Int. J. Environment and Pollution, Vol. 33, No. 4, 2008 401 Copyright © 2008 Inderscience Enterprises Ltd. Shellfish culture in Chile Daniel A. López*, Boris A. López and María L. González Departamento de Acuicultura y Recursos Acuáticos, Universidad de Los Lagos, Avda. Fuchslocher 1305, Osorno, Chile E-mail: [email protected] E-mail: [email protected] E-mail: [email protected] *Corresponding author Abstract: The present situation and future projections regarding shellfish culture in Chile are analysed. Production has increased exponentially, mainly of the ‘Chilean mussel’ Mytilus chilensis (Hupe) and the ‘northern scallop’ Argopecten purpuratus (Lamarck). At present, seven species are cultured, two of which were introduced. The total invertebrate production corresponding to cultures has increased from 1.6% to 38% over the last 15 years. The effects of the following factors on present and future levels of shellfish culture in Chile are discussed: disease, fouling, borers, red tide and environmental variations caused by the cultures themselves. Potential culture of approximately 70 commercially endemic species not only constitutes an economic option, but also favours their conservation. Keywords: shellfish; culture technologies; Chile; environmental effects of aquaculture; diversification; scallop; oyster; mussel. Reference to this paper should be made as follows: López, D.A., López, B.A. and González, M.L. (2008) ‘Shellfish culture in Chile’, Int. J. Environment and Pollution, Vol. 33, No. 4, pp.401–431. Biographical notes: Daniel A. López holds a PhD in biology. He is a Professor and Ex-Principal of the Universidad de Los Lagos, Osorno, Chile. He is the author of more than 50 publications in ISI journals on ecological aspects of aquatic resources (marine and fresh water), and aquaculture. In addition, he is a member of the national study committee and the postgraduate commission on animal health and production pertaining to the national scientific research commission, maximum organisation for the promotion, management and funding of science in Chile, and a member of the national aquaculture commission. Boris A. López graduated in marine biology and professor at the Universidad de Los Lagos. He is involved in research and is co-author of publications in the area of cirriped behaviour and culture strategies for marine invertebrates. María L. González has an MSc, and is currently a Professor at the Universidad de Los Lagos, Osorno, Chile. She specialises in aquatic invertebrate ecophysiology and has studied the metabolic responses and energetic balance of aquatic organisms under different environmental conditions. She is the Director of research projects evaluating the quality of coastal and underground waters in southern Chile. She has published more than 35 publications in 15 specialised journals.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Int. J. Environment and Pollution, Vol. 33, No. 4, 2008 401
Copyright © 2008 Inderscience Enterprises Ltd.
Shellfish culture in Chile
Daniel A. López*, Boris A. López and María L. González Departamento de Acuicultura y Recursos Acuáticos, Universidad de Los Lagos, Avda. Fuchslocher 1305, Osorno, Chile E-mail: [email protected] E-mail: [email protected] E-mail: [email protected] *Corresponding author
Abstract: The present situation and future projections regarding shellfish culture in Chile are analysed. Production has increased exponentially, mainly of the ‘Chilean mussel’ Mytilus chilensis (Hupe) and the ‘northern scallop’ Argopecten purpuratus (Lamarck). At present, seven species are cultured, two of which were introduced. The total invertebrate production corresponding to cultures has increased from 1.6% to 38% over the last 15 years. The effects of the following factors on present and future levels of shellfish culture in Chile are discussed: disease, fouling, borers, red tide and environmental variations caused by the cultures themselves. Potential culture of approximately 70 commercially endemic species not only constitutes an economic option, but also favours their conservation.
Keywords: shellfish; culture technologies; Chile; environmental effects of aquaculture; diversification; scallop; oyster; mussel.
Reference to this paper should be made as follows: López, D.A., López, B.A. and González, M.L. (2008) ‘Shellfish culture in Chile’, Int. J. Environment and Pollution, Vol. 33, No. 4, pp.401–431.
Biographical notes: Daniel A. López holds a PhD in biology. He is a Professor and Ex-Principal of the Universidad de Los Lagos, Osorno, Chile. He is the author of more than 50 publications in ISI journals on ecological aspects of aquatic resources (marine and fresh water), and aquaculture. In addition, he is a member of the national study committee and the postgraduate commission on animal health and production pertaining to the national scientific research commission, maximum organisation for the promotion, management and funding of science in Chile, and a member of the national aquaculture commission.
Boris A. López graduated in marine biology and professor at the Universidad de Los Lagos. He is involved in research and is co-author of publications in the area of cirriped behaviour and culture strategies for marine invertebrates.
María L. González has an MSc, and is currently a Professor at the Universidad de Los Lagos, Osorno, Chile. She specialises in aquatic invertebrate ecophysiology and has studied the metabolic responses and energetic balance of aquatic organisms under different environmental conditions. She is the Director of research projects evaluating the quality of coastal and underground waters in southern Chile. She has published more than 35 publications in 15 specialised journals.

402 D.A. López et al.
1 Introduction
Shellfish culture in Chile was initiated in 1848, with the introduction of the normative governing Aquaculture Maritime Concessions. In 1857, ‘exclusive privileges’ were conceded for the cultivation of ‘Chilean oysters’, Ostrea chilensis Philippi in Valparaíso (33°S) and Chiloé (42ºS). Subsequently in 1943 the Ministry of Agriculture installed a mitilid culture centre in Putemún, Chiloé in order to study and conserve the ‘giant mussel’, Choromytilus chorus (Molina) (Hancock, 1969).
The semi-intensive suspended systems used to develop the majority of invertebrate cultures in Chile were first introduced in 1967, in the Island of Talcán, Chiloé; during the same period rafts were installed in Mejillones Bay in the north of the country (23º06’S) and in Tubildad, Chiloé. The seventies witnessed the large-scale installation of rafts and long-lines, mainly in southern Chile (Winter et al., 1984). This was part of the ‘Chilean oyster’ breeding and marketing development programme in Chiloé that permitted the construction of oyster and mitilid breeding centres in several locations throughout southern Chile, with the support of Spanish, French, and Japanese experts, as well as the Food and Agricultural Organisation of the United Nations (FAO).
Oysters and mitilids were produced during this decade. However there are no reliable statistics available on production levels. In 1982 the National Fisheries Service began to include aquaculture production levels in its statistics. Up to 1987 only harvest data was compiled, subsequently more specific, technical aspects were also recorded. In 1991 the General Fisheries and Aquaculture Law was passed, which, for the first time, incorporated normatives regulating the development of aquaculture activities in Chile. With the approval of a National Aquaculture Policy in 2003, conditions for promoting economic growth through Aquaculture were established which ensured environmental sustainability and fair criteria regulating Aquaculture activities.
From 1982 onwards, the harvests of nine species have been recorded: Aulacomya ater (Molina) ‘ribbed mussel’; Mytilus chilensis (Hupé) ‘Chilean mussel’; Choromytilus chorus (Molina) ‘giant mussel’; Argopecten purpuratus (Lamarck) ‘northern scallop’; Ostrea chilensis Philippi ‘Chilean oyster’; Crassostrea gigas (Thunberg) ‘Pacific oyster’; Haliotis rufescens (Swainson) ‘red abalone’; Haliotis discus hannai Ino ‘Pacific abalone’ and Loxechinus albus (Molina) ‘sea urchin’. Over the last 10 years, shellfish production in culture centres has increased exponentially. Between 1982 and 1992 approximately 25,000 ton were produced; in contrast, between 1992 and 2003 production levels reached approximately 420,000 tons (Sernapesca, 1982–2003). The development of shellfish aquaculture activities in Chile presents various challenges for the future, such as: the diversification of production by cultivating introduced species; the development of commercial cultures for approximately a dozen endemic species; innovative commercialisation strategies aimed at diversifying markets that will signify improved quality and an increase in the quantity of products produced, as well as insertion into foreign markets; increasing aggregate value; modifying restrictive norms to facilitate production and commercialisation; agreements regulating competition in relation to other activities undertaken in the coastal zone; greater requirements with regard to levels of scientific knowledge, principally with regard to physiological, genetic and biotechnological aspects, intensive production technology, hygiene and food handling techniques (Ministry of Economy and Energy, 2003). This study analyses the present state and trends of shellfish culture in Chile. Particular emphasis is placed on
Shellfish culture in Chile 403
diversification using native species and information required for the further development of present cultures systems.
2 Production trends
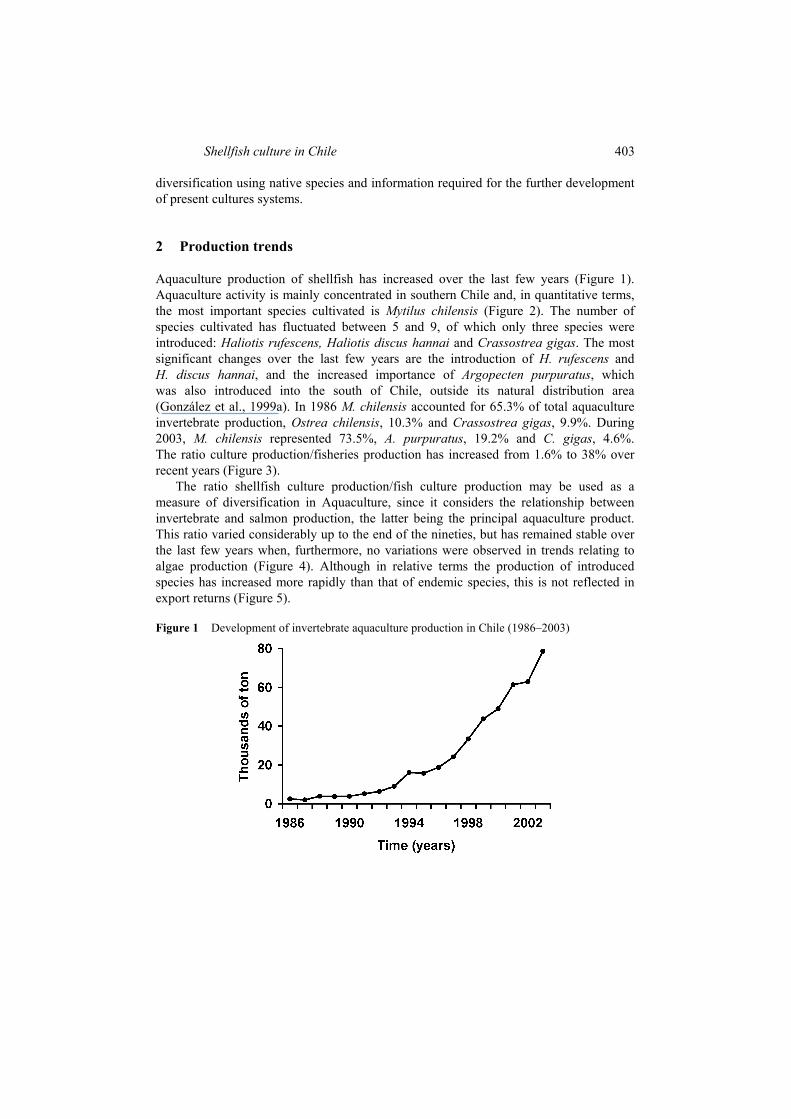
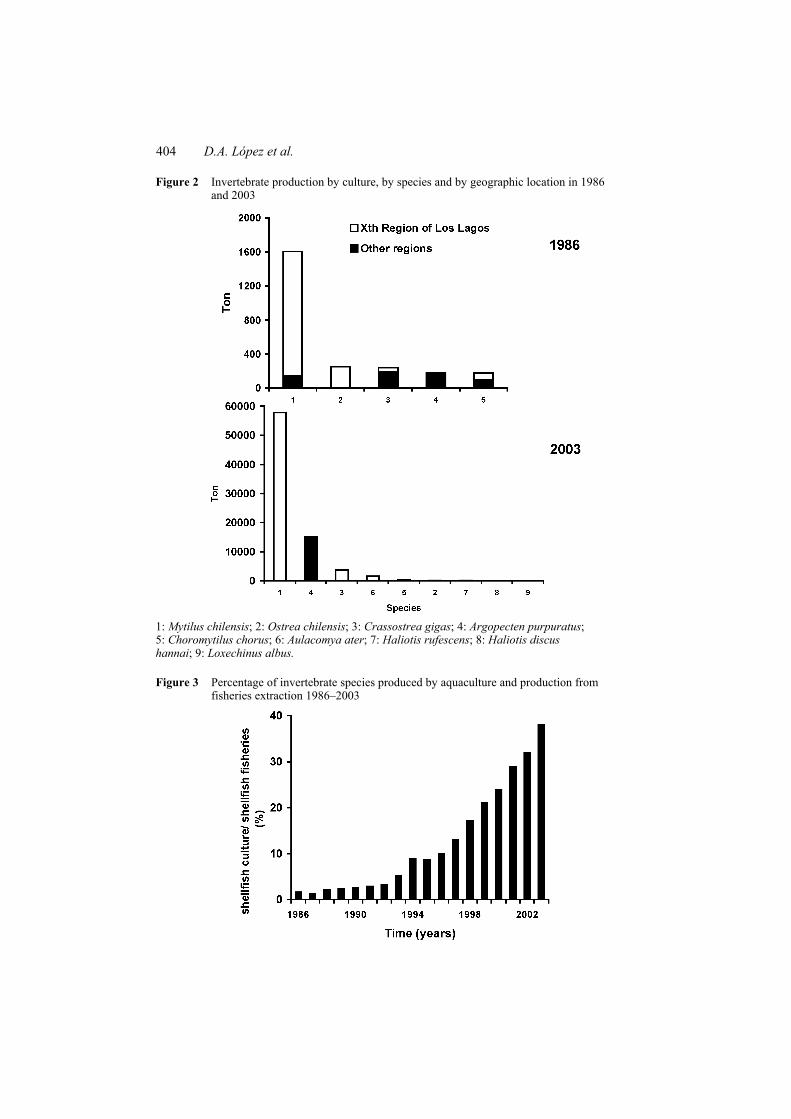
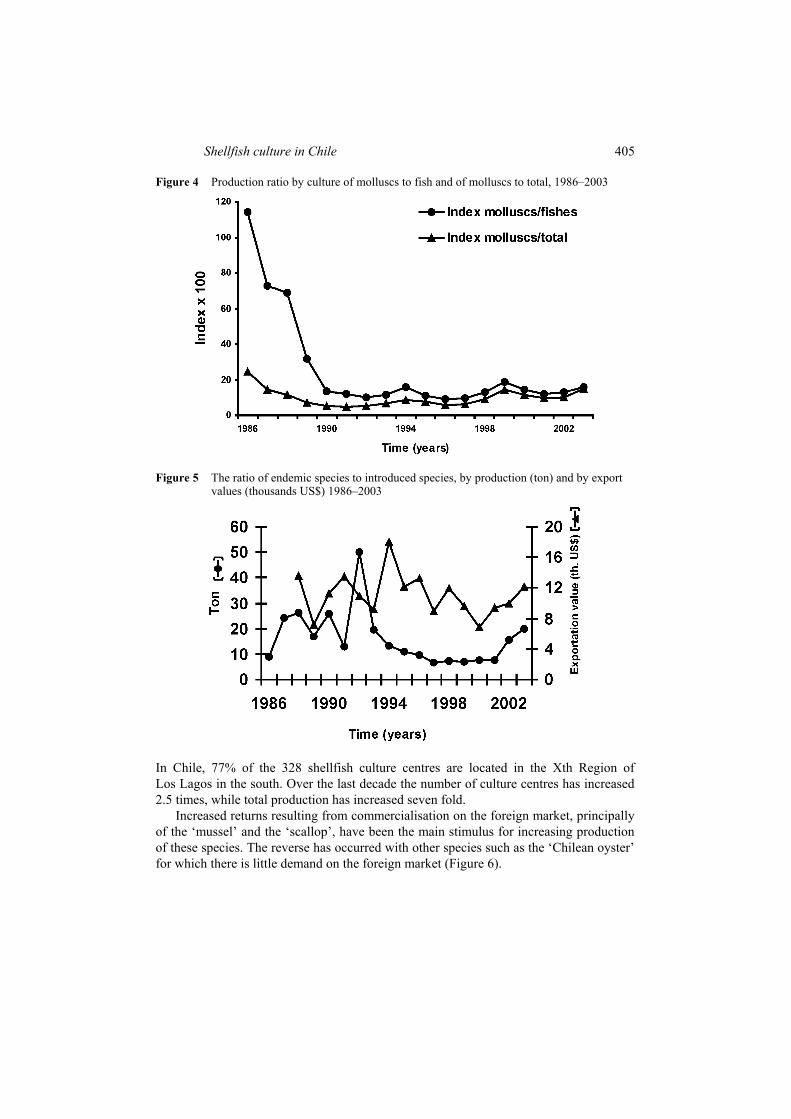
Aquaculture production of shellfish has increased over the last few years (Figure 1). Aquaculture activity is mainly concentrated in southern Chile and, in quantitative terms, the most important species cultivated is Mytilus chilensis (Figure 2). The number of species cultivated has fluctuated between 5 and 9, of which only three species were introduced: Haliotis rufescens, Haliotis discus hannai and Crassostrea gigas. The most significant changes over the last few years are the introduction of H. rufescens and H. discus hannai, and the increased importance of Argopecten purpuratus, which was also introduced into the south of Chile, outside its natural distribution area (González et al., 1999a). In 1986 M. chilensis accounted for 65.3% of total aquaculture invertebrate production, Ostrea chilensis, 10.3% and Crassostrea gigas, 9.9%. During 2003, M. chilensis represented 73.5%, A. purpuratus, 19.2% and C. gigas, 4.6%. The ratio culture production/fisheries production has increased from 1.6% to 38% over recent years (Figure 3).
The ratio shellfish culture production/fish culture production may be used as a measure of diversification in Aquaculture, since it considers the relationship between invertebrate and salmon production, the latter being the principal aquaculture product. This ratio varied considerably up to the end of the nineties, but has remained stable over the last few years when, furthermore, no variations were observed in trends relating to algae production (Figure 4). Although in relative terms the production of introduced species has increased more rapidly than that of endemic species, this is not reflected in export returns (Figure 5).
Figure 1 Development of invertebrate aquaculture production in Chile (1986–2003)
404 D.A. López et al.
Figure 2 Invertebrate production by culture, by species and by geographic location in 1986 and 2003
1: Mytilus chilensis; 2: Ostrea chilensis; 3: Crassostrea gigas; 4: Argopecten purpuratus; 5: Choromytilus chorus; 6: Aulacomya ater; 7: Haliotis rufescens; 8: Haliotis discus hannai; 9: Loxechinus albus.
Figure 3 Percentage of invertebrate species produced by aquaculture and production from fisheries extraction 1986–2003
Shellfish culture in Chile 405
Figure 4 Production ratio by culture of molluscs to fish and of molluscs to total, 1986–2003
Figure 5 The ratio of endemic species to introduced species, by production (ton) and by export values (thousands US$) 1986–2003
In Chile, 77% of the 328 shellfish culture centres are located in the Xth Region of Los Lagos in the south. Over the last decade the number of culture centres has increased 2.5 times, while total production has increased seven fold.
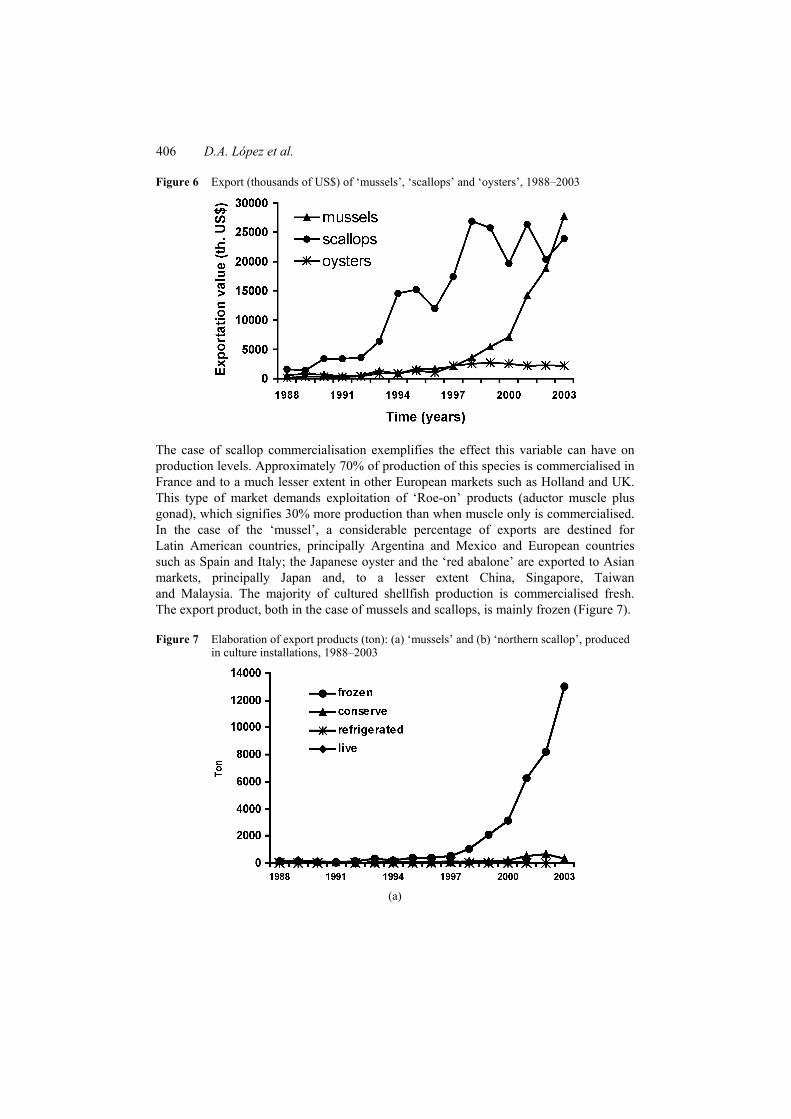
Increased returns resulting from commercialisation on the foreign market, principally of the ‘mussel’ and the ‘scallop’, have been the main stimulus for increasing production of these species. The reverse has occurred with other species such as the ‘Chilean oyster’ for which there is little demand on the foreign market (Figure 6).
406 D.A. López et al.
Figure 6 Export (thousands of US$) of ‘mussels’, ‘scallops’ and ‘oysters’, 1988–2003
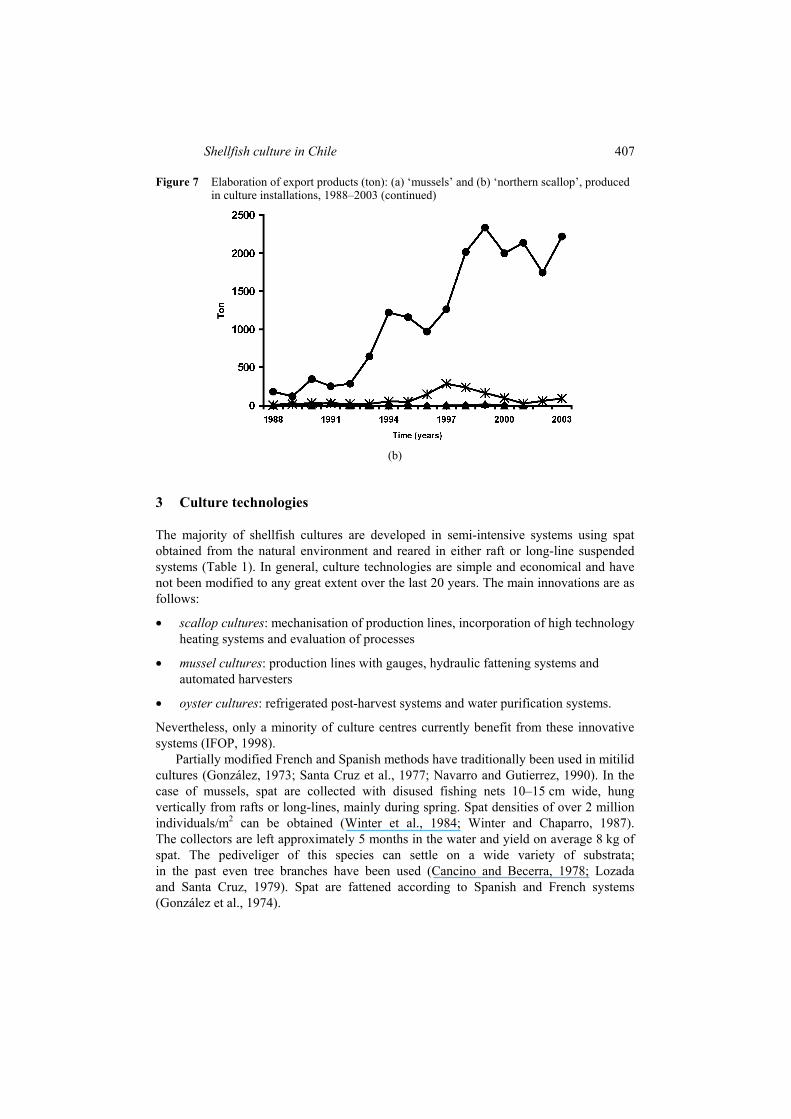
The case of scallop commercialisation exemplifies the effect this variable can have on production levels. Approximately 70% of production of this species is commercialised in France and to a much lesser extent in other European markets such as Holland and UK. This type of market demands exploitation of ‘Roe-on’ products (aductor muscle plus gonad), which signifies 30% more production than when muscle only is commercialised. In the case of the ‘mussel’, a considerable percentage of exports are destined for Latin American countries, principally Argentina and Mexico and European countries such as Spain and Italy; the Japanese oyster and the ‘red abalone’ are exported to Asian markets, principally Japan and, to a lesser extent China, Singapore, Taiwan and Malaysia. The majority of cultured shellfish production is commercialised fresh. The export product, both in the case of mussels and scallops, is mainly frozen (Figure 7).
Figure 7 Elaboration of export products (ton): (a) ‘mussels’ and (b) ‘northern scallop’, produced in culture installations, 1988–2003
(a)
Shellfish culture in Chile 407
Figure 7 Elaboration of export products (ton): (a) ‘mussels’ and (b) ‘northern scallop’, produced in culture installations, 1988–2003 (continued)
(b)
3 Culture technologies
The majority of shellfish cultures are developed in semi-intensive systems using spat obtained from the natural environment and reared in either raft or long-line suspended systems (Table 1). In general, culture technologies are simple and economical and have not been modified to any great extent over the last 20 years. The main innovations are as follows:
• scallop cultures: mechanisation of production lines, incorporation of high technology heating systems and evaluation of processes
• mussel cultures: production lines with gauges, hydraulic fattening systems and automated harvesters
• oyster cultures: refrigerated post-harvest systems and water purification systems.
Nevertheless, only a minority of culture centres currently benefit from these innovative systems (IFOP, 1998).
Partially modified French and Spanish methods have traditionally been used in mitilid cultures (González, 1973; Santa Cruz et al., 1977; Navarro and Gutierrez, 1990). In the case of mussels, spat are collected with disused fishing nets 10–15 cm wide, hung vertically from rafts or long-lines, mainly during spring. Spat densities of over 2 million individuals/m2 can be obtained (Winter et al., 1984; Winter and Chaparro, 1987). The collectors are left approximately 5 months in the water and yield on average 8 kg of spat. The pediveliger of this species can settle on a wide variety of substrata; in the past even tree branches have been used (Cancino and Becerra, 1978; Lozada and Santa Cruz, 1979). Spat are fattened according to Spanish and French systems (González et al., 1974).

408 D.A. López et al.
Table 1 Shellfish culture technologies in Chile
Spat collection Growth
Environment Environment
Species Suspended
systems Sea bed Hatchery Suspended
systems Sea bed In shore
‘Northern scallop’ X X X X X – ‘Chilean mussel’ X – – X – – ‘Ribbed mussel’ X – – X – – ‘Giant mussel’ X – – X – – ‘Chilean oyster X – – X – – ‘Pacific oyster’ – – X X – – ‘Red abalone’ – – X X – X ‘Pacific abalone’ – – X X – X ‘Sea urchin’ – – X X – –
Clusters of spat reach a commercial size of 5–6 cm during the fattening stage in 12–14 months; harvest production fluctuates between 20–50 ton per raft.
The same culture technologies used for the ‘mussel’ are applied with the ‘ribbed mussel’ and the ‘giant mussel’ (Lozada et al., 1974; Santa Cruz, 1976; Aracena, 1983; Winter et al., 1984). ‘Ribbed mussels’ are harvested between 14 and 24 months. ‘Giant mussel’ cultures use collectors that are placed above banks with densities of 11,000 individuals/m2. High mortality rates in specimens over 5 cm can be recorded as a result of detachment and for this reason, specimens are transferred to trays or net bags. The culture period depends on the harvest size required, for example specimens reach 12 cm over a period of 3.7 years (Lozada and Santa Cruz, 1979).
In the case of the ‘Chilean oyster’, the entire supply of spat is collected in the natural environment, mainly between November and January. ‘Necklace’ type collectors are used, where 30–40 ‘ribbed mussel’ or ‘scallop’ valves are joined together by a wire 1.5 m long, and placed in packs in suspended systems. Each of these collectors can produce between 200–400 spat. Plastic plaques in packs are also used and can accumulate between 80 and 100 spat per plaque. The collectors are located close to breeding specimens, either in rafts or tanks, because the pelagic period of competent larvae is short – ranging from minutes to a few hours (Solis, 1973; Chanley and Dinamani, 1980; Toro and Chaparro, 1990). The collectors are left between 6–12 months in the water and are then transferred to suspended or sea bed fattening systems. Collectors open and are dispersed in raft or long-line suspended systems; spat can also adhere directly to polypropelene ropes. The growth period is 18 months for oysters of 5 cm (‘common variety’). However, export-sized specimens take over two years to reach the required size of 6.5 cm. Specimens that grow in suspended systems experience an adjustment stage on the sea bed to permit hardening of the valves (Ferreira, 1975; DiSalvo et al., 1983a, 1983b; DiSalvo and Martinez, 1985).
Scallop production in Chile adapts traditional Japanese technology. Suspended systems are used for the collection and fattening of spat (Illanes, 1988, 1989, 1990) in addition to the production of spat under controlled conditions (10% of production). Spat are obtained from the natural environment using collectors during the maximum

Shellfish culture in Chile 409
maturity period of spring/summer, mainly in December and January (DiSalvo et al., 1984; González et al., 2002). Yields of 300 specimens/collector are achieved. Collectors of 4 mm plastic mesh are placed inside 10 mm conical netlon nets, and suspended in groups of 15–20 along the long-line, for a period of 2–4 months. Specimens are harvested at 10–15 mm and transferred (initial culture) to pearl nets – pyramid-type structures of between 25–30 cm long with an aperture of 2.9 mm. These structures join together forming units of approximately 10 pearl nets. Lantern systems are also used. Lanterns, mesh bags or systems whereby specimens are suspended from their auricles, are used in the final culture stage, when specimens reach 90 mm. Approximately 50,000 scallops are maintained per long-line, reaching a marketable size in 14–16 months. In 55% of culture centres, fattening is undertaken on the sea bed using enclosures protected with nets. Spat production in a controlled environment is carried out using traditional methods: conditioning of breeding specimens; spawning induction; embryo and larvae culture in conical tanks, the latter being fed with commercial stocks of microalgae. Netlon nets are used as collectors in postlarvae cultures (Winkler and Estevez, 2003; Nevejan et al., 2003a).
Traditional methods are used for the culture of exotic species; in the case of the Japanese oyster, techniques described by Breese and Malouf (1975) are used to obtain spat in a controlled environment. Settlement takes place on substrata similar to scallop ‘valves’ over a period of approximately 2 weeks. Fattening up to the 90 mm marketable size is carried out in lanterns or mesh bags on long-lines in suspended systems and using ‘parrón’ (grapevine) techniques in sea bed systems (Hauer, 1988; Illanes, 1989; Hoyl and Uribe, 1990).
Three technology areas have been identified with regard to abalone culture: hatchery technology or spat production; sea growth technology and land-based growth technology. There are two alternative methods for spat production (Californian and Japanese) which differ in the form and size of tanks used, although results obtained with regard to production levels and costs involved are similar. Spawning is artificially induced in breeding specimens and larvae (short planktonic life) are maintained until settlement occurs. The metamorphosis of veliger larvae is induced using GABA and postlarvae are placed in tanks and fed with diatoms until reaching 4 mm (Ebert and Houk, 1984, 1989; Illanes, 1999). Subsequently, juveniles are transferred to cones until reaching approximately 10 mm over a period of around 10 months. Another option is to transfer the larvae to plaques and feed them with macroalgae until they reach 15–20 mm, (Owen et al., 1984). The critical period during spat production is the postlarval stage, between 3 weeks and 3 months, when mortality rates of over 85% are recorded (Uki and Kikuchi, 1984). The fattening stage in the sea is undertaken using ‘barrels’ or cages which vary in cost, density, degree of control and manual labour required. Growth in land-based culture centres takes place in tanks, facilitating culture practices such as instalation, management, supply, monitoring, control and harvest; culture densities are greater and risks (parasites, red tides, storms) are reduced in comparison to sea cultures; the use of artificial feed is also favoured. On the other hand, restrictions include the increased capital necessary for implementation and logistic factors (provision of constant water and air supplies) (Godoy et al., 1992). In Chile, priority areas in abalone culture include the need to develop economical commercial diets – at present macroalgae such as Macrocystis pyrifera and Gracilaria sp are being used; also the need for improved health and pathology control measures, both during the hatchery and growth stages (Godoy and Jerez, 1998).

410 D.A. López et al.
4 Environmental problems
The main problems affecting shellfish culture are the presence of encrusting species and borers, red tides and pathologies.
Encrusting species and valve borers can seriously affect production. A wide variety of epibiont flora and fauna are associated with shellfish cultures, principally: barnacles such as Austromegabalanus psittacus, Notobalanus flosculus and Elminius kingii; tunicates – mainly colonial species and Pyura chilensis and Nemidocarpa robinsonii; gastropods such as Crepidula sp.; the polichaetes Platynereis sp and Terebella sp; bryozoans, in particular Alcyonidium polyoum, and algae including bivalves, decapods, fish, echinoderms, sponges and hydrozoans (Aracena and López, 1973; Osorio et al., 1978). However, no quantitative evaluation of their effects on production has been undertaken.
Calcareous valves can be perforated by various organisms, the most common are: the polichaete Polydora sp, which attacks oysters and scallops (Oliva et al., 1986; Sato-Okoshi and Takatsuka, 2001) and Boccardia proboscidea; the mollusk Petricola rugosa and the phoronid Phoronis ovalis, frequent in scallop valves. Culture systems are also colonised by encrusting species, and the wooden structures perforated by the teredinid Bankia martensi.
Problems generated by micro-organisms and parasites present in marine invertebrates can be categorised as follows: a) diseases, generating mortalities, low growth rates, decreased egg and larvae production, castration or other types of direct and indirect negative effects; b) limitations to commercialisation, in particular with regard to external markets; c) human intoxication following consumption of infected organisms whereby the pathogen (zoonosis) is transmitted. This pathogen is caused by the accumulation of toxins in the tissue of organisms as a result of the proliferation of harmful microalgae (red tides).
Current knowledge about Chilean marine invertebrate pathologies is scarce, and for the most part related to commercially significant species (Carvajal, 1988). The subject matter of research and scientific work published focuses principally on the identification and description of parasites, mostly helminths, but does not consider the negative effects caused by these organisms on cultures. With regard to bacterial diseases Riquelme et al. (1995a), reported high Argopecten purpuratus larval mortality in hatcheries in the northern zone of Chile, caused by the pathogen Vibrio anguillarum, recording concentrations of 104–108 cells/ml. Similar results were obtained in further experiments, thus verifying the pathogenic nature of Pseudonomas sp, Moraxella sp, Aeromonas hydrophila and Vibrio alginolyticus (Riquelme et al., 1995b, 1996a). As a result of problems caused by these microorganisms, strains of native bacteria (Alteromonas haloplanktis and Vibrio sp) with the capacity to inhibit the effects of other harmful bacteria, have been isolated, thus decreasing the negative effects on survival rates of A. purpuratus larvae (Riquelme et al., 1996b, 2000; Jorquera et al., 2000, 2001). Garcia-Tello et al. (2002), detected the presence of bacteria in the hemolinfa. The effectiveness of antibiotics has also been tested, as has that of cloramfenicol on bacterial activity, with favourable results being recorded in northern scallop hatcheries (Uriarte et al., 2001). Lohrmann et al. (2002), reported the presence of rickettsia in adult scallops, with a higher infestation intensity in specimens from natural populations than in culture specimens. Studies on Ostrea chilensis show that the bacteria Arcobacter sp are an important part of the microflora associated with the digestive tract

Shellfish culture in Chile 411
(Romero et al., 2002); the presence of Vibrio anguillarium has been confirmed in the digestive tract of Mytilus chilensis (Vial et al., 1988). In both cases no antecedents are available on pathogenic characteristics. Similarly, Vibrio have been detected in the veliger larvae of Concholepas concholepas, but without detrimental effects. The presence of this pathogen has been also reported in the intestinal and gonadal tissues of adult specimens. Bacteria have also been identified in the sea urchin Loxechinus albus, Pseudomonas sp being most frequent, with no evidence of harmful effects on specimens at concentrations of 3.69 ± 0.48 bacteria per gram of specimen, (Gesche et al., 1978). Recently, the presence of Vibrio parahaemolyticus in shellfish caught in natural banks has caused health problems resulting from human consumption of these shellfish. This is attributed to the sudden, large scale proliferation of this type of bacteria in coastal waters during the summer (Córdova et al., 2002).
As regards viral disease, Enríquez et al. (1992), reported the presence of the hepatitis A virus in the hepatopancreas of Mytilus chilensis, revealing slow elimination of the pathogen from the mitilid. The authors propose the use of these organisms as bioindicators of this type of pathogen agent in the water. The herpes-type virus has been detected in Ostrea chilensis larvae, which would cause high rates of premature mortality (Hine et al., 1998).
In Chile, no information is available on diseases caused by fungi in marine invertebrates, although records exist in the northern hemisphere of larval micosis produced by the fungus Sirolpidium zoophtharium which affects the larvae of the Crassostrea virginica oyster and the Mercenaria mercenaria clam, stunting growth and causing gradual disintegration. Mortality as a result of this phenomenon is extremely high in larval cultures (Sindermann, 1977).
Different types of parasites have been described in invertebrate cultures. Oliva et al. (1986), compiled a list of parasites affecting the northern scallop A. purpuratus, including polichaetes of the Polydora sp genus, which bore into the shell, a herudine in the mantle and cestode larvae in the gonads. A protozoan ciliate (Trichodina sp) has also been identified on the branchia (gills) and digestive glands, being more frequent in cultured individuals than those originating from natural populations (Lohrmann et al., 2002).
The bonamiasis disease, produced by the protozoan Bonamia sp, has been studied in the ‘Chilean oyster’ introduced into New Zealand. This parasite is located inside the hemocytes, provoking hyperplasia in the mantle tissues, gills and digestive glands (Mix and Breese, 1980; Campalans et al., 2000; Hine, 1996, 2002; Hine et al., 2001, 2002; Cochennec-Laureans et al., 2003).
Carvajal (1988), proposed preventative measures to counteract the presence of pathologies in cultures, such as: maintaining the water quality by avoiding pollution; reducing environmental stress; ensuring adequate nutrition of culture organisms; developing vaccinations from stocks resistant to disease to be administered to culture specimens; controlling the possibility of disease introduced by exotic species; applying chemical prophylaxis.
Gonadal damage caused by parasites has been reported in molluscs with culture potential such as Mesodesma donacium ‘pink clams’, Fissurella spp ‘keyhole limpets’ and Concholepas concholepas ‘Chilean abalone’. In Mesodesma donacium, both larval and adult helminth (trematodes and cestodes) have been detected (Bahamonde and López, 1962; Carvajal, 1977; Campbell and Carvajal, 1979; Carvajal et al., 1982). Trematodes (Proctoeces sp), have been reported in gastropods of the Fissurella genus (keyhole limpets) which cause castration, directly affecting the reproductive potential of

412 D.A. López et al.
specimens (Jirón, 1979; Bretos and Jirón, 1980; George-Nascimento and Quiroga, 1983; Osorio et al., 1986; Bretos and Chihuailaf, 1993). The prevalence and intensity of infestation in these molluscs depends both on the geographic distribution of the species (Oliva and Huaquin, 2000), and on its biological characteristics (George-Nascimento et al., 1998). Evidence of castration has also been recorded in Concholepas concholepas as a result of a digenean larval stage (Oliva et al., 1999).
There are no published records on disease in introduced species cultured in Chile. However, bacterial diseases have been identified in the Japanese oyster (Sugumar et al., 1998a, 1998b; Nakamura et al., 1999) and the presence of bacteria and protozoans has been recorded in the ‘abalone’ (Ebert and Houk, 1984; Bower, 1986; Moore et al., 2000, 2001, 2002; Llenart et al., 2003).
The presence of harmful and toxic microalgae in the southern austral zone of Chile has been reported since the early 1970s, (Guzmán and Campodónico, 1975, 1978; Avaria, 1979; Lembeye, 1981; Clément and Pantoja, 1983). Although their effect on shellfish culture has not been studied, information is available from fish cultures (Clément and Lembeye, 1993) and from the incidence of human mortality resulting from consumption of invertebrates originating from natural banks (Avaria, 1989; Lagos, 1998). Negative effects of toxic microalgae on invertebrates have also been recorded in other locations (Smolowitz and Shumway, 1997; Bauder et al., 2001). For this reason phytoplanktonic blooms constitute a potentially negative factor for the culture of filtering species.
The appearance of cases of human intoxication prompted research into the species responsible and the types of toxins that provoke this kind of symtomatology. The dinoflagellate Alexandrium catenella has been cited as the originating agent of the Paralytic Shellfish Poison (PSP), which secretes a saxitoxin – one of the strongest non proteic substances known. (Guzmán and Campodónico, 1978; Andriolo et al., 2002) which adheres to and blocks the Na+ channels in the nerve cells, and can cause death due to respiratory failure (Evans, 1969; Montebruno, 1993). Other isolated compounds in species of the Dinophysis genus are okadaic acid and dinophysistoxins which produce the denominated Diarrhetic Shellfish Poison (DSP), which inhibits phosphatase activity in enzymes associated with important cell reactions, such as water permeability and electrolytes in intestinal epithelium (Zhao et al., 1993; Rivas et al., 2000; Uribe et al., 2001; Garcia et al., 2003). Furthermore, this type of toxin is associated with the appearance of tumours (Garcia et al., 2003). Other diseases reported over the last few years in Chile are those provoked by a diatom of the Pseudonitzschia genus which produces domoic acid and generates the Amnesic Shellfish Poison (ASP) (Araya and Villarroel, 1995; Córdova et al., 2003). Additional phytoplankton species have also been reported, such as cyanobacteria, which also produce toxins (Lagos, 1998) and the role of certain bacteria associated with dinoflagellates in the production of toxins is also under study (Vásquez et al., 2001, 2002).
As a result of the mortalities and at least 300 cases of intoxication associated with the appearance of these phenomena, phytoplankton monitoring and vigilance programmes are being undertaken in the southern austral zone of Chile, in addition to bivalve mollusc health programmes regulating export products. At present the public health authorities periodically analyse samples of bivalves, in particular mitilids (Aulacomya ater and Mytilus chilensis), in order to determine toxin levels. When levels exceed the permitted norm (80 µg/100 g of meat), temporary bans on shellfish extraction are imposed. To date, toxins have been detected in an area from the Seno del Reloncavi (41°S) to

Shellfish culture in Chile 413
Magallanes (56°S); subtoxic levels have been observed in Coquimbo (29°S) (Cortés and Herrera, 1996).
Studies have not only concentrated on filtering molluscs, tissues from secondary consumers have also been analysed, such as the gastropods Concholepas concholepas and Argobuccinum ranelliformes, revealing high levels of paralytic poison in viscera and foot muscle 5 or 6 months after the initial appearance of the phytoplanktonic bloom (Compagnon et al., 1998). Other studies have attempted to determine the effect of culture activities on the appearance of algal blooms, caused by the direct incorporation of nutrients (nitrate and phosphate) into the water column. No association, however, was established (Arzul et al., 1996, 2001).
Research related to these aspects tends to concentrate on the following areas: evaluation and control of encrusting and boring species; detection, monitoring and health measures related to red tides; detection and prevalence of pathologies.
5 Effects of cultures on the environment
Shellfish culture on a large scale produces waste, faeces and ammonium (González et al., 1991) which can affect the water column. Furthermore, in some cases species are introduced (Godoy and Jerez, 1998). Knowledge regarding the effects of shellfish cultures in Chile is related mainly to faecal production and biodeposition (López et al., 1989). Little is known about other aspects (Table 2). Jaramillo et al. (1992) reported that mitilids can be an important source of biodeposition through faeces and pseudo-faeces. López et al. (1988) reported a greater quantity of organic material under the mitilid culture rafts than in the control areas. Under laboratory conditions, González et al. (1991) determined that faecal production rates in several species of invertebrates, none of which are cultured commercially, can vary according to temperature and feeding regime. Extrapolation of this data to the natural habitat would suggest a high production of organic material. The effects of culture biodeposition can be limited (López et al., 1989). Variations do not correlate with the abundance of foraminifers. However, the abundance of three species of crabs increases (López et al., 1988).
Table 2 Knowledge regarding the effects of shellfish culture on the environment
1 2 3
Effects on the water column – – – Nutrient recycling producing changes in coastal productivity – – X Phytoplankton variations – – X Phytoplankton variations in relation to red tides – X – Nitrogen and carbon variations – – X Effects on sediments – – – Biodeposition X – – Changes in chemical composition – X – Physical changes – X – Biological changes – X –

414 D.A. López et al.
Table 2 Knowledge regarding the effects of shellfish culture on the environment (continued)
1 2 3 Changes in populations and biological communities – – – Predator density – X – Populational changes associated with culture practices – X – Infaunal diversity – X – Environmental impact of introduced species – X – Habitat and natural population modification – – X Mitigating by mixed cultures – X –
1: Detailed antecedents. 2: Basic antecedents. 3: No antecedents.
In Chile, the ammonium produced by shellfish cultures has not been measured in situ. Nevertheless, in mixed cultures the addition of nitrogen compounds favours the growth of a red alga in cultures (Cancino et al., 1987; Cancino and Orellana, 1987).
Only limited antecedents are available on the effects of introduced species, such as the red abalone and the Japanese oyster (Buschmann et al., 1996).
In Chile, there are two well-documented cases of species conservation resulting from aquaculture activities (López et al., 1989). The ‘giant mussel’ (Choromytilus chorus) and the ‘Chilean oyster’ (Ostrea chilensis) are endemic to the Chilean coast, and were the most exploited species at the beginning of the 20th century. In 1926, when shellfish and fish landings totalled 20,000 ton, extraction of the ‘giant mussel’ and ‘Chilean oyster’ was recorded at 3,688 and 816 ton, respectively (Reyes, 1986). Fisheries extraction of the ‘giant mussel’ reached a maximum between 1938 and 1944, with an average of 1,700 ton/year. During the period 1945–1986, landings of this species were marginal, averaging only 190 ton/year, presumably due to over-exploitation of natural banks, which occasioned an indefinite fishing ban (Hancock, 1969; Lozada and Santa Cruz, 1979). Extraction during this period was sporadic and production was almost entirely limited to culture centres; culture of this species began in 1965 in Putemún in southern Chile. Cultures of ‘giant mussel’ have used juveniles obtained directly from the natural environment which has probably increased recruitment in areas where these juveniles are cultured. At present, 42% of production of the ‘giant mussel’, between 400–600 ton/year, is produced in culture centres.
The case of the ‘Chilean oyster’ is even more dramatic. Half way through the 20th century, exploitation of this species totalled 420 ton/year. However, production decreased steadily from 1952 onwards and the situation was aggravated by the 1960 earthquake in southern Chile. This led to an intensification of the culture activities initiated in 1943, with the creation of ‘oyster culture’ installations in Pullinque, Chiloé; the main function of this centre was to create a reserve of breeding specimens which would make it possible to obtain juveniles both for production and repopulation purposes (Hancock, 1969). During the period 1967–1980, practically 100% of oyster production was provided by culture centres, with the creation of new centres in southern Chile (Solis and Eberhard, 1979). At present fisheries extraction has increased relatively, reaching an average of 350 ton/year, although the great majority of supplies (approximately 93%) still originate from culture centres (Figure 8).
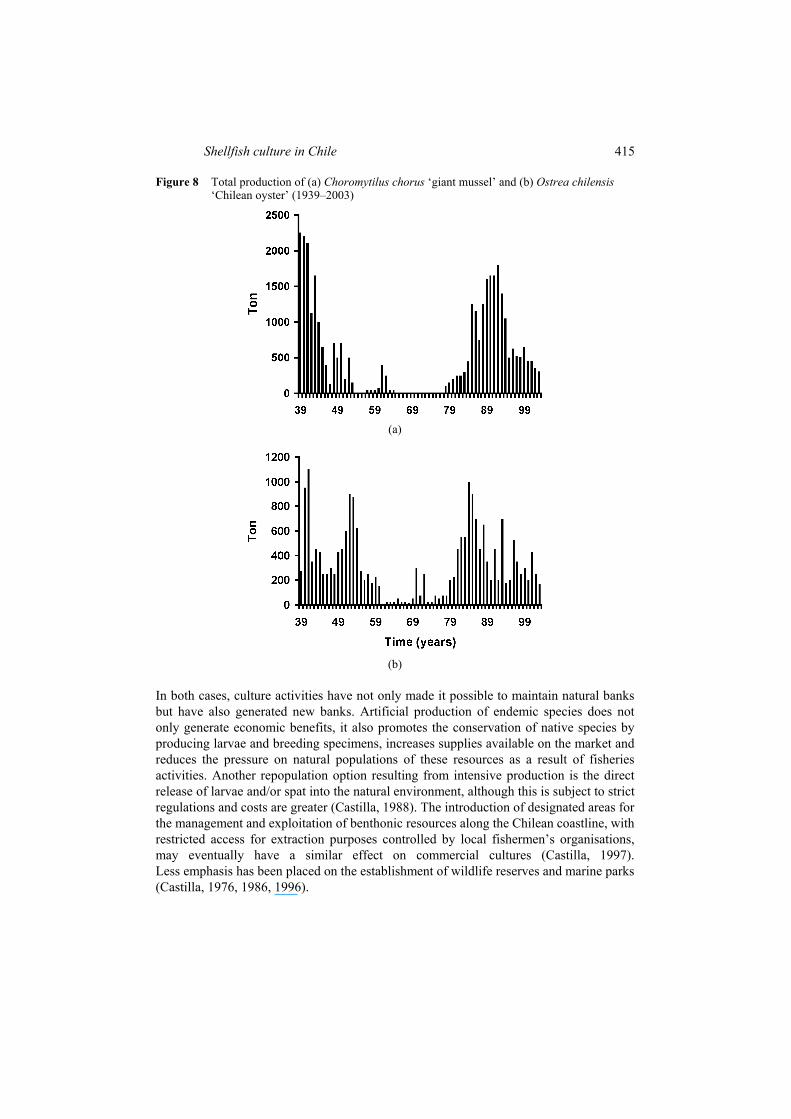
Shellfish culture in Chile 415
Figure 8 Total production of (a) Choromytilus chorus ‘giant mussel’ and (b) Ostrea chilensis ‘Chilean oyster’ (1939–2003)
(a)
(b)
In both cases, culture activities have not only made it possible to maintain natural banks but have also generated new banks. Artificial production of endemic species does not only generate economic benefits, it also promotes the conservation of native species by producing larvae and breeding specimens, increases supplies available on the market and reduces the pressure on natural populations of these resources as a result of fisheries activities. Another repopulation option resulting from intensive production is the direct release of larvae and/or spat into the natural environment, although this is subject to strict regulations and costs are greater (Castilla, 1988). The introduction of designated areas for the management and exploitation of benthonic resources along the Chilean coastline, with restricted access for extraction purposes controlled by local fishermen’s organisations, may eventually have a similar effect on commercial cultures (Castilla, 1997). Less emphasis has been placed on the establishment of wildlife reserves and marine parks (Castilla, 1976, 1986, 1996).

416 D.A. López et al.
6 Normative aspects
Over the last few years, several new norms have been established regulating Aquaculture activities. For the most part they address environmental issues. Several legal bodies are responsible for regulating shellfish culture activities. The General Fisheries and Aquaculture Law (revised text D.S. 430, 1991), establishes general regulatory measures regarding the conservation of hydrobiological resources applied to fisheries extraction, aquaculture, research and sports activities in terrestrial and inner waters, territorial waters and exclusive economic zones. The General Environmental Norms Law (1994) establishes environmental policies and environmental management instruments. Several decrees regulate specific culture aspects: Decree No. 290 (1993) legislates on concessions and authorisations for culture centres; Decree No. 320 (2001) or Environmental Regulations for Aquaculture, covers aspects related to conservation, evaluation of capacities, operational requirements, distance between culture centres and contingency plans. In particular, norms have been developed governing hygiene in shellfish culture techniques and the destination of surplus spat not used for fattening. Decree No. 404 (2003) establishes contents and methodologies for undertaking preliminary site characterisation and obtaining environmental information. This will depend on the centre category, determined in terms of intensity level, production volume, depth and type of sea bed. Decree No. 319 (2001) or Sanitary Regulation establishes measures governing the protection, control and eradication of high risk diseases in hydrobiological species. In the case of exotic species, there are at least three legal instruments which regulate cultures: Decree No. 730 (1995) governs the internment of species imported for the first time and is complemented by Decree No. 96 (1996) which controls procedures for importing hydrobiological species. Finally, Decree No. 626 (2001), dictates norms regulating sanitary certification of imported hydrobiological species. At present, with the approval of the National Aquaculture Policy (2003), various deficiencies or shortcomings have been identified in the legal framework which regulates the development of this activity, as a result of which new guidelines have been proposed to promote decentralisation and the simplification of administrative processes and decisions. These include: – redefinition of procedures regulating initiation of activities in the aquaculture field; facilitating procedures to be followed; reinforcement and creation of control and inspection processes which guarantee that current norms are respected.
7 Evolution and information required
Information published on species cultured at present is heterogeneous. As a result of the post-nineties increase in scallop production, culture technologies have improved (Winkler et al., 1993; Toro et al., 1995a; Uriarte et al., 1996a; Ortiz et al., 2002). Antecedents are available on growth (Mendo and Jurado, 1993; González et al., 1999a; López et al., 2000; Avendaño et al., 2001; Caers et al., 2003) and feeding (Avendaño and Riquelme, 1999; Navarro et al., 2000; González et al., 2002; Palma-Fleming et al., 2002). The species has also been introduced into the south of Chile, outwith its natural distribution area. (Uriarte et al., 1996b; Gonzalez et al., 1999a). However, these improvements are mainly in the areas of reproduction and spat collection (Uriarte and Farias, 1999; Aguilar and Stotz, 2000; Martinez et al., 2000; Avendaño et al., 2001; Farias and Uriarte, 2001; González et al., 2002; Encomendero and Dupré, 2003;

Shellfish culture in Chile 417
Nevejan et al., 2003b; Winkler and Estévez, 2003). 0n the contrary, there are very few recent publications dealing with reproductive aspects of the Chilean oyster, most of them referring to the introduction of this species into New Zealand (Jeffs, 1998, 1999; Jeffs and Hickman, 2000; Jeffs et al., 2002); even fewer records exist with respect to growth and other culture aspects (Toro, 1991; Toro et al., 1992, 1995b). For this reason, information is based mostly on classic studies (Basulto et al., 1967; Padilla and Orrego, 1967; Padilla et al., 1969; Solis, 1973; Winter, 1978). With regard to mitilids, again, little has been published after the 90s: in the case of Mytilus chilensis, publications are associated with growth and feeding (Toro and Sastre, 1995; Toro and Paredes, 1996; Velasco and Navarro, 2002, 2003); for Choromytilus chorus, areas covered include reproductive aspects (Davis and Moreno, 1995; Bellolio et al., 1996), growth and feeding (Navarro and Gutierrez, 1990) and for Aulacomya ater, growth (Navarro and Gutierrez, 1990) and reproductive aspects (Jaramillo and Navarro, 1995; Hernández and Becerra, 1997; Lizarralde, 1998).
Few studies have been published with regard to introduced species. In the case of Haliotis rufescens studies have concentrated on its introduction into Chilean waters and large-scale culture prospects (Owen et al., 1984; Godoy et al., 1992; Godoy and Jerez, 1998). Some evaluation has been made of reproduction and growth in Crassostrea gigas, (Gras et al., 1992; González et al., 1999c; Caers et al., 2000a), in addition to a description of technologies and yield (Hoyl and Uribe, 1990; Caers et al., 2000b).
The type of information required is directly related to the principle limitations affecting shellfish culture production, which include
• ‘Northern scallop’. Problems with spat supplies (low availability of spat from natural banks, the effect of pollution on spat collection and high production costs associated with controlled environments); high production costs in comparison to other invertebrates; culture technology which does not reach standards achieved in other parts of the world; presence of encrusting species and valve borers that affect production; loss of product yield depending on the export market targeted.
• ‘Chilean mussel’. Competition with the product derived from fisheries extraction; presence of encrusting species and borers in culture systems that affect production; lack of selection programs, management and improvement of breeding stocks; products limited mainly to Latin American markets; production processes not vertically integrated.
• ‘Giant mussel’. Commercialisation problems resulting from fluctuating demand and prices and limited supplies; slow growth rate; weak byssus, making the use of trays obligatory during the final stage of the fattening process, which raises production costs; presence of encrusting species and borers in culture systems that affect production; lack of selection programs, management and improvement of breeding stocks and larval settlement processes in the culture systems.
• ‘Ribbed mussel’. Export options on the external market are limited; existence of sexual dimorphism, although not externally visible, does produce considerable variations in product valuation; limited collection of natural spat; large size of first maturity.
418 D.A. López et al.
• ‘Chilean oyster’. Low production levels; slow growth; low fertility levels; difficulties in sea-bed culture due to the presence of predators; limited collection of natural spat; presence of encrusting species and valve borers that affect production; low commercialisation prospects on the external market.
• ‘Pacific oyster’. Presence of encrusting species and borers which affect the final appearance of the product; no collection of natural spat, which raises costs of artificial production; low demand in the internal market.
• ‘Red abalone’ and ‘Pacific abalone’. Lack of manufactured, commercial artificial diets; need for the provision of macroalgae for the diet; provision of spat; no collection of natural spat, which raises costs of artificial production; low growth rates in comparison to other species; deficient knowledge of diseases; more efficient fattening systems in the sea.
• ‘Sea urchin’. Mass production of juveniles; low growth rates; need to optimise feed.
8 Production diversification
The number of invertebrate species cultured is still only a fraction of the total number of commercially viable invertebrate species currently extracted on a small-scale by local fishermen, even though the ratio shellfish culture production/shellfish fisheries production has tended to increase exponentially. There are approximately 70 shellfish species, some of which, such as species of the Fissurella genus, are part of multispecific fisheries activities (Bretos, 1988). Production levels of only two of these species exceed 10,000 ton/year. The majority vary between 8,000 and 6,000 ton/year. A smaller group register production levels of less than 1,000 ton. Advances on an experimental culture have been made in approximately 16 species (Table 3).
Table 3 Small-scale fisheries extraction (ton) of commercially significant marine invertebrates. The vernacular name of each species is given in Spanish and English. Species which have experienced significant changes during the study period are marked in bold
Groups Species 1986 2003 Loxechinus albus ‘erizo’ (sea urchin)* 37,000 42,600 Protothaca thaca y Venus antiqua ‘almejas’ (clams)* 25,000 8,400 Mesodesma donacium ‘macha’ (pink clam)* 11,500 2,300
Group 1: most extracted species
Dosidicus gigas ‘jibia’ (cuttlefish) – 15,100 Aulacomya ater ‘cholga’ (ribbed mussel)** 9,700 3,200 Mytilus chilensis ‘chorito’ (Chilean mussel)** 9,300 2,800 Ensis macha ‘huepo’ (razor clam)* – 4,200 Cancer spp and Homalaspis plana ‘jaibas’ (crabs)* 4,000 5,000 Gari solida ‘culengue’ (clam)* 1,100 4,200 Tagelus dombeii ‘navajuela’ (hard razor clam)* 2,300 3,400 Pleuroncodes monodon ‘langostino colorado’ (squat lobster) 6,000 1,300 Cervimunida johni ‘langostino amarillo’ (squat lobster) 5,400 2,200
Group 2: intermediate level of extraction
Lithodes santolla ‘centolla’ (king crab)* 2,500 3,000
Shellfish culture in Chile 419
Table 3 Small-scale fisheries extraction (ton) of commercially significant marine invertebrates. The vernacular name of each species is given in Spanish and English. Species which have experienced significant changes during the study period are marked in bold (continued)
Groups Species 1986 2003 Concholepas concholepas ‘loco’ (Chilean abalone)* 6,300 3,000 Semele solida ‘tumbao’ (clam)* – 2,400 Fissurella spp ‘lapas’ (keyhole limpet)* 2,100 2,600 Octopus mimus and Enteroctopus megalocyathus ‘pulpo’ (octopus)*
2,500 2,000
Pyura chilensis ‘piure’ (sea squirt) 4,500 1,000 Heterocarpus reedi ‘camarón nailón’ (shrimp) 3,000 3,600 Thais chocolata ‘caracol locate’ (top shell whelk)* 8,300 900 Austromegabalanus psittacus ‘picoroco’ (giant barnacle)* 1,000 200
Group 2: intermediate level of extraction
Chorus giganteus ‘caracol trumulco’ (top shell whelk)* 2,000 100 Paralomis granulosa ‘centollón’ (king crab) 900 3,000 Ostrea chilensis ‘ostra chilena’ (chilean oyster)** 500 2 Tegula atra ‘caracol negro’ (turban snail) 500 11 Athyonidium chilensis ‘pepino de mar’ (sea cucumber)* – 300
Group 3: rare species
Choromytilus chorus ‘choro zapato’ (giant mussel)** 800 130
*Experimental cultures and/or biological research into potential cultures. **With commercial cultures.
Source: Sernapesca (1986, 2003)
Production levels of some of these species have dropped dramatically. This is the case of Mesodesma donacium, an infaunal species inhabiting sandy beaches, which forms banks easily accessed by fishermen and shore collectors (Jaramillo et al., 1994). At the end of the 1980s, landings of this species totalled over 17,000 ton/year, whereas today they barely exceed 1,000 ton/year. Extraction levels of the gastropod Chorus giganteus were first recorded in 1978, and reached over 2,000 ton in 1980, today only 23 ton are landed. In both cases, the drop in production was not the result of demand changes. The extraction of Concholepas concholepas is regulated following over-exploitation in response to increasing demands generated by the external market (Castilla, 1995; Castilla and Fernández, 1998; Leiva and Castilla, 2001). Extraction of the cirriped Austromegabalanus psittacus reached over 4,500 ton/year in the 1970s, decreasing substantially over the following years because fisheries activities related to this species have shifted to different latitudes, probably associated with over-exploitation practices (López et al., 2003b, 2003c).
In addition to using native species, diversification in Aquaculture has been also proposed in terms of species that could be introduced, such as Pecten maximum (‘European scallop’) and Penaeus vannamei (‘Ecuatorian shrimp’).
When considering species with culture potential, a determining factor related to the development of cultures on an industrial scale is the availability of pertinent information, given that the majority of these species are endemic. Fifty two percent of studies published on invertebrate species concentrate on reproductive aspects, 26% on growth and 15.5% on feeding, whereas only 6.1% of studies deal with culture technologies.

420 D.A. López et al.
34.7% of publications are on the ‘Chilean abalone’, 19.2% on the ‘sea urchin’ and the remainder spread over a variety of other species.
Technical reports and Congress communications (grey information) are an important source of information, but are not always easily accessible. In Marine Science-Chile Society congresses between 1990 and 2003 alone, over 353 studies on species with culture potential have been presented. The trend is the same as that detected in the publications. 40.8% of studies refer to reproductive aspects (fecundity, sexual maturity, larval development, larval settlement and recruitment), 31.7% to biological aspects (ecology, physiology, biochemistry, genetics, behaviour, pathologies), 13.9% to growth, 8.8% to fisheries and only 4.8% to culture technologies; 39.9% of studies deal with the ‘Chilean abalone’, 10.8% with ‘keyhole limpets’, which are a substitute of the ‘Chilean abalone’ and 9.3% with the ‘sea urchin’.
Prospects with regard to this group of species with culture potential can be analysed by examining several specific cases. The ‘Chilean abalone’ has been the most researched species because it is highly valued on the external market (López and Varela, 1988; Varela and López, 1989; Campos et al., 1994, Castilla, 1995). The culture of species of ‘keyhole limpets’ has also been studied. However, mainly from a biological perspectiva (Acuña, 1977; Bretos et al., 1988a, 1988b; Huaquin et al., 1998; González et al., 1999b; López et al., 2003a). Recently, the ‘giant barnacle’, because it is easily cultured and has favourable prospects as a luxuty gastronomic product (Arriagada et al., 2003; López et al., 2003b) and two species of ‘octopus’ have been the subject of feasibility studies in southern Chile (Iglesias et al., 1996; Cortez et al., 1999; López et al., 2004).
The development of new culture strategies for native species requires not only a study of market potential, but also the implementation of technologies to obtain spat and accelerating growth (López et al., 2006).
9 Conclusions
• Shellfish culture in Chile has concentrated on only a few species, two of which were introduced. The most significant changes that have occurred over the last few years are the relative increase in culture production in relation to production by fisheries extraction and the expansion of ‘northern scallop’ culture.
• The Information available on cultured species and further information required is heterogeneous, depending on the species. Technology adopted is traditional and spat collection and growth are mostly undertaken in suspended systems. The main environmental problems are the detection and prevalence of diseases, detection, monitoring and health measures with respect to red tides and the evaluation and control of fouling and boring. Knowledge with regard to the effect of cultures on the environment is scarce, particularly in the case of the water column. Nevertheless, antecedents are available on the positive contribution of culture practices to the conservation of populations of the ‘Chilean oyster’ and the ‘giant mussel’.
• There are numerous commercially viable endemic species with culture potential, which have been studied in terms of basic biological aspects. Greatest attention has been focussed on the ‘Chilean abalone’. However problems regarding spat production and the time required to reach a marketable size persist. The main limitations to ‘keyhole limpets’ and ‘octopus’ culture occur at the breeding stage.

Shellfish culture in Chile 421
The commercial projections of the crustacean ‘giant barnacle’ on the external market are unknown. Nevertheless, experimental cultures of all these species have been undertaken and studies have concentrated on adjustment to captivity, feeding and growth.
• The future challenges for shellfish culture development in Chile are associated with: diversification of production by cultivating endemic and exotic species; innovative commercialisation strategies aimed at diversifying markets; increasing aggregate value; modifying restrictive norms to facilitate production and commercialisation; agreements regulating competition in relation to other activities undertaken in the coastal zone, such as navigation or tourism; increased knowledge in the scientific and technological field, principally with regard to native species.
Acknowledgements
We are grateful to the organisers of the International Workshop ‘Aquaculture, Ecology and Economics towards a sustainable paradigm’ held between 9–11 December, 2003 in Puerto Varas, Chile, where the results of this study were presented. This Workshop was organised jointly by the Universidad de Los Lagos, Osorno, Chile and the University of Connecticut, USA. The encouragement and support received from Dr. Alejandro Buschmann and Dr. Boris Bravo-Ureta are particularly appreciated. The contribution of the Universidad de Los Lagos: research projects DIULA 5370 and 5380; FONDECYT (National Fund for Science and Technology): projects 1961302 and 1930040 and the Regional Government of the X ‘Los Lagos’ Region is also acknowledged. The collaboration of Susan Angus in the translation of the manuscript and contributions of María José Garrido, Juan Carvajal, David Friedli, José Uribe and Daniel C. López are also gratefully acknowledged. Finally, our thanks to the two anonymous referees for their valuable suggestions.
References Acuña, E. (1977) ‘Estudio preliminar de edad y crecimiento de Fissurella latemarginata (Sowerby,
1834) en Tocopilla, Chile. (Mollusca, Gastropoda, Fissurellidae)’, Revista de Biología Marina, Vol. 16, No. 2, pp.117–124.
Aguilar, M. and Stotz, W.B. (2000) ‘Settlement sites of juvenil scallops Argopecten purpuratus (Lamarck, 1819) in the subtidal zone at Puerto Aldea, Tongoy Bay, Chile’, Journal of Shellfish Research, Vol. 19, No. 2, pp.749–755.
Andriolo, D., Iglesias, V., García, C. and Lagos, N. (2002) ‘Toxicokinetics and toxicodynamics of gonyautoxins after an oral toxin dose in cats’, Toxicon, Vol. 40, No. 6, pp.699–709.
Aracena, O. and López, M. (1973) ‘Observaciones biológicas en organismos encontrados en sustratos artificiales. Caleta Leandro, Talcahuano. Chile’, V Congreso Lat. Zoología, pp.40–48.
Aracena, O.L. (1983) ‘El cultivo de Choromytilus chorus (Choro zapato) en Chile central y sur’, Mems. Asoc. Latinoam. Acuicultura, Vol. 5, No. 2, pp.231–240.
Araya, A. and Villarroel, O. (1995) ‘Determinación de ácido domoico en moluscos bivalvos’, Resúmenes VIII Jornadas Científicas, Instituto de Salud Pública, Chile, p.1.
Arriagada, S.E., López, B.A. and López, D.A. (2003) ‘Cultivo del cirripedio ‘picoroco’: un recurso potencial para Acuicultura’, Aquanoticias, Vol. 80, pp.63, 64.

422 D.A. López et al.
Arzul, G., Clément, A. and Pinier, A. (1996) ‘Effect on phytoplankton growth of dissolved substances produced by fish farming’, Aquatic Living Resources, Vol. 9, No. 2, pp.95–102.
Arzul, G., Seguel, M. and Clément, A. (2001) ‘Effect of marine animal excretions on differential growth of phytoplankton species’, Ices Journal of Marine Science, Vol. 58, No. 2, pp.386–390.
Avaria, S. (1979) ‘Red tide of the coast of Chile’, in Taylor, D.L. and Selinger, H.H. (Eds.): Toxic Dinoflagellate Blooms, Elsevier North-Holland Inc., New York, pp.161–164, 356.
Avaria, S. (1989) ‘Implicaciones del fenómeno de marea roja en la salud humana’, Revista Médica, Valparaíso Vol. 42, Nos. 1–4, pp 57–69.
Avendaño, R.E. and Riquelme, C.E. (1999) ‘Establishment on mixed culture probiotics and microalgae as food for bivalve larvae’, Aquaculture Research, Vol. 30, Nos. 11–12, pp.893–900.
Avendaño, R.E., Riquelme, C.E., Escribano, R. and Reyes, N. (2001) ‘Sobrevivencia y crecimiento post-larvas de Argopecten purpuratus (Lamarck, 1819) en Bahía Inglesa, Chile: efectos del origen, distribución en la bahía y bacterioflora larval’, Revista Chilena de Historia Natural, Vol. 74, No. 3, pp.669–679.
Bahamonde, N. and López, M.T. (1962) ‘Proboscidosacus mesodesmatis n. sp parásito de Mesodesma donacium Lamarck’, Invest. Zool., Chile, Vol. 8, pp.43–56.
Basulto, S., Orrego, J. and Solis, I. (1967) ‘Experiencia sobre crecimiento de ostras en Pullinque’, Biología Pesquera, Vol. 2, pp.83–87.
Bauder, A.G., Cembella, A.D., Bricelj, V.M. and Quilliam, M.A. (2001) ‘Uptake and fate of diarrhetic shellfish poisoning toxins from the dinoflagellate Prorocentrum lima in the bay scallop Argopecten irradians’, Marine Ecology Progress Series, Vol. 213, pp.39–52.
Bellolio, G.A., Toledo, P. and Dupré, M.E. (1996) ‘Larval development of Choromytilus chorus (Molina, 1782) reared in laboratory’, Scientia Marina, Vol. 60, Nos. 2–3, pp.353–360.
Bower, S.M. (1986) ‘The life cycle and ultrastructure of a new species of Thraustochytrid (Protozooa, Labryrinthomorpha) pathogenic to small abalone’, 2nd. Inter. Colloq. Pathol. Marine Aquac., Portugal, pp.7–11.
Breese, W.P. and Malouf, R.E. (1975) ‘Hatchery manual for the Pacific oyster’, Oregon State University Sea Grant College Program, Publication ORESU-H-75-002, pp.5–21.
Bretos, M. (1988) ‘Pesquería de lapas en Chile’, Medio Ambiente, Vol. 9, No. 2, pp.7–12. Bretos, M. and Chihuailaf, R.H. (1993) ‘Studies on the reproduction and gonadal parasites of
Fissurella pulchra (Gastropoda. Prosobranchia)’, Veliger, Vol. 36, No. 3, pp.245–251. Bretos, M. and Jirón, C. (1980) ‘Trematodes in chilean fissurellid mollusc’, Veliger, Vol. 22,
pp.280–293. Bretos, M., Gutierrez, J. and Espinoza, Z. (1988a) ‘Estudios biólogicos para el manejo de
Fissurella picta’, Medio Ambiente, Vol. 9, No. 1, pp.28–34. Bretos, M., Quintana V. and Ibarrola, V. (1988b) ‘Bases biológicas para el manejo de Fissurella
nigra’, Medio Ambiente, Vol. 9, No. 2, pp.55–62. Buschmann, A.H., López, D.A. and Medina, A. (1996) ‘A review of the environmental effects and
alternative production strategies of marine aquaculture in Chile’, Aquacultural Engineering, Vol. 15, No. 6, pp.397–421.
Caers, M., Coutteau, P. and Sorgeloos, P. (2000a) ‘Impact of starvation and of feeding algal and artificial diets on the lipid content and composition of juvenile oysters (Crassostrea gigas) and clams (Tapes philippinarum)’, Marine Biology, Vol. 136, No. 5, pp.891–899.
Caers, M., Coutteau, P. and Sorgeloos, P. (2000b) ‘Incorporation of different fatty acids, supplied as emulsions or liposomes, in the polar and neutral lipids of Crassostrea gigas spat’, Aquaculture, Vol. 186, Nos. 1–2, pp.157–171.

Shellfish culture in Chile 423
Caers, M., Coutteau, P., Sorgeloos, P. and Gajardo, G. (2003) ‘Impact of algal diets and emulsions on the fatty acid composition and content of selected tissues of adult broodstock of the Chilean Scallop Argopecten purpuratus (Lamarck, 1819)’, Aquaculture, Vol. 217, Nos. 1–4, pp.437–452.
Campalans, M., Rojas, P. and González, M. (2000) ‘Haemocytic parasitosis in the farmed oyster Tiostrea chilensis’, Bulletin of the European Association of Fish Pathologists, Vol. 20, No. 1, pp.31–33.
Campbell, R.A. and Carvajal, J. (1979) ‘Synonyny of the Phyllobothriid genera Rhodobothrium Linton, 1989, Inermiphyllidium Riser, 1955 and Sphaerobothrium Euzet, 1959 (Cestoda: Tetraphyllidea)’, Proc. Helminthol. Soc. Wast., Vol. 46, pp.88–97.
Campos, E.O., Pinto, A., Bustos, E., Rodríguez, S.R. and Inestrosa, N.C. (1994) ‘Metamorphosis of laboratory reared larvae of Concholepas concholepas (Mollusca, Gastropoda)’, Aquaculture, Vol. 126, Nos. 3–4, pp.299–303.
Cancino, J. and Becerra, R. (1978) ‘Antecedentes sobre la biología y tecnología del cultivo de Aulacomya ater (Molina, 1782) (Mollusca: Mytilidae)’, Biología Pesquera, Vol. 10, pp.27–45.
Cancino, J. and Orellana, M. (1987) ‘Interacción Gracilaria verrucosa y su fauna epifita.: perspectivas para la utilización de mitílidos como fuente de nutrientes en cultivos de Gracilaria en piscinas litorales’, in Verreth, J.A.J., carrillo, M., Zanuy, S. and Huisman, E.A. (Eds.): Investigaciones acuícolas en América, Pudoc Press, Wageningen, pp.380–392.
Cancino, J., Muñoz, M. and Orellana, M. (1987) ‘Effects of epifauna on algal growth and quatity of the agar produced by Gracilaria verrucosa (Hudson) Papenfuss’, Hydrobiologia, Vols. 151–152, pp.233–237.
Carvajal, J. (1977) ‘Description of the adult and larva of Caulobothrium myliobatidis sp. n. from Chile’, Journal of Parasitology, Vol. 63, pp.99–103.
Carvajal, J. (1988) ‘Patología de moluscos y repoblación’, Investación Pesquera, Vol. 35, pp.123–128.
Carvajal, J., Barros, C. and Santander, G. (1982) ‘In vitro culture of Rhodobothrium mesodesmatum parasite of a Chilean clam’, Proc. Helminthol. Soc. Wast., Vol. 49, pp.229, 230.
Castilla, J.C. (1976) ‘Parques y reservas marinas chilenas: necesidad de creación, probables localizaciones y criterios básicos’, Medio Ambiente, Vol. 2, pp.70–80.
Castilla, J.C. (1986) ‘Sigue existiendo la necesidad de establecer parques y reservas marítimas en Chile?’, Ambiente y Desarrollo, Vol. 2, pp.53–63.
Castilla, J.C. (1988) ‘La problemática de la repoblación de mariscos en Chile: diagnóstico, estrategias y ejemplos’, Investigación Pesquera, Vol. 35, pp.41–48.
Castilla, J.C. (1995) ‘The sustainability of natural resources as viewed by an ecologist and exemplief by the fishery of the mollusk Concholepas concholepas in Chile’, in Munasinghe, M. and Shearer, W. (Eds.): Defining and Measuring Sustainability, The International Bank for Recontruction and Development, The World Bank, Washington DC, USA, pp.153–159.
Castilla, J.C. (1996) ‘La futura Red Chilena de Parques y Reservas Marinas y los conceptos de conservación, preservación y manejo en la legislación nacional’, Revista Chilena de Historia Natural, Vol. 69, pp.253–270.
Castilla, J.C. (1997) ‘Chilean resources of benthic invertebrates: fishery, collapses, stock rebuilding and role of coastal management areas and national parks’, in Hancock, D.A., Smith, D.C., Grant, A. and Beumer, J.P. (Eds.): Second World Fisheries Congress, CSIRO, Australia, pp.130–135.
Castilla, J.C. and Fernández, M. (1998) ‘Small scale benthic fisheries in Chile: on co-management and sustainable use of the benthic invertebrates’, Ecological Applications, Vol. 8, No. 1, pp.S124–S132.

424 D.A. López et al.
Chanley, P. and Dinamani, P. (1980) ‘Comparative description of some oyster larvae from New Zealand and Chile, and description of a news genus of oyster, Tiostrea. New Zealand’, Journal of Marine Freshwater Research, Vol. 14, No. 2, pp.103–120.
Clément, A. and Lembeye, G. (1993) ‘Phytoplankton monitoring program in the fish farming region of South Chile’, in Smayda, T.J. and Shimuzi, Y. (Eds.): Toxic Phytoplankton Blooms in the Sea, Elsevier Science Publishers Co. Inc., pp.223–228.
Clément, A. and Pantoja, S. (1983) ‘Necesidad de conocimiento químico de los dinoflagelados tóxicos’, Simposio Fitoplancton Marino de Chile, Conocimiento Actual y Perspectivas. Montemar, Universidad de Valparaíso, Chile, pp.45, 46.
Cochennec-Laureau, N., Reece, K.S., Berthe, F.C.J. and Hine, P.M. (2003) ‘Mikrocytos roughleyi taxonomic affiliation leads to the genus Bonamia (Haplosporidia)’, Diseases of Aquatic Organisms, Vol. 54, No. 3, pp.209–271.
Compagnon, D., Lembeye, G., Marcos, N., Ruiz-Tagle, N. and Lagos, N. (1998) ‘Accumulation of paralytic shellfish poisoning toxins in the bivalve Aulacomya ater and two carnivorous gastropods Concholepas concholepas and Argobuccinum ranelliformes during an Alexandrium catenella bloom in southern Chile’, Journal of Shellfish Research, Vol. 17, No. 1, pp.67–73.
Córdova, J., Bustamante, J. and Müller, I. (2003) ‘Caracterización del primer cultivo de diatomea chilena productora de veneno amnésico’, Resúmenes XXIII Congreso de Ciencias del Mar, Universidad de Magallanes, Punta Arenas, Chile, p.107.
Córdova, J.L., Astorga, J., Silva, W. and Riquelme, C. (2002) ‘Characterization by PCR of Vibrio parahaemolyticus isolates collected during the 1997–1998 Chilean outbreak’, Biological Research, Vol. 35, pp.433–440.
Cortés, G. and Herrera, L. (1996) ‘Primer reporte de Veneno Paralítico de los Mariscos (VPM) en áreas de cultivo de la zona de Iquique, norte de Chile’, Resúmenes VI Symposium de Algas Marinas Chilenas y IV Encuentro de Microalgólogos, Puerto Montt, Chile, p.1.
Cortez, T., González, A.F. and Guerra, A. (1999) ‘Growth of cultured Octopus mimus (Cephalopoda, Octopodidae)’, Fisheries Research, Vol. 40, No. 1, pp.81–89.
Davis, A.R. and Moreno, C.A. (1995) ‘Selection of substrata by juvenile Choromytilus chorus (Mytilidae) are chemical cues important’, J. Exp. Mar. Biol. Ecol., Vol. 191, No. 2, pp.167–180.
DiSalvo, L.H. and Martínez, E. (1985) ‘Culture of Ostrea chilensis Philippi, 1845 in a north central Chilean coastal bay’, Biología Pesquera, Vol. 14, pp.16–22.
DiSalvo, L.H., Alarcón, E. and Martínez, E. (1983a) ‘Induced spat production from Ostrea chilensis Philippi 1845 in mid-winter’, Aquaculture, Vol. 30, pp.357–362.
DiSalvo, L.H., Alarcón, E. and Martínez, E. (1983b) ‘Progress in hatchery production of seed of the Chilean oyster Ostrea chilensis (Philippi, 1845)’, Symp. Int. Avances y Perspectivas de la Acuacultura en Chile, pp.269–280.
DiSalvo, L.H., Alarcón, E., Martínez, E. and Uribe, E. (1984) ‘Progress in mass culture of Chlamys (Argopecten) purpurata Lamarck, 1819, with notes on its natural history’, Revista Chilena de Historia Natural, Vol. 57, pp.34–45.
Ebert, E. and Houk, J. (1984) ‘Elements and innovation in the cultivation of the red abalone Haliotis rufescens’, Aquaculture, Vol. 39, pp.375–392.
Ebert, E. and Houk, J. (1989) ‘Abalone cultivation methods used at the California department of fish and games marine resources laboratory’, in Hahn, K.O. (Ed.): CRC Handbook of Culture of Abalone and Other Marine Gastropods, CRC Press, Boca Raton, Fl, pp.239–254, 348.
Encomendero, L. and Dupré, E. (2003) ‘Efecto del sustrato en la intensidad del asentamiento de larvas de Argopecten purpuratus Lamarck 1819 (Bivalvia, Pectinidae) en ambiente controlado’, Investigaciones Marinas, Vol. 31, No. 1, pp.25–32.
Enríquez, R., Frosner, G. G., Hochsteinmintzel, V., Riedemann, S. and Reinhardt, G. (1992) ‘Accumulation and persistence of Hepatitis-A virus in mussels’, Journal of Medical Virology, Vol. 37, No. 3, pp.174–179.

Shellfish culture in Chile 425
Evans, M. (1969) ‘Mechanism of saxitoxin and tetrodotoxion poisoning’, Br. Md. Bull., Vol. 25, No. 3, pp.263–267.
Farias, A. and Uriarte, I. (2001) ‘Effect of microalgae protein on the gonad development and physiological parameters for the scallop Argopecten purpuratus (Lamarck, 1819)’, Journal of Shellfish Research, Vol. 20, No. 1, pp.97–105.
Ferreira, L. (1975) ‘Estudio comparativo de técnicas de cultivo experimental de ostras (Ostrea chilensis)’, Publicaciones Instituto de Fomento Pesquero, Chile, Vol. 58, pp.1–18.
García, C., Pereira, P., Valle, L. and Lagos, N. (2003) ‘Quantitation of diarrhetic shellfish poisoning toxins in Chilean mussel using pyrenyldiazomethane as fluorescent labeling reagent’, Biological Research, Vol. 36, No. 2, pp.171–183.
García-Tello, P., Ardiles, V. and Fajardo, J.P. (2002) ‘Bacterias en la hemolinfa de Argopecten purpuratus (Lamarck, 1819) (Bivalvia: Pectinidae)’, Revista de Biologia Marina y Oceanografía, Vol. 37, No. 2, pp.147–149.
George-Nascimento, M. and Quiroga, G. (1983) ‘Descripción de una nueva especie de trematode, Proctoeces humboldti n. sp. (Digenea: Fellodistomidae), parásito de las lapas Fissurella spp.Bruguière, 1789’, Parasitología al Día, Vol. 7, pp.100–103.
George-Nascimento, M., Balboa, L., Aldana, M. and Olmos, V. (1998) ‘The key-hole limpets Fissurella spp.(Mollusca: Archaeogastropoda) and the clingfish Sicyases sanguineus (Pisces: Gobiesocidae) are sequential hosts of Proctoeces lintoni (Digenea: Fellodistomidae) in Chile’, Revista Chilena de Historia Natural, Vol. 71, No. 2, pp.169–176.
Gesche, E., Montes, L. and Schuler, A.M. (1978) ‘Flora bacteriana aerobia predominante en el erizo comestible (Loxechinus albus, Mol)’, Medio Ambiente, Vol. 3, No. 2, pp.89–92.
Godoy, C. and Jerez, G. (1998) ‘The introduction of abalone in Chile: ten years later’, Journal Shellfish Research, Vol. 17, No. 3, pp.603–605.
Godoy, C., Jerez, G. and Ponce, F. (1992) ‘The introduction of abalone into Chile’, in Sheperd, S.A., Tegner, M. and Guzmán de Proo, S. (Eds.): Abalone of the world. Biology, Fishery and Culture, Blackwell Scientific Publications Inc., pp.485–490, 608.
González, L.E. (1973) ‘Comparación entre el sistema español de encordar mitílidos y el sistema francés actualmente en experimentación’, Circular Instituto de Fomento Pesquero, Chile, Vol. 82, pp.1–10.
González, L.E., Hernández, J.M. and Santa Cruz, S. (1974) ‘Simposio FAO/carpas sobre acuicultura en América Latina’, Algunos aspectos de la Tecnología de los Cultivos Marinos en Chile, p.13.
González, M., Rojas, P. and Campalans, M. (1999c) ‘Anomalías celulares en larvas de ostra Japonesa (Crassostrea gigas) provenientes de cultivo’, Investigaciones Marinas, Vol. 27, pp.111–114.
González, M.L., López, D.A., Pérez, M.C. and Sanhueza, S.E. (1991) ‘Efecto de variaciones en las condiciones ambientales en el aporte de materia orgánica por fecas en invertebrados marinos’, Actas II Congreso Interamericano de Gestión en Recursos Naturales, pp.248–265.
González, M.L., López, D.A., Pérez, M.C., Riquelme, V.A., Uribe, J.M. and Le Pennec, M. (1999a) ‘Growth of the scallop, Argopecten purpuratus (Lamarck, 1819), in southern Chile’, Aquaculture, Vol. 175, Nos. 3–4, pp.307–316.
González, M.L., Pérez, M.C. and López, D.A. (2002) ‘Breeding cycle of the northern scallop, Argopecten purpuratus (Lamarck, 1819) in southern Chile’, Aquaculture Research, Vol. 33, No. 11, pp.847–852.
González, M.L., Pérez, M.C., López, D.A., Uribe, J.M. and Pino, C.A. (1999b) ‘Early development of Fissurella picta (Gmelin, 1791)’, Veliger, Vol. 42, No. 3, pp.275–277.
Gras, N., Thieck, M., Muñoz, L. and Hurtado, S. (1992) ‘Seasonal and geographical variability in some trace elements of pacific oysters (Crassostrea gigas) cultures in 2 different bays of northern Chile’, Journal of Radioanalytical and Nuclear Chemistry Articles, Vol. 161, No. 1, pp.135–146.

426 D.A. López et al.
Guzmán, L. and Campodónico, I. (1975) ‘Marea roja en la región de Magallanes’, Publicaciones del Instituto de la Patagonia, Serie Monografías, Vol. 9, pp.1–44.
Guzmán, L. and Campodónico, I. (1978) ‘Marea roja en Chile’, Interciencia, Vol. 3, No. 3, pp.144–151.
Hancock, A.D. (1969) ‘La pesquería de mariscos en Chile, Publicación No. 45, Instituto de Fomento Pesquero, Chile, p.94.
Hauer, R. (1988) ‘Experiencia de producción de ostras individuales (Crassostrea gigas) en laboratorio’, in Uribe, E. (Ed.): Producción de Larvas y Juveniles de Especies Marinas, Universidad Católica del Norte, Chile, pp.115–118.
Hernández, C. and Becerra, R. (1997) ‘Primeros antecedentes sobre fuerza del biso y capacidad de desplazamiento en cholga Aulacomya ater (Molina, 1782) (Mollusca, Mytilidae)’, Revista de Biologia Marina, Vol. 32, No. 1, pp.1–10.
Hine, P.M. (1996) ‘Southern hemisphere mollusc diseases and overview of associated risk assessment problems’, Revue Scientifique et Technique del Office International des Epizooties, Vol. 15, No. 2, pp.563–577.
Hine, P.M. (2002) ‘Severe apicomplexan infection in the oyster Ostrea chilensis: a possible predisposing factor in bonamiosis’, Diseases of Aquatic Organisms, Vol. 51, No. 1, pp.49–60.
Hine, P.M., Cochennec-Laureau, N. and Berthe, F.C.J. (2001) ‘Bonamia exitiosus n. sp (Haplosporidia) infecting flat oysters Ostrea chilensis in New Zealand’, Diseases of Aquactic Organisms, Vol. 47, No. 1, pp.63–72.
Hine, P.M., Diggles, B.K., Parsons, M.J.D., Pringle, A. and Bull, B. (2002) ‘The effects of stressors on the dynamycs of Bonamia exitiosus Hine, Cochennec-Laureau & Berthe, infections in flat oysters Ostrea chilensis (Philippi)’, Journal of Fish Diseases, Vol. 25, No. 9, pp.545–554.
Hine, P.M., Wesney, B. and Besant, P. (1998) ‘Replication of a herpes-like virus in larvae of the flat oyster Tiostrea chilensis at ambient temperatures’, Diseases of Aquatic Organisms, Vol. 32, No. 3, pp.161–171.
Hoyl, A. and Uribe, E. (1990) ‘Cultivo de la ostra del Pacífico Crassostrea gigas, en Chile (1977–1989)’, in Hernández, A. (Ed.): Cultivo de moluscos en América Latina, pp.265–272.
Huaquin, L., Guerra, R. and Bretos, M. (1998) ‘Identificación del sexo y morfología de gametos de la lapa Fissurella crassa Lamarck, 1822 (Mollusca: Archaeogastropoda)’, Revista de Biología Marina y Oceanografía, Vol. 33, No. 2, pp.223–239.
IFOP (1998) Estado de Situación y Perspectivas de la Acuicultura en Chile, Instituto de Fomento Pesquero, FDI-CORFO, p.194.
Iglesias, J., Sánchez, F.J. and Otero, J.J. (1996) ‘The octopus (Octopus vulgaris Cuvier): a candidate for aquaculture?’, ICES C. M. 1996/F: 10.
Illanes, J. (1988) ‘Experiencias de captación de larvas de ostión (Argopecten purpuratus) en Chile. IV Región’, in Uribe, E. (Ed.): Producción de Larvas y Juveniles de Especies Marinas, Universidad Católica del Norte, Chile, pp.53–57.
Illanes, J. (1989) ‘Tecnología de cultivo de ostras y ostiones en el mundo’, in Flores, H. (Ed.): 5 to Taller de Acuacultura, Universidad Católica del Norte, Chile, pp.6–11.
Illanes, J. (1990) ‘Cultivo del ostión del norte’, Argopecten purpurata’, in Hernández, A. (Ed.): Cultivo de Moluscos en América Latina, pp.211–230.
Illanes, J.E. (1999) ‘Tecnología de cultivo de ‘hatchery’ de semilla de abalón japonés Haliotis discus hannai’, Taller: Cultivo de abalón japonés. Proyecto FONDEF D96/1102, Coquimbo, Chile, p.100.
Jaramillo, E., Bertrán, C. and Bravo, A. (1992) ‘Mussel biodeposition in an estuary in southern Chile’, Marine Ecology Progress Series, Vol. 82, No. 1, pp.85–94.
Jaramillo, E., Pino, M., Filun, L. and González, M. (1994) ‘Longshore distribution of Mesodesma donacium (Bivalvia, Mesodesmatidae) on a sandy beach of the south of Chile’, Veliger, Vol. 37, No. 2, pp.192–200.

Shellfish culture in Chile 427
Jaramillo, R. and Navarro, J. (1995) ‘Reproductive cycle of the Chilean ribbed mussel Aulacomya ater (Molina, 1782)’, Journal of Shellfish Research, Vol. 14, No. 1, pp.165–171.
Jeffs, A.G. (1998) ‘Gametogenic cycle of the Chilean oyster, Tiostrea chilensis (Philippi, 1845), in north-eastern New Zealand’, Invertebrate Reproduction and Development, Vol. 34, Nos. 2–3, pp.109–116.
Jeffs, A.G. (1999) ‘The potential for developing controlled breeding in the Chilean oyster’, Aquaculture International, Vol. 7, No. 3, pp.189–199.
Jeffs, A.G. and Hickman, R.W. (2000) ‘Reproductive activity in a pre-epizootic wild population of the Chilean oyster, Ostrea chilensis, from southern New Zealand’, Aquaculture, Vol. 183, Nos. 3–4, pp.241–253.
Jeffs, A.G., Dunphy, B.J. and Wells, R.M.G. (2002) ‘Experimental effects of water temperature on the gametogenic development of broodstock in the oyster, Ostrea chilensis’, Journal of Shellfish Research, Vol. 21, No. 2, pp.743–747.
Jirón, C. (1979) ‘Presencia de trematodes digenéticos adultos en Fissurella cumingii Reeve, 1849 en Iquique, Chile’, Arch. Biolo. Med. Exp., Vol. 12, pp.520, 521.
Jorquera, M.A., Riquelme, C.E., Loyola, L.A. and Muñoz, L.F. (2000) ‘Production of bactericidal substances by a marine vibrio isolated from cultures of the scallop Argopecten purpuratus’, Aquaculture International, Vol. 7, No. 6, pp.433–448.
Jorquera, M.A., Silva, F.R. and Riquelme, C.E. (2001) ‘Bacteria in the culture of the scallop Argopecten purpuratus (Lamarck, 1819)’, Aquaculture International, Vol. 9, No. 4, pp.285–303.
Lagos, N. (1998) ‘Microalgal blooms: a global issue with negative impacts in Chile’, Biological Research, Vol. 31, No. 4, pp.375–386.
Leiva, G.E. and Castilla, J.C. (2001) ‘A review of the world marine gastropod fishery: evolution of catches management and the Chilean experience’, Reviews in Fish Biology and Fisheries, Vol. 11, No. 4, pp.283–300.
Lembeye, G. (1981) ‘Segunda aparición del Veneno Paralítico de los Mariscos (VPM) asociados Gonyaulax catenella, en Magallanes (Chile)’, Anales del Instituto de la Patagonia, Serie Ciencias Naturales, Vol. 12, pp.273–276.
Lizarralde, Z.I. (1998) ‘Variaciones mensuales en el asentamiento de semilla de cholga (Aulacomya atra atra, Molina) sobre colectores artificiales en el Golfo San José, Argentina’, Biología Pesquera, Vol. 27, pp.31–38.
Llenart, M., Handlinger, J. and Powell, M. (2003) ‘Spionid mudworm infestation of farmed abalone (Haliotis spp)’, Aquaculture, Vol. 221, Nos. 1–4, pp.85–96.
Lohrmann, K.B., Brand, A.R. and Feist, S.W. (2002) ‘Comparison of the parasites and pathogens present in a cultivated and in a wild population of scallops (Argopecten purpuratus Lamarck, 1819) in Tongoy Bay, Chile’, Journal of Shellfish Research, Vol. 21, No. 2, pp.557–561.
López, B.A., Arriagada, S.E. and López, D.A. (2003c) ‘El ‘picoroco’ un recurso chileno de altas potencialidades’, Chile Pesquero, Vol. 131, pp.36–39.
López, B.A., Pérez, M., López, D. and González, M.L. (2004) ‘El pulpo: un promisorio recurso de exportación’, Chile Pesquero, Vol. 141, pp.8–11.
López, D., Buschmann, A. and González, M.L. (1988) ‘Efectos del uso de las zonas costeras por prácticas de acuicultura’, Medio Ambiente, Vol. 9, No. 1, pp.42–54.
López, D., González, M.L. and Buschmann, A. (1989) ‘Efectos ambientales de cultivos de moluscos bivalvos: Revisión general del problema y requerimientos de investigación en Chile’, 5 to taller de Acuacultura. Universidad Católica del Norte, Coquimbo, Chile, p.51.
López, D., López, B. and Arriagada, S. (2006) ‘Strategies of aquaculture diversification by endemic marine invertebrates’, World Aquaculture Magazine, Vol. 37, No. 2, pp.39–42.
López, D.A. and Varela, C. (1988) ‘Manejo de repoductores y posturas de cápsulas en Concholepas concholepas (Bruguière, 1789) (Gastropoda, Muricidae): Una revisión de problemas y requerimientos de investigación’, Biología Pesquera, Vol. 17, pp.21–30.

428 D.A. López et al.
López, D.A., González, M.L. and Pérez, M.C. (2003a) ‘Feeding and growth in the keyhole limpet Fissurella picta (Gmelin, 1791)’, Journal of Shellfish Research, Vol. 22, No. 81, pp.165–169.
López, D.A., López, B.A. and Arriagada, S.E. (2003b) ‘Es posible cultivar picorocos?’, Chile Acuícola, Vol. 14, pp.26–29.
López, D.A., Riquelme, V.A. and González, M.L. (2000) ‘The effects of epibionts and predators on the growth and mortallity rates of Argopecten purpuratus cultures (Lamarck, 1819), in southern Chile’, Aquaculture International, Vol. 8, pp.431–442.
Lozada, E. and Santa Cruz, S. (1979) ‘Choro zapato Choromytilus chorus (Molina) Lamellibranchia Anisomyaria Mytilidae’, Estado Actual de las Principales Pesquerías Nacionales. Bases Para un Desarrollo Pesquero, Moluscos, AP79, pp.18–35.
Lozada, E., Hernández, J.M., Aracena, O. and López, M.T. (1974) ‘Cultivo de la cholga (Aulacomya ater) en Isletilla, estero de Castro (Mollusca; Bivalvia; Mytilidae)’, Boletín de la Sociedad de Biología de Concepción, Vol. 48, pp.331–346.
Martinez, G., Aguilera, C. and Mettifogo, L. (2000) ‘Interactive effects of diet and temperature on reproductive conditioning of Argopecten purpuratus broodstock’, Aquaculture, Vol. 183, Nos. 1–2, pp.149–159.
Mendo, J. and Jurado, E. (1993) ‘Lenght based growth parameter estimates of the Peruvian scallop (Argopecten purpuratus)’, Fisheries Research, Vol. 15, No. 4, pp.357–367.
Ministry of Economy and Energy (2003) Prospectiva Chile 2010. 4. La industria de la Acuicultura, Chile- Innova, pp.1–78.
Mix, M.C. and Breese, W.P. (1980) ‘A cellular proliferative disorder in oysters (Ostrea chilensis) from Chiloé, Chile’, Journal of Invertebrate Pathology, Vol. 36, pp.123, 124.
Montebruno, D. (1993) ‘Anatomopathological study of paralytic shellfish intoxication in the XII Region of Chile’, Revista Médica de Chile, Vol. 121, No. 1, pp.94–97.
Moore, J.D., Finley, C.A., Robbins, T.T. and Friedman, C.S. (2002) ‘Whitering syndrome and restoration of southern California abalone populations’, California Cooperative Oceanic Fisheries Investigations Reports, Vol. 43, pp.112–117.
Moore, J.D., Robbins, T.T. and Friedman, C.S. (2000) ‘Whitering syndrome in farmed red abalone Haliotis rufescens. Thermal induction and association with a gastrointestinal Rickettsiales-like prokaryote’, Journal of Aquatic Animal Health, Vol. 12, No. 1, pp.26–34.
Moore, J.D., Robbins, T.T., Hedrick, R.P. and Friedman, C.S. (2001) ‘Transmision of the Rickettsiales-like prokaryote ‘Candidatus xenohaliotis californiensis’ and its role in withering syndrome of California abalone spp’, Journal of Shellfish Research, Vol. 20, No. 2, pp.867–874.
Nakamura, A., Takahashi, K.G. and Mori, K. (1999) ‘Vibriostatic bacteria isolated from rearing seawater of oyster brood stock: potentiality as biocontrol agents for vibriosis in oyster larvae’, Fish Pathology, Vol. 34, No. 3, pp.139–144.
Navarro, J.M. and Gutiérrez, A. (1990) ‘Sinopsis sobre el estado actual del cultivo de mitílidos en Chile’, Cultivo de Moluscos en América Latina, pp.189–210.
Navarro, J.M., Leiva, G.E., Martínez, G. and Aguilera, C. (2000) ‘Interactive effects of diet and temperature on the scope for growth of the scallop Argopecten purpuratus during reproductive conditioning’, Journal of Experimental Marine Biology and Ecology, Vol. 247, No. 1, pp.67–83.
Nevejan, N., Courtnes, V., Hauva, M., Gajardo, G. and Sorgeloos, P. (2003b) ‘Effects of lipid emulsions on production and fatty acid composition of eggs of the scallop Argopecten purpuratus’, Marine Biology, Vol. 143, No. 2, pp.327–338.
Nevejan, N., Saez, I., Gajardo, G. and Sorgeloos, P. (2003a) ‘Supplementation of EPA and DHA to a Dunaliella tertiolecta diet: effect on growth and lipid composition of scallop larvae, Argopecten purpuratus (Lamarck, 1819)’, Aquaculture, Vol. 217, Nos. 1–4, pp.613–632.
Oliva, M., Herrera, H., Matulic, J. and Severino, B. (1986) ‘Parasitismo en el ostión del norte Chlamys purpuratus (L.)’, Parasitismo al Día, Vol. 8, pp.36–86.

Shellfish culture in Chile 429
Oliva, M.E. and Huaquin, L.G. (2000) ‘Progenesis in Proctoeces lintoni (Fellodistomidae, a parasitic of Fissurella crassa (Archaeogastropoda) in a latitudinal gradient in the Pacific coast of South America’, Journal of Parasitology, Vol. 86, No. 4, pp.768–772.
Oliva, M.E., Olivares, A.N., Diaz, C.D. and Pasten, M.V. (1999) ‘Parasitic castration in Concholepas concholepas (Gastropoda: Muricidae) due to a larval digenean in northern Chile’, Diseases of Aquatic Organisms, Vol. 36, No. 1, pp.61–65.
Ortiz, M., Wolff, M. and Mendo, J. (2002) ‘Quantitative modelling for the development of a sustainable management strategy for the Peruvian scallop Argopecten purpuratus (Lamarck 1819)’, Aquatic Conservation Marine and Freshwater Ecosystems, Vol. 12, No. 3, pp.273–287.
Osorio, C., Aracena, O.L., Lozada, E. and López, M.T. (1978) ‘Fluctuaciones de la fauna acompañante de mitílidos en colectores de Putemún, Chiloé’, Boletín de la Sociedad de Biología de Concepción, Vol. 51, pp.183–192.
Osorio, C., Diaz, H. and Ríos, M. (1986) ‘Prevalencia de Proctoeces humboldti (Digenea: Fellodistomidae) en la población de Fissurella maxima Sowerby, 1835, de los Vilos, Chile’, Revista de Biología Marina, Vol. 22, pp.157–168.
Owen, B., DiSalvo, L., Ebert, E. and Fonk, E. (1984) ‘Culture of the California red abalone Haliotis rufescens Swainson (1822) in Chile’, Veliger, Vol. 27, pp.101–105.
Padilla, M. and Orrego, J. (1967) ‘La fijación larval de ostras sobre colectores experimentales en Quetalmahue 1966–1967’, Boletín Científico del Instituto de Fomento Pesquero, Vol. 3, pp.69–128.
Padilla, M., Mendez, M. and Casanova, F. (1969) ‘Observaciones sobre el comportamiento de la Ostrea chilensis en Apiao’, Boletín Científico del Instituto de Fomento Pesquero, Vol. 10, pp.28–51.
Palma-Fleming, H., Navarro, J.M., Peña, E. and Martinez, G. (2002) ‘Effect of three conditioning diets on the fatty acid composition of gonads and muscle of Argopecten purpuratus’, New Zealand of Marine and Freshwater Research, Vol. 36, No. 3, pp.605–620.
Reyes, E. (1986) ‘Qué pasó con el loco? Crónica de un colapso anunciado’, Ambiente y Desarrollo, Vol. 2, pp.142–146.
Riquelme, C., Araya, R. and Escribano, R. (2000) ‘Selective incorporation of bacteria by Argopecten purpuratus larvae: implications for the use of probiotics in culturing systems of the Chilean scallop’, Aquaculture, Vol. 181, Nos. 1–2, pp.25–36.
Riquelme, C., Hayashida, G., Araya, R., Uchida, A., Satomi, M. and Ishida, Y. (1996b) ‘Isolation of a native bacterial strain from the scallop Argopecten purpuratus with inhibitory effects againts pathogenic vibrios’, Journal of Shellfish Research, Vol. 15, No. 2, pp.369–374.
Riquelme, C., Hayashida, G., Toranzo, A.E., Vilches, J. and Diaz, M.A. (1995a) ‘Pathogenecity studies on a Vibrio anguillarum-related (var) starin causing an epizootic in Argopecten purpuratus larvae cultured in Chile’, Diseases of Aquatic Organisms, Vol. 22, No. 2, pp.135–141.
Riquelme, C., Hayashida, G., Vergara, N., Vásquez, A., Morales, Y. and Chavez, P. (1995b) ‘Bacteriology of the scallop Argopecten purpuratus (Lamarck, 1819) cultured in Chile’, Aquaculture, Vol. 138, Nos. 1–4, pp.49–60.
Riquelme, C., Toranzo, A.E., Barja, J.L., Vergara, N. and Araya, R. (1996a) ‘Association of Aeromonas hydrophila and Vibrio alginolyticus with larval mortalities of scallop (Argopecten purpuratus)’, Journal of Invertebrate Pathology, Vol. 67, No. 3, pp.213–218.
Rivas, M., García, C., Liberona, J.L. and Lagos, N. (2000) ‘Biochemical characterization and inhibitory effects of dinophysistoxin-1, okadaic acid and microcystine1-r on protein phosphatase 2a purified from the mussel Mytilus chilensis’, Biological Research, Vol. 33, Nos. 3–4, pp.197–206.

430 D.A. López et al.
Romero, J., García-Varela, M., Laclette, J.P. and Espejo, R.T. (2002) ‘Bacterial 16S rRNA gene analysis revealed that bacteria related to Arcobacter spp. Constitute an abundant and common component of the oyster microbiota (Tiostrea chilensis)’, Microbial Ecology, Vol. 44, No. 4, pp.365–371.
Santa Cruz, S. (1976) ‘Resultados de algunas experiencias de cultivo de cholgas (Aulacomya ater) en la bahía de Mejillones’, Instituto de Fomento Pesquero. Informes Pesqueros, Vol. 60, pp.1–41.
Santa Cruz, S., González, L.E. and Hernández, J.M. (1977) ‘El cultivo de mitílidos en Chile. Descripción de sistemas’, Serie Informes Pesqueros Instituto de Fomento Pesquero, Chile, Vol. 64, pp.1–57.
Sato-Okoshi, W. and Takatsuka, M. (2001) ‘Polydora and related genera (Polychaeta, Spionidae) around Puerto Montt and Chiloe Island (Chile), with description of a new species of Dipolydora’, Bulletin Marine Science, Vol. 68, No. 3, pp.485–503.
Sernapesca (1982–2003) Anuario Estadístico de Pesca, Servicio Nacional de Pesca, Chile, p.195. Sindermann, C.J. (1977) Diseases Diagnosis and Control in North American Marine Aquaculture,
Elsevier Scientific Publications, New York, USA, p.329. Smolowitz, R. and Shumway, S.E. (1997) ‘Possible cytotoxic effects of the dinoflagelate,
Gyrodinium aureolum, on juvenile bivalve mollusks’, Aquaculture International, Vol. 5, No. 4, pp.291–300.
Solis, I. (1973) ‘Valoración de colectores de larvas de ostras Ostrea chilensis Philippi en Pullinque’, Biología Pesquera, Vol. 6, pp.5–23.
Solis, I. and Eberhard, P. (1979) ‘Ostra’, Estado Actual de las Principales Pesquerías Nacionales. Bases Para un Desarrollo Pesquero, CORFO-IFOP, Moluscos, p.30.
Sugumar, G., Nakai, T., Hirata, Y., Matsubara, D. and Muroga, K. (1998a) ‘Pathogenicity of Vibrio splendidus biovar II, the causative bacterium of bacillary necrosis of Japanese oyster larvae’, Fish Pathology, Vol. 33, No. 2, pp.79–84.
Sugumar, G., Nakai, T., Hirata, Y., Matsubara, D. and Muroga, K. (1998b) ‘Vibrio splendidus biovar II as the causative agent of bacillary necrosis of Japanese oyster Crassostrea gigas larvae’, Diseases of Aquatic Organisms, Vol. 33, No. 2, pp.111–118.
Toro, J. (1991) ‘Respuesta a la selección y estima de heredabilidad realizadas para el carácter longitud de la valva en la ostra chilena Ostrea chilensis, Philippi 1845’, Revista de Biología Marina, Vol. 26, No. 2, pp.351–362.
Toro, J., Sanhueza, M. and Winter, J. (1992) ‘Respuesta a la selección genética divergente en juveniles de Tiostrea chilensis Philippi 1845’, Investigación Pesquera, Vol. 37, pp.89–96.
Toro, J., Vergara, A. and Galleguillos, R. (1995b) ‘Definición de heterocigotos en una cohorte de Ostrea chilensis (Philippi, 1845)’, Biología Pesquera, Vol. 24, pp.3–6.
Toro, J.E. and Chaparro, O.R. (1990) ‘Conocimiento biológico de Ostrea chilensis Philippi 1845. Impacto y perspectivas en el desarrollo de la ostricultura en Chile’, Cultivo de Moluscos de América Latina, pp.231–264.
Toro, J.E. and Paredes, L.I. (1996) ‘Heritability estimates of larval shell length in the Chilean blue mussel Mytilus chilensis, under different food densities’, Aquatic Living Resources, Vol. 9, No. 4, pp.347–350.
Toro, J.E. and Sastre, H.D. (1995) ‘Induced triploidy in the Chilean blue mussel, Mytilus chilensis (Hupe, 1984), and perfomance of triploid larvae’, Journal of Shellfish Research, Vol. 14, No. 1, pp.161–164.
Toro, J.E., Sanhueza, M.A., Paredes, L. and Canello, F. (1995a) ‘Induction of triploid embryos by heat-shock in the Chilean northern scallop Argopecten purpuratus Lamarck, 1819’, New Zealand Journal of Marine and Freshwater Research, Vol. 29, No. 1, pp.101–105.
Uki, N. and Kikuchi, S. (1984) ‘Regulation of maturation and spawning of an abalone, Haliotis (Gastropoda) by external environmental factors’, Aquaculture, Vol. 39, pp.247–254.

Shellfish culture in Chile 431
Uriarte, I. and Farias, A. (1999) ‘The effects od dietary protein content on growth and biochemical composition of Chilean scallop Argopecten purpuratus (L.) postlarvae and spat’, Aquaculture, Vol. 180, Nos. 1–2, pp.119–127.
Uriarte, I., Farias, A. and Castilla, J.C. (2001) ‘Effect of antibiotic treatment during larval development of the Chilean scallop Argopecten purpuratus’, Aquacultural Engineering, Vol. 25, No. 3, pp.139–147.
Uriarte, I., Farias, A. and Muñoz, C. (1996b) ‘Growth and condition index of Argopecten purpuratus (Lamarck) in the South of Chile. Improvement of the Commercial Production of Marine Aquaculture Species’, in Gajardo, G. and Costteau, P. (Eds.): Proceeding of a Workshop on Fish and Mollusc Larviculture, pp.135–143.
Uriarte, I., Farias, A. and Muñoz, C.M. (1996a) ‘Cultivo en hatchery y pre-engorde del ostión del norte, Argopecten purpuratus (Lamarck, 1819), en el sur de Chile’, Revista de Biología Marina, Vol. 31, No. 2, pp.81–90.
Uribe, J.C., García, C., Rivas, M. and Lagos, N. (2001) ‘First report of diarrehetic shellfish toxins in magellanic fjords, southern Chile’, Journal of Shellfish Research, Vol. 20, No. 1, pp.69–74.
Varela, C.E. and López, D.A. (1989) ‘Manejo de reproductores de Concholepas concholepas (Bruguiere) en el diseño de una estrategia de repoblación’, Medio Ambiente, Vol. 10, No. 1, pp.3–12.
Vásquez, M., Gruttner, C., Gallacher, S. and Moore, E.R.B. (2001) ‘Detection and characterization of toxigenic bacteria associated with Alexandrium catenella and Aulacomya ater contaminated with PSP’, Journal of Shellfish Research, Vol. 20, No. 3, pp.1245–1249.
Vásquez, M., Gruttner, C., Moeller, B. and Moore, E.R.B. (2002) ‘Limited selection of sodium channel blocking toxin-producing bacteria from paralytic shellfish toxin-contaminated mussels (Aulacomya ater)’, Research Microbiology, Vol. 153, No. 6, pp.333–338.
Velasco, L.A. and Navarro, J.M. (2002) ‘Feeding physiology of infaunal (Mulinia edulis) and epifaunal (Mytilus chilensis) bivalves under a wide range of concentrations and qualities seston’, Marine Ecology Progress Series, Vol. 240, pp.143–155.
Velasco, L.A. and Navarro, J.M. (2003) ‘Energetic balance of infaunal (Mulinia edulis king, 1831) and epifaunal (Mytilus chilensis Hupe, 1854) bivalves in response to wide variations in concentration and quality of seston’, Journal of Experimental Marine Biology and Ecology, Vol. 296, No. 1, pp.79–92.
Vial, M.V., Teuber, C., Costabal, P., Poblete, T., Donoso, T.G. and Gebauer, M.T. (1988) ‘Presencia de Vibrio anguillarum (Canestrini) en el tracto digestivo de Mytilus chilensis (Hupe)’, Biota, Vol. 4, No. 2, pp.119–124.
Winkler, F., de Guevara, B.L., Estévez, B. and Jollan, L. (1993) ‘Inducción a triploidia en Argopecten purpuratus Lamarck, 1819, mediante citocalasina-B’, Revista de Biología Marina, Vol. 28, No. 2, pp.239–246.
Winkler, F.M. and Estevez, B.F. (2003) ‘Effects of self-fertilization on growth and survival of larvae and juveniles of the scallop Argopecten purpuratus L’, Journal of Experimental Marine Biology and Ecology, Vol. 292, No. 1, pp.93–102.
Winter, J.E. (1978) ‘A review on the knowledge of suspension-feeding in lamellabranchiate bivalves, with special reference to artificial aquaculture systems’, Aquaculture, Vol. 13, pp.1–33.
Winter, J.E. and Chaparro, O.R. (1987) ‘Estado actual necesidades y perspectivas de la Acuicultura de moluscos bivalvos cultivados en aguas templadas del pacífico de Sudamérica’, Proceedings Taller sobre Acuicultura en América Latina, Lima, Perú, pp.261–270.
Winter, J.E., Toro, J.E., Navarro, J.M., Valenzuela, G.S. and Chaparro, O.R. (1984) ‘Recent developments, status, and prospects of molluscan aquaculture on the Pacific coast of South America’, Aquaculture, Vol. 39, pp.95–134.
Zhao, J., Lembeye, G., Cenci, G., Wall, B. and Yasumoto, T. (1993) ‘Determination of okadaic acid and dinophysistoxin-1 in mussels from Chile, Italy and Ireland’, in Smayda, T.J. and Shimuzi, Y. (Eds.): Toxic Phytoplankton Blooms in the Sea, Elsevier Science Publishers Co. Inc., pp.587–592.
Related Documents











