Exp Brain Res (2010) 206:81–91 DOI 10.1007/s00221-010-2400-6 123 RESEARCH ARTICLE Sensory-motor equivalence: manual aiming in C6 tetraplegics following musculotendinous transfer surgery at the elbow Mark A. Robinson · Spencer J. Hayes · Simon J. Bennett · Gabor J. Barton · Digby Elliott Received: 5 June 2010 / Accepted: 18 August 2010 / Published online: 1 September 2010 © Springer-Verlag 2010 Abstract Cervical spinal lesions at C6 result in paralysis of the triceps brachii while leaving deltoid and elbow Xexor function intact. We examined the spatial–temporal charac- teristics of goal-directed aiming movements performed by C6 tetraplegics who had undergone musculotendinous transfer surgery in which the posterior deltoid replaces the triceps as the elbow extensor. On some trials, liquid crystal goggles were used to eliminate vision of the limb and target upon movement initiation. Although tetraplegic partici- pants achieved the same degree of movement accuracy/ consistency as control participants, their movement times were longer regardless of whether the movements were made away from (elbow extension) or towards the body (elbow Xexion). Longer movement times were related to lower peak velocities, and not the symmetry of the aiming proWles. The tetraplegic participants were no more depen- dent on visual feedback for limb regulation than control participants. Although the characteristics of the movement trajectories were surprisingly similar, in both vision condi- tions, tetraplegics required more real and proportional time to reduce spatial variability in the limb’s trajectory for elbow extensions. Our results indicate that the sensorimotor system is adaptable and that the representations governing limb control are not muscle speciWc. Keywords EVector independence · Motor equivalence · Tetraplegia · Tendon transfer · Aiming · Vision · Motor control · Kinematics Introduction Fundamental to the study of human motor control is deter- mining how skilled movements are represented by the central nervous system. Although early models of skilled movement advocated an eVector-speciWc form of repre- sentation, sometimes referred to as a ‘motor program’ (e.g., Keele 1968), over the years it has become clear that at least some of the spatial and temporal characteristics of limb control are independent of the particular muscle groups involved in performing the task (MacKay 1982; Schmidt 1975). Thus, a well-practised set of movements, such as signing one’s name, has kinematic similarities regardless of whether the signature is executed with the preferred hand, with the non-preferred hand or with a pen held in the mouth (Merton 1972; Raibert 1977; cf. Wright 1990). In addition, research involving inter-limb transfer of training indicates that one arm beneWts from practice by the other arm even when deprived of its own opportunity for practice (Park and Shea 2003). The term “eVector independence” has been used to describe this characteristic of general skill representation (e.g., Park and Shea 2002). A related term, ‘motor equivalence’, has been used to capture the phenomenon in which, for a given movement goal, very similar spatial–temporal move- ment patterns are produced regardless of the particular muscle groups recruited to achieve the goal (Marteniuk et al. 2000) or the speciWc force requirements (Levin et al. 2003). These behavioural similarities are also associated with speciWc dynamic patterns of cortical activity (Kelso M. A. Robinson · S. J. Hayes · S. J. Bennett · G. J. Barton · D. Elliott (&) Research Institute for Sport and Exercise Sciences, Faculty of Science, Liverpool John Moores University, Tom Reilly Building, Byrom Street, L3 3AF Liverpool, UK e-mail: [email protected] D. Elliott Department of Kinesiology, McMaster University, Ontario, Canada

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Exp Brain Res (2010) 206:81–91
DOI 10.1007/s00221-010-2400-6RESEARCH ARTICLE
Sensory-motor equivalence: manual aiming in C6 tetraplegics following musculotendinous transfer surgery at the elbow
Mark A. Robinson · Spencer J. Hayes · Simon J. Bennett · Gabor J. Barton · Digby Elliott
Received: 5 June 2010 / Accepted: 18 August 2010 / Published online: 1 September 2010© Springer-Verlag 2010
Abstract Cervical spinal lesions at C6 result in paralysisof the triceps brachii while leaving deltoid and elbow Xexorfunction intact. We examined the spatial–temporal charac-teristics of goal-directed aiming movements performed byC6 tetraplegics who had undergone musculotendinoustransfer surgery in which the posterior deltoid replaces thetriceps as the elbow extensor. On some trials, liquid crystalgoggles were used to eliminate vision of the limb and targetupon movement initiation. Although tetraplegic partici-pants achieved the same degree of movement accuracy/consistency as control participants, their movement timeswere longer regardless of whether the movements weremade away from (elbow extension) or towards the body(elbow Xexion). Longer movement times were related tolower peak velocities, and not the symmetry of the aimingproWles. The tetraplegic participants were no more depen-dent on visual feedback for limb regulation than controlparticipants. Although the characteristics of the movementtrajectories were surprisingly similar, in both vision condi-tions, tetraplegics required more real and proportional timeto reduce spatial variability in the limb’s trajectory forelbow extensions. Our results indicate that the sensorimotorsystem is adaptable and that the representations governinglimb control are not muscle speciWc.
Keywords EVector independence · Motor equivalence · Tetraplegia · Tendon transfer · Aiming · Vision · Motor control · Kinematics
Introduction
Fundamental to the study of human motor control is deter-mining how skilled movements are represented by thecentral nervous system. Although early models of skilledmovement advocated an eVector-speciWc form of repre-sentation, sometimes referred to as a ‘motor program’(e.g., Keele 1968), over the years it has become clear that atleast some of the spatial and temporal characteristics oflimb control are independent of the particular musclegroups involved in performing the task (MacKay 1982;Schmidt 1975). Thus, a well-practised set of movements,such as signing one’s name, has kinematic similaritiesregardless of whether the signature is executed with thepreferred hand, with the non-preferred hand or with apen held in the mouth (Merton 1972; Raibert 1977; cf.Wright 1990). In addition, research involving inter-limbtransfer of training indicates that one arm beneWts frompractice by the other arm even when deprived of its ownopportunity for practice (Park and Shea 2003). The term“eVector independence” has been used to describe thischaracteristic of general skill representation (e.g., Parkand Shea 2002). A related term, ‘motor equivalence’, hasbeen used to capture the phenomenon in which, for agiven movement goal, very similar spatial–temporal move-ment patterns are produced regardless of the particularmuscle groups recruited to achieve the goal (Marteniuk et al.2000) or the speciWc force requirements (Levin et al.2003). These behavioural similarities are also associatedwith speciWc dynamic patterns of cortical activity (Kelso
M. A. Robinson · S. J. Hayes · S. J. Bennett · G. J. Barton · D. Elliott (&)Research Institute for Sport and Exercise Sciences, Faculty of Science, Liverpool John Moores University, Tom Reilly Building, Byrom Street, L3 3AF Liverpool, UKe-mail: [email protected]
D. ElliottDepartment of Kinesiology, McMaster University, Ontario, Canada
123

82 Exp Brain Res (2010) 206:81–91
et al. 1998; Swinnen et al. 2010). Swinnen and colleagues(2010) have suggested that the parietal-premotor areasassociated with this activity are responsible for the abstractmovement encoding and provide the basis for eVectorindependence/motor equivalence.
Although movements performed with diVerent eVectorsystems often have similar spatial characteristics and rela-tive timing patterns (Maraj et al. 1993), there are normallydiVerences in absolute time and/or movement precision.Thus, for example, goal-directed aiming movements per-formed with the non-preferred hand are generally executedmore slowly in order to achieve the same degree of end-point accuracy (e.g., Elliott and Roy 1989). This extra timeis usually associated with the need for time-consumingonline feedback utilization to compensate for less precisecentral planning and force regulation (Carson et al. 1993).This type of result, as well as work conducted involvinginter-limb transfer of learning (e.g., Park and Shea 2003;Vangheluwe et al. 2006), suggests that there are both eVec-tor-independent and eVector-speciWc levels of movementrepresentation.
Much of the work on eVector independence hasinvolved the examination of performance diVerences,transfer of training eVects and interference eVects betweenthe arms or hands. The basic premise is although each arm/eVector system is equipped with its own set of muscles,those muscles, and the neural systems that drive them, areorganized in exactly the same fashion. Over the last coupleof years, members of our research team have been workingwith C6 tetraplegic individuals who provide us with aunique opportunity to examine how movement is repre-sented in the central nervous system. C6 tetraplegic indi-viduals are unable to perform elbow extension movementsagainst gravity or when the clinical examiner blocks theother joints to avoid compensations. Thus, accuracy duringhorizontal elbow Xexion is compromised because the tri-ceps is not available to decelerate the limb when the handreaches the target area (Wierzbicka and Wiegner 1992,1996). Despite this paralysis, C6 tetraplegic individualsare able to perform 3-dimensional (3D) reaching move-ments involving elbow extension with a smooth hand tra-jectory proWle (LaVont et al. 2000, HoVman et al. 2006).Elbow extension during 3D reaching probably resultsfrom dynamical interactions between segments that areanticipated and controlled by rotations of the shoulderjoint (according to a mechanism demonstrated in healthysubjects by Dounskaia 2005).
The procedure of musculotendinous transfer restoresthe ability to perform voluntary elbow extension againstgravity. Following transfer, patients are able to performboth elbow extension and elbow Xexion with accuracy inthe horizontal plane. Shoulder-elbow coordination of 3Dreaching movements is also improved, particularly after a
long period of rehabilitation (Remy-Neris et al. 2003).This improvement could be due to the restoration of theextensor muscle strength and/or of the physiologicalelbow stiVness resulting from the transfer, and probablyalso to more skilled shoulder-elbow coordination (HoVmanet al. 2006).
All Wve tetraplegics in our current study had under-gone musculotendinous transfer surgery in which theposterior deltoid was used to restore elbow extension.Thus, the posterior deltoid takes on the role of the tri-ceps, while more typical biceps function is preserved.Participants in this study performed rapid aiming move-ments from a central home position to small targetscloser to or further away from the body. These move-ments involved elbow Xexion and extension respec-tively. By comparing spatial–temporal characteristics ofgoal-directed aiming movements performed by theseparticipants to a group of age-equivalent control partici-pants who had typical muscle arrangement (i.e., tricepsand biceps intact), it was possible to gain some insightinto underlying movement representation and associatedcontrol processes.1
A strong “eVector independence” view of movementrepresentation predicts a high degree of adaptation to thenew neuromuscular arrangement (HoVman et al. 2006;Remy-Neris et al. 2003). However, with the patient groupexamined here, it was expected that a certain degree ofimpairment would remain relative to control participants(see LaVont et al. 2007). Therefore, our primary hypothesiswas that tetraplegics who had undergone the transfer proce-dure would take more time to achieve the same degree ofmovement precision. With respect to the issue of motorequivalence, we were interested in whether or not the kine-matic proWle of the limb trajectory would be similar in thepatient and control groups, and whether this would dependon whether the posterior deltoid assists as the agonist(extension) or antagonist (Xexion).
Based on the assumption that people with a less precisecentral representation of movement would be more depen-dent on feedback, we also decided to manipulate vision ofthe arm during movement execution. Our secondaryhypothesis was that the elimination of online visual feed-back would interfere with the aiming performance of thetetraplegic participants to a greater degree than the perfor-mance of control participants.
1 In comparison with the triceps brachii, the posterior deltoid has archi-tectural diVerences such as longer muscle Wbres (123 vs. 72 mm) and alarger excursion (150 vs. 100 mm). While these features make the pos-terior deltoid good candidate in terms of available range of motion, theposterior deltoid would only produce around 20% of the triceps muscleforce (Friden and Lieber 2001).
123

Exp Brain Res (2010) 206:81–91 83
Methods
Participants
Five tetraplegic participants (M = 39, SD = 6 years) whohad all suVered traumatic injury to the cervical spinal cord atthe C6 neurological level participated in the present experi-ment. The shoulder and elbow Xexor activation pathwayswere intact, but they were unable to activate the tricepsbrachii to extend the elbow. To replace the lost elbow exten-sor, each individual underwent a posterior deltoid to tricepsmusculotendinous transfer operation. During each opera-tion, the posterior deltoid tendon was released and thenlengthened by attaching the tendon from the tibialis anteriorthat was harvested previously. To allow the posterior deltoidto articulate the elbow, the tendon was then passed througha drilled hole in the olecranon and attached to the aponeuro-sis of the triceps. Following surgery, the elbow was placedin extension to avoid unnecessary strain on the attachmentsite and over the next 6 weeks the range of elbow jointXexion was increased in 10°–15° intervals until the elbowXexion range was restored. All tetraplegics completed a3 month rehabilitation programme, and on completion allwere able to extend the elbow against gravity with resis-tance making them at least grade 4 according to the manualmuscle test scale. Data on the participants’ age of injury, ageof surgery and ASIA motor scores are presented in Table 1.We compared our data to a group of Wve age-matched con-trol participants (M = 38, SD = 7 years). Each participantcompleted an informed consent form and was free to with-draw from the experiment at any time. All procedures wereconducted in accordance with the local ethical guidelines ofthe host University, the National Health Service LocalResearch Ethics Committee and the standards laid down inthe 1964 declaration of Helsinki.
Apparatus and task
Participants sat in front of an experimental table in a wheelchair (tetraplegics) or a chair (controls). On top of the table,we positioned a wooden board that housed a central home
position and two targets (the home and targetdiameters = 15 mm); the targets were positioned 200 mm inthe sagittal plane from the central home positioned in adirection towards or away from the body of the participant(see Fig. 1a). The task required participants to execute anaiming movement using the operated arm (or dominant armfor age-matched controls) to one of the two targets asquickly and accurately as possible. All participants reachedalong the board by rolling a ball bearing that formed part ofa ball transfer unit (LF25, Omnitrack, Woodchester, UK)attached to a wrist guard (see Fig. 1b). The ball transfer unitconsisted of a partially encased 25 mm ball bearing thatrolls on many smaller ball bearings; this product is speciW-cally designed to allow ‘frictionless’ omni-directionalmovement. Attached to the top of the wrist guard was aretroreXective marker that was the referent for the wrist.Five opto-electronic cameras (Qualisys, Gothenburg, Swe-den) with a sampling rate of 120 Hz were used to collectthe positional data of the wrist marker. The cameras werecontrolled by a laptop computer (Dell Latitude D810) run-ning Windows® XP and Qualisys Track Manager (QTM,Qualisys, Gothenburg, Sweden). Plato liquid crystal glasses(Translucent Technologies, Toronto, Canada) were used tomanipulate the presence of vision during the aiming move-ment. These glasses were connected to a separate laptopcomputer (Dell Latitude D810) that controlled the state ofthe LCD using an in-house routine programmed inMATLAB v.7.4.0.287 (The Mathworks, Inc., Natick, USA).This routine also speciWed an auditory go signal (high orlow frequency) and measured reaction time.
Procedure
Each trial began when the participant depressed the centralhome switch with the ball bearing. The two targets werealways present prior to movement execution, following arandom fore period of 500–750 ms, an audio go signal waspresented for 500 ms that indicated the required target.A high-frequency signal (400 Hz) indicated the target posi-tioned away from the body, and the low-frequency signal(200 Hz) indicated the target located near to the body.
Table 1 Functional and surgical details for the Wve tetraplegic participants
ASIA describes the level of impairment according to the American Spinal Injury Association classiWcation
Participant Age (years) Level of injury Time elapsed since ASIA
Skeletal Neurological Injury (years) Operation (years)
1 36 C5/6 C6 21 14 C
2 44 C5 C6 23 13 B
3 36 C5 C6 16 9 A
4 31 C5 C6 10 8 A
5 46 C5 C6 18 13 A
123

84 Exp Brain Res (2010) 206:81–91
There were four blocks of 16 trials. For two blocks of trials,the goggles remained open over the complete course of themovement (full vision condition), while during the no-vision condition the liquid crystal goggles closed once theparticipant released the central home switch on each trial.This ensured that visual information about the position ofthe target and the limb was eliminated until the movementwas complete. Because vision condition was blocked, par-ticipants always knew in advance whether or not visionwould be available for online limb control. Participantswere instructed to remain in their Wnal position until theglasses reopened. Vision and no-vision blocks of trials werealternated with starting order counterbalanced across par-ticipants. Within a block of trials, there were 8 movementsaway from (extension) and 8 movements towards (Xexion)the body. Thus, there were 64 trials in total, with 16 trials ineach of the 2 Vision by 2 Direction conditions. A separaterandom target order was used for each participant.
Data analysis
The position data from the wrist marker were Wltered inVisual 3D v.4.00.20 (C-Motion, Germantown, USA) at10 Hz using a Butterworth fourth-order low pass Wlter andthe Wrst and second derivatives of the wrist marker dis-placements were calculated. Movement start and end weredetermined using a velocity threshold criterion of 20 mm/s.Reaction time was measured from the onset of the auditorysignal to the start of the movement. Movement time wascalculated as the time between the two 20 mm/s thresholdsbeing reached at the start and end of the movement, respec-tively. Additional kinematic and temporal variables wereextracted using Matlab and included peak velocity, percent-age time to peak velocity and percentage time after peakvelocity. We also partitioned each movement into the initialballistic phase (i.e., primary movement) and, where appli-cable, the corrective phase. The end of the primary move-ment was identiWed by a zero crossing in the acceleration
proWle after peak deceleration, or a change from positive tonegative in velocity. These two events indicate a secondacceleration to correct a target undershoot and a reversal tocorrect an overshoot, respectively. For these data, we calcu-lated the proportion of trials with a corrective phase, as wellas the mean time spent completing the primary movementand the mean time spent in the corrective phase of themovement. These two times together equal movement time.
Constant and variable errors were calculated in the pri-mary direction of the movement. Constant error was deW-ned as the signed algebraic mean of individual error scoresand provided an indication of overall target undershooting(negative error) or overshooting (positive error). Variableerror was the standard deviation of the individual signederrors and provides an index of endpoint consistency. Theseperformance and kinematic variables were analysed usingseparate 2 Group (tetraplegics, controls) £ 2 Vision Condi-tion (vision, no vision) £ 2 Direction (away from body,towards body) mixed analysis of variance with repeatedmeasures on the last two factors.
To capture spatial variability over the course of themovement trajectory, we normalized each aiming attemptto total movement time on that trial and then determined thedisplacement of the limb at 10% intervals. We calculatedstandard deviations of these displacements over the 16 tri-als in each condition and conducted a 2 Group £ 2 VisionCondition £ 2 Direction £ 10 Movement Time Intervalmixed analysis of variance with repeated measures on thelast 3 factors. Where appropriate, follow-up tests were con-ducted using the Tukey HSD (P < .05) post hoc procedure.
Results
The reaction time analysis yielded only a main eVect forVision Condition, F(1,8) = 13.49, P < .01, with participantsinitiating their movements more quickly when vision wasavailable (477 ms) than when vision was eliminated upon
Fig. 1 Target and equipment layout (a) and ball transfer unit with wrist guard (b)
123
Exp Brain Res (2010) 206:81–91 85
movement initiation (513 ms). This Wnding is consistentwith the results of a number of other studies involving asimilar visual manipulation (e.g., Elliott and Allard 1985;Hansen et al. 2006). Presumably, people take extra time toprepare their movements when they know in advance thatvision will not be available for online control. This strategicapproach to movement planning was adopted by bothgroups of participants.
As anticipated, the movement time analysis revealed amain eVect for Group, F(1,8) = 6.72, P < .05. The tetraple-gics took signiWcantly more time executing their aimingmovements (673 ms) than control participants (491 ms).Thus, there appears to be at least some temporal costassociated with pathology and transfer surgery (see alsoRobinson et al. 2010 and LaVont et al. 2007). The analysisalso yielded a main eVect for Direction, F(1,8) = 6.60,P < .05, and a Vision Condition by Direction interaction,F(1,8) = 5.28, P < .05. Participants in both groups tookmore time to complete their movements when they wereaiming away from the body (i.e., elbow extension) thantowards the body (i.e., elbow Xexion). The increase inmovement time was particularly evident when both groupsof participants moved without the beneWt of visual feed-back (no vision-away, 623 ms; vision-away, 580 ms; novision-towards, 569 ms; vision-towards, 557 ms).
Partitioning movement time into the time associatedwith primary movement and corrective phase failed to
reveal any further insights. SpeciWcally, there was no diVer-ence between the percentage of trials in which a correctionoccurred for the two groups (tetraplegics = 43%, controls =34%, P > .10). Although there were no group diVerencesin either time to complete the primary submovement(tetraplegics = 540 ms, controls = 447 ms, P = .10) or thecorrective phase of the movement (tetraplegics = 133 ms,controls = 44 ms, P = .10) in both cases, performers in thetetraplegic group took more time. This result indicates thatthe overall movement time eVect reXects the combinedinXuence to the two movement phases.
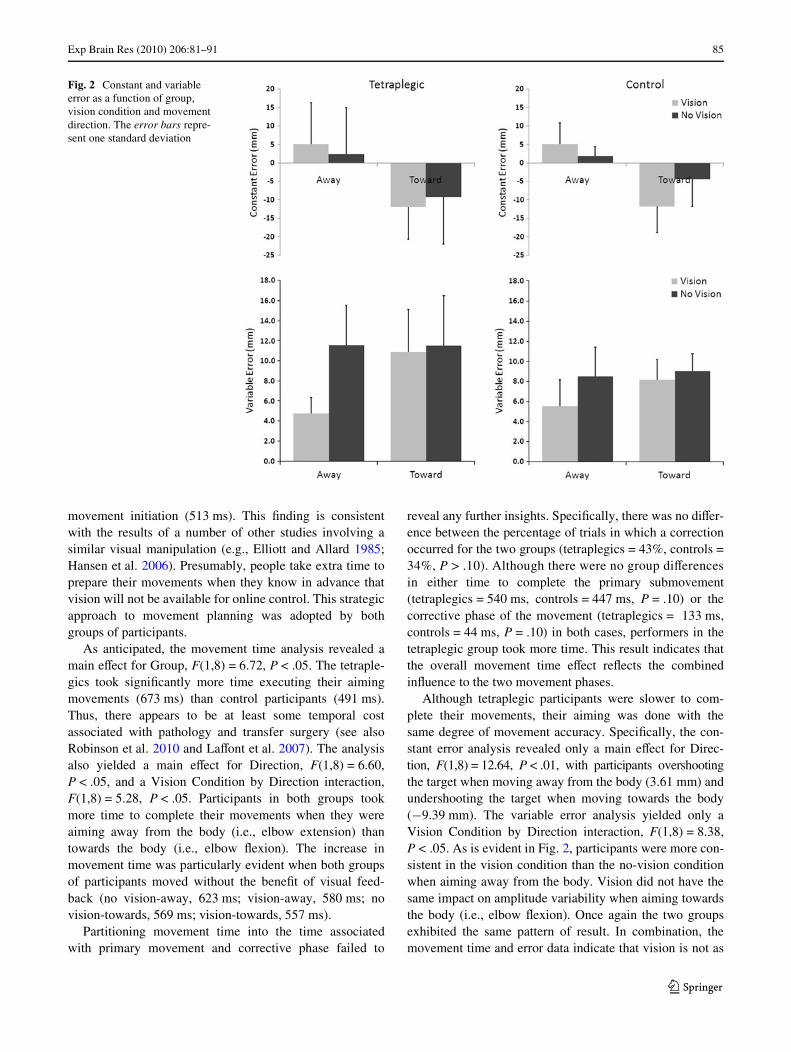
Although tetraplegic participants were slower to com-plete their movements, their aiming was done with thesame degree of movement accuracy. SpeciWcally, the con-stant error analysis revealed only a main eVect for Direc-tion, F(1,8) = 12.64, P < .01, with participants overshootingthe target when moving away from the body (3.61 mm) andundershooting the target when moving towards the body(¡9.39 mm). The variable error analysis yielded only aVision Condition by Direction interaction, F(1,8) = 8.38,P < .05. As is evident in Fig. 2, participants were more con-sistent in the vision condition than the no-vision conditionwhen aiming away from the body. Vision did not have thesame impact on amplitude variability when aiming towardsthe body (i.e., elbow Xexion). Once again the two groupsexhibited the same pattern of result. In combination, themovement time and error data indicate that vision is not as
Fig. 2 Constant and variable error as a function of group, vision condition and movement direction. The error bars repre-sent one standard deviation
123

86 Exp Brain Res (2010) 206:81–91
important for accuracy/consistency when the aimingattempts are made towards the body. This may be becausethe body itself provides stable proprioceptive referent forwhere the limb is in space (Heath and Binsted 2007).
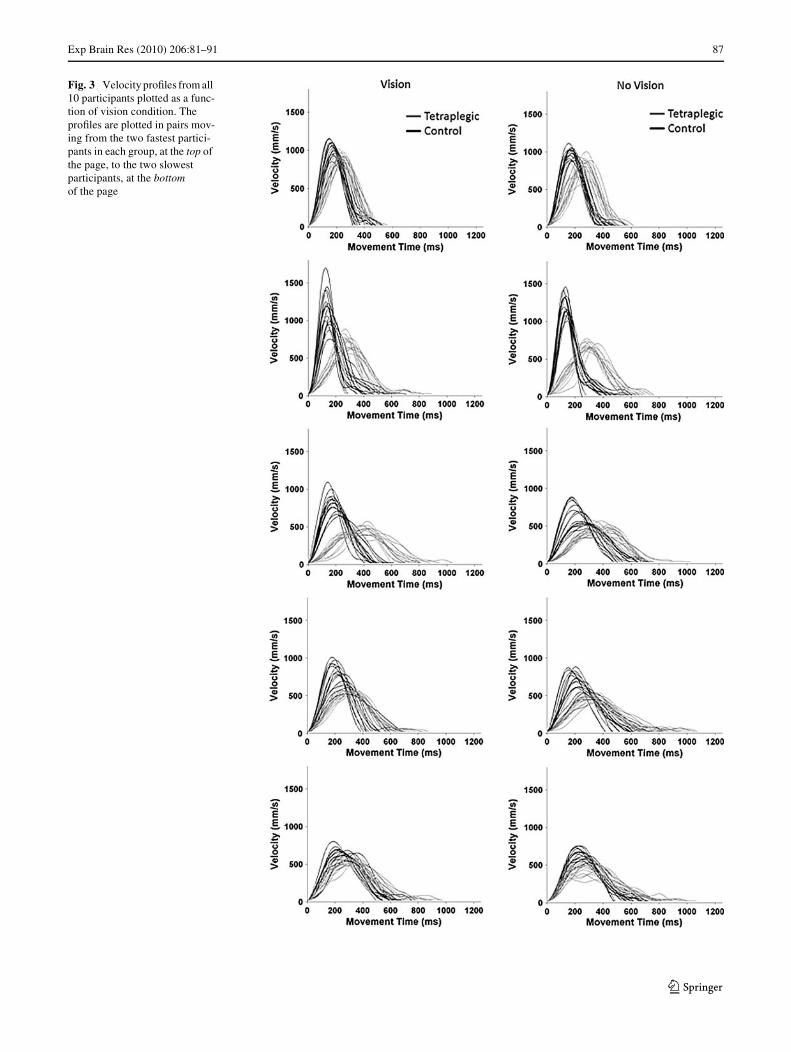
As noted above, the only important performance diVer-ence between the two groups was in movement time. Spe-ciWcally, tetraplegics took more time to execute theirmovements than control participants regardless of visioncondition and direction. The analysis of spatial–temporalcharacteristics of the movement trajectories enables us tobetter understand the basis for this performance diVerence.Velocity proWles from participants when moving awayfrom the body, both with and without vision, are depicted inFig. 3. The Wrst point to note is that at least part of themovement time diVerence between the groups is explainedby a main eVect for Group in peak velocity, F(1,8) = 5.24,P = .05 (tetraplegics = 606 mm/s, controls = 895 mm/s).
The analysis of peak velocity also revealed two interac-tions. A Group by Direction interaction, F(1,8) = 6.00,P < .05, indicated that for movements towards the bodythere was a greater diVerence between the groups (elbowXexion: tetraplegics = 595 mm/s, controls = 923 mm/s)than for movements away from the body (elbow extension:tetraplegics = 617 mm/s, controls = 868 mm/s). Moreover,a Vision Condition by Direction interaction, F(1,8) = 6.82,P < .05, revealed that participants in both groups wereparticularly slow when moving away from the bodywithout vision (away-no vision = 715 mm/s; away-vision =770 mm/s; towards-no vision = 763 mm/s; towards-vision =754 mm/s).
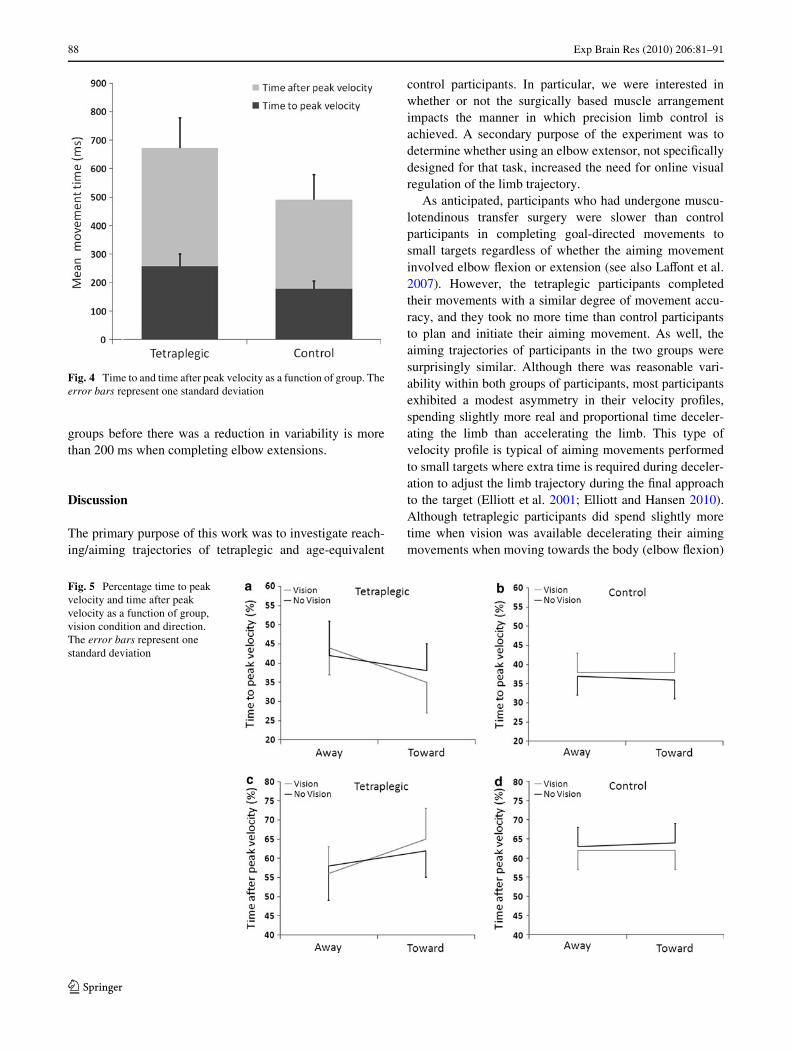
Insight into the limb control strategy adopted by partici-pants can also be obtained by examining the asymmetry ofthe velocity proWle. For movements that require a highdegree of precision, people typically spend a greater per-centage of the overall movement time in the decelerationphase of the movement (see Elliott et al. 2001 for a review).Presumably, this extra time is necessary for error-reducingonline control. In Fig. 4, we can see that movement timediVerences between the two groups are not exclusive toeither the time before or the time after peak velocity. Ratherthe mean percentage of time spent after peak velocity wasreasonably equivalent for the tetraplegic (60.3%) and con-trol participants (62.7%; F < 1.0). The only signiWcanteVect in the percentage time after peak velocity was aGroup by Direction by Vision Condition interaction,F(1,8) = 6.02, P < .05. For control participants, the symme-try of the movement trajectory was not aVected by move-ment direction or the presence or absence of vision (seeFig. 5b and d). Tetraplegic participants, however, tended tospend proportionally more time after peak velocity whenmoving towards the body than away from the body whenvision was available for online control (Tukey HSD,P < .05; see Fig. 5a and c). Importantly, though, diVerences
in the proportional time spent before and after peak velocitywere small compared to the diVerences in peak velocity perse. Thus, the lower peak velocities produced by tetraplegicsappears to do a better job of explaining the increased move-ment time than the symmetry of the velocity proWles.
Another approach to isolating the relative contribution ofprior planning and feedback-dependent processes to limbcontrol involves examining trial-to-trial consistency in theway movement unfolds over the course of a number of aim-ing trajectories (Khan et al. 2006). The idea is that if limbmovements unfold without the beneWt of online control,spatial variability will increase in a linear fashion over thecourse of the movement and be highest when the movementends. The increase in the standard deviation should beroughly proportional to the mean amplitude achieved by thelimb at that point in time. If, however, participants engagein online control, variability should increase up until thepoint where online processes can eVectively reduce vari-ability due to programming error and neuromuscular noise.
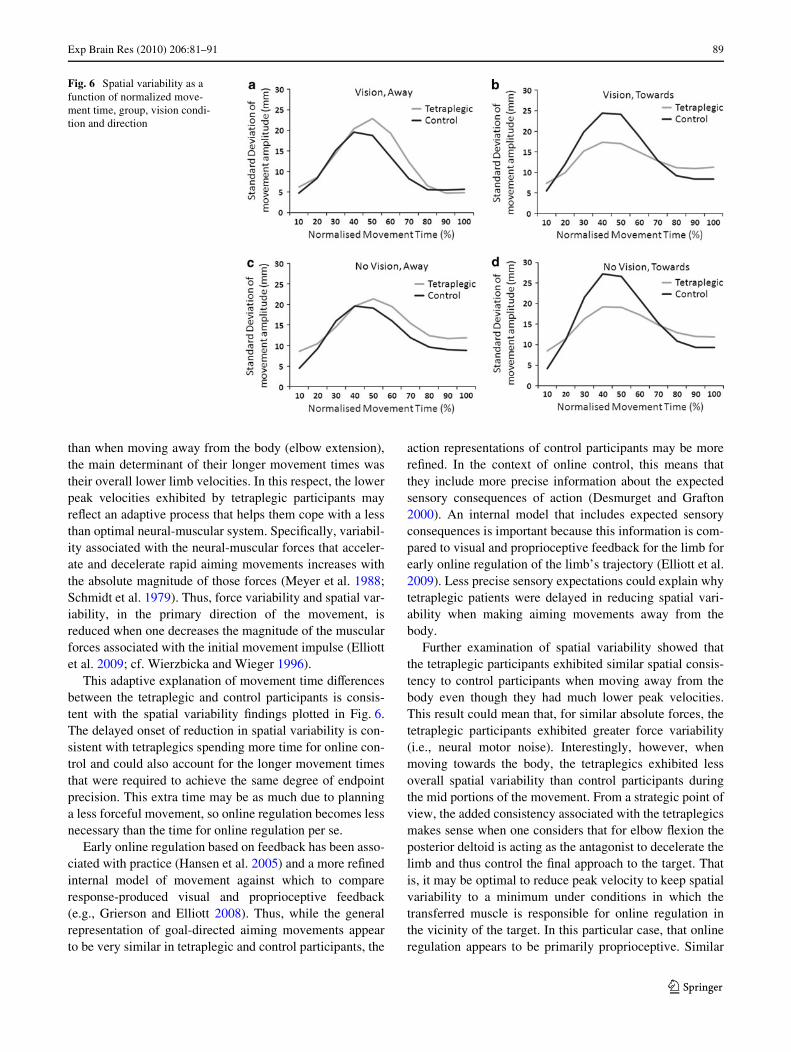
Analysis of the standard deviations in spatial positionyielded a main eVect for Movement Time Interval,F(9,72) = 41.09, P < .001, as well as interactions involvingVision Condition, Direction and Movement Time Interval,F(9,72) = 4.71, P < .001, and Group, Direction and Move-ment Time Interval, F(9,72) = 4.02, P < .001. In Fig. 6, wehave plotted the results as a function of all four variables.The most striking aspect of these data is the large increasein variability over the Wrst 40–60% of the movement fol-lowed by a dramatic decrease thereafter. The Vision Condi-tion by Direction by Movement Time Interval interactionshows that vision plays a more important role in thisdecrease when the limb is moving away from the body.SpeciWcally, there is a more pronounced reduction in spatialvariability between the middle and the end of the move-ment when moving away from the body resulting in greaterconsistency at movement termination (e.g., Fig. 6a vs. c).This diVerential eVect of vision, for movements away fromthe body, was evident in both groups, and the Wnding isconsistent with the variable error results at the end of themovement. However, the Group by Direction by Move-ment Time Interval interaction indicates that, independentof vision, the pattern of variability throughout a trial wasqualitatively diVerent between the groups depending onwhether the movements were made away from (elbowextension, Fig. 6a and c) or towards the body (elbow Xex-ion, Fig. 6b and d). When moving away from the body,the two variability proWles are quite similar except that thereduction in variability is delayed by at least 10% of themovement for tetraplegics relative to control participants.Although this may not seem like a very big diVerence,given that the control participants executed their move-ments, on average, 182 ms more quickly than tetraplegicparticipants, the real-time diVerence between the two
123
Exp Brain Res (2010) 206:81–91 87
Fig. 3 Velocity proWles from all 10 participants plotted as a func-tion of vision condition. The proWles are plotted in pairs mov-ing from the two fastest partici-pants in each group, at the top of the page, to the two slowest participants, at the bottom of the page
123
88 Exp Brain Res (2010) 206:81–91
groups before there was a reduction in variability is morethan 200 ms when completing elbow extensions.
Discussion
The primary purpose of this work was to investigate reach-ing/aiming trajectories of tetraplegic and age-equivalent
control participants. In particular, we were interested inwhether or not the surgically based muscle arrangementimpacts the manner in which precision limb control isachieved. A secondary purpose of the experiment was todetermine whether using an elbow extensor, not speciWcallydesigned for that task, increased the need for online visualregulation of the limb trajectory.
As anticipated, participants who had undergone muscu-lotendinous transfer surgery were slower than controlparticipants in completing goal-directed movements tosmall targets regardless of whether the aiming movementinvolved elbow Xexion or extension (see also LaVont et al.2007). However, the tetraplegic participants completedtheir movements with a similar degree of movement accu-racy, and they took no more time than control participantsto plan and initiate their aiming movement. As well, theaiming trajectories of participants in the two groups weresurprisingly similar. Although there was reasonable vari-ability within both groups of participants, most participantsexhibited a modest asymmetry in their velocity proWles,spending slightly more real and proportional time deceler-ating the limb than accelerating the limb. This type ofvelocity proWle is typical of aiming movements performedto small targets where extra time is required during deceler-ation to adjust the limb trajectory during the Wnal approachto the target (Elliott et al. 2001; Elliott and Hansen 2010).Although tetraplegic participants did spend slightly moretime when vision was available decelerating their aimingmovements when moving towards the body (elbow Xexion)
Fig. 5 Percentage time to peak velocity and time after peak velocity as a function of group, vision condition and direction. The error bars represent one standard deviation
Fig. 4 Time to and time after peak velocity as a function of group. Theerror bars represent one standard deviation
123
Exp Brain Res (2010) 206:81–91 89
than when moving away from the body (elbow extension),the main determinant of their longer movement times wastheir overall lower limb velocities. In this respect, the lowerpeak velocities exhibited by tetraplegic participants mayreXect an adaptive process that helps them cope with a lessthan optimal neural-muscular system. SpeciWcally, variabil-ity associated with the neural-muscular forces that acceler-ate and decelerate rapid aiming movements increases withthe absolute magnitude of those forces (Meyer et al. 1988;Schmidt et al. 1979). Thus, force variability and spatial var-iability, in the primary direction of the movement, isreduced when one decreases the magnitude of the muscularforces associated with the initial movement impulse (Elliottet al. 2009; cf. Wierzbicka and Wieger 1996).
This adaptive explanation of movement time diVerencesbetween the tetraplegic and control participants is consis-tent with the spatial variability Wndings plotted in Fig. 6.The delayed onset of reduction in spatial variability is con-sistent with tetraplegics spending more time for online con-trol and could also account for the longer movement timesthat were required to achieve the same degree of endpointprecision. This extra time may be as much due to planninga less forceful movement, so online regulation becomes lessnecessary than the time for online regulation per se.
Early online regulation based on feedback has been asso-ciated with practice (Hansen et al. 2005) and a more reWnedinternal model of movement against which to compareresponse-produced visual and proprioceptive feedback(e.g., Grierson and Elliott 2008). Thus, while the generalrepresentation of goal-directed aiming movements appearto be very similar in tetraplegic and control participants, the
action representations of control participants may be morereWned. In the context of online control, this means thatthey include more precise information about the expectedsensory consequences of action (Desmurget and Grafton2000). An internal model that includes expected sensoryconsequences is important because this information is com-pared to visual and proprioceptive feedback for the limb forearly online regulation of the limb’s trajectory (Elliott et al.2009). Less precise sensory expectations could explain whytetraplegic patients were delayed in reducing spatial vari-ability when making aiming movements away from thebody.
Further examination of spatial variability showed thatthe tetraplegic participants exhibited similar spatial consis-tency to control participants when moving away from thebody even though they had much lower peak velocities.This result could mean that, for similar absolute forces, thetetraplegic participants exhibited greater force variability(i.e., neural motor noise). Interestingly, however, whenmoving towards the body, the tetraplegics exhibited lessoverall spatial variability than control participants duringthe mid portions of the movement. From a strategic point ofview, the added consistency associated with the tetraplegicsmakes sense when one considers that for elbow Xexion theposterior deltoid is acting as the antagonist to decelerate thelimb and thus control the Wnal approach to the target. Thatis, it may be optimal to reduce peak velocity to keep spatialvariability to a minimum under conditions in which thetransferred muscle is responsible for online regulation inthe vicinity of the target. In this particular case, that onlineregulation appears to be primarily proprioceptive. Similar
Fig. 6 Spatial variability as a function of normalized move-ment time, group, vision condi-tion and direction
123

90 Exp Brain Res (2010) 206:81–91
types of “play-it-safe” behaviours have been reported innormal elderly performers who must also learn to live witha noisier neural motor system (Welsh et al. 2007). It is alsothe case that tetraplegic participants have an impairedcapacity for force production associated with elbow exten-sion and the deceleration of elbow Xexion movements incertain upper arm postures due to the new bi-articulararrangement of the posterior deltoid. The contribution tomotion at two joints means that shortening of the muscleduring shoulder and elbow extension is less favourable forforce production than when the shoulder and elbow areXexed (Kirsch et al. 1996). These length-tension dependen-cies may represent an additional contribution to the lowerpeak velocities exhibited by this group.
Overall, the kinematic Wndings suggest a high degree ofeVector independence and thus are consistent with thenotion that discrete goal-directed movements are repre-sented in a rather general fashion (e.g., MacKay 1982;Swinnen et al. 2010). Irrespective of which muscle groupsact as the agonist and antagonist, there was little diVerencein the proportional timing of tetraplegic and control partici-pants. Instead, it would seem that participants from thepatient group simply scale down the absolute muscularforces associated with limb acceleration and deceleration inorder to reduce the need for online regulation. In this samecontext, tetraplegic participants were no more dependent onvisual feedback for limb regulation than control partici-pants. In fact, when moving towards the body, both groupsof participants terminated their movements with a similardegree of consistency regardless of whether or not visionwas eliminated upon movement initiation. As mentionedabove, this similarity may be related to the tetraplegicsadopting limb control processes designed to reduce impulsevariability and associated spatial variability and thus theneed for visual online regulation (Elliott et al. 2001; Meyeret al. 1988).
Conclusions
In summary, tetraplegics exhibited less forceful but equallyprecise movements to control participants. Thus, for tetra-plegics, the proportional timing of acceleration–decelera-tion was similar to control participants even though theabsolute time to complete the aiming movements waslonger. This outcome is consistent with the notion of eVec-tor independence, and the idea that discrete aiming move-ments is represented in a general fashion. Perhaps thetetraplegic participants moved more slowly because of thereduced force-generating capability of their elbow exten-sors. Alternatively, the tetraplegic participants may havelearned to adopt a limb control strategy that minimizes thespatial variability associated with a noisier neural muscular
system. Thus, the limb performs much the same way a lesspractised eVector system (e.g., the non-preferred arm)might perform in a typical adult who has not suVered spinalcord injury and undergone musculotendinous transfer sur-gery.
Acknowledgments This work was supported by the ResearchInstitute for Sport and Exercise Sciences at Liverpool John MooresUniversity, and the Natural Sciences and Engineering ResearchCouncil of Canada. The authors wish to express their thanks toMrs. Barbara Hoole and Mr. Pradip Sett of the North West RegionalSpinal Injuries Centre, Southport and Ormskirk Hospital NHS Trustfor their assistance with the recruitment of participants.
References
Carson RG, Elliott D, Goodman D, Thyer L, Chua R, Roy EA (1993)The role of impulse variability in manual aiming asymmetries.Psych Res 55:291–298
Desmurget M, Grafton S (2000) Forward modelling allows feedbackcontrol for fast reaching movements. Trends Cogn Sci 4:423–431
Dounskaia N (2005) The internal model and the leading joint hypoth-esis: implications for control of multi-joint movements. ExpBrain Res 166:1–16
Elliott D, Allard F (1985) The utilization of visual feedback informationduring rapid pointing movements. Q J Exp Psych 37A:407–425
Elliott D, Hansen S (2010) Visual regulation of manual aiming: a com-parison of methods. Behav Res Methods (in press)
Elliott D, Roy EA (1989) Manual asymmetries in aimed movements.Q J Exp Psych 41A:501–516
Elliott D, Helsen WF, Chua R (2001) A century later: Woodworth’stwo component model of goal directed aiming. Psych Bull127:342–357
Elliott D, Hansen S, Grierson LE (2009) Optimising speed and energyexpenditure in accurate visually directed upper limb movements.Ergonomics 52:438–447
Friden J, Lieber RL (2001) Quantitative evaluation of the posteriordeltoid to triceps tendon transfer based on muscle architecturalproperties. J Hand Surg 26A:147–155
Grierson LE, Elliott D (2008) Kinematic analysis of goal-directed aimsmade against early and late perturbations: an investigation of therelative inXuence of two online control processes. Hum Move-ment Sci 27:839–856
Hansen S, Tremblay L, Elliott D (2005) Part and whole practice:chunking and online control in the acquisition of a serial motortask. Res Q Exercise Sport 76:60–67
Hansen S, Glazebrook CM, Anson JG, Weeks DJ, Elliott D (2006) TheinXuence of advance information about target knowledge andvisual feedback on movement planning and execution. Can J ExpPsychol 60:200–208
Heath M, Binsted G (2007) Visuomotor memory for target location innear and far reaching space. J Motor Behav 39:169–177
HoVman G, LaVont I, Hanneton S, Roby-Brami A (2006) How toextend the elbow with a weak or paralyzed triceps: control of armkinematics for aiming in C6–C7 quadriplegic patients. Neurosci-ence 139:749–765
Keele SW (1968) Movement control in skilled performance. PsychBull 70:387–403
Kelso JAS, Fuchs A, Lancaster R, Holroyd T, Cheyne D, Weinberg H(1998) Dynamic cortical activity in the human brain revealsmotor equivalence. Nature 392:814–818
Khan MA, Franks IM, Elliott D, Lawrence GP, Chua R, Bernier P-M,Hansen S, Weeks DJ (2006) Inferring online and oZine processing
123

Exp Brain Res (2010) 206:81–91 91
of visual feedback in target directed movements from kinematicdata. Neurosci Biobehav Rev 30:1106–1121
Kirsch RF, Acosta AM, Perreault EJ, Keith MW (1996) Measurementof isometric elbow and shoulder moments: position-dependentstrength of the posterior deltoid-to-triceps muscle tendon transferin tetraplegia. IEEE T Rehabil Eng 4:403–409
LaVont I, Briand E, Dizien O, Combeaud M, Bussel B, Revol M,Rody-Brami A (2000) Kinematics of prehension and pointingmovements in C6 quadriplegic patients. Spinal Cord 38:354–362
LaVont I, HoVman G, Dizien O, Revol M, Roby-Brami A (2007) Howdo C6/C7 tetraplegic patients grasp balls of diVerent sizes andweights? Impact of surgical musculo-tendinous transfers. SpinalCord 45:502–512
Levin O, Wenderoth N, Steyvers M, Swinnen SP (2003) Directionalinvariance during loading-related modulations of muscle activity:evidence for motor equivalence. Exp Brain Res 148:62–76
MacKay D (1982) The problem of Xexibility and Xuency in skilledbehaviour. Psych Review 89:483–506
Maraj BKV, Elliott D, Lee TD, Pollock BJ (1993) Variance and invari-ance in expert and novice triple jumpers. Res Q Exercise Sport64:404–412
Marteniuk RG, Ivens CJ, Bertram CP (2000) Evidence of motor equiv-alence in a pointing task involving locomotion. Mot Control4:165–184
Merton PA (1972) How we control the contraction of our muscles. SciAm 226:30–37
Meyer DE, Abrams RA, Kornblum S, Wright CE, Smith JEK (1988)Optimality in human motor performance: ideal control for rapidaimed movements. Psychol Rev 95:340–370
Park J-H, Shea CH (2002) EVector independence. J Motor Behav34:253–270
Park J-H, Shea CH (2003) EVect of practice on eVector independence.J Motor Behav 35:33–40
Raibert MH (1977) Motor control and learning in a state space model.(Tech. Rep. AI-M-351; Massachusetts Institute of Technology,NTIS AD-A026–960). MIT, Cambridge, MA
Remy-Neris O, Milcamps J, Chiki-Keromost R, Thevenon A, BouttenD, Bouillard S (2003) Improved kinematics of unrestricted armraising in C5–C6 tetraplegic subjects after deltoid-to-tricepstransfer. Spinal Cord 41:435–445
Robinson MA, Barton GJ, Lees A, Sett P (2010) Analysis of tetraple-gic reaching in their 3D workspace following posterior deltoid-triceps tendon transfer. Spinal Cord 48:619–627
Schmidt RA (1975) A schema theory of discrete motor skill learning.Psychol Rev 82:225–260
Schmidt RA, Zelaznik HN, Hawkins B, Frank JS, Quinn JT (1979)Motor output variability: a theory for the accuracy of rapid motoracts. Psychol Rev 86:415–451
Swinnen SP, Vangheluwe S, Wagemans J, Coxon JP, Goble DJ, VanImpe A, Sunaert S, Peeters R, Wenderoth N (2010) Shared neuralresources between left and right inter limb coordination skills: theneural substrate of abstract motor representations. NeuroImage49:2570–2580
Vangheluwe S, Suy E, Wenderoth N, Swinnen SP (2006) Learning andtransfer of bimanual multifrequency patterns: eVector-indepen-dent and eVector-speciWc levels of movement representation. ExpBrain Res 170:543–554
Welsh TN, Higgins L, Elliott D (2007) Are there age-related diVer-ences in learning to optimise speed, accuracy and energy expen-diture? Hum Movement Sci 26:892–912
Wierzbicka MM, Wieger AW (1996) Accuracy of motor responses insubjects with and without control of the antagonist muscle.J Neurophysiol 75:2533–2541
Wierzbicka MM, Wiegner AW (1992) EVects of weak antagoniston fast elbow Xexion movements in man. Exp Brain Res91:509–519
Wright CE (1990) Generalized motor programs: re-examiningclaims of eVector independence in writing. In: Jeannerod M(ed) Attention and performance XIII. Erlbaum, Hillsdale, NJ,pp 294–320
123
Related Documents











