Chemoecology7:13-23 (1996) 0937-7409/96/010013-11 $1.50+ 0.20 © 1996 Birkhäuser Verlag, Basel Sex pheromones and attractants in the Eucosmini and Grapholitini (Lepidoptera, Tortricidae) Peter Witzgall 1, Jean-Pierre Chambon 2, Marie Bengtsson 1, C. Rikard Unelius 3, Monica Appelgren 4, Gyorgy Makranczy s, N. Muraleedharan 6, Darwin W. Reed 7, Klaus Hellrigl s, Hans-Ruedi Buser 9, Eric Hallberg ~°, Gunnar Bergström 4, Miklos Tóth 5, Christer Löfstedt u, and Jan Löfqvist 1 ~Department of Plant Protection Sciences,Swedish University of Agricultural Sciences, S-23053 Alnarp, Sweden 2Department of Zoology, National Institute of Agronomical Research, F-78026 Versailles cedex, France 3Department of Organic Chemistry, Royal Institute of Technology,S-10044 Stockholm, Sweden 4Department of Chemical Ecology, Göteborg University, S-41319 Göteborg, Sweden SPlant Protection Institute, Hungarian Academy of Sciences, H-1525 Budapest, Hungary 6Tea Research Institute, The United Planters' Association of Southern India, Valparai 642127, India 7plant BiotechnologyInstitute, National Research Council, Saskatoon S7N 0W9, Canada 8Wolkensteinstr. 83, 1-39042 Brixen, Italy 9Swiss Federal Research Station, CH-8820 Wädenswil, Switzerland ~°Department of Zoology, Lund University, S-22362 Lund, Sweden 11Department of Ecology, Lund University, S-22362 Lund, Sweden Summary. The geometric isomers (E,E)-, (E,Z) -, (Z,E) -, and (Z,Z)-8,10-dodecadien-l-yl acetate were identified as sex pheromone components or sex attractants in the tribes Eucosmini and Grapholitini of the tortricid sub- family Olethreutinae. Species belonging to the more ancestral Tortricinae were not attracted. Each one isomer was behaviourally active in males of Cydia and Grapholita (Grapholitini), either as main pheromone compound, attraction synergist or attraction inhibitor. Their recipro- cal attractive/antagonistic activity in a number of species enables specific communication with these four com- pounds. Pammene, as weil as other Grapholita and Cydia responded to the monoenic 8- or 10-dodecen-l-yl ace- tates. Of the tribes Olethreutini and Eucosmini, Hedya, Epiblema, Eucosma, and Notocelia trimaculana were also attracted to 8,10-dodecadien-l-yl acetates, but several other Notocelia to 10,12-tetradecadien-l-yl acetates. The female sex pheromones of C. fagiglandana, C. pyrivora, C. splendana, Epiblema foenella and Notocelia roborana were identified. (E,E)- and (E,Z)-8,10-dodecadien-l-yl acetate are produced via a common E9 desaturation pathway in C. splendana. Calling C. nigricana and C. fagiglandana females are attracted to wingfanning males. Key words, sex pheromone - synergist - antagonist - mate recognition - reproductive isolation - chemotax- onomoy - phylogeny - evolution - Lepidoptera - Tortricidae Introduction The comparative analysis of sex pheromones con- tributes to our understanding of the phylogeny of tor- tricid moths and the evolutionary role of mating signals (Roelofs & Brown 1982; Horak & Brown 1991; Phelan 1992; Löfstedt 1993; Linn & Roelofs 1994). To date, sex pheromones have been identified in 10Ö tortricid spe- cies, belonging to 49 different genera (Arn et al. 1992, 1995). This gives a good overview of their pheromone chemistry, but comprehensive studies on closely related and sympatric species are yet sporadic (Cardé et al. 1977; Silk & Kuenen 1988; Foster et al. 1991). The sex pheromones of virtually all economically important tor- tricids are known, but less abundant species have rarely been investigated at all. The species treated here belong to the tribes Olethreutini, Eucosmini and Grapholitini of the mono- phyletic subfamily Olethreutinae. The ancestral Ole- threutini, which comprise oligophagous leafrollers, typically use monoenic 14- or 12-carbon pherome com- pounds. Monophagous, internal feeders are character- istic for the more derived tribes Eucosmini and Grapholitini, 12-carbon pheromones occur in most species. Dienic (A, A) -8,10-dodecadien- 1 -yl (A 8, A 10-12) compounds are found in a number of genera, they are typical for Cydia (Arn et al. 1992, 1995), which is regarded as one of the most advanced genera in the tortricid system (Bradley et al. 1973; Danilevskij & Kuznetsov 1968; Razowski 1976; Horak & Brown 1991). The E,E and E,Z isomers of A8,A10-12Ac have been identified in four C£dia species (Davis et al. 1984; Greenway 1984; Katovich et al. 1989; McDonough et al. 1990; Witzgall et al. 1993). The monoenic (A)-8- and (A)-10-dodecen-l-yl acetates (AS-12Ac,A 10-12Ac) are sex attractants and pheromones in other Cydia and in the closely related genera Grapholita and Pammene. Some Eucosmini, such as Eucoama and Epiblema re-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chemoecology 7:13-23 (1996) 0937-7409/96/010013-11 $1.50 + 0.20 © 1996 Birkhäuser Verlag, Basel
Sex pheromones and attractants in the Eucosmini and Grapholitini (Lepidoptera, Tortricidae) Peter Witzgall 1, Jean-Pierre Chambon 2, Marie Bengtsson 1, C. Rikard Unelius 3, Monica Appelgren 4, Gyorgy Makranczy s, N. Muraleedharan 6, Darwin W. Reed 7, Klaus Hellrigl s, Hans-Ruedi Buser 9, Eric Hallberg ~°, Gunnar Bergström 4, Miklos Tóth 5, Christer Löfstedt u, and Jan Löfqvist 1
~Department of Plant Protection Sciences, Swedish University of Agricultural Sciences, S-23053 Alnarp, Sweden 2Department of Zoology, National Institute of Agronomical Research, F-78026 Versailles cedex, France 3Department of Organic Chemistry, Royal Institute of Technology, S-10044 Stockholm, Sweden 4Department of Chemical Ecology, Göteborg University, S-41319 Göteborg, Sweden SPlant Protection Institute, Hungarian Academy of Sciences, H-1525 Budapest, Hungary 6Tea Research Institute, The United Planters' Association of Southern India, Valparai 642127, India 7plant Biotechnology Institute, National Research Council, Saskatoon S7N 0W9, Canada 8Wolkensteinstr. 83, 1-39042 Brixen, Italy 9Swiss Federal Research Station, CH-8820 Wädenswil, Switzerland ~°Department of Zoology, Lund University, S-22362 Lund, Sweden 11Department of Ecology, Lund University, S-22362 Lund, Sweden
Summary. The geometric isomers (E,E)-, (E,Z) -, (Z,E) -, and (Z,Z)-8,10-dodecadien-l-yl acetate were identified as sex pheromone components or sex attractants in the tribes Eucosmini and Grapholitini of the tortricid sub- family Olethreutinae. Species belonging to the more ancestral Tortricinae were not attracted. Each one isomer was behaviourally active in males of Cydia and Grapholita (Grapholitini), either as main pheromone compound, attraction synergist or attraction inhibitor. Their recipro- cal attractive/antagonistic activity in a number of species enables specific communication with these four com- pounds. Pammene, as weil as other Grapholita and Cydia responded to the monoenic 8- or 10-dodecen-l-yl ace- tates. Of the tribes Olethreutini and Eucosmini, Hedya, Epiblema, Eucosma, and Notocelia trimaculana were also attracted to 8,10-dodecadien-l-yl acetates, but several other Notocelia to 10,12-tetradecadien-l-yl acetates. The female sex pheromones of C. fagiglandana, C. pyrivora, C. splendana, Epiblema foenella and Notocelia roborana were identified. (E,E)- and (E,Z)-8,10-dodecadien-l-yl acetate are produced via a common E9 desaturation pathway in C. splendana. Calling C. nigricana and C. fagiglandana females are attracted to wingfanning males.
Key words, sex pheromone - synergist - antagonist - mate recognition - reproductive isolation - chemotax- onomoy - phylogeny - evolution - Lepidoptera - Tortricidae
Introduction
The comparative analysis of sex pheromones con- tributes to our understanding of the phylogeny of tor-
tricid moths and the evolutionary role of mating signals (Roelofs & Brown 1982; Horak & Brown 1991; Phelan 1992; Löfstedt 1993; Linn & Roelofs 1994). To date, sex pheromones have been identified in 10Ö tortricid spe- cies, belonging to 49 different genera (Arn et al. 1992, 1995). This gives a good overview of their pheromone chemistry, but comprehensive studies on closely related and sympatric species are yet sporadic (Cardé et al. 1977; Silk & Kuenen 1988; Foster et al. 1991). The sex pheromones of virtually all economically important tor- tricids are known, but less abundant species have rarely been investigated at all.
The species treated here belong to the tribes Olethreutini, Eucosmini and Grapholitini of the mono- phyletic subfamily Olethreutinae. The ancestral Ole- threutini, which comprise oligophagous leafrollers, typically use monoenic 14- or 12-carbon pherome com- pounds. Monophagous, internal feeders are character- istic for the more derived tribes Eucosmini and Grapholitini, 12-carbon pheromones occur in most species. Dienic (A, A) -8,10-dodecadien- 1 -yl (A 8, A 10-12) compounds are found in a number of genera, they are typical for Cydia (Arn et al. 1992, 1995), which is regarded as one of the most advanced genera in the tortricid system (Bradley et al. 1973; Danilevskij & Kuznetsov 1968; Razowski 1976; Horak & Brown 1991).
The E,E and E,Z isomers of A8,A10-12Ac have been identified in four C£dia species (Davis et al. 1984; Greenway 1984; Katovich et al. 1989; McDonough et al. 1990; Witzgall et al. 1993). The monoenic (A)-8- and (A)-10-dodecen-l-yl acetates (AS-12Ac,A 10-12Ac) are sex attractants and pheromones in other Cydia and in the closely related genera Grapholita and Pammene. Some Eucosmini, such as Eucoama and Epiblema re-

14 P. Witzgall et al. CHEMOECOLOGY
s p o n d to A 8,A 10-12Ac, wh i l e Notoeel ia a re a t t r a c t e d to ( A , A ) - 1 0 , 1 2 - t e t r a d e c a d i e n y l a c e t a t e s (A 10,A 12-14Ac) ( A r n et al. 1992, 1995).
W e h a v e s c r e e n e d m a l e a t t r a c t i o n to t he g e o m e t r i c i s o m e r s o f t he se c o m p o u n d s . T h e r e su l t s o f t he t r a p tes t s a re c o n f i r m e d b y c h e m i c a l i d e n t i f i c a t i o n o f f e m a l e g l a n d c o m p o u n d s a n d b e h a v i o u r a l s tud ie s in five spe-
cies.
Materials and methods
Chemical analys&
Insects Insects for pheromone gland extracts were field-collected as diapaus- ing last instar larvae. C. fagiglandana were obtained from beecb-nuts (Häckeberga, Sweden); C. leucostoma from tea leaves (Valparai, India); C. millenniana from larch branches (Brixen, Italy); C. nigri- cana from peas (Höör, Sweden); C. pyrivora from pears (Budapest, Hungary); C. splendana from oak glands (Veberöd, Sweden); E. foenella from rootstocks of mugwort (Artemisia vulgaris; Skäne, Sweden); N. roborana from rose buds (Hovs Hallar, Sweden). Adult males of G. funebrana, G. janthinana and G. loóarzewskii Now. were collected near Regensberg (Switzerland). Adults were kept in screen cages (30 l), fresh branches of their respective host plants were supplied daily. Courtship behaviours were video-recorded.
Pheromone extracts Pheromone glands of females were extracted at the onset of the calling period in batches of 12 to 44 (C. splendana; N = 5), 24 to 80 (C. fagiglandana; N = 4), 3 to 21 (C. pyrivora; N = 3), 80 and 92 (C. leucostoma; N =2), 4 to 16 (C. millenniana; N = 3), 18 to 43 (E. foenella; N = 4), 5 to 12 (N. roborana; N = 6), with 5 gl of heptane during 1 min. Hairpencils of G. funebrana, G. janthinana and G. lobarzewskii (8 to 27; N = 3) and scent scales of C. nigrieana males (20 to 87; N = 5) were extracted with 5 gl of hexane during 1 min. The occurrence of scent organs in Cydia males was investigated by scanning electron microscopy (Jeol T330). Insects were fixed and dehydrogenated in ethanol, preparations were critical-point dried and sputtered with 40:60 gold/palladium.
Pheromone biosynthesis in female glands A mixture of (E)-[ll,11,12,12,12-2H»]9-dodecenoic acid (Ds-E9- 12Acid; Löfstedt & Bengtsson 1988) and [ 14,14,14-2H3]-tetradecanoic acid (D3-14Acid; Larodan Fine Chemicals, Malmö, Sweden) in DMSO (0.2 lal; 10 gg/~tl) was applied to pberomone glands of C. splendana and C. nigricana females (N = 3) at the onset of their calling period. The glands were extracted in 5 gl heptane after 15 min of incubation.
The gland extracts were separated by gas chromatography (GC) on a DB-Wax column (see below), deuterium-labelled pheromone component analogues eluted slightly before the non-labelled compo- nents (Löfstedt & Bengtsson 1988). The amounts of labelled and unlabelled pheromonal compounds were estimated by selected ion monitoring on a mass spectrometer (MS; see below). The ions monitored were [M-60] + for native, D3- and D»-labelled dodecyl acetate (12Ac) (m/z 168.15, 171.15, 173.15); for (E)-9-dodecenyl acetate (E9-12Ae) (m/z 166.15, 169.15, 171.15); and M + for the native D 3 and D4-1abelled A8,A 10-12Ac isomers (m/z 224.15, 227.15, 228.15).
Gas chromatography/mass spectrometry Identification of female gland components was done on a Finnigan Mat TSQ700 MS, with electron impact ionization (70 eV/EMV 1200 Ecurr 400), coupled with a HP 5890 GC. Two columns were used, a DB-Wax + (30 m x 0.25 mm; J&W Scientific, Folsom, CA, USA), programmed from 60°C (hold 2min) at 10°/min to 230°C, and a OV-351 (25 m x 0.25 mm; GeneTec, Göteborg, Sweden), pro- grammed from 60°C (hold 2min) at 10°/min to 230°C. Injector
temperature was 200°C, the carrier gas (He) was constant at 0.9 ml/ min. Gland compounds were identified by retention times and mass spectra in comparison with synthetic compounds.
Further analysis of gland extracts and biosynthetic experiments were done on a Hewlett Packard 5970B MS, interfaced with a Hewlett Packard 5890 GC, with a DB-Wax column (30 m x 0.25 mm; J&W Scientifie), programmed from 80°C (hold 2 min) at 10°/min to 230°C. Analysis of isomeric composition was done on a Hewlett Packard 5890 GC with flame ionization detection on a nonpolar SE-54 column (25 m x 0.32 mm; Kupper & Co., Bonaduz, Switzer- land), programmed from 60°C (hold 2 min) at 10°/min to 100°C, and 1.5°/ruin to 230°C.
Field trapping
Chemicals, dispensers and traps All monoenic compounds, E8,E10-12Ac and E10,E12-14Ac were purchased from S. Voerman (Institute for Pesticide Research, Wa- geningen, The Netherlands). E,Z-, Z,E-, and Z,Z-isomers of 8,10- 12Ac and 10,12-14Ac were synthesized, and purified by high-pressure liquid chromatography (HPLC) (Reed & Chisolm 1985; Chisholm et al. 1985; Witzgall et al. 1993). Purity of these compounds, by GC, was >_ 97.3% (overall) and > 99.1% (isomeric).
Compounds in heptanic solution were formulated at 10 gg on red rubber septa (Merck ABS, Dietikon, Switzerland). Tetra traps (Arn et aL 1979) were hung at eye level to green branches, and were ca 5 m apart within one replicate. Traps and septa were exchanged every four to five weeks. The number of males caught was transformed to log(x + 1) and submitted to an analysis of variance, followed by a Tukey-test (P = 0.05).
Trap series and locations E,E-, E,Z-, Z,E-, and z8,zlo-12Ac, and their binary blends were screened in a decidous forest in Touraine (France; N = 3; June to September), in pear orchards at Budapest (Hungary; N = 9; June to August), in tea plantations at Valparai (India; N = 10; March and April), in coniferous forests at Brixen (Italy; N = 5; July to August), in various habitats in Skäne (Sweden; N = 50; May to August, three seasons), and in bushland and forests at Ephesus and Thermessos (Turkey; N =8; May). Trap pairs with E8,E10-12Ac on rubber septum or 3mg of a propheromone compound, tricarbonyl [(8,9,10,11-rl)-8,10-dodecadien-l-yl acetate] iron, releasing isomeri- cally pure E8,E10-12Ac (Streinz et al. 1993) were placed in pea fields (Höör, Sweden; N = 5; July).
E8-12Ac, Z8-12Ac, E10-12Ac, Z10-12Ac, and their binary blends (101ag per septum) were tested in a deciduous forest in Touraine (France; N = 3; June to September) and in Skäne (Sweden; N = 25; May to August, two seasons), adjacent to A8,A 10-12Ac trap series. The E8-/ZIO-12Ac- and Z8-/ElO-12Ac-blends were tested only during one season. E,E-, E,Z-, Z,E-, and Z10,Z12-14Ac, and their binary blends were sereened in Skäne (Sweden; N = 10; May to August), adjacent to A8,A10-12Ac trap series. Eight further trap series with gland compounds identified from C. fagiglandana, C. splendana, E. foenella, N. roborana (N = 10) were done in Skäne (Sweden).
Identification of trapped males Trapped males were in part identified by their wing pattern. Males of uncertain taxonomic status were identified by their genital morphol- ogy. The preparations are conserved at the Laboratoire de Biosystem- atique de l'Insecte (INRA) at Versailles (France).
Results and discussion
Field screening bioassay
T h e t o r t r i c i d species a t t r a c t e d to s ingle c o m p o u n d s a n d b i n a r y b l e n d s o f A 8 , A 1 0 - 1 2 A c ; A 8 - 1 2 A c a n d A10- 12Ac; a n d A 10,A 12 -14Ac a re s h o w n in T a b l e s 1 - 3 . T h e

Vol. 7, 1996 Tortricid pheromones and attractants 15
T a b l e 1 Field attraction of tortricid males to geometric isomers of A8,A 10-12Ac
Compound tag/trap
E8,E10-12Ac 10 10 10 10 E8,Z10-12Ac 10 10 10 10 Z8,EI0-12Ac 10 10 10 10 Z8,Z10-12Ac 10 10 10 10
EE EZ ZE Z Z EE/EZ EE/ZE EE/ZZ EZ/ZE E Z / Z Z Z E / Z Z
Number of males/replicate a'b S c N d L e
O L E T H R E U T I N A E O L E T H R E U T I N I Pseudosciaphila Obr.
brandneriana L. Hedya Hb.
dimidiana CI. dimidioalba Retz. ochroleucana Fröl. pruniana Hb.
EUCOSMINI Gypsonoma Meyr.
dealbana FröI. Notocelia Hb.
rosaecolana Dbld. trimaculana Hw.
Epiblema Hb. confusana H.-S. foenella L.
fuchsiana Rössl. seutulana D.&S. sticticana F.
Eucosma Hb. aemulana Schläg. cana Hw. hohenwartiana D.&S.
G R A P H O L I T I N I Latronympha Meyr.
strigana F. Grapholita Tr.
compositella F. gemmiferana Tr. jungiella C1.
Cydia Hb. blackmoreana Wlsm. duplieana Zett. fagiglandana Z. illutana H.-S. intexta Kuzn. leucostoma Meyr. millenniana Adamcz. nigricana F. pyrivora Danil. servillana Dup. splendana Hb.
strobilella L. succedana D.&S. turcianae Chambon
Dichrorampha Gn. petiverella L.
4 4
16 135
3
6
2
17
3 5
5 7 0 3
10 0
10 0 1 0 1 2 3 2
1 0
i 2 4 1 3 0 0
1
374
1
1 1
1 9
4 0 16
0 17 14 37
1 7
14 1 3 5 0 7 1
18
14 1 1
- 2 S
C 4 S 0 C,M 3 S 1 B 7 S
- 2 S
1 M 4 S
30
- 8 F,S M 2 S
13 2 - 3 F P 3 F
18 1 P 3 H 5 P 17 S 2 - 3 S,T 1 0 4 F,S 1 1 C 4 I ,S
5 3 B 11 F,S 0 0 7 H,S
0 5 4 - 8 H,S
214 17
4 B 10 F,S
3 18
7 5 1 0
1 1 1 1
7 S - 3 H,T
17 S
- 3 T
- 3 S,T P 8 S
8 I,F,S 3 S
I0 In 5 I
P 34 F,S P 4 H - 5 S P 8 S P 3 F - 8 I,S
11 F,H,I,S,T - 3 T
2 2 3 1 1 2 - 2 H,S
EE EZ ZE Z Z EE/EZ EE/ZE EE/ZZ EZ/ZE E Z / Z Z Z E / Z Z S ~ N d L e
a Rounded average number of males per replicate; b Bold-faced numbers significantly different from other captures at P = 0.05 (Tukey test); c Characterization status, according to literature ( A m et al. 1992, 1995) or to separate tests: pheromone (P), off-blend (B), compound missing (C), non-pheromonal compound or mimic (M); d Number of replicates, in which the respective species was trapped; e Trap location: France (F), Hungary (H), Italy (I), India (In), Sweden (S), Turkey (T); fAttract ion to E8,E10-12Ac-propheromone
16 P. Witzgall et al. CHEMOECOLOGY
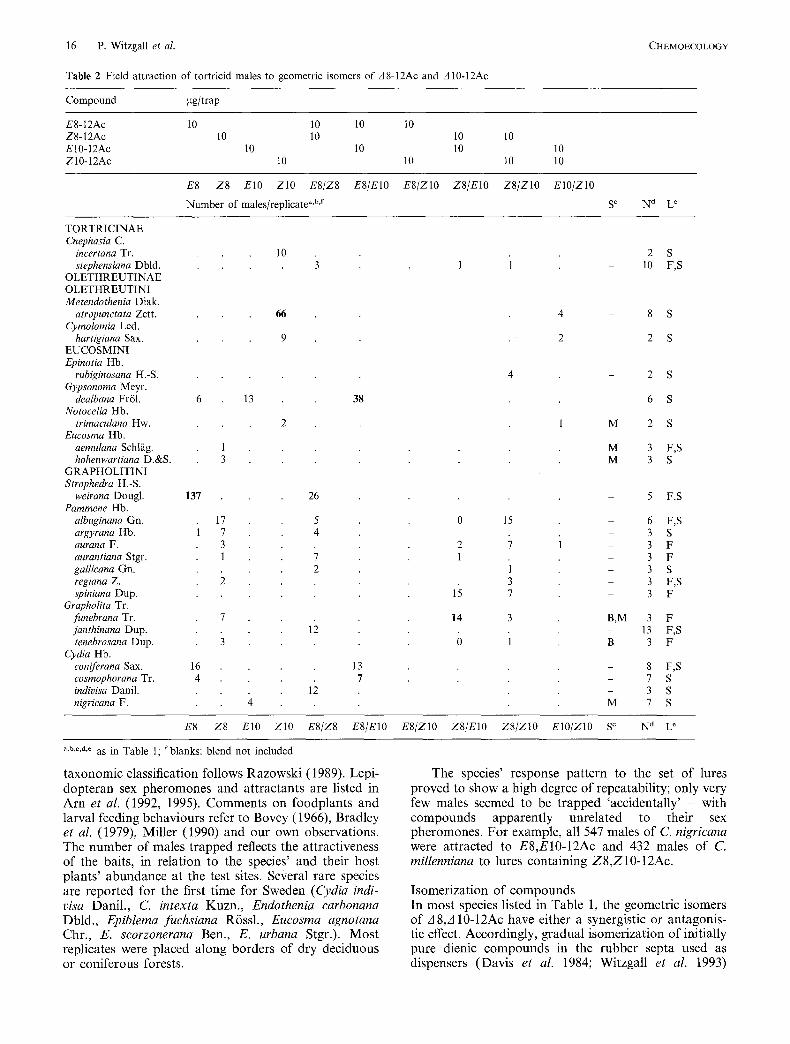
Table 2 Field attraction of tortricid males to geometric isomers of A8-12Ac and A 10-12Ac
Compound gg/trap
ES-12Ac 10 10 10 10 Z8-12Ac 10 10 10 10 E10-12Ac 10 10 10 10 Z10-12Ac 10 10 10 10
E8 Z8 El0 Z10 E8/Z8 E8/EIO E8/ZIO Z8/EIO Z8/ZIO EIO/ZIO
Number of males/replicate a'b'~ S c N d L e
TORTRICINAE Cnephasia C.
incertana Tr. stephensiana Dbld.
OLETHREUTINAE OLETHREUTINI Metendothenia Diak.
atropunctata Zett. Cymolomia Led.
hartigiana Sax. EUCOSMINI Epinotia Hb.
rubiginosana H.-S. Gypsonoma Meyr.
dealbana Fröl. Notocelia Hb.
trimaculana Hw. Eucosma Hb.
aemulana Schläg. hohenwartiana D.&S.
GRAPHOLITINI Strophedra H.-S.
weirana Dougl. Pammene Hb.
albuginana Gn. argyrana Hb. aurana F. aurantiana Stgr. gallicana Gn. regiana Z. spiniana Dup.
Grapholita Tr. funebrana Tr. janthinana Dup. tenebrosana Dup.
Cydia Hb. coniferana Sax. cosmophorana Tr. indivisa Danil. nigricana F.
1 3 7
i
16 4
17 7 3 1
i
13
10
6 6
9
26
5 4
7 2
38
13 7
- 2 S
1 1 - 10 F,S
0 15
2 1
i 3
15 7
1 4 3
0
4 8 S
2 2 S
M
M 3 M 3
- 5
6 3
- 3
- 3
3 3 3
2 S
6 S
2 S
F,S S
F,S
F,S S F F S F,S F
B , M 3 F
- 13 F,S B 3 F
- 8 F , S
- 7 S
- 3 S
M 7 S
E8 Z8 El0 Z10 E8/Z8 ES/EIO ES/ZIO Z8/EIO Z8/ZIO EIO/ZIO S c N d L e
a,b,c,<e as in Table 1; «blanks: blend not included
taxonomic classification follows Razowski (1989). Lepl- dopteran sex pheromones and attractants are listed in Arn et al. (1992, 1995). Comments on foodplants and larval feeding behaviours refer to Bovey (1966), Bradley et al. (1979), Miller (1990) and our own observations. The number of males trapped reflects the attractiveness of the baits, in relation to the species' and their host plants' abundance at the test sites. Se#eral rare species are reported for the first time for Sweden (Cydia indi- visa Danil., C. intexta Kuzn., Endothenia carbonana Dbld., Epiblema fuchsiana Rössl., Eucosma agnotana Chr., E. scorzonerana Ben., E. urbana Stgr.). Most replicates were placed along borders of dry deciduous or coniferous forests.
The species' response pattern to the set of lures proved to show a high degree of repeatability; only very few males seemed to be trapped 'accidentally' - with compounds apparently unrelated to their sex pheromones. For example, all 547 males of C. nigricana were attracted to E8,E10-12Ac and 432 males of C. millenniana to lures containing Z8,Z10-12Ac.
Isomerization of compounds In most species listed in Table 1, the geometric isomers of A 8,A 10-12Ac have either a synergistic or antagonis- tic effect. Accordingly, gradual isomerization of initially pure dienic compounds in the rubber septa used as dispensers (Davis et al. 1984; Witzgall et al. 1993)
V o l . 7, 1 9 9 6
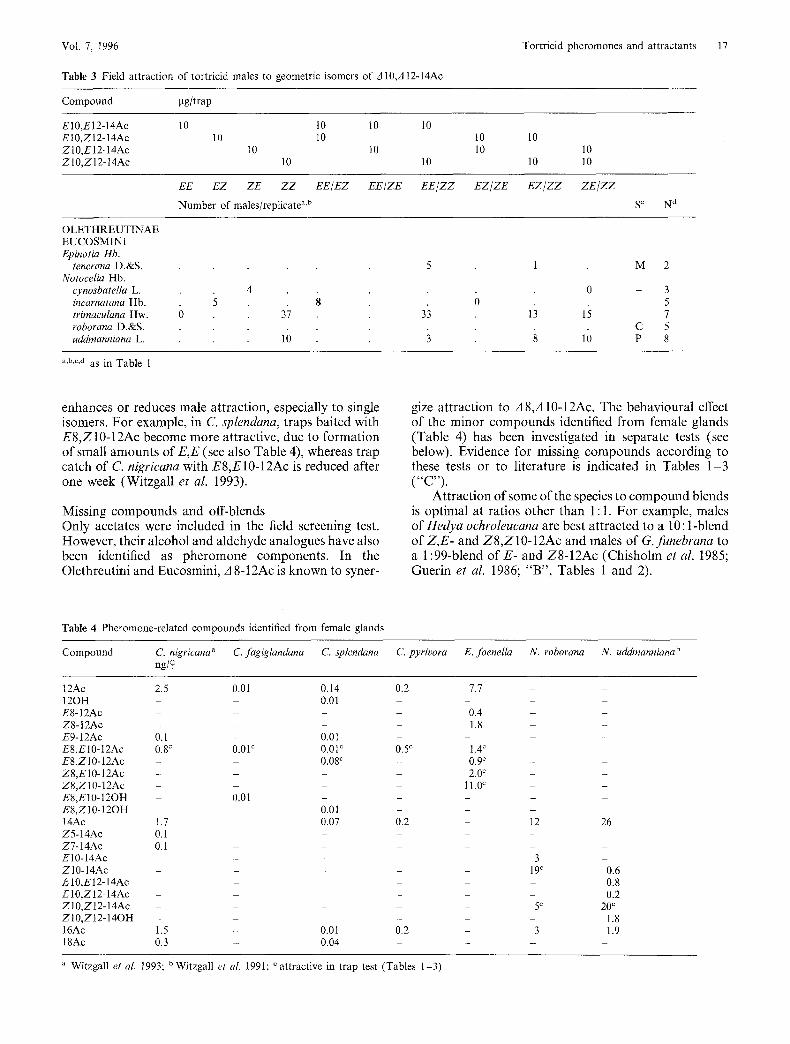
Table 3 Field attraction of tortricid males to geometric isomers of 3 1 0 , A 1 2 - 1 4 A c
Tortricid pheromones and attractants 17
Compound pg/trap
E 1 0 , E 1 2 - 1 4 A c 1 0 1 0 10 10
E 1 0 , Z I 2 - 1 4 A c 1 0 1 0 10 10
Z 1 0 , E I 2 - 1 4 A c I 0 10 10 10
Z 1 0 , Z 1 2 - 1 4 A c 10 10 10 10
EE EZ ZE Z Z EE/EZ EE/ZE E E / Z Z E Z / Z E E Z / Z Z Z E / Z Z
Number of males/replicate a'b S c N a
O L E T H R E U T I N A E
E U C O S M I N I
Epinotia Hb. tenerana D . & S .
Notocelia H b .
cynosbatella L. inearnatana H b .
trimaculana H w .
roborana D . & S .
uddmanniana L.
5
0 3 7
10
5 1 M 2
33
0 - 3
5
13 1; 7 C 5
lO P 8
a,b,c,d as in Table 1
enhances or reduces male attraction, especially to single isomers. For example, in C. splendana, traps baited with E8,Z 10-12Ac become more attractive, due to formation of small amounts of E,E (see also Table 4), whereas trap catch of C. nigricana with E8,E10-12Ac is reduced after one week (Witzgall et al. 1993).
Missing compounds and off-blends Only acetates were included in the field screening test. However, their alcohol and aldehyde analogues have also been identified as pheromone components. In the Olethreutini and Eucosmini, A 8-12Ac is known to syner-
gize attractlon to A 8,A 10-12Ac. The behavioural effect of the minor compounds identified from female glands (Table 4) has been investigated in separate tests (see below). Evidence for missing compounds according to these tests or to literature is indicated in Tables 1-3 ( " c " ) .
Attraction of some of the species to compound blends is optimal at ratlos other than 1 : 1. For example, males of Hedya ochroleucana are best attracted to a 10:1-blend of Z,E- and Z8,Z10-12Ac and males of G. funebrana to a 1:99-blend of E- and Z8-12Ac (Chisholm et al. 1985; Guerin et al. 1986; "B", Tables 1 and 2).
Table 4 Pheromone-related compounds identified from female glands
Compound C. nigricana a C. fagiglandana C. splendana C. pyrivora E. foenella N. roborana N. uddmanniana b n g / ~
1 2 A c 2 . 5 0 . 0 1 0 . 1 4 0 . 2 7 . 7 - -
1 2 O H - - 0 . 0 1
E 8 - 1 2 A c 0 . 4 - -
Z 8 - 1 2 A c . . . . 1 .8 - -
E 9 - 1 2 A c 0 .1 - 0 . 0 1 . . . .
E S , E 1 0 - 1 2 A c 0 . 8 c 0 . 0 1 ° 0 . 0 1 ° 0 . 5 c 1 . 4 ° -
E S , Z I 0 - 1 2 A c - 0 . 0 8 c - 0 . 9 ° -
Z 8 , E 1 0 - 1 2 A c - - 2 . 0 ° -
Z 8 , Z 1 0 - 1 2 A c - 1 1 . 0 ° -
E 8 , E 1 0 - 1 2 O H - 0 . 0 1 . . . .
E S , Z 1 0 - 1 2 O H 0 . 0 1 - - -
1 4 A c 1 .7 0 . 0 7 0 . 2 - 12 2 6
Z 5 - 1 4 A c 0 .1 . . . .
Z 7 - 1 4 A c 0 .1 . . . .
E 1 0 - 1 4 A c - - 3 -
Z 1 0 - 1 4 A c . . . . . 19 c 0 . 6
E 1 0 , E 1 2 - 1 4 A c - - - 0 . 8
E 1 0 , Z t 2 - 1 4 A c . . . . . 0 . 2
Z 1 0 , Z 1 2 - 1 4 A c - - 5 ° 2 0 c
Z 1 0 , Z 1 2 - 1 4 O H . . . . 1 .8
1 6 A c 1 .5 - 0 . 0 1 0 . 2 3 1 .9
1 8 A c 0 . 3 - 0 . 0 4 - -
a Witzgall et al. 1 9 9 3 ; u W i t z g a l l et al. 1 9 9 1 ; ~ attractive in trap test (Tables 1 - 3 )

18 P. Witzgall et al. CNEMOECOLOGY
Synergists and antagonists Most species are tuned to one isomer or to a blend of two isomers, the non-pheromonal isomers being strong attraction antagonists (Tables 1-3). This combination of synergists and antagonists enhances the specificity of the male response and enables communication in a number of species with only a few, biosynthetically related compounds. The response of G. jungiella to E8,Z10-12Ac was synergized by Z,E; but strongly re- duced by E,E and Z,Z, which are attractants for other species.
Moths do not respond to pheromone synergists alone, only to blends of synergists and the main phe- romone component. A majority of species responded accordingly to only one single compound, whereas N. rosaecolana, C. splendana (France), C. strobilella or C. succedana responded to two single isomers, and their blends. This may be explained by isomerization of the pure compounds in the trap dispensers (see above), in combination with an extremely wide male response window (E,E-septa contain at least 2.4% E,Z after orte week; Witzgall et al. 1993).
However, this may also reflect an individual varia- tion of the female production and male response, and even the occurrence of pheromone races. Swedish E. foenella responded to three single isomers identified from batches of female glands (Tables 1 and 4), even though addition of the Z,E isomer did reduce attraction to a blend of Z,Z- and E8,E10-12Ac in a separate test (see below). In Hungary, males responded best to a E,Z/Z,Z-blend, and in France only to the Z,E isomer.
Mimics and cross-attraction Closely related species often use different isomers of the same compounds, but there are examples for transitions between taxa, which are obviously facilitated by the male response to non-pheromonal, but structurally re- lated compounds or "mimics" ("M"; Tables 1-3). Notocelia were attracted to A 10,A12-14Ac, with the exception of N. rosaecolana, which was attracted to A8,A 10-12Ac, like the related Epiblema and Eucosma. Males of N. trimaculana responded to both compounds (Tables 1 and 3).
The monoenic E10-12Ac is used at 3 mg/trap for population monitoring of pea moth, C. nigricana (Wall 1988), instead of its sex pheromone E8,E10-12Ac (Ta- bles 1, 2 and 4). E10-12Ac mimics also E8-12Ac in G. funebrana; using a 99:l-blend of Z8- and E8-12Ac (Guerin et al. 1986; Table 2); and is a sex attractant for several Cydia and Pammene. Dienes mimic monoenes in C. coniferana and P. aurana (Rotundo & Giacometti 1988; Table 2) and in Gypsonoma dealbana and Hedya dimidioalba (nubiferana Hw.) (Tables 1 and 2). The sex pheromone of H. dimidioalba is a blend of E8,E10- 12Ac, Z8- and E8-12Ac (Frérot et al. 1979).
Sex pheromones and sex attractants of Grapholitini
Cydia, Grapholita and Pammene form a rather coherent and well-studied group within the Grapholitini. Most Cydia and some Grapholita are using A8,A10-12 sex
pheromone components, this is corroborated by GC/ MS analysis of female sex glands (Table 4) and male attraction in the field (Table 1). Other Cydia and Grapholita, as well as Pammene, are attracted to mo- noenic A8-12 or A 10-12 compounds (Table 2).
Cydia and Grapholita feeding on Leguminosae A propheromone compound, releasing isomerically pure E8,E10-12Ac (Streinz et al. 1993), was attractive to males of pea moth, C. nigricana (host plants Pisum, Vicia, Lathyrus), and G. compositelIa (Trifolium). How- ever, G. compositelIa was not attracted to E8,E10-12Ac on rubber septum. This suggests that G. compositella may be even more sensitive to the other isomers than C. nigricana (Table 1; Witzgall et al. 1993). In G. gem- miferana (Lathyrus), two males were also attracted to the E,E/Z,E-blend. G. jungiella feeds on leaves and pods of various peas and vetches (Lathyrus, Vicia) and the adults are day-active, as the pea moth and G. gemmiferana. A repellent blend of all four isomers is efficient for mating disruption of pea moth (Bengtsson et al. 1994; Witzgall et al. 1996a).
C. succedana males were best attracted to a E,E/ E,Z-blend, but responded also to both E,E and E,Z alone. In contrast, most other species responding to isomer blends were only attracted to one single com- pound. C. succedana (including its subspecies) is quite variable in size and coloration, and is feeding on seeds of Cytisus, Genista, Ulex, Sarothamnus, and Lotus. The exploitation of some hosts may require adaptations with respect to seasonal flight period, larval behaviour and metabolism. Cyanogenic compounds, for example, are found in Lotus (Bell 1972; Jones 1972). This may point to pheromone races associated with different hosts, rather than an extremely wide male response window. The other European species of the succedana- group are C. cytisanthana Burmann & Pröse (Genista), C. intexta (E,E; Dorycnium), C. ilipulana Wlsm., C. medicaginis Kuzn. (E,E; Bournoville 1979; Medicago), C. ulicetana Hw. (Ulex), C. vallesiaca Sauter (Ononis) (Danilevskij & Kuznetsov 1968; Burmann & Pröse 1988), some of which have been treated as forms and aberrations of C. succedana (Obratsov 1959).
Cydia and Grapholita feeding on seeds of Fagaceae, Pinaceae and Rosaceae The lavae of C. fagiglandaa feed on beech-nuts (Fagus sylvatica) and chestnuts (Castane sativa). In Swedish beech forests, males were attracted to E8,E10-12Ac, trap catch was not entirely reduced by Z8,E10-12Ac (Table 1). In a separate test, addition of 100% E8,E10- 12OH, identified from female glands (Table 4), did not increase trap catch significantly (see also Rotundo & Giacometti 1988). This component is an antagonist for attraction of C. nigricana and C. pyrivora to E8,E10- 12Ac (Witzgall et al. 1993; Makranczy et al. in prep.).
The sex pheromone of pear moth, C. pyrivora, the sibling species of codling moth, C. pomoneIla L., is E8,E10-12Ac (Tables 1 and 4). At a 5% addition, Z8,E10-12Ac had no effect on trap catch; a decline in trap catch with aged rubber septa was due to formation

Vol. 7, 1996 Tortricid pheromones and attractants 19
of the E,Z and Z,Z isomers, but was less pronounced tban in C. nigricana (Makranczy et al. in prep.). At- traction of C. pomonella to codlemone, E8,E10- 12OH, was significantly reduced by a 20%-addition of each of the four A8,A 10-12Ac isomers (Witzgall et al. 1996b).
C. splendana feeds on oak glands (Quercus spec.) and chestnuts (Castanea). In Swedish oak forests, males of the nominate and the melanistic form reau- murana Hein. were best attracted to E,E-/E8,ZIO- 12Ac (Tables 1 and 4). Additional tests showed a 2:10 and 7:10 E,E/E,Z-blend to be significantly more attractive than a 10:2 blend or E,Z alone; addition of E8,Z10-12OH did not enhance trap catch signifi- cantly. The response pattern of C. splendana in an oak/chestnut forest in Western France was different from Swedish oak forests (Table 1). Best attraction in chestnut plantations in Southern France was obtained with a blend of E8,E10-12Ac and E8,E10-12Ald (Frérot et al. 1995).
Of the cone-feeding species, C. strobilella (Picea) showed the same response pattern as C. succedana. C. illutana (Abies, Larix, Picea) responded to the E,E isomer.
Cydia mining in twigs and bark C. millenniana, mining in shoots of Larix, was the only Cydia species attracted to the Z,Z isomer. Trap catch was increased by E,E; even the other isomers may be synergists at lower proportions. The dendrophilous feeding habit and the male response pattern bear resem- blance to Eucosmini species; E. foenella females pro- duce all four isomers of AS,A 10-12Ac (Tables 1 and 4). In the laboratory and field, caged C. millenniana did not mate; gland extracts of non-calling females at the onset of the scotophase did not contain pheromonal com- pounds at > 3 pg/female. C. servillana mines in twigs of Salix, the males were attracted to the E,E isomer.
Of the bark-mining species, both C. coniferana and C. cosmophorana (Pinus) were attracted to E8- and E10-12Ac (Table 2; Booij et aI. 1986). C. indivisa (Picea), the sister species of C. cosmophorana, was attracted to a blend of E8- and Z8-12Ac, as the Grapholita of the subgenus Aspila (Euspila) Steph. (Table 2; Arn et al. 1992, 1995); species of the subgenus Grapholita Tr. use dienic pheromones (Table 1).
Other Cydia The tea flushworm, C. leucostoma, a major pest of tea in Southern India, was trapped with Z8,E10-12Ac. Lures were attractive over more than four weeks and may be used for detection and monitoring. The E,Z isomer had only a weak antagonistic effect. C. leucos- toma did not mate in the laboratory and female glands did not contain detectable amounts of acetates. C. turcianae, a species new to science, was attracted to E,E/E,Z (Chambon et al. 1993). Other species, trapped at only one or two locations, are not included in Table 1 (C. exquisitana Rbl., E,E/E,Z; C. ilipulana Wlsm., E,E; C. oxytropidis Wlsm., E,E; C. pactolana Z., E,E/ E,Z).
Other genera A majority of Pammene uses Z8-12Ac as the main compound, The E-isomer or the analogous A 10-12Ac may be synergists for some of rhein. P. rhediella is the only species attracted to a dienic compound, E8,Z10- 12OH (Guerin et al. 1983). The placing of Strophedra close to the Pammene/Grapholita-group (Danilevskij & Kuznetsov 1968) is corroborated (Table 2).
The position of Latronympha within the Grapholi- tini is not clear. Larval feeding in roots and sterns of Hypericum suggests that Latronympha is close to the Eucosmini; it has been treated as an early offshoot of the Cydia group by Danilevskij & Kuznetsov (1968). A separate test with four E,E/Z,Z blends showed best attraction of L. strigana males at a 1 : 10 ratio. Addition of Z,E to this blend strongly reduced male attraction - although binary Z,E/Z ,Z blends, at a 1:10 and 1:1 ratio did attract nearly as many males as Z,Z alone. This compares to the response pattern of E. foenella (see above; Table 1).
Biosynthetic pathway of female pheromone The female sex pheromone of C. nigricana is E8,EIO- 12Ac (Greenway 1984; Witzgall et al. 1993; Tables 1 and 4). Incorporation of D»-E9-12:Acid into D4- ES,E10-12Ac in female glands demonstrated a E9 de- saturation pathway, as in C. pomonelIa (Löfstedt & Bengtsson 1988). The amount of D4-1abelled acetate was at least twice the amount of unlabelled compound. A small but significant production of D3-E,E-8,10-12Ac from D3-14:Acid occurred; neither D3-14:Acid nor D»- E9-12: Acid were incorporated into 12Ac and E9-12Ac.
In C. splendana, both pheromone compounds, E,E- and ES,Z10-12Ac (Tables 1 and 4) are produced via the same pathway, as evidenced by incorporation of Ds-E9-12: Acid. The amounts of D4-1abelled analogues exceeded those of native isomers. We suggest that A S,A 10-12 aIcohols and acetates of the Olethreutinae are generally biosynthesized by A9 desaturation of 12:acyl derivatives and that a specific conversion of the E9 double bond to the A 8,A 10 system by 1,4-elimina- tion of water (Löfstedt & Bengtsson 1988) may gener- ate different geometric isomers.
Compounds identified from male androconia Abdominal scent organs are commonly found in the Grapholitini, they are absent in the Eucosmini (Ra- zowski 1990). Hair-like scent scales, connected to a sclerotization of the membrane between the eighth ster- nite and the vinculum of the genitalia are characteristic for Grapholita males. Hairpencil compounds and their behavioural activity have been studied in G. molesta Busck. (Baker et al. 1981; Nishida et al. 1982).
A preliminary identification of hairpencil com- pounds in G. funebrana showed eugenol (2-methoxy-4- [2-propenyl]-phenol) and an unknown derivative of abscisic acid (2-cyclohexen- 1-one,4-(3-hydroxy- 1- butenyl)-3,5,5-trimethyl), together with the hydrocar- bons C21 to C30. G. janthinana hairpencils contained methyl salicylate (2-hydroxy benzoic acid methyl ester) and an unidentified compound (m/z 192 M +, 174, 163,

20 P. Witzgall et al. CHEMOECOLOGY
135, 134, 105, 78, 77, 51, 39). G. lobarzewskii Now. hairpencils contained traces of eugenol, all saturated straight-chain hydrocarbons from C15 to C25 and their A9-monounsaturated analogues. The most abundant compound was A 9-heneicosen, probably the E-isomer. Compared to female-produced pheromones, there is a wide variation of male hairpencil compounds even in closely related species, which would in part be se- questered from the larval diet (Nishida et al. 1985; Löfstedt et al. 1989). These compounds are of minor importance for the tortricid phylogeny.
Abdominal scent organs are absent in Cydia, but scent organs were found along the anal vein of the hindwings of C. nigricana and C. fagiglandana. The bulb-shaped glandular structures (ca 10 gm o) are prob- ably transformed scales, with heliciodal thread-like ele- ments connect to a well-developed basal part. Extracts of the glandular region of male C. nigricana hindwings showed traces of an unknown compound which was not present in extracts of genital claspers or forewings. Extracts of ten male hindwings on filter paper elicited wingfanning in resting males and female attraction over a few cm.
Courtship behaviour The courtship sequence of pea moth, C. nigricana, and C. fagiglandana is extremely similar to the one de- scribed for Oriental fruit moth, G. molesta (Baker & Cardé 1979) - despite the entirely different morphology and location of the male androconia (see above).
After upwind orientation flight and landing at sev- eral cm from the pheromone-releasing female, the male wingfanns and rhythmically extends the genital claspers at the tip of the erect abdomen. Within less than one min, the female walks towards the male, and usually touches the male's hind legs or abdomen with her antennae. The male then turns into a parallel, head-to- head position with the female and attempts mating. Male wingfanning immediately elicits a wingfanning response in surrounding, previously inactive males.
In C. splendana, males wingfanned close to the female in 13 of 78 successful courtships. However, these males were not stationary, but rapidly moved in a circle around the female, with the head pointing towards it. The other males followed the same behavioural se- quence as C. pomonella (Castrovillo & Cardé 1980) and C. pyrivora, showing no involvement of male pheromones.
Male pheromones increase the specificity of sexual communication, especially in species which share host plants and apparently use a similar female sex pheromone, such as C. nigricana and C. gemmiferana (see also Phelan 1992). The close relation between Cydia and Grapholita is reemphasized by the corresponding courtship behaviours of G. molesta and C. nigricana.
Sex pheromones and sex attractants of Eucosmini
Epiblema and Eucosma In E. foenella (Artemisia vulgaris), a detailed test with various combinations of compounds extracted from
batches of up to 43 female glands (Table 4) showed a 1 : 1 blend of Z,Z- and ES,E10-12Ac to be more attrac- tive than a 1:10 or 10:1 blend (159, 43, 39 males; N = 10). Addition of Z8-12Ac or E8-12Ac did not augment trap catch. However, a 10%-addition of the female-produced Z,E isomer to the Z,Z/E,E-blend sig- nificantly reduced male attraction (17 males). In spite of that, males were attracted to Z8,E10-12Ac alone in various habitats in Sweden, and males were attracted only to this compound in France; tests in Hungary showed a synergistic role of E8,Z10-12Ac (Table 1).
Analysis of single glands is needed to determine whether this extremely heterogenous response of the male population corresponds to a variation in female production. Several forms have been described in E. foenella, according to a highly variable forewing pattern (Obratsov 1965; Bradley et al. 1979).
Other Epiblema responded also to Z8,Z10-12Ac. However, the species listed in Table 1 are likely to require different blend ratios, or additional compounds. In E. sticticana F. (farfarae Fletch.), ES,E10-12OH or Z8,Z 10-12OH may be an attraction synergist. E. scutu- lana is attracted also to Z8-12Ac (Arn et al. 1992, 1995). The close resemblance of Epiblema and Eueosma is demonstrated by a similar response pattern to A S,A 10-12Ac (Table 1). Eucosma agnotana Chr., E. scorzonerana Ben., E. urbana Stgr. (Sweden) and E. scutana Const. (France) were trapped in small numbers with blends containing Z8,Z10-12Ac.
Notocelia This genus has only recently been separated from Epiblema (Swatschek 1958; Obratsov 1965). The larvae of Epiblema feed in roots and sterns of Compositae, the larvae of NotoceIia mainly on leaves and buds of Rosaceae. In addition, A 10,A 12-14Ac is characteristic for Notocelia (Tables 3 and 4), only N. rosaecolana was trapped with A 8,A 10-12Ac (Table 1).
N. roborana is one of the few tortricid species using a monoene/diene sex pheromone blend. They were best attracted to a 10:2 blend of Z10-14Ac and Z10,Z12- 14Ac, not to either of the single compounds (see Table 3); the addition of E10-14Ac had no effect. Z10-14Ac strongly reduced attraction of N. trimaculana to Z10,Z12-14Ac. By comparison, there is no inter- or intraspecific behavioural role to monoenic gland com- pounds in Cydia. Most Olethreutinae are active be- tween late afternoon and early evening hours. N. roborana females called shortly after midnight (wide- spread wings), N. cynosbatella was active at dawn.
Chemotaxonomy
Taxonomic position and chemistry of the attractant signal are clearly correlated in tortricid species. Males of the subfamily Tortricinae respond mostly to mo- nounsaturated 14-carbon compounds. One exception is the genus Cnephasia, where monounsaturated 12-ac- etates are prevalent. Otherwise, mainly Olethreutinae are attracted to A8- and A 10-12Ac (Arn et al. 1992, 1995; Table 2).

Vol. 7, 1996 Tortricid pheromones and attractants 21
Species responding to A8,A 10-12Ac (Table l) be- long to the Olethreutinae, and most of them to the tribes Eucosmini and Grapholitini, which are consid- ered to be highest in the tortricid system. The use of diunsaturated compounds may be regarded as an ad- vanced character; the combination of synergistic and antagonistic isomers greatly increases pheromone spe- cificity.
Eucosmini and Grapholitini form the supertribe Eucosmidii, and are thought to have evolved from the Olethreutini (Kuznetsov & Stekolnikov 1984; Razowski 1976; Horak & Brown 1991), of which Hedya and possibly Pseudociaphila also use A8,A10-12 com- pounds. The Eucosmini comprise generalized leaf rollers, such as Epinotia and Gypsonoma, attracted mainly to monoenic A9-12 or A 8-12 compounds, and more derived internal feeders, such as Epiblema and Eucosma, at least some of which are attracted to A7,A9-12Ac or A8,A 10-12Ac (Table 1; Arn et al. 1992, 1995). The close relationship of Epiblema and Eucosma (Razowski 1976) and the separation of Epiblerna and Notocelia (Swatschek 1958; Obratsov 1965) are corrob- orated (Tables 1, 3, 4).
Cydia and related genera may have evolved from Eucosma or Epinotia (Brown 1984; Horak & Brown 1991), they use the same pheromone compounds as Hedya, Epiblema and Eucosma. However, male attrac- tion assays or identification of female gland compo- nents do not suffice to confirm the phylogenetic context, as long as we lack proof that these species produce A8,A 1Õ-12Ac via the same pathway.
In Cydia, the E9-12 minor gland component results from a E9 desaturation step, preceding biosynthesis of the A8,A10-12 main compound (Table 4; Arn et al. 1985; Löfstedt & Bengtsson 1988). In comparison, mi- nor A8-12 compounds accompany A8,AI0-12Ac in Epiblema (Table 4) and Hedya (Frérot et al. 1979; Roelofs & Brown 1982). This would hint different pathways in the Eucosmini and Grapholitini. On the other hand, A 7,A 9-12Ac is found in other Eucosrna and Epinotia (Arn et al. 1992, 1995) and A8,A 10-12, A7,A9- 12, as well as A8-12 and A10-12 acetates have been identified in Matsumuraeses falcana Wlsm. (Grapholi- tini) (Wakamura 1985). In addition, three genera of the Laspeyresiae group (Cydia, Grapholita, Pammene) in- clude species that respond to either A8,A 10-12 dienes, or to A8-12 and A10-12 monoenes. This is a strong indication for common biosynthetic pathways in both Eucosmini and Grapholitini.
Grapholita is close to Cydia, the males are distin- guished by the presence of male hairpencils between the 8th and 9th abdominal segment, which are connected to a sclerotization of the membrane (Razowski 1990). The presence of this sclerite in Grapholitini with and with- out hairpencils, following a similar courtship sequence (such as C. nigricana and G. molesta; see above), indi- cates a secondary reduction and questions the taxo- nomic value of this character (Razowski 1989, 1990).
The Grapholita of the subgenus Aspila use blends of E- and Z8-12Ac (Arn et al. 1992, 1995; Table 2) and feed on Rosaceae. In contrast, species of the subgenus
Grapholita using A8,A 10-12Ac (Table 1) all feed on Leguminosae, as the Cydia of the subgenus Endopisa (C. blackmoreana, C. intexta, C. nigricana, C. suc- cedana; Table 1). This indicates that the separation of Grapholita from Cydia is artificial (Obratsov 1959). The division of the Grapholitini into two subtribes, Laspeyresiae (Cydia, Grapholita, Pammene, Latro- nymphs, Strophedra) and Dicroramphae (Dicrorampha) (Danilevskij & Kuznetsov 1968) is supported by our field screening. Only D. petiverella was trapped (Table 1); the dienic acetates may function as pheromone mimics. The female production has not been studied in Dicrorampha.
Conclusion
The geometric isomers of A8,A10-12Ac are sex pheromone components in the tortricid tribes Eucos- mini and Grapholitini. C. splendana females produce two of them, E,E- and E8,Z10-12Ac, via E9 desatura- tion, and we assume that even the biosynthesis of Z,E and Z,Z in other species follows the same pathway.
E. foenella females produce all four isomers (Table 4). Glands of C. pomonella contain, along with several ng of E8,EI0-12OH, a few percent of two non- pheromonal isomers (Arn et al. 1985). Cydia gland extracts investigated in this study contained only very little pheromone (Table 4), but it is conceivable that the females produce also the other isomers, if only in trace amounts. Production of different isomers in closely related species, and the varying degree of male toler- ance towards the nonpheromonal isomers, in species using E8,E10-12Ac (Tables 1 and 4), further substanti- ate this hypothesis.
It is unknown, whether one or more enzymes are involved in the conversion of E9-12:acid into the four isomers, but the perception of the isomers is probably encoded in compound-specific proteins in the male an- tennae (see Bengtsson et al. 1990; Du & Prestwich 1995). In the European corn borer, Ostrinia nubilalis, biosynthetic control of the ratio of a two-component pheromone blend and the electrophysiological response of male olfactory sensilla to these compounds are inde- pendently inherited (Roelofs et al. 1987). However, these findings still leave room for speculations on devel- opmental links between the female gland and the male antennae during sexual differentiation (White et al. 1994) or on the inheritance of protein elements involved in pheromone biosynthesis and perception as coadapted complexes (see Coyne et al. 1988).
The males perceive all four isomers: each one func- tions either as main pheromone compound, attraction synergist or antagonist, and elicits, enhances or pre- vents male attraction (Table 1). The differential re- sponse of related species to the same compounds illustrates that the sensory physiology of attraction synergism and antagonism is very much alike. The respective synergists and antagonists are perceived via specialized antennal receptors (Priesner 1979, 1986), they are processed in the same glomeruli in the olfac-

22 P. Witzgall et al. CHEMOECOLOGY
tory lobe (Christensen & Hildebrand 1994), and they are behaviourally active in blends with the main com- pound (Linn et al. 1986; Witzgall & Priesner 1991). The cross-combination of synergists and antagonists greatly increases the specificity and the diversity of the female signals, which, within phylogenetic groups, are com- posed of only a few, structurally related compounds.
In the more advanced tortricid taxa, we observe a transition from monoenic to dienic pheromones (Arn et al. 1992, 1995). The occurrence of a variety of isomer blends and male perception of both conspecific and heterospecific compounds (Table 1) demonstrates the role of interspecific competition as a selective force (Phelan 1992; Löfstedt 1993; Linn & Roelofs 1994) and the potential for reproductive character displacement in the evolution of pheromone blends (Butlin 1994). The "Recognition Concept of Species" emphasizes the role of the physical environment (Paterson 1985; Lambert et al. 1987; White et al. 1994), but should also accomodate the weight of biotic factors. In the Grapholitini and Eucosmini, divergence in mating signals is promoted by concurrent biosynthetic availability and male percep- tion of the A8,A 10-12 isomers.
Acknowledgements
This study was funded by the Swedish Natural Science Research Council (NFR) and the Erik and Ebba Larssons & Thure Rignells Foundation (Tranemäla). We thank the Department for Environmental Protec- tion, County Administration of Kristianstad, and Rurik Tham for their permission to do field tests at Hovs Hallar, Kivik, Sandhammaren and Häckeberga.
References
Arn H, Rauscher S, Schmid A (1979) Sex attractant formulations and traps for the grape moth Eupoecilia ambiguella Hb. Mitt schweiz entomol Ges 52:49-55
Arn H, Guerin PM, Buser H-R, Rauscher S, Mani E (1985) Sex pheromone blend of the codling moth, Cydia pomonella: evi- dence for a behavioural role of dodecan-l-ol. Experientia 41:1482-1483
Arn H, Tóth M, Priesner E (1992) List of sex pheromones of Lepidoptera and related attractants. 2nd ed. F-Montfavet: Inter- national Organization for Biological and Integrated Control
Arn H, Tóth M, Priesner E (1995) The Pherolist. Internet edition: Http://www.nysaes.cornell.edu/pheronet
Baker TC, Cardé RT (1979) Courtship behavior of the Oriental fruit moth (Grapholitha molesta): experimental analysis and consider- ation of the role of sexual selection in the evolution of courtship pheromones in the Lepidoptera. Ann entomol Soc Am 72:173- 188
Baker TC, Nishida R, Roelofs WL (1981) Close-range attraction of female Oriental fruit moth to herbal scent of male hairpencils. Science 214:1359-1361
Bell EA (1972) Toxic amino acids in the Leguminosae. Pp 163-177 in Harborne JB (ed.) Phytochemical Ecology. GB-London: Aca- demic Press
Bengtsson M, Liljefors T, Hansson BS, Löfstedt C, Copaja SV (1990) Structure-activity relationships for chain-shortened analogs of (Z)-5-decenyl acetate, a pheromone component of the turnip moth, Agrotis segetum. J chem Ecol 16:667-684
Bengtsson M, Karg G, Kirsch PA, Löfqvist J, Sauer A, Witzgall P (1994) Mating disruption of pea moth Cydia nigricana F. (Lepi- doptera: Tortricidae) by a repellent blend of sex pheromone and attraction inhibitors. J chem Ecol 20:871-887
Booij CJH, Voerman S, Willemse LPM (1986) Conifer-infesting tortricids in Western Europe: A search for more potent and selective sex attractants. Z angew Entomol 102:253 259
Bournoville R (1979) Determination, au champ, d'attractifs sexuels de synthèse des mäles de deux Tortricidae (Lep): Laspeyresia medicaginis Kuzn. et Laspeyresia nigricana F., nuisibles aux legumineuses cultivées. Rev Zool agric Pathol vég 78:41-48
Bovey P (1966) Super-famiUe des Tortricoidea. Pp 456-893 in Bala- chowsky AS (ed,) Entomologie Appliquée ä l'Agriculture, Vol. II. F-Paris: Masson
Bradley JD, Tremewan WG, Smith A (1973) British Tortricoid Moths. Cochylidae and Tortricidae: Tortricinae. GB-London: The Ray Society
Bradley JD, Tremewan WG, Smith A (1979) British Tortricoid Moths. Tortricidae: Olethreutinae. GB-London: The Ray Society
Brown RL (1984) Review of Corticivora (Lepidoptera: Tortricidae) with analysis of its tribal relationships and descriptions of new species. Proc entomol Soc Wash 86:278-286
Burmann K, Pröse H (1988) Eine neue Cydia aus den Südalpen: Cydia cytisanthana n. sp. St Trent Sci Nat 64:191-199
Butlin R (1994) Genetic variation in mating signals and responses. Pp 327-366 in Lambert DM, Spencer H (eds) Speciation and the Recognition Concept: Theory and Application. Baltimore/MD: Johns Hopkins Univ Press
Cardé RT, Cardé AM, Hill AS, Roelofs WL (1977) Sex attractant specificity as a reproductive isolating mechanism among the sibling species Archips argyrospilus and mortuanus and other sympatric tortricine moths (Lepidoptera: Tortricidae). J chem Ecol 3:71-84
Castrovillo PJ, Cardé RT (1980) Male codling moth (Laspeyresia pomonella) orientation to visual cues in the presence of pheromone and sequences of courtship behaviors. Ann entomoI Soc Am 73:100-105
Chambon JP, Witzgall P, Bengtsson M (1993) Une nouvelle tordeuse en Turquie, Cydia turcianae n. sp. (Lepidoptera, Tortricidae). Bull Soc entomol France 98:181-183
Chisholm MD, Reed DW, Underhill BW, Palaniswamy P, Wong JW (1985) Attraction of tortricid moths of subfamily Olethreuthinae to field traps baited with dodecadienes. J chem Ecol 11:217-229
Christensen TA, Hildebrand JG (1994) Neuroethology of sexual attraction and inhibition in heliothine moths. Pp 37-46 in Schildberger K, Elsner N (eds) Fortschritte der Zoologie 39: Neural Basis of Behavioural Adaptations. D-Stuttgart: Gustav Fischer
Coyne JA, Orr HA, Futuyma DJ (1988) Do we need a new species concept? Syst Zool 37:190-200
Danilevskij AS, Kuznetsov VI (1968) Listovertki Tortricidae: triba plodozhorki Laspeyresiini in Fauna SSSR, Nasekomye Tcheshuekrylye. Vol. 5. Leningrad: Nauka
Davis HG, McDonough LM, Burditt Jr AK, Bierl-Leonhardt BA (1984) Filbertworm sex pheromone. Identification and field tests of (E,E)- and (E,Z)-8,10-dodecadien-l-ol acetates. J chem Ecol 10:53-61
Du G, Prestwich GD (1995) Protein structure encodes the ligand binding specificity in pheromone binding proteins. Biochemistry 34:8726-8732
Foster SP, Dugdale JS, White CS (1991) Sex pheromones and the status of greenheaded and brownheaded leafroller moths in New Zealand. N Z J Zool 18:63-74
Frérot B, Priesner E, Gallois M (1979) A sex attraetant for the green budworm moth, Hedya nubiferana. Z Naturforsch 34c:1248- 1252
Frérot B, Marro JP, Malosse C (1995) In vitro incubation of sex pheromone gland and identification of pheromone components in Cydia splendana (Hb). C R Acad Sc Paris 318(III):447-451
Greenway AR (1984) Sex pheromone of the pea moth, Cydia nigri- cana (F.) (Lepidoptera: Olethreutidae). J chem Ecol 10:973-982
Guerin PM, Arn H, Blaser C, Lettéré M (1983) Z,E-8,10-dodecadien- 1-ol, attractant for male Pammene rhediella. Entomol exp appl 33:346-347

Vol. 7, 1996 Tortricid pheromones and attractants 23
Guerin PM, Arn H, Buser HR, Charmillot P, Tóth M, Sziräki G (1986) Sex pheromone of Grapholita funebrana. Occurrence of Z-8- and Z-10-tetradecenyl acetate as secondary components. J chem Ecol 12:1361-1368
Hor'ak M, Brown RL (1991) Taxonomy and phylogeny. Pp 23-48 in van der Geest LPS, Evenhuis HH (eds) Tortricid Pests, their Biology, Natural Enemies and Control. NL-Amsterdam: Elsevier
Jones DA (1972) Cyanogenic glycosides and their function. Pp 103- 124 in Harborne JB (ed.) Phytochemical Ecology. GB-London: Academic Press
Katovich SA, Swedenborg PD, Giblin M, Underhill EW (1989) Evidence for (E,Z)-8,10-dodecadienyl acetate as the major compo- nent of the sex pheromone of the eastern pine seedworm, Cydia toreuta (Lepidoptera: Tortricidae). J chem Ecol 15:581 590
Lambert DM, Michaux B, White CS (1987) Are species self-defining? Syst Zool 36:196 205
Linn CE, Roelofs WL (1994) Pheromone communication in moths and its role in the speciation process. Pp 263-300 in Lambert DM, Spencer H (eds) Speciation and the Recognition Concept: Theory and Application. Baltimore/MD: Johns Hopkins Univ Press
Linn Jr CE, Campbell MG, Roelofs WL (t986) Male moth sensitivity to multicomponent pheromones: critical role of female-released blend in determining the functional role of components and active space of the pheromone. J chem Ecol 12:659-668
Löfstedt C (1993) Moth pheromone genetics and evolution. Phil Trans R Soc Lond B 340:167-177
Löfstedt C, Bengtsson M (1988) Sex pheromone biosynthesis of (E,E)-8,10-dodecadienol in codling moth Cydia pomonella in- volves E9 desaturation. J chem Ecol 14:903-915
Löfstedt C, Vickers NJ, Roelofs WL, Baker TC (1989) Diet related courtship success in the Oriental fruit moth, Grapholita molesta (Tortricidae). Oikos 55:402-408
McDonough LM, Eikenbary RD, Smith MT, Davis HG, Smithhisler CL, Hedger GA, Campbell RK, Payne JA, Reid W, McVay JR (1990) Sex pheromone of hickory shuckworm Cydia caryana. Development of an effective field lure. J chem Ecol 16:317-324
Miller WE (1990) Body size and diet quality in the genus Cydia (Tortricidae). J Lep Soc 44:1t3-142
Nishida R, Baker TC, Roelofs WL (1982) Hairpencil pheromone components of male Oriental fruit moths, Grapholita molesta. J chem Ecol 8:947-959
Nishida R, Fukami H, Baker TC, Roelofs WL, Acree TE (1985) Oriental fruit moth pheromone: attraction of females by an herbal essence. Pp 47-63 in Acree TE, Soderlund DM (eds) Semiochemistry: Flavors and Pheromones. D-Berlin: de Gruyter
Obratsov NS (1959) Die Gattungen der palaearktischen Tortricidae. II. Die Unterfamilie Olethreutinae. 2. Teil. Tijdschr Entomol 102:175 215
Obratsov NS (1965) Die Gattungen der palaearktischen Tortricidae. 1I. Die Unterfamilie Olethreutinae. 6. Teil. Tijdschr Entomol 108:365 387
Paterson HEH (1985) The recognition concept of species. Pp 21-29 in Vrba ES (ed.) Species and Speciation. Pretoria: Transvaal Museum Monograph No. 4
Phelan PL (1992) Evolution of sex pheromones and the role of assymetric tracking. Pp 265-314 in Roitberg BD, Isman MB (eds) Insect Chemical Ecology: An Evolutionary Approach. New York: Chapman & Hall
Priesner E (1979) Specificity studies on pheromone receptors of noctuid and tortricid Lepidoptera. Pp 57-71 in Ritter FJ (ed.) Chemical Ecology: Odour Communication in Animals. NL-Am- sterdam: Elsevier
Priesner E (1986) Correlating sensory and behavioural responses in multichemical pheromone systems of Lepidoptera. Pp 225-233 in Payne TL, Birch MC, Kennedy CEJ (eds) Mechanisms in Insect Olfaction. GB-Oxford: Clarendon
Razowski J (1976) Phylogeny and system of Tortricidae (Lepi- doptera). Acta zool Cracov 21:73 120
Razowski J (1989) The genera of Tortricidae (Lepidoptera). Part II: Palaearctic Olethreutinae. Acta zool Cracov 32:107-328
Razowski J (1990) Abdominal scent organs in Tortricidae (Lepi- doptera). Acta zool Cracov 33:521-554
Reed DW, Chisholm MD (1985) Attraction of moth species of Tortricidae, Gelechiidae, Geometridae, Drepanidae, Pyralidae, and Gracillariidae families to field traps baited with conjugated dienes. J chem Ecol 11:1645-I657
Roelofs WL, Brown RL (1982) Pheromones and evolutionary rela- tionships of Tortricidae. Annu Rev Ecol Syst 13:395 422
Roelofs WL, Glover T, Tang X-H, Sreng I, Robbins P, Eckenrode C, Löfstedt C, Hansson BS, Bengtsson B-O (1987) Sex pheromone production and perception in European corn borer moths is determined by both autosomal and sex-linked genes. Proc Natl Acad Sci 84:7585 7589
Rotundo G, Giacometti R (1988) Individuazione di una miscela attrattiva per maschi di Cydiafagiglandana Z. (Lep. Tortricidae) mediante studio di campo. Boll Lab entomol agr Portici 45:81- 97
Silk PJ, Kuenen LPS (1988) Sex pheromones and behavioral biology of the coniferophagous Choristoneura. Annu Rer Entomol 33:83-101
Streinz L, Horak A, Vrkoc J, Hrdy I (1993) Propheromones derived from codlemone. J chem Ecol 19:1-9
Swatschek B (1958) Die Larvalsystematik der Wickler (Tortricidae und Carposinidae). D-Berlin: Akademie-Verlag
Wakamura S (1985) Identification of sex-pheromone components of the podborer, Matsumuraeses falcana (Walsingham) (Lepi- doptera: Tortricidae). Appl Entomol Zool 20:189-198
Wall C (1988) Application of sex attractants for monitoring the pea moth, Cydia nigricana (F.) (Lepidoptera: Tortricidae). J chem Ecol 14:1857-1866
White CS, Lambert DM, Foster SP (1994) ChemicaI signals and the recognition concept. Pp 301-326 in Lambert DM, Spencer H (eds) Speciation and the Recognition Concept: Theory and Ap- plication. Baltimore/MD: Johns Hopkins Univ Press
Witzgall P, Priesner E (1991) Wind tunnel study on an attraction inhibitor in male Coleophora laricella (Lepidoptera: Coleophori- dae). J chem Ecol 17:1355-1362
Witzgall P, Frérot B, Malosse C (1991) Identification du composé majoritaire de ta sécrétion phéromonale de Notocelia uddmanni- ana L. (Lep., Tortricidae). J appl Entomol 112:71-75
Witzgall P, Bengtsson M, Unelius CR, Löfqvist J (1993) Attraction of pea moth Cydia nigricana F. (Lepidoptera: Tortricidae) to female sex pheromone (E,E)-8,10-dodecadien-l-yl acetate, is in- hibited by geometric isomers E,Z, Z,E and Z,Z. J chem Ecol 19:1917-1928
Witzgall P, Bengtsson M, Karg G, Bäckman A-C, Streinz L, Kirsch PA, Blum Z, Löfqvist J (1996a) Behavioral observations and measurements of aerial pheromone concentrations in a mating disruption trial against pea moth Cydia nigricana F. (Lepi- doptera, Tortricidae). J chem Ecol 22:191-206
Witzgall P, Bäckman A-C, Svensson M, Bengtsson M, Unelius CR, Vrkoc J, Kirsch PA, Ioriatti C, Löfqvist J (1996b) Potential of a blend of E8,E10-12OH and E8,E10-12Ac for mating disruption of codling moth, Cydia pomonella L. (Lep., Tortricidae). J appl Entomol: in press
Related Documents











