Selenium (Se) Seed Priming Induced Growth and Biochemical Changes in Wheat Under Water Deficit Conditions Fahim Nawaz & M. Yasin Ashraf & Rashid Ahmad & Ejaz Ahmad Waraich Received: 30 October 2012 / Accepted: 19 November 2012 # Springer Science+Business Media New York 2012 Abstract Insufficient stand establishment at early growth stages in wheat (Triticum aestivum L.) due to drought stress is a major problem that limits overall efficiency and yield of crop. Priming of seed is an effective method for raising seed performance and improving tolerance of crops to abiotic stresses especially drought. The seeds of two local wheat cultivars (Kohistan-97 and Pasban-90) were soaked in distilled water or sodium selenate solu- tions of 25, 50, 75, and 100 μM for 1/2 or 1 h at 25 °C and later re-dried to their original moisture levels before sowing. One-hour priming significantly increased root length stress tolerance index, dry matter stress tolerance index, and total biomass of seedlings; however, no sig- nificant effect of changing duration of Se seed priming was observed on plant height stress tolerance index and shoot/root ratio. Among cultivars, Kohistan-97 was found to be more responsive to Se seed treatment as 1 h priming at 100 μM significantly increased its total biomass by 43 % as compared to control treatment. Although biomass of seedlings was not affected with Se seed priming under normal conditions, but it increased significantly with increase in rates of Se under drought stress conditions. One-hour priming at 75 μM increased the total sugar content and total free amino acids in both wheat cultivars. A more significant decrease in soluble proteins of seedlings was observed by 1 h priming than 1/2 h priming under drought stress conditions. Keywords Selenium seed priming . Mitigation of drought . Crop stand . Physiological indices . Biochemical changes Introduction Drought stress is one of the major limitations to agri- cultural productivity around the globe [1]. It results in impaired germination and seedling growth [2], influen- ces plant growth [3–5], thus reducing fitness and func- tion of plants. A rapid increase in the world population is likely to aggravate the adverse effects of drought [6]. The severity, duration, and timing of water stress are extremely important for better understanding of plant responses to drought stress [7]. The mechanism of osmotic adjustment for drought tolerance by plants has been well documented. It is facilitated by compatible solutes produced at higher levels under limited water conditions [8–10]. The accu- mulation of these compounds in high amounts acts as osmoprotectants of membrane and prevents protein dis- integration [11]. The accumulation of total sugars and other compatible solutes such as polyols is a character- istic feature of most plants under stress [12, 13]. Priming of plant seeds is an easy, low-cost, low-risk, and effective approach to improve plant tolerance under stressful environments [14, 15]. It is a controlled hydra- tion process that promotes metabolic activities before radical protrusion [16, 17]. A number of osmotica have been reported to enhance germination, emergence, M. Y. Ashraf (*) Crop Stress Management Group, Nuclear Institute for Agriculture and Biology (NIAB), Jhang Road, Faisalabad, Pakistan e-mail: [email protected] F. Nawaz : R. Ahmad : E. A. Waraich Department of Crop Physiology, University of Agriculture, Faisalabad, Pakistan Biol Trace Elem Res DOI 10.1007/s12011-012-9556-9

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Selenium (Se) Seed Priming Induced Growth and BiochemicalChanges in Wheat Under Water Deficit Conditions
Fahim Nawaz & M. Yasin Ashraf & Rashid Ahmad &
Ejaz Ahmad Waraich
Received: 30 October 2012 /Accepted: 19 November 2012# Springer Science+Business Media New York 2012
Abstract Insufficient stand establishment at early growthstages in wheat (Triticum aestivum L.) due to droughtstress is a major problem that limits overall efficiencyand yield of crop. Priming of seed is an effective methodfor raising seed performance and improving tolerance ofcrops to abiotic stresses especially drought. The seeds oftwo local wheat cultivars (Kohistan-97 and Pasban-90)were soaked in distilled water or sodium selenate solu-tions of 25, 50, 75, and 100 μM for 1/2 or 1 h at 25 °Cand later re-dried to their original moisture levels beforesowing. One-hour priming significantly increased rootlength stress tolerance index, dry matter stress toleranceindex, and total biomass of seedlings; however, no sig-nificant effect of changing duration of Se seed primingwas observed on plant height stress tolerance index andshoot/root ratio. Among cultivars, Kohistan-97 was foundto be more responsive to Se seed treatment as1 h priming at 100 μM significantly increased its totalbiomass by 43 % as compared to control treatment.Although biomass of seedlings was not affected with Seseed priming under normal conditions, but it increasedsignificantly with increase in rates of Se under droughtstress conditions. One-hour priming at 75 μM increasedthe total sugar content and total free amino acids in bothwheat cultivars. A more significant decrease in soluble
proteins of seedlings was observed by 1 h priming than1/2 h priming under drought stress conditions.
Keywords Selenium seed priming . Mitigation of drought .
Crop stand . Physiological indices . Biochemical changes
Introduction
Drought stress is one of the major limitations to agri-cultural productivity around the globe [1]. It results inimpaired germination and seedling growth [2], influen-ces plant growth [3–5], thus reducing fitness and func-tion of plants. A rapid increase in the world populationis likely to aggravate the adverse effects of drought [6].The severity, duration, and timing of water stress areextremely important for better understanding of plantresponses to drought stress [7].
The mechanism of osmotic adjustment for droughttolerance by plants has been well documented. It isfacilitated by compatible solutes produced at higherlevels under limited water conditions [8–10]. The accu-mulation of these compounds in high amounts acts asosmoprotectants of membrane and prevents protein dis-integration [11]. The accumulation of total sugars andother compatible solutes such as polyols is a character-istic feature of most plants under stress [12, 13].
Priming of plant seeds is an easy, low-cost, low-risk,and effective approach to improve plant tolerance understressful environments [14, 15]. It is a controlled hydra-tion process that promotes metabolic activities beforeradical protrusion [16, 17]. A number of osmotica havebeen reported to enhance germination, emergence,
M. Y. Ashraf (*)Crop Stress Management Group, Nuclear Institute for Agricultureand Biology (NIAB), Jhang Road,Faisalabad, Pakistane-mail: [email protected]
F. Nawaz :R. Ahmad : E. A. WaraichDepartment of Crop Physiology, University of Agriculture,Faisalabad, Pakistan
Biol Trace Elem ResDOI 10.1007/s12011-012-9556-9

growth, and/or grain yield of wheat like potassium chlo-ride [18], polyethylene glycol, and potassium dihydrogenphosphate monobasic solutions [19].
The specific physiological mechanisms that underliethe positive effects of selenium (Se) in plants have notbeen clearly elucidated [20]. It is beneficial to plantsbecause it enhances plant growth [21] and promotesproduction of antioxidants in senescing plants [22]. It ismostly used for the biofortification of crops but has alsobeen reported to play an important role in the adjustmentof plant water status under drought stress [23]. Itimproves germination and radicle elongation in plants[24]. The trace amounts of Se have been reported tostimulate growth in lettuce [20], ryegrass [25], Brassicaoleracea [26], and potato [27]. It is well documented thatsoil pH, organic matter content, precipitation, soil texture,and soil sulfur (S) status adversely affect Se uptake inplants [28, 29], so Se seed treatment can be an effectivetechnique not only to improve the Se concentration in
food crops but can also prove beneficial in alleviating thedrastic effects of drought. Priming of seeds with Se hasbeen reported to increase germinability and antioxidativeresponses in bitter gourd [30], but very little or no workhas been done to evaluate the role of Se seed priming inimproving drought tolerance in wheat.
The present study was carried out with the objectives todetermine the rate of priming wheat seeds with selenium inalleviating the adverse effects of drought stress and to findout optimum rate and duration of selenium priming ofwheat seeds.
Materials and Methods
Seed Material and Experimental Design
Seeds of two local recommended spring wheat cultivarsviz. Kohistan-97 and Pasban-90 were obtained from
Fig. 1 a, b Effect of seedpriming with Se on PHSI inwheat under normal (no stress)and drought (water stress)conditions. Priming treatmentsinclude priming with distilledwater (DW) and priming with25, 50, 75, and 100 μM Se.Values are mean±standard error
Nawaz et al.

Ayub Agricultural Research Institute, Faisalabad, Paki-stan. The experiments were carried out at Nuclear Insti-tute for Agriculture and Biology, Faisalabad, Pakistan.The experiments were laid out in a completely random-ized design with three repeats. Thirty seeds of eachcultivar were sterilized for 5 min with 5 % sodiumhypochlorite solution and washed three times with dis-tilled water. The seeds were soaked in 0, 25, 50, 75,and 100 μM of Na2SeO4 (Sigma-Aldrich, USA) solu-tions for 30 min (1/2 h) or 1 h at 25 °C and later re-dried to their original moisture level. Initially, in eachreplication, ten seeds of each cultivar were sown inplastic pots (15×10 cm) containing sand as growthmedium, but after germination, only five plants were
kept in each replication. The experiment was repeatedthree times to record data regarding physiological indi-ces and total biomass of plants.
Drought stress was imposed by withholding water for2 weeks after seedling emergence in one set of pots,while 100 % field capacity (control) level was main-tained in other set of pots by re-watering the pots. Theplants were harvested after 3 weeks to collect data onroot length, shoot length, and plant biomass. Plant dryweights were recorded after oven drying the samples at70±2 °C.
The following formulae as described by Ashraf et al. [31]were used for the calculation of plant height stress toleranceindex (PHSI), dry matter stress tolerance index (DMSI), and
Fig. 2 a, b Effect of seedpriming with Se on RLSI inwheat under normal (no stress)and drought (water stress)conditions. Priming treatmentsinclude priming with distilledwater (DW) and priming with25, 50, 75, and 100 μM Se.Values are mean±standard error
Selenium (Se) Seed Priming Induced Growth and Biochemical Changes

root length stress tolerance index (RLSI).
PHSI ¼ ½Plant height of stressed plant
=Plant height of control plant� � 100
DMSI ¼ ½Dry matter of stressed plant
=Dry matter of control plant� � 100
RLSI ¼ ½Root length of stressed plant
=Root length of control plant� � 100
Fresh weight of the shoot and roots was recordedimmediately after harvesting to determine total biomass
and shoot/root ratio of seedlings. The shoot/root ratiowas calculated on fresh weight basis.
Determination of Biochemical Attributes
Total sugars were determined according to the methodof Riazi et al. [32], protein as described by Lowry et al.[33], and total free amino acids according to the methodof Hamilton and Van Slyke [34].
Statistical Analysis
Least Significant Difference Test (P<0.05) was used toevaluate the significant differences between cultivars.MSTAT-C software package was used for the statisticalanalysis of the data.
Fig. 3 a, b Effect of seedpriming with Se DMSI in wheatunder normal (no stress) anddrought (water stress)conditions. Priming treatmentsinclude priming with distilledwater (DW) and priming with25, 50, 75, and 100 μM Se.Values are mean±standard error
Nawaz et al.
Results
Seedling Growth
All the physiological indices were significantly (P≤0.05) influenced by the Se (Se) seed priming of wheatunder drought conditions. However, a non-significantdifference was recorded for duration of Se seed primingwith regard to plant height stress tolerance index(PHSI), but a gradual increase in PHSI was noted withincrease in concentration of Se used for seed primingfor both the durations, i.e., 1/2 and 1 h (Fig. 1a, b). Themaximum value for PHSI was recorded in Kohistan-97in case of seed primed with 100 μM of Se solution for1 h (Fig. 1b) which was statistically at par with 50 and75 μM Se in Kohistan-97 and 75 and 100 μM Se inPasban-90 (Fig. 1a, b).
The duration of Se seed priming significantly (P≤0.01) affected the root length stress tolerance index(RLSI). The highest value was obtained in wheat seed-lings where seed priming was done with Se for 1 h(Fig. 2b). The soaking of seeds in Se solutions for 1/2 h
had no significant effect on RLSI of both the wheatcultivars; however, seed soaking for 1 h significantlyincreased the RLSI, and it was the highest in Kohistan-97 seeds primed at 100 μM Se and was statistically atpar with 75 μM Se. Wheat cultivar Pasban-90 gainedthe highest value for RLSI by seed priming with100 μM Se, while other lower rates failed to increasethe RLSI in both the cultivars (Fig. 2a, b). Similar toRLSI, Se seed priming duration significantly (P≤0.01)improved dry matter stress tolerance index (DMSI) andwas the maximum in seedlings where seed priming with100 μM Se was done for both wheat cultivars, whilelower doses of Se and 1/2 h priming did not improvethe DMSI (Fig. 3a, b).
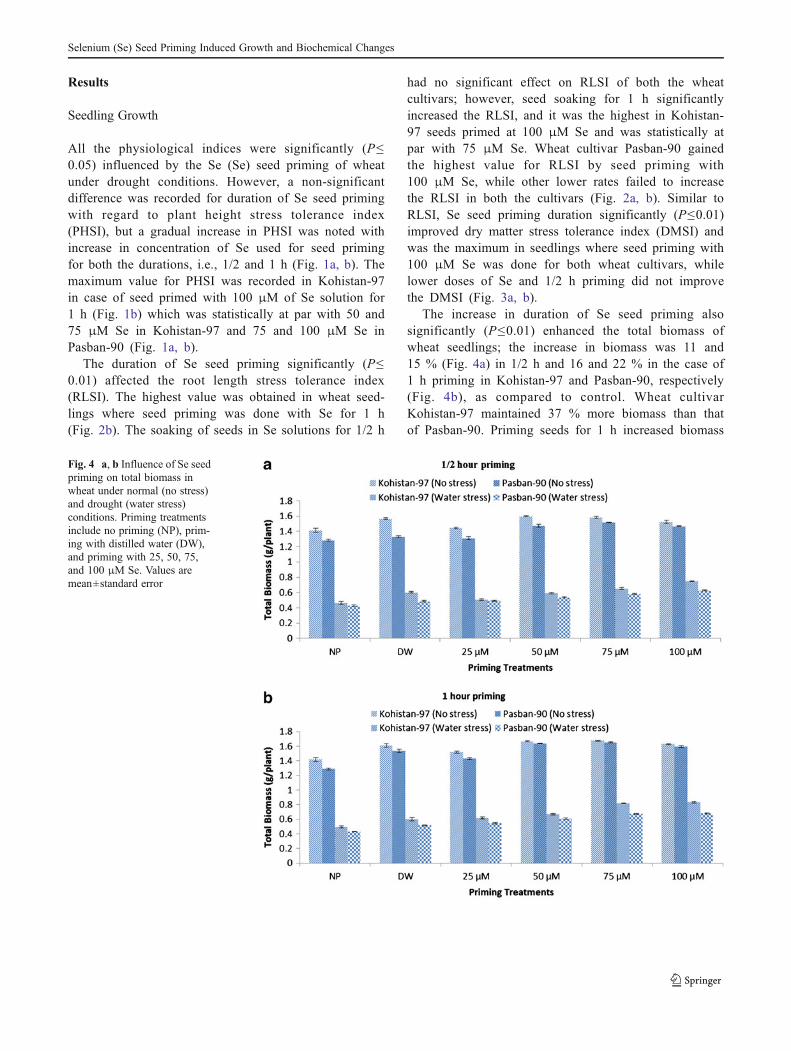
The increase in duration of Se seed priming alsosignificantly (P≤0.01) enhanced the total biomass ofwheat seedlings; the increase in biomass was 11 and15 % (Fig. 4a) in 1/2 h and 16 and 22 % in the case of1 h priming in Kohistan-97 and Pasban-90, respectively(Fig. 4b), as compared to control. Wheat cultivarKohistan-97 maintained 37 % more biomass than thatof Pasban-90. Priming seeds for 1 h increased biomass
Fig. 4 a, b Influence of Se seedpriming on total biomass inwheat under normal (no stress)and drought (water stress)conditions. Priming treatmentsinclude no priming (NP), prim-ing with distilled water (DW),and priming with 25, 50, 75,and 100 μM Se. Values aremean±standard error
Selenium (Se) Seed Priming Induced Growth and Biochemical Changes

of Kohistan-97 up to 43 % when compared withPasban-90. Under normal conditions, seed priming withSe failed to improve the total biomass of seedlings;however, a non-significant (3–4 %) reduction in bio-mass was observed at higher rates of Se for 1/2 and 1 hseed priming durations. But reverse was the case with biomassaccumulation under water stress conditions. Maximum bio-mass was recorded for wheat cultivar Kohistan-97 at 100 μMSe seed priming for 1 h (Fig. 4b) which was statistically at parwith 75 μM Se. Wheat cultivar Kohistan-97 had 18 % morebiomass than Pasban-90 at the same treatment of 100 μM Se,while it was 43 % more as compared to control treatment(Fig. 4b). The Se seed treatments for 1/2 and 1 h had nosignificant effect (P≤0.05) on shoot/root ratio of seedlings ofboth the cultivars (Fig. 5a, b).
Biochemical Attributes
Total free amino acids and soluble sugar contents weresignificantly (P<0.01) increased, but total proteins of theseedlings decreased sharply due to limited water supply in
both the wheat cultivars. Under water deficit conditions,seedlings of wheat cultivars, Pasban-90 and Kohistan-97,developed from the seed primed with 75 μM Se solution,maintained 75 and 64 %, respectively, higher soluble sugarsthan their controls (Fig. 6a, b).
Accumulation of free amino acids was significantlyincreased under drought stress conditions, and Se seedpriming treatment further enhanced the accumulation inthe seedlings. It was 61 % in Kohistan-97 and 64 % inPasban-90 under drought conditions in seedlings raisedfrom seeds primed with 75 μM Se for 1 h; however, thefree amino acids accumulation varied non-significantlybetween both the durations (Fig 7a, b).
The total soluble proteins of the seedlings decreasedsignificantly in both 1/2 and 1 h seed priming durationsat higher rates of selenium seed priming under droughtstress, while lower rates did not affect them significant-ly (Fig 8a, b). Seed priming for 1/2 h with 50 μM Sedecreased total soluble proteins of Kohistan-97 up to32 %, while it was 40 % in seedlings primed with75 μM Se solution under moisture deficit conditions.
Fig. 5 a, b Effect of seedpriming with selenium onshoot/root ratio in wheat undernormal (no stress) and drought(water stress) conditions.Priming treatments include nopriming (NP), priming withdistilled water (DW), andpriming with 25, 50, 75, and100 μM Se. Values are mean±standard error
Nawaz et al.

In wheat cultivar Pasban-90, reduction in protein con-tent increased with the increase in Se concentration ofseed priming solution under drought. The reduction was30 and 40 % under 1/2 and 1 h seed priming, respec-tively, in seedlings of Pasban-90 developed from seedsprimed with 100 μM Se (Fig 8a, b).
Discussion
The results indicate that wheat seedling growth which isadversely influenced by drought stress is due to reduc-tion in germination and seedling establishment [35].Present findings confirm the hypothesis that Se seedpriming is effective in improving drought tolerance inwheat seedlings by enhancing germination. Since Se hasbeen reported to be toxic to plants at relatively lowconcentrations [36] and higher rates of Se may inhibitphotosynthesis, impair nutrient uptake, and transport[37], therefore, the duration of seed priming is kept
for 1/2 and 1 h to evaluate any possible hazardouseffects of Se on growth of seedlings.
Higher concentrations of Se (100 μM) do not affectgrowth of seedlings under normal supply of water, butit significantly increases the growth under drought stressconditions which means that Se improves plant–waterrelations by lowering the osmotic potential of the seed-lings growing under water stress [38]. The differentialresponse of both wheat cultivars may be due to theirgenetic potential [39].
Although, the duration of Se seed priming has non-significant effect on plant height stress tolerance index(PHSI), but Se treatments significantly increase PHSIof both the cultivars as compared to control treatmentwhich indicates the role of Se in activation of planthormones involved in cell expansion and enlargement[40]. These results indicate that Se helps seedlings toregulate their water under drought stress conditionsand comply with the findings of Djanaguiraman [38].An increase in DMSI of seedlings by Se treatment(Fig. 3) under drought depicts its positive role in
Fig. 6 a, b Selenium seedpriming induced changes intotal soluble sugars in wheatunder normal (no stress) anddrought (water stress)conditions. Priming treatmentsinclude no priming (NP), prim-ing with distilled water (DW),and priming with 25, 50, 75,and 100 μM Se. Values aremean±standard error
Selenium (Se) Seed Priming Induced Growth and Biochemical Changes

increasing the dry weight of plants; however, an in-crease in RLSI is only observed at higher doses of Se(75 and 100 μM) in 1 h priming (Fig. 2) which con-firms the findings of Yao et al. [41] that root activity ofwheat seedlings increases by Se supply under droughtstress. Carlson et al. [42] found selenate-treated radiclesto be healthier and vigorous with extensive root hairs. Itis a well-documented fact that there exists a significantrelation between root and seedling dry weight underdrought stress [43, 44], and an increase in dry weightand root length at higher doses of Se supports this fact.It is observed that different rates of Se has no signifi-cant effect on root growth in 1/2 h seed priming, whileSe seed priming for 1 h significantly enhances rootactivity in seedlings under drought. Although a non-significant effect of higher or lower dose of Se onbiomass accumulation is observed by Yao et al. [41] inwheat seedlings under drought stress, but our resultsshow that treatment of seeds with higher doses of Sesignificantly increases biomass of wheat seedlings underdrought stress conditions.
The increase in the concentration of soluble sugars withthe increasing rates of Se seed priming indicates a positive
role of Se in regulating the water status of plants underdrought stress [38], and this sugar flux may act as a signalfor metabolic regulation under drought stress [45]. The highsoluble sugars in both the cultivars may be the result ofamylase activity that decomposes starch under droughtstress and therefore, increases soluble sugar contents [46].
The increase in total free amino acids by Se seed treat-ment under drought stress suggests that it may be due to thedisorder caused by Se in amino acid metabolism [47, 48].These results are in agreement with the findings of Khattab[49] who reported an increase in total free amino acids in thetops of rocket plants exposed to 5 or 100 μM selenate, whileHu et al. [50] reported similar results in selenite-treatedgreen tea.
The decrease in total soluble proteins under droughtstress is in accordance with the findings of Kochaki [51]and Sujin and RayWu [52] who reported that concen-tration of high molecular weight soluble proteinsdecreases while low molecular weight increases inwheat under drought stress and suggested that suchbiochemical attributes like total soluble proteins, totalsugars, and total free amino acids can be used for theselection of drought tolerant cultivars [53].
Fig. 7 a, b Selenium-mediatedchanges in total free aminoacids in wheat under normal (nostress) and drought (waterstress) conditions. Primingtreatments include no priming(NP), priming with distilledwater (DW), and priming with25, 50, 75, and 100 μM Se.Values are mean±standard error
Nawaz et al.

Conclusion
The results of the present study conclude that duration and rateof Se seed priming affect physiological indices, total biomass,and biochemical attributes of wheat seedlings. Lower doses ofSe do not significantly improve the growth of seedlings underlimited water conditions, and a significant improvement is onlyobserved at higher doses. Present study shows that the optimumduration for Se seed priming is 1 h and 75 μM is the optimumrate for enhancing drought tolerance in wheat seedlings
Acknowledgments The authors are grateful to the Higher EducationCommission (HEC) of Pakistan for the financial support provided for thisstudy. We are also thankful to Dr. Rob William, visiting professor fromEngland at National Institute for Biotechnology andGenetic Engineering,Faisalabad, Pakistan, for critically reading the manuscript.
References
1. Waraich EA, Ahmad R, Hur RGM, Ehsanullah, Ahmad A,Mahmood N (2011) Response of foliar application of KNO3 onyield, yield components and lint quality of cotton (Gossypiumhirsutum L.). Afric J Agric Res 24:5457–5463
2. Ashraf CM, Abu-Shakra S (1978) Wheat seed germination underlow temperature and moisture stress. Agron J 70:135–139
3. Jamieson PD, Martin RJ, Francis GS (1995) Drought influences ongrain yield of barley, wheat, and maize. NZL J Crop Horti 23:55–56
4. Tian X, Lei Y (2006) Nitric oxide treatment alleviates droughtstress in wheat seedlings. Biol Plant 50:775–778
5. Xu H, Biswas DK, Li WD, Chen SB, Zhang SB, Jiang GM, Li YG(2007) Photosynthesis and yield responses of ozone-polluted win-ter wheat to drought. Photosynthetica 45:582–588
6. Somerville C, Briscoe J (2001) Genetic engineering and water.Science 292:2217
7. Plaut Z (2003) Plant exposure to water stress during specificgrowth stages. Encyclopedia of Water Science, Taylor & Francis
8. Hasegawa P, Bressan RA, Zhu JK, Bohnert HJ (2000) Plantcellularand molecular responses to high salinity. Ann Rev PlantMol Biol 51:463–499
9. Zhu JK (2000) Salt and drought stress signal transduction in plants.Ann Rev Plant Biol 53:247–273
10. Shao HB, Liang ZS, Shao MA (2005) Changes of anti-oxidativeenzymes and MDA content under soil water deficits among 10wheat (Triticum aestivum L.) genotypes at maturation stage.Colloids Surf B Biointerfaces 45:7–13
11. Yancey PH (1994) Compatible and counteracting solutes. In:Strange K (ed) Cellularand molecular physiology of cell volumeregulation. Boca Raton, CRC Press, pp 81–109
12. Mohammadkhani N, Heidari R (2008) Effects of drought stress onsoluble proteins in two maize varieties. Turk J Bol 32:23–30
Fig. 8 a, b Effect of Se seedpriming on total solubleproteins in wheat under normal(no stress) and drought (waterstress) conditions. Primingtreatments include no priming(NP), priming with distilledwater (DW), and priming with25, 50, 75, and 100 μM Se.Values are mean±standard error
Selenium (Se) Seed Priming Induced Growth and Biochemical Changes

13. Delauney AJ, Verma DPS (1993) Proline biosynthesis and osmo-regulation in plants. Plant J 4:215–223
14. Ashraf M, Foolad MR (2005) Pre-sowing seed treatment—a shot-gun approach to improve germination, plant growth and crop yieldunder saline and non-saline conditions. Adv Agron 88:223–271
15. Wahid A, Shabbir A (2005) Induction of heat stress tolerance inbarley seedlings by pre-sowing seed treatment with glycinebetaine.Plant Growth Regul 46:133–141
16. Sivritepe N, Sivritepe HO, Eris A (2003) The effects of NaClpriming on salt tolerance in melon seedlings grown under salineconditions. Sci Hort 97:229–237
17. Sivritepe HO, Sivritepe N, Eris A, Turhan E (2005) The effects ofNaCl pre-treatment on salt tolerance of melons grown under long-term salinity. Sci Hort 106:568–581
18. Misra NM, Dwibedi DP (1980) Effects of pre-sowing seed treat-ments on growth and dry matter accumulation of high yieldingwheat under rainfed conditions. Ind J Agron 25:230–234
19. Dell’Aquila A, Taranto G (1986) Cell division and DNA synthesisduring osmopriming treatment and following germination in agedwheat embryos. Seed Sci Technol 14:333–341
20. Xue TL, Hartikainen H, Piironen V (2001) Antioxidative andgrowth-promoting effect of selenium on senescing lettuce. PlantSoil 237:555–561
21. Hartikainen H, Xue T (1999) The promotive effect of selenium onplant growth as triggered by ultraviolet irradiation. J Environ Qual28:1272–1275
22. Kuznetsov VV, Kholodova VP, Kuznetsov VIV, Yagodin BA(2003) Selenium regulates the water status of plants exposed todrought. Dok Biol Sci 390:266–268
23. Djanaguiraman M, Durga D, Shanker AK (2005) Selenium—anantioxidative protectant in soybean during senescence. Plant Soil272:77–86
24. Spencer NE, Siegel SM (1978) Effects of sulfur and seleniumoxyanions on Hg-toxicity in turnip seed germination. Water AirSoil Pollut 9:423–427
25. Hartikainen H, Xue T, Piironen V (2000) Selenium as an anti-oxidant and pro-oxidant in ryegrass. Plant Soil 225:193–200
26. Hajiboland R, Amjad L (2007) Does antioxidant capacity of leavesplay a role in growth response to selenium at different sulfurnutritional status? Plant Soil Environ 53:207–215
27. Seppänenn MM, Turakainen M, Hartikainen H (2003) Seleniumeffects on oxidative stress in potato. Plant Sci 165:311–319
28. Voutsa D, Samara C (1998) Dietary intake of trace elements andpolycyclic aromatic hydrocarbons via vegetables grown in anindustrial Greek area. Sci Total Environ 218:203–216
29. Luoma PV, Nayha S, Sikkila K, Hassi J (1995) High serumalpha-tocopherol, albumin, selenium and cholesterol, and lowmortality from coronary heart disease in northern Finland. JInt Med 237:49–54
30. Chen CC, Sung JM (2001) Priming bitter gourd seeds with sele-nium solution enhances germinability and antioxidative responsesunder sub-optimal temperature. Physiol Plant 111:9–16
31. Ashraf MY, Akhtar K, Hussain F, Iqbal J (2006) Screening ofdifferent accessions of three potential grass species fromCholistan desert for salt tolerance. Pak J Bot 38:1589–1597
32. Riazi A, Matruda K, Arslam A (1985) Water stress induceschanges in concentration of proline and other solutes in growingregions. J Exp Bot 36:1716–1725
33. Lowry OH, Rosrbrough NJ, Farr AL, Randall JR (1951) Proteinmeasurement with folin phenol reagent. J Biol Chem 191:265–275
34. Hamilton PB, Van Slyke DD (1943) Aminoacids determinationwith ninhydrin. J Biol Chem 150:231–233
35. Kaya MD, Okçub G, Ataka M, Çıkılıc Y, Kolsarıcıa Ö (2006) Seedtreatments to overcome salt and drought stress during germinationin sunflower (Helianthus annuus L.). Eur J Agron 24:291–295
36. Brown TA, Shrift A (1981) Exclusion of selenium from proteins ofselenium-tolerant Astragalus species. Plant Physiol 67:1051–1053
37. Kahle H (1988). Wirkung von blei und cadmium auf wachstumund mineralstoffhaushalt von jungbuchen (Fagus sylvatica L.) insandkultur. Dissertationes Botanicae 127, Berlin: J. Cramer.
38. Hartikainen H (2005) Biogeochemistry of selenium and its impacton food chain quality and human health. J Trace Elem Med Biol18:309–318
39. Eurola MH, Ekholm PI, Ylinen ME, Koivistoinen PE, Varo PT(1991) Selenium in Finnish foods after beginning the use ofselenate-supplemented fertilizers. J Sci Food Agric 56:57–70
40. Larson KL (1992) Drought injury and resistance of crop plants. In:Gupta SU (ed) Physiological aspects of dry land farming. Oxford& IBH Publishing Co Pvt Ltd, New Delhi, pp 147–162
41. Yao X, Chu J, Wang G (2009) Effects of selenium on wheatseedlings under drought stress. Biol Trace Elem Res 130:283–290
42. Carlson CL, Kaplan DI, Adriano DC (1989) Effects of selenium ongermination and radicle emergence of selected agronomic species.Environ Exp Bot 29:493–498
43. Okçu G, Kaya MD, Atak M (2005) Effects of salt and droughtstresses on germination and seedling growth of pea (Pisum sativumL.). Turk J Agric For 29:237–242
44. Yamur M, Kaydan D (2008) Alleviation of osmotic stress of waterand salt in germination and seedling growth of triticale with seedpriming treatments. Afr J Biotechnol 7:2156–2162
45. Kishor PBK, Sangama S, Amrutha RN, Laxmi PS, Naidu KR, RaoKS (2005) Regulation of proline biosynthesis degradation, uptakeand transport in higher plants: its implications in plant growth andabiotic stress tolerance. Curr Sci 88:424–438
46. Ghasempour HR, Gaff DF, Williams RPW, Gianello RD(1998) Contents of sugars in leaves of drying desiccationtolerant flowering plants, particularly grasses. Plant GrowthRegul 24:185–191
47. Gowily AM, Mahmoud MB, Abdel-Lateef MF, Razak AA,Ramadan TE (1996) Influence of selenium, TBZ and theirmixture on metabolic activities of some fungi. Afr J MycolBiotechol 4:45–56
48. Wu L (1998) Selenium accumulation and uptake by crop andgrassland plant species. In: Willia, T, Frankenberger Jr, EngbergRA (eds.) Environmental Chemistry of Selenium. Marcel Dekker,New York, pp 657–685
49. Khattab H (2004) Metabolic and oxidative responses associatedwith exposure of Eruca sativa (rocket) plants to different levels ofselenium. Int J Agric Biol 6:1101–1106
50. Hu Q, Pan G, Zhu J (2001) Effect of selenium on green teapreservation quality and amino acid composition of tea protein. JHortic Sci Biotechnol 76:344–346
51. Kochaki E (1997) Agronomy and plant breeding dry farming.Ferdosi University, Mashhad, Iran
52. Sujin RW (2004) Stress-inducible synthesis of proline in transgen-ic rice confers faster growth under stress conditions than that withconstitutive synthesis. Plant Sci 166:941–948
53. Ashraf MY (1998) Yield and yield components response of wheat(Triticum aestivum L.) genotypes grown under different soil waterdeficit conditions. Acta Agron Hung 46:45–51
Nawaz et al.
Related Documents