This article was downloaded by: [193.191.134.1] On: 17 January 2012, At: 00:11 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK European Journal of Phycology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tejp20 Sexual reproduction, mating system, chloroplast dynamics and abrupt cell size reduction in Pseudo-nitzschia pungens from the North Sea (Bacillariophyta) Victor A. Chepurnov a , David G. Mann b , Koen Sabbe a , Katrijn Vannerum a , Griet Casteleyn a , Elie Verleyen a , Louis Peperzak c & Wim Vyverman a a Laboratory of Protistology and Aquatic Ecology, Department of Biology, Ghent University, Krijgslaan 281-S8, B-9000 Gent, Belgium b Royal Botanic Garden, Edinburgh EH3 5LR, Scotland, UK c Rijksinstituut voor Kust & Zee, P.O. Box 8039, NL-4330 EA Middelburg, The Netherlands Available online: 20 Feb 2007 To cite this article: Victor A. Chepurnov, David G. Mann, Koen Sabbe, Katrijn Vannerum, Griet Casteleyn, Elie Verleyen, Louis Peperzak & Wim Vyverman (2005): Sexual reproduction, mating system, chloroplast dynamics and abrupt cell size reduction in Pseudo-nitzschia pungens from the North Sea (Bacillariophyta), European Journal of Phycology, 40:4, 379-395 To link to this article: http://dx.doi.org/10.1080/09670260500254735 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was downloaded by: [193.191.134.1]On: 17 January 2012, At: 00:11Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
European Journal of PhycologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tejp20
Sexual reproduction, mating system, chloroplastdynamics and abrupt cell size reduction inPseudo-nitzschia pungens from the North Sea(Bacillariophyta)Victor A. Chepurnov a , David G. Mann b , Koen Sabbe a , Katrijn Vannerum a , GrietCasteleyn a , Elie Verleyen a , Louis Peperzak c & Wim Vyverman aa Laboratory of Protistology and Aquatic Ecology, Department of Biology, GhentUniversity, Krijgslaan 281-S8, B-9000 Gent, Belgiumb Royal Botanic Garden, Edinburgh EH3 5LR, Scotland, UKc Rijksinstituut voor Kust & Zee, P.O. Box 8039, NL-4330 EA Middelburg, The Netherlands
Available online: 20 Feb 2007
To cite this article: Victor A. Chepurnov, David G. Mann, Koen Sabbe, Katrijn Vannerum, Griet Casteleyn, Elie Verleyen,Louis Peperzak & Wim Vyverman (2005): Sexual reproduction, mating system, chloroplast dynamics and abrupt cell sizereduction in Pseudo-nitzschia pungens from the North Sea (Bacillariophyta), European Journal of Phycology, 40:4, 379-395
To link to this article: http://dx.doi.org/10.1080/09670260500254735
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss, actions,claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

Eur. J. Phycol. (2005), 40: 379–395
Sexual reproduction, mating system, chloroplast dynamics
and abrupt cell size reduction in Pseudo-nitzschia pungens
from the North Sea (Bacillariophyta)
VICTOR A. CHEPURNOV1, DAVID G. MANN2, KOEN SABBE1, KATRIJN VANNERUM1,
GRIET CASTELEYN1, ELIE VERLEYEN1, LOUIS PEPERZAK3 AND WIM VYVERMAN1
1Laboratory of Protistology and Aquatic Ecology, Department of Biology, Ghent University, Krijgslaan 281-S8,
B-9000 Gent, Belgium2Royal Botanic Garden, Edinburgh EH3 5LR, Scotland, UK3Rijksinstituut voor Kust & Zee, P.O. Box 8039, NL-4330 EA Middelburg, The Netherlands
(Received 18 October 2004; accepted 12 June 2005)
Clonal cultures of Pseudo-nitzschia pungens were isolated at various times from seven sites in the North Sea. During the mitotic
cell cycle, the two plate-shaped chloroplasts were girdle-appressed during interphase and mitosis. After cytokinesis, the
chloroplasts moved onto the parental valve and remained there during the formation of the new hypovalve and until
separation and re-arrangement of the sibling cells within the cell chain had been completed. Clones were almost always
heterothallic and cultures of opposite mating type isolated from different localities were compatible. Meiosis I was cytokinetic
and accompanied by chloroplast division. Meiosis II involved karyokinesis but not cytokinesis and preceded the
rearrangement and contraction of the two gametes. Sexual reproduction involved physiological anisogamy. With one
exception, gamete behaviour was clone-specific, gametes being active in clones of one mating type but passive in clones of the
other mating type. Auxospore development was accompanied by deposition of a transverse and then a longitudinal
perizonium. Infrequently, triploid auxospores and presumably haploid auxospores were produced. The four chloroplasts of
diploid auxospores did not divide, and behaved synchronously during the two acytokinetic mitotic cycles accompanying
the deposition of the initial thecae. Just before the first division of the initial cell, the chloroplasts shifted onto the valves
(two per valve). The division of the initial cell was not accompanied by chloroplast division and so the two daughter cells
received two chloroplasts each. Two modes of abrupt cell size reduction were detected. One occurred during initial cell
formation when part of the expanded auxospore aborted. The other pattern was more gradual and was observed in
growing cultures; during successive cell divisions a frustule constriction appeared and intensified, one chloroplast split
into two, and part of the protoplast aborted. A simple naming system is proposed for mating types in pennate diatoms.
Key words: abrupt size reduction, auxosporulation, Bacillariophyta, chloroplast, diatoms, heterothally, mating system,
Pseudo-nitzschia, sexual reproduction
Introduction
Species of the fibulate pennate diatom Pseudo-nitzschia H. Peragallo in H. Peragallo et Peragalloare common components of phytoplankton inmarine coastal and open-ocean waters world-wide(e.g. Hasle, 2002). During the last 20 years,Pseudo-nitzschia has attracted much attentionbecause some species produce the neurotoxindomoic acid and blooms can have severeadverse consequences for marine biota andhumans (e.g. Bates, 2000). So far, nine species of
Pseudo-nitzschia have been shown to be actuallyor potentially toxic (Fryxell & Hasle, 2003).The economic significance of Pseudo-nitzschia
species has accelerated study of various aspects oftheir biology and ecology (e.g. Parsons et al.,2002; Maldonado et al., 2002; Fehling et al., 2004;Lundholm et al., 2004; Orellana et al., 2004), tohelp establish a basis for monitoring, understand-ing and countering Pseudo-nitzschia blooms.Consequently, Pseudo-nitzschia species haverapidly become some of the best-studied of alldiatoms. However, as von Stosch commented,‘‘You only know a species if you know its completelife cycle’’ (Elbrachter, 2003, p. 629) and long-termmaintenance of diatoms and completion of the
Correspondence to: Victor Chepurnov.
e-mail: [email protected]
ISSN 0967-0262 print/ISSN 1469-4433 online � 2005 British Phycological Society
DOI: 10.1080/09670260500254735
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

life-cycle in culture require that the characteristicsof the mating system are understood (Mann &Chepurnov, 2004; Chepurnov et al., 2004). In theserespects there is still much to be learned aboutPseudo-nitzschia (Mann, 2002; Mann & Bates,2002). Fortunately, Pseudo-nitzschia species areeasily isolated into culture.Some information on sexual reproduction and
mating systems is available for five Pseudo-nitzschia species, all of them potentially toxic(Fryxell & Hasle, 2003). The best studied isPseudo-nitzschia multiseries (Hasle) Hasle(Davidovich & Bates, 1998; Hiltz et al., 2000),which proved to be heterothallic. Davidovich andBates (1998) also reported heterothallic sexualreproduction in Pseudo-nitzschia pseudodelicatis-sima (Hasle) Hasle from two localities, namely theBlack Sea and the American coast of the AtlanticOcean. Recent re-examination of their cultures byscanning electron microscopy (Lundholm et al.,2003) revealed, however, that the Black Sea strainsbelong to the newly described species Pseudo-nitzschia calliantha Lundholm, Moestrup & Hasle;the Atlantic clones are ‘‘either P. pseudodelica-tissima or P. cuspidata; the identity could not befinally established because the shape of the valve isuncertain’’ (Lundholm et al., 2003). Sexual repro-duction and heterothallic mating behaviour havealso been reported in Pseudo-nitzschia delicatissima(Cleve) Heiden in Heiden et Kolbe from theMediterranean Sea (Amato et al., 2005) andPseudo-nitzschia fraudulenta (Cleve) Hasle fromthe North Sea (Chepurnov et al., 2004). Noinformation, however, is currently available onlife-cycle traits in Pseudo-nitzschia pungens(Grunow ex Cleve) Hasle, which is a very widelydistributed, perhaps cosmopolitan species (Hasle,2002) and which is also potentially toxic (Fryxell &Hasle, 2003). We therefore isolated a series ofmonoclonal cultures from various localities in theNorth Sea where P. pungens is abundant (e.g.Vrieling et al., 1996). Sexual reproduction wassuccessfully initiated experimentally and wedescribe its characteristics, together with someother aspects of the cell and life-cycle that havenot been reported previously for any Pseudo-nitzschia species.
Materials and methods
Sampling, mating experiments and microscopy
Planktonic samples containing living cells of P. pungenswere collected from seven locations in the North Sea(Fig. 1) and 24 clonal cultures were established (Table 1)by isolating single colonies by micropipette. Each colonywas placed in a separate well of a Repli dish (24-wellplates, Greiner bio-one, Frickenhausen, Germany,
No. 662160) with 2.5ml of f2 culture medium(Guillard, 1975), based on filtered and sterilized sea-water (c. 32ø) collected from the North Sea. The cloneswere subsequently grown in 24-well Repli dishes or50-mm Petri dishes in an incubator at 188C with 12:12 hlight-dark period and 25–50 mmol photons m�2 s�1 fromcool-white fluorescent lights. Cells were re-inoculatedinto fresh medium every 7–10 days.
For mating experiments, mixed cultures were pre-pared by inoculating clones (in exponential growthphase) together into Repli wells in all possible pair-wisecombinations. The cultures were examined daily for6–8 days, using a Zeiss Axiovert 135 inverted micro-scope (Zeiss, Jena, Germany), until they reached thestationary phase of growth.
For DAPI (40-6-diamidino-2-phenylindole) staining ofnuclei during gametogenesis, mixed cultures of clones ofopposite mating type were fixed during gametogenesiswith Lugol’s iodine, formalin and sodium thiosulphatesolutions, according to Rassoulzagedan’s method(Sherr & Sherr, 1993). After a few hours’ fixation,cultures were stained with DAPI (0.5 mgml�1) for 20minand filtered gently onto black polycarbonate filters(pore size, 0.2 mm; Isopore GTBP membranes; Millipore,Massachusetts, USA) at low vacuum (<10kPa). Thefilters were then mounted in a drop of low fluorescence(halogen-free) immersion oil (Zeiss, Jena, Germany).
Frustules were cleaned by oxidation with hydrogenperoxide and glacial acetic acid and washed repeatedlywith distilled water before being mounted in Naphrax(PhycoTech, St Joseph, MI, USA). Light microscopical(LM) observations of live cells, cleaned frustules andpreparations stained with DAPI were carried out using
Fig 1. Location of the sampling stations (st 1–st 7) in the
North Sea.
V. A. Chepurnov et al. 380
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

a Zeiss Axioplan 2 Universal microscope (Zeiss, Jena,
Germany) equipped with a digital camera (VIP III,
Hamamatsu Photonics Deutschland, Herrsching,
Germany). The morphometric measurements presented
in Table 2 were made with the aid of ImageJ software
version 1.29x (http://rsb.info.nih.gov/ij/). Scanning
electron microscopy (SEM) was performed using a
JEOL JSM5600LV (JEOL, Tokyo, Japan).Voucher specimens of cleaned material of the original
natural samples and clonal cultures have been deposited
in the Laboratory of Protistology and Aquatic Ecology,
Ghent University, Belgium.
Terminology of mating types
The discovery of heterothally in many pennate diatoms
during the last decade (Chepurnov et al., 2004) has been
a considerable surprise, and terminology has not kept
pace with advances in knowledge. For example, there is
currently no consistency in the naming of mating
types. Mating types have sometimes been referred to
as ‘male’ and ‘female’, where there is a constant
difference between them in the behaviour of the gametes
(e.g. in araphid pennate diatoms: Chepurnov & Mann,
2004; Chepurnov et al., 2004). However, in some
heterothallic diatoms (e.g. Eunotia Ehrenberg: Mann
et al., 2003 and Seminavis D.G. Mann in Round,
Crawford et D.G. Mann: Chepurnov et al., 2002) the
gametes produced by opposite mating types behave alike
and look identical, so that designation as ‘male’ or
‘female’ is impossible. Furthermore, there is no
evidence as to whether, for example, the ‘male’ mating
types are equivalent in different taxa, even though this
seems likely among closely related species. Therefore,
we suggest a simple convention for naming mating
types in diatoms, via an acronym consisting of a few
letters representing the genus and species, followed by
(1) þ or �, if the mating system is simple and
Table 1. List of clones used. Each clone was isolated within one or two days of sampling
Clone Voucher Sampling station Date of sampling
First measurement of cell length
Date Size* (mm)
1 Trnz-1 st 4 24.05.2002 06.06.2002 127–130 (128.6� 0.96)
2 Trnz-2 st 4 24.05.2002 06.06.2002 131–134 (132.3� 1.04)
6 LP-6 st 1 01.05.2000 08.12.2003 78–85 (81.0� 2.22)
14 LP-14 st 2 25.04.2002 08.12.2003 117–120 (119.0� 1.16)
16 LP-16 st 3 22.05.2002 08.12.2003 111–120 (115.7� 2.72)
17 LP-17 st 3 22.05.2002 08.12.2003 115–122 (118.0� 1.77)
18 LP-18 st 3 22.05.2002 08.12.2003 108–114 (111.0� 1.95)
19 LP-19 st 3 22.05.2002 08.12.2003 118–122 (119.5� 1.51)
20 ZL130-20 st 5 30.06.2003 23.09.2003 93–97 (94.2� 1.32)
21 ZL130-21 st 5 30.06.2003 23.09.2003 84–88 (86.2� 1.48)
22 ZL130-22 st 5 30.06.2003 23.09.2003 94–94 (95.4� 1.17)
23 ZL130-23 st 5 30.06.2003 23.09.2003 80–83 (81.4� 1.18)
24 ZL130-24 st 5 30.06.2003 23.09.2003 84–87 (85.1� 0.99)
26 ZL130-26 st 5 30.06.2003 23.09.2003 85–89 (86.8� 1.23)
27 ZL130-27 st 5 30.06.2003 23.09.2003 92–95 (93.3� 0.95)
28 ZL130-28 st 5 30.06.2003 23.09.2003 86–89 (87.7� 1.16)
29 ZL130-29 st 5 30.06.2003 23.09.2003 92–96 (94.2� 1.32)
30 ZL130-30 st 5 30.06.2003 23.09.2003 85–91 (88.4� 2.46)
33 ZL130-33 st 5 30.06.2003 23.09.2003 95–97 (95.8� 1.14)
40 GFC-40 st 6 13.08.2003 24.09.2003 73–77 (75.4� 1.84)
44 GFC-44 st 6 13.08.2003 24.09.2003 82–86 (84.2� 1.55)
50 Ambl-50 st 7 13.09.2003 26.09.2003 98–102 (101.0� 1.33)
52 Ambl-52 st 7 13.09.2003 26.09.2003 90–92 (91.6� 0.84)
57 Ambl-57 st 7 13.09.2003 26.09.2003 90–92 (90.6� 0.84)
*Values are range (means� SD) of 10 measurements.
Table 2. Pseudo-nitzschia pungens: dimensions and stria and fibula densities* from integrated literature data (Fryxell & Hasle,2003), one of our field samples (from station 5) and four clonal cultures
Material analysed Apical length (mm) Width (mm) Striae (in 10 mm) Fibulae (in 10 mm)
Integrated data 74–174 2.4–5.3 9–16 9–16
Sample (N¼ 50) 86.3–160.8 (104.6� 10.42) 3.7–5.3 (4.5� 0.35) 10–13 (11.1� 0.68) 10–16 (12.8� 1.40)
Clone 1 (N¼ 10) 127.0–129.6 (128.6� 0.96) 3.8–4.4 (4.2� 0.18) 11–12 (11.3� 0.35) 11–13 (11.9� 0.57)
Clone 2 (N¼ 10) 130.9–133.7 (132.3� 1.04) 3.8–4.7 (4.3� 0.34) 11–13 (11.9� 0.74) 11–14 (12.1� 1.17)
Clone 24 (N¼ 20) 86.2–94.9 (90.3� 2.00) 3.1–4.4 (3.6� 0.36) 11–13 (12.1� 0.67) 10–15 (13.2� 1.24)
Clone 29 (N¼ 20) 97.1–101.4 (99.0� 1.25) 3.1–4.3 (3.6� 0.29) 11–14 (12.2� 0.71) 12–15 (13.3� 0.93)
*Values are range (means� SD).
Life history and mating system of Pseudo-nitzschia 381
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

gametangia produce either active or passive gametes, or(2) a number, if the gametangia are not obviously
differentiated, or (3) some other symbol that hasmeaning in relation to the characteristics of mating. InP. pungens, the two mating types differ in gametebehaviour and so we propose to designate them as PNpþ
and PNp�, ‘PNp’ being a simple contraction of thespecies binomial.
Results
Morphology and identification
In a freshly collected marine planktonic sample,cells of Pseudo-nitzschia can easily be recognizedby their characteristic motile ‘stepped’ colonies(e.g. Hasle & Syvertsen, 1996; Vrieling et al., 1996;Orsini et al., 2004). In our samples, P. pungens wassometimes accompanied by other Pseudo-nitzschiaspecies already known to occur in the North Sea
(e.g. Vrieling et al., 1996; Hasle, 2002), includingP. fraudulenta, Pseudo-nitzschia turgidula (Hustedt)Hasle and P. delicatissima. However, even in livesamples, it was quite easy to identify P. pungenscolonies. Cells of P. turgidula and P. delicatissimawere generally smaller and when the samples weredisturbed, P. turgidula and P. delicatissima coloniesdisintegrated almost completely into single cells.In contrast, the colonies of P. pungens andP. fraudulenta were robust. Even at low magnifica-tion, P. fraudulenta could be separated fromP. pungens through the extent to which the cellsoverlapped in the stepped colonies (c. 1/5 to 1/3of the cell length in P. pungens but by c. 1/6 to 1/8of the cell length in P. fraudulenta) (Figs 9–12 andsee Throndsen et al., 2003). In addition, the chainsof P. fraudulenta were straight whereas those ofP. pungens were always slightly bent when seen invalve view (Fig. 2). Finally, frustule morphologyand morphometric measurements made with LM
Figs 2–6. Pseudo-nitzschia pungens vegetative cells: LM (Figs 2, 3) or SEM (Figs 4–6). Fig. 2. Stepped chain of cells (live) ingirdle view, clone 18. Scale bar: 50mm. Fig. 3. Valve, clone 29. Note the coarse striae and the absence of a central interspace(the raphe slit is not interrupted by the central nodule). Scale bar: 20mm. Figs 4–6. Valve ultrastructure. Note the presence of
two rows of poroids per stria (Figs 4, 5), the single row of paired poroids on the proximal mantle (thick arrow on Figs 4–6),and single row of poroids (spaced like those of the valve face) on the distal mantle. The girdle of a complete theca consists ofthree bands (Fig. 6, thin arrows), each with a row of single poroids. Figs 4, 5. Clone 2: valve face in internal (Fig. 4) and
external (Fig. 5) views. Fig. 6. Clone 23: frustule in girdle view (the epitheca is marked with an arrowhead). Scale bar: 2mm.
V. A. Chepurnov et al. 382
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

(Table 2, Fig. 3) and SEM (Figs 4–6) showed thatall our isolates could be confidently identifiedas P. pungens (e.g. Hasle, 1995; Hasle & Syvertsen,1996; Fryxell & Hasle, 2003; Throndsen et al.,2003).
Chloroplasts of vegetative cells
The nucleus lay between the chloroplasts in thecentre of the cell and did not migrate at any stageof the mitotic cell cycle. Vegetative cells possessedtwo simple, plate-like chloroplasts, one in eachpolar half of the cell (Figs 7–12). During inter-phase, the chloroplasts lay adjacent to the girdle,with their margins extending slightly beneath thevalves (Figs 7–9). Usually, both were appressed tothe same side of the girdle (Fig. 7), but occasionallythey were located on opposite sides (Fig. 8).During cytokinesis, each chloroplast was splitinto two by the cleavage furrow (Fig. 10), so thateach daughter cell obtained half of each parentalplastid. Following cytokinesis, the chloroplastsshifted onto the valves of the parental frustule(the epivalves of the daughter cells) and remainedvalve-appressed (Fig. 11) until after the new valvehad been completed and the sibling cells hadseparated and slid apart to establish the steppedconfiguration of the mature colony (Fig. 12). Soonafter the translocation of the sibling cells had beencompleted, however, the chloroplasts returned tothe girdle-appressed position illustrated in Fig. 9.
Abrupt cell size reduction
In culture, P. pungens clones generally reduced insize by c. 5 mm (cell length) per month. In addition,however, cell size sometimes reduced abruptly,even by about half the cell length within a division.We observed the process in detail in clone 23.Abrupt size reduction was preceded by progressivechanges during several mitotic divisions. The firstsign was the appearance of a slight irregularity infrustule morphology near the cell centre, normallyin the form of a small bulge (Fig. 8). In subsequentcell divisions, cells became constricted in valveand girdle view, through the formation andprogressive deepening of an indentation (Figs 13,14), which finally caused the neighbouring chlor-oplast to split in two (Fig. 15). Cells containingthree plastids remained viable and continued todivide (Fig. 16). Next, the constriction became sodeep as to divide the entire protoplast into twoparts (Fig. 17), causing abortion of the part thatcontained one product of the divided chloroplastbut no nucleus (Fig. 18). During the next celldivision, therefore, hypovalves were formed thatwere shorter than the epivalves by the length of theaborted part (Fig. 19). Such cells with unequalthecae could not move apart from each other.At subsequent divisions, however, cells wereformed with equal, shortened valves and althoughshortened cells often remained together during afew cell cycles, forming very characteristic clusters
Figs 7–12. Live vegetative cells. Figs 7–9. Interphase chloroplasts. Figs 7, 8. Cells in valve view: the chloroplasts are appressed
to the same (Fig. 7) or opposite sides (Fig. 8) of the girdle. Fig. 9. Cell chain in girdle view. Fig. 10. Chloroplast division atmitotic cytokinesis, girdle view. Fig. 11. Cell division is complete and the chloroplasts have moved onto the epithecae of thenewly formed cells, girdle view. Fig. 12. Two sibling cells after rearrangement into the characteristic stepped configuration,
girdle view. Note that the chloroplasts are still appressed to the epivalves. Scale bars: 20 mm.
Life history and mating system of Pseudo-nitzschia 383
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

Fig 13–22. Abrupt cell size reduction in living vegetative cells, clone 23. Fig. 13. Cell of abnormal shape, valve view. Fig. 14.Two sibling cells of abnormal shape soon after division, girdle view. Note the constriction of the cells at the centre (arrow).Fig. 15. The lower chloroplast has been split into two, valve view. Fig. 16. Two sibling cells just after cell division, each
containing three chloroplasts, girdle view. Fig. 17. Separation of the protoplast into two parts (arrow), girdle view. Fig. 18.The separated protoplast fragment, which contained one product of a divided chloroplast but lacked a nucleus (arrow),has aborted; valve view. Fig. 19. Division of a cell with a partly aborted protoplast has resulted in the formation of short
hypothecae, girdle view. Fig. 20. Typical cluster of abruptly reduced cells formed by three mitoses subsequent to abruptsize reduction, girdle view. Figs 21, 22. Stepped colonies of abruptly reduced cells in valve (Fig. 21) and girdle views (Fig. 22).Scale bars: 20mm.
V. A. Chepurnov et al. 384
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

(Fig. 20), they soon began to form typical motile,stepped colonies (Figs 21, 22).During and immediately after abrupt size reduc-
tion, the chloroplasts were unequal in size and thenucleus shifted from the cell centre towards the endwhere the smaller plastid lay (Figs 19, 20). Later,the cells corrected the asymmetry (Fig. 22) andwhen we isolated a short colony of abruptlyreduced cells and grew it separately (clone 23s)it appeared healthy and grew well, though a littlemore slowly than unreduced clones. Long chainsof the short cells often spiralled.
Mating behaviour
Sexual reproduction never occurred in monoclonalcultures and auxosporulation was initiated only inparticular combinations of clones: P. pungens isheterothallic. Results of repeated crosses of theclones in most of the possible pair-wise combina-tions (only clone 23 was not included in all tests)revealed that 12 clones belonged to one matingtype (PNpþ) and 11 to the opposite mating type(PNp�) (Table 3). However, not all combinationsof PNpþ and PNp� led to auxosporulation andone clone (clone 40) exhibited particularly unusualbehaviour, which is described below.The gametes were morphologically identical,
regardless of mating type, but those produced
by PNpþ gametangia were active (‘male’); theymigrated out of the gametangial frustule tofuse with the gametes of PNp– gametangia,which were passive (‘female’), remaining withinthe compartment formed by the gametangialfrustule (Figs 30, 31).The mating behaviour of clone 40 was anom-
alous. Clone 40, like other clones, was isolated as asingle chain and, as with other clones, neverreproduced sexually in monoclonal cultures, sothat there was no indication of heterogeneitywithin it. However, cells of clone 40 becamesexualized if grown together with any other clone,whether of PNpþ or PNp�. When clone 40 wasmixed with larger-celled clones (see Table 1),e.g., PNpþ clones 6 (Figs 23–26, 28) or 20, orPNp� clones 17 (Figs 27, 29–32) or 52, it waspossible to investigate how cells of each clonebehaved sexually in mixed cultures. These experi-ments showed that cells of clone 40 were able topair amongst themselves and reproduce success-fully, as well as with the PNp� and PNpþ clones.Clone 40 cells behaved as ‘male’ when crossed with‘female’ PNp� clones and as ‘female’ in crosseswith ‘male’ PNpþ clones. In intraclonal pairings,sexual reproduction was physiologically aniso-gamous, as in crosses between normal PNp� andPNpþ clones, so that some of the clone 40 cellsbehaved as male and others as female.
Table 3. Results of mating experiments
Mating type Clone 1 16 17 19 24 29 30 52 28 14 23* 2 6 18 20 21 22 26 27 33 44 50 57 40s 40**
PNp� 1 [0]
PNp� 16 0 [0]
PNp� 17 0 0 [0]
PNp� 19 0 0 0 [0]
PNp� 24 0 0 0 0 [0]
PNp� 29 0 0 0 0 0 [0]
PNp� 30 0 0 0 0 0 0 [0]
PNp� 52 0 0 0 0 0 0 0 [0]
PNp� 28 0 0 0 0 0 0 0 0 [0]
PNp� 14 0 0 0 0 0 0 0 0 0 [0]
PNp� 23* nt nt nt nt 0 nt nt nt nt nt [0]
PNpþ 2 þþ þþ þþ þþ þþ þþ þþ þþ 0 0 nt [0]
PNpþ 6 þ þþ þþ þþ þ þ þ þ þ þþ nt 0 [0]
PNpþ 18 þþ þ þ þ þþ þþ þþ þþ þþ þ nt 0 0 [0]
PNpþ 20 þ þþ þþ þþ þ þ þþ þ þþ 0 nt 0 0 0 [0]
PNpþ 21 þþ þþ þþ þþ þ þþ þ þþ 0 0 nt 0 0 0 0 [0]
PNpþ 22 þ þþ þþ þþ þþ þ þþ þ þþ þþ nt 0 0 0 0 0 [0]
PNpþ 26 þþ þþ þþ þþ þ þ þ þ þ 0 þþ 0 0 0 0 0 0 [0]
PNpþ 27 þ þþ þþ þþ þ þ þ þ 0 0 nt 0 0 0 0 0 0 0 [0]
PNpþ 33 þ þþ þþ þþ þ þ þþ þ 0 0 nt 0 0 0 0 0 0 0 0 [0]
PNpþ 44 þþ þþ þþ þþ þ þþ þ þþ 0 0 nt 0 0 0 0 0 0 0 0 0 [0]
PNpþ 50 þþ þþ þþ þþ þþ þþ þþ þþ 0 þþ nt 0 0 0 0 0 0 0 0 0 0 [0]
PNpþ 57 þ þþ þþ þþ þ þ þ þ 0 0 nt 0 0 0 0 0 0 0 0 0 0 0 [0]
PNpþ 40 s þþ þþ þþ þþ þþ þþ þþ þþ þþ 0 nt 0 0 0 0 0 0 0 0 0 0 0 0 [0]
? 40** þ þ þ þ þ þ þ þ þ þ nt þ þ þ þ þ þ þ þ þ þ þ þ 0 [0]
þ, sexual reproduction occurred in mixed culture; þþ, crosses where we could determine (because of size differences between the clones) that
all matings were interclonal; 0, sexual reproduction absent in mixed culture; [0], sexual reproduction absent in monoclonal culture; nt, cross
not made; *, subculture of clone 23, which was obtained after abrupt size reduction (see text); **, the complex mating behaviour is described
in the text.
Life history and mating system of Pseudo-nitzschia 385
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12
Remarkably, clone 40 was also capable ofintraclonal reproduction in mixed cultures withmonoclonal cultures of either mating type ofP. fraudulenta (which is also heterothallic:Chepurnov et al., 2004) and when grown with astrain of P. multiseries. In these cases, however,unlike in P. pungens crosses, there was nointerclonal pairing; cells belonging to differentspecies could not interbreed.After the anomalous behaviour of clone 40 was
confirmed by the results of repeated set of crosses
(all possible pair-wise combinations with the otherP. pungens clones available (Table 3) and withP. fraudulenta and P. multiseries), we decidedto re-isolate a subclone from a single short(four-cell) colony. Surprisingly, the new culture(subclone 40s) exhibited strictly heterothallicbehaviour and its cells always behaved as ‘males’,like those of a normal PNpþ clone, producinggametangia with active gametes while interactingwith PNp� (Table 3, Figs 30–32). However, in across between subclone 40s and the parental
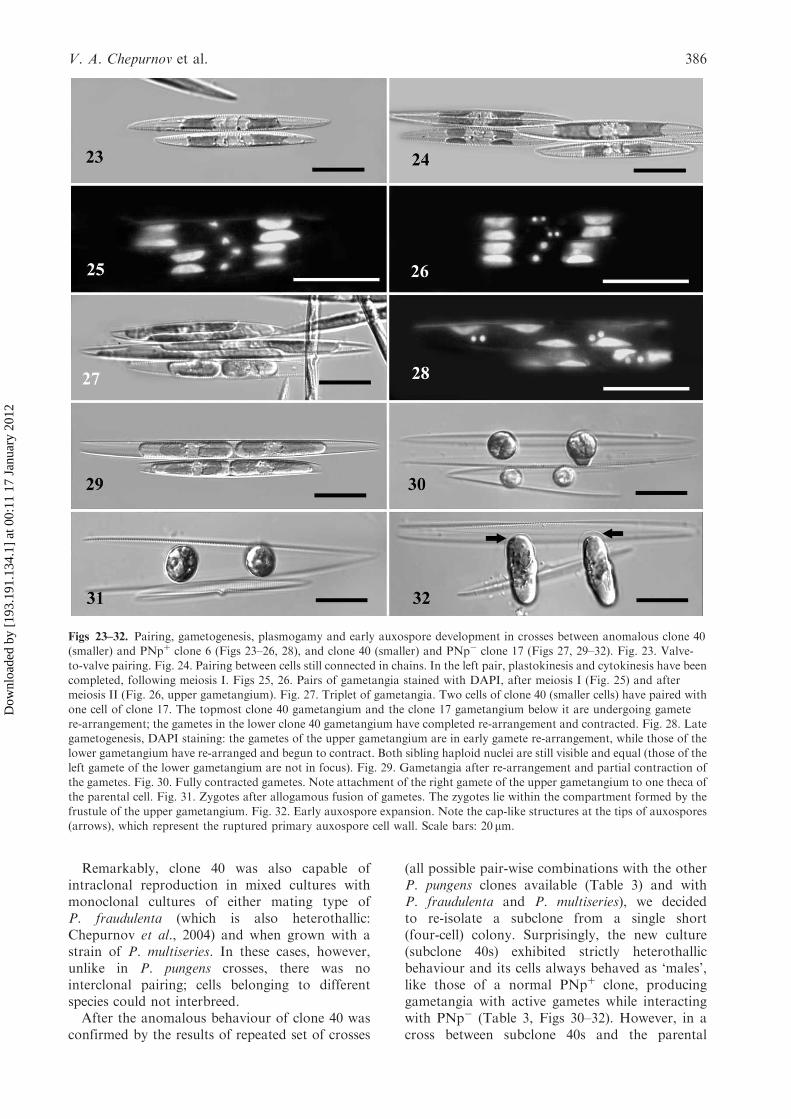
Figs 23–32. Pairing, gametogenesis, plasmogamy and early auxospore development in crosses between anomalous clone 40(smaller) and PNpþ clone 6 (Figs 23–26, 28), and clone 40 (smaller) and PNp� clone 17 (Figs 27, 29–32). Fig. 23. Valve-
to-valve pairing. Fig. 24. Pairing between cells still connected in chains. In the left pair, plastokinesis and cytokinesis have beencompleted, following meiosis I. Figs 25, 26. Pairs of gametangia stained with DAPI, after meiosis I (Fig. 25) and aftermeiosis II (Fig. 26, upper gametangium). Fig. 27. Triplet of gametangia. Two cells of clone 40 (smaller cells) have paired with
one cell of clone 17. The topmost clone 40 gametangium and the clone 17 gametangium below it are undergoing gametere-arrangement; the gametes in the lower clone 40 gametangium have completed re-arrangement and contracted. Fig. 28. Lategametogenesis, DAPI staining: the gametes of the upper gametangium are in early gamete re-arrangement, while those of the
lower gametangium have re-arranged and begun to contract. Both sibling haploid nuclei are still visible and equal (those of theleft gamete of the lower gametangium are not in focus). Fig. 29. Gametangia after re-arrangement and partial contraction ofthe gametes. Fig. 30. Fully contracted gametes. Note attachment of the right gamete of the upper gametangium to one theca ofthe parental cell. Fig. 31. Zygotes after allogamous fusion of gametes. The zygotes lie within the compartment formed by the
frustule of the upper gametangium. Fig. 32. Early auxospore expansion. Note the cap-like structures at the tips of auxospores(arrows), which represent the ruptured primary auxospore cell wall. Scale bars: 20 mm.
V. A. Chepurnov et al. 386
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

clone 40, no signs of sexualization were noted, aswithin clone 40 itself.Clones 14 and 28 were also anomalous. Both
were unisexual PNp� clones (Table 3) but wouldnot interbreed with all of the available PNpþ
clones. Clone 14 bred only with PNpþ clones 6, 18,22 and 50 and never interacted sexually with otherPNpþ clones. Cells of clone 28 were less selectivebut still recognized only PNpþ clones 6, 18, 20, 22,26 and 40 s as compatible. Unsuccessful crosses offemale clones 14 and 28 with PNpþ clones wererepeated at least four times for each pair-wisecombination and were made alongside otherPNpþ�PNp� crosses, using the same subculturesand conditions (usually different wells of a singleRepli dish) as for crosses in which the same clonesexhibited sexual activity.The effect of abrupt size reduction on sexual
competence was examined using clone 23s (seeabove). Clone 23s cells were 42–44 mm in length,in contrast to the original clone 23 cells from whichthey were derived, which were 76–79mm. Despitethis, clone 23s, like clone 23, exhibited strictlyfemale behaviour while crossed with PNpþ clone26 (Table 3), and produced viable offspring.
Pattern of sexual reproduction
The first signs of transition to sexual reproductionusually appeared 2–4 days after compatible cloneswere mixed and involved mass disintegration of thestepped colonies into single cells or (more rarely)short 2–3-cell chains, and sinking of most cellsto the bottom of the container. Hence, pairingwas mostly between single cells (Fig. 23), but thefew cells that remained connected in chains werealso capable of sexualization (Fig. 24). Formationof triplets, where a single cell interacted with twocells of the opposite mating type, also occurred(Fig. 27).The predominant configuration of sexual part-
ners was valve-to-valve (Figs 23, 24); infrequently,valve-to-girdle and girdle-to-girdle configurationswere also observed (not illustrated). Meiosis I ingametangia was accompanied by equal plasto- andcytokinesis (Figs 24–26), which visually resembledthe equivalent mitotic events (Fig. 10). Meiosis IIfollowed shortly after the first meiotic division andinvolved karyokinesis alone (Fig. 26, upper game-tangium). Next, the gametes became rearrangedwithin the parental frustule (Figs 27, 28) andgradually contracted (Figs 27, 29). Finally, theybecame spherical (Fig. 30) and at this stage DAPIstaining revealed that most gametes contained onlyone ‘functional’ nucleus; the other haploid producthad already begun to abort, becoming smaller andcondensed, or had already disappeared completely(not illustrated).
Following the completion of gametogenesis, thegametes from one gametangium migrated towardsthe other gametangium and allogamous fusion ofgametes occurred within the compartment formedby the gametangium (Fig. 31). Sexual reproductionwas therefore physiologically anisogamous and thegametangia could be regarded as ‘male’ (usuallyonly from PNpþ clones) and female (usually onlyfrom PNp� clones). Sometimes, female gametesretained contact with the thecae of the parentalfrustule (Fig. 30) but complete separation of femalegametes from the walls of gametangial frustule,before plasmogamy, was also observed regularly.After a few hours the zygotes started to expand,becoming auxospores. Expansion was bipolar andthe auxospores remained with their tips fixed inthe space between the thecae of the female (PNp�)gametangia (Fig. 32), probably indicating thepresence of mucous material, which, however,could not be detected by LM in our unstainedmaterial. The sibling auxospores tended to expandparallel to each other and perpendicular to theapical axes of the parental gametangia. Theauxospores possessed cap-like structures on theirtips, representing the ruptured organic wall of thezygote (Fig. 32), and expansion was accompaniedby the formation of a perizonium of delicatetransverse bands (Fig. 42).The two functional haploid nuclei inherited from
the gametes became and remained closely asso-ciated with each other at the centre of auxosporesand were easily visible throughout auxosporeexpansion (Fig. 33). Fusion seemed to take placeafter expansion was complete. In the early stages ofexpansion, the four chloroplasts were arrangedapparently at random. Later, they became alignedin a single longitudinal row, with two on either sideof the (still unfused) nuclei (Fig. 33).
Initial cell formation
After expansion, the auxospore contracted awayfrom the transverse perizonium and the initialepivalve was laid down on this side while the line ofchloroplasts lay opposite (Fig. 34). Following this,the chloroplasts shifted onto the girdle region (asdefined by the position of the initial epitheca),stayed there for a while (Fig. 35) and then, still in asingle line, moved synchronously onto the epivalve(Fig. 36). Next, the hypovalve was laid down(Fig. 37) after a second, lesser contraction of theprotoplast. The chloroplasts then moved back tothe girdle (as in the configuration shown for theearlier stage in Fig. 35) but subsequently becamere-arranged within the initial cell, the two apicalchloroplasts shifting onto one valve while the twocentral ones moved onto the other (Figs 38, 39).The first division of the initial cell followed.
Life history and mating system of Pseudo-nitzschia 387
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

This division, unlike other mitotic cytokineses,was not accompanied by plastokinesis (Fig. 40),so that the two daughter cells contained twochloroplasts, just like ordinary vegetative cells.The first division of the initial cell could occur
either before (Fig. 40) or after escape from theperizonium, which ruptured at one pole allowingthe initial cell to slide out (Fig. 41). Once the initialcell had escaped, it became obvious that a lon-gitudinal perizonium is present in P. pungens,
Figs 33–42. Auxospore and initial cell formation, in a cross between clones 40 and 6. Fig. 33. Auxospore at a late stage ofexpansion. Note the two unfused haploid nuclei at the centre. Figs 34–39. Initial cell formation, girdle view. Note the epivalve
is uppermost. Fig. 34. The contents of auxospore have contracted and the epivalve has been laid down. The chloroplasts lieon the cell side opposite to the epitheca. Fig. 35. Shift of the chloroplasts onto the girdle of the future initial cell. Fig. 36.The chloroplasts have moved onto the epivalve. Fig. 37. Completion of initial hypovalve. Fig. 38. Re-arrangement of thechloroplasts: the two marginal plastids have moved onto the hypovalve and the two central plastids onto the epivalve. Fig. 39.
Re-alignment of chloroplasts complete. Fig. 40. Division of initial cell within the transverse perizonium. Fig. 41. Initial cellescaping from the perizonium before the first cell division. Fig. 42. Transverse perizonium (arrowhead) and longitudinalperizonium (arrow), after the escape of the initial cell. Scale bars: 20 mm.
V. A. Chepurnov et al. 388
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

appearing in LM as a single, elongate, smoothstrip (Fig. 42).During initial cell formation, the nucleus was
central. Deposition of each initial valve waspreceded by an acytokinetic mitosis, with quickabortion of one of the products (not illustrated).
‘Cardinal’ points of the life-cycle
The cardinal points represent critical points in thelife-cycle, where changes in physiological statusoccur in relation to cell size (Geitler, 1932;von Stosch, 1965; Chepurnov et al., 2004). Thefirst cardinal point represents the size of the initialcells. 76 normally-formed initial cells (for abnor-malities in initial cell formation, see below) weremeasured, giving a range of 157.1–176.3mm(Table 4). To check for any relationship betweengametangium size and initial cell size, we measuredinitial cells in two mixed cultures of P. pungens, inwhich the gametangia were large (clone 16� clone18: gametangia 101–112mm long) or relativelysmall (clone 6� clone 40: gametangia 61–78 mmlong). The lengths of the initial cells were almostidentical (Table 4).The second cardinal point is the upper size
threshold for sexual reproduction, below whichcells are sexually inducible given appropriategrowth conditions. The largest cells that trans-formed into gametangia were in PNp� clone 19.This clone, isolated when its cells were c. 120mmlong (Table 1), exhibited no signs of sexualizationin repeated crossing experiments until the cells hadreduced to 109.4–115.3mm (112.9� 1.59) long. In across between clone 19 and clone 40s, the largestgametangium of clone 19 was c. 115 mm.The third cardinal point is the critical minimal
size for sexualization, or the critical minimal sizebelow which cells are not viable, whichever comesfirst. In P. pungens, cells died when they werec. 25–30mm long.
Atypical behaviour during sexual reproduction
During the development of the initial cell, partof the protoplast adjacent to one tip of the cellsometimes aborted (Fig. 43). This process, which
did not involve loss of any of the chloroplasts, didnot harm the further development of the initial cell,which was therefore much shorter than theauxospore containing it (Fig. 44).Once, an expanding auxospore was found that
contained only two plastids and a single nucleus(Fig. 45). This auxospore had presumably devel-oped from a single unfertilized gamete. We rarelybut regularly (in almost every vigorously repro-ducing mixed culture that we examined in detail)observed the formation of triploid auxospores,by fusion of three gametes. The triploid cells werecapable of auxospore expansion, even thoughthey contained one more nucleus and two moreplastids than normal auxospores (Fig. 46, compareFig. 33). We did not follow the complete devel-opment of any individual triploid auxospores.However, we saw several initial cells with completefrustules containing six chloroplasts (Fig. 47),instead of the four usually present (Figs 38, 39).These initial cells had presumably developed fromthe triploid auxospores.
Discussion
Chloroplast division and arrangement
Chloroplast division is brought about in P. pungensby inward growth of the cleavage furrow duringcytokinesis. This imposes chloroplast division sothere are two lineages of chloroplasts within eachclone of P. pungens cells, ‘dual’ inheritance sensuMann (1996). Dual inheritance occurs in manyother Bacillariaceae (Mann, 1996 and unpublisheddata), and in some Nitzschia Hassall species(e.g. Nitzschia sigmoidea (Nitzsch) W. Smith andNitzschia linearis W. Smith: Pickett-Heaps &Tippit, 1980; Round et al., 1990) chloroplastdivision is imposed by the cleavage furrow, as inPseudo-nitzschia. Elsewhere, however, chloroplastdivision is at least partly autonomous, constrictionof the chloroplast into two being achieved justbefore the cell is divided by the cleavage furrowin Nitzschia palea (Kutzing) W. Smith and twoother Nitzschia species (Geitler, 1975).A linear arrangement of the chloroplasts, resem-
bling that in P. pungens auxospores (Figs 33–37),has been shown in the young auxospores of N.palea (Geitler, 1928, fig. 11, as Nitzschia subtilis(Kutzing) Grunow: for notes on the identification,see Geitler, 1973), Nitzschia amphibia Grunow(Geitler, 1969, fig. 2), Nitzschia frustulum var.perpusilla (Rabenhorst) Grunow in Van Heurck(Geitler, 1970) and Nitzschia recta Hantzsch exRabenhorst (Mann, 1986, fig. 14). The presence ofa similar arrangement in such a wide spread of taxamay indicate that the arrangement is typical for
Table 4. Parental and initial cell lengths* for two crosses,
clone 6� clone 40 (21.12.2003) and clone 16� clone 18(21.01.2004)
Parental clone Cell length** (mm) Initial cell length (mm)
6 72.3–77.9 (74.4� 2.38) 157.1–176.3
40 61.4–63.5 (62.5� 0.65) (165.6� 4.24) N¼ 50
16 107.1–112.0 (109.5� 1.63) 159.4–175.2
18 101.9–107.8 (104.6� 1.86) (167.7� 4.77) N¼ 26
*Values are range (means� SD); **20 cell measurements per clone.
Life history and mating system of Pseudo-nitzschia 389
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

Bacillariaceae (or at least the biplastidic species).However, further information is desirable. Theorchestrated movements of the chloroplasts fromside to side within the expanded auxospore as theinitial cells form have not been reported previouslyfrom Bacillariaceae, as far as we are aware. Thepattern of translocation of pairs of chloroplastsprior to division of the initial cell indicates tightcontrol on chloroplast inheritance, though wecannot say whether this leads to mixing of thetwo chloroplast lineages from each gametangiumor ensures their segregation.
Size changes during the life-cycle andabrupt size reduction
The maximal length of P. pungens initial cellsobtained experimentally (176.3mm) agrees wellwith the maximum length found in nature, i.e.174mm (Fryxell & Hasle, 2003; see also Table 2).The variation in size of the initial cells (of c. 30mm)is not surprising (e.g. Mann et al., 1999, 2003;
Edlund & Bixby, 2001). In some diatoms, acorrelation has been reported between the sizesof the initial cells and those of the gametangiaproducing them: small gametangia tend toproduce small initial cells (e.g. Roshchin, 1994;Davidovich, 1994, 2001; Nagai et al., 1995; Edlund& Bixby, 2001). However, this is not a universalrule, as demonstrated here in P. pungens and alsoin some other diatoms, such as Tabularia tabulata(C. Agardh) Snoeijs (Roshchin, 1994), where initialcell length remains� constant, despite considerablevariation in gametangium size. In Neidium amplia-tum (Ehrenberg) Krammer, in which each gam-etangium produces one active and one passivegamete (trans physiological anisogamy), the size ofthe initial cell is related to the size of thegametangium in which it forms (i.e. the gametan-gium contributing the passive gamete), but not tothe size of the gametangium contributing the activegamete (Mann & Chepurnov, 2005).As in most diatom studies so far, vegetative
multiplication of P. pungens was generally
Figs 43–47. Anomalous development during auxosporulation, clone 40� clone 6. Figs 43, 44 Abrupt size reduction at initialcell formation. Fig. 43. Partial abortion of auxospore cell contents at one pole (arrow). Fig. 44. The initial cell is shorter than
the developed auxospore and longitudinal perizonium (arrow). Fig. 45. Auxospore containing two chloroplasts and a singlenucleus. Fig. 46. Auxospore containing six chloroplasts and three nuclei (at centre). Fig. 47. Initial cell containing sixchloroplasts. Scale bars: 20 mm.
V. A. Chepurnov et al. 390
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

accompanied by gradual reduction in cell size(according to the Macdonald–Pfitzer rule:Crawford, 1981; Chepurnov et al., 2004).However, abrupt size reduction also occurred.Abrupt cell size reduction has previously beenreported in several lineages of both centric andpennate diatoms (e.g. von Stosch, 1965; Roshchin,1994; Mann et al., 2003; Chepurnov et al., 2004).In diatom cultures, abrupt size reduction can occurspontaneously or can be initiated experimentallyby regulation of the culture density, nutrientlimitation, or delicate ‘surgical intervention’ (e.g.von Stosch, 1965; Chepurnov et al., 2004). Theecological significance of this phenomenon is notunderstood. However, two consequences of thisphenomenon are obvious – reduction of theduration of the life-cycle and, if abrupt sizereduction concerns large cells from the sexually‘insensitive’ part of life-cycle, the possibility ofrapid or immediate sexualization (e.g. Mannet al., 2003).In P. pungens, two methods of abrupt cell size
reduction were detected. The first occurred duringinitial cell formation (see Figs 43, 44) and willshorten the purely vegetative phase of the life-cycle.The second occurred in clones that were alreadywithin the sexual size range (Figs 13–20), althoughwe cannot exclude the possibility that similarreduction may occur in larger cells as well. Theappearance of a constriction that intensified andfinally led to abrupt cell size reduction could beregarded as an artefact of culturing. However, thecells of the original colony from which clone 23was initiated already contained a small but visibleirregularity in shape at the centre and similarabnormalities, as depressions or slight bulges at thecell centre, were also noted (although very infre-quently) in cells from natural samples. Similarabnormalities in cell shape have also beenillustrated in cultures of other Pseudo-nitzschiaspecies, e.g., P. multiseries (Subba Rao et al., 1991,figs 1B and 1C). Thus, abrupt size reduction islikely to occur here as well.
Pattern of sexual reproduction
Sexual reproduction has now been reported for sixspecies of Pseudo-nitzschia (Davidovich & Bates,1998; Chepurnov et al., 2004; Amato et al., 2005).In all of them, the pattern of auxosporulationcorresponds to Geitler’s (1973) type IA2, in whichthe gametangia produce two gametes apiece andare differentiated into ‘male’ (producing activegametes) and ‘female’ (producing passive gametes).Pseudo-nitzschia species are also similar in pairingconfiguration, gametogenesis and the associationof the developing auxospores with the frustule ofthe female gametangium. The species studied
include representatives of all three of the majorclades within the genus that have been detected byanalysis of rDNA sequence data (from the ITS1–5.8S–ITS2 region: Lundholm et al., 2003). Hencewe can predict that type IA2 auxosporulation willbe found to be characteristic of the whole genus.The uniformity of sexual reproduction within asingle, morphologically well-defined, monophyleticgenus is not surprising and has been found, forexample, in Licmophora C. Agardh (Chepurnov& Mann, 2004), Eunotia (Mann et al., 2003),Sellaphora Mereschkowsky (Mann, 1989),Achnanthes C. Agardh (Sabbe et al., 2004),Cocconeis Ehrenberg, Cymbella C. Agardh andGomphonema C. Agardh (Geitler, 1973). However,the uniformity within Pseudo-nitzschia contrastsstrongly with the variation already evident withinthe closely related genus Nitzschia, despite the factthat few Nitzschia species (c. 12) have beenexamined in detail (Geitler 1932, 1973; Mann,1986; Roshchin, 1994). Some Nitzschia species pairside-to-side, as in Pseudo-nitzschia (examples areN. recta and N. longissima (Brebisson ex Kutzing)Grunow: Karsten, 1897; Mann, 1986; Chepurnovin Roshchin, 1994), but N. amphibia pairs end-to-end. Plasmogamy occurs within a diffuse mucilageenvelope in N. fonticola Grunow in Van Heurck(Geitler, 1932) and N. linearis (Mann, unpublisheddata), as in Pseudo-nitzschia, but in the type speciesof Nitzschia, N. sigmoidea, and in several otherspecies, plasmogamy occurs via narrow copulationtubes formed at the centre (N. sigmoidea) or at onepole (N. amphibia) (Geitler, 1969; Mann, 1986).Some species produce one gamete per gametan-gium, others two (Geitler, 1932, 1973). However,the Bacillariaceae, to which Nitzschia and Pseudo-nitzschia belong, is an ancient group that wasalready diverse in the upper Eocene (Schrader,1969; Desikachary & Sreelatha, 1989) and, in thepartial LSU rDNA gene tree presented byLundholm et al. (2002), Nitzschia is paraphyleticwith respect to Pseudo-nitzschia. Hence hetero-geneity in sexual reproduction within Nitzschiamay simply reflect the fact that the revision ofNitzschia by Round et al. (1990), in whichTryblionella W. Smith and Psammodictyon D.G.Mann in Round, Crawford et D.G. Mann as wellas Pseudo-nitzschia and Fragilariopsis Hustedt inA. Schmidt et al. were separated from Nitzschia,did not go far enough.Kaczmarska et al. (2000) made a detailed SEM
investigation of the gametes, auxospores and initialcells of P. multiseries, which is apparently theclosest relative of P. pungens (Lundholm et al.,2003), but found no longitudinal perizonium,only a transverse perizonium. A longitudinalperizonium is present in P. pungens, however
Life history and mating system of Pseudo-nitzschia 391
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

(Figs 42, 44), suggesting that re-examination ofP. multiseries may be worthwhile.
Mating behaviour
Understanding mating systems, including whethermating occurs randomly or assortatively and howsexual partners are related to each other (out-breeding v. inbreeding), is key to the interpretationof data on the genetic structure of species and theirevolutionary potential. Remarkably, all six Pseudo-nitzschia species studied so far are heterothallic(Davidovich & Bates, 1998; Chepurnov et al., 2004;Amato et al., 2005). The only exceptions are theanomalous bisexual behaviour of our P. pungensclone 40 and a report of sexual reproduction in amixture of clones of a single mating type in theotherwise heterothallic P. calliantha (Davidovich &Bates, 1998, as ‘P. pseudodelicatissima’ from theBlack Sea). In contrast, the few other complexes ofclosely related pennate species that have beenstudied intensively with respect to breeding behav-iour, in Sellaphora, Achnanthes and Eunotia, aremore variable in their mating systems, exhibitingboth intra- and interspecific variation in matingbehaviour (Mann, 1999; Chepurnov et al., 2004;Mann et al., 2004; Sabbe et al., 2004).The behaviour of clone 40 raises interesting
questions about mating type and sex determina-tion. The fact that clone 40 never reproduced inmonoclonal culture (either as pure clone 40 or withsubclone 40s) indicates that it was not expressingall features of both mating types. Yet it was able tomate with both PNpþ and PNp� clones in biclonalcultures. Its behaviour as a ‘normal’ male PNpþ
clone after re-isolation (as subclone 40s) from asingle chain is even more curious. Though there islittle secure evidence, most observations made sofar suggest that sex or mating type determinationin pennate diatoms is basically genetic (Chepurnovet al., 2004). The behaviour of clone 40 suggestsperhaps that some mating types are heteroallelicand that some mating type alleles in theseheteroallelic mating types are usually epigeneticallysilenced. Occasionally, however, suppression (orexpression) is incomplete or temporary, leading to‘abnormal’ or inconsistent behaviour. The behav-iour of clones 14 and 28 show further complexities.Clone 14 was among the longest-celled clonesisolated and previous observations (e.g. Mannet al., 1999; Chepurnov et al., 2002) have suggestedthat sexualization becomes progressively easier ascells become significantly smaller than the sexualsize threshold. Hence, some failures might beexpected among tests involving clones such asclone 14, at least initially. However, clone 14 wasinitially similar in size to clone 19, which was ‘wellbehaved’, and clone 28 was relatively small-celled
when isolated. Furthermore, clones 14 and 28 bothfailed to reproduce when mixed with the initiallysmall-celled clone 44.The message from our data, therefore, is that
interesting aspects of the mating system may bemissed if only a few clones are isolated and tested.Although the overall picture is clear � thatP. pungens is heterothallic � there are subtletiesin mating type determination, which may besignificant.Further descriptive studies of mating behaviour
are needed in Pseudo-nitzschia species but signifi-cant progress in understanding the microevolutionand geographical distribution of the genus willdepend more on population genetic studies anddeveloping molecular markers for sex-relatedevents in the natural environment (Armbrust &Galindo, 2001; see also Swanson & Vacquier,2002; Barrett, 2002). However, confirmation ofheterothally (which will enforce outbreeding) in sixPseudo-nitzschia species is consistent with thedemonstration of high genotypic diversity inP. calliantha (Skov et al., 1997, as P. pseudodeli-catissima), P. pungens (Lundholm et al., 2003,p. 811; Evans et al., 2004a) and P. multiseries(Evans et al., 2004b).
What is P. pungens?
A recent review of species-level taxonomy indiatoms concluded that species boundaries haveoften been drawn too widely (Mann, 1999). Theextent to which this is true has been examinedusing various systems, including the freshwatergenus Sellaphora and the marine diatomsSkeletonema Greville, Achnanthes and DiploneisEhrenberg ex Cleve (see Mann, 1999). Knowledgegained from such systems can then be used tohelp interpret variation elsewhere, where timeand money do not permit detailed examinationusing less intensive (and admittedly less sure)approaches. Pseudo-nitzschia has now becomeestablished as another system, because of thegrowing corpus of work stemming from toxico-logical concerns.Recently, two distinctive, widely distributed
‘morphotypes’ of Pseudo-nitzschia have beenstudied for correspondence between genetic andmating data. Based on purely morphological data,definition of these two diatoms – P. pseudodelica-tissima and P. delicatissima – remained ambiguous(Hasle, 2002). Large-scale studies of P. pseudo-delicatissima were therefore undertaken, usingmorphology and molecular data (ITS sequences)and taking into account the results of somecrossing experiments (Davidovich & Bates, 1998).The outcome was the description of two newspecies, P. calliantha and P. caciantha, and
V. A. Chepurnov et al. 392
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

emendation of P. pseudodelicatissima and themorphologically similar Pseudo-nitzschia cuspidata(Hasle) Hasle (Lundholm et al., 2003).Subsequently, intensive studies were made ofP. delicatissima from the Gulf of Naples(Mediterranean Sea), again using a mixture ofmorphological, mating and molecular approaches(Orsini et al., 2004). The data showed thepresence of five distinct lineages within what hadappeared to be a single morphospecies; this crypticdiversity within P. delicatissima requires furtherstudy and may lead to the description of severalnew species.P. pungens is distinctive morphologically, has
been reported worldwide (e.g. Hasle, 2002), andappears to be the sister group to P. multiseries(Lundholm et al., 2003). We undertook the presentwork as a preliminary to a molecular geneticstudy of P. pungens, particularly along the Belgiancoastline but also from other localities. Evans et al.(2004a) have developed microsatellite markers forP. pungens revealing high levels of genetic diversitybut apparently no variation that would indicatecryptic speciation. Among our 24 clonal cultures,isolated at different times and from differentlocalities off the north-western coast of Europe,there was no sign of heterogeneous morphologicalvariation, and the sexual compatibility of clonesand the viability of the F1 progeny (our unpub-lished observations) also suggest that all the clonesinvestigated belong to a single species. We haveobtained ITS1-5.8S-ITS2 sequences from at leastone clone from each of the seven locations ofFig. 1, including clones 14, 28 and 40, and allproved to be identical (Casteleyn et al., 2004).Regular sampling and isolation of multiple (c. 60)clones during blooms of P. pungens along theBelgian coast were undertaken during spring 2004and preliminary analyses of morphology, molecu-lar sequences and results of crossing experimentsgive no indication of any taxonomically significantvariation. We have also investigated clones fromthe Atlantic coast of Spain, New Zealand and thePacific coast of the USA and found all to besexually compatible, with no obvious loss ofviability in the F1 generation.
Polyploidy and haploid parthenogenesis
Polyploidy and haploid parthenogenesis havenot been reported previously in Pseudo-nitzschiabut seem to be not uncommon among diatoms(e.g., Mann, 1994; Chepurnov et al., 2004). Thesignificance and evolutionary consequences of suchchanges in ploidy are not clear, although duringa review of the few chromosome counts availablefor diatoms, Kociolek and Stoermer (1989) sug-gested that polyploidy might play an important
role in diatom evolution. Polyploidy is a regularthough infrequent event in sexually reproducingcultures of P. pungens. It occurred more frequentlyduring mating experiments in P. fraudulenta(unpublished). The ease with which sexuality canbe initiated experimentally in Pseudo-nitzschia andthe regularity with which ploidy changes occurduring auxosporulation in P. pungens and P.fraudulenta make Pseudo-nitzschia an attractivesystem in which to study polyploidy in greaterdetail.
Acknowledgements
Financial support for this research was providedlargely by the Research Programmes G.0292.00and G.0197.05 of the Fund for Scientific Research –Flanders (Belgium), and BOF-project GOA12050398 (Ghent University, Belgium). GrietCasteleyn is funded by the Institute for thePromotion of Innovation by Science andTechnology in Flanders (Belgium). Collaborationwith Drs N. Lundholm (University ofCopenhagen, Denmark), V. Trainer (NorthwestFisheries Science Center, Seattle, WA), M. Lastra(Universidade de Vigo, Spain), L. Rhodes andK. Ponikla (the Cawthron Institute CultureCollection of Micro-algae, Nelson, New Zealand)is greatly appreciated.
References
Amato, A., Orsini, L., D’Alelio, D. & Montresor, M. (2005).
Life-cycle, size reduction patterns, and ultrastructure of the
pennate planktonic diatom Pseudo-nitzschia delicatissima
(Bacillariophyceae). J. Phycol., 41: 542–556.
Armbrust, E.V. & Galindo, H.M. (2001). Rapid evolution of a
sexual reproduction gene in centric diatoms of the genus
Thalassiosira. Appl. Environ. Microbiol., 67: 3501–3513.
Barrett, S.C.H. (2002). The evolution of plant sexual diversity.
Nat. Rev. Genet., 3: 274–284.
Bates, S.S. (2000). Domoic-acid-producing diatoms: another genus
added! J. Phycol., 36: 978–985.
Casteleyn, G., Chepurnov, V.A., Sabbe, K., Mann, D.G.,
Vannerum, K. & Vyverman, W. (2004). Species structure of
Pseudo-nitzschia pungens (Grunow ex Cleve) Hasle: molecular,
morphological and reproductive data. In Abstracts, 18th
International Diatom Symposium (Witkowski, A., Radziejewska,
T., Wawrzyniak-Wydrowska, B., Daniszewska-Kowalczyk, D.
& Ba�k, M., editors), 98. Mie�dzyzdroje, Poland, 2–7 September
2004.
Chepurnov, V.A. & Mann, D.G. (2004). Auxosporulation of
Licmophora communis (Bacillariophyta) and a review of mating
systems and sexual reproduction in araphid pennate diatoms.
Phycol. Res., 52: 1–12.
Chepurnov, V.A., Mann, D.G., Vyverman, W., Sabbe, K. &
Danielidis, D.B. (2002). Sexual reproduction, mating system
and protoplast dynamics of Seminavis (Bacillariophyceae).
J. Phycol., 38: 1004–1019.
Chepurnov, V.A., Mann, D.G., Sabbe, K. & Vyverman, W.
(2004). Experimental studies on sexual reproduction in diatoms.
Int. Rev. Cytol., 237: 91–154.
Life history and mating system of Pseudo-nitzschia 393
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

Crawford, R.M. (1981). Some considerations of size reduction in
diatom cell walls. In Proceedings of 6th International Symposium
on Living and Fossil Diatoms (Ross, R., editor), 253–265. Koeltz
Scientific Books, Koenigstein.
Davidovich, N.A. (1994). Factory, opredelyayuschie razmer
initsial’nykh kletok u diatomovykh vodoroslej. Russ. J. Plant
Physiol., 41: 250–255.
Davidovich, N.A. (2001). Species-specific sizes and size range of
sexual reproduction in diatoms. In Proceedings of the 16th
International Diatom Symposium (Economou-Amilli, A., editor),
191–196. Amvrosiou Press, Athens.
Davidovich, N.A. & Bates, S.S. (1998). Sexual reproduction in
the pennate diatoms Pseudo-nitzschia multiseries and P. pseudo-
delicatissima (Bacillariophyceae). J. Phycol., 34: 126–137.
Desikachary, T.V. & Sreelatha, P.M. (1989). Oamaru diatoms.
Biblioth. diatomol., 19: 1–330.
Edlund, M.B. & Bixby, R.J. (2001). Intra- and inter-specific
differences in gametangial and initial cell size in diatoms. In
Proceedings of the 16th International Diatom Symposium
(Economou-Amilli, A., editor), 169–190. Amvrosiou Press,
Athens.
Elbra« chter, M. (2003). Dinophyte reproduction: progress and
conflicts. J. Phycol., 39: 629–632.
Evans, K.M., Bates, S.S., Medlin, L.K. & Hayes, P.K. (2004a).
Genetic diversity in two Pseudo-nitzschia species. In Abstracts,
18th International Diatom Symposium (Witkowski, A.,
Radziejewska, T. Wawrzyniak-Wydrowska, B., Daniszewska-
Kowalczyk, D. & Ba�k, M., editors), 31. Mie�dzyzdroje, Poland,2–7 September 2004.
Evans, K.M., Bates, S.S., Medlin, L.K. & Hayes, P.K. (2004b).
Microsatellite marker development and genetic variation in the
toxic marine diatom Pseudo-nitzschia multiseries
(Bacillariophyceae). J. Phycol., 40: 911–920.
Fehling, J., Davidson, K., Bolch, C. J. & Bates, S.S. (2004).
Growth and domoic acid production by Pseudo-nitzschia seriata
(Bacillariophyceae) under phosphate and silicate limitation. J.
Phycol., 40: 674–683.
Fryxell, G.A. & Hasle, G.R. (2003). Taxonomy of harmful
diatoms. In Manual on Harmful Marine Microalgae. Monographs
on Oceanographic Methodology (Hallegraeff, G.M., Andersen,
D.M. & Cembella, A.D., editors), 11: 465–509. UNESCO, Paris.
Geitler, L. (1928). Copulation und Geschlechtsverteilung bei
einer Nitzschia-Art. Arch. Protistenk., 61: 419–442.
Geitler, L. (1932). Der Formwechsel der pennaten Diatomeen.
Arch. Protistenk., 78: 1–226.
Geitler, L. (1969). Auxosporenbildung von Nitzschia amphibia.
Osterr. Bot. Z., 117: 404–410.
Geitler, L. (1970). Padogame Automixis und Auxosporenbildung
bei Nitzschia frustulum var. perpusilla. Osterr. Bot. Z., 118:
121–130.
Geitler, L. (1973). Auxosporenbildung und Systematik bei
pennaten Diatomeen und die Cytologie von Cocconeis-Sippen.
Osterr. Bot. Z., 122: 299–321.
Geitler, L. (1975). Lebendbeobachtung der Chromato-
phorenteilung der Diatomee Nitzschia. Plant Syst. Evol.,
123: 145–152.
Guillard, R.R.L. (1975). Culture of phytoplankton for feeding
marine invertebrates. In Culture of Marine Invertebrate Animals
(Smith, W.L. & Chanley, M.H., editors), 29–60. Plenum,
New York.
Hasle, G.R. (1995). Pseudo-nitzschia pungens and P. multiseries
(Bacillariophyceae): nomenclatural history, morphology, and
distribution. J. Phycol., 31: 428–435.
Hasle, G.R. (2002). Are most of the domoic acid-producing
species of the diatom genus Pseudo-nitzschia cosmopolites?
Harmful Algae, 1: 137–146.
Hasle, G.R. & Syvertsen, E.E. (1996). Marine diatoms.
In Identifying marine diatoms and dinoflagellates (Tomas, C.R.,
editor), 5–385. Academic Press, San Diego.
Hiltz, M., Bates, S.S. & Kaczmarska, I. (2000).
Effect of light:dark cycles and cell apical length on
the sexual reproduction of the pennate diatom
Pseudo-nitzschia multiseries (Bacillariophyceae) in culture.
Phycologia, 39: 59–66.
Kaczmarska, I., Bates, S.S., Ehrman, J.M. & Leger, S. (2000).
Fine structure of the gamete, auxospore and initial cell in the
pennate diatom Pseudo-nitzschia multiseries (Bacillariophyta).
Nova Hedwigia, 71: 337–357.
Karsten, G. (1897). Untersuchungen uber Diatomeen III.
Flora, 83: 203–222.
Kociolek, J.P. & Stoermer, E.F. (1989). Chromosome numbers
in diatoms: a review. Diatom Res., 4: 47–54.
Lundholm, N., Daugbjerg, N. & Moestrup, Ø. (2002).
Phylogeny of the Bacillariaceae with emphasis on the genus
Pseudo-nitzschia (Bacillariophyceae) based on partial LSU
rDNA. Eur. J. Phycol., 37: 115–134.
Lundholm, N., Moestrup, Ø., Hasle, G.R. & Hoef-Emden, K.
(2003). A study of the Pseudo-nitzschia pseudodelicatissima/
cuspidata complex (Bacillariophyceae): what is P. pseudodelica-
tissima? J. Phycol., 39: 797–813.
Lundholm, N., Hansen, P.J. & Kotaki, Y. (2004). Effect of pH
on growth and domoic acid production by potentially toxic
diatoms of the genera Pseudo-nitzschia and Nitzschia. Mar. Ecol.
Progr. Ser., 273: 1–15.
Maldonado, M.T., Hughes, M.P., Rue, E.L. & Wells, M.L.
(2002). The effect of Fe and Cu on growth and domoic acid
production by Pseudo-nitzschia multiseries and Pseudo-nitzschia
australis. Limnol. Oceanogr., 47: 515–526.
Mann, D.G. (1986). Methods of sexual reproduction in Nitzschia:
systematic and evolutionary implications (Notes for a mono-
graph of the Bacillariaceae 3). Diatom Res., 1: 193–203.
Mann, D.G. (1989). The diatom genus Sellaphora: separation from
Navicula. Br. Phycol. J., 24: 1–20.
Mann, D.G. (1994). Auxospore formation, reproductive
plasticity and cell structure in Navicula ulvacea and the
resurrection of the genus Dickieia (Bacillariophyta). Eur. J.
Phycol., 29: 141–157.
Mann, D.G. (1996). Chloroplast morphology, movements and
inheritance in diatoms. In Cytology, genetics and molecular
biology of algae (Chaudhary, B.R. & Agrawal, S.B., editors),
249–274. SPB Academic Publishing, Amsterdam.
Mann, D.G. (1999). The species concept in diatoms (Phycological
Reviews 18). Phycologia, 38: 437–495.
Mann, D.G. (2002). Diatom life cycles. In Proceeding of the
LIFEHAB Workshop: Life History of Microalgal Species Causing
Harmful Algal Blooms (Garces, E., Zingone, A., Montresor, M.,
Reguera, B. & Dale, B., editors), 13–17. Commission of the
European Community, www.icm.csic.es/bio/projects/lifehab.
Mann, D.G. & Bates, S.S. (2002). Life cycles in diatoms.
In Proceeding of the LIFEHAB Workshop: Life History of
Microalgal Species Causing Harmful Algal Blooms (Garces, E.,
Zingone, A., Montresor, M., Reguera, B. & Dale, B., editors),
117–120. Commission of the European Community, www.icm.
csic.es/bio/projects/lifehab.
Mann, D.G. & Chepurnov, V.A. (2004). What have the Romans
ever done for us? The past and future contribution of culture
studies to diatom systematics. Nova Hedwigia, 79: 237–291.
Mann, D.G. & Chepurnov, V.A. (2005). Auxosporulation,
mating system, and reproductive isolation in Neidium
(Bacillariophyta). Phycologia, 44: 335–350.
Mann, D.G., Chepurnov, V.A. & Droop, S.J.M. (1999).
Sexuality, incompatibility, size variation, and preferential poly-
andry in natural populations and clones of Sellaphora pupula
(Bacillariophyceae). J. Phycol., 35: 152–170.
Mann, D.G., Chepurnov, V.A. & Idei, M. (2003). Mating
system, sexual reproduction, and auxosporulation in the
anomalous raphid diatom Eunotia (Bacillariophyta). J. Phycol.,
39: 1067–1084.
Mann, D.G., McDonald, S.M., Bayer, M.M., Droop,
S.J.M., Chepurnov, V.A., Loke, R.E., Ciobanu, A. &
du Buf, J. M. H. (2004). Morphometric analysis, ultra-
structure and mating data provide evidence for five new
species of Sellaphora (Bacillariophyceae). Phycologia,
43: 459–482.
V. A. Chepurnov et al. 394
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12

Nagai, S., Hori, Y., Manabe, T. & Imai, I. (1995). Restoration of
cell size by vegetative cell enlargement in Coscinodiscus wailesii
(Bacillariophyceae). Phycologia, 34: 533–535.
Orellana, M.V., Petersen, T.W. & van den Engh, G. (2004).
UV-excited blue autofluorescence of Pseudo-nitzschia multiseries
(Bacillariophyceae). J. Phycol., 40: 705–710.
Orsini, L., Procaccini, G., Sarno, D. & Montresor, M. (2004).
Multiple rDNA ITS-types within the diatom Pseudo-nitzschia
delicatissima (Bacillariophyceae) and their relative abundances
across a spring bloom in the Gulf of Naples. Mar. Ecol. Prog.
Ser., 271: 87–98.
Parsons, M.L., Dortch, Q. & Turner, R.E. (2002).
Sedimentological evidence of an increase in Pseudo-nitzschia
(Bacillariophyceae) abundance in response to coastal eutrophica-
tion. Limnol. Oceanogr., 47: 551–558.
Pickett-Heaps, J.D. & Tippit, D.H. (1980). Light and electron
microscopic observations on cell division in two large pennate
diatoms, Hantzschia and Nitzschia. I. Mitosis in vivo. Eur. J. Cell
Biol., 21: 1–11.
Roshchin, A.M. (1994). Zhiznennye Tsikly Diatomovykh
Vodoroslej. Naukova Dumka, Kiev.
Round, F.E., Crawford, R.M. & Mann, D.G. (1990). The
Diatoms. Biology and morphology of the genera. Cambridge
University Press, Cambridge.
Sabbe, K., Chepurnov, V.A., Vyverman, W. & Mann, D.G.
(2004). Apomixis in Achnanthes (Bacillariophyceae); develop-
ment of a model system for diatom reproductive biology.
Eur. J. Phycol., 39: 327–341.
Schrader, H.-J. (1969). Die pennaten Diatomeen aus dem
Obereozan von Oamaru, Neuseeland. Nova Hedwigia, Beih.,
28: 1–124.
Sherr, E.B. & Sherr, B.F. (1993). Preservation and storage of
samples for enumeration of heterotrophic protists. In Handbook
of Methods in Aquatic Microbial Ecology (Kemp, P.F.,
Sherr, B.F., Sherr, E.B. & Cole, J.J., editors), 207–212. Lewis
Publishers, Boca Raton.
Skov, J., Lundholm, N., Pocklington, R., Rosendahl, S. &
Moestrup, Ø. (1997). Studies on the marine planktonic diatom
Pseudo-nitzschia. 1. Isozyme variation among isolates of
P. pseudodelicatissima during a bloom in Danish coastal
waters. Phycologia, 36: 374–380.
Subba Rao, D.V., Partensky, F., Wohlgeschaffen, G. &
Li, W.K.W. (1991). Flow cytometry and microscopy of
gametogenesis in Nitzschia pungens, a toxic, bloom-forming,
marine diatom. J. Phycol., 27: 21–26.
Swanson, W.J. & Vacquier, V.D. (2002). The rapid evolution of
reproductive proteins. Nat. Rev. Genet., 3: 137–144.
Throndsen, J., Hasle, G. R. & Tangen, K. (2003). Norsk
Kystplanktonflora. Almater Forlag AS, Oslo.
von Stosch, H.A. (1965). Manipulierung der Zellgrosse von
Diatomeen im Experiment. Phycologia, 5: 21–44.
Vrieling, E.G., Koeman, R.P., Scholin, C.A., Scheerman, P.,
Peperzak, L., Veenhuis, M. & Gieskes, W.W.C. (1996). Identi-
fication of a domoic acid-producing Pseudo-nitzschia species
(Bacillariophyceae) in the Dutch Wadden Sea with electron
microscopy and molecular probes. Eur. J. Phycol., 31: 333–340.
Life history and mating system of Pseudo-nitzschia 395
Dow
nloa
ded
by [
193.
191.
134.
1] a
t 00:
11 1
7 Ja
nuar
y 20
12
Related Documents











