Sex Pheromone of the Pine False Webworm Acantholyda erythrocephala Joseph K. Staples & Robert J. Bartelt & Allard A. Cossé & Douglas W. Whitman Received: 24 September 2009 / Revised: 16 December 2009 / Accepted: 16 December 2009 / Published online: 12 January 2010 # Springer Science+Business Media, LLC 2010 Abstract Females of the pine false webworm Acantholyda erythrocephala (L) produce the sex pheromone (Z)-6, 14-pentadecadienal, which attracts flying males in the field. By using gas chromatography coupled with electroantenno- graphic detection (GC-EAD) and mass spectrometry (GC-MS), we detected (Z)-6,14-pentadecadienal in volatile collec- tions and in whole body extracts of female A. erythroce- phala. Females, but not males, also exhibited a 25-carbon cuticular hydrocarbon, ( Z,Z)-1,9,15-pentacosatriene, which can oxidize to (Z)-6,14-pentadecadienal upon exposure to air and sunlight. (Z,Z)-1,9,15-Pentacosatriene and (Z)-6,14-pentadecadienal identifications were corrob- orated by comparison with synthetic standards. (Z)-6, 14-Pentadecadienal is the second pheromone identified for pamphilliid sawflies, and the first to elicit strong field attraction, and thus offer potential as a pheromone lure to aid in control of this forest pest. Keywords Sex pheromone . Pine false webworm . Sawfly . Acantholyda erythrocephala . Cuticular hydrocarbon . Abitotic oxidation . Nonanal . Decanal . (Z)-6,14- pentadecadienal . Pentacosane . (Z)-9-pentacosene . (Z)-1,9-pentacosadiene . (Z,Z)-1,9,15-pentacosatriene . Hymenoptera . Pamphiliidae Introduction Acantholyda erythrocephala (L) (Hymenoptera: Pamphilii- dae), the pine false webworm (PFW), is common to northern Europe and Asia (MiddleKauff 1958). It was reported in North America during the early 1900’ s where it has become a significant pest of white pine (Pinus strobus L.), red pine (P. resinosa Ait), and Scots pine (P. sylvestris L) in both forests and plantations (Syme 1981; Johnson and Lyon 1991; Allen 2000). Outbreaks in the U.S. and Canada can encompass thousands of hectares and persist for 20 yr or more (Asaro and Allen 1999; Allen 2000, personal observa- tion), resulting in a growth reduction and increased mortality of host trees (MiddleKauff 1958; Syme 1981; Moody 1990; Lyons 1994; Asaro and Allen 2001; Mayfield et al. 2005). Numerous strategies considered as a means of control for this pest have met with variable success including: applica- tion of synthetic pesticides (Lyons et al. 1993), natural toxins such as neem extracts (Helson et al. 1999; Lyons et al. 1996, 1998, 2003), biological control (Wilson 1984; Lyons 1999; Kenis and Kloosterman 2001; Thompson et al. 2003). However, many of these strategies require precise knowledge of the distribution and phenology of PFW over large tracts of forest. As such, a pheromone lure could be valuable for monitoring, trapping, or disrupting A. erythrocephala. J. K. Staples (*) Department of Environmental Science, University of Southern Maine, 37 College Avenue, Gorham, ME 04038, USA e-mail: [email protected] R. J. Bartelt : A. A. Cossé Crop Bioprotection Research Unit, USDA, ARS National Center for Agricultural Utilization Research, Peoria, IL 61604, USA D. W. Whitman 4120 School of Biological Sciences, Illinois State University, Normal, IL 61790, USA J Chem Ecol (2009) 35:1448–1460 DOI 10.1007/s10886-009-9736-z

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Sex Pheromone of the Pine False Webworm Acantholydaerythrocephala
Joseph K. Staples & Robert J. Bartelt & Allard A. Cossé &
Douglas W. Whitman
Received: 24 September 2009 /Revised: 16 December 2009 /Accepted: 16 December 2009 /Published online: 12 January 2010# Springer Science+Business Media, LLC 2010
Abstract Females of the pine false webworm Acantholydaerythrocephala (L) produce the sex pheromone (Z)-6,14-pentadecadienal, which attracts flying males in the field.By using gas chromatography coupled with electroantenno-graphic detection (GC-EAD) andmass spectrometry (GC-MS),we detected (Z)-6,14-pentadecadienal in volatile collec-tions and in whole body extracts of female A. erythroce-phala. Females, but not males, also exhibited a 25-carboncuticular hydrocarbon, (Z,Z)-1,9,15-pentacosatriene,which can oxidize to (Z)-6,14-pentadecadienal uponexposure to air and sunlight. (Z,Z)-1,9,15-Pentacosatrieneand (Z)-6,14-pentadecadienal identifications were corrob-orated by comparison with synthetic standards. (Z)-6,14-Pentadecadienal is the second pheromone identified forpamphilliid sawflies, and the first to elicit strong fieldattraction, and thus offer potential as a pheromone lure toaid in control of this forest pest.
Keywords Sex pheromone . Pine false webworm . Sawfly .
Acantholyda erythrocephala . Cuticular hydrocarbon .
Abitotic oxidation . Nonanal . Decanal . (Z)-6,14-pentadecadienal . Pentacosane . (Z)-9-pentacosene .
(Z)-1,9-pentacosadiene . (Z,Z)-1,9,15-pentacosatriene .
Hymenoptera . Pamphiliidae
Introduction
Acantholyda erythrocephala (L) (Hymenoptera: Pamphilii-dae), the pine false webworm (PFW), is common to northernEurope and Asia (MiddleKauff 1958). It was reported inNorth America during the early 1900’s where it has becomea significant pest of white pine (Pinus strobus L.), red pine(P. resinosa Ait), and Scots pine (P. sylvestris L) in bothforests and plantations (Syme 1981; Johnson and Lyon 1991;Allen 2000). Outbreaks in the U.S. and Canada canencompass thousands of hectares and persist for 20 yr ormore (Asaro and Allen 1999; Allen 2000, personal observa-tion), resulting in a growth reduction and increased mortalityof host trees (MiddleKauff 1958; Syme 1981; Moody 1990;Lyons 1994; Asaro and Allen 2001; Mayfield et al. 2005).Numerous strategies considered as a means of control forthis pest have met with variable success including: applica-tion of synthetic pesticides (Lyons et al. 1993), natural toxinssuch as neem extracts (Helson et al. 1999; Lyons et al. 1996,1998, 2003), biological control (Wilson 1984; Lyons 1999;Kenis and Kloosterman 2001; Thompson et al. 2003).However, many of these strategies require precise knowledgeof the distribution and phenology of PFWover large tracts offorest. As such, a pheromone lure could be valuable formonitoring, trapping, or disrupting A. erythrocephala.
J. K. Staples (*)Department of Environmental Science,University of Southern Maine,37 College Avenue,Gorham, ME 04038, USAe-mail: [email protected]
R. J. Bartelt :A. A. CosséCrop Bioprotection Research Unit, USDA,ARS National Center for Agricultural Utilization Research,Peoria, IL 61604, USA
D. W. Whitman4120 School of Biological Sciences, Illinois State University,Normal, IL 61790, USA
J Chem Ecol (2009) 35:1448–1460DOI 10.1007/s10886-009-9736-z

Prior to this research, laboratory studies that used a twochoice olfactometer found that male A. erythrocephalaexhibit a series of characteristic behaviors in response tofemale derived volatiles, as well as to decanal. The lattercan be obtained from aerations and whole body extracts ofboth sexes (Staples 1999). Male behaviors includedantennation, upwind flight, and courtship behavior thatinvolved abdominal flexing accompanied by substratevibration (Staples 1999). These responses were similar tolaboratory and field data from another Pamphilliid sawfly,the web-spinning larch sawfly, Cephalcia lariciphila, inresponse to female derived odors and to a likely pheromonecomponent, ortho-aminoacetophenone (Borden et al. 1978;Baker et al. 1983).
In this paper, we report that female A. erythrocephalaproduce an attractant pheromone, (Z)-6,14-pentadecadienal,to which only males respond. We demonstrate that thesynthetic compound attracts male sawflies in the field, andprovide preliminary evidence indicating that (Z)-6,14-pentadecadienal can be produced via abiotic oxidation ofa long-chain unsaturated hydrocarbon found exclusively infemales.
Methods and Materials
Life History In North America, adult Acantholyda eryth-rocephala emerge from underground in spring (March toMaydepending on latitude) to mate and oviposit (MiddleKauff1958; Lyons 1994; Asaro and Allen 1999). Flight seasonlasts for approximately 3 wk, depending on local climate andweather conditions. Females oviposit on the previous year’sneedles. Larvae hatch approximately 10 d later and feedgregariously from within protective webs for 18–21 d(Middlekauff 1958). In May to July (depending on latitude),last-instars drop to the forest floor and burrow 1–15 cm intothe soil (Jahn 1967; MiddleKauff 1938; Asaro and Allen1999). Larvae overwinter in earthen cells as bright greenpronymphs. Pupation occurs in the following spring (Jahn1967), followed by adult emergence. Adult A. erythroce-phala are strongly phototropic and readily bask in directsunlight on cool days. As such, population densities usuallyare greater along sunny forest edges or in well-lit patches onthe forest floor.
Collection and Rearing of Live Insects Most A. erythroce-phala used in this research were collected as pronymphs inSand Ridge State Forest (Mason County, IL, USA) duringfall 2001, 2002, and 2003, by digging with a shovel underdefoliated trees to a depth of approximately 15 cm. Thebright green pronymphs were identified readily and collectedby hand. Pronymphs were transported to the laboratory andplaced in 31×8×6 cm plastic trays, 100–200 per tray, on
moist, silica sand, 3 cm to 4 cm deep. Trays containing insectswere then placed in growth chambers under total darkness at4°C and ∼70% RH for 2 mo. Pupation was initiated by raisingthe temperature to 12°C for 10–12 d.
Pupae and pre-emergent adults also were collected fromthe Sand Ridge site in mid March through early April andused for chemical analyses, behavioral studies, and fieldtests. Some adults were kept in 26×39×23 cm plastic boxesfitted with aluminum screen covers and maintained in thedark at 4°C and ∼70% RH. Under these conditions, groupsof healthy male or female sawflies (200 individuals perbox) could be kept alive for up to 30 d.
Volatile Collection Volatiles were collected from aerationsof individual and multiple male or female sawflies at theNational Center for Agricultural Utility Research, Peoria,IL, as per Cossé and Bartelt (2000) and Cossé et al. (2001).Sawflies were aerated in 150 ml glass tubes (3×30 cm), andthe resulting volatiles were collected on Super Q traps (80–100 mesh, Alltech Deerfield, IL, USA) (1×0.5 cm diam)plugged with silanized glass wool. These were placed at theinlet and outlet of each chamber. Airflow was held at50 ml min−1 via valve-controlled vacuum. Sawflies weregiven access to water in 2 ml glass vials plugged withcotton and placed inside each chamber. Volatiles werecollected from 1 d to 4 d under florescent light at 27°C anda 16 L:8D photoperiod. Outlet traps from each aerationwere rinsed with 500 μl of HPLC grade hexane (FisherScientific) and concentrated to 50 μl under a steady streamof nitrogen and stored at −70°C.
Whole Body Extracts Whole body extracts were obtainedfrom adult A. erythrocephala by soaking groups of six maleor six female sawflies in 2 ml hexane for 2 min. Theextracts then were concentrated to 200 μl under a stream ofnitrogen. A series of fractions (1.5 ml of each) of increasingpolarity were eluted on a silica gel column (0.6 cm ID×2 cm height, gravity flow), beginning with, hexane,followed by 5%, 10%, and 25% diethyl ether (redistilled)in hexane, and finally 100% ether. Fractions were concen-trated to 30 μl under a stream of nitrogen, and analyzed asdescribed below for specific chemical constituents.
Chemical Analysis Volatile collections and chromatograph-ic fractions of whole body washes were analyzed by usingcoupled gas chromatography-mass spectrometry (GC-MS).Equipment consisted of a Hewlett-Packard model 6890 gaschromatograph interfaced with a Hewlett-Packard model5973 mass selective detector. Injections were made througha split/splitless inlet, operated in splitless mode. Columnsused included DB-5 (30×0.25 mm ID with 0.25 µm film,J&W Scientific, Folsom, CA, USA), DB-1 (15×0.25 mmID with 0.10 µm film, J&W Scientific), and a 30 m EC-5
J Chem Ecol (2009) 35:1448–1460 1449

(0.25 mm ID, 0.25 µm film thickness, Alltech Associates,Deerfield, IL, USA). A typical temperature programwas 50°Cfor 1 min, increasing at 10°Cmin−1 to 300°C, and with a finalhold time of 20 min. GC-MS settings were modified asnecessary for specific analyses; however, the mass spec-trometer scanning range was typically 40 amu to 550 amu(EI, 70 eV), except in analysis of dimethyl disulfide (DMDS)derivatives where the maximum mass was increased to650 amu. The 1995 version of the Wiley mass spectrallibrary was available on the data system (Wiley 1995).
Dimethyl disulfide (DMDS) derivatives of the unsatu-rated cuticular hydrocarbons were prepared and analyzed todetermine double bond locations. The hexane fractions ofthe male and female whole body washes were prepared andanalyzed according to the method in Carlson et al. (1989).Synthetic standards for (Z,Z)-1,9,15-pentacosotriene, (Z)-9,15-pentacosodiene, (Z)-9-pentacososene, and 6-14-pentadecadienal, were analyzed likewise to further verifychemical structures. GC-MS of DMDS derivatives wascarried out on a thin film (0.1 µm) DB-1 column. GC inletand transfer lines were set to 230°C and mass scan upperlimit was set to 650 amu.
Electrophysiology Aerations and polar fractions of wholebody extracts were analyzed by gas chromatographycoupled with electroantennographic detection (GC-EAD)as per Cossé and Bartelt (2000). EAD recordings weremade by severing the head of an adult sawfly at the cervix.The head was placed neck down into a vertical well (3×3 mm) filled with physiological saline located 2 mm fromthe end of a Plexiglass stage. Antennae were extendedhorizontally along two shallow channels that ran from thewell to the end of the stage. Antennae extended beyond theedge of the stage. A ring of dental clay (1 mm thickness)was placed around the well and covered with a Plexiglassplate to reduce evaporation. The plate was secured with ascrew to ensure a water-tight seal. Distal ends of theantennae were severed at the inter-segmental membranenear the 20th antennal segment. EAD connections weremade by inserting the distal ends of each antenna into glasscapillary silver microelectrodes filled with physiologicalsaline. The antenna with the recording electrode waspositioned directly in the effluent stream coming from theGC, and the antennae with the grounding electrode waspositioned out of the effluent stream. Compounds fromeither males or females that elicited antennal activity wereidentified subsequently based on comparisons to retentiontimes and mass spectra of authentic standards, or to libraryspectra (as in the case of common well characterizedcompounds such as nonanal and decanal).
Synthesis 6,14-Pentadecadienal (1), 1,9,15-pentacosatriene(2), and 1,9-pentacosadiene (3) were synthesized as standards
using Wittig condensations as the key step (Fig. 1). Reactionswere monitored by GC-MS. Attempts were not made tooptimize yields. The reagents were obtained from AldrichChemical Co., Milwaukee, WI, USA, and solvents were ofHPLC grade.
Five Wittig salts were prepared by refluxing the corre-sponding alkyl halides with triphenylphosphine in acetonitrile(Sonnet 1974). Three of these (4, 5, and 6) had simple alkylgroups of 1, 10, and 16 carbons, respectively, and two hadfunctionalized alkyl groups (9 and 12). To make 9, 6-bromo-1-hexanol (7) was converted to the tetrahydropyranyl (THP)ether (8) (Miyashita et al. 1977), which was then convertedto the salt. To make 12, 1,6-dibromohexane (10) was firstalkylated with allylmagnesium bromide under catalysis bydilithium tetrachlorocuprate as per Johnson et al. (1994); theGrignard reaction gave only monoalkylation. The resulting9-bromo-1-nonene (11) was then converted to the phospho-nium salt. Two aldehyde intermediates also were prepared:Octanedial (14) was made by oxidizing 1,8-octanediol (13)with pyridinium dichromate (PDC) (Corey and Schmidt1979), and 6,6-dimethoxyhexanal (16) was made fromcyclohexene (15) by ozonolysis, using the procedure ofClaus and Schreiber (1986).
To prepare aldehyde 1, a mixed Wittig reagent was madefrom salts 4 and 9 in tetrahydrofuran (THF) by treatmentwith butyllithium, and then dialdehyde 14 was added. Thereaction was run essentially as per Sonnet (1974), but nodimethylsulfoxide (DMSO) was added (which Sonnet1974, used to increase Z selectivity). Products with severalchain lengths were created, but these differed widely in GCretention, and the THP ether of 6,14-pentadecadien-1-ol(17) was recognized readily from its mass spectrum[diagnostic ions, 308 (M), 222 (M-86), and 85 (intense inTHP ethers)]. Both the 6Z and 6E isomers were formed, but6Z isomer was more abundant and eluted earlier by GC.After purification on silica gel (elution with 5% ether inhexane), the THP ether 17 was converted to the free alcohol18 (Miyashita et al. 1977), and the alcohol was thenoxidized with PDC to aldehyde 1.
When it became clear that 1 was identical to the keycompound from female PFW, the synthesis was repeated bya more efficient route (Fig. 1, bottom). A Wittig reactioninvolving salt 12 and acetal aldehyde 16 (Sonnet 1974)gave just one product chain length, and predominantly theZ isomer (82%). After purification of acetal 19 on silica gel(elution with 10% ether in hexane) and by Kugelrohrdistillation, 19 was readily hydrolyzed to aldehyde 1(Ellison et al. 1975). The final product (140 mg, 70% ofthe theoretical yield) was diluted with hexane and stored at−20°C until needed for field tests.
Standard hydrocarbon triene 2 was prepared in one stepby a Wittig reaction involving the decyl salt (5) andaldehyde 1. Standard hydrocarbon diene 3 was made with a
1450 J Chem Ecol (2009) 35:1448–1460

mixed Wittig reaction involving the methyl and hexadecylsalts (4 and 6) and dialdehyde 14. In both reactions, the Zisomers predominated and eluted slightly earlier than the Eisomers on the DB-1 GC column. The hydrocarbons werepurified by silica gel chromatography (elution with hexane).
Weathering Experiment To determine if (Z,Z)-1-9,15-pen-tacosotriene could in fact oxidize to produce (Z)-6,14-pentadecadienal in the presence of air and ultraviolet light,a 5 μg sample of (Z,Z)-1,9,15-pentacosotriene in 100 μl ofhexane was applied to the inside wall of a quartz cuvette(Beckman, CA, USA) and the solvent allowed to evaporate,leaving the hydrocarbon as a thin oily film. The cuvette wascapped and placed inside a Suntest CPS solar simulator(Atlas, Gainesville, FL, USA), set up as described inMcGuireet al. (2000), and exposed to simulated solar radiation for
4.5 h. The cuvette then was rinsed with 500 μl of hexane,and the solution was concentrated to 100 μl and analyzed byusing GC-MS. Relative peak area of (Z)-6,14-pentadecadie-nal to (Z,Z)-1-9,15-pentacosotriene was determined.
Field Experiments Field testing for male A. erythrocephalaresponse to synthetic and natural chemicals was carried outin the spring of 2002 and 2003 at two sites located incentral Illinois and upstate New York. Both sites containpersistent outbreaks of A. erythrocephala and are remark-ably similar in geology, topography, and host speciescomposition, characterized by mixed and monoculturestands of white, red, and Scots pine.
The Illinois site is located in the Sand Ridge State ForestReserve in Mason County, 50 km southwest of Peoria,Illinois. The reserve encompasses 2,913 ha, made up of
RX
HO(CH2)8OH
Br(CH2)6Br
Br(CH2)6OH
CH3(CH2)8CH
Ph3P+R X-
CH3(CH2)14CH
(MeO)2CH(CH2)4CHO
(CH3O)2CH(CH2)4CH
Br(CH2)6OTHP
CH2Br(CH2)7HC
OHC(CH2)6CHO
THPO(CH2)5CH
CH(CH2)4CH
CH(CH2)6CH
OHC(CH2)4CH
CH(CH2)6CH
CH(CH2)6CH
CH2
CH(CH2)6CH
CH2
CH(CH2)6CH
CH2
CH2
Ph3P+(CH2)6OTHP Br-
CH2
CH2 Br-Ph3P+(CH2)7HC
9
12
7 8
10 11
13 14
1516
4 + 9 + 14
4: R = CH3, X = I5: R = (CH2)9CH3, X =Br6: R = (CH2)15CH3, X = Br
4 + 6 + 14
1
3
1 + 5
2
12 + 16
Wittig salts
Aldehyde intermediates
Standards from Wittig condensations
Alternative synthesis of 1
(+ other Wittig products)
(6E and 6Z isomers)
(E and Z isomers, + other Wittig products)
(E and Z isomers, + other Wittig products)
1 6 14
1915
19
15
25
25
a
b a
ac
d
e
f
g d
f
f
h
17
191
f(82% Z, 18% E)
HO(CH2)5CH CH(CH2)6CH CH218
Fig. 1 Synthesis of 6,14-penta-decadienal (1), 1,9,15-pentaco-satriene (2), and 1,9-pentacosadiene (3). Reactionconditions: a Ph3P, CH3CN,reflux, 10 h; b 2H-3,4-dihydro-pyran, pyridinium p-toluenesul-fonate (PPTS), CH2Cl2, rt, 4 h;c allylmagnesium bromide,Li2CuCl4, ether, 0°C, 3 h; dpyridinium dichromate,CH2Cl2, rt, 4 h; (e) (i) O3,MeOH, CH2Cl2, −78°C, untilexcess O3 present, purge withN2, (ii) TsOH, −78°C to rt,1.5 h, (iii) NaHCO3, rt, 15 min,(iv) Me2S, rt, 12 h; (f) add BuLito salt in THF, then addaldehyde, 0°C, 1 h; (g) PPTS,EtOH, 55°C, 2.5 h; (h) 1:2 50%aqueous CF3COOH:CHCl3,0°C, 6.5 h
J Chem Ecol (2009) 35:1448–1460 1451

1,584 ha of mixed native oak (Quercus sp.) and hickory(Carya sp.) forests and 321 ha of fields and sand prairie.The remaining 1,008 ha consists of monoculture and mixedstands of white pine, red pine, and Scots pine. Althoughlocal conservation personnel noted feeding damage on pinespecies at this site for several years (personal communica-tion), the specific defoliator responsible for the damage wasnot identified until the fall of 2001. This site continues to beinfested as of this writing.
The New York field site is located 16 km southeast ofPotsdam, New York, at High Flats in Saint LawrenceCounty. Species composition, geology, and topography ofthe area are described by Asaro and Allen (1999). Anoutbreak of A. erythrocephala has persisted at this site sincethe late 1980’s and at its peak in the late 1990’s coveredmore than 200,000 ha of mixed forest throughout St.Lawrence and Franklin Counties, New York (Asaro andAllen 1999, 2001). This area continues to be infested,though densities of A. erythrocephala appear to havedeclined in recent years.
Illinois Field Experiments 2002 The first field test of (Z)-6,14-pentadecadienal was carried out at the Illinois sitefrom April 17 through April 20, 2002. Treatments includedindividual rubber septa infused with 50 μg of (Z)-6,14-pentadecadienal, 1 mg of nonanal, or 1 mg of decanal,whole body extracts of virgin male or female sawflies, andblank traps which served as controls.
Treatments containing nonanal and decanal were includ-ed in the 2002 field tests because these compounds elicitstrong antennal activity in both sexes and are also found inaerations and extracts from both sexes. Furthermore, wehave observed that decanal can elicit characteristic court-ship behaviors in males that includes abdominal flexing andsubstrate vibration.
Chemical treatments were prepared 24 h prior to fieldtesting by applying 50 μg of (Z)-6,14-pentadecadienal,1 mg of nonanal, or 1 mg decanal each to individual rubbersepta (Fisher Scientific, Pittsburgh, PA, USA). Treatedsepta were stored at −20°C and transported to the field sitein Styrofoam coolers containing dry ice. At the field site,treated septa were attached to metal paperclips and hunginside wing traps (Trécé model-1C Great Lakes IPM, Inc.,Vestaburg, MI, USA) in single-, double-, or four-compoundcombinations.
Whole-body extracts of unmated sawflies used in theIllinois field experiments were prepared by collecting pre-emergent adults from the field and extracting groups ofeither 400 male or 400 female sawflies in 300 ml of hexanefor 20 min at room temperature. This method producedapproximately 200 ml of extract. Twenty-ml aliquots ofextract (roughly 40 insect equivalents) then were pouredinto plastic Petri dishes (35×10 mm) and allowed to
evaporate at room temperature for 1 h, leaving behind anoily residue inside the dish. Petri dishes with residue werecapped and sealed with Teflon tape and stored at −20°Cbefore being transported to the field 12 h later in coolerscontaining dry ice. At the field site, dishes were uncappedand placed in the bottom center panel of individual wingtraps.
Treatments used in all field experiments were set outfollowing a randomized design with ten replicates pertreatment. Treatments were placed inside wing-traps andsecured to wooden stakes 30 cm above ground. At the endof each day, the bottom panel of each trap was removed, andsawflies were tallied per trap. Total trap data from the entirefield season were transformed using sqrt (y+1) to meetassumptions of homoscedasticity (Bartlet’s test, a=0.05)before performing one-way analysis of variance (ANOVA).Post hoc comparisons of the mean number of males caughtamong treatments were conducted with Tukey’s honestsignificant difference test (Tukey HSD) (α=0.05) usingStatistica version 6.0 (StatSoft Inc. Tulsa, OK, USA 2001).
New York Field Experiments 2002 A second field test wasconducted at the New York site from May 21 through May24, 2002. The goal of the second field test was to verify thatthe synthetic pheromone was indeed active in a differentpopulation of A. erythrocephala. Treatments at the NewYork field site included 500 μg and 50 μg doses of (Z)-6,14-pentadecadienal, whole body extracts, live virgin maleand female sawflies, and blank traps. Because nonanal anddecanal did not appear to influence male trap catches in the2002 Illinois field experiment, they were excluded from theNew York field experiments.
Whole-body extracts from virgin male or female sawflies(roughly 40 insect equivalents), were prepared in Illinois, aspreviously described, 72 h prior to conducting the NewYork field tests. Bulk extracts were transported to the NewYork site in sealed 500 liter Erlenmeyer flasks fitted with arubber stopper sealed with Teflon tape and placed inindividual Styrofoam coolers surround by dry ice. Petridishes containing extracted residue were prepared indoorsnear the New York field site as described above for theIllinois experiments.
Live virgin male and female sawflies were obtained onsite by digging in the ground for pre-emergent adults. Maleand female adults also were collected on site and kept inindividual translucent containers for 8–18 h prior to placingthem in metal screen cylinders (3×5 cm), sealed with corkstoppers. Screen cylinders containing sawflies were hung inthe center of each trap with a metal paperclip. Sawflieswere replaced every morning. Because data did notadequately meet assumptions of normality or heterosceda-sity, differences were analyzed via nonparametric Kruskal-Wallis analysis of variance. Post-hoc comparisons were
1452 J Chem Ecol (2009) 35:1448–1460

conducted via multiple comparisons of mean ranks for allpairs of groups (Siegel and Castellan 1988) via Statisticaversion 6.0 (StatSoft Inc., Tulsa, OK, USA 2001).
Illinois Field Experiments 2003 A third field test wascarried out at the Illinois site in 2003 from April 24–27.Treatments included 1.5 μg, 5 μg, 15 μg, 50 μg, 150 μg,and 500 μg doses of synthetic (Z)-6,14-pentadecadienal,whole body extracts of virgin male or female sawflies, andlive virgin male or female sawflies held in metal cylindricalcages as describe above. Blank wing traps served ascontrols.
Whole body extracts used in the 2003 Illinois fieldexperiment were prepared as described above for the 2002field experiments 6 h prior to placing them in the field. In2003, Petri dishes containing whole body extracts wereuncovered and placed in direct sunlight (∼21°C airtemperature) for 1 h prior to placing them in individualtraps. The purpose of exposing whole body extracts todirect sunlight was to determine if exposure to sunlightwould enhance the attractiveness of the extract relative tothe previous year’s results.
Adult male and female sawflies used in the 2003 fieldexperiments were collected as eonymphs and reared atIllinois State University. Adults were kept in translucent26×39×23 cm plastic containers with wire-screen covers.Containers with fresh sawflies were transported to the fieldevery morning and placed in direct sunlight at approxi-mately 20°C for 1 h prior to placing individual sawflies inscreen cages hung centrally inside each trap. As with thewhole body extracts, the purpose of exposing live extractsto direct sunlight was to assess whether exposing livefemale insects would enhance attractiveness. Cages werereplaced with fresh sawflies every morning for the durationof the experiment. Total trap data from the entire fieldseason were transformed using log (y+1) to meet assump-tions of homoscedasticity (Bartlet’s test, a=0.05) beforeperforming one-way analysis of variance (ANOVA). Posthoc comparisons were conducted as described above.
Results
Chemical Analysis GC-MS analysis of aerations and wholebody extracts obtained from both sexes of Acantholydaerythrocephala primarily contained saturated and unsatu-rated straight chain hydrocarbons, many with a double bondlocated at the Δ9 position. Qualitative and quantitativedifferences between sexes included a 25-carbon triene (MW346), exclusive to females, and a 25-carbon diene (MW348) that was abundant in females, but also occurred intrace amounts in aerations and extracts obtained frommales. A 25-carbon alkene (MW 350) and pentacosane
(MW 352) were present in both male and female aerationsand extracts (Table 1).
The putative pentacosane was verified based on compar-isons to library spectra, GC retention time, and mass spectraof authentic standard (Aldrich Chemical Co., Milwaukee,WI, USA). Preliminary identifications of the -ene, -dien,and -trien, were based on GC-MS analysis of their DMDSderivatives.
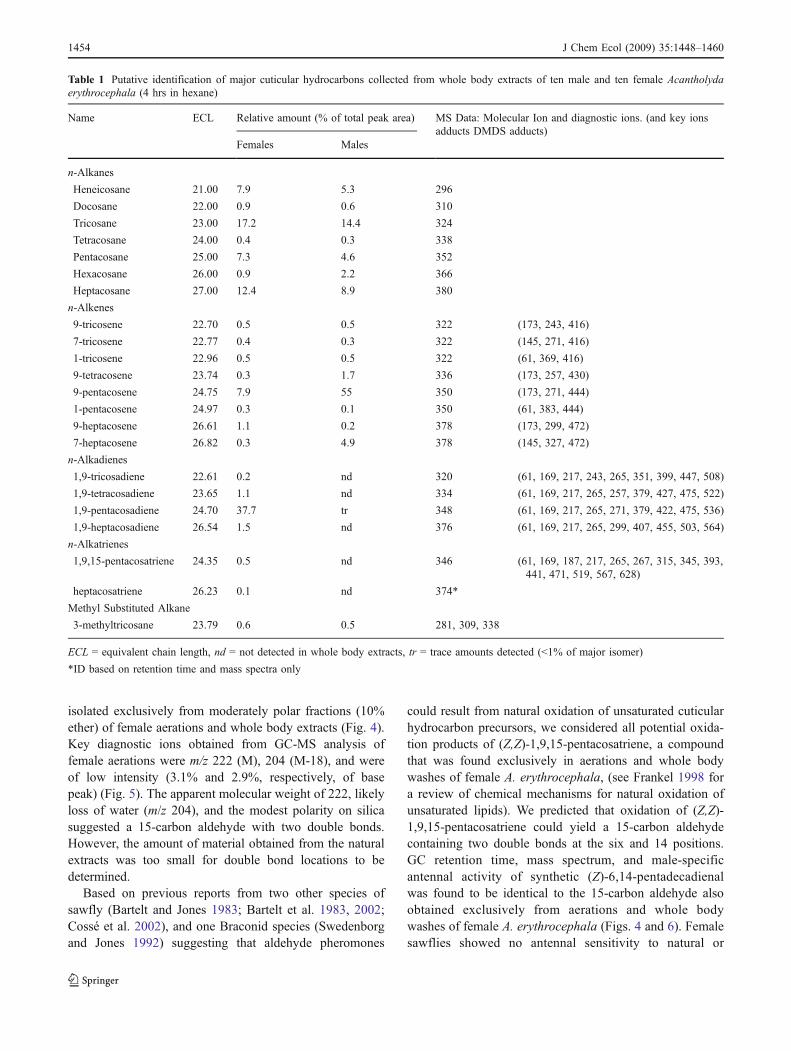
DMDS adducts of pentacosene (Fig. 2a) gave amolecular ion at m/z 444 (350+94), consistent with a 25-carbon chain containing two thiomethyl (CH3S) groups.Diagnostic fragments A+ (m/z 173) and B+ (m/z 271)indicated a double bond at the 9 position (9-pentacosene).
DMDS adducts for pentacosadiene (Fig. 2b) produced amolecular ion at m/z 536 (348 + 188). This was consistentwith a 25-carbon chain containing four CH3S groups. Keydiagnostic ions B+ (m/z 265, 217,169) and D+ (m/z 271)also suggested a double bond at the 9 position. Ionfragments A+ (m/z 61) and C+ (m/z 441, 393, 345) indicatedthe presence of a terminal double bond, resulting in 1,9-pentacosadiene.
DMDS adducts for pentacosatriene (Fig. 2c) produced amolecular ion at m/z 628 (346 + 282), consistent with a 25-carbon chain containing six CH3S groups. Ion fragmentsA+ (m/z 61) and D+ (m/z 567, 519, 471), again indicated aterminal double bond. Fragments B+ (m/z 265, 217, 169) andE+ (383, 315, 267) indicated a double bond at the 9 position.Fragments C+ (441, 393, 345) and F+ (187) suggested a thirddouble bond at the 15 position (1,9,15-pentacosatriene).
Synthetic hydrocarbons corroborated these identifica-tions and helped clarify double bond configurations.Female-derived 1,9-pentacosadiene and 1,9,15-pentacosa-triene showed GC retention times and mass spectraidentical to the earliest-eluting (Z) synthetic isomersindicating that these hydrocarbons existed in the Zconfiguration (i.e., (Z)-1,9-pentacosadiene and (Z,Z)-1,9,15-pentacosatriene). DMDS derivatives of synthetic1,9-pentacosadiene and 1,9,15-pentacosatriene producedmass spectra and GC retention times identical to DMDSderivatives of the natural 1,9-pentacosadiene and 1,9,15-pentacosatriene.
GC-MS analysis of natural and DMDS derivatizedwhole body extracts from ten male and ten female sawfliesin hexane revealed that female sawflies contained a numberof dienes with double bonds at the Δ1 and Δ9 positions.These dienes were either nonexistent in male extracts orpresent only in trace amounts (Table 1).
Electrophysiology Electrophysiological analysis of maleand female aerations showed antennal sensitivity in bothsexes to a series of aldehydes including nonanal, decanal,and possibly 2-undecanal. Male-specific antennal responseswere also recorded for a single unknown compound (Fig. 3)
J Chem Ecol (2009) 35:1448–1460 1453
isolated exclusively from moderately polar fractions (10%ether) of female aerations and whole body extracts (Fig. 4).Key diagnostic ions obtained from GC-MS analysis offemale aerations were m/z 222 (M), 204 (M-18), and wereof low intensity (3.1% and 2.9%, respectively, of basepeak) (Fig. 5). The apparent molecular weight of 222, likelyloss of water (m/z 204), and the modest polarity on silicasuggested a 15-carbon aldehyde with two double bonds.However, the amount of material obtained from the naturalextracts was too small for double bond locations to bedetermined.
Based on previous reports from two other species ofsawfly (Bartelt and Jones 1983; Bartelt et al. 1983, 2002;Cossé et al. 2002), and one Braconid species (Swedenborgand Jones 1992) suggesting that aldehyde pheromones
could result from natural oxidation of unsaturated cuticularhydrocarbon precursors, we considered all potential oxida-tion products of (Z,Z)-1,9,15-pentacosatriene, a compoundthat was found exclusively in aerations and whole bodywashes of female A. erythrocephala, (see Frankel 1998 fora review of chemical mechanisms for natural oxidation ofunsaturated lipids). We predicted that oxidation of (Z,Z)-1,9,15-pentacosatriene could yield a 15-carbon aldehydecontaining two double bonds at the six and 14 positions.GC retention time, mass spectrum, and male-specificantennal activity of synthetic (Z)-6,14-pentadecadienalwas found to be identical to the 15-carbon aldehyde alsoobtained exclusively from aerations and whole bodywashes of female A. erythrocephala (Figs. 4 and 6). Femalesawflies showed no antennal sensitivity to natural or
Table 1 Putative identification of major cuticular hydrocarbons collected from whole body extracts of ten male and ten female Acantholydaerythrocephala (4 hrs in hexane)
Name ECL Relative amount (% of total peak area) MS Data: Molecular Ion and diagnostic ions. (and key ionsadducts DMDS adducts)
Females Males
n-Alkanes
Heneicosane 21.00 7.9 5.3 296
Docosane 22.00 0.9 0.6 310
Tricosane 23.00 17.2 14.4 324
Tetracosane 24.00 0.4 0.3 338
Pentacosane 25.00 7.3 4.6 352
Hexacosane 26.00 0.9 2.2 366
Heptacosane 27.00 12.4 8.9 380
n-Alkenes
9-tricosene 22.70 0.5 0.5 322 (173, 243, 416)
7-tricosene 22.77 0.4 0.3 322 (145, 271, 416)
1-tricosene 22.96 0.5 0.5 322 (61, 369, 416)
9-tetracosene 23.74 0.3 1.7 336 (173, 257, 430)
9-pentacosene 24.75 7.9 55 350 (173, 271, 444)
1-pentacosene 24.97 0.3 0.1 350 (61, 383, 444)
9-heptacosene 26.61 1.1 0.2 378 (173, 299, 472)
7-heptacosene 26.82 0.3 4.9 378 (145, 327, 472)
n-Alkadienes
1,9-tricosadiene 22.61 0.2 nd 320 (61, 169, 217, 243, 265, 351, 399, 447, 508)
1,9-tetracosadiene 23.65 1.1 nd 334 (61, 169, 217, 265, 257, 379, 427, 475, 522)
1,9-pentacosadiene 24.70 37.7 tr 348 (61, 169, 217, 265, 271, 379, 422, 475, 536)
1,9-heptacosadiene 26.54 1.5 nd 376 (61, 169, 217, 265, 299, 407, 455, 503, 564)
n-Alkatrienes
1,9,15-pentacosatriene 24.35 0.5 nd 346 (61, 169, 187, 217, 265, 267, 315, 345, 393,441, 471, 519, 567, 628)
heptacosatriene 26.23 0.1 nd 374*
Methyl Substituted Alkane
3-methyltricosane 23.79 0.6 0.5 281, 309, 338
ECL = equivalent chain length, nd = not detected in whole body extracts, tr = trace amounts detected (<1% of major isomer)
*ID based on retention time and mass spectra only
1454 J Chem Ecol (2009) 35:1448–1460
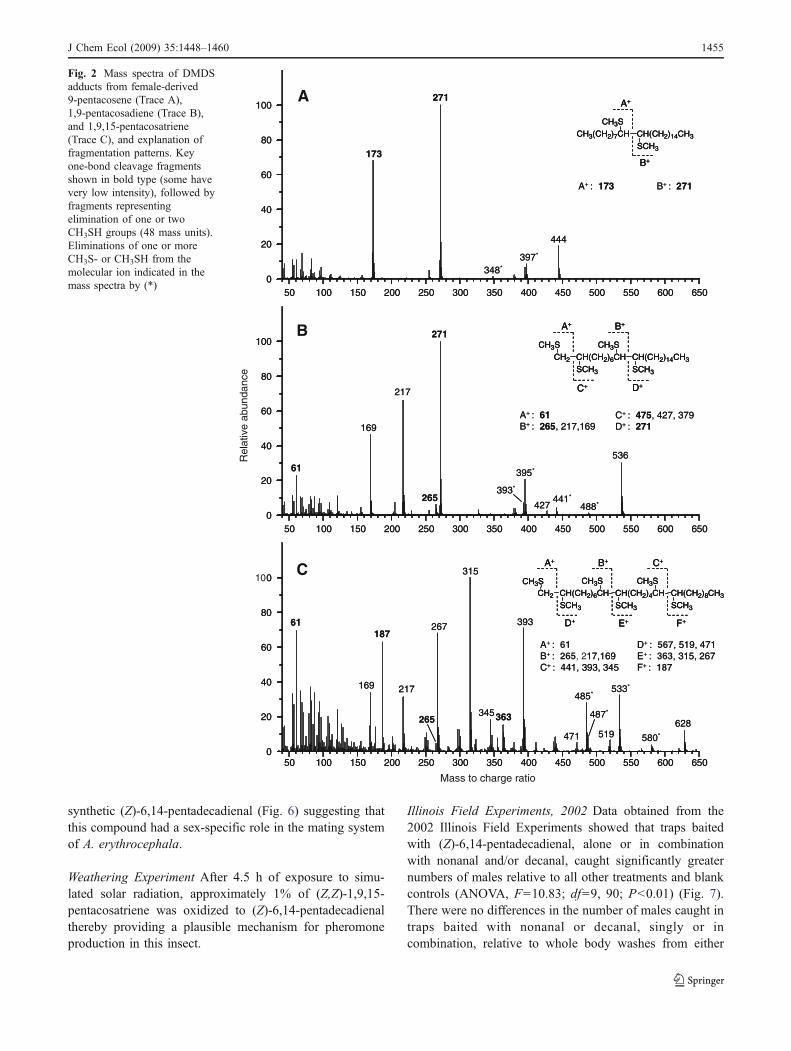
synthetic (Z)-6,14-pentadecadienal (Fig. 6) suggesting thatthis compound had a sex-specific role in the mating systemof A. erythrocephala.
Weathering Experiment After 4.5 h of exposure to simu-lated solar radiation, approximately 1% of (Z,Z)-1,9,15-pentacosatriene was oxidized to (Z)-6,14-pentadecadienalthereby providing a plausible mechanism for pheromoneproduction in this insect.
Illinois Field Experiments, 2002 Data obtained from the2002 Illinois Field Experiments showed that traps baitedwith (Z)-6,14-pentadecadienal, alone or in combinationwith nonanal and/or decanal, caught significantly greaternumbers of males relative to all other treatments and blankcontrols (ANOVA, F=10.83; df=9, 90; P<0.01) (Fig. 7).There were no differences in the number of males caught intraps baited with nonanal or decanal, singly or incombination, relative to whole body washes from either
50 100 150 200 250 300 350 400 450 500 550 600 6500
20
40
60
80
100
50 100 150 200 250 300 350 400 450 500 550 600 6500
20
40
60
80
100
50 100 150 200 250 300 350 400 450 500 550 600 6500
20
40
60
80
100
393
CH(CH2)6CHCH3S
CH(CH2)4CH
SCH3
CH2
SCH3
CH3SCH(CH2)8CH3
SCH3
CH3S
A+ B+ C+
D+ E+ F+
628
533*
580*
485*
519
363
315
217
267187
169
61
345
471
Mass to charge ratio
Rel
ativ
e ab
unda
nce
B
A
C
536
CH(CH2)6CHCH3S
CH(CH2)14CH3
SCH3
CH2
SCH3
CH3S
A+ B+
C+ D+
395*
265
271
217
169
61
427441*
488*
393*
487*
CH3(CH2)7CHCH3S
CH(CH2)14CH3
SCH3
A+
B+173
271
444
397*
348*
265
A+ : 61B+ : 265, 217,169C+ : 441, 393, 345
D+ : 567, 519, 471E+ : 363, 315, 267F+ : 187
A+ : 61B+ : 265, 217,169
C+ : 475, 427, 379D+ : 271
A+ : 173 B+ : 271
50 100 150 200 250 300 350 400 450 500 550 600 6500
20
40
60
80
100
50 100 150 200 250 300 350 400 450 500 550 600 6500
20
40
60
80
100
50 100 150 200 250 300 350 400 450 500 550 600 6500
20
40
60
80
0
393
CH(CH2)6CHCH3S
CH(CH2)4CH
SCH3
CH2
SCH3
CH3SCH(CH2)8CH3
SCH3
CH3S
A+ B+ C+
D+ E+ F+
CH(CH2)6CHCH3S
CH(CH2)4CH
SCH3
CH2
SCH3
CH3SCH(CH2)8CH3
SCH3
CH3S
A+ B+ C+
D+ E+ F+
628
533*
580*
485*
519
363
315
217
267187
169
61
345
471
536
CH(CH2)6CHCH3S
CH(CH2)14CH3
SCH3
CH2
SCH3
CH3S
A+ B+
C+ D+
CH(CH2)6CHCH3S
CH(CH2)14CH3
SCH3
CH2
SCH3
CH3S
A+ B+
C+ D+
395*
265
271
217
169
61
427441*
488*
393*
487*
CH3(CH2)7CHCH3S
CH(CH2)14CH3
SCH3
A+
B+
CH3(CH2)7CHCH3S
CH(CH2)14CH3
SCH3
A+
B+173
271
444
397*
348*
265
A+ : B+ : 17,169C+ : , 393, 345
D+ : , 519, 471E+ : , 315, 267F+ :
A+ : 61B+ : 265, 217,169
C+ : 475, 427, 379D+ : 271
A+ : 173 B+ : 271
Fig. 2 Mass spectra of DMDSadducts from female-derived9-pentacosene (Trace A),1,9-pentacosadiene (Trace B),and 1,9,15-pentacosatriene(Trace C), and explanation offragmentation patterns. Keyone-bond cleavage fragmentsshown in bold type (some havevery low intensity), followed byfragments representingelimination of one or twoCH3SH groups (48 mass units).Eliminations of one or moreCH3S- or CH3SH from themolecular ion indicated in themass spectra by (*)
J Chem Ecol (2009) 35:1448–1460 1455
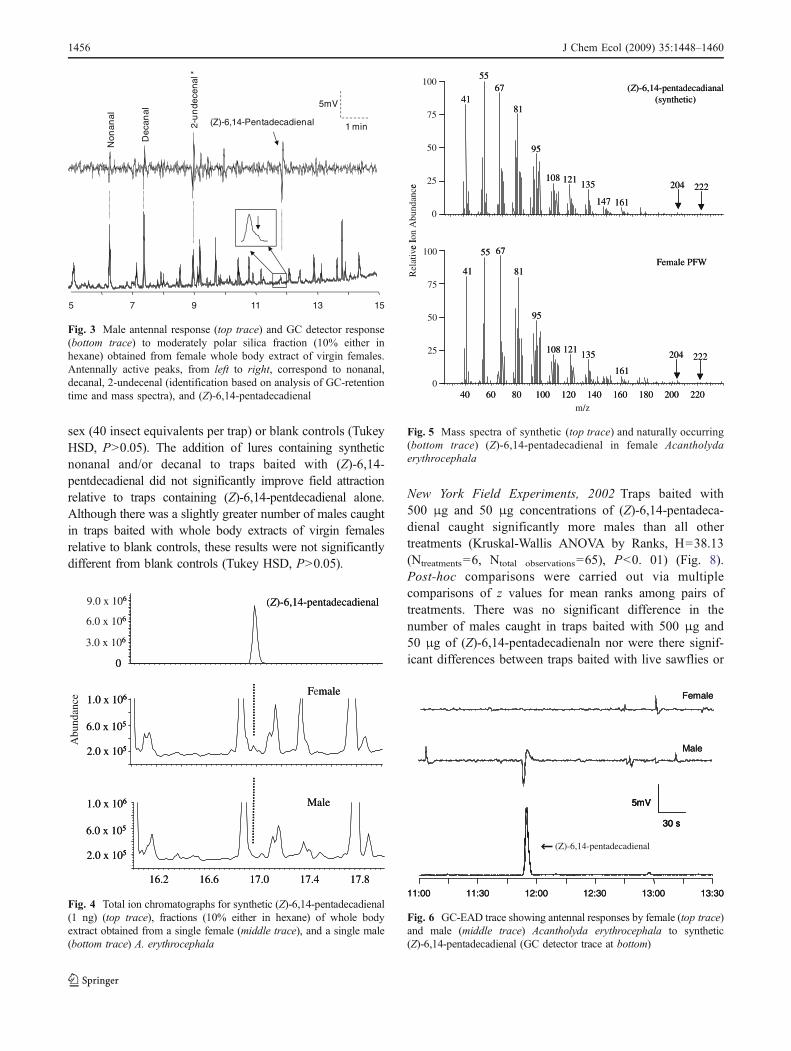
sex (40 insect equivalents per trap) or blank controls (TukeyHSD, P>0.05). The addition of lures containing syntheticnonanal and/or decanal to traps baited with (Z)-6,14-pentdecadienal did not significantly improve field attractionrelative to traps containing (Z)-6,14-pentdecadienal alone.Although there was a slightly greater number of males caughtin traps baited with whole body extracts of virgin femalesrelative to blank controls, these results were not significantlydifferent from blank controls (Tukey HSD, P>0.05).
New York Field Experiments, 2002 Traps baited with500 μg and 50 μg concentrations of (Z)-6,14-pentadeca-dienal caught significantly more males than all othertreatments (Kruskal-Wallis ANOVA by Ranks, H=38.13(Ntreatments=6, Ntotal observations=65), P<0. 01) (Fig. 8).Post-hoc comparisons were carried out via multiplecomparisons of z values for mean ranks among pairs oftreatments. There was no significant difference in thenumber of males caught in traps baited with 500 μg and50 μg of (Z)-6,14-pentadecadienaln nor were there signif-icant differences between traps baited with live sawflies or
11:00 11:30 12:00 12:30 13:00 13:30
5mV
30 s
Female
Male
11:00 11:30 12:00 12:30 13:00 13:30
5mV
30 s
5mV
30 s
Female
Male
(Z)-6,14-pentadecadienal
Fig. 6 GC-EAD trace showing antennal responses by female (top trace)and male (middle trace) Acantholyda erythrocephala to synthetic(Z)-6,14-pentadecadienal (GC detector trace at bottom)
40 60 80 100 120 140 160 180 200 220
Rel
ativ
e Io
n A
bund
ance
Female PFW6755
41 81
95
121108 135
161
204 222
(Z)-6,14-pentadecadianal(synthetic)
5567
4181
95
108 121 135
147 161
100
75
50
25
0
75
50
25
0
204 222
40 60 80 100 120 140 160 180 200 220m/z
e I
e
Female PFW6755
41 81
95
121108 135
161
204 222
(Z)-6,14-pentadecadianal(synthetic)
5567
4181
95
108 121 135
147 161
100
204 222
Fig. 5 Mass spectra of synthetic (top trace) and naturally occurring(bottom trace) (Z)-6,14-pentadecadienal in female Acantholydaerythrocephala
Female
2.0 x 105
6.0 x 105
1.0 x 106
0
3.0 x 106
6.0 x 106
9.0 x 106
16.2 16.6 17.0 17.4 17.8
Male
(Z)-6,14-pentadecadienal
2.0 x 105
6.0 x 105
1.0 x 106
F male
2.0 x 105
6.0 x 105
1.0 x 106
5
5
6
0
6
6
6
16.2 16.6 17.0 17.4 17.8
Abu
ndan
ce
Male
(Z)-6,14-pentadecadienal
2.0 x 105
6.0 x 105
1.0 x 106
5
5
6
Fig. 4 Total ion chromatographs for synthetic (Z)-6,14-pentadecadienal(1 ng) (top trace), fractions (10% either in hexane) of whole bodyextract obtained from a single female (middle trace), and a single male(bottom trace) A. erythrocephala
5 7 9 11 13 15
Non
anal
Dec
anal
2-u
nde
cena
l *
(Z)-6,14-Pentadecadienal
5mV
1 min
Fig. 3 Male antennal response (top trace) and GC detector response(bottom trace) to moderately polar silica fraction (10% either inhexane) obtained from female whole body extract of virgin females.Antennally active peaks, from left to right, correspond to nonanal,decanal, 2-undecenal (identification based on analysis of GC-retentiontime and mass spectra), and (Z)-6,14-pentadecadienal
1456 J Chem Ecol (2009) 35:1448–1460
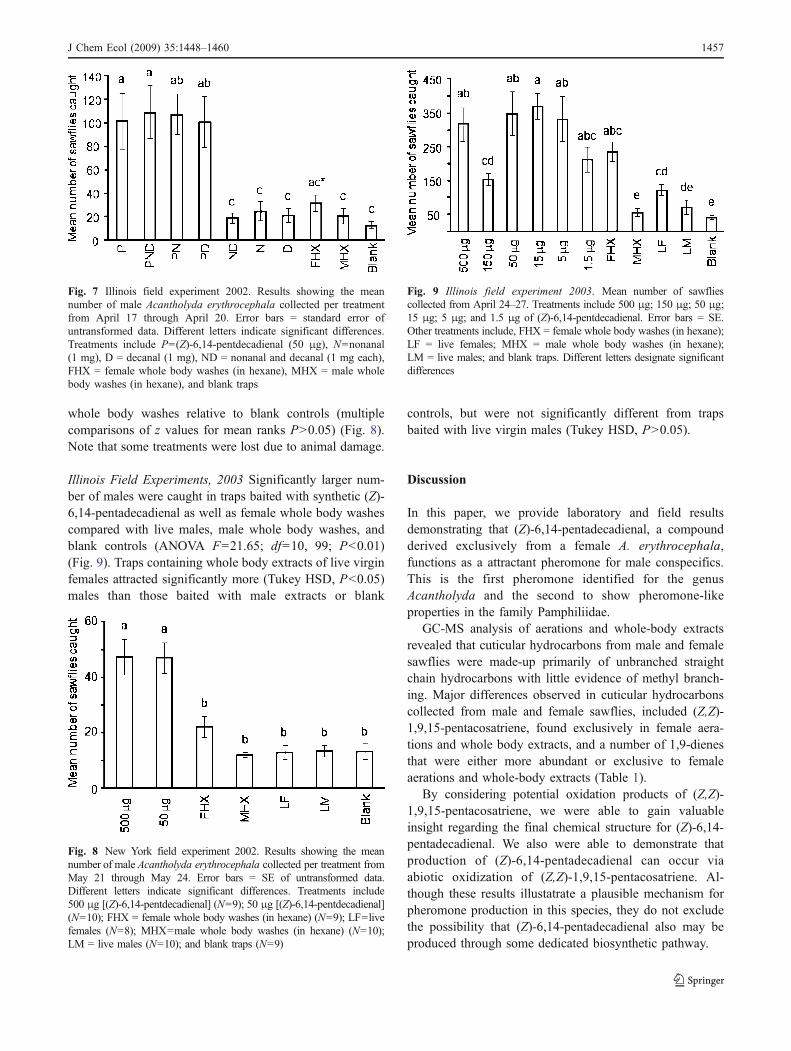
whole body washes relative to blank controls (multiplecomparisons of z values for mean ranks P>0.05) (Fig. 8).Note that some treatments were lost due to animal damage.
Illinois Field Experiments, 2003 Significantly larger num-ber of males were caught in traps baited with synthetic (Z)-6,14-pentadecadienal as well as female whole body washescompared with live males, male whole body washes, andblank controls (ANOVA F=21.65; df=10, 99; P<0.01)(Fig. 9). Traps containing whole body extracts of live virginfemales attracted significantly more (Tukey HSD, P<0.05)males than those baited with male extracts or blank
controls, but were not significantly different from trapsbaited with live virgin males (Tukey HSD, P>0.05).
Discussion
In this paper, we provide laboratory and field resultsdemonstrating that (Z)-6,14-pentadecadienal, a compoundderived exclusively from a female A. erythrocephala,functions as a attractant pheromone for male conspecifics.This is the first pheromone identified for the genusAcantholyda and the second to show pheromone-likeproperties in the family Pamphiliidae.
GC-MS analysis of aerations and whole-body extractsrevealed that cuticular hydrocarbons from male and femalesawflies were made-up primarily of unbranched straightchain hydrocarbons with little evidence of methyl branch-ing. Major differences observed in cuticular hydrocarbonscollected from male and female sawflies, included (Z,Z)-1,9,15-pentacosatriene, found exclusively in female aera-tions and whole body extracts, and a number of 1,9-dienesthat were either more abundant or exclusive to femaleaerations and whole-body extracts (Table 1).
By considering potential oxidation products of (Z,Z)-1,9,15-pentacosatriene, we were able to gain valuableinsight regarding the final chemical structure for (Z)-6,14-pentadecadienal. We also were able to demonstrate thatproduction of (Z)-6,14-pentadecadienal can occur viaabiotic oxidization of (Z,Z)-1,9,15-pentacosatriene. Al-though these results illustatrate a plausible mechanism forpheromone production in this species, they do not excludethe possibility that (Z)-6,14-pentadecadienal also may beproduced through some dedicated biosynthetic pathway.
Fig. 9 Illinois field experiment 2003. Mean number of sawfliescollected from April 24–27. Treatments include 500 μg; 150 μg; 50 μg;15 μg; 5 μg; and 1.5 μg of (Z)-6,14-pentdecadienal. Error bars = SE.Other treatments include, FHX = female whole body washes (in hexane);LF = live females; MHX = male whole body washes (in hexane);LM = live males; and blank traps. Different letters designate significantdifferences
Fig. 8 New York field experiment 2002. Results showing the meannumber of male Acantholyda erythrocephala collected per treatment fromMay 21 through May 24. Error bars = SE of untransformed data.Different letters indicate significant differences. Treatments include500 μg [(Z)-6,14-pentdecadienal] (N=9); 50 μg [(Z)-6,14-pentdecadienal](N=10); FHX = female whole body washes (in hexane) (N=9); LF=livefemales (N=8); MHX=male whole body washes (in hexane) (N=10);LM = live males (N=10); and blank traps (N=9)
Fig. 7 Illinois field experiment 2002. Results showing the meannumber of male Acantholyda erythrocephala collected per treatmentfrom April 17 through April 20. Error bars = standard error ofuntransformed data. Different letters indicate significant differences.Treatments include P=(Z)-6,14-pentdecadienal (50 μg), N=nonanal(1 mg), D = decanal (1 mg), ND = nonanal and decanal (1 mg each),FHX = female whole body washes (in hexane), MHX = male wholebody washes (in hexane), and blank traps
J Chem Ecol (2009) 35:1448–1460 1457

Field results from the Illinois and New York sites in2002 and 2003 demonstrate that (Z)-6,14-pentadecadienalfunctions as an effective male lure. It is also interesting tonote that in the 2003 Illinois field experiments, significantlymore male sawflies were captured in traps baited withwhole body extracts of virgin females relative to trapsbaited with extracts of virgin males after they were exposedto direct sunlight (Fig. 9). These observations fit well withlaboratory experiments that demonstrate that (Z)-6,14-pentadecadienal can indeed arise via abiotic oxidation offemale specific (Z,Z)-1,9,15-pentacosatriene.
Although we recorded antennal responses to nonanal anddecanal in both male and female sawflies, neither of thesecompounds was found to significantly influence maleattraction in the field. Similarly, neither compound exhibitedsynergistic or inhibitory influences on male response to (Z)-6,14-pentadecadienal. Nevertheless, these results do not ruleout a bioactive role for these compounds. Indeed, it isinteresting to note that n-decanal and 2-undecenal, respec-tively, not only stimulate courtship behavior and antennalresponses in male A. erythrocephala, but both compoundsalso are likely oxidation products of (Z,Z)-1,9,15-pentacosa-triene via either α-scission or β-scission of the C15 doublebond. Future planned research will explore the role of2-undecenal in the behavior of A. erythrocephala.
To date, approximately 22 pheromones have beenidentified among 14 families and 8,000 species (Smith1993) of sawfly. Most of these pheromones have beenreported in the Diprionidae and are made up of variousmethyl branched acetate or propionate products of diprionol(Jewett et al. 1976; Ahlgren et al. 1979; Anderbrant et al.1992, 2000, 2005; Bergström et al. 1995, 1998; Chen et al.1997; Tai et al. 1998, 2002; Wassgren et al. 2000; Ostrandet al. 2003; Hedenström et al. 2006). Acantholyda eryth-rocephala differs from diprionids in that its pheromone,(Z)-6,14-pentadecadienal, and likely pheromone precursor,(Z,Z)-1,9,15-pentacosatriene, are both simple unbranchedstraight chain hydrocarbons. Although this simple straightchain structure differs in comparison to active pheromonesof diprionind species, it is not surprising in terms of acuticular precursor or as a pheromone. Indeed, straightchain cuticular hydrocarbons, including those with a doublebond in the Δ-9 position, are quite common among insectcuticular hydrocarbons (Blomquist et al. 1993; Howard1993; Howard and Blomquist 2005; Steiner et al. 2007).Nevertheless, pheromone production by means of abioticoxidation remains a relatively little explored area of studyin insect chemical ecology. Other reports describingpheromone production by means of abiotic oxidation ininsects include the yellow-headed spruce sawfly, Pikonemaalaskensis, (P.) (Tenthredinidae) (Bartelt and Jones 1983),the parasitic wasp of the European corn borer Ostrinianubilalis Hubner, Macrocentrus grandii Goidanich (Hyme-
noptera: Braconidae) (Swedenborg and Jones 1992), and thewheat stem sawfly, Cephus cinctus Norton (Hymenoptera:Cephidae) (Bartelt et al. 2002; Cossé et al. 2002). The resultspresented here, therefore, represent only the fourth report ofpheromone production in an insect species that uses thismechanism.
With the development of a viable pheromone lure for A.erythrocephala, we now offer an additional managementtool for monitoring and possibly controlling this forest pest.
Acknowledgments We thank Robert Behle of the USDA-ARS-NCAUR in Peoria, Illinois for use of the solar simulator, and theIllinois Department of Natural Resources for access to Sand RidgeState Forest.
References
AHLGREN, G., BERGSTRÖM, G., LÖFQVIST, J., JANSSON, A., andNORIN, T. 1979. Trans-perillenal A furanoid monoterpene frompine Saw-Fly, Neodiprion sertifer (Hymenoptera: Diprionidae). J.Chem. Ecol. 5: 309–319.
ALLEN, D. C. 2000. The pine false webworm in New York. pp. 30–32in: LYONS, D. B., JONES G. C., AND SCARR, T. A. (eds.)Proceedings of a Workshop on the Pine False Webworm,Acantholyda erythrocephala (Hymenoptera: Pamphiliidae); 16February 1998; Faculty of Forestry, University of Toronto.Ontario Ministry of Natural Resources of Ontario, NaturalResources Canada, Canadian Forest Service.
ANDERBRANT, O., BENGTSSON, M., LÖFQVIST, J., and BAECKSTRÖM,P. 1992. Field response of the pine sawfly Neodiprion sertifer tocontrolled release of diprionyl acetate, diprionyl propionate andtrans-perillenal. J. Chem. Ecol. 18: 1707–1725.
ANDERBRANT, O., LÖFQVIST, J., HÖGBERG, H.-E., HEDENSTRÖM, E.,BALDASSARI, N., BARONIO, P., KOLMAKOVA, G., LYONS, B.,NAITO, T., ODINOKOV, V., SIMAND, J., SUPATASHVILI, A., TAI, A.,and TOURIANOV, R. 2000. Geographic variation in the fieldresponse of male European pine sawflies, Neodiprion sertifer, todifferent pheromone stereoisomers and esters. Entomol. Exp.Appl. 95: 229–239.
ANDERBRANT, O., ÖSTRAND, F., BERGSTRÖM, G., WASSGREN, A.-B.,AUGER-ROZENBERG, M.-A., GERI, C., HEDENSTRÖM, E., HÖGBERG,H. E., HERZ, A., and HEITLAND, W. 2005. Release of sexpheromone and its precursor in the pine sawfly Diprion pini(Hym., Diprionidae). Chemoecology. 15: 147–151.
ASARO, C., and ALLEN, D. C. 1999. Biology of pine false webworm(Hymenoptera: Pamphiliidae) during an outbreak. Canad. Ento-mologist. 131: 729–742
ASARO, C., and ALLEN, D. C. 2001. History of a pine false webworm(Hymenoptera: Pamphiliidae) outbreak in northern New York.Can. J. For. Res. 31: 181–185
BAKER, R., LONGHURST, C., SELWOOD, D., and BILLANY, D. 1983.Ortho-aminoacetophenone: a component of the sex pheromonesystem of the web-spinning larch sawfly, Cephalcia lariciphilawachtl. Experientia. 39: 993–994.
BARTELT, R. J., and JONES, R. L. 1983. (Z)-10-Nonadecenal: APheromonally Active Air Oxidation Product of (Z,Z) -9, 19Dienes in Yellowheaded Spruce Sawfly. J. Chem. Ecol. 9, 1333–1341.
BARTELT, R. J., JONES, R. L., and KULMAN, H. M. 1982a. Evidencefor a multicomponent sex pheromone in the yellowheaded sprucesawfly. J. Chem. Ecol. 8, 83–94.
1458 J Chem Ecol (2009) 35:1448–1460

BARTELT, R. J., JONES, R. L., and KULMAN, H. M. 1982b.Hydrocarbon components of the yellowheaded spruce saw flysex pheromone: A Series of (Z, Z)-9, 19 Dienes. J. Chem. Ecol.8, 95–114.
BARTELT, R. J., JONES, R. L., and KRICK, T. P. 1983. (Z)-5-Tetradecen-1-ol: A Secondary Pheromone of the YellowheadedSpruce Sawfly, and Its Relationship to (Z)-10-Nonadecenal. J.Chem. Ecol. 9, 1343–1352.
BARTELT, R.J., COSSÉ, A. A., PETROSKI, R.J., and WEAVER, D.K.2002. Cuticular hydrocarbons and novel alkenediol diacetatesfrom wheat stem sawfly (Cephus cinctus): natural oxidation topheromone components. J. Chem. Ecol. 28: 385–405.
BERGSTRÖM, G., WASSGREN, A.-B., ANDERBRANT, O., FAGERHAG, J.,EDLUND, H., HEDENSTRÖM, E., HÖGBERG, H. -E., GERI, C.,AUGER, M. A., VARAMA, M., HANSSON, B. S., and LÖFQVIST, J.1995. Sex pheromone of the pine sawfly Diprion pini (Hyme-noptera: Diprionidae): chemical identification, synthesis andbiological activity. Experientia. 51: 370–380.
BERGSTRÖM, G., WASSGREN, A.-B., ANDERBRANT, O., OCHIENG, S.A., OSTRAND, F., HANSSON, B. S., HEDENSTRÖM, E., andHÖGBERG, H. -E. 1998. The sex pheromone of the pine sawflyMicrodiprion pallipes (Hymenoptera: Diprionidae). Naturwissen-schaften. 85: 244–248.
BLOMQUIST, G. J., TILLMANWALL, J. A., GUO L, QUILICI, D. R., GU
PD, and SCHAL C. 1993. Hydrocarbons and hydrocarbon derivedsex-pheromones in insects – Biochemicstry and EndocrineRegulation. pp. 317–351 in Insect Lipids: Chemistry, Biochem-istry and Biology D. W. Stanley-Samuelson and D. R. Nelson,eds. University of Nebraska Press, Lincoln.
BORDEN J. H., BILLANY, D. J. BRADSHAW, J. W. S., EDWARDS, R. B.,BAKER, R., and EVANS, D.A. 1978. Pheromone response andsexual behavior of Cephacia lariciphila Wachtl (Hymenoptera:Pamphiliidae). Ecol. Entom. 3: 13-23
CARLSON, D. A., ROAN, C.-S., YOST, R. A., and HECTOR, J. 1989.Dimethyl disulfide derivatives of long chain alkenes, alkadienes,and alkatrienes for gas chromatography/mass spectrometry. Anal.Chem. 61: 1564–1571.
CHEN, G.F., ZHANG, Q. H., LI, Z. Y., MIAO, Z. W., WANG, L. Z., andANDERBRANT, O. 1997. A preliminary study on the sexpheromone of Diprion jingyuanensis Xiao et Zhang. Chin. J.Biol. Control. 13: 61–64.
CLAUS, R. E., and SCHREIBER, S. L. 1986. Ozonolytic cleavage ofcyclohexene to terminally differentiated products: Methyl 6-oxohexanoate, 6,6-dimethoxyhexanal, methyl 6,6-dimethoxyhex-anoate. Org. Synth. 64: 150–156.
COREY, E. J., and SCHMIDT, G. 1979. Useful procedures for theoxidation of alcohols involving pyridium dichromate in aproticmedia. Tetrahed. Lett. 5: 399–402
COSSÉ A. A, and BARTELT R. J. 2000. Male-produced aggregationpheromone of Colopterus truncatus: Structure, electrophysiolog-ical, and behavioral activity. J. Chem. Ecol. 26: 1735–1748.
COSSÉ A. A., BARTELT R. J., JAMES, D. G, and PETROSKI R.J. 2001.Identification of a female-specific, antennally active volatile com-pound of the currant stem girdler. J. Chem. Ecol. 27: 1841–1853.
COSSÉ, A. A., BARTELT, R. J., WEAVER, D. K., and ZILKOWSKI, B. W.2002. Pheromone components of the wheat stem sawfly:identification, electrophysiology, and field bioassay. J. Chem.Ecol. 28: 407–423
ELLISON, R. A., LUKENBACH, E. R., and CHIU, C.-W. 1975. Cyclo-penenone synthesis via aldol condensation: Synthesis of a keyprostaglandin intermediate. Tetrahedron Lett. 1975: 499–502.
FRANKEL, E. N. 1998. Lipid Oxidation. Oily Press, Dundee, Scotland.HEDENSTRÖM, E., EDLUND, H., WASSGREN, A.-B., BERGSTRÖM,
G., ANDERBRANT, O., ÖSTRAND, F., SIERPINSKI, A., AUGER-ROZENBERG, M. A., HERZ, A., HEITLAND, W., and VARAMA, M.2006. Sex pheromone of the pine sawfly, Gilpinia pallida:
chemical identification, synthesis, and biological activity. J.Chem. Ecol. 32: 2525-2541.
HELSON, B., LYONS, B., and DE GROOT, P. 1999. Evaluation of neemEC formulations containing azadirachtin for forest insect pestmanagement in Canada. Azadirachta Indica a. Juss. 79–89.
HOWARD, R.W., 1993, Cuticular hydrocarbons and chemical commu-nication, pp. 179–226 in: Insect Lipids: Chemistry, Biochemistryand Biology, D. W. Stanley-Samuelson and D. R. Nelson, eds.University of Nebraska. Press, Lincoln.
HOWARD R, W, and BLOMQUIST G, J. 2005. Ecological, behavioral,and biochemical aspects of insect hydrocarbons. Annu. Rev.Entomol. 50: 371–393.
JAHN, E. 1967. On a population outbreak of the pine false webworm,Acantholyda erythrocephala Chr. in the Steinfeld, Lower Austriain the years 1964–1967. Anz. Schaedingskd. 39: 145–152.
JEWETT, D. M., MATSUMURA, F., and COPPEL, H. C. 1976. Sexpheromone specificity in the pine sawflies: interchange of acidmoieties in an ester. Science. 192: 51–53.
JOHNSON, W. T., and LYON, H H. 1991. Insects that Feed on Trees andShrubs. Comstock Pub. Associates, Ithaca. NY.
JOHNSON, D. K., DONOHOE, J., and KANG, J. 1994. Dilithiumtetrarchlorocuprate catalyzed coupling of allylmagnesium bro-mide with α,ω-dihaloalkanes. Synth. Comm. 24: 1557–1564.
KENIS, M., and KLOOSTERMAN, K. 2001. European Parasitoids of thePine False Webworm (Acantholyda erythrocephala (L.) andTheir Potential for Biological Control in North America.
LYONS. D. B. 1994. Development of the aboreal stages of the pine falsewebworm (Hymenoptera: Pamphiliidae). Env. Entomol. 23: 846–854
LYONS, D. B. 1999. Phenology of the native parasitoid Sinophorusmegalodontis (Hymenoptera: Ichneumonidae) relative to itsintroduced host, the pine false webworm (Hymenoptera: Pam-philiidae). Canad. Entomologist. 131: 787–800.
LYONS D. B., HELSON B. V., JONES G. C., and MCFARLANE J. W. 1993.Development of a chemical control strategy for the Pine FalseWebworm, Acantholyda erythrocephala (L.) (Hymenoptera:Phamphiliidae) Canad. Entomologist. 125: 499–511
LYONS, D. B., HELSON B.V, JONES, G.C., MCFARLANE, J,W., andSCARR, T. 1996. Systemic activity of neem seed extractscontaining azadirachtin in pine foliage for control of the pinefalse webworm, Acantholyda erythrocephala (Hymenoptera:Pamphiliidae). Proc. Entomol. Soc.Ontario 127: 45–55.
LYONS, D. B., HELSON, B.V, JONES G. C, and MCFARLANE J.W. 1998.Effectiveness of Neem- and diflubenzuron-based insecticides forcontrol of the pine false webworm, Acantholyda erythrocephala(L.) (Hymenoptera: Pamphiliidae). Proc. Entomol. Soc.Ontario129: 115–126.
LYONS D. B., HELSON B.V., THOMPSON D. G., MCFARLANE J.W.,ROBINSON A. G., and MICKLE R. E. 2003. Efficacy of ultra-lowvolume aerial application of an azadirachtin-based insecticide forcontrol of the pine false webworm, Acantholyda erythrocephala(L.) (Hymenoptera : Pamphiliidae), in Ontario, Canada. Intl, J.Pest Manag. 49: 1–8,
MAYFIELD A. E., ALLEN D. C., and BRIGGS R. D. 2005. Radialgrowth impact of pine false webworm defoliation on easternwhite pine. Canad. J. Forest Res.-Revue Canadienne DeRecherche Forestiere 35: 1071–1086.
MCGUIRE, M.R., BEHLE, R.W., GOEBEL, H.N., and FRY, T.C. 2000.Calibration of a sunlight simulator for determining solar stabilityof Bacillus thuringiensis and Anagrapha falcifera nuclearpolyhedrovirus Environm. Entomol. 29: 1070–1074
MIDDLEKAUFF, W. W. 1938. Occurrence of a European sawflyAcantholyda. erythrocephala (L.) in New York State. J. Entomol.Soc. New York, 46: 438.
MIDDLEKAUFF, W. W. 1958. North American sawflies of the generaAcantholyda, Cephalcia, Neurotoma (Hymenoptera Pamphilii-dae) Univ. Calif. Publ. Entomol. 14: 51–174
J Chem Ecol (2009) 35:1448–1460 1459

MIYASHITA, N., YOSHIKOSHI, A., and GRIECO, P. A. 1977. Pyridiniump-toluenesulfonate: a mild and efficient catalyst for the tetrahy-dropyranylation of alcohols. J. Org. Chem. 42: 3772–3774.
MOODY, B. H. 1990. Forest insect and disease conditions in Canada,1988. Forest Insect and Disease Survey. Forestry Canada Ottawa,Ont.
OSTRAND, F., ANDERBRANT, O., WASSGREN, A.B., BERGSTRÖM, G.,HEDENSTRÖM, E., HÖGBERG, H.E., BAVU, N., and LARSSON, M.2003. Stereoisomeric composition of the sex pheromone of thepine sawfly Microdiprion pallipes (Hym., Diprionidae). Chemo-ecology. 13: 155–162.
SIEGEL, S. and CASTELLAN, N. J. 1988. Nonparametric Statistics forthe Behavioural Sciences. New York. McGraw-Hill.
SMITH, D. R. 1993. Systematics, life history, and distribution ofsawflies. pp. 3–32 in Sawfly Life History Adaptations to WoodyPlants. Wagner, M. R.; Raffa, K. F (eds).
SONNET, P. E. 1974. cis-Olefins from the Wittig reaction. Org. Prep.Proced. Int. 6: 269–273.
STAPLES, J. K., 1999. Semiochemicals of the Pine False WebowrmAcantholyda erythrocephala (L.) (Hymenoptera: Pamphiliidae).M.S. thesis State University of New York College of Environ-mental Science and Forestry. Syracuse New York.
StatSoft, Inc. (2001). STATISTICA (data analysis software system),version 6. www.statsoft.com.
STEINER, S., MUMM, R., and RUTHER, J. 2007. Courtship Pheromonesin parasitic wasps: Comparison of bioactive and inactivehydrocarbon profiles by multivariate statistical methods. J.Chem. Ecol. 33: 825–838.
SWEDENBORG, P.D., and JONES, R.L. 1992. (Z)-4-tridecenal, apheromonally active air oxidation product from a series of (Z,
Z)-9,13-dienes in Macrocentrus grandii Goidanich (Hymenop-tera: Braconidae). J. Chem. Ecol. 18: 1913–1931.
SYME, P. D. 1981. Occurrence of the introduced sawfly Anantholydaerythrocephala (L.) in Ontario. Dept. of the Env. Can. For. Servc,Attawa, Ont. Res. Notes 1: 4–5
TAI, A., HIGASHIURA, Y., KAKIZAKI, M., NAITO, T., TANAKA, K.,FUJITA, M., SUGIMURA, T., HARA, H., and HAYASHI N. 1998.Field and electroantennogram responses of the pine sawfly,Diprion nipponica, to chiral synthetic pheromone candidates.Biosci. Biotechnol. Biochem. 62: 607–608.
TAI, A., SYOUNO, E., TANAKA, K., FUJITA, M., SUGIMURA, T.,HIGASHIURA, Y., KAKIZAKI, M. HARA, H., NAITO, T. 2002.Regio- and stereochemical study of sex pheromone of pinesawfly; Diprion nipponica. Bull Chem Soc Jpn. 75: 111–121.
THOMPSON, D.G., MICKLE, R.E., LYONS, D.B., HELSON, B.V.,ROBINSON, A.G., CHARTRAND, D.T., and BUSCARINI, T.M.,2003 Deposition of azadirachtin following ultra-low volumeaerial applications to red pine plantations for control of pine falsewebworm, Acantholyda erythrocephala (L.) (Hymenoptera:Pamphiliidae), in Ontario, Canada. Intl. J. Pest Manag. 49: 9–15
WASSGREN, A.B., BERGSTRÖM, G., SIERPINSKI, A., ANDERBRANT, O.,HÖGBERG, H.E., and HEDENSTRÖM, E. 2000. Sex pheromone ofthe pine sawfly Macrodiprion nemoralis (Hymenoptera: Diprio-nidae): Identification of (2S,3R,7R,9S)-3,7,9-trimethyl-2-trideca-nol as the precursor for the active pheromone acetate.Naturwissenschaften. 87: 24–29.
WILEY SPECTRALLIBRARY [CD-Rom], 6th ed 1995,Wiley NewYork.WILSON, G.G. 1984. Infection of the pine false webworm by
Pleistophora schubergi (Microsporida). Canad. For. ServiceRes. Notes. 4: 7–8.
1460 J Chem Ecol (2009) 35:1448–1460
Related Documents











