Selective and divided attention modulates auditory–vocal integration in the processing of pitch feedback errors Ying Liu, 1, * Huijing Hu, 1,2, * Jeffery A. Jones, 3 Zhiqiang Guo, 4 Weifeng Li, 1 Xi Chen, 1 Peng Liu 1 and Hanjun Liu 1,4 1 Department of Rehabilitation Medicine, The First Affiliated Hospital, Sun Yat-sen University, Guangzhou 510080, China 2 Guangdong Provincial Work Injury Rehabilitation Center, Guangzhou, China 3 Psychology Department and Laurier Centre for Cognitive Neuroscience, Wilfrid Laurier University, Waterloo, ON, Canada 4 Department of Biomedical Engineering, School of Engineering, Sun Yat-sen University, Guangzhou, China Keywords: auditory feedback, auditory–motor integration, divided attention, selective attention, speech motor control Abstract Speakers rapidly adjust their ongoing vocal productions to compensate for errors they hear in their auditory feedback. It is cur- rently unclear what role attention plays in these vocal compensations. This event-related potential (ERP) study examined the influence of selective and divided attention on the vocal and cortical responses to pitch errors heard in auditory feedback regard- ing ongoing vocalisations. During the production of a sustained vowel, participants briefly heard their vocal pitch shifted up two semitones while they actively attended to auditory or visual events (selective attention), or both auditory and visual events (divided attention), or were not told to attend to either modality (control condition). The behavioral results showed that attending to the pitch perturbations elicited larger vocal compensations than attending to the visual stimuli. Moreover, ERPs were likewise sensitive to the attentional manipulations: P2 responses to pitch perturbations were larger when participants attended to the audi- tory stimuli compared to when they attended to the visual stimuli, and compared to when they were not explicitly told to attend to either the visual or auditory stimuli. By contrast, dividing attention between the auditory and visual modalities caused suppressed P2 responses relative to all the other conditions and caused enhanced N1 responses relative to the control condition. These find- ings provide strong evidence for the influence of attention on the mechanisms underlying the auditory–vocal integration in the pro- cessing of pitch feedback errors. In addition, selective attention and divided attention appear to modulate the neurobehavioral processing of pitch feedback errors in different ways. Introduction Auditory feedback plays an important role in the acquisition and maintenance of speech motor skills (Houde & Jordan, 1998; Hickok et al., 2011). By exposing speakers to altered auditory feedback (AAF) regarding their ongoing speech production, researchers have demonstrated that speakers adjust their vocal output to compensate for feedback perturbations in fundamental frequency (F 0 ), vocal intensity and formant frequency (Jones & Munhall, 2005; Bauer et al., 2006; Liu et al., 2010; Cai et al., 2011; Scheerer et al., 2013). Researchers have also shown that activity in the auditory cor- tex is suppressed during active vocalisation relative to passive listen- ing when the auditory feedback matches the actual vocal output (Houde et al., 2002; Heinks-Maldonado et al., 2005; Flinker et al., 2010; Behroozmand & Larson, 2011; Wang et al., 2014). By con- trast, when there is a mismatch between auditory feedback and vocal output, vocalisation elicits larger neural responses than during pas- sive listening (Eliades & Wang, 2008; Behroozmand et al., 2009; Chang et al., 2013; Chen et al., 2013). This enhanced cortical activity is thought to reflect a mechanism that serves to compensate for errors perceived during vocalisation (Chang et al., 2013). Although recent research has greatly advanced our understanding of the integration of sensory and motor information during speech, we know little about the role, if any, that attention plays. Outside of the laboratory environment, speakers are typically faced with the task of simultaneously processing auditory feedback in conjunction with other sensory information (e.g. visual, somatosensory, etc.). If auditory feedback processing requires some degree of attention, and given that attention is considered a limited resource (N€ a€ at € anen, 1992), then the processing of perceived motor errors may be modu- lated by the attentional load incurred by processing multimodal stimuli. The cognitive and neural processes that underlie attention have been the subject of intense study for many years. For example, stud- ies of unimodal selective attention have demonstrated enhanced brain activity in the auditory cortex in response to attended vs. unat- tended stimuli (Woldorff et al., 1993; Pugh et al., 1996; Ahveninen et al., 2011). Studies of bimodal (e.g. auditory and visual) selective attention have likewise shown more brain activity in the auditory Correspondence: Hanjun Liu, 1 Department of Rehabilitation Medicine, as above. E-mail: [email protected] * These two authors contributed equally to this study. Received 28 November 2014, revised 28 April 2015, accepted 11 May 2015 © 2015 Federation of European Neuroscience Societies and John Wiley & Sons Ltd European Journal of Neuroscience, pp. 1–10, 2015 doi:10.1111/ejn.12949

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Selective and divided attention modulates auditory–vocalintegration in the processing of pitch feedback errors
Ying Liu,1,* Huijing Hu,1,2,* Jeffery A. Jones,3 Zhiqiang Guo,4 Weifeng Li,1 Xi Chen,1 Peng Liu1 and Hanjun Liu1,41Department of Rehabilitation Medicine, The First Affiliated Hospital, Sun Yat-sen University, Guangzhou 510080, China2Guangdong Provincial Work Injury Rehabilitation Center, Guangzhou, China3Psychology Department and Laurier Centre for Cognitive Neuroscience, Wilfrid Laurier University, Waterloo, ON, Canada4Department of Biomedical Engineering, School of Engineering, Sun Yat-sen University, Guangzhou, China
Keywords: auditory feedback, auditory–motor integration, divided attention, selective attention, speech motor control
Abstract
Speakers rapidly adjust their ongoing vocal productions to compensate for errors they hear in their auditory feedback. It is cur-rently unclear what role attention plays in these vocal compensations. This event-related potential (ERP) study examined theinfluence of selective and divided attention on the vocal and cortical responses to pitch errors heard in auditory feedback regard-ing ongoing vocalisations. During the production of a sustained vowel, participants briefly heard their vocal pitch shifted up twosemitones while they actively attended to auditory or visual events (selective attention), or both auditory and visual events(divided attention), or were not told to attend to either modality (control condition). The behavioral results showed that attendingto the pitch perturbations elicited larger vocal compensations than attending to the visual stimuli. Moreover, ERPs were likewisesensitive to the attentional manipulations: P2 responses to pitch perturbations were larger when participants attended to the audi-tory stimuli compared to when they attended to the visual stimuli, and compared to when they were not explicitly told to attend toeither the visual or auditory stimuli. By contrast, dividing attention between the auditory and visual modalities caused suppressedP2 responses relative to all the other conditions and caused enhanced N1 responses relative to the control condition. These find-ings provide strong evidence for the influence of attention on the mechanisms underlying the auditory–vocal integration in the pro-cessing of pitch feedback errors. In addition, selective attention and divided attention appear to modulate the neurobehavioralprocessing of pitch feedback errors in different ways.
Introduction
Auditory feedback plays an important role in the acquisition andmaintenance of speech motor skills (Houde & Jordan, 1998; Hickoket al., 2011). By exposing speakers to altered auditory feedback(AAF) regarding their ongoing speech production, researchers havedemonstrated that speakers adjust their vocal output to compensatefor feedback perturbations in fundamental frequency (F0), vocalintensity and formant frequency (Jones & Munhall, 2005; Baueret al., 2006; Liu et al., 2010; Cai et al., 2011; Scheerer et al.,2013). Researchers have also shown that activity in the auditory cor-tex is suppressed during active vocalisation relative to passive listen-ing when the auditory feedback matches the actual vocal output(Houde et al., 2002; Heinks-Maldonado et al., 2005; Flinker et al.,2010; Behroozmand & Larson, 2011; Wang et al., 2014). By con-trast, when there is a mismatch between auditory feedback and vocaloutput, vocalisation elicits larger neural responses than during pas-
sive listening (Eliades & Wang, 2008; Behroozmand et al., 2009;Chang et al., 2013; Chen et al., 2013). This enhanced corticalactivity is thought to reflect a mechanism that serves to compensatefor errors perceived during vocalisation (Chang et al., 2013).Although recent research has greatly advanced our understanding
of the integration of sensory and motor information during speech,we know little about the role, if any, that attention plays. Outside ofthe laboratory environment, speakers are typically faced with thetask of simultaneously processing auditory feedback in conjunctionwith other sensory information (e.g. visual, somatosensory, etc.). Ifauditory feedback processing requires some degree of attention, andgiven that attention is considered a limited resource (N€a€at€anen,1992), then the processing of perceived motor errors may be modu-lated by the attentional load incurred by processing multimodalstimuli.The cognitive and neural processes that underlie attention have
been the subject of intense study for many years. For example, stud-ies of unimodal selective attention have demonstrated enhancedbrain activity in the auditory cortex in response to attended vs. unat-tended stimuli (Woldorff et al., 1993; Pugh et al., 1996; Ahveninenet al., 2011). Studies of bimodal (e.g. auditory and visual) selectiveattention have likewise shown more brain activity in the auditory
Correspondence: Hanjun Liu, 1Department of Rehabilitation Medicine, as above.E-mail: [email protected]
*These two authors contributed equally to this study.
Received 28 November 2014, revised 28 April 2015, accepted 11 May 2015
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons Ltd
European Journal of Neuroscience, pp. 1–10, 2015 doi:10.1111/ejn.12949

cortex when participants attend to auditory stimuli while ignor-ing visual stimuli, and more activity in the visual cortex whenparticipants attend to visual stimuli while ignoring auditory stimuli(Woodruff et al., 1996; Johnson & Zatorre, 2005, 2006). In additionto this enhancement in the attended modality, cross-modal inhibitionhas been demonstrated in the form of decreases in activity observedin the ‘ignored’ sensory cortex (Shomstein & Yantis, 2004; Johnson& Zatorre, 2005, 2006).Although attention is typically manifest as a selective focus on
the processing of one aspect of the environment, attention can alsobe divided when we must perform two (or more) tasks simulta-neously. Not surprisingly, the complexity of dividing attentionacross stimuli, or between modalities, involves activity in additionalregions of the brain. For example, in a study of divided visual atten-tion, discrimination between shape, color and speed of a visual stim-ulus activated the anterior cingulate and the prefrontal cortex (PFC)in the right hemisphere (Corbetta et al., 1991). In addition, whenattention has to be divided between auditory and visual stimuli,researchers have found significant decreases in the level of activityin both the auditory and visual cortices compared to when attentionis focused on either modality alone (Klingberg, 1998; Loose et al.,2003; Johnson & Zatorre, 2006). Event-related potential (ERP) stud-ies have similarly shown that divided attention causes processingdelays, which are reflected as increased latencies of the P300 com-ponent (Hohnsbein et al., 1991).The research on cross-modal attention thus indicates that attention
modulates activity in the sensory cortices. As speech motor controlrelies on sensory feedback from audition, somatosensation and kin-esthesis, whether or not the sensorimotor integration that is criticalto speech motor control can be shaped by attention is an importantquestion. Accurate estimation of the dynamic state of the vocal artic-ulators relies on the detection of errors in voice auditory feedback.In addition, considerable evidence has demonstrated that the centralauditory processing of speech sounds is highly dependent on atten-tion. For example, larger ERPs (Hink & Hillyard, 1976; Stevenset al., 2006) or enhanced brain activity (Ahveninen et al., 2006;Sabri et al., 2008) in the auditory cortex is observed in response toattended vs. unattended speech sounds. Moreover, disruption of thelip representation induced by transcranial magnetic stimulationresults in increased left-hemisphere P50m response to attendedspeech sounds (Mottonen et al., 2014), suggesting that the earlierinteractions between auditory and motor cortices depend on atten-tion. Taken together, these studies suggest that attentional mecha-nisms may influence the auditory–motor processing of vocal pitcherrors.Evidence that attention modulates auditory–motor integration
comes from two recent studies. In an ERP study performed by Tum-ber et al. (2014), participants were exposed to pitch perturbationsduring vocalisation while they either passively viewed a rapid serialvisual presentation (RSVP) of letters, or actively identified targetstimuli in the RSVP of letters. The results showed that activelyattending to the RSVP elicited a decrease in the magnitude of vocalcompensation for pitch perturbations relative to passively viewingthe RSVP, whereas the P1-N1-P2 complex elicited by pitch pertur-bations was not modulated by attention. This finding providesbehavioral evidence for the influence of attention on the auditory–motor integration in voice control.In another study conducted by Hu et al. (2015), participants were
exposed to pitch perturbations during vocalisation while they eitherattended to a visual stimulus or attended to the pitch perturbationsin a low (counting the number of perturbations) or high (countingthe type of perturbations) attentional load condition. The results
revealed no systematic change in the vocal compensations for thepitch perturbations, irrespective of whether the perturbations wereattended or not. However, larger P2 responses were observed whenparticipants attended to the pitch perturbations in the low-load atten-tional condition relative to P2 responses observed in the high-loadattentional condition or those observed when the pitch perturbationswere unattended. The authors concluded that attentional load modu-lates auditory cortical responses to perceived vocal pitch errors.Although the above-cited studies provide support for the hypothe-
sis that there is a role for attention in auditory–motor integrationduring speech production, certain shortcomings in these two studieslimit our understanding. For example, Tumber et al. (2014) investi-gated whether taxing central attentional processes affects auditory–motor integration by manipulating the visual attention load but didnot manipulate the attention their participants paid to the pitch per-turbations. Thus, the question of whether dividing attention betweenvisual and auditory stimuli affects the auditory–motor processing ofpitch perturbations remains unanswered. Hu et al. (2015) found themodulatory effect of attention on the cortical processing of vocalpitch feedback errors as a function of load level, but ERP responsesfor these attentional manipulations were not compared to a controlcondition where participants would passively listen to the pitch per-turbations and view the visual stimuli. Thus, Hu et al. (2015) wereunable to determine whether the observed differences in the corticalresponses between attentional conditions reflected enhancement inthe attended modality, inhibition in the unattended modality or both.Therefore, whether and how attention influences the auditory–motorintegration in voice control remains poorly understood.In the present study, we directly compared the influence of selec-
tive attention and divided attention on auditory–motor integrationduring voice control. Participants sustained a vowel phonation whilethey heard their voice pitch-shifted (i.e., auditory stimuli) and simul-taneously saw flashing lights (i.e., visual stimuli). The participants’task was to attend to either the auditory or to the visual stimuli(selective attention), to attend to both modalities (divided attention),or to passively observe both the auditory and visual stimuli (controlcondition). Vocal and cortical (N1-P2 complex) responses to thepitch perturbations in these experimental conditions were measuredand compared. We hypothesised that larger vocal and corticalresponses would occur when participants attended to the pitch per-turbations as compared to when participants were asked to ignorethem. Moreover, we predicted that divided attention would elicitsmaller vocal and ERP responses to the pitch perturbations than inthe selective attention condition.
Materials and methods
Al Subjects
Thirty-three students from Sun Yat-sen University of China partici-pated in the experiment. Three participants were excluded from thefinal sample because of technical problems (N = 2) and poor dataquality (N = 1). Therefore, data from 30 subjects (seven males and23 females) were analysed. All participants were right-handed,native-Mandarin speakers with a mean age of 23 �3 (SD) years. Noparticipant reported a history of speech, hearing, language or neuro-logical disorders. All subjects passed a hearing screening at 25 dBhearing level (HL) for octave intervals of 500–4000 Hz and hadnormal or corrected-to-normal vision. Informed consent wasobtained from all participants. All the procedures, including subjectrecruitment and data acquisition, were approved by the InstitutionalReview Board of The First Affiliated Hospital at Sun Yat-sen Uni-
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–10
2 Y. Liu et al.

versity of China, and were in accordance with the Code of Ethics ofthe World Medical Association (Declaration of Helsinki).
Apparatus
Participants were seated in a sound-attenuated booth throughout theexperiment. Prior to data recording, the experimental system wascalibrated to ensure that the intensity of voice feedback that partici-pants heard was 10 dB (sound pressure level, SPL) higher than theiractual voice output to partially mask the airborne and bone-con-ducted feedback (Behroozmand et al., 2009). The voice signals weretransduced by a dynamic microphone (model DM2200; Takstar Inc.,Huizhou, China) and amplified by a MOTU Ultralite Mk3 FireWireaudio interface (Cambridge, MA, USA). The amplified signals werethen pitch-shifted by an Eventide Eclipse Harmonizer (Little Ferry,NJ, USA), which was controlled by a custom program created withMax/MSP (v.6.0 by Cycling 74, San Francisco, CA, USA). Thisprogram also generated transistor–transistor logic (TTL) pulses thatmarked the onset and offset of the pitch perturbations. The TTLpulses were also sent to the electroencephalograph (EEG) recordingsystem via a FireWire cable. Finally, the pitch-shifted voices wereamplified by an ICON NeoAmp (Middleton, WI, USA) headphoneamplifier and fed back to participants through insert earphones(ER1-14A, Etymotic Research Inc., Elk Grove Village, IL, USA).The original voice, the pitch-shifted feedback and the TTL pulseswere digitised at 10 kHz by a PowerLab A/D converter (modelML880, AD Instruments, Castle Hill, Australia) and recorded usingLabChart software (v.7.0, AD Instruments).
Visual and auditory Stimuli
Visual stimuli
Two circles representing the blue and red indicator lights were gen-erated by Max/MSP software and displayed on the computer screen.The red indicator light began to flash 500 ms after the blue indicatorlight prompted participants to vocalise. Participants were instructedto vocalise when the blue indicator light was turned on and termi-nate their vocalisations when the blue indicator light was turned off,during which they heard their voice auditory feedback unexpectedlyshifted upwards (see Auditory stimuli). During each vocalisation, thered indicator light flashed 1–7 times with variable inter-stimulusintervals (ISIs) ranging from 400 to 1600 ms (400, 600, 800, 1000,1200, 1400 and 1600 ms). The duration of the red light was fixed at200 ms.
Auditory stimuli
The pitch-shift stimulus (PSS) that participants heard was +200cents (100 cents equals 1 semitone) with a fixed duration of200 ms. The number of PSS presented to subjects ranged from oneto four per vocalisation. The first PSS was presented 500–1000 msafter the vocal onset, and the succeeding stimuli occurred with anISI of 700–900 ms. The onsets of visual and auditory stimuli wereasynchronous.
Procedure
The present study included four conditions: (i) attend to the PSS butignore the red indicator light (Auditory Attention); (ii) attend to thered indicator light but ignore the PSS (Visual Attention); (iii) attendto both PSS and red indicator light (Bimodal Attention); and (iv)
passively listen to the PSS and view the lights (Bimodal Passive).Across all conditions, subjects vocalised the vowel sound /u/ for ~5sat a comfortable pitch when cued by the blue indicator light, andsaw the red indicator light flashing. At the end of each vocalisation,subjects were required to take a break of 2–3s prior to initiating thenext vocalisation. Production of ~40 consecutive vocalisationsconstituted one block, which resulted in ~100 trials (i.e. PSS) percondition. As a control condition, the Bimodal Passive conditionwas always the first block, while participants were unaware of theattention-related tasks that were to follow. The order of the otherthree conditions was counterbalanced across all subjects.Across all but the Bimodal Passive condition, an immediate recall
test was performed after each vocalisation to ensure participantsattended to or ignored the stimuli as instructed. Subjects reportedthe number of PSS that they heard in the Auditory Attention condi-tion, the number of red indicator light flashes that they saw in theVisual Attention condition, or both in the Bimodal Attention condi-tion. The percentage of correctly remembered stimuli across thethree conditions was calculated based on these data.
EEG data acquisition and analyses
Participants wore a 64-electrode Geodesic Sensor Net (ElectricalGeodesics Inc., Eugene, OR, USA) on their scalp, and EEG signalswere amplified by a Net Amps 300 amplifier (Electrical GeodesicsInc.) and recorded using NetStation software (v.4.5; Electrical Geo-desics Inc.). EEG signals from all channels were referenced to thevertex (Cz) during the recording and digitised at a sampling fre-quency of 1 kHz. The impedances of individual sensors were main-tained below 50 kΩ throughout the recording (Ferree et al., 2001).After data acquisition, NetStation software was used for off-line
analyses of the EEG signals. Data from all channels were band-pass-filtered at 1–20 Hz and then segmented into epochs rangingfrom 200 ms before to 500 ms after the onset of the PSS. Seg-mented trials contaminated by excessive muscular activity, eyeblinks or eye movements were assessed using the Artifact Detec-tion toolbox in NetStation and eliminated from further analyses.Additional visual inspection on individual trials was performed toensure that artifacts were rejected appropriately. Individual elec-trodes containing artifacts in > 20% of the segments would beexcluded from further analyses. All channels were then re-refer-enced to the average of electrodes on each mastoid, and artifact-free epochs were averaged and baseline-corrected to generate anoverall response across each condition. The amplitude and latencyof both the N1 and P2 components across conditions were mea-sured as the negative and positive peaks in the time windows of80–180 and 160–280 ms after the onset of PSS, and submitted tostatistical analyses.
Vocal data analysis
Voice F0 contours were extracted for each vowel production usingPraat (Boersma, 2001). F0 values in Hz were converted to the centsscale using the formula cents = 100 9 [12 9 log2(F0/reference)],where the reference denotes the arbitrary reference note 195.997 Hz(G4). Each trial was segmented into epochs ranging from 200 msbefore to 700 ms after the onset of the PSS. Each trial was visuallyinspected to ensure that trials with vocal interruptions or signal pro-cessing errors were excluded from further analyses. Artifact-free tri-als for each of the four conditions were then averaged for eachparticipant to generate an overall vocal response. Vocal responselatencies in ms were measured as the time when the F0 trajectory
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–10
Attention effects on voice feedback processing 3

exceeded 2 SDs above or below the pre-stimulus mean followingthe perturbation onset. Vocal response magnitudes in cents were cal-culated by subtracting the pre-stimulus mean from the peak value ofthe voice F0 contour following the response onset. The peak time ofvocal response in ms were determined at the point of greatest devia-tion from the value at the onset of the stimulus.
Statistical analyses
The vocal and neurophysiological response data were subjected torepeated-measures analyses of variance (RM-ANOVAs) in SPSS(v.16.0). Specifically, the magnitudes, latencies and peak times ofvocal responses were analysed using one-way RM-ANOVAs with atten-tion condition as the single factor (i.e. Auditory Attention, VisualAttention, Bimodal Attention and Bimodal Passive). The amplitudeand latency of the N1-P2 complex, extracted from 10 electrodes(FC1, FC2, FCz, FC3, FC4, C1, C2, Cz, C3, C4), were analysedusing three-way RM-ANOVAs (attention condition, anteriority and later-ality). Frontal (FC1, FC2, FCz, FC3, FC4) and central (C1, C2, Cz,C3, C4) electrodes were chosen as an anteriority factor, while lateralleft (FC3, C3), medial left (FC1, C1), midline (FCz, Cz), medial right(FC2, C2) and lateral right (FC4, C4) were used as a laterality factor.Responses from frontal and central electrodes were chosen for statis-tical analyses because neurophysiological responses to PSS aremostly pronounced in these two areas (Hawco et al., 2009; Chenet al., 2012). Subsidiary RM-ANOVAs were calculated if higher-orderinteractive effects reached significance. Probability values wereadjusted using Greenhouse–Geisser correction when the assumptionof sphericity was violated.
Results
Behavioral performance
Participants’ accuracy at counting and recalling the number of PSSin the Auditory Attention condition, the number of red light flashesin the Visual Attention condition, and both of these auditory andvisual stimuli in the Bimodal Attention condition, was calculated asa percentage of correct responses for each of these conditions. Par-ticipants’ accuracy at reporting the number of PSS in the BimodalAttention condition (79 � 2%) (mean � SEM throughout unlessotherwise indicated) was significantly lower than that in the Audi-tory Attention condition (98 � 1%; t29 = 8.470, P < 0.001). Simi-
larly, participants’ accuracy at reporting the number of red lightflashes in the Bimodal Attention condition (80 � 3%) was signifi-cantly lower than that in the Visual Attention condition (95 � 1%;t29 = 6.293, P < 0.001).
Vocal responses
Figure 1 shows the grand-averaged voice F0 contours and the T-barplots of the absolute magnitudes of vocal response to the PSS acrossthe four attention conditions. A one-way RM-ANOVA of the responsemagnitude revealed a significant main effect of attention condition(F3,87 = 2.863, P = 0.041). Post hoc Bonferroni comparison testsrevealed that the Auditory Attention condition (12.5 � 0.8 cents)elicited significantly larger vocal responses than the Visual Attentioncondition (10.3 � 0.8 cents; P = 0.026). Vocal responses during theBimodal Attention condition (12.2 � 1.1 cents) were larger thanduring the Visual Attention condition (11.4 � 0.9 cents) but thisdifference failed to reach significance (P = 0.203). Vocal responsemagnitudes for the Bimodal Passive condition were not significantlydifferent from the responses elicited in the other three conditions(P > 0.5). In addition, vocal response latencies did not differ as afunction of the attention condition (F3,87 = 0.486, P = 0.693; Audi-tory Attention, 86 � 7 ms; Visual Attention, 86 � 9 ms; BimodalAttention, 86 � 7 ms; Bimodal Passive, 91 � 8 ms). In addition,no significant main effect of attention condition was found for thepeak times of vocal responses (F3,87 = 0.697, P = 0.556; AuditoryAttention, 352 � 34 ms; Visual Attention, 348 � 37 ms; BimodalAttention, 324 � 23 ms; Bimodal Passive, 326 � 25 ms).
Neurophysiological responses
Figure 2 shows the grand-averaged ERP waveforms in response tothe PSS for the Auditory Attention (red lines), Visual Attention(blue lines), Bimodal Attention (black lines) and Bimodal Passiveconditions (green lines). Figure 3 shows the topographical distribu-tions of the N1 and P2 components across the four attention condi-tions. As can be seen in Figs 2 and 3, the Auditory Attentioncondition elicited the largest P2 responses and the Bimodal Atten-tion condition elicited the smallest P2 responses, while P2responses were similar in the Visual Attention and Bimodal Pas-sive conditions. In addition, N1 response also appeared to beaffected by attention, as reflected by the larger responses (i.e. morenegative) in the Bimodal Attention condition than in the BimodalPassive condition.
A B
Fig. 1. (A) Grand-averaged voice F0 contours and (B) T-bar plots of the magnitudes of vocal responses to pitch perturbations across the four attention condi-tions. The thick solid line, the sparse dashed line, the dense dashed line and the thin solid line represent the vocal responses in the Auditory Attention, VisualAttention, Bimodal Attention and Bimodal Passive conditions, respectively. *P < 0.05 (P = 0.026) for the size of the vocal responses between the AuditoryAttention and Visual Attention conditions. Error bars represent SEM.
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–10
4 Y. Liu et al.
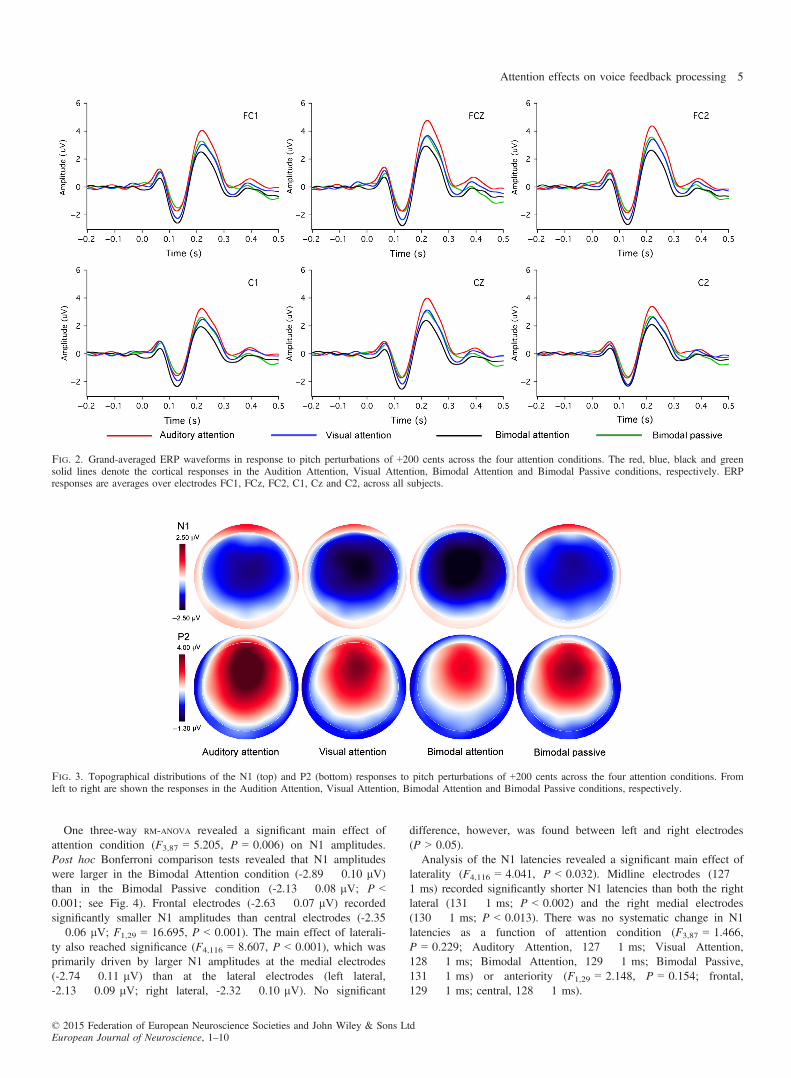
One three-way RM-ANOVA revealed a significant main effect ofattention condition (F3,87 = 5.205, P = 0.006) on N1 amplitudes.Post hoc Bonferroni comparison tests revealed that N1 amplitudeswere larger in the Bimodal Attention condition (-2.89 � 0.10 lV)than in the Bimodal Passive condition (-2.13 � 0.08 lV; P <0.001; see Fig. 4). Frontal electrodes (-2.63 � 0.07 lV) recordedsignificantly smaller N1 amplitudes than central electrodes (-2.35� 0.06 lV; F1,29 = 16.695, P < 0.001). The main effect of laterali-ty also reached significance (F4,116 = 8.607, P < 0.001), which wasprimarily driven by larger N1 amplitudes at the medial electrodes(-2.74 � 0.11 lV) than at the lateral electrodes (left lateral,-2.13 � 0.09 lV; right lateral, -2.32 � 0.10 lV). No significant
difference, however, was found between left and right electrodes(P > 0.05).Analysis of the N1 latencies revealed a significant main effect of
laterality (F4,116 = 4.041, P < 0.032). Midline electrodes (127 �1 ms) recorded significantly shorter N1 latencies than both the rightlateral (131 � 1 ms; P < 0.002) and the right medial electrodes(130 � 1 ms; P < 0.013). There was no systematic change in N1latencies as a function of attention condition (F3,87 = 1.466,P = 0.229; Auditory Attention, 127 � 1 ms; Visual Attention,128 � 1 ms; Bimodal Attention, 129 � 1 ms; Bimodal Passive,131 � 1 ms) or anteriority (F1,29 = 2.148, P = 0.154; frontal,129 � 1 ms; central, 128 � 1 ms).
Fig. 2. Grand-averaged ERP waveforms in response to pitch perturbations of +200 cents across the four attention conditions. The red, blue, black and greensolid lines denote the cortical responses in the Audition Attention, Visual Attention, Bimodal Attention and Bimodal Passive conditions, respectively. ERPresponses are averages over electrodes FC1, FCz, FC2, C1, Cz and C2, across all subjects.
Fig. 3. Topographical distributions of the N1 (top) and P2 (bottom) responses to pitch perturbations of +200 cents across the four attention conditions. Fromleft to right are shown the responses in the Audition Attention, Visual Attention, Bimodal Attention and Bimodal Passive conditions, respectively.
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–10
Attention effects on voice feedback processing 5
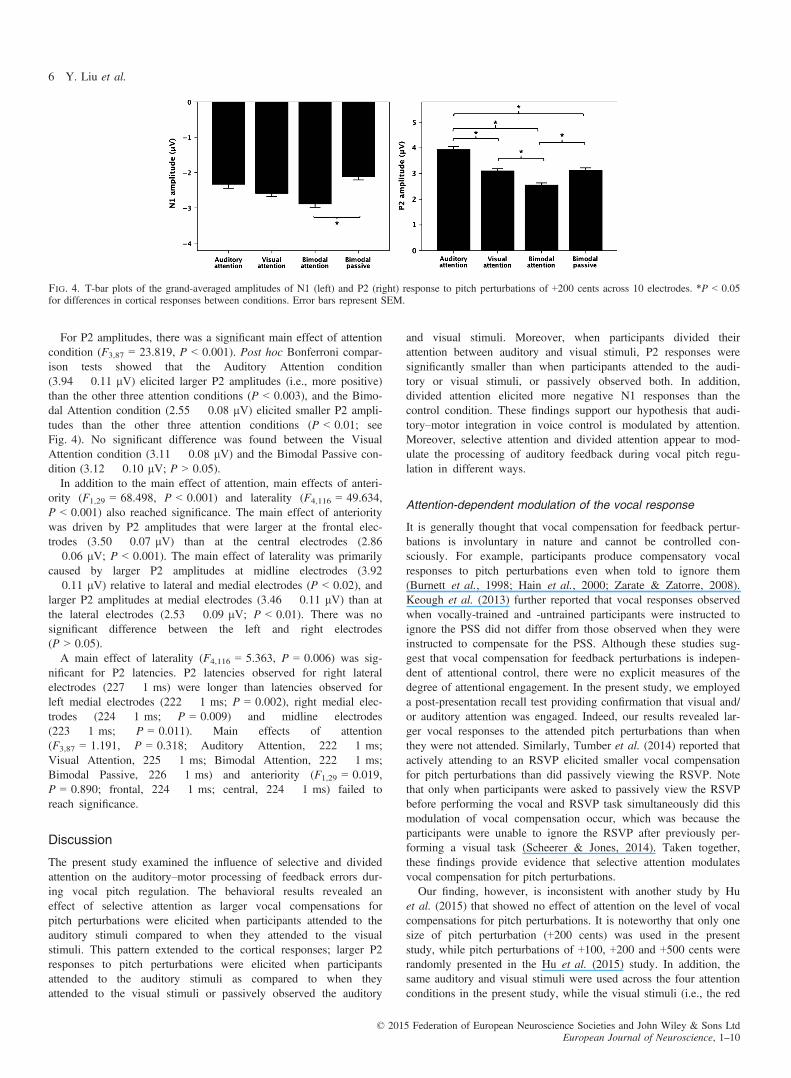
For P2 amplitudes, there was a significant main effect of attentioncondition (F3,87 = 23.819, P < 0.001). Post hoc Bonferroni compar-ison tests showed that the Auditory Attention condition(3.94 � 0.11 lV) elicited larger P2 amplitudes (i.e., more positive)than the other three attention conditions (P < 0.003), and the Bimo-dal Attention condition (2.55 � 0.08 lV) elicited smaller P2 ampli-tudes than the other three attention conditions (P < 0.01; seeFig. 4). No significant difference was found between the VisualAttention condition (3.11 � 0.08 lV) and the Bimodal Passive con-dition (3.12 � 0.10 lV; P > 0.05).In addition to the main effect of attention, main effects of anteri-
ority (F1,29 = 68.498, P < 0.001) and laterality (F4,116 = 49.634,P < 0.001) also reached significance. The main effect of anterioritywas driven by P2 amplitudes that were larger at the frontal elec-trodes (3.50 � 0.07 lV) than at the central electrodes (2.86� 0.06 lV; P < 0.001). The main effect of laterality was primarilycaused by larger P2 amplitudes at midline electrodes (3.92� 0.11 lV) relative to lateral and medial electrodes (P < 0.02), andlarger P2 amplitudes at medial electrodes (3.46 � 0.11 lV) than atthe lateral electrodes (2.53 � 0.09 lV; P < 0.01). There was nosignificant difference between the left and right electrodes(P > 0.05).A main effect of laterality (F4,116 = 5.363, P = 0.006) was sig-
nificant for P2 latencies. P2 latencies observed for right lateralelectrodes (227 � 1 ms) were longer than latencies observed forleft medial electrodes (222 � 1 ms; P = 0.002), right medial elec-trodes (224 � 1 ms; P = 0.009) and midline electrodes(223 � 1 ms; P = 0.011). Main effects of attention(F3,87 = 1.191, P = 0.318; Auditory Attention, 222 � 1 ms;Visual Attention, 225 � 1 ms; Bimodal Attention, 222 � 1 ms;Bimodal Passive, 226 � 1 ms) and anteriority (F1,29 = 0.019,P = 0.890; frontal, 224 � 1 ms; central, 224 � 1 ms) failed toreach significance.
Discussion
The present study examined the influence of selective and dividedattention on the auditory–motor processing of feedback errors dur-ing vocal pitch regulation. The behavioral results revealed aneffect of selective attention as larger vocal compensations forpitch perturbations were elicited when participants attended to theauditory stimuli compared to when they attended to the visualstimuli. This pattern extended to the cortical responses; larger P2responses to pitch perturbations were elicited when participantsattended to the auditory stimuli as compared to when theyattended to the visual stimuli or passively observed the auditory
and visual stimuli. Moreover, when participants divided theirattention between auditory and visual stimuli, P2 responses weresignificantly smaller than when participants attended to the audi-tory or visual stimuli, or passively observed both. In addition,divided attention elicited more negative N1 responses than thecontrol condition. These findings support our hypothesis that audi-tory–motor integration in voice control is modulated by attention.Moreover, selective attention and divided attention appear to mod-ulate the processing of auditory feedback during vocal pitch regu-lation in different ways.
Attention-dependent modulation of the vocal response
It is generally thought that vocal compensation for feedback pertur-bations is involuntary in nature and cannot be controlled con-sciously. For example, participants produce compensatory vocalresponses to pitch perturbations even when told to ignore them(Burnett et al., 1998; Hain et al., 2000; Zarate & Zatorre, 2008).Keough et al. (2013) further reported that vocal responses observedwhen vocally-trained and -untrained participants were instructed toignore the PSS did not differ from those observed when they wereinstructed to compensate for the PSS. Although these studies sug-gest that vocal compensation for feedback perturbations is indepen-dent of attentional control, there were no explicit measures of thedegree of attentional engagement. In the present study, we employeda post-presentation recall test providing confirmation that visual and/or auditory attention was engaged. Indeed, our results revealed lar-ger vocal responses to the attended pitch perturbations than whenthey were not attended. Similarly, Tumber et al. (2014) reported thatactively attending to an RSVP elicited smaller vocal compensationfor pitch perturbations than did passively viewing the RSVP. Notethat only when participants were asked to passively view the RSVPbefore performing the vocal and RSVP task simultaneously did thismodulation of vocal compensation occur, which was because theparticipants were unable to ignore the RSVP after previously per-forming a visual task (Scheerer & Jones, 2014). Taken together,these findings provide evidence that selective attention modulatesvocal compensation for pitch perturbations.Our finding, however, is inconsistent with another study by Hu
et al. (2015) that showed no effect of attention on the level of vocalcompensations for pitch perturbations. It is noteworthy that only onesize of pitch perturbation (+200 cents) was used in the presentstudy, while pitch perturbations of +100, +200 and +500 cents wererandomly presented in the Hu et al. (2015) study. In addition, thesame auditory and visual stimuli were used across the four attentionconditions in the present study, while the visual stimuli (i.e., the red
Fig. 4. T-bar plots of the grand-averaged amplitudes of N1 (left) and P2 (right) response to pitch perturbations of +200 cents across 10 electrodes. *P < 0.05for differences in cortical responses between conditions. Error bars represent SEM.
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–10
6 Y. Liu et al.

indicator light) were not presented when participants were asked toattend to the pitch perturbations in the Hu et al. (2015) study.Therefore, the inconsistency in the effect of selective attention onthe vocal pitch monitoring between these two studies may be attrib-uted to the differences in the presentation of the auditory and visualstimuli across the attention conditions.As expected, participants performed more poorly during the
Bimodal Attention condition than the Auditory Attention conditionin terms of their recall for the number of PSS they heard duringeach utterance. This finding is in line with other cross-modal studiesthat have shown behavioral deficits when participants divided theirattention between modalities as compared to selectively attending toone modality and ignoring the other (Bonnel & Hafter, 1998; Jolico-eur, 1999). One might therefore hypothesise that the Bimodal Atten-tion condition should elicit smaller vocal compensatory responses topitch perturbations than the Auditory Attention condition. However,the vocal responses did not differ across these two conditions, whichsuggests that divided attention does not affect the vocal responses.Nonetheless, selectively attending to or ignoring the auditory modal-ity did affect the vocal responses. Moreover, the vocal responses didnot differ across the Visual Attention, Bimodal Attention and Bimo-dal Passive conditions. In addition, vocal responses were similar inthe Auditory Attention and Bimodal Passive conditions. Together,this pattern of results suggests that the attention effect on vocalcompensation for feedback perturbations may be dependent on thedegree of attentional engagement. In other words, only when pitchperturbations were fully attended vs. completely ignored did atten-tion have an impact on the vocal compensation.
Selective attention effect on the neurophysiological responses
The present study found larger P2 responses were elicited when par-ticipants attended to the pitch perturbations (i.e., Auditory Attentioncondition) than when they ignored the pitch perturbations (i.e.,Visual Attention condition) or were not asked to attend to the audi-tory stimuli or the visual stimuli (i.e., Bimodal Passive condition),indicating that selective attention modulates cortical processing ofauditory feedback during vocal pitch regulation. This aspect of ourfinding is consistent with one recent study that showed larger P2responses to attended vs. unattended pitch perturbations in voiceauditory feedback (Hu et al., 2015). Similarly, previous research onauditory perception showed enhanced brain activity in the auditorycortex (Grady et al., 1997; Petkov et al., 2004), larger P2 responses(Picton & Hillyard, 1974; Woldorff & Hillyard, 1991; Neelon et al.,2006) and mismatch negativity (N€a€at€anen et al., 1993; Alain &Woods, 1997; Woldorff et al., 1998; Sussman et al., 2007) elicitedby attended vs. unattended sounds. Our results are also consistentwith the effects of selective attention on bimodal sensory processing(e.g., auditory and visual) reported in other neuroimaging studiesthat showed more activity in the auditory cortex while participantsattended to an auditory stimulus than when the same stimulus wasignored (Kawashima et al., 1999; Loose et al., 2003; Johnson &Zatorre, 2005, 2006).There are theories that account for the enhanced P2 responses to
attended vs. unattended pitch perturbations. One theory is the gain-based theory of selective attention (Hillyard et al., 1998) that positsthat paying attention to one modality while ignoring anotherincreases the gain of neurons that are sensitive to the attended modal-ity, which results in decreased perceptual thresholds for that modal-ity. In the context of this theory then, attending to the pitchperturbations while ignoring the flashing lights in the Auditory Atten-tion condition may lead to an increased gain for neurons involved in
auditory–motor integration that facilitates the detection/correction ofpitch errors in voice auditory feedback, resulting in enhanced P2responses to attended vs. unattended pitch perturbations.An alternative explanation for the enhanced P2 responses we
observed may be that cortical responses to the unattended pitchperturbations decreased. There is evidence that suggests thatmodality-specific selective attention results primarily fromdecreased processing in the unattended modality rather thanincreased processing in the attended modality (Mozolic et al.,2008). Thus, the selective attention effect observed in the presentstudy may have resulted from inhibited activity in the sensorycortices that subserve the processing of pitch perturbations, therebyfreeing central attentional resources for the processing of flashinglights in the Visual Attention condition.There is also considerable evidence that both cross-modal facilita-
tion and inhibition underlie attentional modulation of the sensorycortices, such that attention to one modality leads not only toincreased activity in the sensory cortical area involved in the pro-cessing of that modality but also to suppressed activity in regionsassociated with other modalities (Loose et al., 2003; Johnson & Za-torre, 2005, 2006; Mozolic et al., 2008). This leaves open the possi-bility that changes in cortical responses may result from not onlyenhanced response to the attended pitch perturbations but alsodecreased responses to the unattended pitch perturbations.As the above interpretations involve the comparison of the audi-
tory and visual selective attention conditions with each other, ratherthan to the control condition, it is unclear whether changes in thecortical responses reflect only enhancement in the attended modality,inhibition in the unattended modality, or both. However, our resultsrevealed that cortical responses to pitch perturbations in the VisualAttention condition did not differ from those observed when partici-pants passively observed the bimodal stimuli (i.e., Bimodal Passivecondition). Similarly, Tumber et al. (2014) reported that the P1-N1-P2 complex to pitch perturbations did not change when participantsactively attended to the RSVP as compared to when they passivelyviewed the RSVP. In other words, increased attention to the visualstimuli, relative to the control condition, did not appear to suppressthe cortical processing of auditory stimuli during the online controlof vocal production. This finding does not support the hypothesisthat the processing of attended information leads to the inhibition ofprocessing of unattended information. Thus, the observed effect ofselective attention may be primarily due to increased gain in neuronsthat are sensitive to the detection and/or correction of feedback per-turbation during vocal production.
Divided attention effect on the neurophysiological response
The present study found smaller P2 responses in the Bimodal Atten-tion condition than in the Auditory Attention condition. This findingsuggests that divided attention leads to decreased auditory corticalprocessing of vocal pitch perturbations relative to selective attention.These results are in line with previous bimodal studies that showeddecreased activity in the sensory cortex (i.e., auditory or visual) dur-ing divided attention relative to the levels of activity observed dur-ing selective attention (Loose et al., 2003; Johnson & Zatorre,2006). This pattern of results fits the classical theory of limitedattention resource (N€a€at€anen, 1992), according to which fewer neu-ral resources are allocated for the processing of auditory stimuliwhen dividing attention to auditory and visual stimuli than whenselectively focusing attention on auditory stimuli. However, anotherpossible mechanism responsible for this divided attention effect isthat the auditory and visual systems reciprocally inhibit the process-
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–10
Attention effects on voice feedback processing 7

ing of visual and auditory stimuli, respectively (Loose et al., 2003).In the light of this hypothesis, activity in the sensory cortices thatsubserve the processing of pitch perturbations may have been sup-pressed due to the activation of sensory cortices that subserve theprocessing of flashing lights in the Bimodal Attention condition,which led to decreased cortical responses to pitch perturbations inthe divided attention condition relative to the selective attention con-dition.If the above interpretations were taken, one might hypothesise
that larger cortical responses would be elicited by pitch perturba-tions in the Bimodal Attention condition than in the Visual Atten-tion condition, because more neural resources should be devoted toauditory processing of the pitch perturbations when participantsattend to both the auditory and visual stimuli rather than the visualstimuli alone. The present findings, however, revealed smaller P2responses in the Bimodal Attention condition than in the VisualAttention condition, which is counter to this hypothesis. We specu-late that the different cortical responses elicited by the pitch pertur-bations in the divided and selective attention conditions may berelated to differences in working memory load. As compared toselectively attending to a single modality, dividing attentionbetween two modalities requires more working memory (Johnson &Zatorre, 2006). The PFC is generally thought to support executivefunctions such as the manipulation of information in working mem-ory (Petrides, 2000; Curtis & D’Esposito, 2004). For example, sev-eral neuroimaging studies have shown that the PFC is active duringdivided attention, while selective attending did not recruit frontalareas (Loose et al., 2003; Johnson & Zatorre, 2005, 2006). More-over, at least one study has shown that the requirement to divideattention between auditory and visual stimuli decreased recognitionmemory scores when the dorsolateral prefrontal cortex was tempo-rally disrupted (Johnson et al., 2007). It is thus possible that thePFC exerts top-down control over the primary sensory areas andreduces their activity during divided attention. This hypothesis issupported by several ERP studies that showed increased auditory-evoked and somatosensory-evoked responses in patients with focalPFC damage (Knight et al., 1989; Yamaguchi & Knight, 1990),suggesting an inhibitory influence of the PFC on sensory activity.Johnson & Zatorre (2006) also reported that a divided attention con-dition elicited decreased activity in the sensory cortices butincreased activity in the dorsolateral PFC relative to a selectiveattention condition. Under this view, decreased cortical responses topitch perturbations during divided attention relative to selectiveattention may be the result of inhibited activity in sensory areas dueto an increased involvement of PFC for the processing of multipleinputs to working memory. This speculation is further supported bythe smaller P2 responses we observed during the divided attentioncondition than during the control condition in which participantspassively observed the auditory and visual stimuli. Future researchusing other neuroimaging techniques or paradigms should be con-ducted to verify this speculation.Note also that the Bimodal Attention condition also elicited larger
(more negative) N1 responses to the pitch perturbations than the N1responses observed for the Bimodal Passive condition. Nonetheless,there was no systematic change in the N1 responses as a function ofselective attention. The finding that the attention effect was primarilyreflected by P2 rather than N1 may indicate that different mecha-nisms underlie the N1-P2 complex in auditory–motor processing ofpitch feedback errors. It has been suggested that N1 reflects theautomatic detection of mismatches between the predicted input andthe actual feedback in the auditory–motor processing of pitch feed-back errors, while P2 reflects interactions between the auditory and
motor system that demand higher level of cognitive processes (Behr-oozmand et al., 2014; Hu et al., 2015). On the other hand, observ-ing more negative N1 responses only during divided attention butlarger vocal responses during selective attention may reflect differen-tial effects of selective and divided attention on the auditory–motorintegration in voice control.
Conclusion
The present study investigated the influence of selective anddivided attention on the auditory–motor processing of pitch feed-back errors during vocal regulation. The results revealed the effectof selective attention by showing larger vocal compensations andlarger P2 amplitudes in response to attended vs. unattended pitchperturbations. The divided attention instruction did not lead tochanges in the magnitude or latency of the vocal response com-pared to the other conditions. However, divided attention did elicitsmaller P2 responses than selective attention and the control condi-tion. In addition, more negative N1 responses were found whenparticipants actively attended to both auditory and visual stimulithan when participants passively observed them. These findingsdemonstrate the attention-driven modulation of auditory–motor inte-gration in vocal pitch regulation. Selective attention and dividedattention appear to have differential effects on the neurobehavioralprocessing of pitch perturbations in auditory feedback during vocalpitch regulation.
Acknowledgements
This study was funded by grants from National Natural Science Foundationof China (Nos 31070990, 31371135, 81301675 and 81472154), GuangdongNatural Science Funds for Distinguished Young Scholars (No.S2013050014470) and the Fundamental Research Funds for the CentralUniversities (No. 13ykzd05). Y.L. and H.H. contributed equally to this study.All authors declare they have no competing interests.
Abbreviations
AAF, altered auditory feedback; EEG, electroencephalograph; ERP, event-related potential; F0, fundamental frequency; HL, hearing level; ISI, inter-stimulus interval; PFC, prefrontal cortex; PSS, pitch-shift stimulus; RM-ANOVA,repeated-measures analyses of variance; RSVP, rapid serial visual presenta-tion; SPL, sound pressure level.
References
Ahveninen, J., Jaaskelainen, I.P., Raij, T., Bonmassar, G., Devore, S., Hama-lainen, M., Levanen, S., Lin, F.H., Sams, M., Shinn-Cunningham, B.G.,Witzel, T. & Belliveau, J.W. (2006) Task-modulated “what” and “where”pathways in human auditory cortex. Proc. Natl. Acad. Sci. USA, 103,14608–14613.
Ahveninen, J., Hamalainen, M., Jaaskelainen, I.P., Ahlfors, S.P., Huang,S., Lin, F.H., Raij, T., Sams, M., Vasios, C.E. & Belliveau, J.W.(2011) Attention-driven auditory cortex short-term plasticity helps segre-gate relevant sounds from noise. Proc. Natl. Acad. Sci. USA, 108,4182–4187.
Alain, C. & Woods, D.L. (1997) Attention modulates auditory pattern mem-ory as indexed by event-related brain potentials. Psychophysiology, 34,534–546.
Bauer, J.J., Mittal, J., Larson, C.R. & Hain, T.C. (2006) Vocal responses tounanticipated perturbations in voice loudness feedback: An automaticmechanism for stabilizing voice amplitude. J. Acoust. Soc. Am., 119,2363–2371.
Behroozmand, R. & Larson, C.R. (2011) Error-dependent modulation ofspeech-induced auditory suppression for pitch-shifted voice feedback.BMC Neurosci., 12, 54.
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–10
8 Y. Liu et al.

Behroozmand, R., Karvelis, L., Liu, H. & Larson, C.R. (2009) Vocalization-induced enhancement of the auditory cortex responsiveness during voiceF0 feedback perturbation. Clin. Neurophysiol., 120, 1303–1312.
Behroozmand, R., Ibrahim, N., Korzyukov, O., Robin, D.A. & Larson,C.R. (2014) Left-hemisphere activation is associated with enhancedvocal pitch error detection in musicians with absolute pitch. Brain Cog-nition, 84, 97–108.
Boersma, P. (2001) Praat, a system for doing phonetics by computer. GlotInt., 5, 341–345.
Bonnel, A.M. & Hafter, E.R. (1998) Divided attention between simultaneousauditory and visual signals. Percept. Psychophys., 60, 179–190.
Burnett, T.A., Freedland, M.B., Larson, C.R. & Hain, T.C. (1998) Voice F0Responses to Manipulations in Pitch Feedback. J. Acoust. Soc. Am., 103,3153–3161.
Cai, S., Ghosh, S.S., Guenther, F.H. & Perkell, J.S. (2011) Focal manipula-tions of formant trajectories reveal a role of auditory feedback in theonline control of both within-syllable and between-syllable speech timing.J. Neurosci., 31, 16483–16490.
Chang, E.F., Niziolek, C.A., Knight, R.T., Nagarajan, S.S. & Houde, J.F.(2013) Human cortical sensorimotor network underlying feedback controlof vocal pitch. Proc. Natl. Acad. Sci. USA, 110, 2653–2658.
Chen, Z., Liu, P., Wang, E.Q., Larson, C.R., Huang, D. & Liu, H. (2012)ERP correlates of language-specific processing of auditory pitch feedbackduring self-vocalization. Brain Lang., 121, 25–34.
Chen, Z., Jones, J.A., Liu, P., Li, W., Huang, D. & Liu, H. (2013) Dynamicsof vocalization-induced modulation of auditory cortical activity at mid-utterance. PLoS One, 8, e60039.
Corbetta, M., Miezin, F.M., Dobmeyer, S., Shulman, G.L. & Petersen, S.E.(1991) Selective and divided attention during visual discriminations ofshape, color, and speed: functional anatomy by positron emission tomogra-phy. J. Neurosci., 11, 2383–2402.
Curtis, C.E. & D’Esposito, M. (2004) The effects of prefrontal lesions onworking memory performance and theory. Cogn. Affect. Behav. Ne., 4,528–539.
Eliades, S.J. & Wang, X. (2008) Neural substrates of vocalization feedbackmonitoring in primate auditory cortex. Nature, 453, 1102–1106.
Ferree, T.C., Luu, P., Russell, G.S. & Tucker, D.M. (2001) Scalp electrodeimpedance, infection risk, and EEG data quality. Clin. Neurophysiol., 112,536–544.
Flinker, A., Chang, E.F., Kirsch, H.E., Barbaro, N.M., Crone, N.E. &Knight, R.T. (2010) Single-trial speech suppression of auditory cortexactivity in humans. J. Neurosci., 30, 16643–16650.
Grady, C.L., Van Meter, J.W., Maisog, J.M., Pietrini, P., Krasuski, J. &Rauschecker, J.P. (1997) Attention-related modulation of activity in pri-mary and secondary auditory cortex. NeuroReport, 8, 2511–2516.
Hain, T.C., Burnett, T.A., Kiran, S., Larson, C.R., Singh, S. & Kenney,M.K. (2000) Instructing subjects to make a voluntary response reveals thepresence of two components to the audio-vocal reflex. Exp. Brain Res.,130, 133–141.
Hawco, C.S., Jones, J.A., Ferretti, T.R. & Keough, D. (2009) ERP correlatesof online monitoring of auditory feedback during vocalization. Psycho-physiology, 46, 1216–1225.
Heinks-Maldonado, T.H., Mathalon, D.H., Gray, M. & Ford, J.M. (2005)Fine-tuning of auditory cortex during speech production. Psychophysiol-ogy, 42, 180–190.
Hickok, G., Houde, J.F. & Rong, F. (2011) Sensorimotor integration inspeech processing: computational basis and neural organization. Neuron,69, 407–422.
Hillyard, S.A., Vogel, E.K. & Luck, S.J. (1998) Sensory gain control (ampli-fication) as a mechanism of selective attention: electrophysiological andneuroimaging evidence. Philos. T. Roy. Soc. B., 353, 1257–1270.
Hink, R.F. & Hillyard, S.A. (1976) Auditory evoked potentials during selectivelistening to dichotic speech messages. Percept. Psychophys., 20, 236–242.
Hohnsbein, J., Falkenstein, M., Hoormann, J. & Blanke, L. (1991) Effects ofcrossmodal divided attention on late ERP components. I. Simple andchoice reaction tasks. Electroen. Clin. Neuro., 78, 438–446.
Houde, J.F. & Jordan, M.I. (1998) Sensorimotor adaptation in speech pro-duction. Science, 279, 1213–1216.
Houde, J.F., Nagarajan, S.S., Sekihara, K. & Merzenich, M.M. (2002)Modulation of the auditory cortex during speech: an MEG study. J. Cogni-tive Neurosci., 14, 1125–1138.
Hu, H., Liu, Y., Guo, Z., Li, W., Liu, P., Chen, S. & Liu, H. (2015)Attention modulates cortical processing of pitch feedback errors in voicecontrol. Sci. Rep., 5, 7812.
Johnson, J.A. & Zatorre, R.J. (2005) Attention to simultaneous unrelatedauditory and visual events: behavioral and neural correlates. Cereb. Cor-tex, 15, 1609–1620.
Johnson, J.A. & Zatorre, R.J. (2006) Neural substrates for dividing andfocusing attention between simultaneous auditory and visual events.NeuroImage, 31, 1673–1681.
Johnson, J.A., Strafella, A.P. & Zatorre, R.J. (2007) The role of the dorsolat-eral prefrontal cortex in bimodal divided attention: two transcranial mag-netic stimulation studies. J. Cognitive Neurosci., 19, 907–920.
Jolicoeur, P. (1999) Restricted attentional capacity between sensory modali-ties. Psychon. B. Rev., 6, 87–92.
Jones, J.A. & Munhall, K.G. (2005) Remapping auditory-motor representa-tions in voice production. Curr. Biol., 15, 1768–1772.
Kawashima, R., Imaizumi, S., Mori, K., Okada, K., Goto, R., Kiritani, S.,Ogawa, A. & Fukuda, H. (1999) Selective visual and auditory attentiontoward utterances-a PET study. NeuroImage, 10, 209–215.
Keough, D., Hawco, C. & Jones, J.A. (2013) Auditory-motor adaptation tofrequency-altered auditory feedback occurs when participants ignore feed-back. BMC Neurosci., 14, 25.
Klingberg, T. (1998) Concurrent performance of two working memorytasks: potential mechanisms of interference. Cereb. Cortex, 8, 593–601.
Knight, R.T., Scabini, D. & Woods, D.L. (1989) Prefrontal cortex gating ofauditory transmission in humans. Brain Res., 504, 338–342.
Liu, H., Wang, E.Q., Chen, Z., Liu, P., Larson, C.R. & Huang, D. (2010)Effect of tonal native language on voice fundamental frequency responsesto pitch feedback perturbations during vocalization. J. Acoust. Soc. Am.,128, 3739–3746.
Loose, R., Kaufmann, C., Auer, D.P. & Lange, K.W. (2003) Human prefron-tal and sensory cortical activity during divided attention tasks. Hum. BrainMapp., 18, 249–259.
Mottonen, R., van de Ven, G.M. & Watkins, K.E. (2014) Attention fine-tunes auditory-motor processing of speech sounds. J. Neurosci., 34,4064–4069.
Mozolic, J.L., Joyner, D., Hugenschmidt, C.E., Peiffer, A.M., Kraft, R.A.,Maldjian, J.A. & Laurienti, P.J. (2008) Cross-modal deactivations duringmodality-specific selective attention. BMC Neurol., 8, 35.
N€a€at€anen, R. (1992) Attention and Brain Function. Psychology Press, Hills-dale, NJ.
N€a€at€anen, R., Paavilainen, P., Tiitinen, H., Jiang, D. & Alho, K. (1993)Attention and mismatch negativity. Psychophysiology, 30, 436–450.
Neelon, M.F., Williams, J. & Garell, P.C. (2006) The effects of auditoryattention measured from human electrocorticograms. Clin. Neurophysiol.,117, 504–521.
Petkov, C.I., Kang, X., Alho, K., Bertrand, O., Yund, E.W. & Woods, D.L.(2004) Attentional modulation of human auditory cortex. Nat. Neurosci.,7, 658–663.
Petrides, M. (2000) The role of the mid-dorsolateral prefrontal cortex inworking memory. Exp. Brain Res., 133, 44–54.
Picton, T.W. & Hillyard, S.A. (1974) Human auditory evoked potentials. II.Effects of attention. Electroen. Clin. Neuro., 36, 191–199.
Pugh, K.R., offywitz, B.A., Shaywitz, S.E., Fulbright, R.K., Byrd, D., Skud-larski, P., Shankweiler, D.P., Katz, L., Constable, R.T., Fletcher, J., Laca-die, C., Marchione, K. & Gore, J.C. (1996) Auditory selective attention:an fMRI investigation. NeuroImage, 4, 159–173.
Sabri, M., Binder, J.R., Desai, R., Medler, D.A., Leitl, M.D. & Liebenthal,E. (2008) Attentional and linguistic interactions in speech perception. Neu-roImage, 39, 1444–1456.
Scheerer, N.E. & Jones, J.A. (2014) The predictability of frequency-alteredauditory feedback changes the weighting of feedback and feedforwardinput for speech motor control. Eur. J. Neurosci., 40, 3793–3806.
Scheerer, N.E., Liu, H. & Jones, J.A. (2013) The developmental trajectory ofvocal and ERP responses to frequency altered auditory feedback. Eur. J.Neurosci., 38, 3189–3200.
Shomstein, S. & Yantis, S. (2004) Control of attention shifts between visionand audition in human cortex. J. Neurosci., 24, 10702–10706.
Stevens, C., Sanders, L. & Neville, H. (2006) Neurophysiological evidencefor selective auditory attention deficits in children with specific languageimpairment. Brain Res., 1111, 143–152.
Sussman, E.S., Horvath, J., Winkler, I. & Orr, M. (2007) The role ofattention in the formation of auditory streams. Percept. Psychophys., 69,136–152.
Tumber, A.K., Scheerer, N.E. & Jones, J.A. (2014) Attentional demandsinfluence vocal compensations to pitch errors heard in auditory feedback.PLoS One, 9, e109968.
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–10
Attention effects on voice feedback processing 9

Wang, J., Mathalon, D.H., Roach, B.J., Reilly, J., Keedy, S.K., Sweeney,J.A. & Ford, J.M. (2014) Action planning and predictive coding whenspeaking. NeuroImage, 91, 91–98.
Woldorff, M.G. & Hillyard, S.A. (1991) Modulation of early auditory pro-cessing during selective listening to rapidly presented tones. Electroen.Clin. Neuro., 79, 170–191.
Woldorff, M.G., Gallen, C.C., Hampson, S.A., Hillyard, S.A., Pantev, C.,Sobel, D. & Bloom, F.E. (1993) Modulation of early sensory processingin human auditory cortex during auditory selective attention. Proc. Natl.Acad. Sci. USA, 90, 8722–8726.
Woldorff, M.G., Hillyard, S.A., Gallen, C.C., Hampson, S.R. & Bloom, F.E.(1998) Magnetoencephalographic recordings demonstrate attentional
modulation of mismatch-related neural activity in human auditory cortex.Psychophysiology, 35, 283–292.
Woodruff, P.W., Benson, R.R., Bandettini, P.A., Kwong, K.K., Howard,R.J., Talavage, T., Belliveau, J. & Rosen, B.R. (1996) Modulation ofauditory and visual cortex by selective attention is modality-dependent.NeuroReport, 7, 1909–1913.
Yamaguchi, S. & Knight, R.T. (1990) Gating of somatosensory input byhuman prefrontal cortex. Brain Res., 521, 281–288.
Zarate, J.M. & Zatorre, R.J. (2008) Experience-dependent neural substratesinvolved in vocal pitch regulation during singing. NeuroImage, 40, 1871–1887.
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons LtdEuropean Journal of Neuroscience, 1–10
10 Y. Liu et al.
Related Documents











