Short and Prolonged Periods of Maternal Separation and Voluntary Ethanol Intake in Male and Female Ethanol-Preferring AA and Ethanol-Avoiding ANA Rats Erika Roman, Lisa Gustafsson, Petri Hyytiä, and Ingrid Nylander Background: Genetic as well as environmental factors can affect the propensity for psychopathology and/or drug dependence. Maternal separation represents an animal experimental model that is useful in studies of effects of early life experiences. The authors have established a protocol for short and prolonged periods of maternal separation to study adult neurochemistry, behavior, and ethanol intake and have previously reported alterations in ethanol intake in Wistar rats and ethanol-preferring rats. The aim of the current study was to more thoroughly study how early life experiences affect an inherited propensity for high and low ethanol intake, respectively, in male and female ethanol-preferring AA (Alko alcohol) and ethanol-avoiding ANA (Alko, Non-Alcohol) rats. Methods: AA and ANA pups were assigned to one of three different rearing conditions: 15 min (MS15) or 360 min (MS360) of daily maternal separation in litters or normal animal facility rearing (AFR) during postnatal days 1 to 21. In adulthood, voluntary ethanol intake was investigated using the two-bottle free choice paradigm. Results: In male ethanol-preferring AA rats, MS15 resulted in a lower intake and fewer high-preferring animals at 8% and 10% ethanol compared with MS360 rats. The male MS360 rats had a higher ethanol intake at 8% and 10% ethanol in comparison with AFR rats. In contrast, the female AA MS15 and MS360 rats had a lower ethanol intake and a lower preference for the 10% ethanol solution compared with the female AA AFR rats. In male and female ANA rats, no major separation-induced effects were found. Conclusions: The current results show that genetic inheritance can be affected by environmental ma- nipulations in AA rats with an inherent high ethanol intake. The findings in female ethanol-preferring AA rats give further evidence of a differential outcome of maternal separation in male and female rats, as previously shown. Key Words: Handling, Maternal Deprivation, Alcohol, Estrous Cycle, Sex Differences. T HE SIGNIFICANCE OF genetic factors in the devel- opment of alcohol dependence has been described for humans as well as experimental animals (Crabbe and Phillips, 2004; Dick and Foroud, 2003; Gianoulakis, 2001; Schumann et al., 2003). Furthermore, the importance of environmental factors in the development and maintenance of drug abuse has gained increasing interest. In humans, for example, ad- verse experiences early in life can affect the individual vulner- ability to develop psychopathology (e.g., Carlson and Earls, 1997; Caspi et al., 2002, 2003; Gilmer and McKinney, 2003; Langeland et al., 2004; Nemeroff, 1998) and affect intake of drugs of abuse (e.g., Gordon, 2002; Langbehn et al., 2003; Langeland and Hartgers, 1998; Spak et al., 2001) later in life. However, the neurobiological mechanisms are not fully un- derstood. In recent studies in nonhuman primates, it was demonstrated that a genetic profile could affect ethanol in- take as a consequence of environmental manipulations (Barr et al., 2003, 2004). Furthermore, environmental factors may also provide protection against adult psychopathology (Caspi et al., 2002, 2003, 2004; Kim-Cohen et al., 2004), which further demonstrates the importance of studying the interaction be- tween environmental and genetic factors. In experimental animals, early life experiences can be studied using the maternal separation paradigm. Newborn and young rat pups are dependent on their nursing mothers for survival and normal development. Separation of rat pups from their mothers affects mother-infant interaction and alters neuronal development, resulting in a variety of neurochemical, hormonal, and behavioral changes, which may persist into adulthood. Short periods of maternal sep- aration (3–20 min), also known as handling, result in ani- From the Department of Pharmaceutical Biosciences, Division of Phar- macology, Uppsala University, Uppsala, Sweden (ER, LG, IN); and the Department of Mental Health and Alcohol Research, National Public Health Institute, Helsinki, Finland (PH). Received for publication June 17, 2004; accepted January 1, 2005. Supported by Grant K2002-04X-12588-05A from The Swedish Research Council, Grant 98/21:4 from the Alcohol Research Council of the Swedish Alcohol Retailing Monopoly, and grants from the AFA and the Facias Foundation. ER and LG contributed equally to this article. Reprint requests: Erika Roman, PhD, Department of Pharmaceutical Bio- sciences, Division of Pharmacology, Uppsala University, P.O. Box 591, SE-751 24 Uppsala, Sweden; Fax: 46-18-471-4847; E-mail: [email protected] Copyright © 2005 by the Research Society on Alcoholism. DOI: 10.1097/01.ALC.0000158933.70242.FC 0145-6008/05/2904-0591$03.00/0 ALCOHOLISM:CLINICAL AND EXPERIMENTAL RESEARCH Vol. 29, No. 4 April 2005 Alcohol Clin Exp Res, Vol 29, No 4, 2005: pp 591–601 591

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Short and Prolonged Periods of Maternal Separationand Voluntary Ethanol Intake in Male and Female
Ethanol-Preferring AA and Ethanol-Avoiding ANA RatsErika Roman, Lisa Gustafsson, Petri Hyytiä, and Ingrid Nylander
Background: Genetic as well as environmental factors can affect the propensity for psychopathologyand/or drug dependence. Maternal separation represents an animal experimental model that is useful instudies of effects of early life experiences. The authors have established a protocol for short and prolongedperiods of maternal separation to study adult neurochemistry, behavior, and ethanol intake and havepreviously reported alterations in ethanol intake in Wistar rats and ethanol-preferring rats. The aim of thecurrent study was to more thoroughly study how early life experiences affect an inherited propensity forhigh and low ethanol intake, respectively, in male and female ethanol-preferring AA (Alko alcohol) andethanol-avoiding ANA (Alko, Non-Alcohol) rats.
Methods: AA and ANA pups were assigned to one of three different rearing conditions: 15 min (MS15)or 360 min (MS360) of daily maternal separation in litters or normal animal facility rearing (AFR) duringpostnatal days 1 to 21. In adulthood, voluntary ethanol intake was investigated using the two-bottle freechoice paradigm.
Results: In male ethanol-preferring AA rats, MS15 resulted in a lower intake and fewer high-preferringanimals at 8% and 10% ethanol compared with MS360 rats. The male MS360 rats had a higher ethanolintake at 8% and 10% ethanol in comparison with AFR rats. In contrast, the female AA MS15 and MS360rats had a lower ethanol intake and a lower preference for the 10% ethanol solution compared with thefemale AA AFR rats. In male and female ANA rats, no major separation-induced effects were found.
Conclusions: The current results show that genetic inheritance can be affected by environmental ma-nipulations in AA rats with an inherent high ethanol intake. The findings in female ethanol-preferring AArats give further evidence of a differential outcome of maternal separation in male and female rats, aspreviously shown.
Key Words: Handling, Maternal Deprivation, Alcohol, Estrous Cycle, Sex Differences.
THE SIGNIFICANCE OF genetic factors in the devel-opment of alcohol dependence has been described for
humans as well as experimental animals (Crabbe and Phillips,2004; Dick and Foroud, 2003; Gianoulakis, 2001; Schumannet al., 2003). Furthermore, the importance of environmentalfactors in the development and maintenance of drug abusehas gained increasing interest. In humans, for example, ad-verse experiences early in life can affect the individual vulner-ability to develop psychopathology (e.g., Carlson and Earls,
1997; Caspi et al., 2002, 2003; Gilmer and McKinney, 2003;Langeland et al., 2004; Nemeroff, 1998) and affect intake ofdrugs of abuse (e.g., Gordon, 2002; Langbehn et al., 2003;Langeland and Hartgers, 1998; Spak et al., 2001) later in life.However, the neurobiological mechanisms are not fully un-derstood. In recent studies in nonhuman primates, it wasdemonstrated that a genetic profile could affect ethanol in-take as a consequence of environmental manipulations (Barret al., 2003, 2004). Furthermore, environmental factors mayalso provide protection against adult psychopathology (Caspiet al., 2002, 2003, 2004; Kim-Cohen et al., 2004), which furtherdemonstrates the importance of studying the interaction be-tween environmental and genetic factors.
In experimental animals, early life experiences can bestudied using the maternal separation paradigm. Newbornand young rat pups are dependent on their nursing mothersfor survival and normal development. Separation of ratpups from their mothers affects mother-infant interactionand alters neuronal development, resulting in a variety ofneurochemical, hormonal, and behavioral changes, whichmay persist into adulthood. Short periods of maternal sep-aration (3–20 min), also known as handling, result in ani-
From the Department of Pharmaceutical Biosciences, Division of Phar-macology, Uppsala University, Uppsala, Sweden (ER, LG, IN); and theDepartment of Mental Health and Alcohol Research, National Public HealthInstitute, Helsinki, Finland (PH).
Received for publication June 17, 2004; accepted January 1, 2005.Supported by Grant K2002-04X-12588-05A from The Swedish Research
Council, Grant 98/21:4 from the Alcohol Research Council of the SwedishAlcohol Retailing Monopoly, and grants from the AFA and the FaciasFoundation. ER and LG contributed equally to this article.
Reprint requests: Erika Roman, PhD, Department of Pharmaceutical Bio-sciences, Division of Pharmacology, Uppsala University, P.O. Box 591, SE-75124 Uppsala, Sweden; Fax: 46-18-471-4847; E-mail: [email protected]
Copyright © 2005 by the Research Society on Alcoholism.
DOI: 10.1097/01.ALC.0000158933.70242.FC
0145-6008/05/2904-0591$03.00/0ALCOHOLISM: CLINICAL AND EXPERIMENTAL RESEARCH
Vol. 29, No. 4April 2005
Alcohol Clin Exp Res, Vol 29, No 4, 2005: pp 591–601 591

mals with decreased emotional reactivity as well as in-creased ability to cope with and adapt to stressful stimuli asadults. However, prolonged periods of maternal separation(� 1 hr) result in effects that sometimes are opposite tothose observed after short periods of maternal separation(Hall, 1998; Ladd et al., 2000; Lehmann and Feldon, 2000;Newport et al., 2002; Pryce and Feldon, 2003), althoughresults are somewhat contradictory (Lehmann and Feldon,2000; Pryce and Feldon, 2003). Furthermore, maternal sep-aration has been shown to affect ethanol intake behavior inthe adult animal (Fahlke et al., 2000; Higley et al., 1991;Hilakivi-Clarke et al., 1991; Huot et al., 2001; Jaworski etal., 2004; Weinberg, 1987). The time course of developmentdiffers between females and males (Andersen, 2003), and,as a consequence, maternal separation may induce sex-dependent effects. Evidence for sex-dependent outcome ofmaternal separation has been presented (Papaioannou etal., 2002a, 2002b; Park et al., 2003). We have previouslyshown that short and prolonged periods of maternal sepa-ration, using the same experimental protocol as in thecurrent study, altered voluntary ethanol intake in maleWistar rats (Ploj et al., 2003a). Interestingly, no effectswere found in female Wistar rats (Gustafsson et al., 2005;Roman et al., 2004).
In the current study, the ethanol-preferring AA (Alko,Alcohol) and ethanol-avoiding ANA (Alko, Non-alcohol)rats were used. These animal lines were developed byselective breeding and show a large difference in volun-tary ethanol intake. AA rats are considered to drink etha-nol for its reinforcing effects, whereas ANA rats avoidethanol (Eriksson, 1973; Eriksson 1968, 1971; Hilakivi etal., 1984; Hyytiä et al., 1996; Sinclair, 1974). We havepreviously shown maternal separation–induced effects onvoluntary ethanol intake in male ethanol-preferring AArats (Roman et al., 2003). However, in that study, thenumber of animals was limited, and the period of access toethanol was relatively short. The aim of the current exper-iment was to more thoroughly investigate the impact ofmaternal separation on acquisition of voluntary ethanolintake and the subsequent ethanol intake in rats with agenetic inheritance. To further broaden the knowledgeabout the influence of early life experiences on later etha-nol intake, ethanol-avoiding ANA rats, the counterparts toAA rats, were included. ANA rats have a well-documentedlow voluntary ethanol intake and, since 360 min of maternalseparation has been shown to result in a high voluntaryethanol intake, it was of interest to investigate whether thiswould be true also for the ANA rats. The use of both malesand females within the same experiment allows furtherexamination of sex differences. Male and female AA andANA pups were subjected to daily periods of maternalseparation for 15 min (MS15) or 360 min (MS360) orhoused under normal animal facility rearing conditions(AFR) during the first 3 weeks of life. In adulthood, therats had continuous access to ethanol to study the effects of
short and prolonged periods of maternal separation onvoluntary intake.
MATERIALS AND METHODS
Animals and Housing
Time-mated pregnant AA and ANA rats (F85 generation) were sup-plied from the National Public Health Institute, Helsinki, Finland. Onarrival, the dams were singly housed in standard macrolon cages (59 � 38� 20 cm) containing wood-chip bedding material and nesting material.The rats were maintained on a 12-hr light/dark cycle with lights on at 0600hr and standard pellet food (R36 Labfor; Lactamin, Vadstena, Sweden)and water ad libitum. The rats were housed in a temperature- (22 � 1°C)and humidity-controlled (50 � 10%) animal room used only for thisexperiment. All animal experiments were performed under a protocolapproved by the local ethical committee and in accordance with theSwedish Animal Protection Legislation.
Maternal Separation
The protocol for short and prolonged periods of maternal separation isnow well established in our group (Gustafsson et al., 2005; Ploj et al., 2002,2003a, 2003b; Roman et al., 2003, 2005). On the day of birth (day 0), thepups were sexed by measurement of anogenital distance and cross-fostered into new litters. The cross-fostering procedure was used due to alimited number of pregnant AA and ANA females available. The litter-mates were, as far as possible, spread among the dams so that the newlitters contained a mix of pups from several dams. This procedure resultsin the same maternal influences impacting on pups of different geneticbackgrounds. The arranged litters contained eight to nine pups with ahomogenous sex distribution, as far as possible. The litters (AA n � 14,ANA n � 8) were thereafter randomly assigned to one of three rearingconditions: 15 min (MS15; AA n � 5, ANA n � 2) or 360 min (MS360; AAn � 5, ANA n � 3) of maternal separation or normal animal facilityrearing (AFR; AA n � 4, ANA n � 3). The separations occurred oncedaily during postnatal days (PNDs) 1 to 21. First the dam and then thewhole litter was removed from the home cage. The pups were at all timeskept close together in the respective litters, and the pups were neverindividually handled. Each litter was placed in macrolon cages (26 � 20 �14 cm) containing wood-chip bedding material and moved to an adjacentroom (25 � 0.4°C) for 15 or 360 min, respectively. The cages in which thelitters were placed during the separation were changed every day. Thedams of the MS360 litters were returned to their home cages duringthe separation procedure but taken out before the return of the litters. Thedams of the MS15 litters were transferred to another cage duringthe separation, and the litters were returned to the home cages before thedams. The separation sessions were performed in the same room andat the same time every day, starting at 0900 hr for the MS15 groupsand at 0930 hr for the MS360 groups. The litters of the MS15 andMS360 groups were inspected daily to determine the day for furringand eye opening as a control for normal development. The MS15 andMS360 litters were weighed on PNDs 1, 4, 7, 10, 13, 16, and 19, whereasAFR litters were weighed on PNDs 7 and 16, when cage changes wereperformed for AFR litters as well as for MS15 and MS360 litters.During cage changes, a small part of the old bedding material wasmixed with the clean bedding material, and the old nesting material wastransferred to the new cage to limit potential stressful experiences. TheAFR litters were handled only when the cages were changed. The sameperson, who was the only person with permission to enter the animalroom during the period from the arrival of the pregnant dams untilPND 22, performed the cage changes as well as the separations. Gloveswere used in all contact with the animals. All litters were weighed at thetime for weaning on PND 22, both as whole litters as well as each litterdivided by sex. After weaning, the rats were left undisturbed, exceptfrom normal animal care, and were housed in standard macrolon cages
592 ROMAN ET AL.

(59 � 38 � 20 cm) in the same strain, sex, and experimental groups oftwo to five rats per cage.
Voluntary Ethanol Intake
Starting on PNDs 77 to 86, the AA rats (male MS15 n � 20, femaleMS15 n � 19, male MS360 n � 20, female MS360 n � 20, male AFR n �15, female AFR n � 14) weighing 201 to 367 g (males) and 155 to 220 g(females) and the ANA rats (male MS15 n � 8, female MS15 n � 8, maleMS360 n � 12, female MS360 n � 8, male AFR n � 8, female AFR n �9) weighing 288 to 405 g (males) and 206 to 291 g (females) were singlyhoused in macrolon cages (42 � 26 � 18 cm) and were introduced toethanol by giving them continuous access to a bottle containing an ethanolsolution in addition to the water bottle (plastic 150 ml-bottles [males] or250 ml-bottles [females] with ball-valve nipples, Scanbur BK AB, Sollen-tuna, Sweden). Bottle positions were randomly changed to avoid positionpreferences. For the AA rats, the ethanol concentrations (v/v) were grad-ually increased (4 days with 2%, 4 days with 4%, 4 days with 6%, 4 dayswith 8%); thereafter the rats had access to 10% ethanol for 27 days. Forthe ANA rats, the ethanol concentrations were gradually increased as forthe AA rats (4 days with 2%, 4 days with 4%, 4 days with 6%, 4 days with8%), but because all ANA rats had a very low intake at 8% ethanol, therats were maintained on 6% ethanol, instead of 10%, for the remaining 27days before the end of the experiment on PNDs 120 to 129. Measurementsof ethanol and water intake were taken daily, and food intake and bodyweight were measured every third day. For practical reasons, all measure-ments in female rats were made in the morning, whereas measurements inmale rats were made in the afternoon.
Estrous Cycle
For analyses of ethanol intake during the different phases of the estrouscycle in the female rats, vaginal smears were collected from all animals fivedays a week, starting on the 17th day of the ethanol intake period andthroughout the experiment. The samples were taken between 0900 and1130 hr. A small amount of saline was flushed into the vaginal canal usinga plastic pipette with the edge rounded off to avoid injuring the mucous ofthe vagina. The fluid, containing vaginal cells, was then instantly retrieved,transferred to a slide, and left to dry. A May-Grünwald-Giemsa stainingwas used for histologic evaluation. The dry smears were fixed in methanol(5 min, room temperature), then transferred to May-Grünwald solution (5min, room temperature) for staining of the cytoplasm and granulae, andfinally left in Giemsa solution for 15 min (1:50 in water, room tempera-ture) for staining of nuclei and cytoplasm. The smears were then rinsedunder water and left to dry. The smears were analyzed under a microscope(magnification � 100), and the stages identified were diestrus (leuko-cytes), proestrus (nucleated epithelial cells), and estrus (cornified epithe-lial cells).
Statistical Analyses
The mean pup weights during PNDs 1 to 21, at the time for weaning onPND 22, as well as body weights during the period of access to ethanolwere analyzed with one-factor ANOVA followed by Fisher protected leastsignificant difference (PLSD) test. The fluid intake data and the foodintake data did not show a normal distribution, and nonparametric statis-tics, Kruskal-Wallis test followed by Mann-Whitney U test, was thereforeused for the analyses. The number of high-preferring rats in each exper-imental group was analyzed using �2 test. Whenever significant effectswere found with the Mann-Whitney U test or the �2 test, statisticalsignificances were further corrected using the Bonferroni-Holm test(Holm, 1979) to correct for type I errors. Analyses of sex differences involuntary ethanol intake were performed using the Mann-Whitney U test.The ethanol intake during the different phases of the estrous cycle wasanalyzed with the nonparametric Friedman test. StatView 5.0.1 (SASInstitute Inc., Cary, NC) computer software was used for all statisticalanalyses. Differences were considered statistically significant at p � 0.05.
RESULTS
There were no differences in normal development interms of furring or eye opening in AA or ANA pups duringthe postnatal period.
In ANA rats but not AA rats, a digging and buryingbehavior was noted when cages were changed during theperiod of access to ethanol, even though some of the oldbedding material was mixed with clean bedding material tominimize potential stress. In male ANA rats, this behaviorwas more pronounced in the beginning of the period ofaccess to ethanol but could be seen more or less during theentire period, whereas the digging and burying behaviorattenuated during the end of the drinking period in thefemale ANA rats, most probably because of the increasedhandling related to the collection of vaginal smears.
Effects of Maternal Separation on Body Weight
A significant difference in mean pup weight (g) in the AAMS15, MS360, and AFR litters was found on PND 7 [F(2,11)� 4.4, p � 0.05], whereas no difference was found on PND 16[F(2,11) � 2.9, not significant]. On PND 7, the mean pupweight was significantly lower (p � 0.05) in AA MS360 litterscompared with AA AFR litters. No differences were observedin mean pup weights in the MS15 and MS360 litters at any ofthe seven measurements during PNDs 1 to 21. For ANAlitters, no significant differences were found among MS15,MS360, and AFR pups. The weight gain during PNDs 1 to 21showed a similar pattern for all experimental groups in theAA and ANA litters, respectively.
At the time for weaning, on PND 22, no differences werefound in mean pup weight between experimental groups,either in AA litters [F(2,11) � 1.7, not significant] or inANA litters [F(2,5) � 1.2, not significant]. Furthermore,there were no statistically significant differences when ei-ther AA and ANA males only [F(2,11) � 2.0, not signifi-cant and F(2,5) � 0.8, not significant, respectively] or AAand ANA females only [F(2,11) � 1.1, not significant andF(2,5) � 1.4, not significant, respectively] were comparedin each litter and experimental group. On PND 22, thefollowing weights were recorded for male pups: AA MS15,39–51 g; AA MS360, 40–49 g; AA AFR, 45–56 g; ANAMS15, 45–49 g; ANA MS360, 46–57 g; ANA AFR, 48–57g. The female pups weights were as follows: AA MS15,39–49 g; AA MS360, 41–47 g; AA AFR, 41–55 g; ANAMS15, 44–48 g; ANA MS360, 46–54 g; ANA AFR, 47–55 g.
When the male rats were weighed for the first time asadults, no differences in body weight (g) between the ex-perimental groups were found, either for AA rats [F(2,52)� 2.6, not significant] or for ANA rats [F(2,25) � 3.3, notsignificant]. However, in male AA rats, the AFR rats hadan overall higher body weight than AA MS15 and AAMS360 rats during the period of access to ethanol. BothAA MS15 and AA MS360 rats weighed significantly less at10 out of a total of 15 measures in comparison with the AAAFR rats. No differences were found between male AA
MATERNAL SEPARATION AND ETHANOL INTAKE IN AA AND ANA RATS 593

MS15 and AA MS360 rats at any time. In the three exper-imental male ANA groups, ANA MS360 rats had an overallhigher body weight during the entire period of continuousaccess to ethanol, and this higher body weight reached thelevel of significance at 7 out of a total of 15 measures incomparison with ANA MS15 rats but not in comparisonwith ANA AFR rats. No differences were found whencomparing male ANA MS15 and ANA AFR rats or ANAMS360 and ANA AFR rats. The body weight gain showedsimilar patterns for the respective male AA and ANAexperimental groups.
The first weighing in adulthood in the female AA ratsindicated statistically significant differences in mean bodyweight in the AA rats [F(2,50) � 5.0, p � 0.05]. The femaleAA MS360 group showed a significantly lower mean bodyweight than the MS15 and AFR groups (p � 0.01 and p �0.05, respectively). This difference then remained duringthe period of access to ethanol. However, in the femaleANA rats, no differences were found in mean body weight[F(2,22) � 2.1, not significant] between the experimentalgroups on the first weighing. However, the ANA MS15group showed a significantly lower mean body weight thanthe MS360 group at 10 out of a total of 15 measures. Thebody weight gain showed similar patterns for the respectivefemale AA and ANA experimental groups.
Both male and female ANA rats had an overall higherbody weight in comparison with the respective male andfemale AA rats during the entire period of access to etha-nol, as previously shown (Sinclair et al., 1989).
Effects of Maternal Separation on Fluid Intake in AA Rats
The design of the voluntary ethanol intake and the dailyethanol drinking patterns (g/kg/day) in male and female AAMS15, MS360, and AFR rats during the entire period ofcontinuous access to ethanol are illustrated in Fig. 1A and 1B.The male AA MS360 rats had an overall higher ethanol intakecompared with both AA MS15 and AA AFR rats throughoutthe experiment (Fig. 1A). In the female AA MS15, MS360,and AFR rats (Fig. 1B), the ethanol intake was similar in thethree experimental groups at the lower ethanol concentra-tions, but during the period with 8% ethanol solution, theintake curves started to deviate. During the first 13 days with10% ethanol, the intake slightly increased in both males andfemales, followed by a stabilization during the last 14 days.Therefore, the period with access to 10% ethanol was alsodivided into two phases, the first 13 days and the last 14 days,in the statistical analyses.
Figure 2 shows the ethanol intake (g/kg/day) in male andfemale AA MS15, MS360, and AFR rats during the differ-ent ethanol concentrations used. In the male AA rats, thestatistical analyses revealed that the AA MS15 rats had asignificantly lower ethanol intake at 8% ethanol in compar-ison with AA MS360 rats. Also, during the entire period of10% ethanol, significant group differences were found, withAA MS15 rats having a significantly lower intake compared
with AA MS360 rats. When dividing the period of access to10% ethanol in two parts (data not shown), it was foundthat the ethanol intake was significantly lower during thefirst 13 days (U � 93.0, p � 0.01) but not during the last 14days (U � 133.5, not significant). In the male MS360 rats,a statistically significant higher ethanol intake was foundduring 8% ethanol as well as during the entire period of10% ethanol, during both the first 13 days (U � 60.0, p �0.01) and the last 14 days (U � 53.0, p � 0.01) of access to10% ethanol (data not shown) compared with AFR rats.No differences were found between male AA MS15 andAFR rats at any concentration. The ethanol preference(percent of total fluid intake; data not shown) was in con-cordance with the ethanol intake, with the male AA MS360rats having the highest preference. However, afterBonferroni-Holm correction, no differences reached thelevel of significance. Water intake (g/kg/day; data notshown) and total fluid intake (g/kg/day; data not shown) didnot show a significant difference between the male AAgroups at any time during the period of access to ethanol.
In the female AA rats, the statistical analyses did notreveal significant differences between the experimentalgroups at 2–8% ethanol. Differences between the threegroups were obtained when analyzing the entire periodwith 10% ethanol (H � 6.4, p � 0.05) but not after theBonferroni-Holm correction. During the first 13 days with10% ethanol solution, the MS15 group showed a signifi-cantly lower ethanol intake (U � 66.0, p � 0.05) in com-parison with the AFR group, whereas no differences couldbe observed during the last 14 days (U � 74.0, not signifi-cant). In the female AA rats, no statistically significantdifferences in ethanol preference (percent of total fluidintake; data not shown) were detected at 2–8%. During theperiod with access to 10%, differences were observed be-tween the experimental groups (H � 8.5, p � 0.05). Theethanol preference was lower in both MS15 and MS360 rats(U � 65.0, p � 0.05 and U � 65.0, p � 0.01, respectively)compared with AFR rats. The lower ethanol preference inthe MS360 group was observed during the first 13 days (U� 73.0, p � 0.05) as well as during the last 14 days (U �74.0, p � 0.05) with 10% ethanol. Also, in the MS15 group,the ethanol preference was lower during both periods.However, after the Bonferroni-Holm correction, the etha-nol preference just failed statistical significance the firstperiod (U � 68.0, not significant), whereas the intake dur-ing the last 14 days with 10% ethanol was significantlylower (U � 66.0, p � 0.05). No statistically significantdifferences were seen in total fluid intake (g/kg/day; datanot shown) among the female experimental groups. How-ever, in water intake (g/kg/day; data not shown), there weredifferences found during the period with 10% ethanol so-lution (H � 7.9, p � 0.05), where the MS15 and MS360groups showed a higher water intake than the AFR group(U � 64.0, p � 0.05 and U � 70.0, p � 0.05, respectively).
The number of animals with a high ethanol preference ineach group was further investigated using a limit of 60%, with
594 ROMAN ET AL.
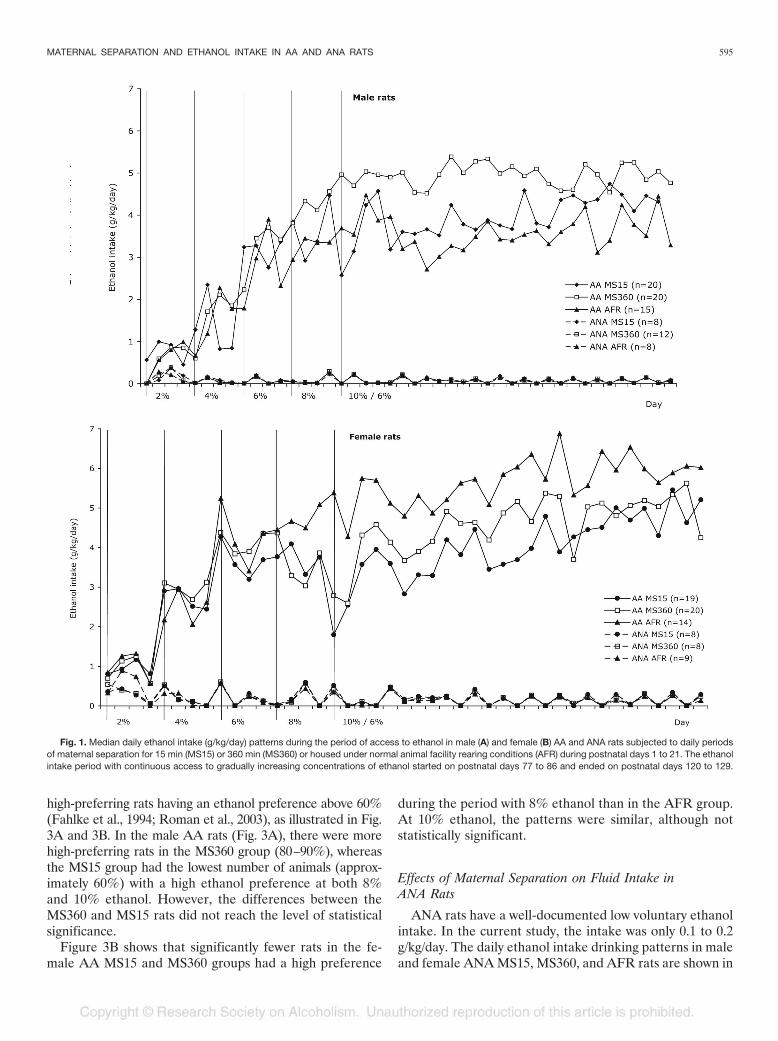
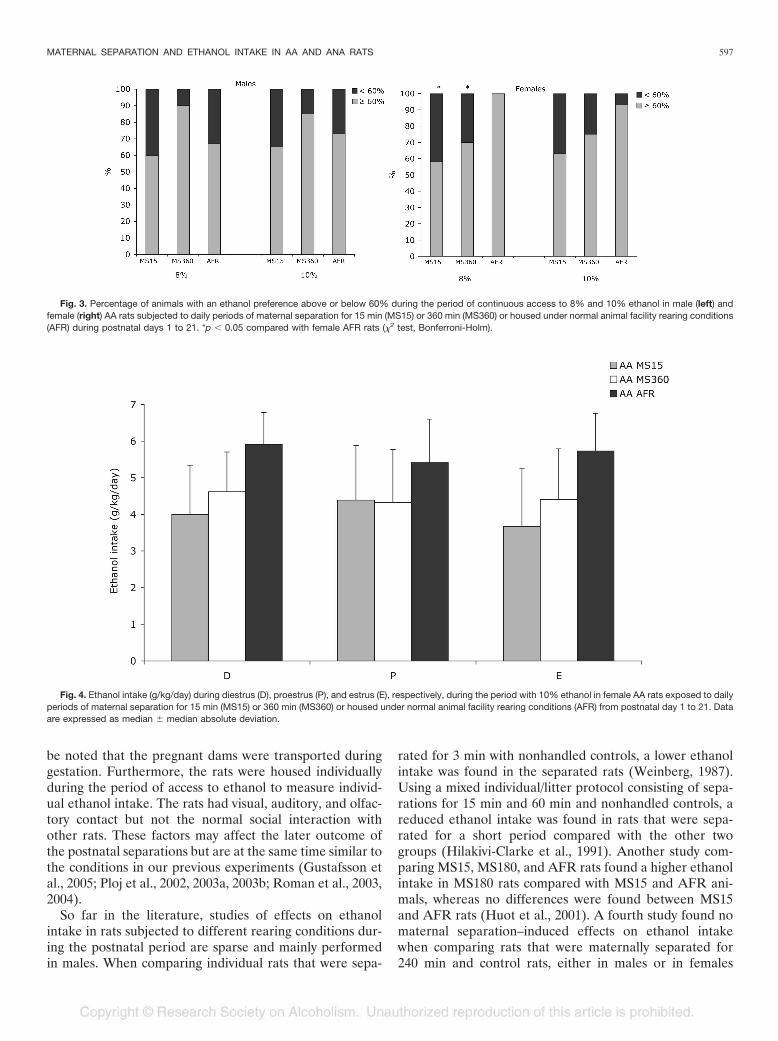
high-preferring rats having an ethanol preference above 60%(Fahlke et al., 1994; Roman et al., 2003), as illustrated in Fig.3A and 3B. In the male AA rats (Fig. 3A), there were morehigh-preferring rats in the MS360 group (80–90%), whereasthe MS15 group had the lowest number of animals (approx-imately 60%) with a high ethanol preference at both 8%and 10% ethanol. However, the differences between theMS360 and MS15 rats did not reach the level of statisticalsignificance.
Figure 3B shows that significantly fewer rats in the fe-male AA MS15 and MS360 groups had a high preference
during the period with 8% ethanol than in the AFR group.At 10% ethanol, the patterns were similar, although notstatistically significant.
Effects of Maternal Separation on Fluid Intake inANA Rats
ANA rats have a well-documented low voluntary ethanolintake. In the current study, the intake was only 0.1 to 0.2g/kg/day. The daily ethanol intake drinking patterns in maleand female ANA MS15, MS360, and AFR rats are shown in
Fig. 1. Median daily ethanol intake (g/kg/day) patterns during the period of access to ethanol in male (A) and female (B) AA and ANA rats subjected to daily periodsof maternal separation for 15 min (MS15) or 360 min (MS360) or housed under normal animal facility rearing conditions (AFR) during postnatal days 1 to 21. The ethanolintake period with continuous access to gradually increasing concentrations of ethanol started on postnatal days 77 to 86 and ended on postnatal days 120 to 129.
MATERNAL SEPARATION AND ETHANOL INTAKE IN AA AND ANA RATS 595
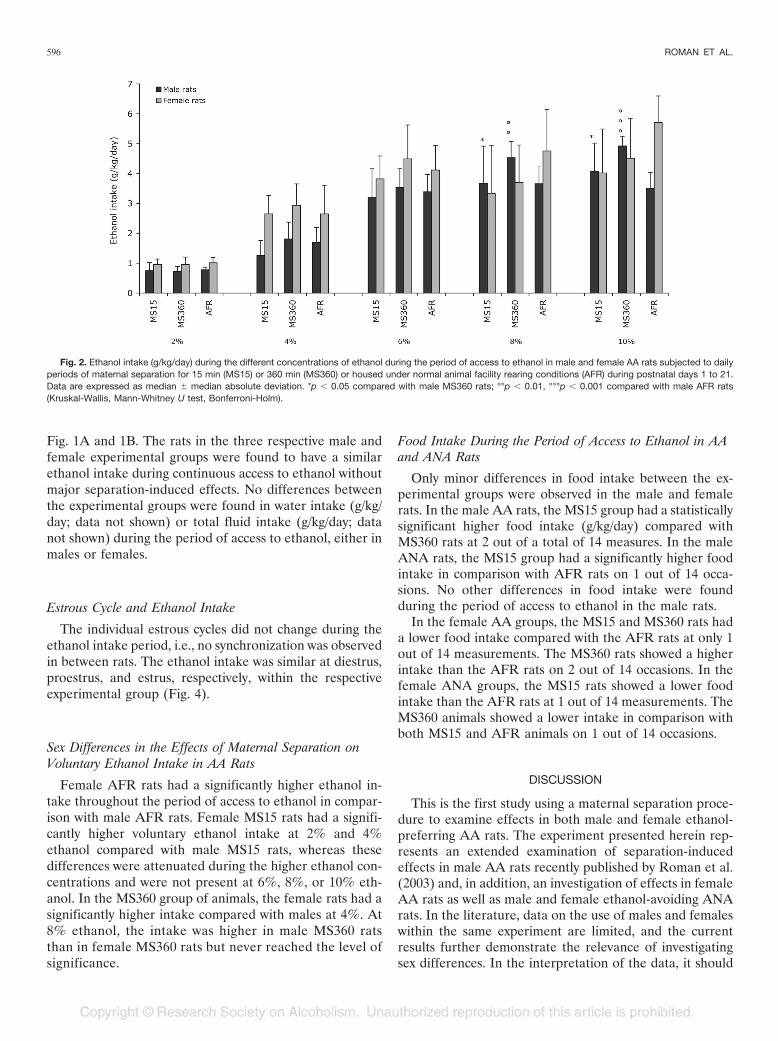
Fig. 1A and 1B. The rats in the three respective male andfemale experimental groups were found to have a similarethanol intake during continuous access to ethanol withoutmajor separation-induced effects. No differences betweenthe experimental groups were found in water intake (g/kg/day; data not shown) or total fluid intake (g/kg/day; datanot shown) during the period of access to ethanol, either inmales or females.
Estrous Cycle and Ethanol Intake
The individual estrous cycles did not change during theethanol intake period, i.e., no synchronization was observedin between rats. The ethanol intake was similar at diestrus,proestrus, and estrus, respectively, within the respectiveexperimental group (Fig. 4).
Sex Differences in the Effects of Maternal Separation onVoluntary Ethanol Intake in AA Rats
Female AFR rats had a significantly higher ethanol in-take throughout the period of access to ethanol in compar-ison with male AFR rats. Female MS15 rats had a signifi-cantly higher voluntary ethanol intake at 2% and 4%ethanol compared with male MS15 rats, whereas thesedifferences were attenuated during the higher ethanol con-centrations and were not present at 6%, 8%, or 10% eth-anol. In the MS360 group of animals, the female rats had asignificantly higher intake compared with males at 4%. At8% ethanol, the intake was higher in male MS360 ratsthan in female MS360 rats but never reached the level ofsignificance.
Food Intake During the Period of Access to Ethanol in AAand ANA Rats
Only minor differences in food intake between the ex-perimental groups were observed in the male and femalerats. In the male AA rats, the MS15 group had a statisticallysignificant higher food intake (g/kg/day) compared withMS360 rats at 2 out of a total of 14 measures. In the maleANA rats, the MS15 group had a significantly higher foodintake in comparison with AFR rats on 1 out of 14 occa-sions. No other differences in food intake were foundduring the period of access to ethanol in the male rats.
In the female AA groups, the MS15 and MS360 rats hada lower food intake compared with the AFR rats at only 1out of 14 measurements. The MS360 rats showed a higherintake than the AFR rats on 2 out of 14 occasions. In thefemale ANA groups, the MS15 rats showed a lower foodintake than the AFR rats at 1 out of 14 measurements. TheMS360 animals showed a lower intake in comparison withboth MS15 and AFR animals on 1 out of 14 occasions.
DISCUSSION
This is the first study using a maternal separation proce-dure to examine effects in both male and female ethanol-preferring AA rats. The experiment presented herein rep-resents an extended examination of separation-inducedeffects in male AA rats recently published by Roman et al.(2003) and, in addition, an investigation of effects in femaleAA rats as well as male and female ethanol-avoiding ANArats. In the literature, data on the use of males and femaleswithin the same experiment are limited, and the currentresults further demonstrate the relevance of investigatingsex differences. In the interpretation of the data, it should
Fig. 2. Ethanol intake (g/kg/day) during the different concentrations of ethanol during the period of access to ethanol in male and female AA rats subjected to dailyperiods of maternal separation for 15 min (MS15) or 360 min (MS360) or housed under normal animal facility rearing conditions (AFR) during postnatal days 1 to 21.Data are expressed as median � median absolute deviation. *p � 0.05 compared with male MS360 rats; °°p � 0.01, °°°p � 0.001 compared with male AFR rats(Kruskal-Wallis, Mann-Whitney U test, Bonferroni-Holm).
596 ROMAN ET AL.
be noted that the pregnant dams were transported duringgestation. Furthermore, the rats were housed individuallyduring the period of access to ethanol to measure individ-ual ethanol intake. The rats had visual, auditory, and olfac-tory contact but not the normal social interaction withother rats. These factors may affect the later outcome ofthe postnatal separations but are at the same time similar tothe conditions in our previous experiments (Gustafsson etal., 2005; Ploj et al., 2002, 2003a, 2003b; Roman et al., 2003,2004).
So far in the literature, studies of effects on ethanolintake in rats subjected to different rearing conditions dur-ing the postnatal period are sparse and mainly performedin males. When comparing individual rats that were sepa-
rated for 3 min with nonhandled controls, a lower ethanolintake was found in the separated rats (Weinberg, 1987).Using a mixed individual/litter protocol consisting of sepa-rations for 15 min and 60 min and nonhandled controls, areduced ethanol intake was found in rats that were sepa-rated for a short period compared with the other twogroups (Hilakivi-Clarke et al., 1991). Another study com-paring MS15, MS180, and AFR rats found a higher ethanolintake in MS180 rats compared with MS15 and AFR ani-mals, whereas no differences were found between MS15and AFR rats (Huot et al., 2001). A fourth study found nomaternal separation–induced effects on ethanol intakewhen comparing rats that were maternally separated for240 min and control rats, either in males or in females
Fig. 3. Percentage of animals with an ethanol preference above or below 60% during the period of continuous access to 8% and 10% ethanol in male (left) andfemale (right) AA rats subjected to daily periods of maternal separation for 15 min (MS15) or 360 min (MS360) or housed under normal animal facility rearing conditions(AFR) during postnatal days 1 to 21. *p � 0.05 compared with female AFR rats (�2 test, Bonferroni-Holm).
Fig. 4. Ethanol intake (g/kg/day) during diestrus (D), proestrus (P), and estrus (E), respectively, during the period with 10% ethanol in female AA rats exposed to dailyperiods of maternal separation for 15 min (MS15) or 360 min (MS360) or housed under normal animal facility rearing conditions (AFR) from postnatal day 1 to 21. Dataare expressed as median � median absolute deviation.
MATERNAL SEPARATION AND ETHANOL INTAKE IN AA AND ANA RATS 597

(Marmendal et al., 2004). In a fifth study comparing MS15,MS180, and AFR male rats, it was demonstrated that MS15rats consumed significantly less ethanol compared withMS180 rats, but neither group differed from the AFRgroup of rats (Jaworski et al., 2004). Using the currentexperimental protocol, it was demonstrated that MS15 re-sulted in a lower ethanol intake in male Wistar rats incomparison with AFR and MS360 rats, whereas MS360increased voluntary ethanol intake compared with MS15and AFR rats (Ploj et al., 2003a). Interestingly, no effects ofMS15 and MS360 on adult ethanol intake were detected infemale Wistar rats (Gustafsson et al., 2005; Roman et al.,2004). Taken together, results from a variety of maternalseparation protocols have been presented and the interpre-tation and comparisons of these compiled data are some-times rather complicated. The use of AFR or nonhandledrats as a comparison group further complicates the inter-pretations because these two groups often differ from eachother (Ladd et al., 2000; Lehmann and Feldon, 2000; Pryceand Feldon, 2003), which elegantly was demonstrated in arecent study investigating effects on voluntary ethanol in-take (Jaworski et al., 2004).
The results from the previous studies in male rats indi-cate that short periods of maternal separation could serveas protection against a high voluntary ethanol intake laterin life. In the current study, AA MS15 rats showed a lowerethanol intake and ethanol preference during continuousaccess to ethanol as compared with MS360 rats. Further-more, fewer high-preferring animals (ethanol preference �60%) were present in the MS15 group than in the MS360group. When comparing the ethanol preference in thecurrent study, similar numbers of high-preferring animalswere found in the MS15 and AFR groups, except towardthe end of the experiment, where the number of AFR ratswith a high preference had increased. The difference be-tween AA MS15 and AFR rats was not as pronounced aspreviously shown (Roman et al., 2003) but was similar toprevious reports in nonpreferring rats (Huot et al., 2001;Jaworski et al., 2004). This different outcome in AA ratsfrom two different experiments could possibly be due to thefact that the AA rats in the previous study (Roman et al.,2003) were younger when they had access to ethanol thanin the current study; however, the discrepancies must befurther investigated.
In this first study investigating the effects of maternalseparation in female ethanol-preferring AA rats, the etha-nol intake, as well as the ethanol preference, was lower inthe MS15 rats than in the AFR rats. This is in accord withthe hypothesis that MS15 could reduce the propensity forexcessive ethanol intake later in life. Therefore, in ethanol-preferring rats, this proposed protective environment re-sulted in a reduced ethanol intake, but this effect was morepronounced in female AA rats than in males.
Prolonged periods of maternal separation have previ-ously been shown to increase ethanol intake in adulthood.However, AA rats subjected to MS360 showed less pro-
nounced effects, with only a trend toward a higher ethanolintake compared with AFR rats (Roman et al., 2003),possibly caused by too few rats in the experimental groups.There have also been good reasons to believe that AA ratsmay be resistant to manipulations to further increase analready high ethanol intake. Most attempts to increase theethanol intake in AA rats have been studies of ethanoldeprivation. In Wistar, Long-Evans, and ethanol-preferringP (P for preferring) rats, deprivation-induced drinking in-creased with the length of the deprivation. However, in AArats, up to 24 hr of deprivation increased drinking maxi-mally, with no further increases with longer periods ofdeprivation (Sinclair and Li, 1989). A more recent studyshowed significant deprivation-induced increases in etha-nol intake in Wistar rats and P rats, whereas no effect wasobserved in AA rats and HAD (high-alcohol-drinking) rats(Vengeliene et al., 2003). Therefore, it is an interestingfinding that MS360 can increase voluntary ethanol intakealso in AA rats with an inherent high voluntary ethanolintake, as shown in the current extended study.
In the current study, female AA rats subjected to MS360had a similar ethanol intake as the female MS15 rats, i.e.,rats in the MS360 group also had a lower ethanol intakethan the AFR rats. Therefore, in contrast to findings inmale AA rats, short and prolonged periods of maternalseparation seem to induce similar effects in female AA rats.These new findings are not easily explained, but one reasonfor the similar MS15- and MS360-induced effects could bethat the separation per se is not critical in these female AArats. Instead, it is the handling, or nonhandling, of thelitters that seems to be the critical factor, i.e., daily handlingof the litters during the separation procedure, that the AFRrats are not exposed to.
In male and female ANA rats with an inherent lowvoluntary ethanol intake, no specific separation-inducedeffects on voluntary ethanol intake could be found. Thedigging and burying behavior observed during cagechanges, as previously reported in these rats (Sandbak etal., 1998), supports the common notion that the ANA ratshave a different behavioral profile than AA rats (Badishtovet al., 1995; Fahlke et al., 1993; Knapp et al., 1997; Korpi etal., 1988; Möller et al., 1997; Overstreet et al., 1997; Sali-mov et al., 1996; Sandbak et al., 1998; Tuominen et al.,1990; Viglinskaya et al., 1995). The general picture emerg-ing from these studies is that the ANA rats are moresusceptible to stress-induced behavioral changes than theAA rats, suggesting that the ANA line of rats could alsorespond to manipulations such as maternal separation, andthis was therefore worth investigating. However, it is alsopossible that the main factor controlling ethanol consump-tion in ANA rats is the accumulation of acetaldehyde dur-ing ethanol exposure (Eriksson, 1973; Hilakivi et al., 1984;Sinclair et al., 1989), which may limit the effect of anymanipulation that would increase ethanol intake in otherrats without this metabolic trait.
Taken together, the protective effect of short periods of
598 ROMAN ET AL.

daily maternal separation in terms of high ethanol intakeseems to be a general phenomenon, previously shown inmale nonpreferring and preferring rats (Hilakivi-Clarke etal., 1991; Huot et al., 2001; Jaworski et al., 2004; Ploj et al.,2003a; Roman et al., 2003; Weinberg, 1987) and here inmale and female AA rats. It is noteworthy, however, thatfemale preferring and nonpreferring rats are differentiallyaffected by short separations. In contrast to Wistar females(Gustafsson et al., 2005; Marmendal et al., 2004; Roman etal., 2004), the AA rats showed a low adult ethanol intake.Prolonged periods of maternal separation were here shownto further increase adult voluntary ethanol intake in maleAA rats as previously has been shown in nonpreferring rats(Huot et al., 2001; Ploj et al., 2003a), and MS360 mighttherefore serve as a risk factor for high ethanol intake inmale rats. Interestingly, the results indicated that femaleethanol-preferring AA rats had a lower ethanol intake alsoafter prolonged periods of maternal separation, in contrastto the unaffected intake in female Wistar rats (Gustafssonet al., 2005; Marmendal et al., 2004; Roman et al., 2004).Sex differences in various aspects of intake of drugs ofabuse have recently been summarized (Roth et al., 2004).In the two-bottle free choice paradigm, female rats havebeen shown to have a higher voluntary ethanol intake thanmales (Cailhol and Mormede, 2001; Juarez and Barrios deTomasi, 1999; Lancaster and Spiegel, 1992). This was truealso for the AA AFR group of animals, where the femaleAFR rats had a significantly higher ethanol intake through-out the experiment compared with male AFR rats. In thetwo different rearing groups, MS15 and MS360, sex differ-ences were observed during the lower concentrations ofethanol, where female AA rats had a higher voluntaryethanol intake than males. However, during 8% and 10%ethanol, these differences were no longer present, indicat-ing different effects of MS15 and MS360 on ethanol intakein male and female AA rats. The current results thereforeadd further evidence for sex differences in the outcome ofmaternal separation on voluntary ethanol intake. The un-derlying mechanisms for the sex differences observed arenot known. Several studies describe sex differences in long-term effects of different maternal separation proceduresduring the postnatal period (e.g., Kalinichev et al., 2001,2002; Lehmann et al., 1999; McIntosh et al., 1999; Papaio-annou et al., 2002a, 2002b; Park et al., 2003; Wigger andNeumann, 1999). Possible explanations may include differ-ent separation-induced neurochemical effects in males andfemales, hormone-dependent effects, and/or sex-dependentmother-infant behavior during the postnatal period. Anearly study found that mother rats interacted differentlywith male and female offspring (Moore and Morelli, 1979),whereas a recent study found no such differences (Cham-pagne et al., 2003). It has also been reported that naturalvariations in maternal behavior can result in sex-specificneurochemical changes in adulthood (Francis et al., 2002).Another factor of importance in terms of ethanol intakethat might have an effect is housing. Male rats were found
to have higher corticosterone levels under crowded condi-tions, whereas female rats were found to have the highestlevels when individually housed (Brown and Grunberg,1995). It has also been demonstrated that housing condi-tions can affect reward sensitivity in rats (van der Harst etal., 2003). Another factor of importance is the hormonalinfluences in female rats. The ethanol intake in femalesmay vary across the estrous cycle, although previous resultspresent a complex picture. It has been shown that theestrous cycle was without influence in rats (Ford et al.,2002; Roberts et al., 1998) and in humans across the men-strual cycle (Holdstock and de Wit, 2000), whereas syn-chronized rats showed a lower ethanol intake during estrus(Roberts et al., 1998). Here, we demonstrate that (1) therats continue their respective individual cycles throughoutthe experiment and (2) the rats within as well as betweeneach experimental group are in separate individual cycles.This is in accord with the findings of Lau et al. (1996) andalso shows that the maternal separation-induced effects onethanol intake presented herein were not secondary to ahormonal influence.
In our previous experiments, using the same experimen-tal protocol, no differences in body weight have been foundbetween the groups during the period of access to ethanol(Gustafsson et al., 2005; Ploj et al., 2003a; Roman et al.,2003, 2004). In the current study, no differences in bodyweight were observed on PND 22, but during the period ofaccess to ethanol, differences occurred in both males andfemales. Worth noting is that the differences in body weightwere not due to alterations in food intake, because onlyminor changes in food intake were found. Differences inbody weight could potentially influence the ethanol intakeresults because the intake is presented as g/kg/day. How-ever, similar differences between groups were obtained forboth males and females when comparing the ethanol intakeas g/day (data not shown) instead of g/kg/day. Therefore,the differences in body weight had no influence on thedifferences in ethanol intake presented herein.
In conclusion, the current results provide evidence thatenvironmental manipulations early in life can affect thegenetic high ethanol intake in adult ethanol-preferring AArats. Maternal separation affects adult voluntary ethanolintake differently in male and female AA rats, giving fur-ther evidence of sex-dependent effects of maternal separa-tion. In ANA rats, with an inherent low ethanol intake, nomaternal separation-induced effects on ethanol intakecould be seen, either in males or in females. The effectsinduced by daily maternal separation are therefore depen-dent on both the genetic propensity for high ethanol intakeand sex.
ACKNOWLEDGMENTS
The authors thank Ms. Marita Berg, Uppsala University, Upp-sala, Sweden, for technical assistance; Dr. Johan Lindbäck, Stat-istician, Uppsala Clinical Research Center, University Hospital,
MATERNAL SEPARATION AND ETHANOL INTAKE IN AA AND ANA RATS 599

Uppsala, Sweden, for advice in statistical matters; and Ms. MaijaSarviharju, National Public Health Institute, Helsinki, Finland,for assistance with the mating and transportation of the AA andANA rats.
REFERENCES
Andersen SL (2003) Trajectories of brain development: Point of vulner-ability or window of opportunity? Neurosci Biobehav Rev 27:3–18.
Badishtov BA, Overstreet DH, Kashevskaya OP, Viglinskaya IV,Kampov-Polevoy AB, Seredenin SB, Halikas JA (1995) To drink or notto drink: Open field behavior in alcohol-preferring and -nonpreferringrat strains. Physiol Behav 57:585–589.
Barr CS, Newman TK, Becker ML, Champoux M, Lesch KP, Suomi SJ,Goldman D, Higley JD (2003) Serotonin transporter gene variation isassociated with alcohol sensitivity in rhesus macaques exposed to early-life stress. Alcohol Clin Exp Res 27:812–817.
Barr CS, Newman TK, Lindell S, Shannon C, Champoux M, Lesch KP,Suomi SJ, Goldman D, Higley JD (2004) Interaction between serotonintransporter gene variation and rearing condition in alcohol preferenceand consumption in female primates. Arch Gen Psychiatry 61:1146–1152.
Brown KJ, Grunberg NE (1995) Effects of housing on male and femalerats: Crowding stresses male but calm females. Physiol Behav 58:1085–1089.
Cailhol S, Mormede P (2001) Sex and strain differences in ethanol drink-ing: Effects of gonadectomy. Alcohol Clin Exp Res 25:594–599.
Carlson M, Earls F (1997) Psychological and neuroendocrinological se-quelae of early social deprivation in institutionalized children in Roma-nia. Ann NY Acad Sci 807:419–428.
Caspi A, McClay J, Moffitt TE, Mill J, Martin J, Craig IW, Taylor A,Poulton R (2002) Role of genotype in the cycle of violence in mal-treated children. Science 297:851–854.
Caspi A, Moffitt TE, Morgan J, Rutter M, Taylor A, Arseneault L, TullyL, Jacobs C, Kim-Cohen J, Polo-Tomas M (2004) Maternal expressedemotion predicts children’s antisocial behavior problems: Usingmonozygotic-twin differences to identify environmental effects on be-havioral development. Dev Psychol 40:149–161.
Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H,McClay J, Mill J, Martin J, Braithwaite A, Poulton R (2003) Influenceof life stress on depression: Moderation by a polymorphism in the5-HTT gene. Science 301:386–389.
Champagne FA, Francis DD, Mar A, Meaney MJ (2003) Variations inmaternal care in the rat as a mediating influence for the effects ofenvironment on development. Physiol Behav 79:359–371.
Crabbe JC, Phillips TJ (2004) Pharmacogenetic studies of alcohol self-administration and withdrawal. Psychopharmacology (Berl) 174:539–560.
Dick DM, Foroud T (2003) Candidate genes for alcohol dependence: Areview of genetic evidence from human studies. Alcohol Clin Exp Res27:868–879.
Eriksson CJP (1973) Ethanol and acetaldehyde metabolism in rat strainsgenetically selected for their ethanol preference. Biochem Pharmacol22:2283–2292.
Eriksson K (1968) Genetic selection for voluntary alcohol consumption inthe albino rat. Science 159:739–741.
Eriksson K (1971) Rat strains specially selected for their voluntary alcoholconsumption. Ann Med Exp Biol Fenn 49:67–72.
Fahlke C, Eriksson CJP, Hård E (1993) Audiogenic immobility reactionand open-field behavior in AA and ANA rat lines. Alcohol 10:311–315.
Fahlke C, Hård E, Thomasson R, Engel JA, Hansen S (1994)Metyrapone-induced suppression of corticosterone synthesis reducesethanol consumption in high-preferring rats. Pharmacol Biochem Be-hav 48:977–981.
Fahlke C, Lorenz JG, Long J, Champoux M, Suomi SJ, Higley JD (2000)Rearing experiences and stress-induced plasma cortisol as early risk
factors for excessive alcohol consumption in nonhuman primates. Al-cohol Clin Exp Res 24:644–650.
Ford MM, Eldridge JC, Samson HH (2002) Microanalysis of ethanolself-administration: Estrous cycle phase-related changes in consump-tion patterns. Alcohol Clin Exp Res 26:635–643.
Francis DD, Young LJ, Meaney MJ, Insel TR (2002) Naturally occurringdifferences in maternal care are associated with the expression ofoxytocin and vasopressin (V1a) receptors: Gender differences. J Neu-roendocrinol 14:349–353.
Gianoulakis C (2001) Influence of the endogenous opioid system on highalcohol consumption and genetic predisposition to alcoholism. J Psy-chiatry Neurosci 26:304–318.
Gilmer WS, McKinney WT (2003) Early experience and depressive dis-orders: Human and non-human primate studies. J Affect Disord 75:97–113.
Gordon HW (2002) Early environmental stress and biological vulnerabil-ity to drug abuse. Psychoneuroendocrinology 27:115–126.
Gustafsson L, Ploj K, Nylander I (2005) Effects of maternal separation onvoluntary ethanol intake and brain peptide systems in female Wistarrats. Pharmacol Biochem Behav, in press.
Hall FS (1998) Social deprivation of neonatal, adolescent, and adult ratshas distinct neurochemical and behavioral consequences. Crit Rev Neu-robiol 12:129–162.
Higley JD, Hasert MF, Suomi SJ, Linnoila M (1991) Nonhuman primatemodel of alcohol abuse: Effects of early experience, personality, andstress on alcohol consumption. Proc Natl Acad Sci USA 88:7261–7265.
Hilakivi L, Eriksson CJP, Sarviharju M, Sinclair JD (1984) Revitalizationof the AA and ANA rat lines: Effects on some line characteristics.Alcohol 1:71–75.
Hilakivi-Clarke LA, Turkka J, Lister RG, Linnoila M (1991) Effects ofearly postnatal handling on brain beta-adrenoceptors and behavior intests related to stress. Brain Res 542:286–292.
Holdstock L, de Wit H (2000) Effects of ethanol at four phases of themenstrual cycle. Psychopharmacology (Berl) 150:374–382.
Holm S (1979) A simple sequentially rejective multiple test procedure.Scand J Stat 6:65–70.
Huot RL, Thrivikraman KV, Meaney MJ, Plotsky PM (2001) Develop-ment of adult ethanol preference and anxiety as a consequence ofneonatal maternal separation in Long Evans rats and reversal withantidepressant treatment. Psychopharmacology (Berl) 158:366–373.
Hyytiä P, Schulteis G, Koob GF (1996) Intravenous heroin and ethanolself-administration by alcohol-preferring AA and alcohol-avoidingANA rats. Psychopharmacology (Berl) 125:248–254.
Jaworski JN, Francis DD, Kuhar MJ (2004) Variations in maternal sep-aration affect ethanol intake in adult rats. Program No. 572.2. Societyfor Neuroscience, San Diego.
Juarez J, Barrios de Tomasi E (1999) Sex differences in alcohol drinkingpatterns during forced and voluntary consumption in rats. Alcohol19:15–22.
Kalinichev M, Easterling KW, Holtzman SG (2001) Repeated neonatalmaternal separation alters morphine-induced antinociception in malerats. Brain Res Bull 54:649–654.
Kalinichev M, Easterling KW, Plotsky PM, Holtzman SG (2002) Long-lasting changes in stress-induced corticosterone response and anxiety-like behaviors as a consequence of neonatal maternal separation inLong-Evans rats. Pharmacol Biochem Behav 73:131–140.
Kim-Cohen J, Moffitt TE, Caspi A, Taylor A (2004) Genetic and envi-ronmental processes in young children’s resilience and vulnerability tosocioeconomic deprivation. Child Dev 75:651–668.
Knapp DJ, Kampov-Polevoy AB, Overstreet DH, Breese GR, RezvaniAH (1997) Ultrasonic vocalization behavior differs between lines ofethanol-preferring and nonpreferring rats. Alcohol Clin Exp Res 21:1232–1240.
Korpi ER, Sinclair JD, Kaheinen P, Viitamaa T, Hellevuo K, Kiianmaa K(1988) Brain regional and adrenal monoamine concentrations and be-havioral responses to stress in alcohol-preferring AA and alcohol-avoiding ANA rats. Alcohol 5:417–425.
600 ROMAN ET AL.

Ladd CO, Huot RL, Thrivikraman KV, Nemeroff CB, Meaney MJ,Plotsky PM (2000) Long-term behavioral and neuroendocrine adapta-tions to adverse early experience. Prog Brain Res 122:81–103.
Lancaster FE, Spiegel KS (1992) Sex differences in pattern of drinking.Alcohol 9:415–420.
Langbehn DR, Cadoret RJ, Caspers K, Troughton EP, Yucuis R (2003)Genetic and environmental risk factors for the onset of drug use andproblems in adoptees. Drug Alcohol Depend 69:151–167.
Langeland W, Draijer N, van den Brink W (2004) Psychiatric comorbidityin treatment-seeking alcoholics: The role of childhood trauma andperceived parental dysfunction. Alcohol Clin Exp Res 28:441–447.
Langeland W, Hartgers C (1998) Child sexual and physical abuse andalcoholism: A review. J Stud Alcohol 59:336–348.
Lau C, Klinefelter G, Cameron AM (1996) Reproductive developmentand functions in the rat after repeated maternal deprivation stress.Fundam Appl Toxicol 30:298–301.
Lehmann J, Feldon J (2000) Long-term biobehavioral effects of maternalseparation in the rat: Consistent or confusing? Rev Neurosci 11:383–408.
Lehmann J, Pryce CR, Bettschen D, Feldon J (1999) The maternalseparation paradigm and adult emotionality and cognition in male andfemale Wistar rats. Pharmacol Biochem Behav 64:705–715.
Marmendal M, Roman E, Eriksson CJP, Nylander I, Fahlke C (2004)Maternal separation alters maternal care, but has minor effects onbehavior and brain opioid peptides in adult offspring. Dev Psychobiol45:140–152.
McIntosh J, Anisman H, Merali Z (1999) Short- and long-periods ofneonatal maternal separation differentially affect anxiety and feeding inadult rats: gender-dependent effects. Brain Res Dev Brain Res 113:97–106.
Moore CL, Morelli GA (1979) Mother rats interact differently with maleand female offspring. J Comp Physiol Psychol 93:677–684.
Möller C, Wiklund L, Thorsell A, Hyytiä P, Heilig M (1997) Decreasedmeasures of experimental anxiety in rats bred for high alcohol prefer-ence. Alcohol Clin Exp Res 21:656–660.
Nemeroff CB (1998) The neurobiology of depression. Sci Am 278:42–49.Newport DJ, Stowe ZN, Nemeroff CB (2002) Parental depression: Animal
models of an adverse life event. Am J Psychiatry 159:1265–1283.Overstreet DH, Halikas JA, Seredenin SB, Kampov-Polevoy AB, Viglin-
skaya IV, Kashevskaya O, Badishtov BA, Knapp DJ, Mormede P,Kiianmaa K, Li TK, Rezvani AH (1997) Behavioral similarities anddifferences among alcohol-preferring and -nonpreferring rats: Confir-mation by factor analysis and extension to additional groups. AlcoholClin Exp Res 21:840–848.
Papaioannou A, Dafni U, Alikaridis F, Bolaris S, Stylianopoulou F(2002a) Effects of neonatal handling on basal and stress-induced mono-amine levels in the male and female rat brain. Neuroscience 114:195–206.
Papaioannou A, Gerozissis K, Prokopiou A, Bolaris S, Stylianopoulou F(2002b) Sex differences in the effects of neonatal handling on theanimal’s response to stress and the vulnerability for depressive behav-iour. Behav Brain Res 129:131–139.
Park MK, Hoang TA, Belluzzi JD, Leslie FM (2003) Gender specificeffect of neonatal handling on stress reactivity of adolescent rats. J Neu-roendocrinol 15:289–295.
Ploj K, Roman E, Nylander I (2002) Effects of maternal separation onbrain nociceptin/orphanin FQ peptide levels in male Wistar rats. Phar-macol Biochem Behav 73:123–129.
Ploj K, Roman E, Nylander I (2003a) Long-term effects of maternalseparation on ethanol intake and brain opioid and dopamine receptorsin male Wistar rats. Neuroscience 121:787–799.
Ploj K, Roman E, Nylander I (2003b) Long-term effects of short and longperiods of maternal separation on brain opioid peptide levels in maleWistar rats. Neuropeptides 37:149–156.
Pryce CR, Feldon J (2003) Long-term neurobehavioural impact of thepostnatal environment in rats: Manipulations, effects and mediatingmechanisms. Neurosci Biobehav Rev 27:57–71.
Roberts AJ, Smith AD, Weiss F, Rivier C, Koob GF (1998) Estrous cycleeffects on operant responding for ethanol in female rats. Alcohol ClinExp Res 22:1564–1569.
Roman E, Hyytiä P, Nylander I (2003) Maternal separation alters acqui-sition of ethanol intake in male ethanol-preferring AA rats. AlcoholClin Exp Res 27:31–37.
Roman E, Ploj K, Nylander I (2004) Maternal separation has no effect onvoluntary ethanol intake in female Wistar rats. Alcohol 33:31–39.
Roth ME, Cosgrove KP, Carroll ME (2004) Sex differences in the vul-nerability to drug abuse: A review of preclinical studies. NeurosciBiobehav Rev 28:533–546.
Salimov RM, McBride WJ, Sinclair JD, Lumeng L, Li T (1996) Perfor-mance in the cross-maze and slip funnel tests of four pairs of rat linesselectively bred for divergent alcohol drinking behavior. Addict Biol1:273–280.
Sandbak T, Murison R, Sarviharju M, Hyytiä P (1998) Defensive buryingand stress gastric erosions in alcohol-preferring AA and alcohol-avoiding ANA rats. Alcohol Clin Exp Res 22:2050–2054.
Schumann G, Spanagel R, Mann K (2003) Candidate genes for alcoholdependence: Animal studies. Alcohol Clin Exp Res 27:880–888.
Sinclair JD (1974) Rats learning to work for alcohol. Nature 249:590–592.Sinclair JD, Le AD, Kiianmaa K (1989) The AA and ANA rat lines,
selected for differences in voluntary alcohol consumption. Experientia45:798–805.
Sinclair JD, Li TK (1989) Long and short alcohol deprivation: Effects onAA and P alcohol-preferring rats. Alcohol 6:505–509.
Spak F, Allebeck P, Spak L, Thundal KL (2001) Gothenburg study ofwomen and alcohol: Problems during childhood and youth an importantrisk factor. Lakartidningen 98:1109–1114.
Tuominen K, Hilakivi LA, Päivärinta P, Korpi ER (1990) Behavior ofalcohol-preferring AA and alcohol-avoiding ANA rat lines in tests ofanxiety and aggression. Alcohol 7:349–353.
van der Harst JE, Baars AM, Spruijt BM (2003) Standard housed rats aremore sensitive to rewards than enriched housed rats as reflected by theiranticipatory behaviour. Behav Brain Res 142:151–156.
Vengeliene V, Siegmund S, Singer MV, Sinclair JD, Li TK, Spanagel R(2003) A comparative study on alcohol-preferring rat lines: Effects ofdeprivation and stress phases on voluntary alcohol intake. Alcohol ClinExp Res 27:1048–1054.
Viglinskaya IV, Overstreet DH, Kashevskaya OP, Badishtov BA,Kampov-Polevoy AB, Seredenin SB, Halikas JA (1995) To drink or notto drink: Tests of anxiety and immobility in alcohol-preferring andalcohol-nonpreferring rat strains. Physiol Behav 57:937–941.
Weinberg J (1987) Effects of early experience on responsiveness to eth-anol: A preliminary report. Physiol Behav 40:401–406.
Wigger A, Neumann ID (1999) Periodic maternal deprivation inducesgender-dependent alterations in behavioral and neuroendocrine re-sponses to emotional stress in adult rats. Physiol Behav 66:293–302.
MATERNAL SEPARATION AND ETHANOL INTAKE IN AA AND ANA RATS 601
Related Documents











