Neuroscience Vol. 41, NO. 2/3, pp. 381-390, 1991 Printed in Great Britain 0306-4522/91 $3.00 + 0.00 Pergamon Press plc 0 1991 IBRO SEPTAL GABAERGIC NEURONS INNERVATE INHIBITORY INTERNEURONS IN THE HIPPOCAMPUS OF THE MACAQUE MONKEY A. I. GULY&* L. SERW,~ K. T&H,* L. ACSADY,* M. ANTAL~and T. F. FREIJND*$ *Department of Functional Neuroanatomy, Institute of Experimental Medicine, Hungarian Academy of Sciences, Budapest, P.O. Box 67, H-1450, Hungary; and 1st Department of Anatomy, Semmelweis University, Medical School, Budapest, H-1450, Hungary tDepartment of Physiology, University Medical School, Pets, H-7624, Hungary SDepartment of Anatomy, University Medical School, Debrecen, H-4012, Hungary Abstract-The septohippocampal projection was visualized in three Mucaca mulatta monkeys by anterograde transport of Phaseolus oulgarb leucoagglutinin. Following injections of the lectin into the medial septal nucleus, P. &garb leucoagglutinin-labehed fibres were found in the hippocampal complex, mainly in stratum oriens of the CA1 subfield, throughout the CA3 subfield, and in the hilus and stratum moleculare of the dentate gyrus. The majority of labelled axons were varicose, and formed multiple contacts with cell bodies and dendrites of calbindin D,,- and parvalbumin-immunoreactive non-pyrami- dal cells. GABA immunoreactivity of P. oulgaris leucoagglutinin-labelled axons and their postsynaptic targets was investigated by sectioning varicose axon segments for correlated light and electron microscopy, and processing alternate ultrathin sections for postembedding immunogold staining for GABA. All P. oulguris leucoagglutinin-labelled boutons examined were GABA-immunoreactive and the majority of them formed symmetrical synapses with GABA-immunoreactive cell bodies and dendrites. The results demonstrate that a GABAergic septohippocampal pathway exists in the monkey, and, similar to the rat, terminates on different types of GABAergic neurons, including the parvalbumin- and calbindin D,,,-containing non-pyramidal cells. Studies of the human and monkey hippocampal electroencephalogram (EEG) revealed a considerable difference between primates and subprimates in the pattern and behavioural correlation of the EEG.3,‘8 The most prominent difference is the rare occurrence of rhythmic slow activity (RSA, or theta activity) in the primates compared to the lower mammalian species where theta activity can be detected in corre- lation with certain behaviours, like running, walking, rearing, manipulation of objects and during the para- dox phase of sleep. ‘Og4’ Although there are reports about some forms of theta activity related to particu- lar hippocampal function in primates,3*Ms45 a close correlation of theta activity with the ongoing be- haviour has not yet been established.” However, lesion studies and human pathological cases suggest that the basic functions of the hippocampus are likely to be similar throughout the species from rodents to primates. 5,14,18,23,46,47,51 $To whom correspondence should be addressed. Abbreviations: ABC, avidin-biotinylated horseradish per- oxidase complex; BSA, bovine serum albumin; CaBP, calbindin Dzsl;; DAB, 3,3’-diaminobenxidine; DG, den- tate gyrus; EEG, electroencephalogram; ms-dbB, medial septum-diagonal band of Broca complex; NGS, normal goat serum; PB, phosphate buffer; PHAL, Phaseolus vulgaris leucoagglutinin; RSA, rhythmic slow activity; TBS, Tris-buffered saline. The role of the medial septal area in the generation of hippocampal theta activity in rodents is well established.10~38~44~4s~47 The finding that rhythmic activity can be elicited in the monkey hippocampus by septal stimulation’* suggests that the septal area may be able to efficiently control large populations of principal cells as is also reflected by EEG patterns in primates. The septohippocampal projection medi- ating this effect’ is also partly cholinergic in the monkey.29*” In the rat the non-cholinergic com- ponent2s49 of the septohippocampal pathway was shown to be largely GABAergic,30 and to selectively innervate different types of GABAergic interneurons in the hippocampal formation.“*25 These interneurons are known to control large populations of principal cells through their extensive local axon arborizations, and thereby could serve as ideal targets for subcorti- cal pathways, which exercise a global control over hippocampal activity.“i2~L5~22*38~47*48 The question arises as to whether differences in behaviour-associated EEG patterns between primate and subprimate species is paralleled by differences in the connectivity or neurotransmitter composition of the septohippocampal projection. In particular, the goals of this primate study were to determine (i) whether GABAergic neurons also participate in the septohippocampal projection in addition to the choline&c component, and (ii) whether the termin- ation of this projection is similar to that found in the rat. 381

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Neuroscience Vol. 41, NO. 2/3, pp. 381-390, 1991 Printed in Great Britain
0306-4522/91 $3.00 + 0.00 Pergamon Press plc
0 1991 IBRO
SEPTAL GABAERGIC NEURONS INNERVATE INHIBITORY INTERNEURONS IN THE HIPPOCAMPUS OF
THE MACAQUE MONKEY
A. I. GULY&* L. SERW,~ K. T&H,* L. ACSADY,* M. ANTAL~ and T. F. FREIJND*$
*Department of Functional Neuroanatomy, Institute of Experimental Medicine, Hungarian Academy of Sciences, Budapest, P.O. Box 67, H-1450, Hungary; and 1st Department of Anatomy,
Semmelweis University, Medical School, Budapest, H-1450, Hungary tDepartment of Physiology, University Medical School, Pets, H-7624, Hungary
SDepartment of Anatomy, University Medical School, Debrecen, H-4012, Hungary
Abstract-The septohippocampal projection was visualized in three Mucaca mulatta monkeys by anterograde transport of Phaseolus oulgarb leucoagglutinin. Following injections of the lectin into the medial septal nucleus, P. &garb leucoagglutinin-labehed fibres were found in the hippocampal complex, mainly in stratum oriens of the CA1 subfield, throughout the CA3 subfield, and in the hilus and stratum moleculare of the dentate gyrus. The majority of labelled axons were varicose, and formed multiple contacts with cell bodies and dendrites of calbindin D,,- and parvalbumin-immunoreactive non-pyrami- dal cells. GABA immunoreactivity of P. oulgaris leucoagglutinin-labelled axons and their postsynaptic targets was investigated by sectioning varicose axon segments for correlated light and electron microscopy, and processing alternate ultrathin sections for postembedding immunogold staining for GABA. All P. oulguris leucoagglutinin-labelled boutons examined were GABA-immunoreactive and the majority of them formed symmetrical synapses with GABA-immunoreactive cell bodies and dendrites.
The results demonstrate that a GABAergic septohippocampal pathway exists in the monkey, and, similar to the rat, terminates on different types of GABAergic neurons, including the parvalbumin- and calbindin D,,,-containing non-pyramidal cells.
Studies of the human and monkey hippocampal electroencephalogram (EEG) revealed a considerable difference between primates and subprimates in the pattern and behavioural correlation of the EEG.3,‘8 The most prominent difference is the rare occurrence of rhythmic slow activity (RSA, or theta activity) in the primates compared to the lower mammalian species where theta activity can be detected in corre- lation with certain behaviours, like running, walking, rearing, manipulation of objects and during the para- dox phase of sleep. ‘Og4’ Although there are reports about some forms of theta activity related to particu- lar hippocampal function in primates,3*Ms45 a close correlation of theta activity with the ongoing be- haviour has not yet been established.” However, lesion studies and human pathological cases suggest that the basic functions of the hippocampus are likely to be similar throughout the species from rodents to primates. 5,14,18,23,46,47,51
$To whom correspondence should be addressed. Abbreviations: ABC, avidin-biotinylated horseradish per-
oxidase complex; BSA, bovine serum albumin; CaBP, calbindin Dzsl;; DAB, 3,3’-diaminobenxidine; DG, den- tate gyrus; EEG, electroencephalogram; ms-dbB, medial septum-diagonal band of Broca complex; NGS, normal goat serum; PB, phosphate buffer; PHAL, Phaseolus vulgaris leucoagglutinin; RSA, rhythmic slow activity; TBS, Tris-buffered saline.
The role of the medial septal area in the generation of hippocampal theta activity in rodents is well established.10~38~44~4s~47 The finding that rhythmic activity can be elicited in the monkey hippocampus by septal stimulation’* suggests that the septal area may be able to efficiently control large populations of principal cells as is also reflected by EEG patterns in primates. The septohippocampal projection medi- ating this effect’ is also partly cholinergic in the monkey.29*” In the rat the non-cholinergic com- ponent2s49 of the septohippocampal pathway was shown to be largely GABAergic,30 and to selectively innervate different types of GABAergic interneurons in the hippocampal formation.“*25 These interneurons are known to control large populations of principal cells through their extensive local axon arborizations, and thereby could serve as ideal targets for subcorti- cal pathways, which exercise a global control over hippocampal activity.“i2~L5~22*38~47*48
The question arises as to whether differences in behaviour-associated EEG patterns between primate and subprimate species is paralleled by differences in the connectivity or neurotransmitter composition of the septohippocampal projection. In particular, the goals of this primate study were to determine (i) whether GABAergic neurons also participate in the septohippocampal projection in addition to the choline&c component, and (ii) whether the termin- ation of this projection is similar to that found in the rat.
381
3x2 A. I. Gu1.rAs (‘I 01
EXPERIMENTAL PROCEDURES
Animals and surgical procedures
Three adult (four years, 5.5 kg) female Macaca mulatta monkeys (from the colony of University Medical School, WCS, Hungary) were used for the experiments. For the operation the monkeys were anaesthetized with an intra- muscular injection of a cocktail of IOmgjkg ketamine, 1 mg/kg diazepam, and 0.5 mg/kg lidocain chloride. Sup- plementary doses were given approximately every 45 min. The projection from the medial septum-diagonal band of Broca complex (ms-dbB) to the hippocampus was visualized by iontophoretic injection of an anterograde tracer, Phase- 01u.s vulgaris leucoagglutinin (PHAL; 2.5%, Vector).” The coordinates of the injections were calculated from atlases based on M. fuscata and M. mulatta brains.““’ The ms-dbB region was injected at 18 sites along six tracks (at 17, 19 and 2 1 mm rostra1 to the earbar; 0 mm and 1 mm lateral to the midline) at three dorsoventral coordinates (20-24 mm below the level of the dura). PHAL was iontophoretically applied for 10 min at each site with 5.0 PA direct current (7 s onoff cycle).r4
The monkeys survived for seven (monkey 1) or 10 days (monkeys 2 and 3) and then were anaesthetized with the same anaesthetic and perfused through the heart with 500 mI saline (3-4 min) followed by 4 1 Iixative containing 1% ghrtaraldehyde, 2.5% paraformaldehyde and 0.2% picric acid in 0.1 M phosphate buffer (PB; pH 7.4):’ Blocks of the septal area, the hippocampal complex of both hemi- spheres and the entorhinal cortex were dissected and trans- ferred into 10 and 20% sucrose until they sank. The blocks were then freeze-thawed in liquid nitrogen, sectioned on a Vibratome (80-pm septal sections, 60-pm hippocampal sections) and washed in several changes of 0.1 M PB.
lmmunocytochemisrry
Double immunostainingW was used to visualize the PHAL-labelled septohippocampal fibres and the calcium- binding protein-containing cells (calbindin D,,, and parv- albumin) in the same hippocampal sections. In order to enhance the immunoreactivity for the proteins the sections were treated with 1% NaBH, (dissolved in distilled water) for 30 min followed by several washes in 0.1 M PB. First the sections were incubated in a mixture of the primary anti- bodies for two days; biotinylated goat anti-PHAL (1: 200, Vector) was mixed either with rabbit anti-parvalbumin (1: 1000, code R 301) or rabbit anti-calbindin (1: 1000, codes R 8701 and R202). 6~11 The second layer (overnight) was a mixture of avidin-biotinylated horseradish peroxidase complex (ABC; 1: 100, Vector) and goat anti-rabbit IgG (1: 50, ICN). The PHAL-labelled axons were visualized with a 3,3’-diaminobenzidine (DAB) reaction intensified by ammonium-nickel sulphate (deep blue to black reaction product). The third layer was rabbit peroxidaseanti- peroxidase complex (1: 100, DAKOPATT; overnight). The calcium-binding protein-containing cells were visualized with DAB alone, giving a brown end-product. For the dilution of the antisera and for all the washes 50mM Tris-buffered saline (TBS; pH 7.4) containing 1% normal goat serum (NGS) was used. In the case of sections pro- cessed for light microscopy the solutions contained 0.5% Triton X-100 to enhance the penetration of the antibodies.
The sections processed for light microscopy were thoroughly washed, mounted on gelatine-coated slides, de- hydrated and covered with XAM neutral medium. Sections prepared for electron microscopy were treated with 1% 0~0, (in PB) for 1 h, dehydrated, and embedded in Durcu- pan (ACM, Fluka). From sections immunostained only for PHAL, areas containing labelled axons were re-embedded
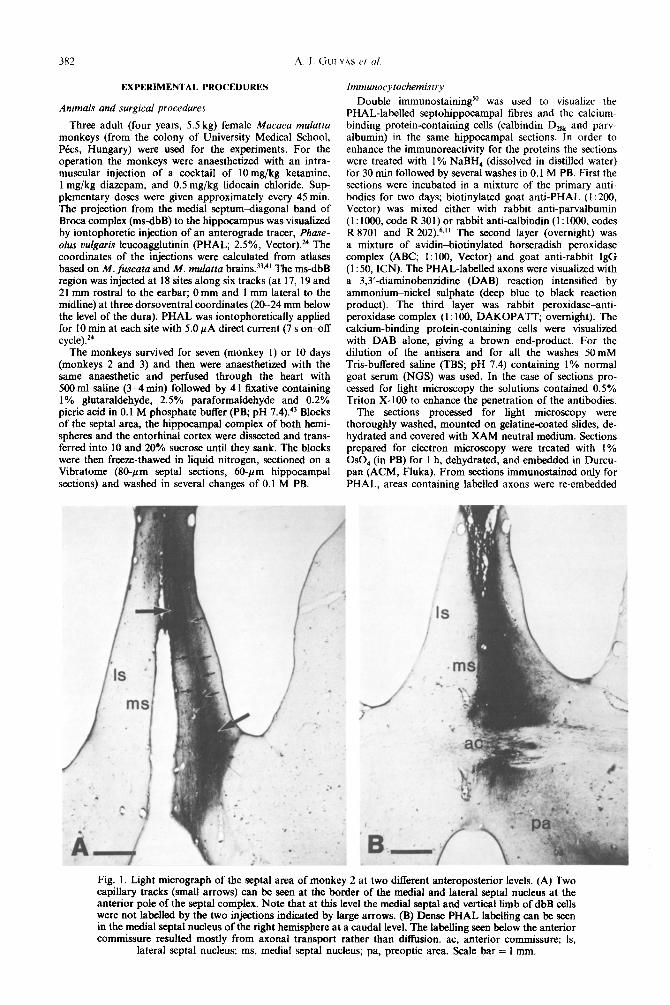
Fig. 1. Light micrograph of the septal area of monkey 2 at two different anteroposterior levels. (A) Two capillary tracks (small arrows) can be seen at the border of the medial and lateral septal nucleus at the anterior pole of the septal complex. Note that at this level the medial septal and vertical limb of dbB cells were not 1abeIled by the two injections indicated by large arrows. (B) Dense PHAL labelhng can be seen in the medial septal nucleus of the right hemisphere at a caudal level. The labelling seen below the anterior commissure resulted mostly from axonal transport rather than diffusion. ac, anterior commissure; Is,
lateral scptal nucleus; ms, medial septal nucleus; pa, preoptic area. Scale bar = 1 mm.

Septal innervation of hippocampal interneurons in monkey 383
Fig. 2. Light ~cro~aph of ~pp~ampal sections double-steno for pa~lbu~n IpV, brown) and PHAL (black). (A) Several large varicosities (arrows) of a PHAL-immunoreactive septohippocampal ftbre terminate on the proximal dendrites and cell body of a parvalbumin-positive non-pyramidal cell in stratum oriens of the CA3 region. (B) Three p~~b~in-imm~or~ctive Nan-pyramidal cells are contacted by thin ~~tohipp~arn~l axons studded with several small boutons (arrows) in stratum oriens of the CA1
region. Scale bars = 20 pm,
for ultra~n sectioning. Alternate sections were cut and mounted on copper and nickel grids, and the latter grids were processed for immunogold staining for GABA.
Sections of the septal area were processed for PHAL i~un~yt~he~stry alone, to check for the location and spread of the PHAL injection sites.
The immunogold staining procedure followed that de- scribed by Somogyi and Hodgsor? with small modih- cations, using a well~haracte~z~ antiserum against GABA.” The steps were carried out on droplets of Milli- pore-filtered solutions in humid Petri dishes, as follows: 1% periodic acid (H,IO,) for 10 min; wash by dipping in several changes of double~istill~ water; 2% sodium metaperio- date (NaIO,, BDH) for lo-30min; wash as before: three times 2 mitt’ in TBS (pH 7.4); 30 min in 1% ovalbumin dissolved in TBS; three times 10 mitt in TBS containing 1% NGS; l-2 h in a rabbit anti-GABA antiserum (code no. 9, diluted 1:IOOO in NGS-TBS);” two times 10 min TBS; 10 min in 50 mM Tris buffer (DH 7.41 containine 1% bovine serum albumin (BSA) and O.jblo Tween 20; go2 anti-rabbit IgG-coated colloidal gold (15 nm, Janssen) for 2 h {dilute 1:lO in the same buffer); two times 5-min wash in double- distilled water; saturated uranyl acetate for 30 min; wash in four changes of double-distills water; staining with lead citrate; wash in distilled water.
RESULTS
Injection sites
Examination of the PHAL~i~~unostain~ sec- tions of the septal area revealed that the injections were most successful in monkey 2. In the rostra1 part of the septal area the injections were restricted to the
right hemisphere, and included the lateral part of the medial septal nucleus and the lateral septal nucleus (Fig. 1A). More caudally the injected area involved the medial septal nucleus of the right he~sphere with negligible spread of the tracer to the other hemisphere (Fig. 1B). PHAL-la~ll~ cells were found in the core of the injection, surrounded by homogeneous PHAL immunoreactivity. In monkey 1 PHAL was iontophorized mainly into the lateral septal nucleus, with a small PHAL billing in the lateral part of the media1 septal nucleus. In monkey 3 the injection tracks passed through the lateral septal nucleus, close to the lateral ventricle, and one track through the ventricular edge of the caudate nucleus.
Distribution of Phaseolus vulgaris ~eucooggiut~~i~- ~obeI~e~ sepal axons in the hi~poc~~~
In monkeys I and 2 the density of labelled axons in the hip~ampal fo~ation was less than that visualized with similar techniques in the rat.z1~25 The PHAL-positive fibres showed an uneven distribution along the lon~tudinal axis of the ~pp~amp~s. In monkey 1, which survived for seven days after the operation and where only a few neurons of the medial septal area were su~ssfully injected with PHAL, substantially fewer labelied fibres were found than in monkey 2, which survived for 10 days, and the PHAL injection was more on target. In monkey 3, in which the injections were in the lateral septal area and the caudate nucleus, no PHAL-labelled fibres were found in any regions of the hippocampal formation.

384 A. 1. GJLYAS P/ 01.
PHAL-labelled axons were frequently seen in stra- tum moleculare, stratum granulosum and the hilus of the dentate gyrus, in all layers of the CA3 region and in stratum oriens of the CA2 and CA1 regions. The highest density of labelled axons was seen in stratum oriens of the CA3-CAl region and in the alveus.
The PHAL-labelled axons were varicose and frequently formed baskets around cell bodies (Figs 2, 3A), or followed single dendrites for over IOOpm. With regard to their light microscopic fea- tures, the fibres were heterogeneous, some of them were thick with large varicosities (Fig. 2A), while others were thin, had less branches studded with small boutons and took a rather straight course in the neuropil (Fig. 2B). The distinction between the two types was not as obvious as in the rat,22.25,37 and axons having intermediate properties were also frequently encountered. Axons with large boutons occurred most frequently in stratum oriens and alveus of CAl-CA3 regions where they usually surrounded cell bodies of interneurons (see below). Axons with small varicosities were more frequent in the molecular layer of the dentate gyrus.
Distribution of parvalbumin - and calbindin -immune - reactive cells
The overall distribution of cells containing these calcium-binding proteins was similar to that de- scribed in the rat hippocampus.6,s’.40 Parvalbumin immunoreactivity was present exclusively in inter- neurons (i.e. non-principal cells), in the granule cell layer and hilus of the dentate gyrus (DG), as well as in stratum pyramidale and stratum oriens of the CAI-CA3 regions. A small number of parvalbumin- positive cells were also seen in stratum moleculare of the DG. Calbindin D,,, (CaBP) immunoreactivity was found in all granule cells and in a large pro- portion of pyramidal cells in the CA1 region. Pyrami- dal cells of the CA3 subfield were always negative for CaBP. Interneurons immunoreactive for CaBP were found mainly in stratum oriens and stratum radiatum of the CA3 region, in stratum oriens of the CA1 region and occasionally in the stratum molecu- lare of the dentate gyrus. The only major difference from the rat with respect to the distribution of these cells was the sparse occurrence of CaBP-positive interneurons in stratum radiatum of the CA1 region. A detailed description of the distribution and types of
parvalbumin- and CaBP-immunoreactive inter- neurons in the monkey will be dealt with separately.3”
Termination of septohippocampal axons on identified interneurons
The examination of hippocampal sections double- stained for PHAL and one of the calcium binding proteins revealed that both parvalbumin- and CaBP- immunoreactive cells were among the targets of the septohippocampal projection. Septohippocampal axons formed multiple contacts (five to 17 boutons per cell) with cell bodies, proximal and distal den- drites of mostly parvalbumin-positive but also of CaBP-positive neurons (Fig. 2). Due to the low density of PHAL-labelled fibres it was impossible to give a quantitative account of the ratio of the con- tacted parvalbumin- and CaBP-immunoreactive cells in the different regions. Contacted cells were found in all layers and regions of the hippocampus where parvalbumin- and CaBP-immunoreactive non- pyramidal cells occurred. The most frequent targets were the parvalbumin-immunoreactive cells in stra- tum oriens of the CA3 area (Fig. 2A). Neurons embedded among myelinated axons in the alveus were also often innervated by septal axons.
To provide direct evidence that septohippocampal axons form synaptic contacts with GABAergic non- pyramidal cells in the hippocampus and that the afferents themselves are also GABAergic, PHAL- labelled varicose axons from stratum oriens and radiatum of the CA3 region were serially sectioned for correlated light and electron microscopy. Alternate grids were processed for postembedding immunogold staining for GABA and examined in the electron microscope. Both thick (Fig. 3) and thin (Fig. 4) axons were found to be immunoreactive for GABA, and to establish symmetrical synapses on somata (Fig. 3) and on dendritic shafts (Fig. 4). The cell bodies postsynaptic to the labelled axons were always GABA-immunoreactive (three somata for the thin axons and six for the thick axons), and received four to 11 synapses per cell from septal boutons (Fig. 3). Somata of the contacted cells showed charac- teristic features of non-pyramidal cells, e.g. several mitochondria and polyribosomes in the cytoplasm, invaginated nucleus occasionally containing an intra- nuclear rod, and a dense somatic input. The postsyn- aptic dendrites often received synaptic contacts from
Fig. 3. (A) Boutons (b,_,) of a PHAL-labelled septohippocampal axon surround a cell body (S) in stratum oriens of the CA3 region. (B) Low power electron micrograph of the cell shown at the light microscopical level in A. The cell (S) contacted by three boutons (b,_,, arrows) in this section shows the characteristic features of non-pyramidal cells. Synapses formed by boutons b, and b, are shown at higher magnification in C and D. (C) Bouton b, (marked in A), shown here at high power, forms a symmetric synaptic contact (arrow) with the soma of the cell, shown in A and B. (D,E) Adjacent ultrathin sections of a PHAL-labelled bouton (b,, framed in B) which is in symmetrical synaptic contact (arrows) with the cell body shown in B. The section in E has been immunostained for GABA using the immunogold procedure. Both the ccl1 body (S) and the PHAL-labelled bouton (b,) were GABA immunoreactive, as indicated by the accumulation of colloidal gold particles. Other GABA-positive profiles (asterisks) and GABA-negative boutons with round synaptic vesicles (stars) are also indicated. Scale bars = 10 pm in A; 2 pm in B;
0.25pm in C. D, E.

Fig. 3.

Fig.
4.
(A
) A
den
driti
c sh
aft
from
st
ratu
m
orie
ns
of
the
CA
3 re
gion
is
in
sym
met
rioa
l sy
napt
ic
cont
act
(arr
ow)
with
a
thin
PH
AL
-lab
elle
d se
ptoh
ippo
cam
pal
axon
. (B
) B
oth
the
post
syna
ptic
de
ndri
te
(d oA
BA
) and
the
PHA
L-l
abel
led
bout
on
are
imm
unor
eact
ive
for
GA
BA
as
sho
wn
on
this
im
mun
ogol
d-st
aine
d se
ctio
n,
adja
cent
to
tha
t se
en i
n A
. T
wo
othe
r G
AB
A-i
mm
unor
eact
ive
term
inal
s ar
e sh
own
(ast
eris
ks),
on
e of
the
m
is
in c
onta
ct
with
the
den
drite
. G
AB
A-n
egat
ive
bout
ons
(sta
rs)
are
mak
ing
asym
met
rica
l sy
naps
es,
and
serv
e to
ind
icat
e th
e le
vel
of b
ackg
roun
d st
aini
ng.
Scal
e ba
rs =
0.2
5 pm
.

Septal innervation of hippocampal intemeurons in monkey 387
several other unlabelled boutons. In the case of the thick axons the majority of postsynaptic dendrites were GABA-immunoreactive (15 out of 18). The ratio of GABA-immunoreactive postsynaptic den- drites was somewhat lower for the thin axons (three out of six), although the sample in this case is too small due to the sparse occurrence of thin axons and synaptic varicosities along them.
DISCUSSION
In the present study we demonstrated (i) the existence of a GABAergic septohippocampal pro- jection in the monkey, and (ii) that synaptic boutons of this projection terminate preferentially on GABA- ergic interneurons-including the parvalbumin- and CaBP-containing non-pyramidal cells-f the hippo- campal formation. Thus, we found no qualitative difference between the rat and the monkey with respect to the synaptic organization of the GABA- ergic septohippocampal pathway.
Technical considerations
When evaluating results obtained by anterograde transport we have to consider the possibility of labelling fibres of passage arising from different sub- cortical structures and passing through the septal area (e.g. those from the raphe nuclei and locus coeruleus).
PHAL is known as an anterograde tracer if applied iontophoretically,” but it can be transported by fibres of passage as well. ” However, in the latter case the labelling was found to be of shorter range, the axons were few in number and were relatively weakly stained. The length of the septohippocampal pathway in the monkey is several centimetres long, thus the possibility of labelling fibres that originate outside the septal area is minimal.
After injection of PHAL into the septal area we found a relatively small number of labelled fibres in the hippocampus of the monkeys, compared to our previous experiments in the rat.“*25 Furthermore, the labelled fibres were unevenly distributed along the longitudinal axis of the hippocampus; some sec- tions contained no labelled axons at all. A possible reason for the weak anterograde labelling is that the axonal transport of PHAL is relatively slow, only 4-6 mm/day,24 and in the monkey the septo- hippocampal pathway is much longer than in the rat. Longer survival times may result in better labelling, but the possibility of degeneration-induced early neurochemical changes will have to be considered when doing immunocytochemistry on the labelled axons. The fact that we found an uneven labelling along the longitudinal axis of the hippocampus after injection of PHAL into the medial septal area suggests that the projection to the hippocampus is topographical in the monkey, as described in the rat by Nyakas et al.” Our sample is, however, far too
small to make any conclusions about the possible topography of the septohippocampal projection.
In monkey 3, in which the lateral septum was injected by PHAL, there were no labelled axons at any anteroposterior levels, or in any subfields of the hippocampus. This confirms previous data that there is no projection from the lateral septal nuclei into the hippocampal formation.’ We found no evidence either for a projection from the lateral septum to the medial septum in this animal.
Identity of the Phaseolus vulgaris leucoagglutinin- labeled jibres
The majority of the neurons participating in the projection from the ms-dbB region to the hippo- campus was shown to be choline@ both in the rat2.49 and in the monkey.29su In the monkey, this path- way arises from the medial septum where lo%, and from the vertical limb of the dbB where 70%, of the neurons are immunoreactive for choline acetyltrans- ferase.29*” Both small and large neurons of these areas were shown to project to the hippocampal formation;’ however, the choline acetyltransferase- positive cells are generally small in size?9s” This suggested that-similar to the rat2~49-non-cholin- ergic neurons also participate in this pathway. In the rat, the non-choline@ component of the septo- hippocampal pathway was shown to be largely GABAergic,” and to represent the most target selec- tive pathway so far reported in the brain, i.e. it selectively innervates GABAergic interneurons in the hippocampus and DG. 22*25 In the present study we provide direct evidence that such a GABAergic projection from the septal area to the hippocampal formation also exists in the macaque monkey. However, the finding that most, if not all, of the PHAL-labelled axons in the present study were GABA-immunoreactive is somewhat unexpected and requires discussion. A likely explanation is that the cholinergic component-which is likely to be the numerically dominant component-of the septo- hippocampal pathway was not labelled by PHAL. A careful examination of the injection sites suggests that anterograde labelling took place largely, if not exclusively, from neurons of the medial septal nucleus, where only 10% of the neurons are cholin- ergic, as opposed to the vertical limb of the dbB, where they represent over 70% of the projecting neurons.34 In addition, the haemorrhage along the midline of the medial septum caused by the injection probably reduced the number of anterogradely labelled fibres, and this may have critically effected the cholinergic axons gathering in this region before going to the hippocampus. The course taken by the GABAergic pathway is not known, although it is unlikely to be different. A preferential transport of the lectin by GABAergic neurons has been suggested for the septohippocampal neurons in the rat,21*u and for GABAergic basket cells in the cat neo- cortex (Kisvirday, Gulyas, Freund and Somogyi,

388 PI. I. GULYAS et (II
unpublished observations). This preferential labelhng may be explained by the relatively large diameter of the GABAergic axons, and/or a preferential binding and/or uptake of PHAL by GABAergic neurons as shown for Viciu uillosa lectin.3” The septohippocam- pal pathway in the monkey is very long; therefore the large diameter GABAergic fibres may have a great advantage in transporting sufficient amount of lectin to the hippocampus to reach the threshold for immunocytochemical detection.
The possibility that some of the GABA-immuno- reactive septal afferents may contain acetylchohne has also to be considered. In the cat neocortex Beaulieu et ~1.‘~~ demonstrated that 8% of the choline acetyltransferase-immunoreactive axon ter- minals were positive for GABA. If such coexistence of GABA and acetylcholine would exist in some septohippocampal axons of the monkey, it could account for the observed anatomical heterogeneity of PHAL-labelled fibres in our material. The rather sparse, thin axons share some characteristics of the type II, possibly chohnergic, axons described in the rat 22,25.37 for example; they also contacted GABA- negative dendrites belonging to presumed pyramidal cells (three out of six boutons). Thus, at least some of these axons may contain both GABA and acetyl- choline. Immunocytochemical co-localization studies are required to determine the exact proportion of terminals (or neurons) containing both acetylcholine and GABA.
The third possibility, namely that our GABA antiserum would recognize something other than GABA in the PHAL-labelled terminals, can be re- jected on the basis of our earlier and present control experiments. The solid phase adsorption of the anti- serum to GABA resulted in the disappearance of specific staining all over the tissue, including the PHAL-labelled terminals.2’~22 No staining was found either when sections were incubated with normal rabbit serum (1: 500) instead of the primary anti- serum. In the double-labelled terminals (PHAL- and GABA-positive) the gold particles were usually con- centrated in the mitochondria, similar to GABA- immunoreactive profiles, which did not stain for PHAL (see also Refs 21, 22, 25). However, the lectin or the immunoreaction end-product of the pre- embedding staining does not penetrate mitochondria; thus the possibility that our GABA antiserum cross- reacts with certain epitopes of the pre-embedding immune complex can be excluded.
Septohippocampal axons innervate parvalbumin- and calbindin D,,,-immunoreactive non -pyramidal cells
The distribution of parvalbumin- and CaBP-con- taining neurons in the hippocampus is characteristic and only partially overlaps, as demonstrated in the rat6.26,3’,4” and in our recent work in the monkey.39 Anatomical studies in rats have shown that the parvalbumin- and CaBP-immunoreactive non- pyramidal neurons are GABAergic,‘6*31 and form two
separate cell populations.‘9,26 However, CaBP is also present in two populations of principal cells, in the dentate granule cells and in “superficial layer” pyra- midal cells of the CA1 region. In the hippocampus” and neocortexr6 parvalbumin is present exclusively in interneurons; in particular, in the axoaxonic cells and the basket cells.28 The extensive axon arborizations of these interneurons terminate on the axon initial segments and in the perisomatic region of large populations of principal cells (pyramidal and gran- ule cells), probably exerting a powerful inhibition of the cells’ firing. The septohippocampal pathway is able, by establishing multiple contacts with these non-pyramidal cells, to gain control over large populations of principal cells.
Functional implications
The synaptic organization of the GABAergic sep- tohippocampal projection in the monkey described in the present study is consistent with a possible dis- inhibitory function proposed earlier in the rat.9.22*25.32 During different behavioural states associated with different EEG patterns, the GABAergic septo- hippocampal pathway may play a crucial role in the synchronization or desynchronization of large popu- lations of principal cells via the inhibition of the parvalbumin- and CaBP-containing interneurons. This concept is favoured by the observation that stimulation of the septal region (or of fibres passing through that area) elicits population synchrony in the monkey hippocampus.‘*
Our study was initiated by the observed difference between the primate and subprimate EEG, mainly with respect to theta activity. The present results demonstrate that there is a great similarity between the rat and the monkey in the synaptic organization of the GABAergic septohippocampal pathway. Thus, other factors, such as differences in the postsynaptic targets of the cholinergic component, or differences in the firing pattern of septohippocampal neurons in relation to behaviour, may have to be considered. One may speculate that-as suggested by the pres- ent anatomical and earlier electrophysiological studies’*-the septohippocampal pathway in the monkey is capable of producing a powerful disin- hibitory and/or feed-forward inhibitory effect similar to that in the rat,9,‘2,‘5.20~32 but the pattern of pace- maker neurons active at any one time-point may be more specific in its behaviour dependence during theta-associated behaviours. Consequently, its global effect on hippocampal electrical activity patterns may be more subtle and specific in space and time.
Sharp waves are known to be present both in the primate and subprimate EEG,3,‘8 and are likely to be the result of a rapidly synchronizing population burst of CA3 pyramidal cells. 13,15 Recruiting via the exten- sive recurrent collateral system of CA3 pyramidal cells is under a powerful local inhibitory control.35 One may speculate that the GABAergic septo- hippocampal pathway is responsible for disinhibiting

Septal innervation of hippocampal interneurons in monkey 389
recurrent excitatory circuits in the CA3 region, which calbindin Dzak. to Dr P. Somogyi (MRC Anatomical Neuro-
may lead to rapid synchronization and the occurrence pharmacology Unit, Oxford, U.K.) for the antiserum
of sharp waves. against GABA, and to Dr G. Buzs.&ki for helpful discussions and comments on the manuscrint. We also thank Drs Zs. Horvath, G. Jando and L. L&rd for their help with the monkey surgery, and MS K. Boczko and MS M. Paleszter
Acknowledgements-We are grateful to Dr K. G. for excellent technical assistance. This study was supported Bairnbridge (University of British Columbia, Vancouver, by grants from the Hungarian Academy of Sciences and the Canada) for a gift of antisera against parvalbumin and FIDIA Ischemia Research Fund.
REFERENCES
1. Amaral D. G. and Cowan W. M. (1980) Subcortical afferents to the hippocampal formation in the monkey. J. camp. Neural. 189, 573-591.
2. Amaral D. G. and Kurz J. (1985) An analysis of the origins of the cholinergic and noncholinergic septal projections to the hippocampal formation of the rat. J. camp. Neurol. 240, 37-59.
3. Amolds D. E. A. T., Lopes da Silva F. H., Aitnik J. W., Kamp A. and Boejinga P. (1980) The spectral properties of hippocampal EEG related to behavior in man. Electroenceph. clin. Neurophysiol. SO, 324-328.
4. Assaf S. Y. and Miller J. J. (1978) The role of a raphe scrotonin system in the control of septal unit activity and hippocampal desynchronization. Neuroscience 3, 539-550.
5. Bachevalier J., Saunders R. C. and Mishkin M. (1985) Visual recognition in monkeys: effects of transection of fomix. Expl Brain Res. 57, 547-553.
6. Baimbridge K. G., Miller J. J. and Parkes C. 0. (1982) Calcium-binding protein distribution in the rat brain. Brain Res. 239, 519-525.
7. Bcaulieu C. and Somogyi P. (1989) Neurochemical properties and postsynaptic targets of cholinergic synapses in cat visual cortex. Sot. Neurosci. Abstr. 15, 1107.
8. Beaulieu C. and Somogyi P. (1991) Enrichment of choline@ synaptic terminals on GABAergic neurons and coexistence of immunoreactive GABA and choline acetyltransferase in the same synaptic terminals in cortex. Eur. J. Neurosci. (in press).
9. Bilkey D. K. and Goddard G. V. (1985) Medial septal facilitation of hippocampal granule cell activity is mediated by inhibition of inhibitory interneurons. Brain Res. 361, 99106.
10. Bland B. H. (1986) The physiology and pharmacology of hippocampal formation theta rhythms. Prog. Neurobiol. 26, l-54.
Il. Buchan A. M. J. and Bairnbridge K. G. (1988) Distribution and co-localization of calbindin D-28k with VIP and neuropcptide Y but not somatostatin, galanin and substance P in the enteric nervous system of the rat. Peptides 9, 333-338.
12. Buzsaki G. (1984) Feed-forward inhibition in the hippocampal formation. Prog. Neurobiol. 22, 131-153. 13. Buzsdki G. (1986) Hippocampal sharp waves: their origin and significance. Brain Res. 398, 242-252. 14. Buzslki G. (1989) Two-stage model of memory trace formation: a role for “noisy” brain states. Neuroscience
31, 551-570. 15. Buzsaki G., Leung L.-W. S. and Vanderwolf C. H. (1983) Cellular bases of hippocampal EEG in the behaving rat.
Brain Res. Rev. 6, 139-171. 16. Celio R. M. (1986) Parvalbumin in most r-aminobutyric acid containing neurons of the rat cerebral cortex. Science
231, 995-997. 17. Cliffer K. D. and Giesler G. J., Jr (1988) PHA-L can be transported anterogradely through fibers of passage. Brain Res.
458, 185-191. 18. Crowne D. P. and Radcliffe D. D. (1975) Some characteristics of functional relations of the electrical activity of the
primate hippocampus and hypotheses of hippocampal function. In The Hippocampus (eds Isaacson R. R. and Pribram K. H.), Vol. 2, pp. 185-206. Plenum Press, New York.
19. Dgnos P., Freund T. F. and Frotscher M. (1991) Fine structure and synaptic input of rat hippocampal non-pyramidal cells are not related to chemical characteristics. Brain Res. (in press).
20. Fantie B. D. and Goddard G. V. (1982) Septal modulation of the population spike in the fascia dentata produced by perforant path stimulation in the rat. Brain Res. 252, 227-237.
21. Freund T. F. (1989) GABAergic septohippocampal neurons contain parvalbumin. Brain Res. 478, 375-381. 22. Freund F. T. and Antal M. (1988) GABA-containing neurons in the septum control inhibitory interneurons in the
hippocampus. Nature 336, 170-173. 23. Gaffan D.. Saunders R. C.. Gaffan E. A.. Harrison S.. Shields C. and Owen M. J. (1984) Effects of fornix
transsection upon associativememory in monkeys: role of the hippocampus in learned action: Q. i exp. Psychol. 36B, 173-221.
24. Gerfen C. R. and Sawchenko P. E. (1984) An anterograde neuroanatomical tracing method that shows the detailed morphology of neurons, their axons and terminals: immunohistochemical localization of an axonally transported plant lectin, Phaseolus oulgaris leucoagglutinin (PHA-L). Brain Res. 290, 219-238.
25. Gulyis A. I., Gijrcs T. J. and Freund T. F. (1990) Innervation of different peptide-containing neurons in the hippocampus by GABAergic septal afferents. Neuroscience 37, 31-44.
26. Gulyas A. I., T&h K., Danos P. and Freund T. F. (1991) Subpopulations of GABAergic neurons containing parvalbumin, calbindin D,,, and cholecystokinin in the rat hippocampus. J. camp. Neural. (in press).
27 Hodgson A. J., Penke B., Erdei A., Chubb I. V. and Somogyi P. (1985) Antisera to r-aminobutyric acid. I. Production and characterization using a new model system. J. Histochem. Cytochem. 33, 229-239.
28. Katsumaru H., Kosaka T., Heizmann C. W. and Hama K. (1988) Immunocytochemical study of GABAergic neurons containing the calcium-binding protein parvalbumin in the rat hippocampus. Expl Brain Res. 72, 347-362.

390 A.I. Gu~vlis et al
29. Kitt C. A., Mitchell S. J., DeLong M. R., Wainer B. H. and Price L. D. (1987) Fiber pathways of basal forebram cholinergic neurons in monkeys. Brain Res. 406, 192-206.
30. Kdhler C., Chan-Palay V. and Wu J.-Y. (1984) Septal neurons containing glutamic acid decarboxylase immunoreactivity project to the hippocampal region in the rat brain. Anat. Embryol. 169, 41 -44.
31. Kosaka T., Katsumaru H., Hama K., Wu J.-Y. and Heinzmann C. W. (1987) GABAergic neurons containing the Ca*+-binding protein parvalbumin in the rat hippocampus. Bruin Res. 419, 119-130.
32. Krnjevic K., Ropert N. and Casullo J. (1988) Septohippocampal disinhibition. Brain Res. 438, 182-192. 33. Kusama T. and Mabuchi M. (1970) Stereotaxic Arias of the Bruin ofMacaca fuscata. University of Tokyo Press, Tokyo. 34. Mesulam M. M., Mufson E. J., Levey A. I. and Wainer B. H. (1983) Cholinergic innervation of cortex by the basal
forebrain: cytochemistry and cortical connection of the septal area, diagonal band nuclei, nucleus basalis (substantia innominata), and hypothalamus in the rhesus monkey. J. camp. Neural. 214, 170-197.
35. Miles R. and Wong R. K. S. (1987) Inhibitory control of local excitatory circuits in the guinea-pig hippocampus. J. Physiol., Lond. 388, 61 l-629.
36. Nakagawa F., Schulte B. A. and Spicer S. S. (1986) Selective cytochemical demonstration of glycoconjugate-containing terminal N-acetylgalactosamine on some brain neurons. J. camp. Neurof. 243, 280-290.
37. Nyakas C., Luiten P. G. M., Spencer D. G. and Traber J. (1987) Detailed projection patterns of septal and diagonal band efferents to the hippocampus in the rat with emphasis on innervation of CA1 and dentate gyrus. Brain Res. BUN. 18, 533-545.
38. Petsche H., Stumpf C. and Gogolak G. (1962) The significance of the rabbit’s septum as a relay station between the midbrain and the hippocampus. The control of hippocampal arousal activity by septum cells. Electroenceph. clin. Neurophysiol. 14, 202-2 1 I.
39. Seress L., Gulyls A. I. and Freund T. F. (1991) Distribution of parvalbumin- and calbindin D,,,-containing neurons in the primate hippocampus. J. camp. Neural. (in press).
40. Sloviter R. (1989) Calcium-binding protein (calbindin-D,,,) and parvalbumin immunoeytochemistry: localixation in the rat hippocampus with specific reference to the selective vulnerability of hippocampal neurons to seizure activity. J. camp. Neural. 280, 183-196.
41. Snider R. S. and Lee J. C. (1961) A Siereofaxic Atlas of the Monkey Brain (Macaca mulatta). University of Chicago Press, Chicago.
42. Somogyi P. and Hodgson A. J. (1985) Antisera to T-aminobutyric acid. III. Demonstration of GABA in Golgi-impreg- nated neurons and in conventional electron microscopic sections of cat striate cortex. J. His&&em. Cytochem. 33, 249-257.
43. Somogyi P. and Takagi H. (1982) A note on the use of picric acid-paraformaldehyde-glutaraldehyde fixative for correlated light and electron microscopic immunocytochemistry. Neuroscience 7, 1779-1783.
44. Stewart M. and Fox S. E. (1989) Monkeys have hippocampal theta activity. Sot. Neurosci. Abstr. 15, 1250. 45. Stewart M. and Fox S. E. (1989) Do septal neurons pace the hippocampal theta rhythm? Tren& Neurosci. 13, 163-168. 46. Thompson R. F. (1986) The neurobiology of learning and memory. Science 233, 941-947. 47. Vanderwolf C. H. (1969) Hippocampal electrical activity and voluntary movement in the rat. Electroenceph. clin.
Neurophysiol. 26, 407-418. 48. Vanderwolf C. H., Leung L. W. S., Baker G. B. and Stewart D. J. (1989) The role of serotonin in the control of cerebral
activity: studies with intracerebral 5,7dihydroxytryptamine. Bruin Res. 504, 181-191. 49. Wainer B. H., Levey A. I., Rye D. B., MesuIam M. M. and Mufson E. J. (1985) Cholinergic and non-cholinergic
septohippocampal pathways. Neurosci. L&r. 54, 45-52. 50. Wouterlood F. G., Bol J. G. J. M. and Steinbush H. W. M. (1987) Double-label immunocytochemistry: combination
of anterograde neuroanatomical tracing with Phuseolus vulguris leucoaggiutinin and enzyme immunocytoehemistry of target neurons. J. Hisrochem. Cyrochem. 35, 817-823.
51. Zola-Morgan S., Squire L. R. and Amaral D. G. (1986) Human amnesia and the medial temporal region: enduring memory impairment following a bilateral lesion limited to field CA1 of the hippocampus. J. Neurosci. 6, 2950-2967.
(Accepted 4 October 1990)
Related Documents











