showed significant site by year
interactions that did not correspond with significant site
or year effects alone. The most abundant fish family in
both years was Acanthuridae; but the average abundance
across all sites did not change pre- and post- bleaching
(t = 0.17, df = 131, p = 0.86); in 2002 abundance
averaged 23.7 fish per 154 m2; in 2005, 24.5 fish per
154 m2 for each point count. A two-way ANOVA
revealed a significant year effect, whereby the overall
abundance of all fish combined significantly declined
from 2002 to 2005 (F = 8.765, df = 1,1 p = 0.0038).
However, there was also significant site-to-site variabil-
ity (Fig. 3; F = 3.484, df = 8,8, p = 0.0014) and a
significant year by site interaction (F = 3.346, df = 8,8,
p = 0.002), since not all sites showed a decline in mean
abundance of all fish combined. This difference was
maintained across sites (Fig. 3; paired t = 3.41, df = 8,
p = 0.0093), with all sites declining in fish abundance
between 2002 and 2005 except for Windward Wing.
Exploring possible causes for changes in fish
abundance using targeted fish families
To test hypotheses that might explain the differences
in fish abundances over time, we explored the effect of
island type (atoll versus island), and change in percent
coral cover in the subset of three fish families that
changed over time regardless of site (Fig. 4; Table 2).
For Epinephelinae and all fish, island type did
not explain the differences between years as there
were no island type or island type * year interactions
(Table 2A). However, there was a significant island
type effect and/or a significant island type*year
interaction for Carangidae and Chaetodontidae
(Table 2A). Correspondingly, abundances of Chae-
todontidae varied significantly with declines in coral
cover (Table 2B; Fig. 4). Although the decline of live
coral cover was observed at all sites (Fig. 2, insets), no
other significant relationships between fish family
abundance and coral decline were found.
Fig. 3 Mean abundance of all fish families shown by site,
between years from 2002 and 2005
Fig. 2 Mean abundance of fish families (±standard error) per
transect (154 m2) organized by permanent monitoring sites in
the Phoenix Islands in 2002 (black bars) and 2005 (white bars).
O Orona Atoll, N Nikumaroro Island, M Manra Atoll, K Kanton
Atoll, E Enderbury Island, R Rawaki Island. Exposure is
denoted as L or W for Leeward or Windward respectively,
Habitat is denoted as A or I for atoll or island. Results from
Haemulidae and Sphyraenidae families were not presented
because of their low density. Upper right inset depicts mean
(±standard error) percent hard coral cover for 2002 in black
(n = 10 transects), and for 2005 in white (n = 5 transects)
b
Rev Fish Biol Fisheries (2014) 24:505–518 511
123

Discussion
This paper aims to describe family-level changes in fish
abundance over time from a relatively isolated and near-
pristine reef; it is among a growing number of studies
that use isolated reefs as benchmarks for benthic
degradation and recovery, and resulting changes in
community structure (e.g. Halford and Caley 2009;
Chong-Seng et al. 2012; Wernberg et al. 2012a, b;
Gilmour et al. 2013). This paper contributes several
Fig. 4 Exploration of factors (habitat and coral cover) that may
have influenced post-bleaching fish abundance (per transect; or
per 154 m2) for families that experienced a significant change
between 2002 (black bars) and 2005 (white bars), and for all fish
(representing 13 families)
512 Rev Fish Biol Fisheries (2014) 24:505–518
123

confirming insights into how fish families on near-
pristine reefs change in the short-term, following a
severe coral bleaching event. Firstly, there is dramatic
variation in site-level response. Because the Phoenix
Islands are isolated from local anthropogenic impacts,
site-to-site variation is likely indicative of naturally-
varying factors such as nutrient availability, benthic
structure, oceanographic features, recruitment dynam-
ics and/or population connectivity. Despite the baseline
differences pre-bleaching (in 2002), all sites experi-
enced declines in coral cover in 2005, suggesting that
despite this variation, no sites were immune to severe
thermal stress. In contrast, mean total fish abundance
varied from site-to-site, and significant site-specific
family-level changes in fish community composition
were observed. The documentation of these site- and
family-specific differences in the Phoenix Islands is the
main contribution of this paper.
Reef response over time
Consistent with a large and global body of literature
dedicated to the consequences of increased ocean
temperatures (e.g. Hoegh-Guldberg et al. 2007), corals
of the Phoenix Islands bleached and experienced
subsequent mortality following extreme thermal stress
(Fig. 2 insets) (Alling et al. 2007; Obura et al. 2011;
Obura and Mangubhai 2011). It is important to note,
however, that this thermal stress was among the largest
reported for any reef worldwide, at any time, peaking at
21 degree heating weeks (Obura and Mangubhai 2011).
Extreme and discrete thermal stress events are increas-
ing in frequency and magnitude (Kerr 2011), and have
been shown to cause ecological changes such as
diversity loss via shifting species distributions (Smale
and Wernberg 2013). However, even with such thermal
severity, we observed site-to-site variation in coral
cover decline (Fig. 2 insets; see also Table 3 and Fig. 3
in Obura and Mangubhai 2011), which is consistent
with previous studies indicating site- or species-level
variation in response to severe bleaching due to
differences in reef habitat (Grimsditch et al. 2010),
species resilience (Pratchett et al. 2010), and/or
repeated thermal exposure (Williams et al. 2010). Such
variation suggests that while extreme stress events can
cause ecological change, such change is not necessarily
Table 2 Exploring hypotheses for changes in fish family abundance between 2002 and 2005
Fish family A. Habitat (Island vs. Atoll) B. Coral Cover (% change)
Effect SS F df p SS F df P
Carangidae Habitat 1.514 4.67 1 0.033
Year 4.346 13.4 1 0.0004 Model 0.003 5.19 1 0.072
Habitat*Year 0.082 0.25 1 0.616
Error 41.85 Error 0.006 5
Chaetodontidae Habitat 0.238 2.79 1 0.097
Year 0.790 9.27 1 0.003 Model 1.451 6.45 1 0.039
Habitat*Year 0.632 7.42 1 0.007
Error 16.41 Error 1.576 5
Epinephelinae Habitat 0.0001 0.003 1 0.959
Year 0.208 2.79 1 0.097 Model 0.046 0.379 1 0.379
Habitat*Year 0.024 0.33 1 0.568
Error 9.62 Error 0.609 5
All Fish Habitat 0.343 1.44 1 0.230
Year 0.773 3.25 1 0.072 Model 0.001 0.003 1 0.956
Habitat*Year 0.858 3.61 1 0.058
Error 410.3 Error 0.872 5
(A) Two-way ANOVA of log-transformed fish abundances exploring habitat (island versus atoll) and year effects for each family
identified as having a significant decrease over time. (B) Linear regression of log-transformed changes in fish abundance and coral
cover (% change over time)
For both (A) and (B), all fish includes all fish families surveyed
Rev Fish Biol Fisheries (2014) 24:505–518 513
123

uniform and is highly influenced by interacting local
factors.
In this study, we found that overall, total fish
abundance and live coral cover both declined from
2002 to 2005 and family- and site- specific changes in
the fish community over time were observed. Similar to
other bleaching events and non-structural reef distur-
bances (e.g. Pratchett et al. 2010), the 2002 to 2003
bleaching event in the Kiribati Phoenix Islands caused
massive tissue mortality but left coral skeletons largely
intact. Thus, at least for the short-term, topographic
complexity was not immediately compromised, which
may have maintained higher organismal diversity and
abundance compared to disturbances where topographic
complexity and heterogeneity were immediately
reduced (Sano et al. 1987; Graham et al. 2006). Along
these lines, we found that many families of fish did not
significantly change in abundance despite the severe
thermal event and corresponding coral loss (Fig. 2;
Table 1) Over longer periods of time, coral mortality
leaves the reef vulnerable to erosion, facilitating the loss
of habitat structure (Sheppard et al. 2002; Graham et al.
2006), which may be a driver of reef fish decline over the
longer term (Wilson et al. 2006; Garpe et al. 2006).
Changes in fish family abundance over time
We examined 13 fish families across 9 sites in 2002
and 2005, and found marked variability in abundance
across all. Three of the fish families declined, regard-
less of site (Carangidae, Chaetodontidae, and serranid
subfamily Epinephelinae). Despite major coral mor-
tality, Lethrinidae, Labridae, Sphyrinidae, and Pom-
acanthidae did not differ in abundance across sites or
years. Such consistent abundances may be tied to their
natural history; for example, species within Lethrin-
idae often prefer sandy and/or rubble habitats and are
often omnivorous, displaying trophic flexibility that
may allow stable abundances even in variable envi-
ronments. The most abundant fish family pre- and
post-bleaching was Acanthuridae, of which many
species are widely regarded as important herbivores
that contribute to overall reef resilience (e.g. Burkepile
and Hay 2008). Our findings are consistent with a
previous study that also found little/no change in
relative Acanthurid abundance over time (Halford and
Caley 2009). We found a decline in Chaetodontidae
over time, which is consistent with post-bleaching
decline observed at other locations (Bozec et al. 2005;
Wilson et al. 2008; Halford and Caley 2009; Pratchett
et al. 2011).
Examining hypotheses to explain differences
in fish family abundance
We explored two hypotheses that might explain the
significant declines of Carangidae, Chaetodontidae, and
subfamily Epinephelinae abundance post-bleaching:
the impact of island type and/or changes in coral cover.
For Epinephelinae, neither of these hypotheses ade-
quately explained the observed changes in abundance at
a family level. These fish families may have a more
nuanced response to bleaching than has previously been
appreciated, perhaps related to other habitat character-
istics, or their trophic, species-specific, reproductive, or
behavioral characteristics—all of which could be
examined with finer taxonomic resolution.
Changes in the abundance of fishes in the family
Chaetodontidae over time were most dramatic on atolls,
and were also associated with changes in live coral
cover. Atolls may have a significant impact on coral
cover, since they have been shown to be more sensitive
to thermal stress and bleaching severity as compared to
non-lagoonal islands (Obura and Mangubhai 2011).
Similarly, leeward sites near lagoonal waters also
experienced high mortality, especially on Kanton and
Nikumaroro. The highest mortality rates occur closest
to the mouth and decrease with distance, suggesting that
these atoll lagoons influence water quality and nutrient
characteristics, which may have downstream impacts
on coral resilience, recovery, and growth. Pre-bleach-
ing, there was a high density of corals in shallow
lagoons, particularly of large table Acropora colonies.
Anecdotally, butterflyfishes commonly found in
2002—including the obligate corallivore Chaetodon
trifascialis—were only rarely sighted in 2005 post-
bleaching. The combination of habitat and nutritional
specificity likely explains our finding that Chaetodonti-
dae abundance differed by island type.
In addition to location considerations, changes in
Chaetodontid abundance over time were also associ-
ated with declines in live coral cover. Chaetodontidae
feed primarily on living tissue from scleractinian and
alcyonacean corals (Anderson et al. 1981); conse-
quently, their abundance and performance are usually
tied to live coral cover (Findley and Findley 1985;
Williams 1986; Pratchett et al. 2004; Bozec et al.
2005). Pre-bleaching, we found that Chaetodontidae
514 Rev Fish Biol Fisheries (2014) 24:505–518
123

abundance was high, likely reflective of both coral
community composition and/or coral nutritional value.
Post-bleaching, we found a decline in Chaetodontidae
abundance across sites, consistent with previous studies
that found obligate coral-feeding fish declined substan-
tially following a severe bleaching event (Graham et al.
2006) or a severe decrease in live coral cover (Bozec
et al. 2005). Chaetodontids comprise the highest
number of species of obligate corallivores for any fish
family (Cole et al. 2008; Rotjan and Lewis 2008), and
our findings are consistent with a recent study that
flagged obligate corallivores as being especially sensi-
tive to climate vulnerability, displaying the highest risk
of local extinction compared to other functional groups
(Graham et al. 2011).
In contrast to Chaetodontidae, changes in the
abundance of fishes in the family Carangidae over
time were most dramatic on islands, and were not
associated with changes in live coral cover. Many
species in the family Carangidae are known to be
highly mobile, and are not known to be among the fish
families that are tightly coral-associated (Pratchett
et al. 2008). Variation in fish counts has been
acknowledged in the literature (e.g. McClanahan
et al. 2007), and such variation is likely to be
augmented in highly mobile species. Our data also
show high site-to-site variation in Carangidae abun-
dance (Fig. 2), suggesting that there may be more
nuanced site-level drivers of Carangidae abundance.
Other potential drivers of changing fish abundance
include nutrient availability, other aspects of benthic
structure, oceanographic features, recruitment dynam-
ics and/or population connectivity. We found a variety
of interactions between site and year for other fish
families that could be explained by some of these
alternative drivers. Furthermore, the significant time
effect that we found for Epinephelinae is indicative of
other factors at play, given that neither island type nor
changes in hard coral cover explained the observed
differences over time. Given the opportunistic nature
of this dataset, a comprehensive analysis of each factor
was not possible, but should be a priority for the future.
Taxonomic approach
We focused primarily on fish abundance and diversity
organized by taxonomic family, as the initial purpose
for monitoring was to establish long term monitoring
stations for national purposes consistent with guidance
under the Global Coral Reef Monitoring Network
(Salvat 2002; English et al. 1997). This ‘‘higher
taxon’’ approach is considered sufficient for some
systems such as birds, intertidal molluscs, plants, and
insects (e.g. Gaston and Williams 1993; Kallimanis
et al. 2012; Gladstone and Alexander 2005). We found
this approach to be adequate in determining broad
patterns in response to bleaching in a logistically-
constrained location, and our results are similar to
other recent studies assessing the influence of benthic
condition on reef fish assemblages (reviewed by
Pratchett et al. 2011). However, there are clear
limitations to family level data (Green and Bellwood
2009; Obura and Grimsditch 2009). For example, it is
known that high species diversity and abundance
within functional groups helps to reduce the impact of
disturbance at the community level (Schmitz et al.
2000). The species diversity within family is known
for the Phoenix Islands (Allen and Bailey 2011), and
assessing the ecological, species-specific contribu-
tions to reef resilience and/or recovery is a priority for
future studies (Wilson et al. 2010a).
Abundance, diversity, and biomass changes
in response to bleaching
Assessing changes in fish diversity or biomass in
response to disturbance is a growing area of research
and is far from straightforward. For example, Wilson
et al. (2009) found that reef fish species richness was
maintained in 9 out of 10 sites on the Great Barrier
Reef, despite frequent disturbances and subsequent
coral declines of 46–96 %. Thus, patchy and/or severe
changes to the benthos do not necessitate immediate
losses in fish species richness (Wilson et al. 2009) or
uniform family-level loss of abundance (this study).
Despite a non-uniform response, there is an abundant
literature on how habitat degradation drives fish
community changes (e.g. Feary et al. 2009; Graham
et al. 2008; Wilson et al. 2008; Pratchett et al. 2011;
Chong-Seng et al. 2012) or local human influence (e.g.
Wilson et al. 2006; DeMartini et al. 2008). Taken
together, it seems that fishing pressure, coastal devel-
opment, and/or habitat loss are all important contrib-
utors to overall reef fish decline; the major
contribution of this study is to demonstrate how, at
least in the short-term, site-to-site variation in habitat
degradation can impact fish family abundance
changes, even in the absence of other pressures.
Rev Fish Biol Fisheries (2014) 24:505–518 515
123

Long-term impacts
While this study examined the response of fish families
over the short-term (3-years) following severe bleach-
ing in the absence of local human influence, long-term
impacts remain to be investigated. There are several
delayed ecosystem responses that may help to promote
or prevent reef recovery, and the fish community
composition may change as a result (Graham et al.
2008). For example, coral-associated juvenile fish may
suffer consequences from bleaching with a resulting lag
in adult fish decline; previous studies have shown that
coral mortality can decrease settlement rates for up to
65 % of reef fishes (Jones et al. 2004). Similarly,
McCormick et al. (2010) found that 10 times as many
fish settled to healthy coral compared to sub-lethally
bleached coral. Thus, following coral mortality, long-
term recruitment may be hindered even though few
immediate effects are observed in the adult population.
Long-term recovery may also be hindered by repeated
or ongoing disturbances, with resulting consequences
for the community of coral-dependent fishes (Berumen
and Pratchett 2006).
The Phoenix Islands present a unique opportunity
to assess global disturbance events in the absence of
local human impacts. During the time of this study
(2002–2005), the Phoenix Islands were not yet a
marine protected area and species-specific monitoring
was not yet implemented. However, the reefs were
considered a ‘‘de facto’’ marine protected area due to
its extreme isolation. We are reasonably confident that
any change in Phoenix Islands reef fish assemblages
between 2002 and 2005 was likely due directly to
severe thermal stress, or indirectly due to the cascad-
ing impact of thermal stress on related habitat.
However, the possibility remains that changes in fish
abundances (or lack of) were completely independent,
and instead reflect natural variation. The isolation and
protection of the Phoenix Islands should hopefully
shed light on the natural variation of fish families
during intervals without major thermal stress.
The establishment of the Phoenix Islands Protected
Area (PIPA) so soon post-bleaching will ensure that
the future trajectory of PIPA reefs should be unim-
paired by fishing-induced loss of fish diversity,
abundance or biomass. However, continued thermal
stress and associated habitat loss will continue to be
important drivers of PIPA reef status. A 2010 bleach-
ing event in the U.S. Phoenix Islands (Vargas-Angel
et al. 2011) and corresponding high temperature event
in the Kiribati Phoenix Islands (Mangubhai et al.
2012) will likely impact the recovery trajectory of the
Phoenix Islands, but will also heighten the importance
of this region for the opportunity to examine long-term
reef response to repeated disturbance in the absence of
local human stressors.
Acknowledgments The authors would like to thank Stuart
Sandin and Les Kaufman for their thoughtful comments on this
manuscript. We are grateful to Kerry Lagueux for producing the
maps depicted in Fig. 1, and to Tania Lemos Eskin for drawing
the fish sketches used in Table 2 and Fig. 3. This study is part of
a larger effort examining reef recovery and resilience processes
on isolated atoll reefs in the central Pacific; this effort has been
funded and supported by the New England Aquarium, the
National Geographic Society, Conservation International
(Global Conservation Fund), the Government of Kiribati, the
Akiko Shiraki Dynner Fund for Ocean Exploration and
Conservation, and the Nai’a (Rob Barrel and Cat Holloway).
Findings from this manuscript were presented at the World
Fisheries Congress in 2012.
References
Allen GR (2007) Conservation hotspots of biodiversity and
endemism for Indo-Pacific coral reef fishes. Aquatic Con-
serv Mar Fresh Eco 18:541–556
Allen GR, Bailey S (2011) Reef fishes of the Phoenix Islands,
Central Pacific Ocean. Atoll Res Bull 589:83–118
Alling A, Doherty O, Logan H, Feldman L, Dustan P (2007)
Catastrophic coral mortality in the remote central Pacific
Ocean: Kirabati (sic) Phoenix Islands. Atoll Res Bull 551:
1–18
Anderson GRV, Ehrlich AH, Ehrlich PR, Roughgarden JD,
Russell BC, Talbot FH (1981) Community Struct Coral
Reef Fishes. Am Nat 117(4):476–495
Balmford A, Bennun L, Ten Brink B (2005) The convention on
biological diversity’s 2010 target. Science 307:212–213
Berumen ML, Pratchett MS (2006) Recovery without resilience:
persistent disturbance and long-term sifts in the structure of
fish and coral communities at tiahura Reef, Moorea. Coral
Reefs 25:647–653
Bonin MC, Munday PL, McCormick MI, Srinivasan M, Jones GP
(2009) Coral-dwelling fishes resistant to bleaching but not to
mortality of host corals. Mar Ecol Prog Ser 394:215–222
Bozec YM, Doledec S, Kulbicki M (2005) An analysis of fish-
habitat associations on disturbed coral reefs: chaetodontic
fishes in New Caledonia. J Fish Biol 66:966–982
Burkepile DE, Hay ME (2008) Herbivore species richness
and feeding complementarity affect community structure
and function on a coral reef. Proc Natl Acad Sci 105:
16201–16206
Chong-Seng KM, Mannering TD, Pratchett MS, Bellwood DR,
Graham NAJ (2012) The influence of coral reef benthic
condition on associated fish assemblages. PLoS ONE
7:e42167
516 Rev Fish Biol Fisheries (2014) 24:505–518
123

Christensen N, Bartuska A, Brown J, Carpenter S, Da C, Francis
R, Franklin JF, MacMahon JA, Noss RF, Parsons DJ,
Peterson CH, Turner MG, Woodmansee RG (1996) The
report of the ecological society of America committee on
the scientific basis for ecosystem management. Ecol App
9:1266–1277
Cole AJ, Pratchett MS, Jones GP (2008) Diversity and func-
tional importance of coral-feeding fishes on tropical coral
reefs. Fish Fish 9(3):286–307
DeMartini EE, Friedlander AM, Sandin SA, Sala E (2008)
Differences in fish-assemblage structure between fished
and unfished atolls in the northern Line Islands, central
Pacific. Mar Ecol Prog Ser 365:199–215
English C, Wilkinson C, Baker V (1997) Survey manual for
tropical marine resources 2nd edn. Australian Institute of
Marine Sciences ISBN 0 642 2594 4
Feary DA, McCormick MI, Jones GP (2009) Growth of reef
fishes in response to live coral cover. J Exp Mar Biol Ecol.
doi:10.1016/j.jembe.2009.03.002
Findley JS, Findley MT (1985) A search for pattern in butterf-
lyfish communities. Am Nat 126:800–816
Garpe KC, Yahya SAS, Lindahl U, Ohman MC (2006) Long-
term effects of the 1998 coral bleaching even on reef fish
assemblages. Mar Ecol Prog Ser 315:237–247
Gaston KJ, Williams PH (1993) Mapping the world’s species -
the higher taxon approach. Biodivers Lett 1:2–8
Gilmour JP, Smith LD, Heyward AJ, Baird AH, Pratchett MS
(2013) Recovery of an isolated coral reef system following
severe disturbance. Science 340:69–70
Gladstone W, Alexander T (2005) A test of the higher-taxon
approach in the identification of candidate sites for marine
reserves. Biodivers Conserv 14:3151–3168
Graham NAJ, Wilson SK, Jennings S, Polunin NVC, Bijoux JP,
Robinson J (2006) Dynamic fragility of oceanic coral reef
ecosystems. Proc Nat Acad Sci 103:8425–8429
Graham NAJ, McClanahan TR, MacNeil MA, Wilson SK,
Polunin NVC et al (2008) Climate warming, marine pro-
tected areas and the ocean-scale integrity of coral reef
ecosystems. PLoS ONE 3(8):e3039. doi:10.1371/journal.
pone.0003039
Graham NAJ, Chabanet P, Evans RD, Jennins S, Letourner Y,
MacNeil MA, McClanahan TR, Ohman MC, Polunin
NVC, Wilson SK (2011) Extinction vulnerability of coral
reef fishes. Ecol Lett 14:341–348
Green AL, Bellwood DR (2009) Monitoring functional groups
of herbivorous reef fishes as indicators of coral reef resil-
ience – A practical guide for coral reef managers in the
Asia Pacific region. IUCN working group on climate
change and coral reefs, IUCN, Gland, Switzerland, 70 pp
Grimsditch G, Mwaura JM, Kilonzo J, Amiyo N (2010) The
effects of habitat on coral bleaching responses in Kenya.
Ambio 39:195–304
Halford AR, Caley MJ (2009) Towards an understanding of
resilience in isolated coral reefs. Glob Change Biol 15:
3031–3045
Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS,
Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ,
Caldeira K, Knowlton N, Eakin CM, Iglesias-Prieto R,
Muthiga NA, Bradbury RH, Dubi A, Hatziolos ME (2007)
Coral reefs under rapid climate change and ocean acidifi-
cation. Science 318(5857):1737–1742
Jackson JBC (2001) What was natural in the coastal oceans?
Proc Nat Acad Sci 98(10):5411–5418
Jones GP, Syms C (1998) Disturbance, habitat structure and the
ecology of fishes on coral reefs. Aust J Ecol 23:287–297
Jones GP, McCormick MI, Srinivasan M, Eagle JV (2004) Coral
decline threatens fish biodiversity in marine reserves. Proc
Nat Acad Sci 101:8251–8253
Kallimanis AS, Mazaris AD, Tsakanikas D, Dimopoulos P, Pantis
JD, Sgardelis SP (2012) Efficient biodiversity monitoring:
which taxonomic level to study? Ecol Indic 15:100–104
Kerr RA (2011) Humans are driving extreme weather; time to
prepare. Science 334:1040
Lieske E, Myers R (1994) Coral reef fishes: Indo-Pacific and
Caribbean. Harper Collins, London
Lindahl U, Ohman MC, Schelten CK (2001) The 1997/1998
mass mortality of corals: effects on fish communities on a
Tanzanian coral reef. Mar Poll Bull 42(2):127–131
Mangubhai S, Rotjan RD, Obura DO (2012) Phoenix Islands
Protected Area 2012 Expedition Report. New England
Aquarium, Boston, MA, USA, 44 p
McClanahan TR, Graham NAJ, Maina J, Chabanet P, Brugge-
mann JH, Polunin NVC (2007) Influence of instantaneous
variation on estimates of coral reef fish populations and
communities. Mar Ecol Prog Ser 340:221–234
McCormick MI, Moore JAY, Munday PL (2010) Influence of
habitat degradation on fish replenishment. Coral Reefs
29:537–546
Munday PL, Jones GP, Pratchett MS, Williams AJ (2008) Cli-
mate change and the future for coral reef fishes. Fish Fish
9:261–285
Obura DO, Grimsditch G (2009) Resilience assessment of coral
reefs – Assessment protocol for coral reefs, focusing on
coral bleaching and thermal stress. IUCN working group
on climate change and coral reefs, IUCN, Gland, Swit-
zerland, 70 pp
Obura DO, Mangubhai S (2011) Coral mortality associated with
thermal fluctuations in the Phoenix Islands, 2002-2005.
Coral Reefs 30:607–619
Obura DO, Stone GS (2002) Phoenix Islands: Summary of
marine and terrestrial assessments conducted in the
Republic of Kiribati: June 5-July 10, 2002. New England
Aquarium, Boston
Obura D, Stone G, Mangubhai S, Bailey S, Yoshinaga A, Hol-
loway C, Barrel R (2011) Baseline marine biological sur-
veys of the Phoenix Islands, July 2000. Atoll Res Bull
589:1–62
Pandolfi JM, Bradbury RH, Sala E, Hughes TP, Bjorndal KA,
Cooke RG, McArdle D, McClenachan L, Newman M,
Paredes G, Warner RR, Jackson JBC (2003) Global tra-
jectories of the long-term decline of coral reef ecosystems.
Science 301:955–958
Petraitis PS, Latham RE, Niesenbaum RA (1989) The mainte-
nance of species diversity by disturbance. Q Rev Biol
64:393–418
Pickett STA, White PS (eds) (1986) The ecology of natural
disturbance and patch dynamics. Academic Press, Ontario
Pratchett MS, Wilson SK, Berumen ML, McCormick MI (2004)
Sublethal effects of coral bleaching on an obligate coral
feeding butterflyfish. Coral Reefs 23(3):352–356
Pratchett MS, Munday PL, Wilson SK, Graham NAJ, Cinner JE,
Bellwood DR, Jones GP, Polunin NVC, McClanahan TR
Rev Fish Biol Fisheries (2014) 24:505–518 517
123

(2008) Effects of climate-induced coral bleaching on coral-
reef fishes: ecological and economic consequences. Oce-
anogr Mar Biol Ann Rev 46:251–296
Pratchett MS, Trapon N, Berumen ML, Chong-Seng K (2010)
Recent disturbances augment community shifts in coral
assemblages in Moorea, French Polynesia. Coral Reefs
online first. doi:10.1007/s00338-00010-00678-00332
Pratchett MS, Hoey AS, Wilson SK, Messmer V, Graham NAJ
(2011) Changes in the biodiversity and functioning of reef
fish assemblages following coral bleaching and coral loss.
Diversity 3:424–452
Rotjan RD, Lewis SM (2008) Impact of coral predators on
tropical reefs. Mar Ecol Prog Ser 367:73–91
Salvat B (2002) Status of southeast and central Pacific coral
reefs ‘Polynesia Mana Node’: Cook Islands, French Poly-
nesia, Kiribati, Niue, Tokelau, Tonga, Wallis and Futuna.
In: Wilkinson C (ed) Status of coral reefs of the world:
2002. Australian Institute of marine Science, Townsville,
pp 203–216
Sandin SA, Smith JE, deMartini EE, Dinsdale EA, Donner SD,
Friedlander AM, Konotchick T, Malay M, Maragos JE,
Obura D, Pantos O, Paulay G, Ritchie M, Rohwer F,
Schroeder RE, Walsh S, Jackson JBC, Knowlton N, Sala E
(2008) Baslines and degredation of coral reefs in the
Northern Line Islands. PLoS ONE 3(2):w1548
Sano M, Shimizu M, Nose Y (1987) Long-term effects of
destruction of hermatypic corals by Acanthaster planci
infestation on reef fish communities at Iriomote Island,
Japan. Mar Ecol Prog Ser 37:191–199
Schmitz OJ, Hamback PA, Beckerman AP (2000) Trophic
cascades in terrestrial systems: a review of the effects of
carnivore removals on plants. Am Nat 155:141–153
Sheppard CRC, Spalding MD, Bradshaw C, Wilson SK (2002)
Erosion vs. recovery of coral reefs after 1998 El Nino:
Chagos reefs. Ambio 31:40–48
Smale DA, Wernberg T (2013) Extreme climatic event drives
range contraction of a habitat-forming species. Proc Roy
Soc B 280:20122829
Sousa WP (1984) The role of disturbance in natural communi-
ties. Ann Rev Ecol Sys 5:353–391
Thomas CD, Cameron A, Green RE (2004) Extinction risk from
climate change. Nature 416:389–395
Vargas-Angel B, Looney EE, Vetter OJ, Coccagna EF (2011)
Severe, widespread El Nino associated coral bleaching in
the US Phoenix Islands. Bull Mar Sci 87:623–638
Wernberg T, Smale DA, Thomsen MS (2012a) A decade of cli-
mate change experiments on marine organisms: procedures,
patterns, and problems. Glob Change Biol 18:1491–1498
Wernberg T, Smale DA, Tuya F, Thomsen MS, Langlois TJ,
deBettignes T, Bennett S, Rousseaux CS (2012b) An
extreme climatic event alters marine ecosystem structure in
a global biodiversity hotspot. Nat Clim Chang 3:78–82
Williams DM (1986) Temporal variation in the structure of reef
slope fish communities (central great barrier reef): short-
term effects of Acanthaster planci infestation. Mar Ecol
Prog Ser 28:157–164
Williams GJ, Knapp IS, Maragos JE, Davy SK (2010) Modeling
patterns of coral bleaching at a remote Central Pacific atoll.
Mar Poll Bull 60:1467–1476
Wilson SK, Graham NAJ, Pratchett MS, Jones GP, Polunin
NVC (2006) Multiple disturbances and the global degre-
dation of coral reefs: are reef fishes at risk or resilient? Glob
Change Biol 12:2220–2234
Wilson SK, Burgess S, Cheal A, Emslie M, Fisher R, Miller I,
Polunin NVC, Sweatman HPA (2008) Habitat utilisation
by coral reef fish: implications for specialists vs. generalists
in a changing environment. J Anim Ecol 77:220–228
Wilson SK, Dolman AM, Cheal AJ, Emslie MJ, Pratchett MS,
Sweatman HPA (2009) Maintenance of fish diversity on
disturbed coral reefs. Coral Reefs 28:3–14
Wilson SK, Adjeroud M, Bellwood DR, Berument ML, Booth
D, Bozec Y-M, Chabanet P, Cheal AJ, Cinner J, Dep-
czynski M, Feary DA, Gagliano M, Graham NAJ, Halford
AR, Halpern BS, Harbone AR, Hoey AS, Holbrook SJ,
Jones GP, Kulbiki M, Letourneur Y, De Loma TL,
McClanahan T, McCormick MI, Meekan MG, Mumby PJ,
Munday PL, Ohman MC, Pratchett MS, Riegl B, Sano M,
Schmitt RJ, Syms C (2010a) Crucial knowledge gaps in
current understanding of climate change impacts on coral
reef fishes. J Exp Biol 213:894–900
Wilson SK, Fisher R, Pratchett MS, Graham NAJ, Dulvy NK,
Turner RA, Cakacaka A, Polunin NVC (2010b) Habitat
degredation and fishing effects on the size structure of coral
reef fish communities. Ecol App 20:442–451
518 Rev Fish Biol Fisheries (2014) 24:505–518
123
Related Documents











