Separating subjective emotion from the perception of emotion-inducing stimuli: An fMRI study Amy S. Garrett a, * and Richard J. Maddock b a Stanford University School of Medicine, Department of Psychiatry, 401 Quarry Road, Stanford, CA 94305-5795, USA b University of California, Davis Medical Center, Department of Psychiatry, CA 95814, USA Received 15 April 2005; revised 3 May 2006; accepted 8 May 2006 fMRI was used to dissociate neural responses temporally associated with the subjective experience of emotion from those associated with the perception of emotion-inducing stimuli in order to better define the emotion-related functions of the amygdala, lateral orbital frontal cortex (OFC), and hippocampus. Subjects viewed aversive pictures followed by an extended post-stimulus period of sustained subjective emotion. Brain regions showing activation paralleling the period of sustained subjective emotion were distinguished from those showing activation limited to the period of aversive picture presentation. Behavioral results showed that subjective ratings of emotion remained elevated for 20 s after offset of the aversive pictures. fMRI results showed that viewing aversive pictures activated the amygdala, lateral OFC, and hippocampus. Subjective emotion (present both during and after aversive pictures) was temporally associated with activation in the right lateral OFC and left hippocampus but not the amygdala. Ratings of subjective emotion were correlated with activation in the right lateral OFC and left hippocampus. The results support direct amygdala involvement in emotion perception but suggest that amyg- dala activation is not temporally associated with subjective emotion that occurs after the offset of emotion-related stimuli. The results are consistent with a general role for the lateral OFC in monitoring or reflecting on internal experience and show that hippocampal activation is sustained during a period of subjective emotion, possibly related to enhanced memory encoding for the aversive pictures. D 2006 Elsevier Inc. All rights reserved. Keywords: Emotion; fMRI; Amygdala; Orbitofrontal; Hippocampus; Sustained Introduction The emotional response to an affectively relevant stimulus is comprised of multiple processes, including perception, attention, evaluation, memory, autonomic responses, cognitive responses, behavior, and subjective experience (Cacioppo and Gardner, 1999). While medial temporal, prefrontal, and other brain regions have been consistently implicated in the emotional response, important questions remain about the specific processes they mediate. The various components of an emotional response differ in their temporal characteristics. Emotional processes involving attention and perception are typically limited to the time the evoking stimulus is present, while the subjective experience of emotion, autonomic responses, cognitive responses, and behaviors such as facial expressions are often sustained beyond the offset of the evoking stimulus (Sirota et al., 1987; Bradley et al., 1996; Baker et al., 1997; Pizzagalli et al., 1999; Garrett and Maddock, 2001). These temporal characteristics might be used to identify brain regions mediating these various components. Previously, we showed that subjects report negative subjective emotion while viewing aversive pictures and for about 16 s after the offset of aversive pictures (Garrett and Maddock, 2001). Thus, while picture perception is confined to the duration of picture presentation, subjective emotion can be measured over a relatively sustained period of time. Using this method, we can test whether regions such as the amygdala, OFC, and hippocampus are involved primarily in emotion perception and/or emotional responses that parallel the timecourse of subjective emotional experience. The amygdala appears to be critical for lending affective significance to stimuli during perception (Adolphs and Tranel, 1999a,b; Adolphs and Tranel, 1999a,b; Adolphs et al., 1999; Hamann et al., 1999; Anderson and Phelps, 2001; Amaral, 2002; Amaral et al., 2003), but its role in the more sustained process of subjective emotion is uncertain. Mood and subjective emotional experience are generally unaffected in patients with amygdala lesions (Anderson and Phelps, 2002), suggesting that these functions are independent of the amygdala. In contrast, neuro- imaging studies often show amygdala activation during induced emotion (Phan et al., 2002; Levesque et al., 2003), and increased amgydala activation has been shown in a group of 5 subjects instructed to maintain a negative subjective emotion after picture presentation (Schaefer et al., 2002). However, these studies have not examined the unmanipulated subjective experience of emotion as distinct from the perception of an emotion-inducing stimulus throughout the entire brain. 1053-8119/$ - see front matter D 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.neuroimage.2006.05.024 * Corresponding author. E-mail address: [email protected] (A.S. Garrett). Available online on ScienceDirect (www.sciencedirect.com). www.elsevier.com/locate/ynimg NeuroImage 33 (2006) 263 – 274

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/ynimg
NeuroImage 33 (2006) 263 – 274
Separating subjective emotion from the perception of
emotion-inducing stimuli: An fMRI study
Amy S. Garrett a,* and Richard J. Maddockb
aStanford University School of Medicine, Department of Psychiatry, 401 Quarry Road, Stanford, CA 94305-5795, USAbUniversity of California, Davis Medical Center, Department of Psychiatry, CA 95814, USA
Received 15 April 2005; revised 3 May 2006; accepted 8 May 2006
fMRI was used to dissociate neural responses temporally associated
with the subjective experience of emotion from those associated with
the perception of emotion-inducing stimuli in order to better define the
emotion-related functions of the amygdala, lateral orbital frontal
cortex (OFC), and hippocampus. Subjects viewed aversive pictures
followed by an extended post-stimulus period of sustained subjective
emotion. Brain regions showing activation paralleling the period of
sustained subjective emotion were distinguished from those showing
activation limited to the period of aversive picture presentation.
Behavioral results showed that subjective ratings of emotion remained
elevated for 20 s after offset of the aversive pictures. fMRI results
showed that viewing aversive pictures activated the amygdala, lateral
OFC, and hippocampus. Subjective emotion (present both during and
after aversive pictures) was temporally associated with activation in the
right lateral OFC and left hippocampus but not the amygdala. Ratings
of subjective emotion were correlated with activation in the right
lateral OFC and left hippocampus. The results support direct
amygdala involvement in emotion perception but suggest that amyg-
dala activation is not temporally associated with subjective emotion
that occurs after the offset of emotion-related stimuli. The results are
consistent with a general role for the lateral OFC in monitoring or
reflecting on internal experience and show that hippocampal activation
is sustained during a period of subjective emotion, possibly related to
enhanced memory encoding for the aversive pictures.
D 2006 Elsevier Inc. All rights reserved.
Keywords: Emotion; fMRI; Amygdala; Orbitofrontal; Hippocampus;
Sustained
Introduction
The emotional response to an affectively relevant stimulus is
comprised of multiple processes, including perception, attention,
evaluation, memory, autonomic responses, cognitive responses,
behavior, and subjective experience (Cacioppo and Gardner,
1999). While medial temporal, prefrontal, and other brain regions
1053-8119/$ - see front matter D 2006 Elsevier Inc. All rights reserved.
doi:10.1016/j.neuroimage.2006.05.024
* Corresponding author.
E-mail address: [email protected] (A.S. Garrett).
Available online on ScienceDirect (www.sciencedirect.com).
have been consistently implicated in the emotional response,
important questions remain about the specific processes they
mediate.
The various components of an emotional response differ in their
temporal characteristics. Emotional processes involving attention
and perception are typically limited to the time the evoking
stimulus is present, while the subjective experience of emotion,
autonomic responses, cognitive responses, and behaviors such as
facial expressions are often sustained beyond the offset of the
evoking stimulus (Sirota et al., 1987; Bradley et al., 1996; Baker et
al., 1997; Pizzagalli et al., 1999; Garrett and Maddock, 2001).
These temporal characteristics might be used to identify brain
regions mediating these various components. Previously, we
showed that subjects report negative subjective emotion while
viewing aversive pictures and for about 16 s after the offset of
aversive pictures (Garrett and Maddock, 2001). Thus, while picture
perception is confined to the duration of picture presentation,
subjective emotion can be measured over a relatively sustained
period of time. Using this method, we can test whether regions
such as the amygdala, OFC, and hippocampus are involved
primarily in emotion perception and/or emotional responses that
parallel the timecourse of subjective emotional experience.
The amygdala appears to be critical for lending affective
significance to stimuli during perception (Adolphs and Tranel,
1999a,b; Adolphs and Tranel, 1999a,b; Adolphs et al., 1999;
Hamann et al., 1999; Anderson and Phelps, 2001; Amaral, 2002;
Amaral et al., 2003), but its role in the more sustained process of
subjective emotion is uncertain. Mood and subjective emotional
experience are generally unaffected in patients with amygdala
lesions (Anderson and Phelps, 2002), suggesting that these
functions are independent of the amygdala. In contrast, neuro-
imaging studies often show amygdala activation during induced
emotion (Phan et al., 2002; Levesque et al., 2003), and increased
amgydala activation has been shown in a group of 5 subjects
instructed to maintain a negative subjective emotion after picture
presentation (Schaefer et al., 2002). However, these studies have
not examined the unmanipulated subjective experience of emotion
as distinct from the perception of an emotion-inducing stimulus
throughout the entire brain.

A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274264
In addition to amygdala activation, hippocampal activation is
often reported in studies of emotion, primarily related to enhanced
encoding of emotional stimuli (McGaugh, 2000; Pare, 2003),
through the interaction between the amygdala and hippocampus
(Dolcos et al., 2004) (Richardson et al., 2004). Therefore, we might
predict that hippocampal activation would be parallel to amygdala
activation, that is, limited to the period of perception of emotion-
related stimuli.
A number of brain regions have been proposed to have a role in
the subjective experience of emotion, including the medial and
orbital prefrontal cortices, the insular cortex, and the somatosensory
cortex (Damasio, 1999; Mayberg et al., 1999; Lane, 2000; LeDoux,
2000; Craig, 2002; Hornak et al., 2003; Phillips et al., 2003).
Brodmann area (BA) 47, in the lateral OFC, is the prefrontal region
most commonly activated in studies of emotion (Steele and Lawrie,
2004). This region also has been implicated in emotional and non-
emotional meta-cognitive processes of self-monitoring and self-
reflection and emotion regulation (Hariri et al., 2000; Lieberman et
al., 2004). It has been linked to reflective awareness of subjective
emotion {Lieberman et al., 2004 #121} as well as self-reflection
associated with meta-memory phenomena (Maril et al., 2001; Kikyo
et al., 2002). Previous studies have suggested an association between
the lateral OFC and ratings of subjective responses in several
sensory modalities, including the subjective response to tastes
(Kringelbach et al., 2003; Small et al., 2003), odors (Anderson et al.,
2003), the affective component of pain (Rolls et al., 2003), and sad
feelings induced by watching films (Levesque et al., 2003). Taken
together, these findings suggest that the lateral OFC is activated
when subjects reflect on and assess a variety of subjective
experiences. Studies of emotion suggest that subjective emotion is
often accompanied by spontaneous self-reflection {Schooler, 2002
#129; Lieberman et al., 2004 #121}. If so, then the lateral OFC may
exhibit activation that is temporally associated with the subjective
experience of emotion, that is, observed during the presentation of
aversive pictures and also after the offset of the pictures.
The current fMRI study uses epochs of aversive pictures
followed by extended post-stimulus epochs of subjective emotion
to identify brain activation temporally associated with aversive
picture perception as well as those temporally associated with the
experience of subjective emotion. The hypothesis that amygdala
activation is associated with perception of emotion-related stimuli
but not subjective emotional experience predicts that its activation,
as well as that of the hippocampus, will be limited to the time the
aversive stimuli are present. The hypothesis that the lateral OFC
mediates spontaneous self-reflection that is temporally associated
with subjective emotion predicts that its activation will be observed
during the presentation of aversive pictures and also during the
subsequent epoch of continuing subjective emotion.
Fig. 1. Design of the emotion task. NES=n
Methods
Subjects
The Human Subjects Committee at University of California,
Davis, approved all protocols. Nine volunteers (4 male) were
recruited by advertisement from the local community and paid for
participation. All subjects gave informed consent and underwent a
screening interview to rule out psychiatric or neurological
conditions and medications affecting neural or cerebrovascular
function. All subjects reported that they were right-hand dominant.
Ages ranged from 27 to 50 (average = 38.67, standard deviation
(SD) = 8.7), education ranged from 12 to 18 years (average =
14.22, SD = 2.68) and ethnicity included 5 White, 2 Hispanic, 1
African American, and 1 African American/White individual.
Stimuli
Fifty-two pictures were selected from the International Affec-
tive Picture Set (Lang et al., 1997) and from additional highly
aversive pictures generously provided by our colleagues (Liberzon
et al., 2000) and previously described in detail (Garrett and
Maddock, 2001). Briefly, 26 aversive pictures were chosen to be as
unpleasant and arousing as possible, in order to evoke a lasting
emotional response, including scenes of violence, mutilation, and
injury. The remaining 26 pictures were chosen to be neutral, having
medium levels of arousal and valence. All pictures were chosen
based on published ratings and were equated on measures of
luminosity and complexity.
Emotion task
An abbreviated version of the Negative Emotional State (NES)
rating scale (Garrett and Maddock, 2001) was used by subjects to
indicate the intensity of their negative emotions while in the
scanner. Ratings for the abbreviated version ranged from 1 to 5,
where a rating of 1 corresponded to ‘‘no negative feelings’’ and the
scale descriptors ‘‘not upset, not uneasy, not bothered’’. A rating of
3 corresponded to ‘‘moderately negative feelings’’ and the
descriptors ‘‘uncomfortable, uneasy, bothered’’. A rating of 5
corresponded to ‘‘extremely negative feelings’’ and the descriptors
‘‘upset, shocked, queasy’’. Each subject practiced rating the
intensity of his/her current negative feelings following the viewing
of a sample set of neutral pictures.
For the emotion task, 20-s picture epochs were followed by 26-s
rating epochs (Fig. 1). During the picture epochs, subjects were
instructed to look carefully at the pictures and to let any emotions
arise naturally if they occurred. For the rating epochs, the subject
egative emotion scale; sec=seconds.

A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274 265
was instructed not to rate the pictures. Specifically, the subject was
told to rate his/her current negative feelings at that moment, whether
or not the feelings had been elicited by the previous pictures or his/
her own thoughts and memories. Subjective reports of the current
emotional state prompted by cues have been suggested to be a
relatively accurate estimate of ongoing experience, more accurate,
for example, than ratings referring to a previous time (Hurlburt and
Heavey, 2001). Finally, the subject was told that he/she would be
asked to make several ratings of emotion in succession, to assess
whether or not their emotions changed over time. The subject was
not biased to report changes in ratings over time, as we explained
that ‘‘sometimes emotions do not change over time, and sometimes
emotions do change over time. Please let us know what you
experience’’.
The task was presented, and ratings were collected using
Eprime software (Psychology Software Tools, Pittsburgh, PA).
During each picture epoch, 4 pictures were presented at the rate of
5 s per picture. During the rating epochs, 4 consecutive ratings
were collected, at the rate of 1 rating per 6 s. During the rating
epochs, a fixation cross was presented. The fixation cross
intermittently turned to a star (*) to signal the subject to make a
rating by pressing a button. In addition, 2 s was allowed at the
beginning of each rating period in order to give the subject a brief
interval to transition to the rating epoch. Six aversive and six
neutral picture epochs were presented. Each picture epoch was
followed by a rating epoch. An additional neutral picture epoch
and rating epoch were used as an initial baseline but were not
analyzed, as they were designed to allow the subject to acclimate to
the scanner and the task.
Imaging protocol
Images were obtained with a 1.5-T magnetic resonance imaging
system (Signa Advantage: General Electric, OS version 8.4) with a
GE birdcage head coil. For anatomical localization in each subject,
a high-resolution Fast Spin Echo scan was acquired in the coronal
plane (TR = 3100; effective TE = 17 and 36; matrix = 256 � 256;
FOV = 22 cm; slice thickness = 6 mm with a 2 mm gap; 24 slices).
Functional images were acquired using a T2*-weighted gradient
recalled echo planar imaging (EPI) pulse sequence with shim (TR =
2000 ms; effective TE = 40 ms; flip angle = 90-; matrix = 64 � 64;
FOV = 22 cm; slice thickness = 6 mm with 2 mm gap; 18 coronal
slices). Activation was detected by the blood oxygenation level
dependent (BOLD) contrast. Head motion was constrained by foam
padding.
Picture ratings
Following the end of the scanning session, subjects rated each
picture using the Self-Assessment Mannikin (SAM, Bradley and
Lang, 1994). Each picture was rated on dimensions of valence and
arousal in order to verify that the subjects perceived the pictures as
intended.
Functional image analysis
Images were reconstructed using a Fourier transform-based
algorithm with removal of N/2 ghost artifacts (Buonocore and Gao,
1997). SPM99 (http://www.fil.ion.bpmf.ac.uk/spm) was used for
image analysis, including adjustment for timing of slice acquisi-
tion. Realignment for movement correction used a least squares
minimization without higher order corrections for spin history.
Images were normalized to the Montreal Neurological Institute
template and resampled by sinc interpolation into 2 � 2 � 2 mm
voxels. Spatial smoothing was performed using a 4-mm Gaussian
kernel.
Whole-brain statistical analyses were performed using the
theory of Gaussian random fields (Holmes and Friston, 1998).
For each subject, confounding effects of fluctuation in global mean
were removed by proportional scaling, where each voxel was
scaled by the global mean at each time point. Low-frequency noise
was removed using a high-pass filter. A canonical hemodynamic
response function was used to model the BOLD response.
Comparisons of interest were performed for each subject by
computing voxel-wise t statistics normalized to Z scores to provide
a statistical measure of activation independent of sample size. A
random effects model was used to combine individual subject data
into a group analysis. This model estimates the error variance for
each condition of interest across subjects, rather than across scans,
providing better generalization to the population. Statistical
significance was determined as clusters that passed a joint
threshold of height (Z > 1.67; P < 0.05) and extent (cluster size
corresponding to P < 0.05) after correction for multiple compar-
isons (Poline et al., 1997).
Analysis of the emotion task
Brain regions associated with the perception of negative
emotional stimuli were identified as those activated during the
aversive picture epochs (compared to the neutral picture epochs)
but not during the post-aversive rating epochs (compared to the
post-neutral rating epochs). Regional activations temporally
associated with the subjective experience of negative emotion
were identified as those activated in both the aversive picture
contrast and the post-aversive rating contrast. This conjunction
analysis was used instead of a reference vector that modeled the
emotional response across the entire pictures and ratings epochs in
order to avoid biasing the results toward regions activated during
the picture epochs. Strong activation during the picture epochs
leads to false positive activation when considering the picture and
ratings epochs together. Therefore, the conjunction analysis is the
most conservative method for identifying activation temporally
associated with subjective emotion both during and after the
picture blocks.
This analysis is different than the mixed blocked/event-related
design described previously to separate transient and sustained
activity in fMRI (Visscher et al., 2003). First, responses to the
pictures were not analyzed as events because the goal was to build
a negative emotional response across the block and not to address
the question of responses to individual aversive pictures. Secondly,
the established method of analyzing sustained activation is
designed to isolate components of the hemodynamic response that
last throughout the series of pictures but are not maintained beyond
the picture block. The current study, by contrast, is interested in
identifying activation that continues in the absence of picture
stimuli, just as the subjective experience of negative emotion
continues beyond the termination of picture stimuli.
To increase our sensitivity to activation associated with
negative subjective emotion, the rating block having the lowest
NES ratings for each individual subject was excluded from the
analysis (Canli et al., 2002). Also, we truncated emotion rating
blocks to 20 s to isolate the time of elevated negative emotion.
A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274266
Therefore, the first 20 s of the aversive emotion rating epochs were
compared to the first 20 s of the neutral emotion rating epochs.
Twenty seconds was chosen based on the ratings of subjective
emotion reported by the subjects in this study (reported below).
In a separate analysis, correlations between ratings and brain
activation were performed in SPM99 by specifying each subject’s
vector of NES ratings as a regressor in the linear model. Activation
related to the ratings was chosen as the contrast of interest.
Results
Picture ratings
Repeated measures ANOVA confirmed that subjects rated the
aversive pictures as significantly higher in arousal (F(1,46) =
158.28, P < 0.0001) and significantly more negative in valence
(F(1,46) = 102.14, P < 0.0001) compared to the neutral pictures.
Subjective emotion ratings
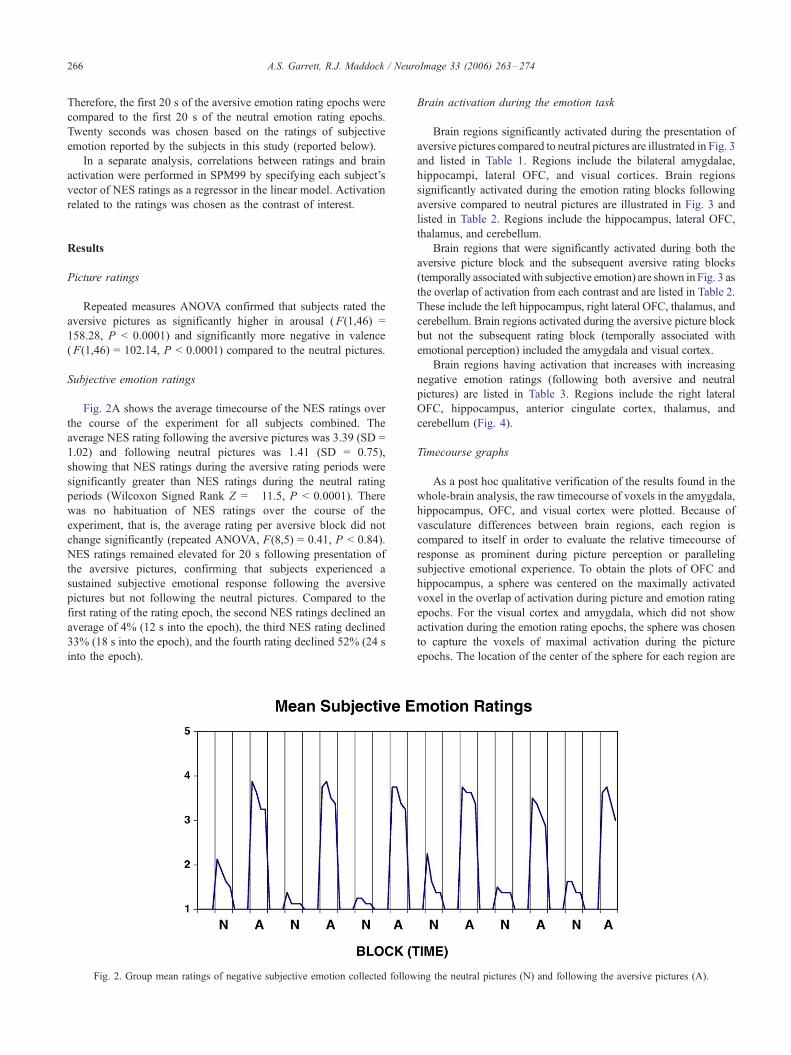
Fig. 2A shows the average timecourse of the NES ratings over
the course of the experiment for all subjects combined. The
average NES rating following the aversive pictures was 3.39 (SD =
1.02) and following neutral pictures was 1.41 (SD = 0.75),
showing that NES ratings during the aversive rating periods were
significantly greater than NES ratings during the neutral rating
periods (Wilcoxon Signed Rank Z = �11.5, P < 0.0001). There
was no habituation of NES ratings over the course of the
experiment, that is, the average rating per aversive block did not
change significantly (repeated ANOVA, F(8,5) = 0.41, P < 0.84).
NES ratings remained elevated for 20 s following presentation of
the aversive pictures, confirming that subjects experienced a
sustained subjective emotional response following the aversive
pictures but not following the neutral pictures. Compared to the
first rating of the rating epoch, the second NES ratings declined an
average of 4% (12 s into the epoch), the third NES rating declined
33% (18 s into the epoch), and the fourth rating declined 52% (24 s
into the epoch).
Fig. 2. Group mean ratings of negative subjective emotion collected follow
Brain activation during the emotion task
Brain regions significantly activated during the presentation of
aversive pictures compared to neutral pictures are illustrated in Fig. 3
and listed in Table 1. Regions include the bilateral amygdalae,
hippocampi, lateral OFC, and visual cortices. Brain regions
significantly activated during the emotion rating blocks following
aversive compared to neutral pictures are illustrated in Fig. 3 and
listed in Table 2. Regions include the hippocampus, lateral OFC,
thalamus, and cerebellum.
Brain regions that were significantly activated during both the
aversive picture block and the subsequent aversive rating blocks
(temporally associatedwith subjective emotion) are shown in Fig. 3 as
the overlap of activation from each contrast and are listed in Table 2.
These include the left hippocampus, right lateral OFC, thalamus, and
cerebellum. Brain regions activated during the aversive picture block
but not the subsequent rating block (temporally associated with
emotional perception) included the amygdala and visual cortex.
Brain regions having activation that increases with increasing
negative emotion ratings (following both aversive and neutral
pictures) are listed in Table 3. Regions include the right lateral
OFC, hippocampus, anterior cingulate cortex, thalamus, and
cerebellum (Fig. 4).
Timecourse graphs
As a post hoc qualitative verification of the results found in the
whole-brain analysis, the raw timecourse of voxels in the amygdala,
hippocampus, OFC, and visual cortex were plotted. Because of
vasculature differences between brain regions, each region is
compared to itself in order to evaluate the relative timecourse of
response as prominent during picture perception or paralleling
subjective emotional experience. To obtain the plots of OFC and
hippocampus, a sphere was centered on the maximally activated
voxel in the overlap of activation during picture and emotion rating
epochs. For the visual cortex and amygdala, which did not show
activation during the emotion rating epochs, the sphere was chosen
to capture the voxels of maximal activation during the picture
epochs. The location of the center of the sphere for each region are
ing the neutral pictures (N) and following the aversive pictures (A).
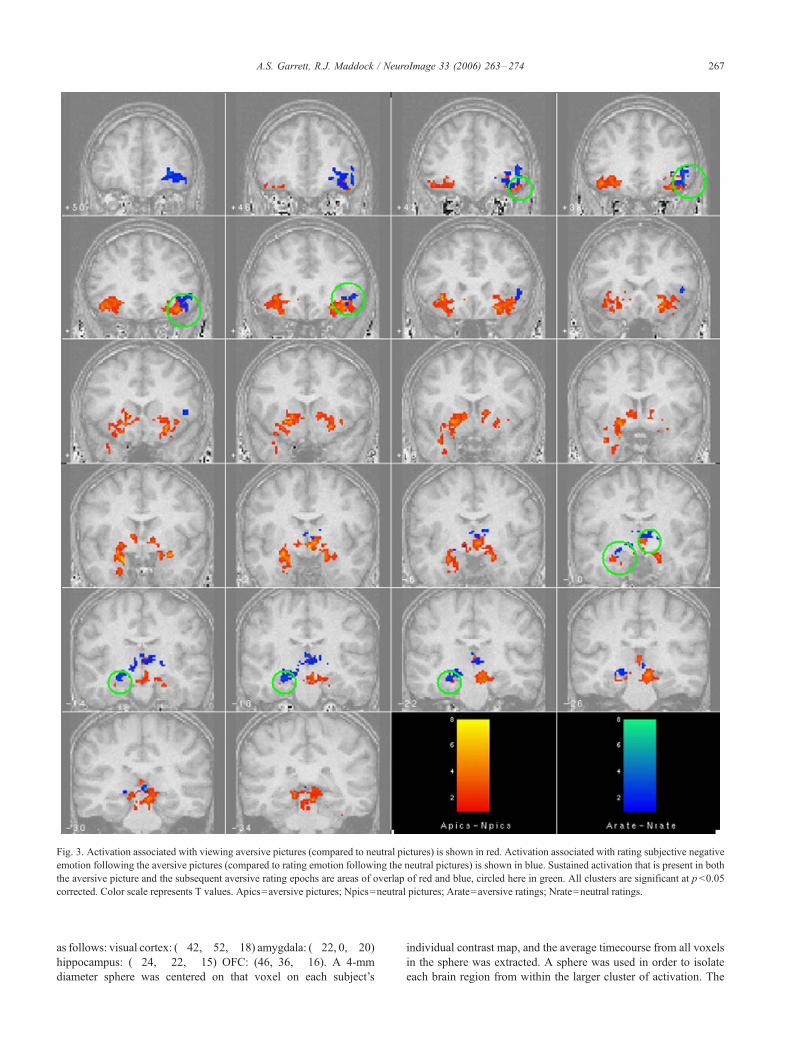
Fig. 3. Activation associated with viewing aversive pictures (compared to neutral pictures) is shown in red. Activation associated with rating subjective negative
emotion following the aversive pictures (compared to rating emotion following the neutral pictures) is shown in blue. Sustained activation that is present in both
the aversive picture and the subsequent aversive rating epochs are areas of overlap of red and blue, circled here in green. All clusters are significant at p<0.05
corrected. Color scale represents T values. Apics = aversive pictures; Npics = neutral pictures; Arate = aversive ratings; Nrate = neutral ratings.
A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274 267
as follows: visual cortex: (�42,�52,�18) amygdala: (�22, 0,�20)hippocampus: (�24, �22, �15) OFC: (46, 36, �16). A 4-mm
diameter sphere was centered on that voxel on each subject’s
individual contrast map, and the average timecourse from all voxels
in the sphere was extracted. A sphere was used in order to isolate
each brain region from within the larger cluster of activation. The

Table 1
Areas of significant activation to aversive pictures compared to neutral pictures
Region (Brodmann’s area) # Voxels (corrected
P of cluster)
Z score of peak
voxel in region
Talairach coordinates,
X, Y, Z
Uncorrected P of
peak voxel in region
L middle occipital gyrus (19) 3531 4.41 �48, �77, 15 0.001
L inferior occipital gyrus (19) (P = 0.001) 4.19 �36, �72, �3 0.001
L fusiform gyrus (19) 3.47 �40, �66, �8 0.001
L inferior temporal gyrus (37) 3.49 �50, �62, �2 0.001
L middle temporal gyrus (37/21) 3.62 �48, �54, 6 0.001
R inferior temporal gyrus (37) 4366 4.41 51, �64, 3 0.001
R inferior occipital gyrus (19) (P = 0.001) 3.92 42, �74, �8 0.001
R middle occipital gyrus (18/19) 4.36 30, �78, 24 0.001
R fusiform gyrus (19) 4.03 46, �67, �10 0.001
R cerebellum 2.78 40, �44, �26 0.003
R middle temporal gyrus (37) 3.92 50, �56, 1 0.001
R superior temporal sulcus (21/22) 2.23 55, �48, 19 0.013
L amygdala 4424 4.21 �22, 3, �17 0.001
R amygdala (P = 0.001) 2.87 20, �2, �10 0.002
L parahippocampal gyrus 2.95 �14, �31, �5 0.002
L hippocampus 2.44 �30, �16, �16 0.007
R hippocampus 3.11 22, �10, �15 0.001
Midbrain 3.63 6, �32, �12 0.001
R thalamus 3.30 4, �15, 4 0.001
R/L hypothalamus 3.98 2, �2, �5 0.001
R putamen 2.64 26, 13, �4 0.004
L putamen 3.72 �24, 11, �6 0.001
R caudate 2.83 12, 12, 1 0.002
L caudate 4.10 �16, 17, �1 0.001
R anterior insula 2.86 38, 11, �4 0.002
R lateral OFC (11/47) 3.75 40, 29, �10 0.001
L lateral OFC (11/47) 3.84 �34, 25, �8 0.001
R OFC (10/11) 3.71 26, 26, �15 0.001
L OFC (10/11) 3.39 �38, 34, �15 0.001
OFC = orbital frontal cortex; inf = inferior; R = right; L = left.
A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274268
timecourses were averaged across blocks, i.e., the aversive blocks
(including both picture and subsequent ratings blocks) were
averaged into a single timecourse per subject, as were the neutral
blocks. Individual subject timecourse data were normalized, to
remove differences in relative intensity values, by dividing each
point in the timecourse by the average of the first 2 timepoints (4 s) of
that same timecourse, which corresponds to a baseline. This method
is commonly used in the analysis of scalp-recorded event-related
potentials (Dawson, 1954). After normalization, all subjects’
average timecourse data were combined into a group average and
graphed as the change in intensity over time.
Table 2
Areas of significant activation during negative emotion ratings following aversive compared to neutral pictures
Region (Brodmann’s area) # Voxels (corrected
P of cluster)
Z score of peak
voxel in region
Talairach coordinates
X, Y, Z
Uncorrected P of
peak voxel in region
Cerebellar vermis 1786 4.35 4, �70, �32 0.001
R/L cerebelluma (P = 0.001) 3.94 �6, �78, �10 0.001
R/L lingual gyrus (18) 3.14 4, �82, �4 0.001
R/L thalamusa 624 3.82 4, �29, 1 0.001
L hippocampusa (P = 0.03) 2.77 �14, �22, �7 0.003
R/L hypothalamusa 2.90 �2, �4, �7 0.002
R lateral OFC (11/47)a 841 3.40 50, 36, �14 0.001
R OFC (11) (P = 0.004) 2.94 30, 48, �6 0.002
R inferior frontal gyrus (45/47) 2.89 48, 36, �5 0.002
R frontal operculum/anterior insula 2.54 48, 21, 1 0.006
OFC = orbital frontal cortex; R = right; L = left.a Region showing sustained activation (activation overlapped that seen in the aversive compared to neutral pictures contrast).
Fig. 5 shows the graph of the timecourses. For all regions,
activation is delayed about 3 TR, consistent with a standard
hemodynamic lag. Note that activation in the visual cortex is
transient for both the aversive and neutral blocks. Amygdala
activation is elevated for a longer time during the aversive
compared to neutral blocks but still declines approximately at the
end of picture presentation during the aversive blocks. However,
hippocampal activation is sustained well into the rating epochs for
the aversive but not neutral blocks. Activation in the lateral OFC is
also sustained into the rating epoch for the aversive compared to
the neutral blocks. These graphs confirm the results of the

Table 3
Areas of activation significantly correlated with ratings of negative emotions
Region (Brodmann’s area) # Voxels (corrected P of cluster) Z score of peak voxel Talairach coordinates X, Y, Z P value of peak voxel
Cerebellar vermis 3336 4.41 4, �71, �30 0.001
R cerebellum (P = 0.001) 4.00 32, �42, �26 0.001
R hippocampus 4.02 16, �20, �12 0.001
L hippocampus 3.33 �14, �16, �9 0.001
R thalamus 3.09 8, �19, 8 0.001
L thalamus 3.05 �6, �11, 8 0.001
R lateral OFC (11/47) 810 4.17 26, 52, �18 0.001
R anterior insula (P = 0.01) 3.29 50, 32, �17 0.001
L precentral gyrus (4) 1277 3.96 �36, �7, 56 0.001
L postcentral gyrus (1/2) (P = 0.001) 3.39 �32, �13, 47 0.001
L middle frontal (6) 3.16 �28, 9, 59 0.001
L medial frontal (6/32) 756 3.57 �8, 12, 49 0.001
R medial frontal (6/32) (P = 0.015) 4.58 6, 12, 40 0.001
R anterior cingulate (24) 3.02 2, 23, 32 0.001
L anterior cingulate (24) 1.80 �8, 9, 33 0.036
OFC = orbital frontal cortex; R = right; L = left.
A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274 269
quantitative analysis and illustrate that the lateral OFC and
hippocampal activations begin during the presentation of the
aversive pictures, and remain above baseline concurrent with the
subjective experience of negative emotion.
Discussion
This study shows that the neural response to highly arousing,
unpleasant (aversive) pictures can be separated into activation
temporally associated with emotional perception and activation
temporally associated with the subjective experience of emotion.
We identified brain regions in which the response to aversive
pictures was limited to the period of stimulus presentation and
brain regions in which the response was sustained beyond the
period of stimulus presentation while subjects continued to
experience negative emotion. During the presentation of aversive
pictures, activation was seen in predicted regions, including the
visual cortex, amygdala, hippocampus, and lateral OFC. During
the subsequent epoch, while subjects’ ratings indicated an ongoing
negative emotional state, activation continued in the lateral OFC
and the hippocampus but not in the amygdala. These data suggest
that the amygdala is primarily activated during the perception of
emotion-related stimuli, but not during subjective emotion that
occurs after the offset of the emotion-related stimuli. Activation in
the lateral OFC during the aversive pictures continued after the
offset of the pictures while subjects reported ongoing subjective
emotion, indicating that this region may have a role in subjective
emotion. However, other studies suggest that this region is more
likely involved in the self-monitoring or self-reflection that often
accompanies subjective emotion. Finally, the hippocampus showed
sustained activation throughout the presentation of stimuli and the
subjective emotion rating epochs and may be related to enhanced
encoding of aversive stimuli.
The findings suggest that the amygdala plays a relatively
transient role in the emotional response to aversive pictures.
Amygdala activation increased during the perception of aversive
pictures but did not persist during subjective reports of sustained
negative emotion. Furthermore, ratings of subjective emotion
collected after picture offset did not show a correlation with
amygdala activation. While the role of the amygdala is emotional
processes in general is well established, the role of the amygdala
specifically in subjective emotion has been debated. Previous
neuroimaging studies have shown amygdala activation during a
variety of emotion-related paradigms (see Phan et al., 2002 for a
review), including during induced emotional states (Levesque et
al., 2003), correlated with ratings of subjective emotion, including
pain (Ketter et al., 1996; Schneider et al., 2001), and amygdala
activation that was up or down-modulated while subjects con-
sciously up or down regulated subjective negative emotion
(Ochsner et al., 2004). However, these studies have not separated
the effects of emotion-inducing stimuli from those of subjective
emotion. Schaefer et al. (2002) observed increased amygdala
activation while subjects consciously maintained a negative mood,
in the absence of negative picture stimuli. However, this study did
not address the role of the amygdala during the natural timecourse
of subjective emotion. Moreover, human subjects with lesions of
the amygdala report normal subjective experience of emotion
(Anderson and Phelps, 2002) and amygdala lesions in humans
impair the perception of fearful facial expressions but do not
disrupt the expression of fear (Anderson and Phelps, 2000). The
current study supports the hypothesis that the amygdala partic-
ipates in perception of emotion-related stimuli but is not directly
involved in the subjective negative emotion that persists after the
offset of the evoking stimulus.
As predicted, the lateral OFC exhibited activation during
presentation of the aversive pictures that continued during the
post-stimulus period, during which subjects reported ongoing
negative emotion. Further, individual ratings of emotion were
significantly correlated with activation in this region. Together,
these findings suggest that the lateral OFC mediates some
component(s) of the emotional response that is temporally
associated with the subjective experience of emotion. Neuroana-
tomical evidence supports this idea, as the lateral OFC receives
sensory information, insular projections, and limbic input from the
amygdala and entorhinal cortex (Price, 1999) and therefore could
integrate, monitor, or regulate these components as part of a
multifaceted emotional response.
Many neuroimaging studies have shown activation of the
lateral OFC in response to emotion-inducing stimuli. In a recent
meta-analysis, Steele and Lawrie (2004) reviewed 30 published
experiments showing emotion-related activation along the orbital,
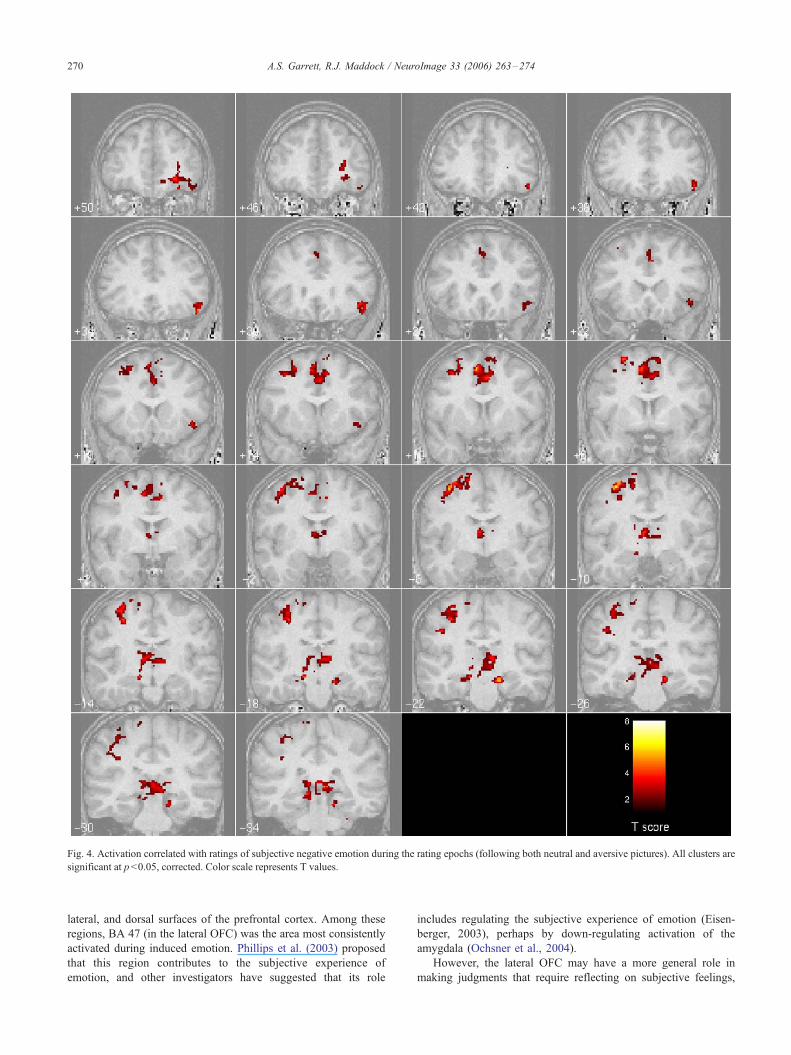
Fig. 4. Activation correlated with ratings of subjective negative emotion during the rating epochs (following both neutral and aversive pictures). All clusters are
significant at p < 0.05, corrected. Color scale represents T values.
A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274270
lateral, and dorsal surfaces of the prefrontal cortex. Among these
regions, BA 47 (in the lateral OFC) was the area most consistently
activated during induced emotion. Phillips et al. (2003) proposed
that this region contributes to the subjective experience of
emotion, and other investigators have suggested that its role
includes regulating the subjective experience of emotion (Eisen-
berger, 2003), perhaps by down-regulating activation of the
amygdala (Ochsner et al., 2004).
However, the lateral OFC may have a more general role in
making judgments that require reflecting on subjective feelings,

Fig. 5. Timecourses of activation in the left hippocampus, right lateral OFC, amygdala, and visual cortex during aversive, compared to neutral, pictures and
ratings. This graph is not adjusted for the hemodynamic delay, thus the hemodynamic response begins approximately 3 TR after the neural response. (a)
Activation that is transient relative to the comparison condition includes the visual cortex and left amygdala. (b) Activation that is sustained relative to the
comparison condition includes the left hippocampus and right lateral OFC.
A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274 271
including value judgments {Zysset, 2002 #85}, feelings of sadness
(Levesque et al., 2003), enjoyment while eating chocolate (Small et
al., 2001), pleasant and painful touch (Rolls et al., 2003), and taste
(Kringelbach et al., 2003). Also, neuroimaging studies without an
affective component show activation in BA 47 related to meta-
memory phenomena (Maril et al., 2001; Kikyo et al., 2002), active
retrieval in working memory (Wager and Smith, 2003), long-term
memory (Ranganath et al., 2003; Wig et al., 2004), semantic
retrieval (Wagner et al., 2001), and reasoning tasks (Goel et al.,
2004). Together, these studies suggest that BA 47 mediates a range
of executive functions, including reflecting on current subjective
experience, and that these functions can be flexibly recruited to
serve either emotional or non-emotional processes.
The subjective experience of emotion has been conceptualized
as including both phenomenal experience (specific feelings and
sensations) and self-reflection (meta-cognitive appraisal of phe-
nomenal experience) (Lane et al., 1997; Schooler, 2002; Lieber-
man et al., 2004). Although self-reflection does not accompany
all emotional responses, it is often spontaneously evoked during
strong, negative emotional responses and may have an important
role in the regulation of negative emotional states (Schooler,
2002; Lieberman et al., 2004). Lieberman et al. (2004) proposed
that the self-reflection evoked by negative emotion is mediated by
the right ventrolateral prefrontal cortex. The current results are
consistent with this formulation. It is worth noting that our design
allowed for both spontaneous self-reflection (accompanying
subjective emotion) and deliberate self-reflection (while making
self-ratings). However, the latter occurred only during the rating
epoch. Therefore, only self-reflection that spontaneously accom-
panied subjective emotion would have been captured by our
conjunction analysis. Experiments specifically designed to exam-
ine the role of self-reflection in subjective emotion will be
necessary to test the hypothesis that the right lateral OFC
mediates this process.
A follow-up analysis of data from our laboratory confirms that
the lateral OFC participates in non-emotional processes as well as
emotion-related evaluation. To test this, a second experiment,
called the memory-rating task, used the same subjects and a
similar experimental design. It was implemented as a control
experiment to identify activation related to the meta-cognitive
process of deliberately reflecting on and rating a non-emotional
internal experience and to determine whether activation during the
rating epochs was related to memory retrieval of the previously
presented pictures. The timing of the memory-rating task was

A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274272
identical to the emotion task. Subjects viewed blocks of neutral
pictures, followed by blocks in which they made ratings. For the
picture epochs, subjects were instructed to look carefully at the
pictures but not to make any specific effort to remember them, as
they would not be tested on their memory of the pictures. For the
rating epochs, subjects were instructed to rate the clarity of their
memory at that moment for any of the previously shown pictures,
without explicitly trying to remember the pictures. These
instructions were used to discourage effortful strategies of
encoding and retrieval and to direct the subject to rate the clarity
of memories retrieved with minimal effort. The subjects were told
that they would be asked to make several ratings in succession, so
that we would know whether or not the clarity of their memory
changed over time. Picture/Rating epochs were compared with
epochs in which the subject was shown unrecognizable scrambled
pictures and then rated his/her subjective emotion using the NES
scale. Results showed that the lateral OFC (but not the amygdala
or hippocampus) was activated while subjects performed the non-
emotional memory-rating task. These results support our hypoth-
esis that BA 47 activation is not specific to emotion-related
processes but is related to the process of self-reflection, including
the deliberate self-reflection that occurs during the rating task and
the spontaneous self-reflection that occurs during subjective
emotion. The hippocampus was not activated during the rating
epochs, possibly because subjects were not asked to try to
remember the pictures and therefore no effortful encoding took
place, and the pictures were not emotion-related or otherwise
highly interesting.
We did not observe sustained medial prefrontal or anterior
cingulate cortex (ACC) responses to the aversive pictures.
However, ratings of subjective emotion were significantly corre-
lated with activation of the ACC. Several studies have suggested
that the ACC has an important role in the subjective experience of
negative emotion (Mayberg et al., 1999; Lane, 2000; Hornak et al.,
2003). One study (Taylor et al., 2003) showed that the process of
rating emotional pictures increased medial prefrontal and ACC
activation compared to passive viewing. Another (Lane et al.,
1998) reported that attention to subjective experience is associated
with increased anterior cingulate activation. The lack of sustained
ACC activation in this study may indicate variability in its
response to different experimental conditions or insufficient
statistical power. However, note that we cannot comment on the
lack of activation in portions of the ventral prefrontal cortex. An
analysis of signal dropout due to susceptibility artifact showed that
signal loss in the anterior ventromedial PFC extends to approx-
imately Z = �15 in the anterior PFC and about Z = �10 in the
posterior PFC. This is in line with the signal loss observed in other
studies. However, it would interfere with detecting activation in
the ventral portion of BA 25 (the subgenual and subcallosal
cortices), as well as more anterior parts of the ventromedial
prefrontal cortex.
Our finding of hippocampal activation is consistent with earlier
studies of negatively valenced emotional stimuli (Phillips et al.,
1998; Lane, 2000; Maratos et al., 2001; Williams et al., 2001;
Maddock et al., 2003; Phillips et al., 2003). However, this is the
first demonstration of emotional stimulus-induced hippocampal
activation that is sustained following the offset of the inducing
stimulus. Activation of the hippocampal region has been associ-
ated with a subsequent memory effect (better subsequent memory
for stimuli evoking greater activation at encoding) (Brewer et al.,
1998; Fernandez et al., 2002; Strange et al., 2002; Davachi et al.,
2003; Stark and Okado, 2003; Jackson and Schacter, 2004).
Animal, lesion, pharmacological and imaging studies show that
modulation of hippocampal activity by the amygdala mediates
enhanced memory for emotionally arousing stimuli (Anderson and
Phelps, 2001; Hamann, 2001; Kilpatrick and Cahill, 2003).
Although we did not assess memory in this study, the hippocampal
and amygdala activation may reflect enhanced memory encoding
for the aversive pictures. If so, our findings suggest that the
emotional memory processes mediated by the amygdala and
hippocampus may have different temporal dynamics, with only
hippocampal activity sustained during the post-stimulus period.
Fear conditioning studies suggest contrasting functions for the
amygdala and the hippocampus, with associations between
specific stimuli mediated by the amygdala and contextual
associations mediated by the hippocampus (Maren, 2001;
Matus-Amat et al., 2004). Explicit memory for emotional stimuli
and events may involve similar interactions between the
amygdala and hippocampus (McGaugh et al., 1996; Cahill and
McGaugh, 1998; Richardson et al., 2004). Contextual information
typically persists after the offset of aversive stimuli, as was the
case in our experiment. Encoding of contextual information may
have contributed to the sustained activation observed in the
hippocampus.
In the current study, the cerebellum was found to be activated
during picture perception epochs as well as emotion rating epochs
and to be correlated with ratings of negative emotion. This is
consistent with the suggestion that the cerebellum is involved in
some way in the subjective experience of emotion. Previous studies
of emotion have observed cerebellar activation, for example, in
association with happy moods in men (Habel et al., 2005), in
mothers viewing photos of their infants (Nitschke et al., 2004), and
in bereaved women viewing photos of the deceased (Gundel et al.,
2003). In addition, cerebellar lesions in humans can produce
emotional flattening and impaired autonomic activity to negative
stimuli (Annoni et al., 2003). However, no studies have sought to
clarify the specific role of the cerebellum in normal human emotion.
A recent paper by Sacchetti et al. (2005) reviews evidence that the
cerebellum participates in fear learning, and that inactivation of the
cerebellar vermis in animals during the consolidation of fear
conditioning impairs fear memory, through its interconnections
with the hypothalamus, amygdala, and hippocampus. These studies
suggest that cerebellar activation in the current study may be linked
to the subjective experience of emotion as part of the autonomic
response to negative emotional states.
Several limitations of this study should be noted. The sample
size is small and includes variations in age, gender, race, and
education level that could decrease our chances of detecting
significant activation. In particular, gender differences in
responses to emotion-related stimuli have been observed (Wager
et al., 2003). However, the small sample size did not prohibit
testing our hypotheses, as significant activation was found in all
contrasts. It is not clear why activation was seen in the left
hemisphere amygdala and hippocampus and in the right
hemisphere lateral OFC. A review by Wager et al. (2003)
concludes that there is no consistent pattern of lateralization of
activation to emotion-related stimuli. Combined with the small
sample size of the current study, we hesitate to interpret the
hemispheric patterns seen in the current study.
Although the timing of activation in the right lateral OFC
paralleled that of subjective emotion and the magnitude of
activation correlated with ratings of subjective emotion, we cannot

A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274 273
be certain that this activation indicates a single neural process
temporally associated with the subjective experience of emotion. It
is possible that right lateral OFC activation during the emotion
rating period indicated only the explicit self-rating task. However,
activation in the same location was seen while subjects viewed the
emotional pictures without any explicit self-rating and has been
widely reported in other studies of emotion-inducing stimuli
without explicit self-rating tasks (Steele and Lawrie, 2004).
Requiring subjects to rate their subjective experience during
scanning may have diminished some components of their
emotional response (Liberzon et al., 2000; Lieberman et al.,
2004). Adding physiological measures of emotion to subjective
reports would provide a more complete assessment of emotional
responses. Furthermore, subjective ratings of emotion are an
imprecise measure of the emotional state of the subject due to
variability in reporting skills. However, cued reports of current
emotional state are more reliable and accurate than retrospective or
summary ratings (Hurlburt and Heavey, 2001), and subjective
ratings are essential for experimental investigations of the
subjective experience of emotion.
References
Adolphs, R., Tranel, D., 1999. Intact recognition of emotional prosody
following amygdala damage. Neuropsychologia 37 (11), 1285–1292.
Adolphs, R., Tranel, D., 1999. Preferences for visual stimuli following
amygdala damage. J. Cogn. Neurosci. 11 (6), 610–616.
Adolphs, R., Tranel, D., et al., 1999. Recognition of facial emotion in nine
individuals with bilateral amygdala damage. Neuropsychologia 37 (10),
1111–1117.
Amaral, D.G., 2002. The primate amygdala and the neurobiology of social
behavior: implications for understanding social anxiety. Biol. Psychiatry
51 (1), 11–17.
Amaral, D.G., Behniea, H., et al., 2003. Topographic organization of
projections from the amygdala to the visual cortex in the macaque
monkey. Neuroscience 118 (4), 1099–1120.
Anderson, A.K., Phelps, E.A., 2000. Expression without recognition:
contributions of the human amygdala to emotional communication.
Psychol. Sci. 11 (2), 106–111.
Anderson, A.K., Phelps, E.A., 2001. Lesions of the human amygdala impair
enhanced perception of emotionally salient events. Nature 411 (6835),
305–309.
Anderson, A.K., Phelps, E.A., 2002. Is the human amygdala critical for the
subjective experience of emotion? Evidence of intact dispositional
affect in patients with amygdala lesions. J. Cogn. Neurosci. 14 (5),
709–720.
Anderson, A.K., Christoff, K., et al., 2003. Dissociated neural representa-
tions of intensity and valence in human olfaction. Nat. Neurosci. 6 (2),
196–202.
Annoni, J.M., Ptak, R., et al., 2003. Decoupling of autonomic and
cognitive emotional reactions after cerebellar stroke. Ann. Neurol. 53
(5), 654–658.
Baker, S.C., Frith, C.D., et al., 1997. The interaction between mood and
cognitive function studied with PET. Psychol. Med. 27 (3), 565–578.
Bradley, M.M., Lang, P.J., 1994. Measuring emotion: the self-assessment
manikin and the semantic differential. J. Behav. Ther. Exp. Psychiatry
25 (1), 49–59.
Bradley, M.M., Cuthbert, B.N., et al., 1996. Picture media and emotion:
effects of a sustained affective context. Psychophysiology 33 (6),
662–670.
Brewer, J.B., Zhao, Z., et al., 1998. Making memories: brain activity that
predicts how well visual experience will be remembered. Science 281
(5380), 1185–1187.
Buonocore, M.H., Gao, L., 1997. Ghost artifact reduction for echo planar
imaging using image phase correction. Magn. Reson. Med. 38 (1),
89–100.
Cacioppo, J.T., Gardner, W.L., 1999. Emotion. Annu. Rev. Psychol. 50,
191–214.
Cahill, L., McGaugh, J.L., 1998. Mechanisms of emotional arousal and
lasting declarative memory. Trends Neurosci. 21 (7), 294–299.
Canli, T., Sivers, H., et al., 2002. Amygdala response to happy faces as a
function of extraversion. Science 296 (5576), 2191.
Craig, A.D., 2002. How do you feel? Interoception: the sense of the phy-
siological condition of the body. Nat. Rev., Neurosci. 3 (8), 655–666.
Damasio, A.R., 1999. The Feeling of What Happens: Body and Emotion in
the Making of Consciousness. Harcourt Brace, New York.
Davachi, L., Mitchell, J.P., et al., 2003. Multiple routes to memory: distinct
medial temporal lobe processes build item and source memories. Proc.
Natl. Acad. Sci. U. S. A. 100 (4), 2157–2162.
Dawson, G.D., 1954. A summation technique for the detection of small
evoked potentials. Electroencephalogr. Clin. Neurophysiol., Suppl. 6
(1), 65–84.
Dolcos, F., LaBar, K.S., Cabeza, R., 2004 Jun 10. Interaction between the
amygdala and the medial temporal lobe memory system predicts better
memory for emotional events. Neuron 42 (5), 855–863.
Fernandez, G., Klaver, P., et al., 2002. Human declarative memory
formation: segregating rhinal and hippocampal contributions. Hippo-
campus 12 (4), 514–519.
Garrett, A.S., Maddock, R.J., 2001. Time course of the subjective emotional
response to aversive pictures: relevance to fMRI studies. Psychiatry
Res. 108 (1), 39–48.
Goel, V., Makale, M., et al., 2004. The hippocampal system mediates
logical reasoning about familiar spatial environments. J. Cogn. Neuro-
sci. 16 (4), 654–664.
Gundel, H., O’Connor, M.F., et al., 2003. Functional neuroanatomy of
grief: an fMRI study. Am. J. Psychiatry 160 (11), 1946–1953.
Habel, U., Klein, M., et al., 2005. Same or different? Neural correlates of
happy and sad mood in healthy males. NeuroImage 26 (1), 206–214.
Hamann, S., 2001. Cognitive and neural mechanisms of emotional memory.
Trends Cogn. Sci. 5 (9), 394–400.
Hamann, S.B., Ely, T.D., et al., 1999. Amygdala activity related to
enhanced memory for pleasant and aversive stimuli. Nat. Neurosci. 2
(3), 289–293.
Hariri, A.R., Bookheimer, S.Y., et al., 2000. Modulating emotional
responses: effects of a neocortical network on the limbic system.
NeuroReport 11 (1), 43–48.
Holmes, A.P., Friston, K.J., 1998. Generalizability, random effects, and
population inference. NeuroImage, 754.
Hornak, J., Bramham, J., et al., 2003. Changes in emotion after circum-
scribed surgical lesions of the orbitofrontal and cingulate cortices. Brain
126 (Pt 7), 1691–1712.
Hurlburt, R.T., Heavey, C.L., 2001. Telling what we know: describing inner
experience. Trends Cogn. Sci. 5 (9), 400–403.
Jackson, O. III, Schacter, D.L., 2004. Encoding activity in anterior medial
temporal lobe supports subsequent associative recognition. NeuroImage
21 (1), 456–462.
Ketter, T.A., Andreason, P.J., et al., 1996. Anterior paralimbic mediation of
procaine-induced emotional and psychosensory experiences. Arch. Gen.
Psychiatry 53 (1), 59–69.
Kikyo, H., Ohki, K., et al., 2002. Neural correlates for feeling-of-knowing:
an fMRI parametric analysis. Neuron 36 (1), 177–186.
Kilpatrick, L., Cahill, L., 2003. Amygdala modulation of parahippocampal
and frontal regions during emotionally influenced memory storage.
NeuroImage 20 (4), 2091–2099.
Kringelbach, M.L., O’Doherty, J., et al., 2003. Activation of the human
orbitofrontal cortex to a liquid food stimulus is correlated with its
subjective pleasantness. Cereb. Cortex 13 (10), 1064–1071.
Lane, R., 2000. Neural correlates of conscious emotional experience. In:
Lane III, R., Nadel, L. (Eds.), Cognitive Neuroscience of Emotion.
Oxford Univ. Press, New York.

A.S. Garrett, R.J. Maddock / NeuroImage 33 (2006) 263–274274
Lane, R.D., Reiman, E.M., et al., 1997. Neuroanatomical cor
relates of happiness, sadness, and disgust. Am. J. Psychiatry 154
(7), 926–933.
Lane, R.D., Reiman, E.M., et al., 1998. Neural correlates of levels of
emotional awareness. Evidence of an interaction between emotion and
attention in the anterior cingulate cortex. J. Cogn. Neurosci. 10 (4),
525–535.
Lang, P.J., Bradley, M.M., et al., 1997. International Affective
Picture System (IAPS): Technical Manual and Affective Ratings.
NIMH Center for the Study of Emotion and Attention, Gains-
ville, FL.
LeDoux, J., 2000. Listen to the brain. In: Lane, R., Nadel, L. (Eds.),
Cognitive Neuroscience. Oxford Univ. Press, New York.
Levesque, J., Eugene, F., et al., 2003. Neural circuitry underlying voluntary
suppression of sadness. Biol. Psychiatry 53 (6), 502–510.
Liberzon, I., Taylor, S.F., et al., 2000. Limbic activation and psycho-
physiologic responses to aversive visual stimuli. Interaction with
cognitive task. Neuropsychopharmacology 23 (5), 508–516.
Lieberman, M.D., Jarcho, J.M., et al., 2004. The neural correlates
of placebo effects: a disruption account. NeuroImage 22 (1),
447–455.
Maddock, R.J., Garrett, A.S., et al., 2003. Posterior cingulate cortex
activation by emotional words: fMRI evidence from a valence decision
task. Hum. Brain Mapp. 18 (1), 30–41.
Maratos, E.J., Dolan, R.J., et al., 2001. Neural activity associated with
episodic memory for emotional context. Neuropsychologia 39 (9),
910–920.
Maren, S., 2001. Neurobiology of Pavlovian fear conditioning. Annu. Rev.
Neurosci. 24, 897–931.
Maril, A., Wagner, A.D., et al., 2001. On the tip of the tongue: an event-
related fMRI study of semantic retrieval failure and cognitive conflict.
Neuron 31 (4), 653–660.
Matus-Amat, P., Higgins, E.A., et al., 2004. The role of the dorsal
hippocampus in the acquisition and retrieval of context memory
representations. J. Neurosci. 24 (10), 2431–2439.
Mayberg, H.S., Liotti, M., et al., 1999. Reciprocal limbic–cortical function
and negative mood: converging PET findings in depression and normal
sadness. Am. J. Psychiatry 156 (5), 675–682.
McGaugh, J.L., 2000. Memory-a century of consolidation. Science 287
(5451), 248–251.
McGaugh, J.L., Cahill, L., et al., 1996. Involvement of the amygdala in
memory storage: interaction with other brain systems. Proc. Natl. Acad.
Sci. U. S. A. 93 (24), 13508–13514.
Nitschke, J.B., Nelson, E.E., et al., 2004. Orbitofrontal cortex tracks
positive mood in mothers viewing pictures of their newborn infants.
NeuroImage 21 (2), 583–592.
Ochsner, K.N., Ray, R.D., et al., 2004. For better or for worse: neural
systems supporting the cognitive down- and up-regulation of negative
emotion. NeuroImage 23 (2), 483–499.
Pare, D., 2003. Role of the basolateral amygdala in memory consolidation.
Prog. Neurobiol. 70 (5), 409–420.
Phan, K.L., Wager, T., et al., 2002. Functional neuroanatomy of emotion: a
meta-analysis of emotion activation studies in PET and fMRI. Neuro-
Image 16 (2), 331–348.
Phillips, M.L., Young, A.W., et al., 1998. Neural responses to facial and
vocal expressions of fear and disgust. Proc. R. Soc. Lond., B Biol. Sci.
265 (1408), 1809–1817.
Phillips, M.L., Drevets, W.C., et al., 2003. Neurobiology of emotion
perception: I. The neural basis of normal emotion perception. Biol.
Psychiatry 54 (5), 504–514.
Pizzagalli, D., Regard, M., et al., 1999. Rapid emotional face processing in
the human right and left brain hemispheres: an ERP study. NeuroReport
10 (13), 2691–2698.
Poline, J.B., Worsley, K.J., et al., 1997. Combining spatial extent and peak
intensity to test for activations in functional imaging. NeuroImage 5 (2),
83–96.
Price, J.L., 1999. Prefrontal cortical networks related to visceral function
and mood. Ann. N. Y. Acad. Sci. 877, 383–396.
Ranganath, C., Johnson, M.K., et al., 2003. Prefrontal activity associated
with working memory and episodic long-term memory. Neuropsycho-
logia 41 (3), 378–389.
Richardson, M.P., Strange, B.A., et al., 2004. Encoding of emotional
memories depends on amygdala and hippocampus and their interac-
tions. Nat. Neurosci. 7 (3), 278–285.
Rolls, E.T., O’Doherty, J., et al., 2003. Representations of pleasant and
painful touch in the human orbitofrontal and cingulate cortices. Cereb.
Cortex 13 (3), 308–317.
Sacchetti, B., Scelfo, B., et al., 2005. The cerebellum: synaptic changes and
fear conditioning. Neuroscientist 11 (3), 217–227.
Schaefer, S.M., Jackson, D.C., Davidson, R.J., Aguirre, G.K., Kimberg,
D.Y., Thompson-Schill, S.L., 2002 Aug 15. Modulation of amygdalar
activity by the conscious regulation of negative emotion. J. Cogn.
Neurosci. 14 (6), 913–921.
Schneider, F., Habel, U., et al., 2001. Subjective ratings of pain correlate
with subcortical – limbic blood flow: an fMRI study. Neuropsychobio-
logy 43 (3), 175–185.
Schooler, J.W., 2002. Re-representing consciousness: dissociations between
experience and meta-consciousness. Trends Cogn. Sci. 6 (8), 339–344.
Sirota, A.D., Schwartz, G.E., et al., 1987. Facial muscle activity during
induced mood states: differential growth and carry-over of elated versus
depressed patterns. Psychophysiology 24 (6), 691–699.
Small, D.M., Zatorre, R.J., et al., 2001. Changes in brain activity related
to eating chocolate: from pleasure to aversion. Brain 124 (Pt 9),
1720–1733.
Small, D.M., Gregory, M.D., et al., 2003. Dissociation of neural
representation of intensity and affective valuation in human gustation.
Neuron 39 (4), 701–711.
Stark, C.E., Okado, Y., 2003. Making memories without trying: medial
temporal lobe activity associated with incidental memory formation
during recognition. J. Neurosci. 23 (17), 6748–6753.
Steele, J.D., Lawrie, S.M., 2004. Segregation of cognitive and emotional
function in the prefrontal cortex: a stereotactic meta-analysis. Neuro-
Image 21 (3), 868–875.
Strange, B.A., Otten, L.J., et al., 2002. Dissociable human perirhinal,
hippocampal, and parahippocampal roles during verbal encoding.
J. Neurosci. 22 (2), 523–528.
Taylor, S.F., Phan, K.L., et al., 2003. Subjective rating of emotionally
salient stimuli modulates neural activity. NeuroImage 18 (3), 650–659.
Visscher, K.M., Miezin, F.M., et al., 2003. Mixed blocked/event-related
designs separate transient and sustained activity in fMRI. NeuroImage
19 (4), 1694–1708.
Wager, T.D., Smith, E.E., 2003. Neuroimaging studies of working memory:
a meta-analysis. Cogn. Affect Behav. Neurosci. 3 (4), 255–274.
Wager, T.D., Phan, K.L., et al., 2003. Valence, gender, and lateralization of
functional brain anatomy in emotion: a meta-analysis of findings from
neuroimaging. NeuroImage 19 (3), 513–531.
Wagner, A.D., Pare-Blagoev, E.J., et al., 2001. Recovering meaning: left
prefrontal cortex guides controlled semantic retrieval. Neuron 31 (2),
329–338.
Wig, G.S., Miller, M.B., et al., 2004. Separable routes to human memory
formation: dissociating task and material contributions in the prefrontal
cortex. J. Cogn. Neurosci. 16 (1), 139–148.
Williams, L.M., Phillips, M.L., et al., 2001. Arousal dissociates
amygdala and hippocampal fear responses: evidence from simulta-
neous fMRI and skin conductance recording. NeuroImage 14 (5),
1070–1079.
Related Documents



![The Neural Representation of Rhythm, Non-Rhythm and ......Persian Music, Functional Neuroimaging, fMRI, Rhythm, Non-Rhythm, Melody. M of emotion that it arouses [3]. Although music](https://static.cupdf.com/doc/110x72/6134296ddfd10f4dd73b8de2/the-neural-representation-of-rhythm-non-rhythm-and-persian-music-functional.jpg)







