Comparative Biochemistry and Physiology Part C 124 (1999) 211 – 220 Serotonergic responses of the siphons and adjacent mantle tissue of the zebra mussel, Dreissena polymorpha Jeffrey L. Ram a, *, Denise Moore a , Sashi Putchakayala a , Azrael A. Paredes a , Daniel Ma a , Roger P. Croll b a Department of Physiology, Wayne State Uni6ersity, Detroit, MI 48201 USA b Department of Physiology and Biophysics, Dalhousie Uni6ersity, Halifax, NS, Canada Received 19 January 1999; received in revised form 28 July 1999; accepted 3 August 1999 Abstract Bivalve siphons have important functional roles in nutritional physiology, defense, and reproductive mechanisms, yet little is known about their neuromuscular control. In the present study, tension measurements and video observations of siphons and adjacent mantle tissue were used to investigate responses to serotonin (5-HT). 5-HT caused relaxation at 10 -6 M and 10 -5 M, contraction or biphasic responses at 10 -4 M and 10 -3 M, and siphon opening at 10 -4 M and 10 -3 M. Responses were slow and lasted 5–10 min after much shorter (20 s) applications of 5-HT. The relaxation phase was enhanced in high potassium medium. Contractile responses could be mimicked by a-1-methyl-5-HT and 2-methyl-5-HT but not by 8-OH-DPAT. The responses were not affected by methiothepin, TFMPP, 1-(1-naphthyl)piperazine, metergoline, NAN-190, mianserin, cyprohep- tadine, and ketanserin. The pharmacology of the 5-HT receptor(s) mediating these siphon/mantle responses is, therefore, different from previously described 5-HT receptors involved in spawning. The siphon/mantle contains previously undescribed longitudinal muscle fibers in the mantle and circular muscle fibers around the siphons. Serotonergic innervation of the siphon margins and mantle tissue was observed by immunohistology. The presence of 5-HT in the siphon/mantle tissue and the responsiveness of these preparations to 5-HT suggest that 5-HT may be a physiological regulator of mantle and siphon movements in the zebra mussel. © 1999 Elsevier Science Inc. All rights reserved. Keywords: Contraction; Dreissena polymorpha ; 5-hydroxytryptamine; Immunohistology; Mantle; Serotonin; Siphon; Zebra mussel www.elsevier.com/locate/cbpc 1. Introduction The inhalant and exhalant siphons of bivalves play an important role in the nutritional physiology, defense, and reproductive mechanisms of bivalves, yet little is known about their neuromuscular control. Although the actual filtration of phytoplankton and other partic- ulates takes place on the gills [22], the siphons can regulate the influx of water into the branchial cavity, the back pressure in the branchial cavity produced by ‘the bivalve pump,’ and the exit velocity and range of the exhalant water jet [6,7]. Siphons extend from the animal and move actively during filtering. In zebra mussels, the exhalant siphon decreases in average aper- ture in response to increases in filtrate concentration [10]. Changes in branchial cavity pressure in response to changes in the openings of inhalant and exhalant siphons during filtering may play a role in filtration efficiency [5] and in hydrodynamics of mixing near the mussel bed. These hydrodynamics can affect food and feces recycling [14], larval settlement [1], and distance of gamete dispersion [7]. Furthermore, closing and with- drawal of siphons into the shells may have an impor- tant role in defense and avoidance of toxic substances. Despite the important functional role of the siphons in bivalves, virtually nothing was known until recently about their neuromuscular control. In a recent study from this laboratory on the pharmacological control of the siphons in the zebra mussel, Ram et al. [16] showed that acetylcholine and FMRFamide contract the siphons and adjacent mantle region. In the present study, the effects of serotonin (5-hydroxytryptamine, * Corresponding author. Tel.: +1-313-5771558; fax: +1-313- 5775494. E-mail address: [email protected] (J.L. Ram) 0742-8413/99/$ - see front matter © 1999 Elsevier Science Inc. All rights reserved. PII:S0742-8413(99)00068-7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Comparative Biochemistry and Physiology Part C 124 (1999) 211–220
Serotonergic responses of the siphons and adjacent mantle tissueof the zebra mussel, Dreissena polymorpha
Jeffrey L. Ram a,*, Denise Moore a, Sashi Putchakayala a, Azrael A. Paredes a,Daniel Ma a, Roger P. Croll b
a Department of Physiology, Wayne State Uni6ersity, Detroit, MI 48201 USAb Department of Physiology and Biophysics, Dalhousie Uni6ersity, Halifax, NS, Canada
Received 19 January 1999; received in revised form 28 July 1999; accepted 3 August 1999
Abstract
Bivalve siphons have important functional roles in nutritional physiology, defense, and reproductive mechanisms, yet little isknown about their neuromuscular control. In the present study, tension measurements and video observations of siphons andadjacent mantle tissue were used to investigate responses to serotonin (5-HT). 5-HT caused relaxation at 10−6 M and 10−5 M,contraction or biphasic responses at 10−4 M and 10−3 M, and siphon opening at 10−4 M and 10−3 M. Responses were slowand lasted 5–10 min after much shorter (20 s) applications of 5-HT. The relaxation phase was enhanced in high potassiummedium. Contractile responses could be mimicked by a-1-methyl-5-HT and 2-methyl-5-HT but not by 8-OH-DPAT. Theresponses were not affected by methiothepin, TFMPP, 1-(1-naphthyl)piperazine, metergoline, NAN-190, mianserin, cyprohep-tadine, and ketanserin. The pharmacology of the 5-HT receptor(s) mediating these siphon/mantle responses is, therefore, differentfrom previously described 5-HT receptors involved in spawning. The siphon/mantle contains previously undescribed longitudinalmuscle fibers in the mantle and circular muscle fibers around the siphons. Serotonergic innervation of the siphon margins andmantle tissue was observed by immunohistology. The presence of 5-HT in the siphon/mantle tissue and the responsiveness of thesepreparations to 5-HT suggest that 5-HT may be a physiological regulator of mantle and siphon movements in the zebra mussel.© 1999 Elsevier Science Inc. All rights reserved.
Keywords: Contraction; Dreissena polymorpha ; 5-hydroxytryptamine; Immunohistology; Mantle; Serotonin; Siphon; Zebra mussel
www.elsevier.com/locate/cbpc
1. Introduction
The inhalant and exhalant siphons of bivalves playan important role in the nutritional physiology, defense,and reproductive mechanisms of bivalves, yet little isknown about their neuromuscular control. Althoughthe actual filtration of phytoplankton and other partic-ulates takes place on the gills [22], the siphons canregulate the influx of water into the branchial cavity,the back pressure in the branchial cavity produced by‘the bivalve pump,’ and the exit velocity and range ofthe exhalant water jet [6,7]. Siphons extend from theanimal and move actively during filtering. In zebramussels, the exhalant siphon decreases in average aper-
ture in response to increases in filtrate concentration[10]. Changes in branchial cavity pressure in responseto changes in the openings of inhalant and exhalantsiphons during filtering may play a role in filtrationefficiency [5] and in hydrodynamics of mixing near themussel bed. These hydrodynamics can affect food andfeces recycling [14], larval settlement [1], and distance ofgamete dispersion [7]. Furthermore, closing and with-drawal of siphons into the shells may have an impor-tant role in defense and avoidance of toxic substances.
Despite the important functional role of the siphonsin bivalves, virtually nothing was known until recentlyabout their neuromuscular control. In a recent studyfrom this laboratory on the pharmacological control ofthe siphons in the zebra mussel, Ram et al. [16] showedthat acetylcholine and FMRFamide contract thesiphons and adjacent mantle region. In the presentstudy, the effects of serotonin (5-hydroxytryptamine,
* Corresponding author. Tel.: +1-313-5771558; fax: +1-313-5775494.
E-mail address: [email protected] (J.L. Ram)
0742-8413/99/$ - see front matter © 1999 Elsevier Science Inc. All rights reserved.PII: S 0 7 4 2 -8413 (99 )00068 -7

J.L. Ram et al. / Comparati6e Biochemistry and Physiology, Part C 124 (1999) 211–220212
5-HT) have been investigated. 5-HT is of interest be-cause it is known to have important roles in othermolluscan tissues, notably in stimulating gamete releasefrom gonads [12,19], regulating surface area and waterchannels in gills [13], and having excitatory, inhibitory,or modulatory effects on muscles in the cardiovascularsystem [3] and byssal control system [24,27] of variousbivalves. Yet, the roles of 5-HT in regulating move-ments of bivalve siphons and mantle have not previ-ously been investigated. The present paper investigatesthe effects and presence of 5-HT in the siphons andadjacent mantle tissue of the zebra mussel.
The zebra mussel, Dreissena polymorpha, is a small(B3 cm in length) freshwater bivalve that is readilyobtained in the Great Lakes region ([2,18]; see also TheZebra Mussel Page, http://www.science.wayne.edu/�jram/zmussel.htm). Its tendency to live in dense aggre-gates [9,23] has caused economic problems as a result ofbiofouling [9]. In addition, biofiltering by high densitiesof zebra mussels have caused ecological changes be-cause of increased water clarity and deposit of feces andpseudofeces in the benthos [11]. Increased knowledge ofthe mechanisms regulating zebra mussel behavior, in-cluding siphon and mantle movements, might lead tounderstanding the processes mediating the environmen-tal impacts of zebra mussels and to the development ofnew control methods.
The findings described here, with both tension mea-surements and video observations of siphons andmantle in in vitro preparations, show a complex set ofresponses to 5-HT. The responses include both contrac-tion and relaxation of siphon/mantle preparations, aswell as changes in the apertures of both inhalant andexhalant siphons. In addition, the presence of 5-HT inthe siphon region is demonstrated immuno-histologically.
2. Materials and methods
2.1. Animals and dissection
Zebra mussels were collected by scraping from bulk-heads on Belle Isle Park in the Detroit River. Musselswere transported back to the laboratory in an ice-cooled container in less than 1 h, cleaned of debris, andthen maintained at 12°C with an LD cycle of 16 h light:8 h dark.
All experiments utilized the isolated siphon/mantlepreparation, described previously [16], and illustrated inthe video images in this paper. Sharpened forceps in-serted between the valves along one shell in the siphonregion cut the attachment of the mantle to one shell,and upon opening the shells the siphon/mantle prepara-tion was dissected free of the other shell and tissues.The preparation consisted of both siphons, mantle tis-
sue between the siphons and along their sides, as well asmantle tissue extending several mm beyond each siphonto provide tissue for attaching threads or pins, asappropriate. The overall dimensions of these siphon/mantle preparations are approximately 2 mm wide and1 cm long.
2.2. Tension measurements
Siphon/mantle preparations were mounted in a mus-cle chamber (volume 0.3 ml), anchored by one thread atthe bottom of the chamber and the other attached to aGrass FT.03 force transducer. Responses were recordedon a Grass Polygraph Model 79D. Initial tension of thepreparations was usually set equivalent to the forceexerted by approximately 200 mg. Preparations werecontinuously superfused with zebra mussel Ringer (0.5mM KCl and 5 mM each of CaCl2, NaCl, NaHCO3,Na2SO4, and HEPES, pH 7.8) or other solutions, whichwere pumped through the chamber at 3.5 ml/min atambient temperatures. At various intervals, indicated inResults, the perfusate was changed for brief periods(usually 20 s) to solutions containing 5-HT or otherchemicals dissolved in zebra mussel Ringer or othersolutions. A high potassium Ringer (‘Na:K 80:20medium’) was made by replacing 20% of the sodiumsalts with their corresponding potassium salts (giving atotal potassium concentration of 4.5 mEquivalents/l).The methodology and apparatus are similar to proce-dures previously described to study Aplysia buccal mus-cle contraction [21].
2.3. Video and isotonic contraction measurements
Dissected siphon/mantle preparations were mountedin a small well (volume 0.3 ml) carved out of Sylgard.One end of the preparation was immobilized by beingpinned down to the Sylgard, while the other end of thepreparation was attached by a thread to an isotonictransducer (Gould Metripak Model 793341), amplifiedby a Gould Model 11 DC Amplifier, and recorded on aBrush 220 pen recorder. The siphon region of thepreparation was visualized by video recording througha dissecting microscope (JVC Compact VHS camcorderrecording through one eyepiece of a dissecting micro-scope). Video images were captured either by using aTarga framegrabber and HALO Desktop Imager soft-ware (as described previously [17]), or by usingSNAPPY (Play, Inc.).
2.4. Histology
Zebra mussels were fixed in 4% paraformaldehydebuffered in 0.1 M phosphate buffer (pH 7.4), dehy-drated through an ascending ethanol series, embeddedin paraffin, and then sectioned at 10 mm. Following

J.L. Ram et al. / Comparati6e Biochemistry and Physiology, Part C 124 (1999) 211–220 213
deparaffination and rehydration, the sections were ei-ther stained with hematoxylin and eosin or processedfor immunocytochemistry. Immunocytochemical proce-dures for use in bivalve species have been describedpreviously [16,18,26]. For this study, the primary anti-bodies were affinity-purified, polyclonal anti-5-HT anti-bodies raised in rabbit (IncStar, Stillwater, MN, USA).FITC- and rhodamine goat-anti-rabbit secondary anti-bodies were used to detect the anti-5-HT antibodies.Peroxidase-anti-peroxidase detection was also used tolocalize the primary antibodies, as described previously[8,16]. Positive controls consisted of simultaneous pro-cessing of Lymnaea central ganglia known to contain5-HT. As a negative control, tissues were processed asdescribed above but incubation in the anti-5-HT serumwas omitted from the procedure.
3. Results
3.1. Tension measurements
A 20-s application of 5-HT caused both relaxationand contraction of siphon/mantle preparations testedwith isometric contraction measurements (Fig. 1). 5-HTcaused relaxation at low concentrations (10−5 M, andsometimes as low as 10−6 M; see Fig. 1B and Fig. 3).At 10−4 M, 5-HT typically caused a biphasic response,with an initial contraction followed by a slower relax-ation (Fig. 1A); an initial, but brief contractile phasewas also occasionally seen at 10−5 M (Fig. 1A). 10−3
M 5-HT caused siphon/mantle preparations to con-tract, occasionally with fast twitches superimposed on a
Fig. 2. Summary of the relaxation and contraction responses toserotonin (5-HT) from 17 preparations which were tested at 10−5 M,10−4 M, and 10−3 M 5-HT, similar to Fig. 1A. Latencies to reachthe peak contractions and/or relaxations were measured and areindicated by the points with horizontal error bars (mean9SEM).Response magnitudes were calculated as percent of the maximalcontraction to 10−3 M 5-HT and plotted as mean9SEM (verticalbars). Magnitudes were measured at peak relaxation or contractionand also, for preparations that had no relaxation phase, at 5.2 minafter 5-HT application (the time of peak relaxation for preparationsthat relaxed in response to 10−4 M). Data for 10−4 M is plotted intwo curves: one is the average of nine preparations that had no netrelaxation phase; the other is the average of eight preparations thatdid relax. Curves were fitted by eye to pass through plotted pointsand to match approximately the shapes of observed tension record-ings.
tonic response (Fig. 1A). This pattern of responses hasbeen observed in more than 25 preparations. No suchchanges in tension were elicited by changing perfusionmedium to zebra mussel Ringer with no added chemical(a negative control; data not shown), lower concentra-tions of 5-HT (data not shown), or a variety of otherchemicals that have no apparent direct effects onsiphon/mantle tension (several examples shown later inthis paper).
The peak responses and latencies to 20- or 25-s pulsesof 10−5 M, 10−4 M, and 10−3 M 5-HT were measuredin 17 preparations (Fig. 2). In all preparations, 5-HTwas applied in ascending concentrations, at intervals ofat least 10 min. As a percent of the peak contraction at10−3 M 5-HT, the responses to 10−5 M contracted nomore than approximately 5%, and the peak contractionto 10−4 M averaged 32%94% (mean9SEM). Con-tractions in response to 10−3 M 5-HT took an averageof 3 min to reach their peak. Responses to lowerconcentrations peaked earlier, probably because of theonset of the slower relaxation phase at these lowerconcentrations. Relaxation was maximal at approxi-mately 3–5 min after the 5-HT application. All prepa-rations treated with 10−5 M 5-HT exhibited arelaxation phase. Only about half of the preparationstreated with 10−4 M 5-HT relaxed below the initialtension, and their average relaxation and latency is
Fig. 1. Relaxation and contraction in response to various concentra-tions of serotonin (5-HT) applied to zebra mussel siphon/mantlepreparations. Preparations were continuously superfused with normalRinger which was changed to Ringer containing various concentra-tions of 5-HT for 20 s at the times indicated by the arrows. (A)Typical responses to 10−5 M, 10−4 M, and 10−3 M 5-HT. (B)Another preparation. This preparation was more sensitive than most,showing relaxation responses to 5-HT concentrations as low as 10−6
M.

J.L. Ram et al. / Comparati6e Biochemistry and Physiology, Part C 124 (1999) 211–220214
plotted in a separate curve from those that did notexhibit net relaxation (Fig. 2).
5-HT responses were slow and long-lasting: when5-HT was applied for just 20 or 25 s, as in the aboveexperiments, the tonic responses, whether relaxation orcontraction, lasted 5–10 min beyond the peak re-sponses. Longer applications of 5-HT similarly causedrelaxation and contraction while 5-HT was present,indicating the responses were not rebound responses toremoval of 5-HT. An example of the response to longapplications (10–12 min) of 10−6 M, 10−5 M, and10−3 M 5-HT is shown in Fig. 3. A 10-min applicationof 10−6 M 5-HT produced a relaxation that was moresustained than that produced by a 25-s application.With 10−5 M 5-HT, a 25-s application sometimesproduced a biphasic contraction (e.g. as quantitated inFig. 2), and the 10-min application of 10−5 M pro-duced a contraction that partially relaxed during the5-HT application. Contractile responses that partiallyrelaxed during 10-min applications of 10−5 M 5-HTwere observed in five of seven preparations exposed for10 min to 10−5 M 5-HT; the other two preparationsgave sustained relaxation throughout the application of10−5 M 5-HT. At 10−3 M 5-HT, three of three prepa-rations gave larger contractions than in response to 25s of 10−3 M 5-HT, but, as illustrated in Fig. 3, thecontraction mostly relaxed during the 10-min exposureand was not followed by further relaxation.
3.2. Video and isotonic contraction measurements
To determine the types of movements that mightcorrespond to the tension measurements describedabove, we used video micrography with simultaneous
Fig. 4. Relaxation and contraction of siphon/mantle preparationsrecorded with an isotonic transducer. Serotonin (5-HT) was appliedby continuous superfusion at the indicated times for 30 s. (A)Relaxation and contraction responses to 10−5 M and 10−3 M 5-HT,respectively. (B) Recording from preparation that was simultaneouslyobserved by videomicrography, as illustrated in Fig. 5. Letters A, B,C, and D correspond to the time points illustrated in Fig. 5. 5-HTsuperfusion for 30 s began at the time point indicated by ‘A’.
isotonic contraction measurements. Under constanttension, preparations relaxed slowly. Responses to 5-HT were superimposed on this slow relaxation. At10−5 M, 5-HT typically produced an increase in relax-ation rate lasting several minutes (Fig. 4); 10−3 Mcaused contraction of the preparations. These resultscorresponded well with the responses seen with isomet-ric measurements described above.
Video observations showed that a high concentration(10−3 M) of 5-HT caused opening of both inhalant andexhalant siphons. A typical response is illustrated in theimages in Fig. 5. Opening of the siphons was frequentlyaccompanied by twitch-like movements of the siphonmargin or, for the inhalant siphon, twitches of thetentacles. As with the tension and contraction measure-ments, the latency of the opening responses of thesiphons was long, with maximal opening occurring onaverage 1–2 min (n=9) after a 30-s pulse of 5-HT usedto activate the response. As with the tension measure-ments, long applications of 5-HT also caused activationof siphon opening, with similar latency, showing thatthe opening is not a ‘rebound’ elicited by washout ofthe 5-HT. With long applications, 10−4 M was alsoconsistently able to activate opening responses.
By recording isotonic contraction and video imagessimultaneously on the same preparations, it was some-times possible to identify specific movements that corre-sponded to the measured contraction. The video imagesin Fig. 5 showed a clear shortening of the preparation,corresponding to the measured isotonic contraction(Fig. 4B). Detailed measurements of video images cor-responding to contractions recorded on several siphon/
Fig. 3. Relaxation and contraction of a siphon/mantle preparation inresponse to long applications of serotonin (5-HT). The two lines arefrom a continuous recording, separated by about 8 min. 5-HT wasapplied by continuous superfusion at the indicated concentrationsthroughout the periods shown by the horizontal black bars. An initialshort application of 10−6 M 5-HT for 25 s demonstrated that thepreparation responded normally to a short application of 5-HT.

J.L. Ram et al. / Comparati6e Biochemistry and Physiology, Part C 124 (1999) 211–220 215
mantle preparations, measuring incurrent siphon widthand length, the distance between siphons, as well as thedistances between identifiable marks on either end ofthe video field, indicated that the shortening did notcorrespond to any decrease in siphon diameter but,rather, was a general shortening of the entire prepara-tion. Indeed, as is evident in Fig. 5, the shortening ofthe preparation occurred while siphons were gettinglarger and, in fact, followed a different time coursefrom the overall contraction.
Fig. 6. Lack of effect of 8-OH-DPAT, an effective activator ofspawning in zebra mussels, on tension of a siphon/mantle prepara-tion. Prior to 8-OH-DPAT, application of 10−5 M 5-HT showed theusual relaxation response (top trace). 8-OH-DPAT was applied atsuccessively increasing concentrations for 20 s at 8-min intervals(middle three traces). Subsequent application of 10−3 M 5-HT(bottom trace) showed the usual contractile response.
Fig. 5. Videomicrograph of siphon/mantle responses to 10−3 Mserotonin. Images in A, B, C, and D correspond to time pointsillustrated in Fig. 4B. ‘In’ and ‘Ex’ labels are located above theinhalant and exhalant siphons, respectively. Ventral (v) is to the left,and dorsal (d) is to the right. Calibration mark: 1 mm.
Although we were able to identify isotonic contrac-tion measurements with a general shortening of thepreparation, the much smaller isotonically-recordedrelaxation responses, obtained in response to 10−5 M5-HT, yielded no reliably identifiable movements ofthe preparation. Thus, at this concentration neithersiphon opening nor preparation lengthening wereevident within the resolution offered by the video im-ages.
3.3. Pharmacology of siphon/mantle serotonergicresponses
The pharmacology of 5-HT receptors involved inzebra mussel spawning behavior has previously beeninvestigated [20]. To compare the 5-HT receptor(s)causing siphon/mantle responses to those mediatingspawning in zebra mussels, we tested several serotoner-gic compounds that had previously been tested onspawning. 8-OH-DPAT, which is as effective as 5-HTin causing spawning, had no effect on siphon/mantletension (Fig. 6). Similarly, no effects on siphon apertureor isotonic contraction were seen with 8-OH-DPAT atconcentrations up to 10−3 M (data not shown). Me-thiothepin is the most effective antagonist of spawning;however, methiothepin blocked neither relaxation norcontractile responses of the siphon/mantle to 5-HT(Fig. 7). Other compounds known to affect spawningthat were without effect on siphon/mantle tension in-cluded TFMPP, 1-(1-naphthyl)piperazine, metergoline,NAN-190, mianserin, cyproheptadine, and ketanserin.All of these compounds were tested for effects (relax-ation, contraction or, for inhibitors, blocking 5-HTresponses) on at least two preparations each.

J.L. Ram et al. / Comparati6e Biochemistry and Physiology, Part C 124 (1999) 211–220216
Fig. 7. Lack of effect of methiothepin, the most effective antagonistof 5-HT elicited spawning in zebra mussels, on 5-HT elicited relax-ation and contraction of siphon/mantle preparations. (A) Relaxationin response to 10−5 M 5-HT was similar whether 10−5 M methio-thepin was applied 2 min preceding and concommitantly with 5-HT(first application) or 5-HT was applied without methiothepin (secondapplication). (B) Contractile response to 10−3 M 5-HT was notgreatly reduced when 5-HT was applied following 2-min applicationand concommitantly with 10−4 M methiothepin.
Fig. 9. Contractile responses of siphon/mantle preparation to 2-methyl-5-HT (2-Me-5HT). Following application of 2-Me-5HT, bothrelaxation and contraction responses to successively increasing con-centrations of 5-HT were demonstrated. The two lines are from acontinuous recording, separated by approximately 3 min betweenupper and lower traces.
medium was larger than in normal medium even whenthe initial tension in the presence of 80:20 Na:Kmedium was adjusted to the same initial value (200 mg)as in normal medium prior to 5-HT application. Undersuch conditions, relaxation in 80:20 Na:K medium, as apercent of the maximal contractile response to 10−3 M5-HT in normal medium, averaged −2791% at 10−5
M, −95942% at 10−4 M, and −65%91% at 10−3
M 5-HT, with latencies to maximal relaxation of 3, 6,and 11 min, respectively (n=3 preparations; compareto values in Fig. 2). The contractile phases of theresponses to 10−4 M and 10−3 M 5-HT still occurredin high potassium medium but were reduced in magni-tude by about 30% from their levels in normal medium.
3.4. Morphology
Previous descriptions of the muscle fibers of zebramussel mantle [15] had shown only fibers at the edge ofthe mantle, oriented towards the midline (the pallialretractor muscles), which is perpendicular to the direc-tion of contraction and relaxation being measured inthe present study. To ascertain the orientation of mus-cle fibers in the siphon region, this region was sectionedand stained with hematoxylin-eosin (Fig. 10). The mi-crographs clearly showed long cylindrical cells (putativemuscle cells) in several orientations: [1] around thesiphons themselves, fibers ran circularly around theouter margins of the siphons (Fig. 10A); [2] for most ofthe length of the mantle regions of the preparations,fibers ran longitudinally (i.e. along the axis of ourmeasured contractions (Fig. 10B and C); and [3] alongthe edges of the preparations, a comparatively smallnumber of fibers were oriented across the preparation
In contrast, two serotonergic compounds that havelittle or no effect on spawning, a-1-methyl-5-HT (Fig.8) and 2-methyl-5-HT (Fig. 9), mimicked the contractileeffects of 5-HT. With video observations, neither ofthese compounds activated siphon opening at 10−3 M.Neither a-1-methyl-5-HT nor 2-methyl-5-HT mimickedthe relaxation effect of 5-HT on the siphon prepara-tions. Indeed, 5-HT is the only compound that we havefound, so far, that causes relaxation of siphon/mantlepreparations.
The only manipulation that consistently modified the5-HT-elicited relaxation of these preparations wastreatment with high potassium medium. In 80:20 Na:Kmedium, 5-HT elicited greater relaxation over abroader concentration range (as high as 10−3 M 5-HT)than in normal medium. Relaxation in high potassium
Fig. 8. Contractile responses of siphon/mantle preparation to a-1-methyl-5-HT (alpha-1-Me5HT). The procedure was similar to Fig. 6.

J.L. Ram et al. / Comparati6e Biochemistry and Physiology, Part C 124 (1999) 211–220 217
towards the midline (Fig. 10C). The longitudinal mus-cle fibers (Fig. 10B and C) are typical of muscle fibersin the mantle regions located both dorsally and ven-trally from the siphons, beginning within 1 mm of thenearest siphon. Smaller bundles of longitudinally-ori-
ented fibers also occur between the siphons and themantle edge (not shown). The fibers oriented towardsthe midline can occur all along the mantle edge (de-pending on the intactness of this region after dissec-tion), both in the region of the siphons themselves (notshown) and the adjacent mantle area (as in Fig. 10C).
The diameters of these cylindrical cells ranged from3–8 mm. The lengths of these fibers was difficult toascertain as entire cells were not necessarily all in thesame focal plane or even completely within a section;however, we were able to trace single fibers for lengthsof more than 50 mm (e.g. several are apparent in Fig.10C).
3.5. Immunohistology of 5-HT in the siphon region
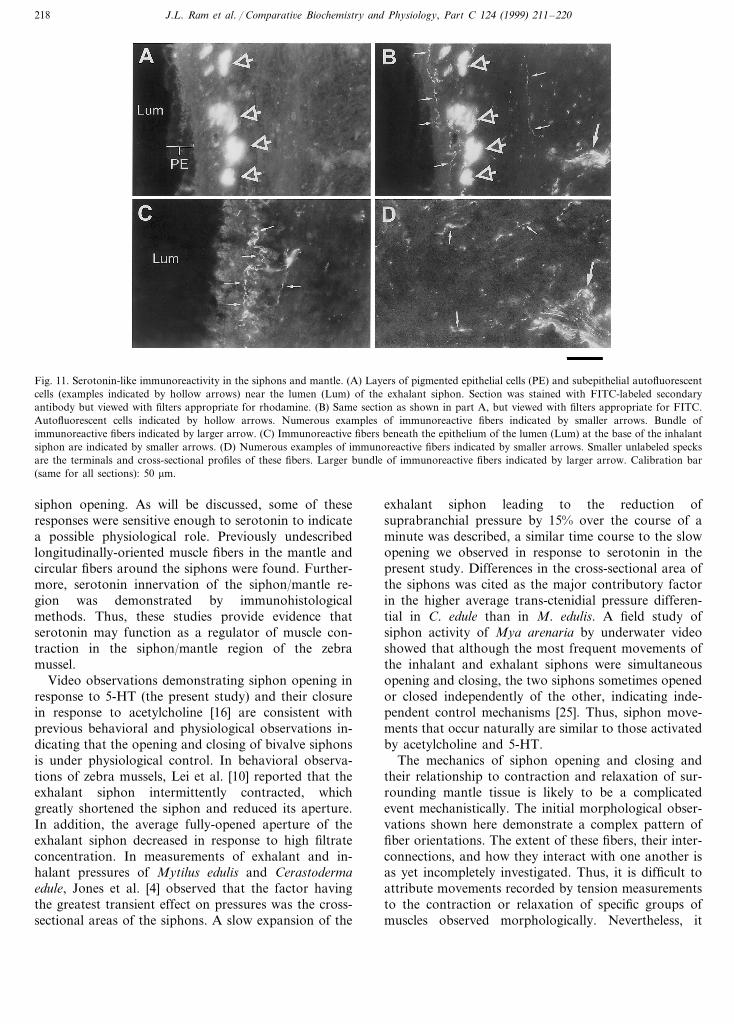
Both FITC- and rhodamine-labeled secondary anti-bodies yielded identical staining patterns whereas omis-sion of the primary antibody from the protocoleliminated all staining except for a layer of autofluores-cent cells lying beneath the pigmented epithelia withinthe lumina of both the exhalant and inhalant siphons.These autofluorescent cells were also visible using eitherepifluorescence filter cube, regardless of which sec-ondary antibody was used (Fig. 11A). Serotonin-likeimmunoreactive fibers and their terminals were concen-trated beneath the pigmented epithelium in the vicinityof the autofluorescent cells of the exhalant siphon (Fig.11B). These 5-HT immunoreactive processes originatedfrom larger bundles of serotonergic fibers which ap-proached the lumen from the mantle. A similar stainingpattern was also observed in the inhalant siphon, exceptat the base of this siphon both the pigmented epithelialcells and the subepithelial autofluorescent cells becameless abundant, thus offering unobscured views of theimmunoreactive fibers (Fig. 11C). Immunoreactivefibers and terminals were generally less concentratedwithin the mantle than around the lumina of the twosiphons. In certain regions of the mantle, however,terminals and immunoreactive fiber bundles were abun-dant, appearing to originate from nerves embedded inthe mantle tissue (Fig. 11D).
Similar staining patterns in both siphons and themantle were also obtained using PAP for detection ofserotonin-like immunoreactivity, except that theautofluorescent subepithelial cells stained light brownand, therefore, again partially obscured the immunore-active fibers and terminals (data not shown).
4. Discussion
This study investigated the effects and presence ofserotonin in the siphon/mantle region of the zebramussel. At least three different types of responses toserotonin were observed: contraction, relaxation, and
Fig. 10. Muscle fibers in siphon/mantle sections stained with hema-toxylin-eosin. All sections in this illustration have the longitudinalaxis oriented horizontally. (A) Muscle fibers (arrows) going aroundthe border of a siphon. (B) Muscle fibers between the arrows areoriented longitudinally in the mantle regions of the preparation. (C)Muscle fibers near the edge of the mantle, showing longitudinal fibers(e.g. arrow) and presumed pallial retractor muscle fibers perpendicu-lar to them (e.g. arrow head). Calibration bars: (A) 100 mm, (B) 100mm, (C) 50 mm.
J.L. Ram et al. / Comparati6e Biochemistry and Physiology, Part C 124 (1999) 211–220218
Fig. 11. Serotonin-like immunoreactivity in the siphons and mantle. (A) Layers of pigmented epithelial cells (PE) and subepithelial autofluorescentcells (examples indicated by hollow arrows) near the lumen (Lum) of the exhalant siphon. Section was stained with FITC-labeled secondaryantibody but viewed with filters appropriate for rhodamine. (B) Same section as shown in part A, but viewed with filters appropriate for FITC.Autofluorescent cells indicated by hollow arrows. Numerous examples of immunoreactive fibers indicated by smaller arrows. Bundle ofimmunoreactive fibers indicated by larger arrow. (C) Immunoreactive fibers beneath the epithelium of the lumen (Lum) at the base of the inhalantsiphon are indicated by smaller arrows. (D) Numerous examples of immunoreactive fibers indicated by smaller arrows. Smaller unlabeled specksare the terminals and cross-sectional profiles of these fibers. Larger bundle of immunoreactive fibers indicated by larger arrow. Calibration bar(same for all sections): 50 mm.
siphon opening. As will be discussed, some of theseresponses were sensitive enough to serotonin to indicatea possible physiological role. Previously undescribedlongitudinally-oriented muscle fibers in the mantle andcircular fibers around the siphons were found. Further-more, serotonin innervation of the siphon/mantle re-gion was demonstrated by immunohistologicalmethods. Thus, these studies provide evidence thatserotonin may function as a regulator of muscle con-traction in the siphon/mantle region of the zebramussel.
Video observations demonstrating siphon opening inresponse to 5-HT (the present study) and their closurein response to acetylcholine [16] are consistent withprevious behavioral and physiological observations in-dicating that the opening and closing of bivalve siphonsis under physiological control. In behavioral observa-tions of zebra mussels, Lei et al. [10] reported that theexhalant siphon intermittently contracted, whichgreatly shortened the siphon and reduced its aperture.In addition, the average fully-opened aperture of theexhalant siphon decreased in response to high filtrateconcentration. In measurements of exhalant and in-halant pressures of Mytilus edulis and Cerastodermaedule, Jones et al. [4] observed that the factor havingthe greatest transient effect on pressures was the cross-sectional areas of the siphons. A slow expansion of the
exhalant siphon leading to the reduction ofsuprabranchial pressure by 15% over the course of aminute was described, a similar time course to the slowopening we observed in response to serotonin in thepresent study. Differences in the cross-sectional area ofthe siphons was cited as the major contributory factorin the higher average trans-ctenidial pressure differen-tial in C. edule than in M. edulis. A field study ofsiphon activity of Mya arenaria by underwater videoshowed that although the most frequent movements ofthe inhalant and exhalant siphons were simultaneousopening and closing, the two siphons sometimes openedor closed independently of the other, indicating inde-pendent control mechanisms [25]. Thus, siphon move-ments that occur naturally are similar to those activatedby acetylcholine and 5-HT.
The mechanics of siphon opening and closing andtheir relationship to contraction and relaxation of sur-rounding mantle tissue is likely to be a complicatedevent mechanistically. The initial morphological obser-vations shown here demonstrate a complex pattern offiber orientations. The extent of these fibers, their inter-connections, and how they interact with one another isas yet incompletely investigated. Thus, it is difficult toattribute movements recorded by tension measurementsto the contraction or relaxation of specific groups ofmuscles observed morphologically. Nevertheless, it

J.L. Ram et al. / Comparati6e Biochemistry and Physiology, Part C 124 (1999) 211–220 219
seems likely that contraction and relaxation detected byforce and movement transducers in response to 5-HT isa result mainly of contractions of the mantle regions.As illustrated in Fig. 5, the contractile response oc-curred with a different time course than siphon open-ing, and furthermore, siphon opening was a movementthat would tend to have the opposite effect on theoverall length of the preparation. Siphon opening alsoprobably was not the movement that was measuredwhen relaxation was observed, because relaxation wasseen at lower 5-HT concentrations, at which littlechange in siphon dimensions occurred. The motivesource of these movements may be the previously unde-scribed muscle fibers running longitudinally within themantle. In further support of this notion, additionalexperiments on pieces of mantle adjacent to (but notincluding) siphons, showed both contractile and relax-ation phases in response to 5-HT (data not shown).Although these longitudinal movements were quitereadily measured in these in vitro preparations, theoccurrence of such movements and their function hasnot been studied in intact bivalves. Possibly, relaxationof these fibers may enable siphons to extend further outof the zebra mussel’s shells, and conversely, contractionmay assist in tightening up the mantle when valve gapeis reduced. Consistent with this suggestion are observa-tions by Jorgensen et al. [7] that application of 10−5 M5-HT to intact M. edulis was reported ‘to overextendthe mantle and siphon, indicating extreme relaxation ofthe muscles.’ It should be noted that the measurementsin this paper examine only the longitudinal tensionalong the mantle and not the tension across the mantle,corresponding in the intact animal to tension betweenthe two shells of the bivalve. Thus, the function andpharmacology of the muscle fibers oriented across thepreparations towards the midline and their interactionwith the circular and longitudinal fibers remain unex-plored. Further studies are needed of the function ofthe siphon and mantle muscles and their regulation byneurotransmitters.
How likely is it that these siphon/mantle movementselicited by 5-HT reflect a physiological function of5-HT? The presence of 5-HT in the siphon/mantleregion, clearly demonstrated by immunohistology, sup-ports a possible physiological role. Secondly, the sensi-tivity of at least some of the responses to 5-HT was inthe expected range for physiological responses. Thus,the relaxation response occurred in response to concen-trations as low as 10−6 M, (Figs. 1 and 3). Thecontractile response typically was not seen in responseto brief pulses of 10−5 M, possibly because of thepredominance of relaxation at this concentration inmost preparations; however, longer applications of 5-HT (e.g. Fig. 3) usually showed contraction at 10−5 M.On the other hand, siphon opening movements in re-sponse to 5-HT were consistently observed only at
10−3 M 5-HT (also seen with long applications of 10−4
M 5-HT; data not shown). Other molluscan musclesthought to be under physiological control by 5-HT aresensitive to 5-HT at concentrations less than 10−5 M(e.g. anterior byssus retractor muscle of Mytilus [27];bivalve hearts [3]; Aplysia buccal muscles [21]). Thus,the relaxation response appears to be elicited in thephysiological range, and the contractile response mayalso be; however, the opening responses of the siphonsappear to be too insensitive. Possibly, the siphon re-sponses are physiological, but diffusion to the appropri-ate siphon opening muscles is restricted. Alternatively,5-HT may be acting at receptors that ordinarily areactivated by other, possibly related agents. The preciselocations of the receptors mediating responses in thesepreparations is unknown, because 5-HT could be actingdirectly on muscle fibers or indirectly through effects onperipheral neurons or presynaptic endings of motorneu-rons. A further consideration about whether these re-sponses to 5-HT may be related to physiologicalfunction is our failure, in general, to find serotonergicinhibitors of their receptors. This implies either thatunique 5-HT receptors are involved or that the physio-logical transmitter(s) may not be serotonergic. Identifi-cation of effective blockers of any of these responseswould be helpful in identifying the physiological regula-tors and ascertaining their functional roles.
Whatever receptor is involved in mediating the re-sponses to 5-HT demonstrated here, it is clearly differ-ent from the serotonergic receptor(s) that mediatespawning in the zebra mussel. None of the blockersthat inhibit zebra mussel spawning had any effect onthe contractile responses of the siphons. Furthermore,two serotonergic agonists that have no effect on spawn-ing did mimic the effect of 5-HT on contraction but didnot elicit relaxation nor siphon opening. The experi-ments with high potassium provide a method for en-hancing the relaxation effect of 5-HT and may be usefulin further analyses of the receptors mediating theresponse.
In summary, the presence of 5-HT in the siphon/mantle tissue and the responsiveness of this tissue to5-HT application suggest that 5-HT may be a physio-logical regulator of mantle and siphon movements inthe zebra mussel. However, activity of the serotonergicsiphon innervation, the types of receptors involved, andtheir roles in behavior need to be explored in futurestudies.
Acknowledgements
This publication is a result of work sponsored, inpart, by NSF Grant IBN9631008, a grant from Michi-gan Sea Grant, and NIH grant GM-08167 to J.L. Ram,and NSERC (Canada) Operating Grant cOPG38863

J.L. Ram et al. / Comparati6e Biochemistry and Physiology, Part C 124 (1999) 211–220220
to R.P. Croll. The authors gratefully acknowledge as-sistance from Janette Nason and Christine Cupps inhistology experiments.
References
[1] Ertman SC, Jumars PA. Effects of bivalve siphonal currents onthe settlement of inert particles and larvae. J Mar Res1988;46:797–813.
[2] Hebert PDN, Muncaster BW, Mackie GL. Ecological and ge-netic studies on Dreissena polymorpha (Pallas): a new mollusc inthe Great Lakes. Can J Fish Aquat Sci 1989;46:1587–91.
[3] Hill RB, Welsh JH. Heart, circulation, and blood cells. In:Wilbur KM, Yonge M, editors. Physiology of Mollusca, vol. 2.New York: Academic Press, 1966:126–74.
[4] Jones HD, Allen JR. Inhalant and exhalant pressures in Mytilusedulis L. and Cerastoderma edule (L.). J Exp Mar Biol Ecol1986;98:231–40.
[5] Jorgensen CB. Bivalve Filter Feeding: Hydrodynamics, Bioen-ergetics, Physiology and Ecology. Fredensborg, Denmark: Olsenand Olsen, 1990.
[6] Jorgensen CB, Famme P, Kristensen HS, Larsen PS, Mohlen-berg F, Riisgard HU. The bivalve pump. Mar Ecol Prog Ser1986;34:69–77.
[7] Jorgensen CB, Larsen PS, Mohlenberg F, Riisgard HU. Themussel pump: properties and modelling. Mar Ecol Prog Ser1988;45:205–16.
[8] Kemenes G, Elekes K, Hiripi L, Benjamin PR. A comparison offour techniques for mapping the distribution of serotonin andserotonin-containing neurons in fixed and living ganglia of thesnail, Lymnaea. J Neurocytol 1989;18:193–208.
[9] Kovalak WP, Longton GD, Smithee RD. Infestation of powerplant water systems by the zebra mussel (Dreissena polymorpha).In: Nalepa TF, Schloesser DW, editors. Zebra Mussels: Biology,Impact and Control. Boca Raton, FL: Lewis Publishers,1993:359–80.
[10] Lei J, Payne BS, Wang SY. Filtration dynamics of the zebramussel, Dreissena polymorpha. Can J Fish Aquat Sci 1996;53:29–37.
[11] MacIsaac HJ. Potential abiotic and biotic impacts of zebramussels on the inland waters of North America. Am Zoolist1996;36:287–99.
[12] Matsutani T, Nomura T. Induction of spawning by serotonin inthe scallop, Patinopecten yessoensis (Jay). Mar Biol Lett1982;3:353–8.
[13] Medler S, Silverman H. Functional organization of intrinsic gillmuscles in zebra mussels, Dreissena polymorpha (Mollusca:Bivalvia) and response to transmitters in vitro. Invertebr Biol1997;116:200–12.
[14] Monismith SG, Koseff JR, Thompson JK, O’Riordan CA, NepfHM. A study of model bivalve siphonal currents. LimnolOceanogr 1990;35:680–96.
[15] Morton B. The anatomy of Dreissena polymorpha and the evolu-tion and success of the heteromyarian form in the Dreissenoidea.In: Nalepa TF, Schloesser DW, editors. Zebra Mussels: Biology,Impacts and Control. Ann Arbor: Lewis Publishers, 1993:185–215.
[16] Ram JL, Baidoun F, Ram ML, Croll RP. Cholinergic andpeptidergic regulation of siphon/mantle function in the zebramussel, Dreissena polymorpha. Comp Biochem Physiol1997;117C:275–82.
[17] Ram JL, Fares MA, Standley PR, Therell LL, ThyagaranjanRV, Sowers JR. Insulin inhibits vasopressin-elicited contractionof vascular smooth muscle cells. J Vasc Biol Med 1993;4:250–5.
[18] Ram JL, Fong P, Croll RP, Nichols SJ, Wall D. The zebramussel (Dreissena polymorpha), a new pest in North America:reproductive mechanisms as possible targets of control strategies.Invertebr Reprod Dev 1992;22:77–86.
[19] Ram JL, Fong PP, Garton DW. Physiological aspects of zebramussel reproduction: maturation, spawning and fertilization. AmZoologist 1996;36:326–38.
[20] Ram JL, Fong PP, Kyozuka K. Serotonergic mechanisms medi-ating spawning and oocyte maturation in the zebra mussel,Dreissena polymorpha. Invertebr Reprod Dev 1996;30:29–37.
[21] Ram JL, Shukla UA, Ajimal GS. Serotonin has both excitatoryand inhibitory modulatory effects on feeding muscles in Aplysia.J Neurobiol 1981;12:613–21.
[22] Silverman H, Lynn JW, Dietz TH. Particle capture by the gills ofDreissena polymorpha : structure and function of latero-frontalcirri. Biol Bull 1996;191:42–54.
[23] Stanczykowska A, Lewandowski K. Thirty years of studies ofDreissena polymorpha ecology in Mazurian Lakes of Northeast-ern Poland. In: Nalepa TF, Schloesser DW, editors. ZebraMussels: Biology, Impacts and Control. Boca Raton, FL: LewisPublishers, 1993:3–37.
[24] Sugi H, Yamaguchi T. Activation of the contractile mechanismin the anterior byssal rectractor muscle of Mytilus edulis. JPhysiol 1976;257:531–47.
[25] Thorin S, Bourdages H, Vincent B. Study of siphon activity inMya arenaria (L.) in the intertidal zone by means of an underwa-ter video camera. J Exp Mar Biol Ecol 1998;224:205–24.
[26] Too CKL, Croll RP. Detection of FMRFamide-like immunore-activity in the sea scallop Placopecten magellanicus by immuno-histochemistry and Western blot analysis. Cell Tissue Res1995;281:295–304.
[27] Twarog BM. The nature of catch and its control. In: Pepe FA,Sanger JW, Nachmias VT, editors. Motility in Cell Function.New York: Academic Press, 1979:231–41.
.
Related Documents











