Journal of Plankton Research Vol.20 no.S pp.831-846, 1998 Selective feeding by protozoa: model and experimental behaviors and their consequences for population stability Suzanne L.Strom and Harilaos Loukos 1 Shannon Point Marine Center, Western Washington University, 1900 Shannon Point Road, Anacortes, WA 98221 and 'Joint Institute for the Study of the Atmosphere and Ocean (JISAO), Box 354235, University of Washington, Seattle, WA 98195, USA Abstract. Selective feeding by zooplankton can have profound consequences for the stability of grazer and prey populations, as demonstrated by the behavior of plankton dynamics models. We present an analytical approach—calculation of prey ratio trajectories—that reveals unambiguously whether selective feeding behavior is stabilizing (i.e. provides a refuge for preferred prey species at low prey concentrations) or destabilizing (i.e. results in elimination of prey populations). Prey ratio trajectories were calculated for three modeled selective feeding behaviors. Constant selection was consistently destabilizing, while selection behaviors that changed in response to either prey ratio or prey abundance could be stabilizing. Batch culture experiments with four protozoan grazer species (three ciliates, one heterotrophic dinofiagellate) demonstrated that protozoa fed selectively in every case, weakly preferring the larger of the two algal species offered. Stabilizing selection was observed only in the experiment with Favella sp., however, meaning that only this species altered its selection behavior in response to changing experimental conditions. Because protozoa are the major grazers of phytoplankton in many planktonic systems, our findings indicate that the use of selective feeding behaviors to stabilize plankton dynamics models needs careful evaluation. The modeling and graphical techniques presented here are a tool for linking further exploration of selective feeding behaviors with the development of plankton dynamics models. Introduction 'The value of information is generally finite, and partial reductions in ambiguity often may be good enough.' (Stephens and Krebs, 1986, p. 79) 'God is in the details.' (L. Mies van der Rohe) Selective feeding has long fascinated ecologists, from the details of individual feeding behavior to the implications for predator-prey dynamics and co- evolution. Selective feeding is denned as an imbalance between the proportion of prey types in a predator's diet and the proportion in the environment. The phenomenon is a compelling one because it suggests that predator feeding behavior may regulate not only the biomass, but also the composition of the prey community. In marine planktonic ecosystems, selective feeding by protozoa should be of particular importance given that these grazers are thought to consti- tute the major source of bacterial and-phytoplankton mortality (Pomeroy, 1974; Pace, 1988; Sherr and Sherr, 1993). Thus, selective feeding by planktonic proto- zoa could strongly influence the size and composition of marine bacterial and phytoplankton communities. The phenomenon of selective feeding is also important in the context of prey population stability. This can be seen in an examination of ecosystem dynamics models. Such models attempt to replicate and predict natural temporal patterns in generalized biomass compartments (e.g. primary producers, herbivores) and © Oxford University Press 831 by guest on July 17, 2011 plankt.oxfordjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Plankton Research Vol.20 no.S pp.831-846, 1998
Selective feeding by protozoa: model and experimental behaviorsand their consequences for population stability
Suzanne L.Strom and Harilaos Loukos1
Shannon Point Marine Center, Western Washington University, 1900 ShannonPoint Road, Anacortes, WA 98221 and 'Joint Institute for the Study of theAtmosphere and Ocean (JISAO), Box 354235, University of Washington, Seattle,WA 98195, USA
Abstract. Selective feeding by zooplankton can have profound consequences for the stability of grazerand prey populations, as demonstrated by the behavior of plankton dynamics models. We present ananalytical approach—calculation of prey ratio trajectories—that reveals unambiguously whetherselective feeding behavior is stabilizing (i.e. provides a refuge for preferred prey species at low preyconcentrations) or destabilizing (i.e. results in elimination of prey populations). Prey ratio trajectorieswere calculated for three modeled selective feeding behaviors. Constant selection was consistentlydestabilizing, while selection behaviors that changed in response to either prey ratio or preyabundance could be stabilizing. Batch culture experiments with four protozoan grazer species (threeciliates, one heterotrophic dinofiagellate) demonstrated that protozoa fed selectively in every case,weakly preferring the larger of the two algal species offered. Stabilizing selection was observed onlyin the experiment with Favella sp., however, meaning that only this species altered its selectionbehavior in response to changing experimental conditions. Because protozoa are the major grazersof phytoplankton in many planktonic systems, our findings indicate that the use of selective feedingbehaviors to stabilize plankton dynamics models needs careful evaluation. The modeling andgraphical techniques presented here are a tool for linking further exploration of selective feedingbehaviors with the development of plankton dynamics models.
Introduction'The value of information is generally finite, and partial reductions in ambiguity oftenmay be good enough.' (Stephens and Krebs, 1986, p. 79)
'God is in the details.' (L. Mies van der Rohe)
Selective feeding has long fascinated ecologists, from the details of individualfeeding behavior to the implications for predator-prey dynamics and co-evolution. Selective feeding is denned as an imbalance between the proportionof prey types in a predator's diet and the proportion in the environment. Thephenomenon is a compelling one because it suggests that predator feedingbehavior may regulate not only the biomass, but also the composition of the preycommunity. In marine planktonic ecosystems, selective feeding by protozoashould be of particular importance given that these grazers are thought to consti-tute the major source of bacterial and-phytoplankton mortality (Pomeroy, 1974;Pace, 1988; Sherr and Sherr, 1993). Thus, selective feeding by planktonic proto-zoa could strongly influence the size and composition of marine bacterial andphytoplankton communities.
The phenomenon of selective feeding is also important in the context of preypopulation stability. This can be seen in an examination of ecosystem dynamicsmodels. Such models attempt to replicate and predict natural temporal patternsin generalized biomass compartments (e.g. primary producers, herbivores) and
© Oxford University Press 831
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

S.LStrom and H.Loukos
ecosystem function (e.g. nutrient flux, material export). The basis for the modelsoften constitutes a balance between experimentally determined organismresponses and capabilities, on the one hand, and parameter adjustments neededto replicate field observations, on the other. A long-standing conundrum in suchmodels (e.g. Steele, 1974) is the mechanism by which predators avoid eliminat-ing their prey populations. Certain types of selective feeding can provide a solu-tion to the problem. For example, Fasham et al. (1990) chose a prey ratio-basedselection function (see below) to describe zooplankton grazing on bacteria,detritus and phytoplankton in their North Atlantic model. Because prey ratio-based selection relieves grazing pressure on the rarest prey types, the behaviorcreates a refuge for prey populations, and stabilizes both predator and preyabundances.
There is a good deal of evidence suggesting that planktonic protozoa arecapable of selective feeding (see reviews by Stoecker, 1988; Verity, 1991b). On amechanical level, some protozoans feed most efficiently (i.e. exhibit highest clear-ance rates) over a narrow prey size range, indicating that many potential preyparticles may be excluded from the diet simply by virtue of size (summarized byHansen et al., 1994). Other morphological attributes (e.g. spines) can similarlyrestrict availability. On a behavioral level, ciliates and flagellates clearly havechemosensory abilities, and can locate and remain near preferred prey types (e.g.Levandowsky and Kaneta, 1987; Sibbald etai, 1987; Verity, 1991a). Furthermore,high-speed videos show that ciliates can actively reject less preferred preyparticles even after they are captured (Taniguchi and Takeda, 1988; Stoecker etal, 1995). Studies comparing diet with the composition of available prey provideadditional evidence for selective feeding: protozoans have been shown variouslyto prefer live over dead cells, 'high-quality' over 'low-quality' foods, algae overglass beads, etc. (summarized in Verity, 1991b).
This paper presents a critical look at both model formulation of selectivefeeding as a stabilizing predation behavior, and at experimental data supportingthis hypothesis. By extending previous modeling efforts, we show that stabilizingbehavior can be obtained through abundance-based selection. A new method forthe analysis of data from simple batch culture experiments is described; thismethod reveals whether protozoan grazers exhibited stabilizing or destabilizingpredation behavior during experiments, although it does not unambiguouslyreveal the behavioral basis of that selection. We conclude that, on the one hand,the repertoire of grazer behaviors that tend to stabilize plankton dynamicsmodels should be extended to include abundance-based selection. On the otherhand, the limited available evidence suggests that stabilizing selection behavioramong protozoan grazers is uncommon, pointing to the need for further experi-mental and theoretical work on this important ecological phenomenon.
Selective feeding mechanisms: behavioral basisConstant selectionWith constant selection, grazer preference does not vary with changes in preyavailability; rather, grazing is continuously more effective on one prey type than
832
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

Protozoan selective feeding
on another. Selection is an invariant function of grazer feeding capability andprey morphological or physiological state (hence 'constant'); the terms 'mechan-ical selection' and 'passive selection' have also been used (e.g. Fenchel, 1986;Monger and Landry, 1991; Verity, 1991b). Differences in ability to feed couldarise at the level of prey capture, handling, ingestion, or some combination of allthree. The net result is that the availability of prey to the grazer is not the sameas it would appear from a count of prey abundance in the environment. Size andshape are probably the most obvious cause of differences in prey availability, andmany studies have reported size selectivity by protozoa (e.g. Andersson et ai,1986; Chrzanowski and Simek, 1990; Kivi and Setala, 1995). There may also bedifferences in the predator's ability to detect various prey types: some may besensed at a greater distance from the grazer, increasing the encounter ratebetween grazer and prey (Gerritsen and Strickler, 1977; Price and Paffenhofer,1983). Constant selection does not involve behavioral changes on the part of thegrazer, although longer term morphological or physiological adaptation could, ofcourse, alter the range of available prey. In the short term, however, constantselection does not vary with variations in the abundance or composition of theprey community.
Prey ratio-based selection
This and the abundance-based selection described below take as their premisethe idea that protozoa can feed selectively, in response to differences in prey size,nutrient or toxin content, or any of a host of other variables. They go on todescribe how selection might vary as prey availability changes—hence theirimportance for plankton dynamics models.
In prey ratio-based selection, selective feeding is dependent on the proportion(ratio) of prey types in the environment. The most commonly invoked form ofratio-based selection has been termed 'switching', defined by Murdoch (1969, p.336) as follows: 'Given that both prey (types) are eaten when presented alone toa predator, does the predator switch from the one prey when it becomes rare tothe other which is more abundant?' By providing a refuge from predation whena prey type becomes rare, this behavior can prevent the elimination of any preytype and can stabilize prey abundance (but see May, 1977). At equal prey abun-dances, a grazer may or may not exhibit a preference for one prey type overanother. The phenomenon is more robust when selection is strong (i.e. prefer-ence for the more abundant prey type is marked), and weak selection can, undercertain sets of conditions, have counter-intuitive destabilizing effects (Hutson,1984).
Ratio-based selection requires behavioral changes on the part of the grazer.Somehow, the grazer must sense the composition of the prey field and alterfeeding behavior in response; in consequence, this and the abundance-basedselection described below are sometimes termed 'active selection'. For protozoa,the underlying biological mechanisms are not clear, although it seems possiblethat information on prey composition obtained during digestion, for example,might feed back to other cellular components, triggering alterations in cell surface
833
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

S.I~Strom and H.Loukos
properties that affect encounter, capture or handling capabilities. In support ofthis, Tarran (1991) found that the heterotrophic dinoflagellate Oxyrrhis marinaapparently 'learned' to avoid latex beads when offered in a mixture with algae,showing no selectivity at the beginning of the experiment, but feeding only onalgal cells after several hours of exposure to the mixture.
Other data suggestive of prey ratio-based selection exist for protozoan grazers.Oxyrrhis marina has been shown to switch from a diet of Isochrysis galbana tocannibalism as I.galbana concentrations decreased (Flynn etal, 1996). Grazing apreferred alga to low concentration, followed by a switch to disproportionategrazing on a more abundant algal species, was found in studies of thechrysomonad flagellate Paraphysomonas imperforata (Goldman and Dennett,1990) and the dinoflagellate Gymnodinium sp. (Strom, 1991).
Abundance-based selectionThis mechanism postulates that selective feeding is a function of prey abundance.Abundance-based selection has been formalized in the basic prey model of forag-ing theory (summarized by Stephens and Krebs, 1986). The behavioral argumentpostulates that a grazer will feed preferentially on the highest energy-yieldingprey when it is abundant, because in this case ingestion of alternate prey willlower net energy assimilation by 'getting in the way'. When the highest energy-yielding prey is sparse, however, ingestion of alternate prey is predicted becausethis will increase net energy assimilation (Lehman, 1976; Stephens and Krebs,1986).
Under the most rigorous set of assumptions (i.e. encounter with individual preyitems is sequential and random; energetic costs of prey rejection are zero; search-ing and handling are mutually exclusive), the prey model predicts that a lesspreferred prey type (Q) should either always be ingested or always be ignoredwhen encountered ('all-or-nothing' selection). However, either individual vari-ance in the abundance of preferred prey (P) giving rise to ingestion of Q, or nutri-ent and toxin constraints (e.g. Q is an important source of a limiting nutrient, Pand/or Q contain a toxin that can only be tolerated in limited amounts) will leadto partial selection (Stephens and Krebs, 1986). Similarly, significant energeticcosts associated with rejection can lead to selection that is dependent on the abun-dance of both prey types, rather than just the abundance of preferred prey(Lehman, 1976; DeMott, 1990). Our model of abundance-based selection (below)includes these variants on 'all-or-nothing' selection.
As for prey ratio-based selection, abundance-based selection requires behav-ioral changes on the part of the grazer. There is evidence for abundance-basedselection in a recent study by Jiirgens and DeMott (1995), which examinedfeeding on a mix of prey types by the flagellates Bodo saltans and Spumella sp.Selection strength was directly proportional to the concentration of bacterialprey. Similarly, Heinbokel (1978; his Figure 10) found that the ciliate Eutintinnuspectinus fed preferentially on l.galbana when the concentration of that alga washigh, but exhibited no selectivity at low l.galbana concentrations.
834
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

Protozoan selective feeding
Method
Grazing experimentsExperimental procedures are described in detail in Strom and Morello (1998).Briefly, four species of protozoan grazers (three ciliates, one heterotrophic dino-flagellate) isolated from coastal Washington and Oregon (USA) waters weretested in simple growth assays to determine which paired combination of phyto-plankton species supported the highest protozoan growth rates. These phyto-plankton species were grown to late exponential phase in stock cultures; eachexperiment was initiated by adding the two phytoplankton species to sterilized0.2-um-filtered sea water in each of six or eight replicate 23 1 polycarbonatecarboys (Table I). Protozoan grazers were then added to half (three or four) ofthese carboys at initial concentrations of 0.1-19.7 cells ml"1. The initial volume incarboys was 11-23 1, while initial total algal concentration was 10 400-29 600 cellsml"1 (194-271 ug C I"1) (Table I). Carboys were then incubated at 13°C in dimlight (<1 umol photons nr2 s"1) with a 12:12 h light:dark cycle. After gentlemixing, samples were withdrawn at 12-24 h intervals for 6-10 days for determin-ation of algal abundance (by epifluorescence microscopy) and protozoan grazerabundance (by inverted microscopy). Algal biomass was estimated from algalabundances and cell volumes using the equations of Montagnes et al. (1994; seealso Strom and Morello, 1998).
Protozoan grazer abundance during each sampling interval was calculated froma logarithmic average (e.g. Heinbokel, 1978); clearance and ingestion rates werecalculated using the equations of Frost (1972). These equations account forchanges in control algal abundance in determining clearance and ingestion rates.
Selective feeding modelsThe approach used here stems from the work of Chesson (1978,1983), and hasbeen applied in various forms to plankton dynamics and food web models by Paceet al. (1984), Evans (1988), Fasham et al. (1990), Armstrong (1994), and possibly
Table I. Grazer and algal species, and initial concentrations (IC) for selective grazing experiments.Initial concentrations are in cells ml'1 for grazers and in pg C I'1 for algae
Exp.
1
2
3
4
Grazer
Favella sp.
StrombidinopsisacuminatumAmphidinium sp.
Uronema sp.
IC
0.1
0.4
14.9
19.7
AlgaP
Species
GymnodiniumsimplexProrocentrumminimumPyrenomonassalinaPyrenomonassalina
IC
154
49
156
204
AlgaQ
Species
PyrenomonassalinaPyrenomonassalinaIsochrysisgalbanaIsochrysisgalbana
IC
117
145
82
49
Alga P, preferred algal species; alga Q, less preferred algal species.
835
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

SJLStrom and H.Loukos
others. In particular, model development follows that of Fasham et al. (1990; theirAppendix A), although the arguments here are developed more extensively. Inthis approach, the probability of ingesting any particular prey type is proportionalto the combined probabilities of encountering and capturing that prey type. Thiscombined probability (Chesson's a, here p for preferred prey type P, q for lesspreferred prey type Q) can be used to model feeding success by multiplying it bythe prey abundance NP or NQ. In this case, 'One can think of the predator behav-ing as if there were pNP individuals of prey type P instead of NP and the proba-bility of ingestion is the probability of selecting, at random, a prey of type P fromthis distorted population' (Chesson, 1978, p. 212, our notation).
In the models described below, preference values p and q were considered todescribe the relationship between the proportion of a prey type in the environ-ment (NyJNtola\, where Ntotai = total Prey abundance) and the proportion in thediet (£>p/£\otai. where £>,oui = total prey in the diet) when NP = NQ, such that:
£Wotal andP
Thus, the total (p + q) sums to the total number of prey types in the system (here2) when NP = NQ, and the preference values can be used directly, as shown below,to scale grazing (e.g. in the no preference case, p = q = 1, and pNf = NP). Notethat p and q are prescribed values that are constants for a given model run, andcould be measured readily in a simple experimental system.
The specific grazing rate (G) was modeled using the Michaelis-Menten equa-tion:
where g is the maximum specific grazing rate, kg is the half-saturation constantfor grazing and A = pNP + qNQ = the total phytoplankton concentration asperceived by the grazers (see Table II for a summary of model notation). Grazingon a particular prey type (e.g. Q) is then scaled to the perceived abundance of Qby substitution of preference-scaled prey concentrations into equation (2):
Table II. Model variables and initial conditions
Variable
Maximum specific grazing rateGrazing half-saturation constantGross growth efficiencyGrazing saturation level (= 2 &g)Constant selection coefficientsPrey ratio-based selection coefficientsAbundance-based selection coefficientsInitial conditionsConcentration of preferred algal speciesConcentration of less preferred algal speciesGrazer concentration
Symbol
8kJi•"salP. 9P*,<7*
N?NQH
Value
4100
0.25200
250250
2
Units
day1
ugCI-1
ugCl-1
ugCI-1
MgCMugCI-'
836
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

.g
Protozoan selective feeding
(3)~ u kg + (pNP + qNQyThe simplest case, constant selection, is represented by equation (3). In the case
of prey ratio-based selection, preferences are denoted by p* and q*, whichdepend on prey proportion in the environment:
p* =pNP
pNP + qNQand pNP + qNQ
(4)
Substitutingp* and q* for p and q in equation (3), grazing on prey type Q is thendescribed by:
GQ = g (5)
Abundance-based selection was modeled in two different ways, correspondingto the predictions of foraging theory under different energy- or nutrient-limitation scenarios (see above). First, grazing on the less preferred species (Q)was modeled as 'all or nothing' (e.g. full selection or no selection), with the tran-sitional prey abundance a function of NP. The rigorous (and biologically unreal-istic) 'all-or-nothing' assumption was relaxed by assuming some degree ofvariance around the transitional prey abundance; below, we illustrate selectionpatterns corresponding to high and low levels of variance. This variance repre-sents partial selection, and could correspond to differing perceptions of the tran-sitional prey abundance, among individuals in a grazer population. Second,selection was modeled as directly proportional to prey abundance. In this case,selection becomes progressively weaker as prey abundance decreases, rather thanalternating between 'all' and 'nothing' states. Throughout, /?** and q** are used
O.Oq-100 200 300 400
Abundance of P (^gC liter1)500
Fig. L Logistic curves used to model variance around 'all-or-nothing' transitional prey abundance(see the text for a description of the abundance-based selection model). Slope values of 30 and 6 usedto represent transition point variances of ±10% i4sat and ±50% ^sat, respectively.
837
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

S.L*Stroin and H.Loukos
to denote abundance-based preferences, and were substituted for p and q [equa-tion (3)] in the various model runs.
Grazer selectivity in the case of 'all or nothing' abundance-based selection wassimulated using a logistic curve with asymptotic minimum = 0 and asymptoticmaximum = q (Figure 1):
(6)
where b = slope of logistic curve and the transitional prey abundance = Aax = thelowest prey concentration at which grazing is saturated. This formulation assumesthat selection will occur only when prey concentrations are high enough to satur-ate grazing, an assumption that could easily be varied. Slope (b) values of 6 and30 were used to simulate grazer populations in which 90% of the individuals tran-sitioned between 'all' and 'nothing' behavior within ±50% Asat (a 'messy' popu-lation) and ±10% Aat (a 'tidy' population), respectively (Figure 1).
Selection proportional to the entire range of prey abundance was simulated intwo ways.1. For selection proportional to preferred prey abundance Q?** and q** <* NP):
lfNP<Asat,p**=l-^-(l-p) and q** = l-^L(l-q) (7)•"sat -"sat
, If NP>Asat,p**=p,q** = q
2. For selection proportional to the abundance of each individual prey type {p**oc Nj>, q** « NQ), P** is defined as in equation (7), while q** is defined analo-gously:
lfNQ<Asat,q** = l-^(l-q) (8)•"sat
IiNQ>:Asat,q** = q
Models were run as a series of 0.1 day time steps. Ingestion (/, ug C'h1) wascalculated separately for P and Q during each time interval At according to:
(9)
(and analogously for Q), where the grazer concentration (H) was given by:
// (0 = £(/„,_„ + /Q ( , - D) + / /( , . i) (10)For model runs, values of the maximum specific grazing rate g (4 day1) and grossgrowth efficiency E (0.25) were based on average values from these grazingexperiments (Strom and Morello, 1998; see Table II). The half-saturationconstant kg (100 ug C I"1) was estimated based on literature values (Heinbokel,1978; Verity, 1985). Note that the only effect of changing g or E in this model isto change the time required for the grazer population to remove all phyto-plankton. Since model output is normalized to time (see below), values of g and
838
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

Protozoan selective feeding
E ultimately have no effect on prey ratio trajectories and assessments of stability.Model runs were initialized with conditions (phytoplankton and grazer abun-dances) resembling those at the start of actual grazing experiments (Tables I andII), and allowed to run until all phytoplankton were consumed. For simplicity,phytoplankton growth during model runs was 0 (as in grazing experiments); theeffect of varying phytoplankton growth rate could be investigated readily withthis model formulation.
ResultsSelective feeding was observed during all four of the grazing experiments, as indi-cated by continuous decreases in the relative abundance of the preferred prey
over time (Figure 2). Changes in A/pWQ in control carboys were
4 6Time (d)
Fig. 2. Prey composition, grazer and prey abundance during grazing experiments. Upper panels:Np:NQ = preferred prey abundance:less preferred prey abundance (ug C I"'). Lower panels: • , grazerabundance; A, Pyrenomonos salina abundance; O, abundance of Gymnodinium simplex (A), Proro-centrum minimum (B) or Isochrysis galbana (C, D). An asterisk indicates the preferred algal speciesin each experiment.
839
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

S.L-Strom and H.Loukos
Table III. Dimensions of algal species used in experiments. Measurements are averages of 40observations; all cells preserved in 5% acid Lugol's. Length (L) and width (W) in um, ER is theeffective radius = (L + W)I2. p = logC/yFjJ/logCf/fig/f/?,™) (Monger and Landry, 1991), where Fisthe clearance rate. F\JFsm was calculated as the average of clearance rate ratios from all carboys andall time intervals except the first two of each experiment. As the larger alga was always preferred,ERflERQ = ERJER
Gymnodinium simplexPyrenomonas salinaProrocentrum minimum'Prorocentrum minimum*1
Pyrenomonas salinaPyrenomonas salinaIsochrysis galbanaPyrenomonas salinaIsochrysis galbana
11.28.0
11.116.19.09.44.09.23.8
6.74.7
10.014.64.95.03.75.03.6
9.06.4
na15.47.07.23.97.13.7
Exp. Alga L W ER ERp/ER0 0
1 Gymnodinium simplex 11.2 6.7 9.0 1.4 1.5
2.2 1.5
1.8 0.4
1.9 1.03.8 3.6 3.7
••Dimensions of cell contents.^Dimensions of thecae.
non-directional and slight by comparison (data not shown). Protozoan grazersshowed a consistent preference for the larger of the two prey species offered.Note that Pyrenomonas salina was offered in every experiment; it was thepreferred prey, however, only during experiments 3 and 4 when it was the largerof the two algal species (Table III). Furthermore, the larger species was preferredregardless of whether it was initially more or less abundant than the smallerspecies (Table I).
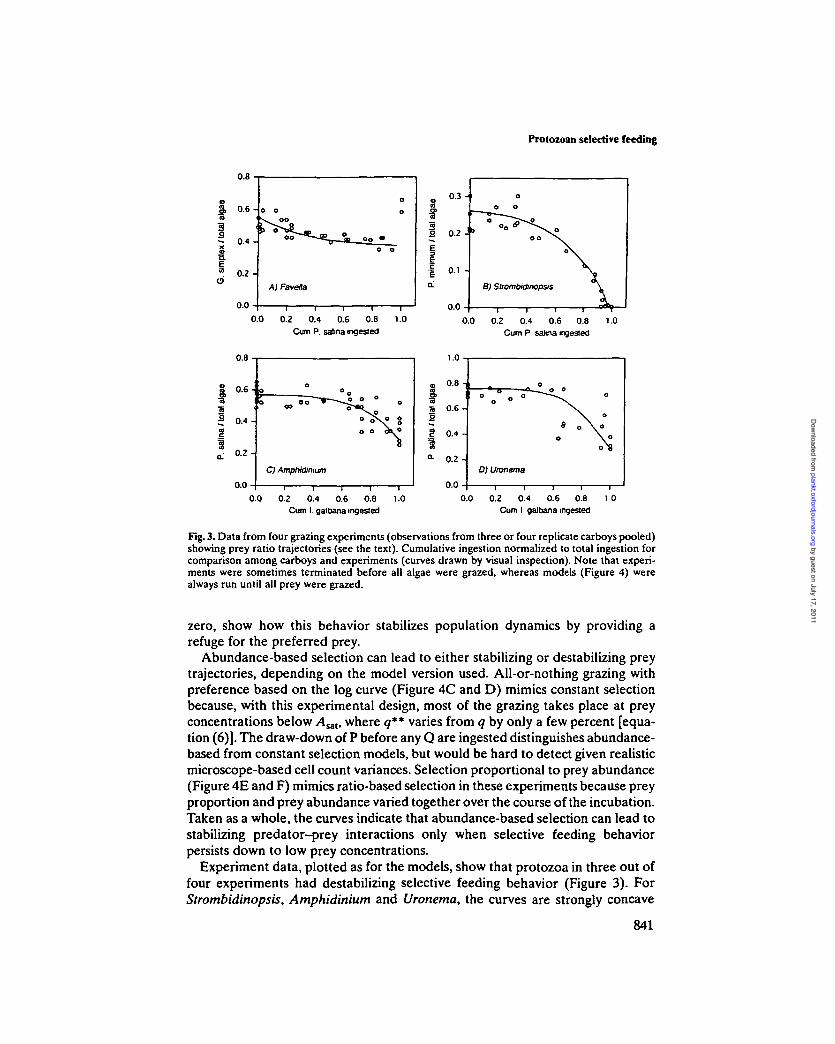
No major differences among the four grazing experiments were seen whendata were viewed in a conventional manner (i.e. as changes in prey abundanceor prey ratio over time; Figure 2). We explored several alternative methods ofevaluating changes in prey ratio [Np/(NP + NQ)] over the course of each experi-ment. It seemed likely that the trajectory of the change could be related to bothpreference strength and the selection mechanism. As the independent variable(Figure 3), we used the cumulative amount of algae ingested, rather than time,to normalize among experiments with different time courses. Cumulative algaeingested was expressed as a fraction of total algae ingested to normalize amongreplicate carboys within an experiment. This approach (Figure 3) shows how theprey ratio changes as increasing amounts of algae are ingested by grazers. Plot-ting against cumulative Q ingested (as opposed to cumulative P, or cumulativeP + Q) provided the best separation between the three grazing models, as wellas among the varying degrees of preference (e.g. magnitudes of p and q) for agiven model.
The three selective feeding models are distinguished by fundamental differ-ences in the behavior of the prey ratio when plotted as a function of cumulativeQ ingested. The strongly concave-downward trajectories of the constant selectionmodel (Figure 4A), with values of the prey ratio going to zero (i.e. all preferredprey removed by grazers), show that, as modeled here, this feeding mechanism isfundamentally destabilizing. In contrast, the concave-upward trajectories of theratio-based model (Figure 4B), in which the prey ratio always remains above
840
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from
Protozoan selective feeding
0.6-
0.4 -
0.2 -
0.0-
o o\ ^ oo
A) Favetla
oo
_ <p On »
o o
0.0 0.2 0.4 0.6 0.8Cum P. salina ingested
1.0
0.2 0.4 0.6 0.8 1.0Cum I. galbana ingested
fa2
0.3 -
0.2 -
0.2 0.4 0.6 0.8 1.0Cum P salina ingested
a§
2%Sia.
0.8 -
0.6-
0.4 -
0.2 -
0.0-
oo o ^ ^ ^ ^
a ° ^ \\
" oo
0) UronemaV " 1 "" 1 I "
o
o\ o
\ °o o
,—0.0 0.2 0.4 0.6 0.8
Cum I galbana ingested1 0
Fig. 3. Data from four grazing experiments (observations from three or four replicate carboys pooled)showing prey ratio trajectories (see the text). Cumulative ingestion normalized to total ingestion forcomparison among carboys and experiments (curves drawn by visual inspection). Note that experi-ments were sometimes terminated before all algae were grazed, whereas models (Figure 4) werealways run until all prey were grazed.
zero, show how this behavior stabilizes population dynamics by providing arefuge for the preferred prey.
Abundance-based selection can lead to either stabilizing or destabilizing preytrajectories, depending on the model version used. All-or-nothing grazing withpreference based on the log curve (Figure 4C and D) mimics constant selectionbecause, with this experimental design, most of the grazing takes place at preyconcentrations below A^, where q** varies from q by only a few percent [equa-tion (6)]. The draw-down of P before any Q are ingested distinguishes abundance-based from constant selection models, but would be hard to detect given realisticmicroscope-based cell count variances. Selection proportional to prey abundance(Figure 4E and F) mimics ratio-based selection in these experiments because preyproportion and prey abundance varied together over the course of the incubation.Taken as a whole, the curves indicate that abundance-based selection can lead tostabilizing predator-prey interactions only when selective feeding behaviorpersists down to low prey concentrations.
Experiment data, plotted as for the models, show that protozoa in three out offour experiments had destabilizing selective feeding behavior (Figure 3). ForStrombidinopsis, Amphidinium and Uronema, the curves are strongly concave
841
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

SJLStrom and H.Loukos
oo 02 04 os oeOmQngnM
00 0.2 04 OS OSCumOmgraad
Fig. 4. Prey ratio trajectories as predicted by different selective feeding models. Trajectories repre-sent prey ratios [Np/(Np + NQ)] as a function of the cumulative ingestion of the less preferred preyspecies (Q). Cumulative ingestion normalized to total ingestion for comparison with experimentresults. Lines show trajectories for values of q (preference for algal species Q) ranging from 0.1 to 1.0(see the text). (A) Constant selection. (B) Prey ratio-based selection. (C + D) 'All-or-nothing' abun-dance-based selection with logistic curve slope values equal to (C) 30 and (D) 6. (E + F) Abundance-based selection with (E) selection proportional to abundance of the preferred prey type P; (F)selection for P proportional to abundance of P, selection for less preferred prey type Q proportionalto abundance of Q.
downward. Not all experiments were run until prey were completely removed, sonot all curves go to prey ratios of zero. All three do, however, decrease to lowprey ratios relative to initial ratios. Further, the shapes of the curves indicate thatselection against the less-preferred prey type was weak (i.e. by comparison withmodel curves, values of q were only slightly <1).
The remaining experiment, with the tintinnid Favella sp. as grazer, resulted in afundamentally different type of selection behavior. The weakly concave-upward
842
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

Protozoan selective feeding
curve and the slight decrease in prey ratio (Figure 3A) demonstrate a stabilizingselection mechanism. Further, Favella showed a complete reversal in preferenceduring the last time interval of the experiment when the abundance of thepreferred Gymnodinium simplex became very low (average 15 ug C I"1). A tran-sition to preferential grazing on Rsalina during this last time interval resulted in asharp increase in the prey ratio. Removal of preferred prey to this level or lowerin other experiments did not result in similar preference reversals.
DiscussionAll protozoan grazers tested showed selective feeding, with consistent preferencefor the larger of the two prey species offered. Though common, preference forlarger prey is by no means universal (Hansen et al, 1994), and may simply indi-cate that prey characteristics likely to override size (e.g. ease of capture, nutrientor toxin content) were eliminated by the use of optimized diets in our experi-ments.
More importantly, selection against smaller prey was weak in all four experi-ments. That is, prey ratio trajectories most closely resembled model curves withq values close to one. This finding agrees with the prediction of the Force-Balancemodel (Monger and Landry, 1990,1991), which predicts that clearance rates (F)should be roughly proportional to the 0.7-1.0 power of prey radius (/?). Thecoefficient p, which indicates the strength of clearance rate size dependence, isgiven by:
(£) &Y <»>(Monger and Landry, 1991). Values of P calculated for our experiments rangedfrom 0.4 to 1.5 (Table III), substantiating the idea that size-based preferences areroughly proportional to prey radius and lead to weak selection.
Selective feeding was destabilizing in three out of the four experiments, asshown unequivocally by prey ratio trajectories (Figure 3). This means that threeof the protozoan species, while capable of feeding selectively, did not alter theirselection behavior in response to changing conditions in the batch cultures.Alteration of selection behavior is required to produce stabilizing prey trajec-tories, i.e. to create a behavioral refuge for the preferred prey when its concen-tration is low. It is important to note that weak selection can still be destabilizing,as is clear from both actual and modeled prey ratio trajectories (Figures 3and 4).
Uniquely among these protozoa, the tintinnid Favella sp. did exhibit stabiliz-ing selection behavior. Selective feeding behavior by this ciliate must havechanged during the course of the incubation, in response to changing prey ratios,prey abundances, or some combination of both. Behavioral shifts by Favella arealso indicated by the complete reversal in algal preference during the last timeinterval of this experiment, a phenomenon not seen when other grazers experi-enced similar variations in prey availability.
Two high-speed video studies of Favella (Taniguchi and Takeda, 1988; Stoecker
843
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

S.LStrom and H.Loukos
et al., 1995) indicate that this ciliate can actively reject particles after capture.Stoecker et al. (1995) also showed that Favella usually ingests larger particles athigher rates, due in part to enhanced capture efficiency in the outer and middleciliary zones. In other words, Favella is better able to capture large particles withthe entire ciliary apparatus, and has the ability to reject less desirable particlesafter capture. It is also suggestive that rejection behavior was more frequent whenoverall food densities were higher, in agreement with an abundance-based selec-tion mechanism. Thus, one possible explanation for Favella's unique selectionbehavior is that this ciliate has an unusual ability to modify its feeding behavior.An alternative hypothesis is that phenotypic changes over the incubation periodcaused changes in feeding behavior in this, but not the other, protozoan speciesstudied.
The modeling technique presented here shows that abundance-based selectivefeeding can be stabilizing under certain conditions. Specifically, grazer behaviormust change in response to prey abundance even at very low prey concentrations.The 'all-or-nothing' abundance response predicted under the most basic assump-tions of foraging theory is not stabilizing because the abundance response 'turnsoff' at relatively high food levels. On the other hand, selection behavior thatresponds to the entire range of prey abundance provides a behavioral refuge forthe preferred prey that is essential for stability. We propose that abundance-basedselection is an alternative hypothesis for the real-world temporal stability of manyplanktonic ecosystems, and should be explored further in ecosystem models andgrazing experiments.
Stabilizing selective feeding behavior was not common among the protozoathat we studied. This calls into question the legitimacy of using such behaviors asstabilizing mechanisms in plankton dynamics models. Because the nature ofherbivore-phytoplankton interaction is so critical to model stability, we urgefurther exploration of both the behavioral repertoire of protozoan herbivores andthe range of approaches that can provide model stability. An additional unre-solved question is whether stabilizing selection behaviors can be used in modelsas proxies for whole community responses (e.g. Fasham et al., 1990). In otherwords, to what extent can the grazer species assemblage—the 'meta-popu-lation'—alter in response to changes in prey availability, and can such alterationsgive rise to stabilizing selection behaviors that are not achievable by singlespecies? Our modeling and graphical technique is a tool for closer articulation ofmodel and experiment development. Use of this theoretical framework to designand optimize experiments, and translation of results directly into ecosystem-levelplankton dynamics models, should speed progress in this research area.
AcknowledgementsThe Uronema sp. culture was graciously provided by B. and E.Sherr. We thankT.A.Morello, W.Arthurs, J.Holmes and K.Thompson for assistance with culturemaintenance, cell counts and grazing experiments. Helpful comments wereprovided by G.Wolfe, B.Frost, the SPMC Plankton Group and two anonymousreviewers. This research was supported by NSF grant OCE 9301698 (to S.S.). H.L.
844
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

Protozoan selective feeding
was supported by a JISAO fellowship under a cooperative agreement withNOAA (NA67RJ0155). This publication is JISAO contribution no. 478.
ReferencesAndersson,A., Larsson.U. and Hagstrom,A. (1986) Size-selective grazing by a microflagellate on
pelagic bacteria. Mar. EcoL Prog. Ser., 33, 51-57.Armstrong,R. A. (1994) Grazing limitation and nutrient limitation in marine ecosystems: Steady state
solutions of an ecosystem model with multiple food chains. Limnol. Oceanogr., 39,597-608.ChessonJ. (1978) Measuring preference in selective predation. Ecology, 59, 211-215.ChessonJ. (1983) The estimation and analysis of preference and its relationship to foraging models.
Ecology, 64,1297-1304.Chrzanowski.T.H. and Simek.K. (1990) Prey-size selection by freshwater flagellated protozoa.
Limnol. Oceanogr., 35,1429-1436.DeMott.W.R. (1990) Retention efficiency, perceptual bias, and active choice as mechanisms of food
selection by suspension-feeding zooplankton. In Hughes.R.N. (ed.), Behavioural Mechanisms ofFood Selection. Springer-Verlag, Berlin, pp. 569-594.
Evans.G.T. (1988) A framework for discussing seasonal succession and coexistence of phytoplanktonspecies. Limnol Oceanogr., 33,1027-1036.
FashamAU.R., Ducklow.H.W. and McKelvie.S.M. (1990) A nitrogen-based model of planktondynamics in the oceanic mixed layer. / Mar. Res., 48,591-639.
Fenchel,T. (1986) Protozoan filter feeding. Prog. Protistoi, 1,65-113.Flynn.KJ., Davidson.K. and Cunningham.A. (1996) Prey selection and rejection by a microflagellate;
implications for the study and operation of microbial food webs. J. Exp. Mar. Biol. EcoL, 196,357-372.
Frost,B.W. (1972) Effects of size and concentration of food particles on the feeding behavior of themarine planktonic copepod Calanus pacificus. Limnol. Oceanogr., 17, 805-815.
Gerritsen j . and StricklerJ.R. (1977) Encounter probabilities and community structure in zooplank-ton: a mathematical model. J. Fish. Res. Board Can., 34,73-82.
Goldman J.C. and Dennett.M.R. (1990) Dynamics of prey selection by an omnivorous flagellate. Mar.Ecol. Prog. Ser., 59,183-194.
Hansen.B., Bjornsen,P.K. and Hansen,PJ. (1994) The size ratio between planktonic predators andtheir prey. Limnol. Oceanogr, 39,395-403.
Heinbokel J.F. (1978) Studies on the functional role of tintinnids in the Southern California Bight. I.Grazing and growth rates in laboratory cultures. Mar. Biol., 47, 177-189.
Hutson.V. (1984) Predator mediated coexistence with a switching predator. Math. Biosci, 68,233-246.
JUrgens.K. and DeMott.W.R. (1995) Behavioral flexibility in prey selection by bacterivorous nano-flagellates. Limnol. Oceanogr., 40,1503-1507.
Kivi.K. and Setala.O. (1995) Simultaneous measurement of food particle selection and clearance ratesof planktonic oligotrich ciliates (Ciliophora: Oligotrichina). Mar. Ecol. Prog. Ser., 119, 125-137.
Lehman J.T. (1976) The filter-feeder as an optimal forager, and the predicted shapes of feedingcurves. Limnol. Oceanogr., 21,501-516.
Levandowsky,M. and Kaneta,P.J. (1987) Behavior in dinoflagellates. In TaylorJFJ.R. (ed.). TheBiology of Dinoflagellates. Blackwell Scientific, Oxford, pp. 360-397.
May.R.M. (1977) Predators that switch. Nature, 269,103-104.Monger.B.C. and Landry.M.R. (1990) Direct-interception feeding by marine zooflagellates: the
importance of surface and hydrodynamic forces. Mar. EcoL Prog. Ser, 65, 123-140.Monger,B.C. and Landry,M.R. (1991) Prey-size dependency of grazing by free-living marine flagel-
lates. Mar. EcoL Prog. Ser., 74,239-248.MontagnesJ5J.S., BergesJ., HarrisonJ'J. and TaylorJ.R. (1994) Estimating carbon, nitrogen,
protein, and chlorophyll a from volume in marine phytoplankton. LimnoL Oceanogr, 39,1044-1060.
Murdoch,W.W. (1969) Switching in general predators: experiments on predator specificity andstability of prey populations. EcoL Monogr., 39, 335-354.
Pace,M.L. (1988) Bacterial mortality and the fate of bacterial production. Hydrobiologia, 159,41-49.Pace,M.L., GlasserJ.E. and Pomeroy,L.R. (1984) A simulation analysis of continental shelf food
webs. Mar. BioL, 82,47-63.Pomeroy,L.R- (1974) The ocean's food web, a changing paradigm. Bioscience, 24, 499-504.
845
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from

S.L.Srrom and ILLoukos
Price.HJ. and Paffenhofer,G.-A. (1983) Modes of cell capture in calanoid copepods. LimnoLOceanogr., 28,116-123.
SherT.E.B. and Sherr.B.F. (1993) Bacterivory and herbivory: key roles of phagotrophic protists inpelagic food webs. Microb. EcoL, 28,223-235.
Sibbald.MJ., Albright.LJ. and Sibbald.P.R. (1987) Chemosensory responses of a heterotrophicmicroftagellate to bacteria and several nitrogen compounds. Mar. EcoL Prog. Ser., 36,201-204.
SteeleJ.H. (1974) The Structure of Marine Ecosystems. Harvard University Press, Cambridge.Stephens.D.W. and KrebsJ.R. (1986) Foraging Theory. Princeton University Press, Princeton.Stoecker,D.K. (1988) Are marine planktonic ciliates suspension-feeders?/ ProtozooL, 35,252-255.Stoecker,D.K., Gallager,S.M., Langdon.CJ. and DavisJLH. (1995) Particle capture by Favella sp.
(Ciliata, Tintinnina). J. Plankton Res., 17,1105-1124.Strom.S.L. (1991) Growth and grazing rates of the herbivorous dinoflagellate Cymnodinium sp. from
the open subarctic Pacific Ocean. Mar. EcoL Prog. Ser., 78,103-113.Strom.S.L. and Morello.T.A. (1998) Comparative growth rates and yields of ciliates and heterotrophic
dinoflagellates. / Plankton Res., in press.Taniguchi.A. and Takeda.Y. (1988) Feeding rate and behavior of the tintinnid ciliate Favella
taraikaensis, observed with a high speed VTR system. Mar. Microb. Food Webs, 3,21-34.Tarran.G.A. (1991) Aspects of the feeding behaviour of the marine dinoflagellate Oxyrrhis marina
Dujardin. PhD Thesis, Southampton University, Southampton.VerityJ".G. (1985) Grazing, respiration, excretion, and growth rates of tintinnids. LimnoL Oceanogr,
30,1268-1282.Verity.P.G. (1991a) Aggregation patterns of ciliates from natural assemblages in response to different
prey. Mar. Microb. Food Webs, 5,115-128.Verity,P.G. (1991b) Feeding in planktonic protozoans: evidence for non-random acquisition of prey.
/ ProtozooL, 38, 69-76.
Received on December 4,1996; accepted on December 9, 1997
846
by guest on July 17, 2011plankt.oxfordjournals.org
Dow
nloaded from
Related Documents











