1 Sex-specific behavioral syndromes allow the independent evolution of behavioral dimorphism Raphael Royauté 1,2,4 Ann Hedrick 3 Ned A. Dochtermann 2 1 Movement Ecology Group, Senckenberg Biodiversity and Climate Research Centre (SBiK- F), Frankfurt, Germany (current address) 2 Department of Biological Sciences; North Dakota State University, Fargo, USA 3 Department of Neurobiology, Physiology, and Behavior; University of California, Davis, USA 4 corresponding author: [email protected] Running Title: Sex-specific syndromes and behavioral sexual dimorphism Keywords: Behavioral syndromes, evolutionary constraints, G matrix, personality, behavioral ecology, sexual dimorphism, intralocus sexual conflicts Author Contributions: All authors conceived the project and supervised the gathering of data. R.R conducted behavioral trials and analyzed the data. R.R. wrote the first draft of the manuscript and all authors contributed to subsequent versions. Conflicts of interests: The authors declare no conflict of interests Data Accessibility Statement: All data, code and analyses are available at: https://osf.io/pnug5/ Acknowledgements: We thank Monica Berdal, Katelyn Cannon, Jeremy Dalos, Sarah Felde, 1 Brady Klock, Ishan Joshi, Hannah Lambert, Jenna LaCoursiere and Alondra Neunsinger for 2 assistance in conducting behavioral trials and in rearing and care of the crickets and 3 Martori Farms, David Lightfoot, Scott Bundy, Nico Franz, Sangmi Lee, Cameron Jones, 4 Kenny Chapin, Ti Eriksson, Meranda Feagins, Charlotte Mulloney, Melody Martinez, Allyson 5 Richins, Mauriel Rodriguez, Helen Vessels and David Wikman for assistance in collecting 6 the crickets. We thank for J. Sztepanacz and A.J. Wilson for their advice on statistical 7 analyses and K. Laskowski for comments on an early draft of this manuscript. This work 8 was supported by US NSF IOS grants 1557951 and 1558069 to N.A.D. and A.H. respectively. 9

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
Sex-specificbehavioralsyndromesallowtheindependentevolutionofbehavioraldimorphism
RaphaelRoyauté1,2,4AnnHedrick3
NedA.Dochtermann2
1MovementEcologyGroup,SenckenbergBiodiversityandClimateResearchCentre(SBiK-F),Frankfurt,Germany(currentaddress)2DepartmentofBiologicalSciences;NorthDakotaStateUniversity,Fargo,USA3DepartmentofNeurobiology,Physiology,andBehavior;UniversityofCalifornia,Davis,USA4correspondingauthor:[email protected]:Sex-specificsyndromesandbehavioralsexualdimorphismKeywords:Behavioralsyndromes,evolutionaryconstraints,Gmatrix,personality,behavioralecology,sexualdimorphism,intralocussexualconflictsAuthorContributions:Allauthorsconceivedtheprojectandsupervisedthegatheringofdata.R.Rconductedbehavioraltrialsandanalyzedthedata.R.R.wrotethefirstdraftofthemanuscriptandallauthorscontributedtosubsequentversions.Conflictsofinterests:TheauthorsdeclarenoconflictofinterestsDataAccessibilityStatement:Alldata,codeandanalysesareavailableat:https://osf.io/pnug5/Acknowledgements:WethankMonicaBerdal,KatelynCannon,JeremyDalos,SarahFelde,1
BradyKlock,IshanJoshi,HannahLambert,JennaLaCoursiereandAlondraNeunsingerfor2
assistanceinconductingbehavioraltrialsandinrearingandcareofthecricketsand3
MartoriFarms,DavidLightfoot,ScottBundy,NicoFranz,SangmiLee,CameronJones,4
KennyChapin,TiEriksson,MerandaFeagins,CharlotteMulloney,MelodyMartinez,Allyson5
Richins,MaurielRodriguez,HelenVesselsandDavidWikmanforassistanceincollecting6
thecrickets.WethankforJ.SztepanaczandA.J.Wilsonfortheiradviceonstatistical7
analysesandK.Laskowskiforcommentsonanearlydraftofthismanuscript.Thiswork8
wassupportedbyUSNSFIOSgrants1557951and1558069toN.A.D.andA.H.respectively. 9

2
ABSTRACT10
Whenselectiondiffersbysex,thecapacityforsexestoreachoptimalphenotypescanbe11
constrainedbythesharedgenomeofmalesandfemales.Becausephenotypictraitsare12
oftencorrelated,thisdifferenceextendsacrossmultipletraitsandunderlyinggenetic13
correlationscanfurtherconstrainevolutionaryresponses.Behaviorsarefrequently14
correlatedasbehavioralsyndromes,andthesecorrelationsoftenhaveageneticbasis.15
However,whethercross-sexandacrossbehaviorcorrelationsleadtoconstrainedevolution16
remainsunknown.Here,weshowthataboldness-activitysyndromeisstronglysex-17
specificatthegeneticlevelinthewesternfieldcricket(Gryllusinteger)andthatemergence18
fromashelterisgeneticallyindependentbetweenmalesandfemales.However,male19
activityisstronglyrelatedtofemaleshelteremergence,creatingthepotentialforbiased20
responsestoselection.Ourresultsshowthatthesex-specificgeneticarchitectureof21
behavioralsyndromescanshapetheevolutionofbehavioralphenotypes.22
23

3
INTRODUCTION24
Malesandfemalessharethesamegenomebutoftenrelyondifferentstrategiestoincrease25
fitness.Thiscanleadtointralocussexualconflictswhereselectionactingonasharedtrait26
displacesonesexfromitsoptimum(HedrickandTemeles1989;Bondurianskyand27
Chenoweth2009).Thissexualconflictisfurthercomplicatedbytheconstrainingpotential28
ofgeneticcorrelations(Lande1980;WalshandBlows2009).Forexample,behaviorsare29
oftencorrelatedascomponentsofabehavioralsyndrome(Sihetal.2004)andthese30
behavioralsyndromeshavethepotentialtoalterevolutionaryoutcomes(Dochtermann31
andDingemanse2013).32
Ingeneral,anysustainedselectionfavoringdifferentoptimabetweensexes,i.e.33
sexuallydiscordantselection,willeventuallyresolvethesexualconflictandattenuate34
geneticconstraintsovertime,thusallowingsexestoevolveindependently(Delphetal.35
2011).Thisconstraintcanbeformallymeasuredasthecross-sexcorrelationcoefficientrmf,36
withvalues<1increasingtherapidityatwhichsexualdimorphismcanevolve.Cross-sex37
correlationsaregenerallylargebuttendtodecreaseinspecieswithstrongersexual38
dimorphism(Poissantetal.2010).Additionalconstraintscanemergewhengeneralizingto39
multiplephenotypesexpressedinmalesandfemalesandtheirinteractions(Lande1980).40
Bydecomposingtheadditivegeneticcovariancematrixintoitssex-specific(Gm,Gf)and41
cross-sexsub-matrices(B),onecanestimateifgeneticcorrelationsacrosssexesandtraits42
createconstrainedevolutionaryoutcomes(Fig.1).43
Thisdecompositionapproachhasbeenusedformorphologicaltraits,revealingthat44
cross-sexcovariances(i.e.theBmatrix,(Lande1980))canprofoundlyaltertheevolution45
ofsexualdimorphism(Gosdenetal.2012;Bergeretal.2014;GosdenandChenoweth46
2014),especiallywhenselectionfavorsdifferentaveragetraitvaluesforeachsex(Long47
andRice2007;Bergeretal.2014).However,theGm,Gf,andBhaverarelybeenestimated48
forbehaviors.Howmuchthegeneticarchitectureofbehavioralsyndromescanaffect49
evolutionaryoutcomesunderintralocussexualconflictremainsunclear.Thisisan50
importantomissionbecausebehaviorfrequentlyimpactsbothsurvivalandfitness(Moiron51
etal.2020)andsexualdimorphismiscommonforbehaviors(Blanckenhorn2005;Aragón52

4
2011;Mainwaringetal.2011;Kokrasetal.2012).Despitemanyconceptualargumentsfor53
whybehavioralsyndromesshoulddifferbysexes(Schuettetal.2010;Hämäläinenetal.54
2018;Immonenetal.2018),sex-differencesintheexpressionofbehavioralcorrelations55
arefrequentlyignoredinpractice.Asaresult,thedegreetowhichcross-sexgenetic56
correlationsmightbeshapedbysexualconflictforbehavioralphenotypesremains57
unknownandtheimportanceofthesecorrelationsfortheevolutionofbehaviorsis58
similarlyunknown. 59
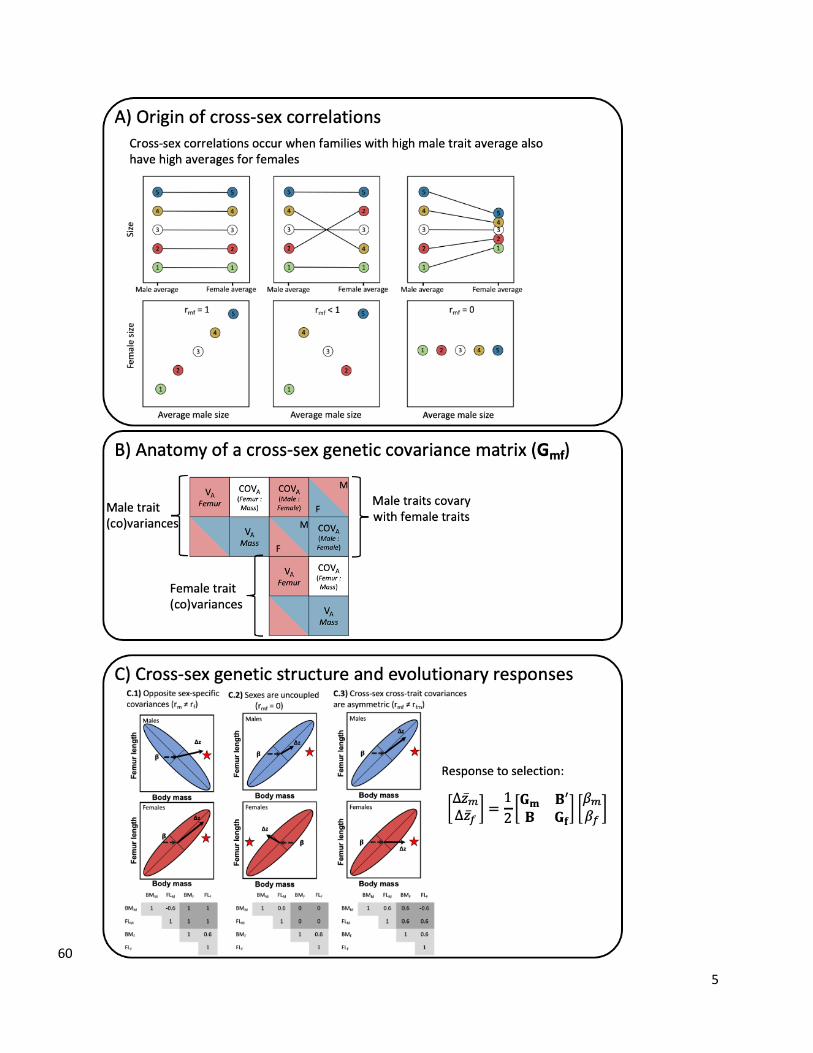
5
60

6
Figure1.Thegeneticarchitectureofcross-sexcorrelationscanhavenon-intuitive61 consequencesonevolutionaryresponsesandsexualdimorphism.A)Highvaluesofcross-62 sexcorrelationsforthesametraits(here,bodysize)indicatethatmaleandfemaleaverages63 arepositivelycorrelatedamongfamilies.B)Thecross-sexcovariancematrix(Gmf)allows64 tocomparepatternsofgeneticcovarianceswithinsexes(herebetweenbodymassand65 bodysize)aswellasunderstandthestrengthofcross-traitcorrelationswithinandamong66 traits.C)Cross-sexcross-traitcorrelation(rmf)canproducenon-intuitiveresponsesto67 selectivepressures.Hereweshow3scenariosindicativeofthevarietyofresponsesto68 selection(Δz)thatcanoccurdependingonthedirectionofselection(β)andthemagnitude69 andorientationofrmf.Theellipsesrepresentthebivariate(co)variationinfemurlength70 andmass,withthepopulationaverageatthevertexofthe(thinsolid)linesindicatingthe71 directionsinbivariatespacewiththemostvariation.Dashedlinesrepresentthedirection72 selection(β)ispushingapopulation’saverageandthesolidthickarrowsshowthe73 directionandmagnitude(arrowtip)ofselectionresponses.InbothscenariosC.1andC.2,74 selectionisconcordant—i.e.actingthesame—betweensexesandmalesandfemaleshave75 thesamefitnessoptimum.InscenarioC.3,selectionissexuallydiscordantandfavors76 increasedsexualdimorphism.ScenarioC.1showsthatstrongcross-sexcrosstrait77 correlationscanbiassexesawayfromtheiroptimum.InscenarioC.2,nullrmfresultsin78 independenttrajectoriesbysexes.InscenarioC.3,theeffectoffathersontheirdaughters’79 phenotypeisoppositeofthatofmother’sontheirsons’,resultinginasymmetricrmfand80 biasedresponsestoselectioninmales. 81

7
Intralocussexualconflictshouldbeparticularlystronginthefieldcrickets82
(Gryllidae).Inmanyfieldcrickets,malesandfemalesdifferintheirbehaviorsand83
reproductivestrategies(HedrickandKortet2012).Forexample,femalesexploretheir84
environmentstosamplemateswhilemalesremainatburrowsfromwhichtheysignal85
(FrenchandCade1987).Further,aboldness-activitysyndromehasaconservedgenetic86
basisinatleastonespecies,theWesternstutter-trillingfieldcricket(Gryllusinteger),87
whichseemstohaveconstrainedbehavioraldivergence(Royautéetal.2020).Whetherthis88
syndromeissex-specificiscurrentlyunknownbutthecombinationofconserved89
syndromestogetherwithalackofsex-specificitywouldfurtherconstraintheabilityof90
populationstorespondtolocalselectivepressures.91
Hereweusedbehavioralmeasurementsofover960Gryllusintegerindividualsto92
estimatetheinfluenceofcross-sexgeneticcorrelationsonevolutionaryresponses.We93
predictedthatcricketswouldexhibitdimorphisminaverageactivitygiventhemate-94
samplingbehavioroffemalesandthatgeneticvarianceforthistraitwouldbelowerfor95
femalecricketsduetoselectionfavoringincreasedtraveling.Wealsopredictedsex-96
specificityinthebehavioralsyndrome,withastrongerpositivegeneticcorrelation97
betweenantipredatorresponseandactivityinfemales.Sinceexitfromrefugiaandactivity98
shouldhaveastrongerinfluenceonfemalefitness,wealsoexpectedastrongercorrelation99
betweenshelteremergencewithactivityandantipredatorresponseinfemales.Although100
therearefewestimatesforcross-sexcorrelationsofbehaviors,thoseavailablesuggest101
behaviorsareundersimilarconstraintsasotherphenotypes(Poissantetal.2010).We102
thereforeexpectedthatcross-sexgeneticcorrelations(rmf)wouldnotdepartsignificantly103
from1.Wetestedthesepredictionsbyestimatingthecross-sexcovariancematrix,i.e.Gmf,104
decomposedintoitssex-specificandcross-sexsub-matrices(Gm,GfandBsubmatrices,105
Figure1).Wecomparedthestrengthofcovariancesamongsexesandtheeffectsofthese106
covariancesonresponsestoselectionusingrandomskeweranalysis.107
108

8
METHODS109
Wecollectedadultfemalecricketsfromfourpopulationsthroughoutthesouthwesternand110
westernUS:Socorro,NM;LasCruces,NM;Aguila,AZ;andDunnigan,CA(Figure2)during111
thesummerof2017andhousedthemandinourlaboratoryfacilitiesatNorthDakotaState112
University.Femaleswerehousedindividuallyin0.71Lcontainersandprovidedwithad113
libitumfood(PurinaChickStarter)andwater(waterwasprovidedinglassvialscapped114
withcotton).Eachcricketwasalsoprovidedwithasmallpieceofcardboardeggcartonfor115
shelter.Thecrickethousingroomwasmaintained~27Cona12:12dark:lightcycle116
reversedsuchthattheroomwasdarkduringdaytimehours.Werunoffspringofthis117
parentalgenerationthroughmultiplebehavioraltrialsbeforematingindividualsatrandom118
withineachpopulation.Werepeatedthisprocessfortwoadditionalgenerations.119
Behavioraltesting120
Latencytoemergefromshelter121
Gryllidcricketsusesmallburrowsandnaturalcracksasrefugestowhichtheyretreatwhen122
underthreat.Thetimetakentoemergefromashelterafterdisturbancecanthereforebe123
consideredaproxyforrisk-takingbehavioror“boldness”(KortetandHedrick2007).We124
transferredindividualsfromtheirhomecontainersintosmallartificialburrows(40cm3)125
placedwithina34.6×21cmarena.Weleftthecricketstorestfortwominutesafterwhich126
weremovedthecapfromtheburrowandletindividualsemerge.Wethenrecordedhow127
longittookforanindividualtoemerge(inseconds)foruptosixminutesandthirty128
seconds.Individualsthatdidnotemergeweregivenamaximumlatencyof390seconds.129
Openfieldexploratorybehavior130
Weusedopenfieldteststomeasureactivityandexploratorypropensityina30×30cm131
plexiglassarena.Thesetestsareclassicbehavioralassayacrosstaxa(WalshandCummins132
1976)andcanrevealstrongamong-individualdifferencesinexplorationpatterns,133
includingincrickets(Royautéetal.2015,2019;RoyautéandDochtermann2017).134
Individualsthatmovethroughmoreofthearenaareconsideredmorethoroughexplorers135
(Réaleetal.2007).Weintroducedindividualsintothearenaandleftthemtorestundera136

9
smallcontainerfor30seconds.Attheendofthis30seconds,weremovedthecontainer137
andthecricketwasallowedtoexplorethearenafor3minutesand40seconds.Thearena138
wascleanedwithisopropylalcoholbetweentrialstoremoveanychemosensorycuesfrom139
thearena.WeusedEthovisionXTtorecordthetotaldistancetheindividualmovedduring140
thetrial(cm).141
Responsetocuesofpredatorpresence142
Wemeasuredtheresponsetocuesofpredatorpresence,abehavioralassaycommonly143
usedinGryllidspeciestodetermineantipredatorresponse(RoyautéandDochtermann144
2017;Royautéetal.2019).Specifically,individualswereintroducedintoa15cmdiameter145
circulararena(7.5cmheight),thefloorofwhichwascoveredwithdryfilterpaperthathad146
beensoakedwithdilutedexcretafromleopardgeckos(Eublepharismacularius).Crickets147
respondtoexposuretoleopardgeckocuesbyincreasingactivityandindividualswith148
higherdistancemovedareconsideredmoreresponsivetothecue(Royautéand149
Dochtermann2017;Royautéetal.2019).Weintroducedcricketstoaportionofthearena150
withoutpredatorcueandleftthemtorestunderasmallshelterfor30seconds.Wethen151
removedtheshelterandallowedtheindividualallowedtofreelymovethroughoutthe152
arenafor3minutesand40seconds.WethenusedEthovisionXTtorecordthetotal153
distanceanindividualmovedduringthetrial(cm).154
155
Statisticalanalyses156
AllanalyseswereperformedusingRversion4.0.3(Rcitation)usingtheMCMCglmm157
package (Hadfield2010).158
Estimationofcross-sexgeneticcovariances(Gmf)159
Weusedamulti-responsemixedeffectanimalmodels(Kruuk2004;Wilsonetal.2010)to160
estimategeneticvariancesandcovariances(i.e.theGmfmatrix).Weincludedtheeffectsof161
temperature,dayandtimeoftestinginthebehavioralarenaroomalongwithsex,life-stage162
andmassoftheindividualasfixedeffects.Weusedtheindividualrelatednessmatrix163

10
(basedontheknownpedigree)asarandomeffectandthefollowingbehavioraltraitswere164
includedasresponsevariables:(i)thelatencythatanindividualemergedfromtheshelter165
duringthetrial(modeledascensoredGaussian),(ii)thedistancemovedduringtheopen166
fieldtrial(Gaussian),(iii)thedistanceanindividualmovedduringthepredatorcue167
responsetrial(Gaussian).Toestimatebothsex-specificandcross-sexcovariances,we168
treatedthebehaviorofeachsexasaseparatetrait-resultingintheestimationofa6×6169
covariancematrix.Weranthemulti-responsemodelwithanMCMCchainof4.8×106170
iterations,withan800,000burn-inperiodandathinningintervalof4,000andweuseda171
parameterexpandedpriorthatwasminimallyinformativeforbothvariancesand172
covariances.Allvariancesandcovarianceswereestimatedattheadditivegeneticleveland173
onthelatentscale.174
175
Estimationofbehavioraldimorphism176
Wetestedfortheexistenceofsexualdimorphisminbehavioralexpressionbycomparing177
linearcoefficientforthesexfixedeffectincludedinourmultivariateanimalmodelandbase178
ourstatisticalinferenceontheirBayesianprobability(Pmcmc).Thismetricvariesbetween179
0.5and1andindicatestheprobabilityofasignificantdifferencebasedonthenumberof180
posteriorestimatesoverlappingwith0.Pmcmcvalues>0.95werejudgedassignificant.181
182
Comparisonofsex-specificcovariances(GmandGfmatrices)183
Weusedatwo-stepapproachtocomparetheintensityofthedifferenceingenetic184
covariancesamongsexes.First,wecalculatedthedifferenceincovariancebetweenmales185
andfemales(ΔCOVA)foreachpairofbehaviorsandtheirassociatedBayesianprobabilities186
(Pmcmc).Next,wetestedwhethersex-specificcovariancesweresimilarlyoriented.Todo187
so,wecalculatedthevectorcorrelation(r°)betweenaxescontainingthehighestamountof188
geneticvariationusingeigenvaluedecomposition.Wethenestimatedwhetherthe189
resultingvectorcorrelationsamongeigenvectorsofGdifferedsubstantiallyfrom190
expectationsof0(noalignmentofgeneticvariationamongsexes)and1(perfect191

11
alignment).Becausevectorcorrelationsareboundedby0and1,weestimatedtheRegion192
ofPosteriorEquivalence(ROPE)whichwedefinedastheintervals[0.0;0.1]and[0.9;1.0].193
EstimatesfallingwithintheseROPEregionsarejudged“practicallyequivalent”tovector194
correlationsof0and1respectively.Weconvertedtheproportionofestimatesfalling195
outsidetheseROPEregionsintoPmcmcvaluestoinfersignificance,withPmcmc>0.95196
indicatingsignificantdeparturefromthesenullhypotheses.197
198
Estimatingthegeneticconstraintimposedbythecross-sexcovariancematrix(B)199
Wefirsttestedwhethercross-sexcovarianceswithintraitsdifferedsignificantlyfrom0and200
1.Cross-sexcovariancesarerepresentedonthediagonalelementsoftheBmatrix.A201
covarianceof0indicatecompletegeneticindependenceamongsexes,whichwe202
determinedusingBayesianprobabilities(Pmcmc).Totestforadeparturefromcomplete203
geneticcouplingamongsexes,weconvertedthecovariancestocorrelationcoefficients204
(rmf)andusedtheROPEtestdescribedabove.Wetheninvestigatedwhethercross-sex205
cross-traitscovariances–representedontheoff-diagonalelementsofB–differed206
significantlyfromoneanotherbycalculatingtheposteriordifferenceincovarianceas207
described(ΔCOVA).208
Next,wecomparedtheevolutionarytrajectoriesofsexeswithascenarioweresexes209
evolvedindependentlywithoneweresexeswerefullyconstrained(Coxetal.2017).Todo210
so,wesimulated500selectiongradientsbasedonamultivariatenormaldistributionthat211
wescaledtounitlength.Wegeneratedconcordantselectiongradientsbyassigningthe212
samevaluestomaleandfemalegradients(βf=βm)anddiscordantselectionbysettingβf=-213
βm.Weappliedeachselectiongradienttoall1,000posteriorcovariancematricesofGmf214
estimatedfromourmultivariateanimalmodel,thusensuringwetooktheuncertaintyin215
estimatesforward.Wethencalculatedtheresultingresponsetoselection(𝛥𝑧̅)byapplying216
themultivariatebreeder’sequationforcross-sexcovariance(Lande1980):217
$𝛥𝑧!̅𝛥𝑧"̅
% = #$$𝐆𝐟 𝐁′𝐁 𝐆𝐦
% $𝛽!𝛽"% (equation1)218

12
Wecontrastedtheseresponsestoselectiontocaseswherewesetallcross-sexcovariances219
0andbyfullyconstrainingcross-sexcorrelationsto1.Toachievethislaststep,we220
convertedGmftoacorrelationmatrixandreplacedBelementsby1.Wethenback-221
convertedthismodifiedmatrixintocovariancesbyreplacingthediagonalelementsofBby222
thegeometricmeanofmaleandfemalegeneticvariances(i.e.thediagonalelementsofGf223
andGmrespectively).Wethenestimatedthevectorcorrelationbetweentheresponse224
calculatedfromourestimatedmatrixandthoseestimatedwithunconstrained(rB=0)and225
constrainedmatrices(rB=1).Next,wecomparedhowconsistentmaleandfemaleresponse226
waswhenevolvingtowardthesameoptimum(concordantselection)andwhenselection227
wasdiscordant.Thiswasachievedbycalculatingthevectorcorrelationbetweenthe228
directionofselectionforeachsexandthecorrespondingresponse(rβ×Δz).229
RESULTS230
Malesandfemalesshowedlittleevidenceforsexualdimorphisminaveragebehavior(all231
Pmcmc<0.57,TableS1,S2).WedidfindevidenceofaG×Sexinteraction,andfemaleshad232
lowerheritabilityandevolvabilitycomparedtomales(meanfemaleh2=0.10;meanmale233
h2=0.47;meanfemaleI=3.04%;meanmaleI=15.73%).Thisisconfirmedbythefact234
thattheadditivegeneticvariancewaslowerinfemalescomparedtomalesforallthree235
behaviors(posteriormedian[89%CI];shelteremergence:ΔVA(females–males)=-50.12[-236
110.18;8.18],Pmcmc=0.92;activity:ΔVA=-43.38[-89.01;-9.19],Pmcmc=0.99;237
antipredatorresponse:ΔVA=-17.63[-44.47;5.45],Pmcmc=0.92)(Figure2,3;TableS3).238
Wealsofoundstrongevidenceforthesex-specificexpressionofbehavioral239
syndromes,withweakergeneticcorrelationsbetweenbehaviorsinfemales.Malesand240
femalesdifferedprimarilyinhowshelteremergencerelatedtoopen-fieldactivityand241
antipredatorresponse(Figure2),whiletherelationshipbetweenopen-fieldactivityand242
antipredatorresponsewasstablebetweensexes(females:rmf=0.65[0.20;0.98],Pmcmc=243
0.94,Pmcmc=0.91;males:rmf=0.64[0.27;0.92],Pmcmc=0.98).Femalesthatstayed244
longerintheshelterhadlowerantipredatorresponseandtravelledfurtherintheopen-245
field(shelteremergence×open-field:rmf=-0.45[-0.92;0.11],Pmcmc=0.86;shelter246
emergence×antipred.:rmf=-0.48[-0.90;-0.01],Pmcmc=0.91).Incontrast,maleswith247

13
slowshelteremergencehadhigheractivityandantipredatorresponse(shelteremergence248
×open-field:rmf=0.47[0.06;0.87],Pmcmc=0.93;shelteremergence×antipred.:rmf=249
0.50[-0.01;0.84],Pmcmc=0.92).Inaddition,thecorrelationbetweenmajoraxesof250
geneticvariation(gmax)wassignificantly<1,providinganotherlineofevidenceforthe251
presenceofasex-specificsyndrome(vectorcorrelationr°=0.36[0.00;0.76],Pmcmc≠1=252
0.97)(TableS2,S3).253
254
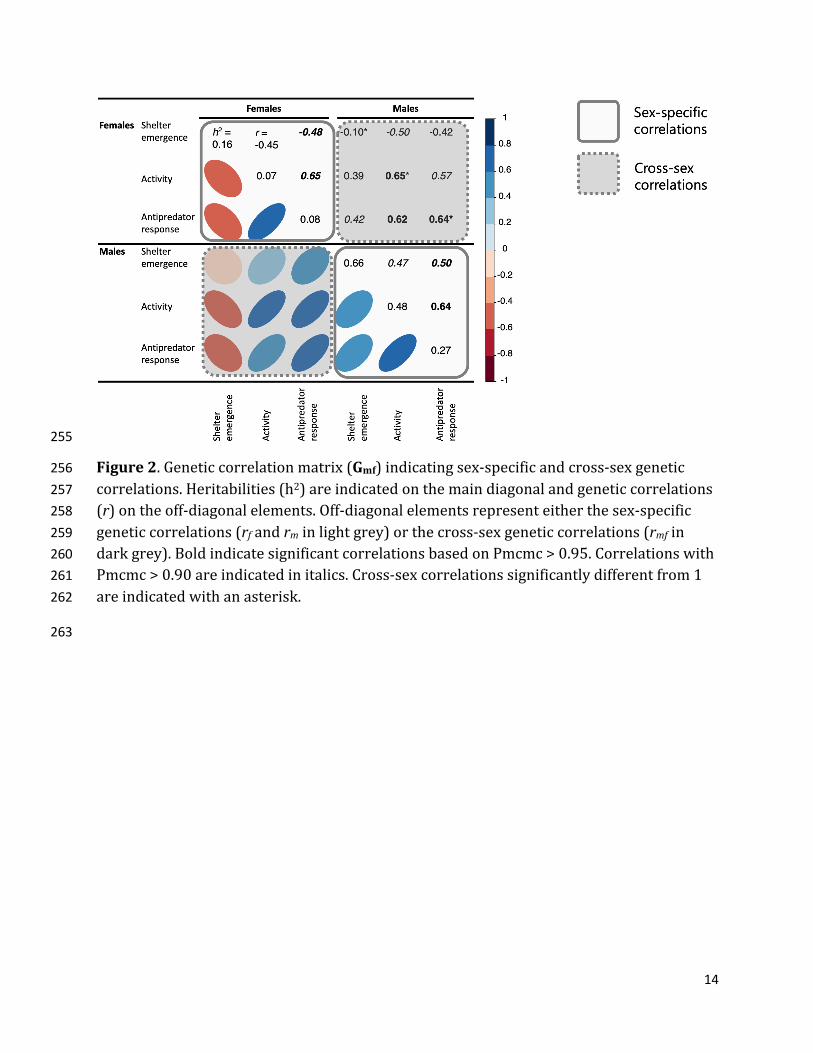
14
255
Figure2.Geneticcorrelationmatrix(Gmf)indicatingsex-specificandcross-sexgenetic256 correlations.Heritabilities(h2)areindicatedonthemaindiagonalandgeneticcorrelations257 (r)ontheoff-diagonalelements.Off-diagonalelementsrepresenteitherthesex-specific258 geneticcorrelations(rfandrminlightgrey)orthecross-sexgeneticcorrelations(rmfin259 darkgrey).BoldindicatesignificantcorrelationsbasedonPmcmc>0.95.Correlationswith260 Pmcmc>0.90areindicatedinitalics.Cross-sexcorrelationssignificantlydifferentfrom1261 areindicatedwithanasterisk.262
263

15
Allbehaviorsshowedweaktomoderatecross-sexcorrelationsandwere264
significantlybelow1(shelteremergence:rmf=-0.09[-0.59;0.39],Pmcmc≠1=1.00;activity:265
rmf=0.65[0.22,0.97],Pmcmc≠1=0.94;antipredatorresponse:rmf=0.64[0.26;0.94],266
Pmcmc≠1=0.96)(TableS3).Inaddition,sexeshadbiasedexpressionofdifferenttrait267
combinations,asindicatedbythehighproportionofasymmetryinB—thecross-sex268
covariancematrix(proportionofskew-symmetry=0.25[0.04,0.42]).Thisdegreeof269
asymmetrywasmostpronouncedforthecross-sexcorrelationsbetweenshelter270
emergenceandactivity(ΔCOVA=-21.39[-43.72;-0.56],Pmcmc=0.97)andbetween271
shelteremergenceandantipredatorresponse(ΔCOVA=-14.8[-32.33;1.46],Pmcmc=272
0.95).Incontrast,thecorrelationbetweenactivityandantipredatorresponsedidnotdiffer273
amongsexes(ΔCOVA=-3.02[-11.73;6.73],Pmcmc=0.71)(Figure3).274
Thismeansthathighlyactivefathersproduceddaughterswithfastershelter275
emergence(rmf=-0.50[-0.85;-0.06],Pmcmc=0.94)andhigherantipredatorresponse(rmf276
=0.57[0.12;0.95],Pmcmc=0.93).Incontrast,activemothersonlyweaklycontributedto277
theirsons’shelteremergence(rmf=0.39[-0.14;0.85],Pmcmc=0.85)andantipredator278
response(rmf=0.42[-0.01;0.85],Pmcmc=0.90).279
280
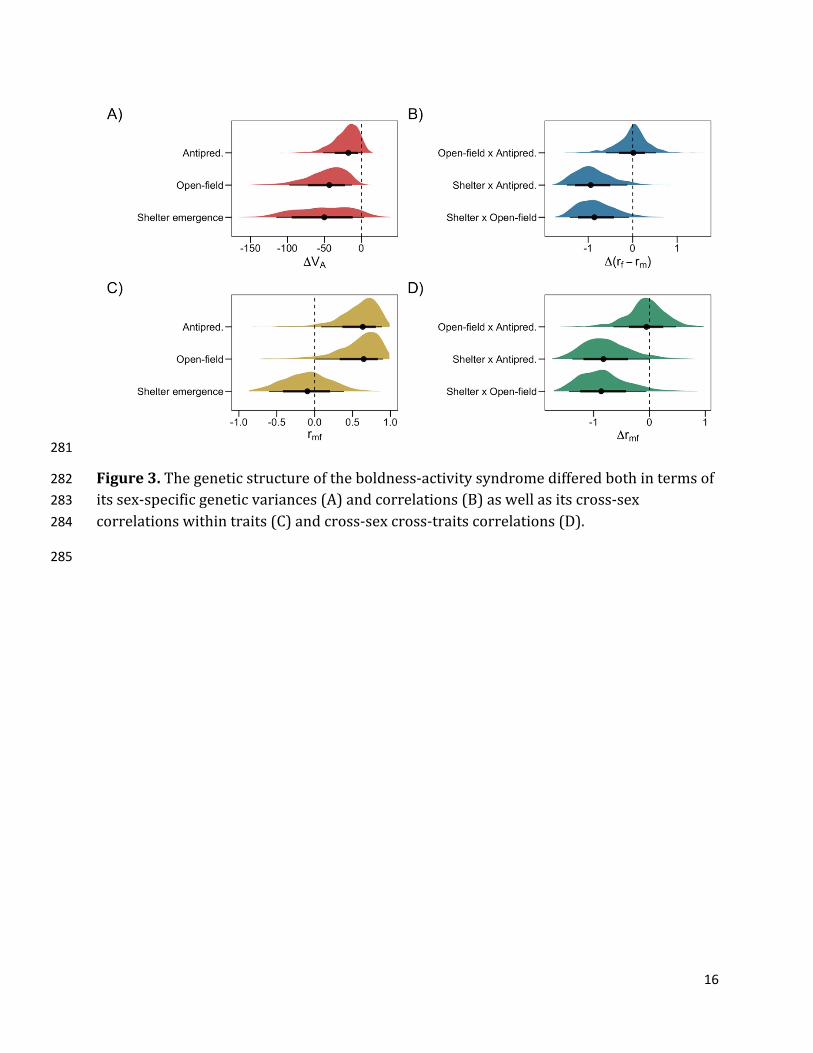
16
281
Figure3.Thegeneticstructureoftheboldness-activitysyndromedifferedbothintermsof282 itssex-specificgeneticvariances(A)andcorrelations(B)aswellasitscross-sex283 correlationswithintraits(C)andcross-sexcross-traitscorrelations(D).284
285

17
Finally,bysimulatingresponsestoselection,wefoundthatevolutionarytrajectories286
weremoreconsistentwithbehaviorsbeingsexuallyindependentthanwithaconstrained287
expressionofbehaviors.Thiswasthecaseregardlessofwhetherselectionfavoredsex-288
specificoptima(i.e.discordantselection:rB=0= 0.95 [0.73;1.00];rB=1=0.28[0.00;0.56];Δr289
=0.64[0.24;0.98],Pmcmc=0.96)orwhenbothsexeshadthesameoptimum(i.e.290
concordantselection:rB=0=0.95[0.74;1.00],Δr=0.23[-0.04;0.66],Pmcmc=0.95)(Figure291
4).Wenextcomparedtheagreementbetweensimulatedselectiongradientsandpredicted292
responsetoselection(rβ×Δz)betweensexes.Malesandfemaleshadequallyconsistent293
responsestoconcordantselection(femalerβ×Δz=0.72[0.29;1.00];malerβ×Δz=0.75[0.47;294
1.00];Δrβ×Δz=-0.19[-0.67;0.07],Pmcmc=0.88).However,femaleresponsetendedtobe295
biasedawayfromselectioncomparedtomaleswhenselectionwasdiscordant(femalerβ×Δz296
=0.50[0.06;0.95];malerβ×Δz=0.77[0.46;1.00],Δrβ×Δz=-0.27[-0.75;0.11],Pmcmc=0.86).297
Thiscouldmeanthatthemagnitudeofthegeneticconstraintisexpressedmorestronglyin298
males,whichisconsistentwiththeasymmetricnatureofthecross-sexcovariancematrix.299
However,thislastresultshouldbeinterpretedwithcautiongiventhewidecredible300
intervalsaroundtheseestimates. 301
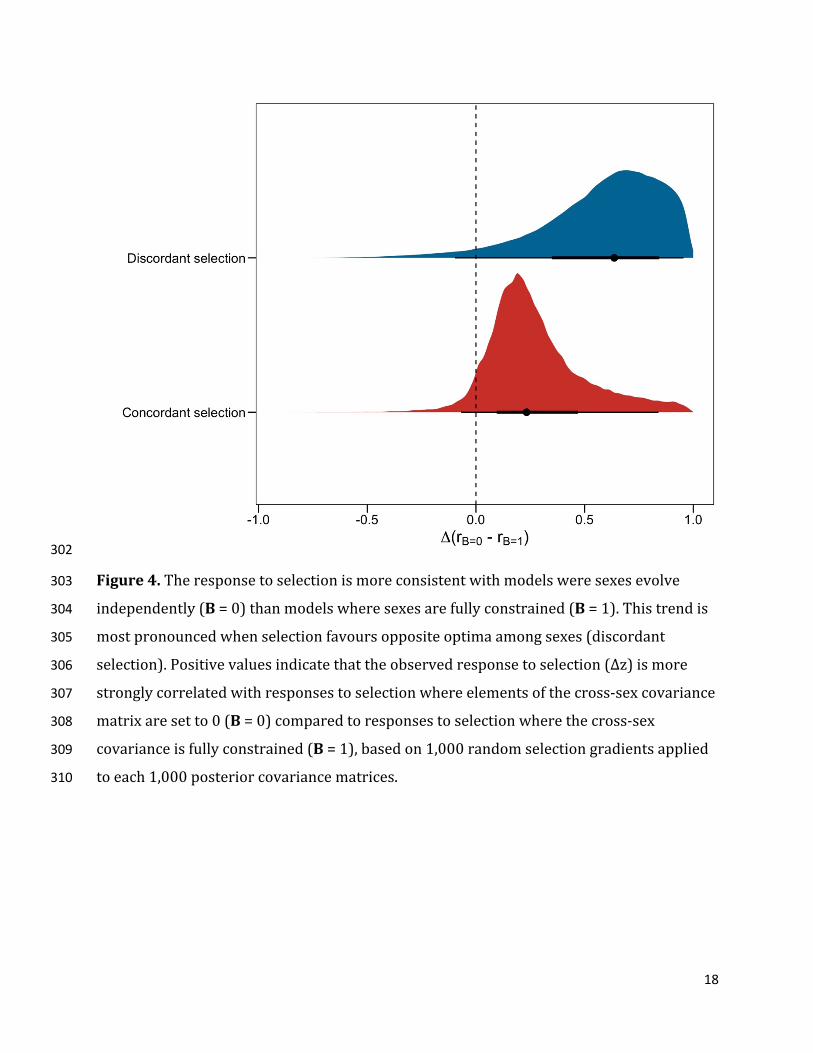
18
302
Figure4.Theresponsetoselectionismoreconsistentwithmodelsweresexesevolve303
independently(B=0)thanmodelswheresexesarefullyconstrained(B=1).Thistrendis304
mostpronouncedwhenselectionfavoursoppositeoptimaamongsexes(discordant305
selection).Positivevaluesindicatethattheobservedresponsetoselection(Δz)ismore306
stronglycorrelatedwithresponsestoselectionwhereelementsofthecross-sexcovariance307
matrixaresetto0(B=0)comparedtoresponsestoselectionwherethecross-sex308
covarianceisfullyconstrained(B=1),basedon1,000randomselectiongradientsapplied309
toeach1,000posteriorcovariancematrices. 310

19
DISCUSSION311
Ourresultsshowthatmalesandfemalesdiffersubstantiallyintheirbehavioralsyndromes312
atthegeneticlevel.Shelteremergencewasgeneticallyindependentbetweenmalesand313
females,whereasthegeneticconstraintforactivityandantipredatorresponsewas314
strongerbutstilldepartedfrom1.Theabsenceofastronggeneticconstraintlinkingthe315
sexesissurprisinggiventhatthebehaviorswemeasuredarenottypicallyconsidered316
distinctsex-specifictraits.Inthecaseofshelteremergence,across-sexcorrelation317
approachingzeroimpliesthatthesamebehaviorisunderpinnedbycompletely318
independentsetsofgenesinmalesandfemales.Suchgeneticuncouplingmeansthatthis319
behaviorcanfulfilldifferentfunctionsineachsexandcanevolveindependently.Our320
resultssuggestthatsexualconflictmayhavebeenresolvedinthisspecieseveninabsence321
ofobservablebehavioraldimorphism.322
Byapplyingaquantitativegeneticapproach,wewereabletouncovermultipleways323
inwhichsexesdifferedintheirbehaviors.Whilemalesandfemalesdidnotdifferinmean324
behaviors,weuncoveredasignatureofbehavioraldimorphismintheamountofgenetic325
variationexpressedbyeachsex.ThisG×Sexeffectwascharacterizedbyfemalebehavior326
beinglessheritablethanobservedformales.Inourcase,traitswithlowerheritabilities327
alsohadlowerevolvabilities,indicatingthatfemalesarelessresponsivetoselective328
pressuresthanmalesinthisspecies.Thisstrongdifferenceingeneticvariancebetween329
sexescouldresultfromstrongerstabilizingselectioninfemaleserodinggeneticvariation330
intraitsrelatedtoexplorationandrisktakingwhilemaintainingasimilaroptimumineach331
sex.Multivariatestabilizingselectionhasindeedbeenshowntobeanimportantdriverof332
traitevolution,forexampleingenitalmorphologyinarthropods(Arnqvist1997;Houseet333
al.2020).Anotherpossibilityisthatrearingindividualsinlaboratoryconditionswithad-lib334
accesstofoodandsheltercouldalleviatetheresourceacquisitionandallocationtrade-offs335
thatmaybenegotiateddifferentlybyeachsex.Thisexplanationisespeciallyplausibleif336
malesandfemalesmodifytheirphenotypestodifferentdegreeinresponsetocaptivity.337
Notetoothattheenvironmentalcontributiontofemales’behavioralphenotypewasmuch338
greaterthanthatofmales,leavingspacefordifferentialadjustmentofbehaviorbysexesto339
occur.340

20
Malesandfemalesdifferedsubstantiallyinhowtheyexpressedaboldness-activity341
syndrome.Contrarytoourpredictions,inbothsexesactivegenotypeswerealsomore342
sensitivetopredatorycues,withnochangetothemagnitudeofthegeneticcorrelation.The343
primarydifferencewasthereforeinhowshelteremergencerelatedtoactivityand344
antipredatorresponse.“Bold”females–i.e.femaleswithfastemergencefromtheshelter–345
hadhigheractivityandantipredatorresponse.Incontrast,activemalesweremore346
sensitivetopredatorcuesandbehavedcautiouslywhenemergingfromtheshelter.This347
resultprovidesimportantinsightintohowmaleandfemalecricketshandlerisky348
situations.Femalemusttravelthroughriskyenvironmentsinordertolocatemates.349
Therefore,boldandactivegenotypesmayneedtocompensatefortheseriskybehaviorsby350
beingmorereactivetothepresenceofpredatorcues.Males,incontrast,stayclosetotheir351
shelterbutproducecourtshipsignalsthatmakethemthetargetofpredatorsandparasites.352
Asaresult,boldermalesmayignorethepresenceofpredatorycuesifthisstrategyyields353
higherfrequencyofencounterswithfemales.Thistypeofriskcompensationstrategyhas354
alreadybeenshowninpreviousstudieswheremaleswithmoreattractivesongsstay355
longerinshelters(Hedrick2000).356
Ourfindingsthatmalesandfemalesdifferedinthegeneticexpressionbehavioral357
syndromesandthatcross-sexcorrelationsareuncoupledsuggestthatrisk-takingis358
regulatedbydifferentphysiologicalpathwaysineachsex.Incrickets,severalmonoamine359
neurohormonesareinvolvedintheregulationofmultiplebehaviorstiedtoa“fightor360
flight”response,includingaggression,courtship,dispersalandresponsetosimulated361
predationexposure(Adamoetal.1995,2013;StevensonandRillich2016;Adamo2017;362
Lundgrenetal.2021).However,experimentstendtoeitherfocusonmalesexclusivelyor363
arenotdesignedtoaddresssex-differencesinphysiologicalmechanisms.However,sex-364
differencesinasimilaraxisofvariation,theproactive/reactiveaxis,havealsobeen365
describedinseveralvertebratetaxa(Kokrasetal.2012;Immonenetal.2018).366
Thegeneticstructureofbehavioralsyndromesishighlyconservedinthisspecies,367
evenacrossisolatedpopulations(Royautéetal.2020).Thisindicatesthatbehavioral368
syndromesmaynotvarymuchwhenexposedtodifferentselectivepressures.Instead,369
geneticconstraintsresultingfrompleiotropywasthemorelikelyexplanationsforthe370

21
presenceofbehavioralsyndromeinthisspecies.Here,weshowthatbehavioralsyndromes371
differedmorestronglybetweensexesthanamongpopulations.Alikelyexplanationisthat372
malesandfemalesexpresssex-specificbehaviorsregardlessofthepopulationoforigin.As373
aresult,selectionmaybemorelikelytodifferbetweensexesthanbetweenpopulations.374
Whilecomparisonsofdifferencesinselectionamongsexesandpopulationsarescarce,375
previousstudiessuggestthatsexuallydiscordantselectioniscommoninthewildand376
tendstobestrongerintraitsinwhichsexualdimorphismispronounced(Coxetal.2009).377
Whilerarelyinvestigated,sexspecificdifferencesinbehavioralsyndromeshave378
beendemonstratedinsomespecies.However,mostofthesestudieshavefocusedon379
comparisonsofbehavioralrepeatabilitiesbetweenmalesandfemalesinsingletraits380
(Jenkins2011;Debeffeetal.2015)ratherprovidingabroaderexplorationofmultivariate381
patternsofcross-sexcorrelationinbehavioraltraits,aswedidhere.Interestingly,(Hedrick382
andKortet2012)previouslyidentifiedsexdifferencesintherepeatabilityofshelter383
emergenceinaseparatepopulationofG.integer.Contrarytowhatwedetected,maleshad384
lowerrepeatabilitycomparedtofemales.Notethatrepeatabilityandheritabilityarenot385
directlycomparablebecauserepeatabilityalsoincludessourcesofvariationduetothe386
“permanentenvironment”whileheritabilityonlyincludesadditivegeneticvariation.More387
recentlyseveralstudieshavealsocomparedsyndromestructureamongsexeswithmixed388
results.Somestudiesreportedlargedifferencesinbehavioralsyndromesbetweenmales389
andfemales(Fresneauetal.2014;Hanetal.2015;Royauté2015;Wayetal.2015)while390
otherssupportaconservedsyndromestructurebetweensexes(Michelangelietal.2016;391
Gouletetal.2021).Ourresultssupplyanadditionallineofevidenceinfavorofsex-specific392
syndromes.393
Thereisonlylimitedexplorationofmultivariatepatternsofcross-sexcorrelationin394
behavioraltraits.Moststudiesreportedfrom(Poissantetal.2010)concernsingletraits395
andpointtostrongcross-sexcorrelationswithrelativelyweakdimorphismforbehavior.396
Themajorityoftheseestimateswere,however,fromstudiesinterestedinsexualselection,397
with<4%ofestimatescomingfromexplicitlybehavioralstudies.Becausewealso398
estimatedthecross-sexcorrelationattheadditivegeneticlevel,wewereabletodetermine399
thatbehavioraltraitsmaybelessconstrainedbysexthanpreviouslythought.Indeed,our400

22
meanestimateforrmfwasmuchweakerthanobservedinpreviousstudiesevaluatingintra-401
locussexualconflictsinbehavior(meanrmf=0.46vs.0.77,Poissantetal.2010).Our402
resultssuggestthatbehavioraltraitsmaycommonlyfulfilldifferentfunctionsbetween403
sexesandshouldthereforebeanalyzedasseparatetraitsbetweenthesexesinbehavioral404
syndromeandanimalpersonalitystudies.405
Veryfewstudieshavebeenabletoestimatethegeneticcontributionofsexesto406
behavioralsyndromesaswehavedonehere.Intheorb-weavingspiderNuctenea407
umbratical,cross-sexcorrelationsforactivityandaggressionwereinthesamerangeasour408
estimatesforactivityandantipredatorresponse(rmf=0.50)(Kralj-Fišeretal.2019).In409
Gryllusbimaculatus,whichiscloselyrelatedtoG.integer,explorationandaggressionare410
morestronglycorrelatedinfemalesandthesetraitshaveasymmetriccontributionsacross411
sexes(Hanetal.2019).However,cross-sexcorrelationswithintraitswerequalitatively412
strongerthanobservedinourstudy(meanrmf=0.70vsmeanrmf=0.46),pointingtoa413
strongergeneticconstraintbetweensexesthanweobservedinGryllusinteger.Finally,414
Whiteetal.(2019)showeddifferencesbetweensex-specificcovariancesinguppiesto415
similarG.integer.However,theyreportedamuchweakersignalofasymmetryinB.The416
factthatthesefewstudiesdifferwidelyinconclusionsshowsthatthereisnoconsensusyet417
onhowbehavioralsyndromesdifferbetweensexesandevencloselyrelatedspeciesmay418
showstrongdifferencesintheexpressionofbehavioraldimorphism.419
Otherstudies,conductedprimarilyonmorphologicaltraits,pointtostrong420
constraintsimposedbygeneticcross-sexcovariances(Gosdenetal.2012;Gosdenand421
Chenoweth2014;SztepanaczandHoule2019).Inourcase,theconstrainingeffectofcross-422
sexcovarianceswasweakatbestandresponsestoselectionweremoreconsistentwith423
independentevolutionarytrajectoriesforeachsex.Thisissimilartoapreviousstudyon424
sexuallydimorphicornamentsinbrownanoleswhichshowedthatthecross-sex425
covarianceamongornamentsdidnotsubstantiallyconstraintheevolutionofdimorphism426
(Coxetal.2017).427
Wefoundahighdegreeofasymmetryinthecross-sexcross-traitscomponentsof428
thegeneticcovariancematrix.Inparticular,maleactivityhadastrongercontributionto429

23
femaleshelteremergencethanfemaleactivityonmaleshelteremergence.Thistypeof430
asymmetriccontributiontothephenotypeisexpectedtobiasresponsesawayfrom431
selection.InDrosophilaforexample,wingshapeevolutionisprimarilyshapedbycross-sex432
covariancesforthesametrait(i.e.thediagonalelementsofB)ratherthancross-sexcross-433
traitcovariances(SztepanaczandHoule2019).Thiswasthecaseeventhoughupperand434
lowerelementsoftheBmatrixfrequentlyhadoppositesignsanddespiteageneral,though435
weaker,signalofasymmetrysuchaswereporthere(5%vs.25%).Despitestrong436
asymmetryinourestimateofcross-sexcovariances,wefoundonlyweakevidencefor437
biasedresponsestoselection.Instead,oursimulationsoftheresponsestoselections438
showedthattheevolutionofbehavioraldimorphismswasmoreconsistentwithscenarios439
inwhichsexesaregeneticallyuncoupledandallowedtoevolveindependentlycomparedto440
afullyconstrainedcross-sexgeneticarchitecture.Thismeansthattheindependent441
evolutionarytrajectoriesbysexeswereportarelikelyaresultoftheweakcross-sex442
correlationswithintraitswedetected(meanrmf<0.50).443
Weshowthattraitsinvolvedinstudiesofanimalpersonalityhaveasex-specific444
geneticarchitecture.Thesebehaviorsaregeneticallyuncoupledbetweensexesandallow445
sexestofollowindependentevolutionarytrajectories.Ourresultsareintriguingbecause446
theorypredictsthatcross-sexcovarianceswillactasconstraintsandlimittheabilitiesof447
sexestoreachtheiroptimum.Thetypeofgeneticindependencewefoundsuggestsapast448
historyofdiscordantselectionthathasstronglyshapedbehavioraldimorphismandhas449
resolvedsexualconflictinthisspecies.450
451

24
LITTERATURECITED452
Adamo,S.A.2017.Thestressresponseandimmunesystemshare,borrow,andreconfigure453
theirphysiologicalnetworkelements:Evidencefromtheinsects.Horm.Behav.454
88:25–30.455
Adamo,S.A.,I.Kovalko,andB.Mosher.2013.Thebehaviouraleffectsofpredator-induced456
stressresponsesinthecricket(Gryllustexensis):theupsideofthestressresponse.457
J.Exp.Biol.216:4608–4614.TheCompanyofBiologistsLtd.458
Adamo,S.A.,C.E.Linn,andR.R.Hoy.1995.Theroleofneurohormonaloctopamineduring459
“fightorflight”behaviourinthefieldcricketGryllusbimaculatus.J.Exp.Biol.460
198:1691–1700.TheCompanyofBiologistsLtd.461
Aragón,P.2011.Theresponsetothesocialenvironmentrevealssex-dependent462
behaviouralsyndromesintheBosca’snewt(Lissotritonboscai).J.Ethol.29:79–83.463
Arnqvist,G.1997.Theevolutionofanimalgenitalia:distinguishingbetweenhypothesesby464
singlespeciesstudies.Biol.J.Linn.Soc.60:365–379.465
Berger,D.,E.C.Berg,W.Widegren,G.Arnqvist,andA.A.Maklakov.2014.Multivariate466
intralocussexualconflictinseedbeetles.Evolution68:3457–3469.467
Blanckenhorn,W.U.2005.BehavioralCausesandConsequencesofSexualSize468
Dimorphism.Ethology111:977–1016.469
Bonduriansky,R.,andS.F.Chenoweth.2009.Intralocussexualconflict.TrendsEcol.Evol.470
24:280–288.471
Cox,R.M.,R.A.Costello,B.E.Camber,andJ.W.McGlothlin.2017.Multivariategenetic472
architectureoftheAnolisdewlaprevealsbothsharedandsex-specificfeaturesofa473
sexuallydimorphicornament.J.Evol.Biol.30:1262–1275.474

25
Debeffe,L.,J.F.Lemaître,U.A.Bergvall,A.J.M.Hewison,J.M.Gaillard,N.Morellet,M.475
Goulard,C.Monestier,M.David,H.Verheyden-Tixier,L.Jäderberg,C.Vanpé,andP.476
Kjellander.2015.Short-andlong-termrepeatabilityofdocilityintheroedeer:sex477
andagematter.Anim.Behav.109:53–63.478
Delph,L.F.,J.C.Steven,I.A.Anderson,C.R.Herlihy,andE.D.B.Iii.2011.Eliminationofa479
GeneticCorrelationBetweentheSexesViaArtificialCorrelationalSelection.480
Evolution65:2872–2880.481
Dochtermann,N.A.,andN.J.Dingemanse.2013.Behavioralsyndromesasevolutionary482
constraints.Behav.Ecol.24:806–811.OxfordAcademic.483
French,B.W.,andW.H.Cade.1987.Thetimingofcalling,movement,andmatinginthe484
fieldcricketsGryllusveletis,G.pennsylvanicus,andG.integer.Behav.Ecol.485
Sociobiol.21:157–162.486
Fresneau,N.,E.Kluen,andJ.E.Brommer.2014.Asex-specificbehavioralsyndromeina487
wildpasserine.Behav.Ecol.25:359–367.488
Gosden,T.P.,andS.F.Chenoweth.2014.TheEvolutionaryStabilityofCross-Sex,Cross-489
TraitGeneticCovariances.Evolution68:1687–1697.490
Gosden,T.P.,K.-L.Shastri,P.Innocenti,andS.F.Chenoweth.2012.TheB-MatrixHarbors491
SignificantandSex-SpecificConstraintsontheEvolutionofMulticharacterSexual492
Dimorphism.Evolution66:2106–2116.493
Goulet,C.T.,W.Hart,B.L.Phillips,J.Llewelyn,B.B.M.Wong,andD.G.Chapple.2021.No494
behavioralsyndromesorsex-specificpersonalitydifferencesinthesouthern495
rainforestsunskink(Lampropholissimilis).Ethology127:102–108.496

26
Hadfield,J.D.2010.MCMCmethodsformulti-responsegeneralizedlinearmixedmodels:497
theMCMCglmmRpackage.J.Stat.Softw.33:1–22.498
Hämäläinen,A.,E.Immonen,M.Tarka,andW.Schuett.2018.Evolutionofsex-specificpace-499
of-lifesyndromes:causesandconsequences.Behav.Ecol.Sociobiol.72:50.500
Han,C.S.,T.P.Gosden,andN.J.Dingemanse.2019.Proteindeprivationfacilitatesthe501
independentevolutionofbehaviorandmorphology.Evolution73:1809–1820.502
Han,C.S.,P.G.Jablonski,andR.C.Brooks.2015.Intimidatingcourtshipandsexdifferences503
inpredationriskleadtosex-specificbehaviouralsyndromes.Anim.Behav.109:177–504
185.505
Hedrick,A.V.,andR.Kortet.2012.Sexdifferencesintherepeatabilityofboldnessover506
metamorphosis.Behav.Ecol.Sociobiol.66:407–412.507
Hedrick,A.V.,andE.J.Temeles.1989.Theevolutionofsexualdimorphisminanimals:508
Hypothesesandtests.TrendsEcol.Evol.4:136–138.509
House,C.,P.Tunstall,J.Rapkin,M.J.Bale,M.Gage,E.delCastillo,andJ.Hunt.2020.510
Multivariatestabilizingsexualselectionandtheevolutionofmaleandfemalegenital511
morphologyintheredflourbeetle*.Evolution74:883–896.512
Immonen,E.,A.Hämäläinen,W.Schuett,andM.Tarka.2018.Evolutionofsex-specificpace-513
of-lifesyndromes:geneticarchitectureandphysiologicalmechanisms.Behav.Ecol.514
Sociobiol.72:60.515
Jenkins,S.H.2011.Sexdifferencesinrepeatabilityoffood-hoardingbehaviourofkangaroo516
rats.Anim.Behav.81:1155–1162.517

27
Kokras,N.,C.Dalla,A.C.Sideris,A.Dendi,H.G.Mikail,K.Antoniou,andZ.Papadopoulou-518
Daifoti.2012.Behavioralsexualdimorphisminmodelsofanxietyanddepression519
duetochangesinHPAaxisactivity.Neuropharmacology62:436–445.520
Kortet,R.,andA.Hedrick.2007.AbehaviouralsyndromeinthefieldcricketGryllusinteger:521
intrasexualaggressioniscorrelatedwithactivityinanovelenvironment.Biol.J.522
Linn.Soc.91:475–482.523
Kralj-Fišer,S.,K.L.Laskowski,andF.Garcia-Gonzalez.2019.Sexdifferencesinthegenetic524
architectureofaggressivenessinasexuallydimorphicspider.Ecol.Evol.9:10758–525
10766.526
Kruuk,L.E.B.2004.Estimatinggeneticparametersinnaturalpopulationsusingthe‘animal527
model.’Philos.Trans.R.Soc.Lond.B.Biol.Sci.359:873–890.528
Lande,R.1980.SexualDimorphism,SexualSelection,andAdaptationinPolygenic529
Characters.Evolution34:292–305.[SocietyfortheStudyofEvolution,Wiley].530
Long,T.A.F.,andW.R.Rice.2007.Adultlocomotoryactivitymediatesintralocussexual531
conflictinalaboratory-adaptedpopulationofDrosophilamelanogaster.Proc.R.Soc.532
BBiol.Sci.274:3105–3112.RoyalSociety.533
Lundgren,K.A.,R.N.Abbey-Lee,L.C.Garnham,A.Kreshchenko,S.Ryding,andH.Løvlie.534
2021.Manipulatingmonoaminesreducesexplorationandboldnessof535
Mediterraneanfieldcrickets.Behav.Processes183:104298.536
Mainwaring,M.C.,J.L.Beal,andI.R.Hartley.2011.Zebrafinchesarebolderinanasocial,537
ratherthansocial,context.Behav.Processes87:171–175.538

28
Michelangeli,M.,D.G.Chapple,andB.B.M.Wong.2016.Arebehaviouralsyndromessex539
specific?Personalityinawidespreadlizardspecies.Behav.Ecol.Sociobiol.70:1911–540
1919.541
Moiron,M.,K.L.Laskowski,andP.T.Niemelä.2020.Individualdifferencesinbehaviour542
explainvariationinsurvival:ameta-analysis.Ecol.Lett.23:399–408.543
Poissant,J.,A.J.Wilson,andD.W.Coltman.2010.Sex-SpecificGeneticVarianceandthe544
EvolutionofSexualDimorphism:ASystematicReviewofCross-SexGenetic545
Correlations.Evolution64:97–107.546
Réale,D.,S.M.Reader,D.Sol,P.T.McDougall,andN.J.Dingemanse.2007.Integrating547
animaltemperamentwithinecologyandevolution.Biol.Rev.82:291–318.548
Royauté,C.,R.,.Buddle,C.M.,.Vincent.2015.Undertheinfluence:Sublethalexposuretoan549
insecticideaffectspersonalityexpressioninajumpingspider.29.550
Royauté,R.,andN.A.Dochtermann.2017.Whenthemeannolongermatters:551
Developmentaldietaffectsbehavioralvariationbutnotpopulationaveragesinthe552
housecricket(Achetadomesticus).Behav.Ecol.28:337–345.553
Royauté,R.,C.Garrison,J.Dalos,M.A.Berdal,andN.A.Dochtermann.2019.Currentenergy554
stateinteractswiththedevelopmentalenvironmenttoinfluencebehavioural555
plasticity.Anim.Behav.148:39–51.556
Royauté,R.,K.Greenlee,M.Baldwin,andN.A.Dochtermann.2015.Behaviour,metabolism557
andsize:phenotypicmodularityorintegrationinAchetadomesticus?Anim.Behav.558
110:163–169.559

29
Royauté,R.,A.Hedrick,andN.A.Dochtermann.2020.Behaviouralsyndromesshape560
evolutionarytrajectoriesviaconservedgeneticarchitecture.Proc.R.Soc.BBiol.Sci.561
287:20200183.RoyalSociety.562
Schuett,W.,T.Tregenza,andS.R.X.Dall.2010.Sexualselectionandanimalpersonality.563
Biol.Rev.85:217–246.564
Sih,A.,A.Bell,andJ.C.Johnson.2004.Behavioralsyndromes:anecologicaland565
evolutionaryoverview.TrendsEcol.Evol.19:372–378.566
Stevenson,P.A.,andJ.Rillich.2016.Controllingthedecisiontofightorflee:therolesof567
biogenicaminesandnitricoxideinthecricket†.Curr.Zool.62:265–275.568
Sztepanacz,J.L.,andD.Houle.2019.Cross-sexgeneticcovarianceslimittheevolvabilityof569
wing-shapewithinandamongspeciesofDrosophila.Evolution73:1617–1633.John570
Wiley&Sons,Ltd.571
Walsh,B.,andM.W.Blows.2009.AbundantGeneticVariation+StrongSelection=572
MultivariateGeneticConstraints:AGeometricViewofAdaptation.Annu.Rev.Ecol.573
Evol.Syst.40:41–59.574
Way,G.P.,A.L.Kiesel,N.Ruhl,J.L.Snekser,andS.P.McRobert.2015.Sexdifferencesina575
shoaling-boldnessbehavioralsyndrome,butnolinkwithaggression.Behav.576
Processes113:7–12.577
Wilson,A.J.,D.Réale,M.N.Clements,M.M.Morrissey,E.Postma,C.A.Walling,L.E.B.578
Kruuk,andD.H.Nussey.2010.Anecologist’sguidetotheanimalmodel.J.Anim.579
Ecol.79:13–26.580
581
Related Documents











