Facies (2005) 50:463–475 DOI 10.1007/s10347-004-0026-5 ORIGINAL PAPER Humberto Astibia · Aitor Payros · Xabier Pereda Suberbiola · Javier Elorza · Ana Berreteaga · Nestor Etxebarria · Ainara Badiola · Josep Tosquella Sedimentology and taphonomy of sirenian remains from the Middle Eocene of the Pamplona Basin (Navarre, western Pyrenees) Received: 27 April 2004 / Accepted: 16 September 2004 / Published online: 3 November 2004 # Springer-Verlag 2004 Abstract Sirenian vertebrae and ribs have been recently discovered from two Middle Eocene localities of the Pamplona Basin, Navarre (western Pyrenees). These outcrops correspond to different lower Bartonian lithos- tratigraphic units: the lower part of the Pamplona Marl Formation (Uztarrotz site) and the upper part of the Ar- danatz Sandstone (Ardanatz site). The former represents a deep and low-energy sea floor far away from a deltaic slope; the Ardanatz environment probably corresponds to a semi-closed deltaic bay periodically affected by catas- trophic floods (i.e., fluvial hyperpycnal flows). The presence of epibiontic activity suggests that the bones were exposed for a while prior to the burial. The histo- logical structures are well preserved except in the pe- ripheral region, where tubular-like microstructures filled by pyrite and iron oxides probably correspond to micro- bial bioerosion. The major mineral component of the fossil bones is francolite (carbonate fluorapatite). In the Ardanatz samples there is evidence of secondary franco- lite due to the late replacement of original carbonate fluorapatite through internal fractures. The Ardanatz and Uztarrotz sirenian fossils do not show any evidence of reelaboration. They have similar sum of rare earth ele- ments (REE) concentrations relative to the host rock, but comparatively lower than in other vertebrate fossil bones. This feature may be due to the dense compact structure of pachyosteosclerotic sirenian bones. Keywords Sirenians · Taphonomy · Geochemistry · Sedimentology · Eocene · Western Pyrenees Introduction Sirenian fossils are relatively abundant in the Eocene formations of the Mediterranean Tethys (Sagne 2001a, 2001b). In the Pyrenean Realm, sirenian remains have been described in a number of Middle and Upper Eocene localities from the southern foothills of the Catalonian Basin (Pilleri et al. 1989; Galobart et al. 1992). On the contrary, the sirenian remains hitherto found in the western Pyrenees are rather scarce. In the Jaca Basin (Huesca), Pilleri et al. (1989) described a caudal vertebra from the Middle Eocene of Santa Cilia de Jaca, and GɃ- mez (1998) listed a mandible with teeth and postcranial bones from the Middle to Upper Eocene BelsuȖ-AtarȖs Formation of Oroel and Used. Moreover, Astibia et al. (1999) described two isolated caudal vertebrae from the Middle Eocene of Santa Cilia de Jaca in Huesca and from Ardanatz de Egues in Navarre (Fig. 1A). Recently, additional sirenian remains have been found in the Middle Eocene of the Pamplona Basin (Ardanatz and Uztarrotz localities, Navarre). The material, which consists of disarticulated vertebrae and ribs, will be de- scribed in detail elsewhere. The present work focuses on the sedimentological and taphonomic aspects, with spe- cial emphasis in petrological and geochemical features of the fossils. H. Astibia ( ) ) · A. Payros · X. P. Suberbiola · A. Berreteaga · A. Badiola Departamento de Estratigrafȷa y Paleontologȷa, Universidad del Paȷs Vasco/EHU, Apartado 644, 48080 Bilbao, Spain e-mail: [email protected] J. Elorza Departamento de Mineralogȷa y Petrologȷa, Universidad del Paȷs Vasco/EHU, Apartado 644, 48080 Bilbao, Spain N. Etxebarria Departamento de Quȷmica Analȷtica, Universidad del Paȷs Vasco/EHU, Apartado 644, 48080 Bilbao, Spain J. Tosquella Departamento de GeodinƁmica y Paleontologȷa, Universidad de Huelva, Campus del Carmen, 21071 Huelva, Spain

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Facies (2005) 50:463–475DOI 10.1007/s10347-004-0026-5
O R I G I N A L P A P E R
Humberto Astibia · Aitor Payros ·Xabier Pereda Suberbiola · Javier Elorza ·Ana Berreteaga · Nestor Etxebarria · Ainara Badiola ·Josep Tosquella
Sedimentology and taphonomy of sirenian remainsfrom the Middle Eocene of the Pamplona Basin(Navarre, western Pyrenees)Received: 27 April 2004 / Accepted: 16 September 2004 / Published online: 3 November 2004� Springer-Verlag 2004
Abstract Sirenian vertebrae and ribs have been recentlydiscovered from two Middle Eocene localities of thePamplona Basin, Navarre (western Pyrenees). Theseoutcrops correspond to different lower Bartonian lithos-tratigraphic units: the lower part of the Pamplona MarlFormation (Uztarrotz site) and the upper part of the Ar-danatz Sandstone (Ardanatz site). The former represents adeep and low-energy sea floor far away from a deltaicslope; the Ardanatz environment probably corresponds toa semi-closed deltaic bay periodically affected by catas-trophic floods (i.e., fluvial hyperpycnal flows). Thepresence of epibiontic activity suggests that the boneswere exposed for a while prior to the burial. The histo-logical structures are well preserved except in the pe-ripheral region, where tubular-like microstructures filledby pyrite and iron oxides probably correspond to micro-bial bioerosion. The major mineral component of thefossil bones is francolite (carbonate fluorapatite). In theArdanatz samples there is evidence of secondary franco-lite due to the late replacement of original carbonate
fluorapatite through internal fractures. The Ardanatz andUztarrotz sirenian fossils do not show any evidence ofreelaboration. They have similar sum of rare earth ele-ments (REE) concentrations relative to the host rock, butcomparatively lower than in other vertebrate fossil bones.This feature may be due to the dense compact structure ofpachyosteosclerotic sirenian bones.
Keywords Sirenians · Taphonomy · Geochemistry ·Sedimentology · Eocene · Western Pyrenees
Introduction
Sirenian fossils are relatively abundant in the Eoceneformations of the Mediterranean Tethys (Sagne 2001a,2001b). In the Pyrenean Realm, sirenian remains havebeen described in a number of Middle and Upper Eocenelocalities from the southern foothills of the CatalonianBasin (Pilleri et al. 1989; Galobart et al. 1992). On thecontrary, the sirenian remains hitherto found in thewestern Pyrenees are rather scarce. In the Jaca Basin(Huesca), Pilleri et al. (1989) described a caudal vertebrafrom the Middle Eocene of Santa Cilia de Jaca, and G�-mez (1998) listed a mandible with teeth and postcranialbones from the Middle to Upper Eocene Belsu�-Atar�sFormation of Oroel and Used. Moreover, Astibia et al.(1999) described two isolated caudal vertebrae from theMiddle Eocene of Santa Cilia de Jaca in Huesca and fromArdanatz de Egues in Navarre (Fig. 1A).
Recently, additional sirenian remains have been foundin the Middle Eocene of the Pamplona Basin (Ardanatzand Uztarrotz localities, Navarre). The material, whichconsists of disarticulated vertebrae and ribs, will be de-scribed in detail elsewhere. The present work focuses onthe sedimentological and taphonomic aspects, with spe-cial emphasis in petrological and geochemical features ofthe fossils.
H. Astibia ()) · A. Payros · X. P. Suberbiola · A. Berreteaga ·A. BadiolaDepartamento de Estratigraf�a y Paleontolog�a,Universidad del Pa�s Vasco/EHU,Apartado 644, 48080 Bilbao, Spaine-mail: [email protected]
J. ElorzaDepartamento de Mineralog�a y Petrolog�a,Universidad del Pa�s Vasco/EHU,Apartado 644, 48080 Bilbao, Spain
N. EtxebarriaDepartamento de Qu�mica Anal�tica,Universidad del Pa�s Vasco/EHU,Apartado 644, 48080 Bilbao, Spain
J. TosquellaDepartamento de Geodin�mica y Paleontolog�a,Universidad de Huelva,Campus del Carmen, 21071 Huelva, Spain

Geological setting
The current distribution of the Eocene rocks in the SouthPyrenean zone shows the geometry of the original basin(Fig. 1A; also see Pujalte et al. (2002), for an updatedsynthesis). In Eocene time, the South Pyrenean zone wasa NW-trending foreland basin developed in front of thetectonically uplifting orogen to the north, and it remainedas a narrow arm of the sea entering from the Bay ofBiscay. However, its palaeogeography changed in re-sponse to the increasing tectonic deformation. Thus, inEarly-Middle Eocene (Ypresian-Lutetian) time the SouthPyrenean basin corresponded to a deep marine trough,flanked in the south by extensive shallow-water carbonateramps and axially filled up by siliciclastic sediments de-rived from the Pyrenean axial zone. During this stage, themain denudation site was located in the emergent easternPyrenees, where large fluviodeltaic systems developedand fed westwards the so-called Hecho deep-sea subma-rine fan. This palaeogeographic configuration progres-sively encroached westwards, so that denudation of the
orogen began in the western Pyrenees by Middle-LateEocene (Bartonian-Priabonian) time, and the fluviodeltaicsedimentary context became widespread throughout theforeland basin.
This tectonosedimentary evolution is clearly recordedin the Eocene succession of the Pamplona Basin (Figs. 1Band 2). Ypresian to Lutetian shelfal limestones and deep-sea flysch deposits (lower part of the Hecho Group ofMutti (1984)) are overlain by Bartonian to Priabonianmarly deposits, which have been divided into lithostrati-graphic units on the basis of interbedded sandy intervals(Fig. 1B). The marly succession starts with the Erize andMendiorotz formations, attributed to the lower BartonianP12 planktic foraminiferal Zone of Berggren et al. (1995)(see Payros 1997). The Erize Formation is composed ofhemipelagic marls interspersed with thin calciturbidites,and correspond to distal carbonate ramp deposits con-nected to the SW with shallow-water limestones. Theygrade to the NE into hemipelagic marls and thin, fine-grained siliciclastic turbidites of the Mendiorotz Forma-tion (upper part of the Hecho Group).
Fig. 1 Simplified geologicalmaps of the Pyrenees ( A), withlocation of the study area in thewestern part of the South Pyre-nean zone ( SPZ), and of thePamplona Basin ( B). The sire-nian localities (Ardanatz andUztarrotz, Navarre) are high-lighted in boxes and other sitesreferred to in the text have beenplaced for situation purposes
464

The Erize Formation is overlain by a discontinuoussandy unit (Ezkaba Formation (Payros 1997; Payros et al.1997) dated as lower Bartonian (P13 Zone). The EzkabaFormation is lacking to the east of the Pamplona Basin,but a glauconitic silty unit (Urrotz level (Puigdefabregas1975)) occurs at the same stratigraphic position. TheEzkaba Formation is interpreted as a channel-levee tur-bidite system fed from the orogen to the north; therefore,it records the onset of the tectonically induced emergence(and denudation) of the western Pyrenean axial zone.
The Pamplona Formation (Mangin 1959–60) is com-posed of homogeneous blue marls with sparse thin-bed-ded, fine-grained sandy turbidites. Regional studies haveshown that the Pamplona Formation amply extendseastwards. In the east of Navarre it shows very similarfeatures to those in the Pamplona Basin but the amount ofturbidites increases (Puigdefabregas 1975). Further east-wards, in the Jaca Basin, similar and coeval marls havebeen interpreted as shelfal (Arguis Formation) andprodeltaic (Pamplona Formation) deposits, the latterhaving been fed from a deltaic system to the ESE repre-sented nowadays by the Belsu�-Atar�s Sandstone For-mation (Puigdefabregas 1975).
A second sandy unit, not formally defined yet butcommonly referred to as the Ardanatz Sandstone (e.g.Ruiz de Gaona 1947), occurs on top of the PamplonaFormation in the Pamplona Basin. The Ardanatz Sand-stone is composed of alternating sandstones and siltymarls that extend from the Etxauri Valley in the west ofthe Pamplona basin to the Egues and Aranguren valleys inthe east, where the unit outlines the NW-trending Ara-nguren syncline. In this areas the marly interbeds yieldabundant shallow-marine microfossils and macrofossils(Ruiz de Gaona 1947; Ruiz de Gaona and Colom 1950;Mangin 1959–60), including nummulitids that, accordingto Astibia et al. (1999), pertained to the early Bartonian(upper part of the SBZ17 Zone of Serra-Kiel et al.(1998)). The Ardanatz Sandstone can be traced further tothe east, bordering the Izaga syncline, but it shows dif-ferent sedimentological features. Here the sandstone bedsare graded and contain abundant mud clasts, and they arerestricted to large-scale, lens-shaped lithosomes. Ac-cording to these features, they were interpreted as tur-biditic channels (Puigdefabregas 1975).
The Ardanatz Sandstone gradually passes upwards intothe Ilundain Marls Formation (Mangin 1959–60), whichdiffers form the Pamplona Formation in higher silt con-tent. Small-sized bivalves classified as Dymia pamplo-nensis (Calzada and Astibia 1996), most probably op-portunistic species, are locally abundant in the transitionfrom the Ardanatz Sandstone to the Ilundain Formation.All these characteristics suggest shallower and more re-stricted marine environments than the Pamplona Forma-tion. The regional study by Puigdefabregas (1975) dem-onstrated that the Ilundain Formation is related to theprogradation of the deltaic Belsu�-Atar�s Formation fromthe ESE.
The Priabonian Gendulain Formation (Puigdefabregas1975) is mostly composed of coastal deposits divided into
Fig. 2 Representative columnar section of the Upper Eocene marlysuccession in the Pamplona Basin (Navarre), with its constituentlithostratigraphic units and the depositional environments so farproposed in the literature (see text for explanations). The strati-graphic position of the levels containing sirenian remains in Ar-danatz (A) and Uztarrotz (U) is indicated
465

three distinct members: lower evaporites, middle marls,and upper sandstones. Lateral variations in facies andthicknesses record the synsedimentary tectonic deforma-tion of the Pamplona Basin (Cendon et al. 1998; Payros etal. 2000), as it progressively became a detached piggy-back basin atop of southward displacing thrust sheets.Owing to the concomitant tectonic uplift, the GendulainFormation records the last episode of deltaic progradationfrom the ESE, the closure of the South Pyrenean Sea, andthe definite evolution of the Pamplona Basin into terres-trial conditions.
The sirenian fossils in the Pamplona Basin were col-lected in two outcrops (Uztarrotz and Ardanatz; Fig. 1B)located in the Egues Valley, few kilometres east of thePamplona/Iru�ea city, displaying different lithostrati-graphic units. The fossils from Uztarrotz come from thelower part of the Pamplona Formation and, therefore, theyare lower Bartonian in age (on the basis of age data fromthe under- and overlying units, the Pamplona Formationcorresponds to the P14 Zone of Berggren et al. (1995)).The sirenian fossils from Ardanatz come from a marlyinterval within the upper part of the Ardanatz Sandstone,just in the transition zone into the overlying IlundainFormation. According to Astibia et al. (1999), the Ar-danatz fossils are early Bartonian (upper part of theSBZ17 Zone of Serra-Kiel et al. (1998)) in age.
Sedimentology and palaeoenvironments
Uztarrotz outcrop
Description
The marly deposits in Uztarrotz share the sedimentologi-cal characteristics of the lower part of the PamplonaFormation elsewhere. They are monotonous and homo-geneous blue marls, with few fine-grained sandstone thinbeds interspersed. Except for the sirenian remains studiedhere, the only macroscopic fossils are centimetre-scalefragments of crinoid stems with pentameral shapedcolumnar plates (isocrinids, ? Isselicrinus sp.), quitecommon throughout the lower Pamplona Formation. Ac-cording to Ruiz de Gaona and Colom (1950) andPuigdefabregas (1975), the micropalaeontological assem-blage in the lower part of the Pamplona Formation istypified by ostracods, gastropods (mostly pteropods), highpercentage of planktic foraminifers, and diversified bent-hic foraminifers.
Interpretation
The marly deposits are formed by the mixture of biogenic,open-marine carbonate particles with siliciclastic mud,certainly terrestrial in origin. The latter was probablycarried by hypopycnal buoyant plumes (sensu Wright1977) emanating from river mouths. The importance ofhypopycnal plumes as mud-suppliers in tectonically ac-
tive settings, such as the Bartonian Pamplona Basin, hasbeen amply discussed by Nemec (1995). The envisagedenvironment corresponds to a low-energy sea floor, belowstorm-wave base, where sediments settled quietly. Thetype and scarcity of macrofossils confirm such an inter-pretation and suggest an environment below the photiczone, as crinoids are not abundant in shallow seas andpedunculate crinoids are most common in deep waters,recent ones usually living deeper than 100 m (Sprinkleand Kier 1987; Clarkson 1992). Similarly, according tothe microforaminiferal content, Ruiz de Gaona and Co-lom (1950) proposed that the lower Pamplona Formationis composed of deep-water marls, while Puigdefabregas(1975) specified that deposition occurred deeper than300 m in a prodelta environment. Indeed, our revision ofthe benthic microforaminifers listed and the benthic/planktic foraminiferal ratio provided by those authorsconfirms that the assemblages and values are comparableto present day outer shelf and slope environments (A.Cearreta, personal communication). However, the scarcityof sandy thin beds in the Uztarrotz outcrop, which areinterpreted as episodic deposits from diluted and ex-panded turbidity currents, and the lack of any depositfrom rheologically plastic sediment gravity flows (e.g.mud flows, debris flows, slumps or slides) suggest a de-positional site far away from the deltaic slope whereresedimentation processes initiated. Therefore, a deep andeven sea floor is here considered.
Ardanatz outcrop
Description
The Ardanatz Sandstone in the Aranguren Syncline canbe divided into two distinct intervals. The lower intervalcrops out in the southern limb of the syncline, where itreaches 20 m in thickness. It is composed of coarse-grained sandstones with abundant imbricate fragments ofred algae and bryozoans. The bioclastic sandstones formamalgamated sheet-like beds, up to 0.5 m thick, withgradational transitions.
The upper interval crops out in both limbs of theAranguren Syncline, but it is best exposed in the Ardanatzarea and the following description corresponds to thoseoutcrops. Silty marls with carbonaceous plant remainsand vertical and horizontal burrows are dominant. Theirmicropalaeontological association is dominated by bent-hic foraminifers (e.g. Trochammina, Rotalina, Brizalina,Bulimina, Cibicides, Nonion, Haplophragmoides, Re-ophax), ostracods and charophyte gyrogonites (M. Lam-olda, personal communication; also see Ruiz de Gaona1947; Ruiz de Gaona and Colom 1950). In addition, theycontain macrofossils of corals, bryozoans, tubicolous an-nelids and molluscs (many of them epiphytic), andaquatic vertebrate remains (shark teeth, turtle platesstudied by Astibia et al. (1999), as well as sirenian ver-tebrae and ribs). The invertebrate assemblage is charac-
466

terised by the great abundance of remains, the small sizeof the specimens and the low specific diversity.
The silty marls contain interbedded sandstones withabundant carbonaceous plant remains and some bioclasts(e.g. Nummulites). In general, these beds are tabular anddisplay sharp bases with occasional groove casts (indi-cating ENE palaeocurrent direction), whereas their upperboundaries are gradational. They show a high degree ofbioturbation with burrowing structures attributable toThalassinoides and Ophiomorpha. When bioturbation hasnot completely destroyed the original depositional fea-tures, the sandstone beds show a lower part where normalgrading is the only visible feature and an upper one withparallel lamination; cross lamination has been observed insome beds.
The thickness and grain size of the sandstone bedsrhythmically vary along the succession. Thus, severalthickening- and coarsening-upward sequences, rangingfrom some metres to a few decametres in scale, can bedistinguished. The lower parts of the sequences aredominated by the silty marls and contain just a few in-terspersed sandstone beds, which are thin (10–30 cm),fine-grained and pervasively burrowed; the Ardanatzsirenian remains described below were collected in thelower part of one sequence. The upper parts of the se-quences contain medium-grained sandstones, the degreeof bioturbation is lower, the beds are thicker (50–70 cm)and they are commonly amalgamated in packages up to2 m in thickness.
Interpretation
The mixed composition of the lower Ardanatz Sandstoneindicates that an autochthonous, shallow-marine carbon-ate sediment production and a terrestrial siliciclasticsediment flux coexisted. The coarse grain size and theimbricate grain fabric suggest that the depositional settingwas under the effect of continuous high-energy currents,above fair weather wave base.
In the upper Ardanatz Sandstone the predominant siltymarls with abundant marine fossils point out to a low-energy sea floor. Most of the invertebrate macrofossilscorrespond to organisms whose current relatives mainlylive in shallow (in general less than 60 m deep) marineenvironments covered by sea-grass meadows. Therefore,they indicate a relatively shallow sea floor, within thephotic zone, where sea-grasses played a significant role indissipating the action of marine currents. The invertebratemacrofossil assemblages show low specific diversity andsmall-sized specimens if compared to other Bartoniansites in Europe, such as Barisseuse in the Paris Basin(Morellet and Morellet 1945; Patzkowsky and Bezusko-Layou 1999) or Barton-on-Sea in England (West 2002;Morton 2003), where the fossil assemblages are com-posed of hundreds of invertebrate species. These charac-teristics suggest some kind of palaeoecological andpalaeoenvironmental stresses in the Ardanatz Sea. It wasprobably consequence of some geographical restriction
that left the area apart from the open ocean. On the otherhand, the high terrigenous content, the abundance ofcarbonaceous plant remains and the occurrence ofcharophyte gyrogonites readily indicate terrestrial sedi-ment supply and suggest certain proximity to rivermouths. Indeed, it is considered that the large amount ofterrestrial fine-grained sediment was carried in suspensionby steady hypopycnal buoyant effluents (Nemec 1995).The microforaminiferal association also suggests innerplatform environments with high terrestrial input (A.Cearreta, personal communication).
The tabular sandstone layers record occasional, butfrequent, episodes of comparatively sudden input ofcoarser-grained siliciclastic sediment. Each event wasrelated to a high-energy current that sharply burst into thedepositional site and progressively waned. Therefore, thetransport and deposition of the sandy sediments was theresult of non-confined, high-concentration turbulentflows. Taking into account that the sandstone beds areencased within restricted, shallow-marine marls with ahigh content of terrestrially derived particles, it is con-sidered that the sandstone beds could also have formed inresponse to fluvial supplies, not in normal conditions,though, but in flood periods. Under these conditions theinertia of hyperconcentrated, sediment-laden effluentswas great enough to overcome the deceleration producedby friction with the sea floor and by partial mixing withthe ambient seawater. Thus, they continued flowing andcarrying sand into the sea as hyperpycnal jets (inverselybuoyant plumes; Wright 1977; Mulder et al. 2003). Theresult of each flooding event would be a sandstone bed.The common presence of marly interbeds argues againstsustained hyperpycnal flows generated by “dirty rivers”(sensu Mulder and Syvitski 1995; Mulder et al. 2003),which would have deposited amalgamated sandstonebeds, but for “cleaner rivers” capable of generating hy-perpycnal flows only during major flood events. The or-igin and characteristics of hyperpycnal fluvial effluentshave attracted increasing sedimentological interest in re-cent years (e.g. Martinsen 1990; Sparks et al. 1993;Mulder and Syvitski 1995; McLeod et al. 1999; Mulder etal. 2003). However, the role of catastrophic floods insupplying sediment to marine depositional sites as well asthe stratigraphic importance of their accumulations arestill largely overlooked and underestimated (Mutti et al.1996, 2000, 2003), few further case studies having beenpublished hitherto (e.g. Haines et al. 2001; Mellere et al.2002; Plink-Bj�rklund and Steel 2004).
Successive thickening and coarsening-upward se-quences record the punctuated progradation of flood-re-lated submarine sand bodies. Taking into account thatthese sand bodies were sourced by expanded and non-confined, flood-related hyperpycnal flows emanatingfrom river mouths, that they formed in a generally quietmarine zone, and that they experienced a punctuatedprogradational evolution, it can be inferred that theyprobably corresponded to lobe-like accumulations. Thus,they share many characteristics with the so-called “flood-generated delta-front sandstone lobes” (Mutti et al. 2000,
467

2003). According to these authors, these lobes form inflood-dominated fluviodeltaic systems (i.e. “small andimmature fluvial systems with relatively high-elevationcatchment areas and short and high-gradient transferzones in mountainous regions best developed in tectoni-cally active settings”) that supply a high sediment fluxinto the sea, mostly during catastrophic flood stages thatgenerate hyperpycnal flows entering the marine deposi-tional zone. Such a palaeogeographical setting broadlyfits that envisaged above for the Bartonian PamplonaBasin with the synorogenic progradation of the deltaicBelsu�-Atar�s Formation delta from the ESE.
In summary, the palaeoenvironment of the Ardanatzarea corresponds to a shallow restricted sea in a tectoni-cally active basin. It was probably a semi-closed deltaicbay protected from the influence of open marine currentsbut exposed to strong fluvial influences. The bay marginswere covered by bioclastic sands similar to those in thelower Ardanatz Sandstone. Autochthonous carbonatebioclasts and siliciclastic grains emanating from rivermouths were continuously reworked and mixed by littoralcurrents. The central part of the bay, on the basis of theupper Ardanatz Sandstone, was a deeper, but still shallow,quiet zone covered by sea-grass meadows. Autochthonouscarbonate mud with abundant biogenic remains was thepredominant type of sediment. In addition, terrestrialsupply also occurred, as steady hypopycnal buoyantplumes flowing out from river mouths carried silt grainsand plant remains. Episodic, flood-related hyperpycnaljets provided coarser grained sediments that formedprogradational, lobe-shaped sand bodies. The ENE-di-rected paleocurrent indicators in the Ardanatz Sandstone
beds and the general distribution of facies (shallow-waterdeposits in the western half of the Pamplona Basin, andturbiditic channels to the east) show the palaeopolarity ofthe Ardanatz Bay and suggest that the main deltaic bodylay to the south of the present day outcrops.
Taphonomy
Material and methods
The sirenian material consists of disarticulated vertebrae and ribs(Fig. 3). The Uztarrotz outcrop has yielded a number of anteriorthoracic vertebrae, mainly neural arches, and rib fragments(Fig. 3A–C). The accurate position of the remains in the strataremains unknown. The fossils were found in anatomical relation, sothey presumably come from the same individual. The Ardanatzfossils consist of fragmentary dorsal, sacral and caudal vertebrae, aswell as ribs (Fig. 3D–F). Most of the remains were collected from amarly layer intercalated within the upper part of the ArdanatzSandstone and probably belong to a single individual. Additionalremains come from another close marly outcrop in the same Ar-danatz area. All this material is provisionally referred to as Sireniaindet. Pending their study, the sirenian remains are kept in theDepartamento de Estratigraf�a y Paleontolog�a of the Universidaddel Pa�s Vasco (UPV/EHU).
The mineralogical nature of the fossil bones was assessed inseveral ways: petrological techniques, X-ray diffraction (XRD) andgeochemical analyses (ICP-AES, ICP-MS). Fossil bones used forthis study were prepared as thin sections for histological analysis.Thin sections were examined under reflected and transmitted light.The Cathodoluminescence (CL) work employed a Technosyn ColdCathodoluminescence System, model 8200 Mk II, mounted on anOlympus triocular research microscope with a maximum magnifi-cation capability of �400, utilising universal stage objectives.Standard operating conditions included an accelerating potential of12 kV and 0.5–0.6 mA beam current with a beam diameter of
Fig. 3 Sirenian remains fromthe Middle Eocene of the Pam-plona Basin, Navarre. A–B Uz1.6, anterior dorsal vertebrafrom Uztarrotz in anterior andleft lateral views; C Uz 1.22,fragmentary dorsal rib fromUztarrotz in dorsal view; D Ar1.6, posterior dorsal vertebrafrom Ardanatz in posteriorview; E Ar 1.9, sacral vertebrafrom Ardanatz in posteriorview; F Ar 1.22, fragmentarydorsal rib in dorsal view. Scaleline in cm
468
approximately 5 mm. Fragment fossil bones were photographed intransmitted and reflected light and under CL observation.
X-ray diffraction analyses were performed with a PhillipsPW1710 diffractometer using Cu Ka radiation monochromated bygraphite with generator conditions of 40 kV, 20 mA, with a stepsize of 0.02 (2 q) and a time per step of 1 s. The crystallinity index(CI) was determined from X-ray diffractograms, and measured asthe value in degrees 2 q of the width of the (002) peak measured athalf the peak height (Blumenthal et al. 1975; Sillen 1989).
Density is one of the most important physical properties of solidmaterials. There are at least six general methods available for de-termining the density of the solids. One of them is known as the
Archimedes method, adapted for microfragment samples by Romanand Gutierrez-Zorrilla (1985). The density of six sirenian sampleshas been calculated following this method (Table 1).
Ten sirenian remains (six from Ardanatz and four from Uztar-rotz) were analysed for their content in major, minor and traceelements and REEs (Table 2). The samples correspond to sirenianribs (Uz 1.24, 1.25, 1.26, 1.27, Ar 1.26, 1.27, Ar 1.28, Ar 2.1 and Ar2.2). A turtle plate (Ar 3.1) was also analysed for comparison.Fossils were ground to powder with an agate mortar and pestle and/or mill and placed in clean glass sample vessels. The major elementmeasurements (wt%) were determined by inductively coupledplasma emission spectrometry (ICP-AES) using an ARL 3410model at the Departamento de Qu�mica Anal�tica of the UPV/EHUof Bilbao. The trace elements and REEs of the fossil samples weremeasured by inductively coupled plasma mass spectrometry anal-ysis (ICP-MS) at the Adirondak Laboratory of the Parque Tec-nol�gico de Zamudio (Biscay). All the REE results have beennormalized to PAAS (Post-Archean Australian Shales) (Taylor andMcLennan 1985).
The statistical test of Mann-Whitney U was carried out with theaid of the SPSS/PC version 10.0. Statistical methods of principalcomponents analysis (PCA) were conducted using Unscrambler 7.5(Camo AS).
Table 1 Crystallinity index (CI) and density of the sirenian fossilsfrom Uztarrotz and Ardanatz (Navarre)
Samples C.I. Density
Ar 1.26 0.307 2.94Ar 1.30 0.270 2.93Ar 1.31 0.258 2.90Ar 2.2 0.251 2.87Uz 1.27 0.267 2.97Uz 1.28 0.275 2.93
Table 2 Summary of all the results of geochemical analyses (ma-jor, minor, trace and REEs) of the fossils from Ardanatz (Ar) andUztarrotz (Uz), Navarre. All the samples are sirenian ribs except Ar
3.1 (turtle plate) and Uz S1 (host rock). Major element are ex-pressed in % and minor, trace and REEs in ppm
ppm Uz 1.26 Uz 1.25 Uz 1.24 Uz 1.27 Ar 1.28 Ar 1.26 Ar 2.1 Ar 2.2 Ar 3.1 Uz.S1
Ba 901.19 3,772.93 4,438.96 2,295.91 4,354.60 5,894.68 961.28 927.83 111.74 309.30Ce 111.86 101.27 309.49 297.80 94.23 82.62 161.63 116.61 262.46 93.94Co 10.21 12.27 11.95 9.77 12.86 3.81 11.13 2.00 11.99 6.13Cr 10.52 8.74 7.28 7.64 4.47 4.76 5.97 5.88 6.01 67.55Dy 17.83 12.96 60.09 62.30 12.97 10.67 23.84 16.60 56.68 4.92Eu 4.07 3.75 11.00 10.65 4.02 3.79 5.96 4.47 9.77 2.93Ga 11.63 24.39 28.70 15.37 22.37 27.67 9.25 9.11 7.04 17.66Gd 13.15 20.46 71.72 60.17 26.75 28.41 27.94 16.78 46.74 1.39Ho 5.93 5.05 13.01 13.35 4.68 4.35 6.33 5.48 12.14 3.21La 46.38 35.94 135.33 130.22 28.85 24.75 66.37 49.44 125.60 24.26Lu 3.69 3.56 4.36 4.47 3.32 3.32 3.56 3.52 4.12 3.25Nd 32.53 24.95 162.95 160.00 28.56 21.94 74.35 42.18 138.24 20.99Ni 41.28 56.66 30.87 29.09 54.42 22.52 27.46 11.60 93.23 30.93Pb 19.57 14.35 196.19 25.01 27.38 11.43 11.87 11.60 19.80 14.11Pr 10.07 8.37 40.39 38.77 8.92 7.34 18.73 11.93 33.97 7.65Sm 9.72 8.42 37.94 38.28 9.66 8.35 19.38 12.14 34.02 6.74Th 0.15 1.71 1.13 0.67 0.84 0.36 1.29 0.98 0.23 11.54Tb 4.04 3.46 10.56 10.71 3.57 3.15 5.29 4.11 10.01 2.41Tm 3.68 3.41 5.12 5.31 3.16 3.16 3.60 3.41 4.80 2.87U 4.90 5.78 7.35 7.61 28.17 29.16 29.63 21.09 15.79 3.57V 178.02 150.58 137.13 140.01 120.30 137.80 163.96 146.25 178.45 181.32Y 164.69 117.17 416.61 428.79 78.43 68.56 140.74 127.73 364.11 19.09Yb 8.22 6.68 14.00 15.15 4.93 5.04 6.73 6.39 12.19 3.93Zn 160.34 174.83 121.12 120.38 165.76 129.68 158.12 148.16 655.16 115.09Zr 2.58 209.60 60.99 206.34 41.67 9.30 37.54 75.42 12.54 409.75(La/Sm) 4.77 4.27 3.57 3.40 2.99 2.97 3.42 4.07 3.69(La/Yb) 5.64 5.38 9.66 8.60 5.85 4.91 9.86 7.73 10.30S REE 267.49 234.87 870.84 841.87 230.46 203.72 420.10 289.66 745.94 175.61%
Al2O3 0.01 0.01 0.00 0.01 0.04 0.00 0.00 0.01 0.06 9.91CaO 40.93 41.20 42.35 41.34 39.69 41.65 39.56 40.51 38.74 22.70Fe2O3 2.51 2.64 1.77 2.15 4.20 2.26 3.35 1.80 4.79 3.72MgO 0.31 0.18 0.16 0.17 0.19 0.16 0.19 0.18 0.33 2.19MnO 0.01 0.01 0.01 0.01 0.00 0.00 0.00 0.00 0.02 0.04Na2O 0.82 0.74 0.65 0.79 0.69 0.77 0.79 0.78 0.48 0.34P2O5 31.10 30.44 29.87 31.23 29.22 29.94 28.13 31.54 22.65 0.06SiO2 0.41 0.40 0.44 0.52 0.55 0.47 0.48 0.59 0.73 27.21TiO2 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.44
469

Petrological characteristics, X-ray diffraction,crystallinity indices, density and cathodoluminescence
Uztarrotz fossils
Description
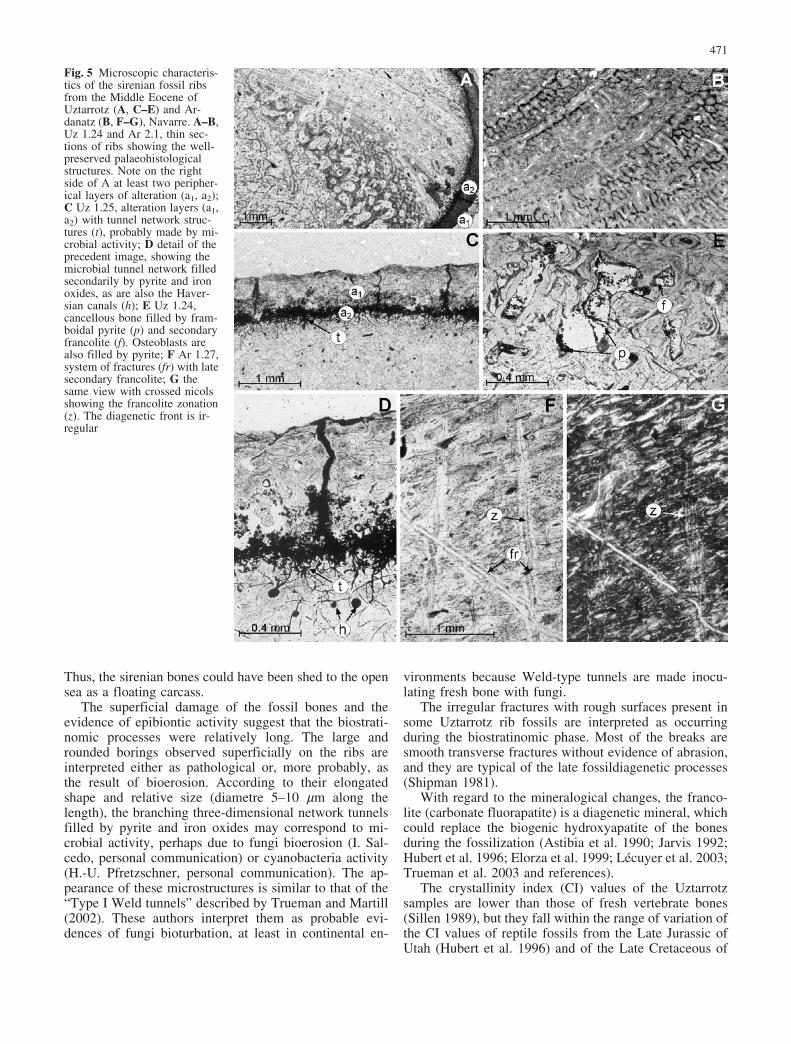
The fossil bones are eroded, especially the ends of thevertebral apophyses and the rib surfaces (Fig. 3A–C). Theexternal layer of the ribs is mostly missing. In somepieces, there is evidence of epibiontic activity, e.g. cal-careous tubes of annelids and small bivalves (Fig. 4A).
Several ribs show externally (superficially) hemi-spheric borings whose diameter is mainly between 1 and3 cm (Fig. 4B). The ribs are generally broken and occur asfragments about 1–5 cm long. Some of the fractures areirregular and moderately abraded. Most of them, how-ever, are smooth transverse fractures without abrasion.Epibionts are distributed on the borings and on the su-perficial cracks, and so they probably colonized the sub-stratum once eroded the bones.
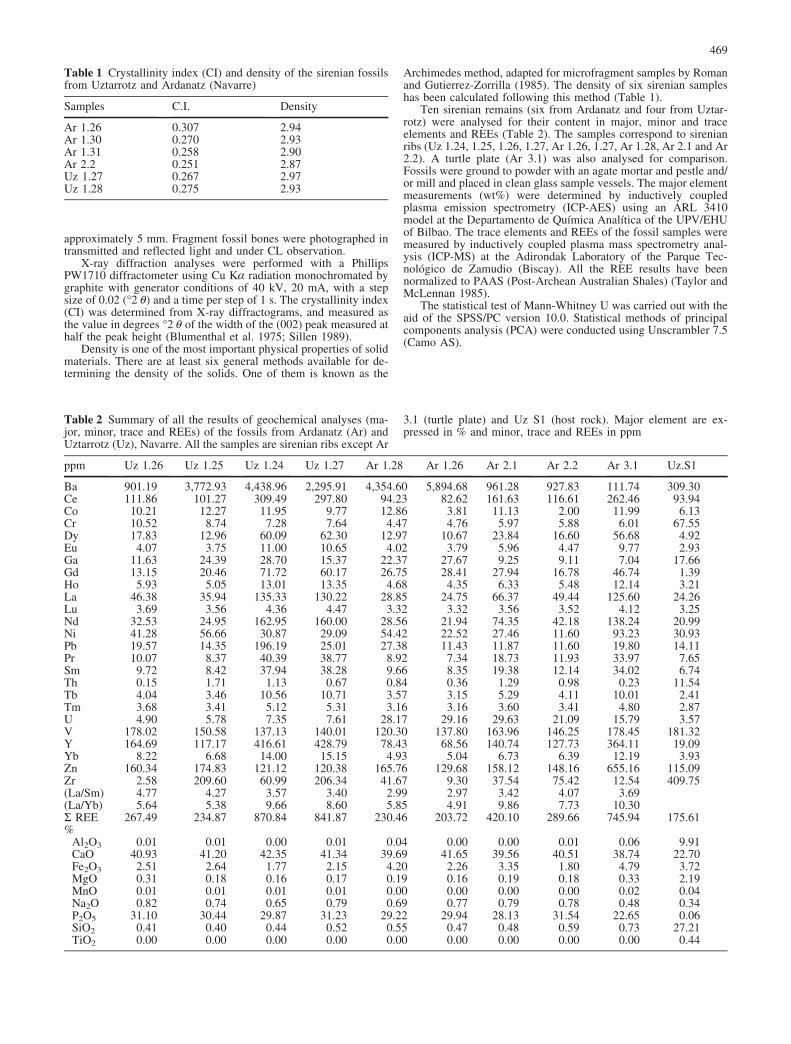
Histologically, the first and second order structures(e.g., compact bone, secondary osteons, lines of arrestedgrowth; see Francillon-Vieillot et al. 1990) are well pre-served. The thin sections of the ribs show the micrometre-scale histological features of bones with a pachyos-teosclerotic structure, as is typical in mammals welladapted to life in water as sirenians (see Ricqls andBuffr�nil 2001). The ribs are very thick and compact,without evidence of the medullary cavity (Fig. 5A). Themicrofractures, as well as the vascular canals and othercavities, are filled by pyrite or iron oxides and carbonates.In the peripheral region of the ribs, the palaeohistologicalstructures are highly altered and two concentric zones canbe differentiated, which reach approximately 0.7 to 1 mmin thickness (Fig. 5C–D): an external, irregular layerabout 300–350 mm thick, where the vascular canal mi-crostructure of the osteons is still visible with plane po-larised light, but not with polar crossed; and a complexalteration layer just under the external one, where thevascular canals of the osteons are hardly recognizableeven under plane polarised light; internally, there is abranching three-dimensional tunnel network, which isfilled by authigenic pyrite or iron oxides. This tunnelnetwork is characterized by relatively shallow (approxi-mately 0.7 mm) penetration into the massive mi-crostructure of the bone. The pyrite is produced across abig and spacing tunnel system, more or less perpendicularto the external bone surface (Fig. 5E).
The petrographical analyses of two rib samples madein an X-ray diffractometre indicate that the major mineralcomponent is francolite (carbonate fluorapatite [Ca5 (PO4,CO3)3 (F)]). In addition, the diffractograms show peakscorresponding to pyrite, iron oxides and calcite. Thecrystallinity index (CI) is about 0.27, and the densityreaches 2.9 (see Table 1). Thin sections analysed bycathodoluminiscence (CL) methods do not show evidenceof luminescence.
Interpretation
Generally, sirenians live in fluvial and shallow coastalenvironments where they graze the vegetation of the seafloor. In contrast, the sedimentological features of thePamplona Marl Formation in Uztarrotz and their pala-eontological content (microforaminifers, invertebrates)indicate a deep marine environment. Therefore, it is likelythat the Uztarrotz sirenian remains correspond to anademic organism (sensu Fern�ndez L�pez 1990, 2000).
Fig. 4 Epibiontic activity and bioerosion on sirenian ribs from theMiddle Eocene of Navarre. A small bivalves (Uz 1.23); B hemi-spheric borings (Uz 1.22); C annelid tubes (Ar 1.24); D bryozoancolonies (Ar 1.29). A–B from the Uztarrotz site, C–D from Ar-danatz. Note the eroded surface of the ribs (A–B) and the thinlongitudinal cracks (C). Scale lines 5 cm
470
Thus, the sirenian bones could have been shed to the opensea as a floating carcass.
The superficial damage of the fossil bones and theevidence of epibiontic activity suggest that the biostrati-nomic processes were relatively long. The large androunded borings observed superficially on the ribs areinterpreted either as pathological or, more probably, asthe result of bioerosion. According to their elongatedshape and relative size (diametre 5–10 mm along thelength), the branching three-dimensional network tunnelsfilled by pyrite and iron oxides may correspond to mi-crobial activity, perhaps due to fungi bioerosion (I. Sal-cedo, personal communication) or cyanobacteria activity(H.-U. Pfretzschner, personal communication). The ap-pearance of these microstructures is similar to that of the“Type I Weld tunnels” described by Trueman and Martill(2002). These authors interpret them as probable evi-dences of fungi bioturbation, at least in continental en-
vironments because Weld-type tunnels are made inocu-lating fresh bone with fungi.
The irregular fractures with rough surfaces present insome Uztarrotz rib fossils are interpreted as occurringduring the biostratinomic phase. Most of the breaks aresmooth transverse fractures without evidence of abrasion,and they are typical of the late fossildiagenetic processes(Shipman 1981).
With regard to the mineralogical changes, the franco-lite (carbonate fluorapatite) is a diagenetic mineral, whichcould replace the biogenic hydroxyapatite of the bonesduring the fossilization (Astibia et al. 1990; Jarvis 1992;Hubert et al. 1996; Elorza et al. 1999; L�cuyer et al. 2003;Trueman et al. 2003 and references).
The crystallinity index (CI) values of the Uztarrotzsamples are lower than those of fresh vertebrate bones(Sillen 1989), but they fall within the range of variation ofthe CI values of reptile fossils from the Late Jurassic ofUtah (Hubert et al. 1996) and of the Late Cretaceous of
Fig. 5 Microscopic characteris-tics of the sirenian fossil ribsfrom the Middle Eocene ofUztarrotz (A, C–E) and Ar-danatz (B, F–G), Navarre. A–B,Uz 1.24 and Ar 2.1, thin sec-tions of ribs showing the well-preserved palaeohistologicalstructures. Note on the rightside of A at least two peripher-ical layers of alteration (a1, a2);C Uz 1.25, alteration layers (a1,a2) with tunnel network struc-tures (t), probably made by mi-crobial activity; D detail of theprecedent image, showing themicrobial tunnel network filledsecondarily by pyrite and ironoxides, as are also the Haver-sian canals (h); E Uz 1.24,cancellous bone filled by fram-boidal pyrite (p) and secondaryfrancolite (f). Osteoblasts arealso filled by pyrite; F Ar 1.27,system of fractures (fr) with latesecondary francolite; G thesame view with crossed nicolsshowing the francolite zonation(z). The diagenetic front is ir-regular
471

the Iberian Peninsula (Elorza et al. 1999). This highcrystallinity of the Uztarrotz fossils is probably related todiagenetic factors.
Ardanatz fossils
Description
The surface of the rib fragments shows thin longitudinalcracks, which are perpendicular to the long axis of thebones. Most of the fractures are not eroded. There is noevidence of abrasion. However, a number of ribs exhibittraces of epibiontic activity, mainly bryozoan coloniesand annelid tubes (Fig. 4C–D).
The thin sections of the ribs show well-preservedhistological microstructures (Fig. 5E). As in the Uztarrotzsirenian ribs, there is internally a nearly peripheral, ap-proximately 1 mm thick, alteration layer exhibiting abranching three-dimensional network tunnels filled bypyrite and/or iron oxides. The Ardanatz samples showa larger number of fractures than those of Uztarrotz.Some of these fractures reach the external surface of thebone, where they show a replacement by secondaryfrancolite, with different light-coloured layers on eachside (Fig. 5F–G).
The X-ray diffraction analyses of four Ardanatz sam-ples also indicate that the fossil bone is mostly composedof francolite. The crystallinity index (CI) is between 0.25and 0.30, and the density reaches 2.91 as mean value(Table 1).
Interpretation
As noted above, the Ardanatz deposits correspond to ashallower environment than the Pamplona Marl Forma-tion in the Uztarrotz outcrop, and so they seem more inagreement with the sirenian way of life. The thin longi-tudinal cracks observed on the surface of the ribs areindicative of early erosion. This fact and the presence ofepibiontic activity suggest, as in the Uztarrotz fossils, thatthe bones were exposed on the sea floor for a while priorthe burial.
The peripheral complex alteration layer of the Ar-danatz fossils is similar to that described above for theUztarrotz ribs. The branching three-dimensional networktunnels correspond to the “Type I Weld tunnels” ofTrueman and Martill (2002), regarded by them as bio-erosion evidences of fungic attack. The crystallinity indexand density are high, comparable to the values describedfor Mesozoic reptile bones (Hubert et al. 1996; Elorza etal. 1999). As a singular feature, the compact bone mi-crostructure is affected by a dense system of fractures,which have allowed the passage of late diagenetic phos-phate-rich fluids, and laterally replacing the massive di-agenetic francolite by secondary francolite. The replace-ment extends from the fractures and progresses fromundulating advance front toward the internal part of the
bone. The internal structure is mostly preserved (as“ghosts”) in areas close to the fracture surface. This latefracture system, which affects the vascular canals filledby pyrite and iron oxides, could be owing to the peculiaroccurrence of the bones in marly levels located betweensandstones, and so to a different and fragile response ofthe strata to the compaction and subsequent tectonicstrain.
Geochemical composition
Uztarrotz and Ardanatz fossils
Description
The oxides CaO and P2O5 are the most abundant elements(mean value 70.4%); Fe2O3, Na2O and SiO2 are present inmuch lower concentrations (less than 4%). The Uztarrotzsediment (Uz S1 sample) is rich in CaO, SiO2 and Al2O3(Table 2). There are no significant differences betweenthe major elements of both Uztarrotz and Ardanatz fossilsamples, except that those of the former locality are richerin Mn (according to Mann-Whitney U’s test at confidentlevels p<0.05). The principal component analysis (PCA)suggests three PCs that explain 90% of the total variance(Fig. 6). The first component can separate clearly a po-rous sample (Ar 3.1, turtle plate), richer in Al, Si and Fe,relative to the remainder (sirenian samples), which con-tain mainly Ca, P and Na. The second component analysisseparates the samples from the two sites, having those ofUztarrotz more positive values of Mn than those of Ar-danatz (Fig. 6).The most abundant trace elements are Ba, Zn, Y and Ce(Table 2). There are no significant differences in theconcentrations of Cr, Lu, Tm, U, Y, and Yb between thesamples. In the Uztarrotz sediment, Ba, Zn and Zr are alsoabundant.
Fig. 6 Principal component analysis (PCA) of major elements ofthe sirenian fossils from the Middle Eocene of Uztarrotz and Ar-danatz (Navarre). The first principal component separates thesample Ar 3.1 (turtle plate) from other samples richer in Ca and P;the second principal component separates the Uztarrotz samples,richer in Mn, from the Ardanatz samples
472

The REE concentrations are very low and display asimilar pattern (Fig. 7). There is evidence of higher valuesof MREE and HREE than LREE in the samples.
Interpretation
Because of the mineralogical composition of the samples(francolite: Ca5 (PO4, CO3)3 (F)), the Ca and P are themost abundant elements. The high concentration of Fecould be owing to the presence of pyrite and iron oxidesfilling the fractures and microcavities of the bones. Thepyritization process needs enough concentration of Fe andsulfures and a low potential redox (reduction zones).These conditions are common in coastal deposits, wherethe pyritization is a usual process (Martill 1991). Fol-lowing Pfretzschner (2000, 2001a), the pyrite is one of themost common palaeohistological cavity-filling mineralsduring the early fossildiagenesis, especially in marineenvironments, where there is a high concentration in Fe.On the other hand, the presence of pyrite and associatediron oxides in the tubular-like cavities could correspond toa later diagenetic phase, as pointed out by Pfretzschner(2001b).
The higher concentration of Mn in the Uztarrotz fossilsrelative to those of the Ardanatz site could be due to adeeper burial environment under reducing conditions.
As apatite has a very strong affinity for a wide range oftrace elements (Trueman 1999), the vertebrate bonesshow generally REE enrichment from the microenviron-
ment fluids during the early diagenesis, being the con-centrations of REEs in fossil apatite higher than in thehost rock. On the contrary, the studied fossil sirenianbones and sediment samples have a similar sum of REEs(Table 2). This could be due to the dense compactstructure of the sirenian bones relative to other verte-brates.
In Ardanatz and Uztarrotz, the REE distribution of thesirenian fossils is regarded as quite homogeneous (Fig. 7),displaying a flat REE pattern in both sites that is known as‘flat-shaped’ (L�cuyer et al. 2003). This suggests that theremains have suffered early diagenetic processes: the ratiovalues La/Sm versus La/Yb in both sites are in agreementwith a REE adsorption mechanism (Reynard et al. 1999).
The Ardanatz remains were deposited in shallow ma-rine deposits; they do not show any evidence of abrasion.These remains are here regarded as belonging to an au-tochthonous assemblage, probable from demic organisms(sensu Fern�ndez L�pez 1990, 2000). The similar REEtrends and crystallinity index (CI) observed in the sireniansamples suggest that the diagenetic process was probablyuniform and unique. As far as known, there is no evidenceof reelaboration for the Ardanatz fossils.
Conclusions
Recently discovered sirenian fossils from the PamplonaBasin (Navarre) consist of vertebrae and ribs from twosuccessive, but sedimentologically different, Middle Eo-
Fig. 7 Rare earth elements (REE) pattern of the sirenian fossilsfrom the Middle Eocene of Uztarrotz and Ardanatz (Navarre)compared with a turtle plate (Ar 3.1) from Ardanatz (Navarre) andmarine Mesozoic fossil vertebrate bones from the Upper Triassic ofEngland: A1, A4, A5, A8 and A13 correspond to Pachystropheus(reptile) bones and AS7 from an indeterminate vertebrate, both at
Aust Cliff; W7 from an indeterminate vertebrate at WestburyGarden Cliff (all the samples correspond to dense and/or cancellouscompact bone but the rib W7; after Trueman et al. 2003). All theREE results have been normalized to PAAS (Post-Archean Aus-tralian Shales) (Taylor and McLennan 1985)
473

cene (Bartonian) lithostratigraphic units. The Uztarrotzfossils were collected from the deep-marine PamplonaMarl Formation, and they are interpreted as coming froman ademic organism (i.e., transported over a distance fromthe original habitat). The Ardanatz fossils accumulated ina shallow-water, semi-closed muddy deltaic bay coveredby extensive sea-grass meadows and affected by episodic,flood-related hyperpycnal jets. Therefore, the Ardanatzsirenian remains, which are also thought to belong to asingle individual, are regarded as nearly autochthonous,probably deposited in their natural habitat or scarcelytransported.
The ribs are thick and show a pachyosteoscleroticstructure, as is typical in sirenians. The superficial dam-age of the bones from the two Navarrese localities and theevidence of epibiontic activity suggest relatively longbiostratinomic phases. The Uztarrotz remains are moreeroded than those of Ardanatz, perhaps because they wereexposed in the sea floor for a longer time prior to theburial as result of a lower sedimentation rate in deepmarine environments.
The fossil-bone mineral is well-crystallized francolite(carbonate fluorapatite). The palaeohistologic structuresare well preserved except in the peripheral region, wherethere is an altered layer filled by pyrite and oxides thatmay correspond to microbial bioturbation.
Both Uztarrotz and Ardanatz sites exhibit a ‘flat-shaped’ distribution of rare earth elements (REE) whichsuggests that the remains suffered early diagenetic pro-cesses. The similar REE trend indicates that there is noevidence of mixing reworked bones. Although derivedfrom different sedimentary environments, the sirenianfossils from Uztarrotz and Ardanatz have similar sum ofREE concentrations relative to the host rock, higher thanin recent bones but lower than in most other vertebratefossils. This low concentration of REEs could be relatedto the dense compact structure of pachyosteoscleroticsirenian bones.
Acknowledgements We are grateful to Mr. Jos� Luis Belzunegi forcollecting and supplying the Ardanatz fossils and to Mr GustavoGaspar for lending to study the Uztarrotz remains and showing usthe outcrop. Special thanks to Dr Alejandro Cearreta (UPV/EHU,Dpto. Estratigraf�a y Paleontolog�a) for his interpretation of mi-cropalaeontological data, Dr Isabel Salcedo (UPV/EHU, Dpto.Biolog�a Vegetal y Ecolog�a, Laboratorio de Bot�nica) for heruseful comments about microbial activity, and to the referees Prof.Dr. Andr� Freiwald (Universit�t Erlangen) and Prof. Dr. H.-U.Pfretzschner (Universit�t T�bingen) for their advice on the manu-script. We gratefully acknowledge the financial support providedby the Ministerio de Ciencia y Tecnolog�a of Spain (BOS2000-1369 and BTE2002-03806) and by the Universidad del Pa�s Vasco/EHU (9/UPV 00121.310-15303/2003 and 14455/2002). The re-search work of the third author (X.P.S.) was supported by thePrograma Ram�n y Cajal of the Ministerio de Ciencia y Tec-nolog�a, and that of the fifth author (A.B.) by a Ph.D. grant of theUniversidad del Pa�s Vasco/EHU.
References
Astibia H, Herrero JM and Elorza JJ (1990) An example of pet-rographic microscopy and X-ray diffraction techniques as toolsin vertebrate taphonomic analysis of some Spanish fossil ma-terial. In: Fernandez S (ed) Comunicaciones Reuni�n Ta-fonom�a y Fosilizacion, Univ Complutense Madrid, 27–39
Astibia H, Murelaga X, Payros A, Pereda Suberbiola X, Tosquella J(1999) Tortugas y sirenios f�siles en el Eoceno marino deNavarra y Cuenca de Jaca. Geogaceta 25:15–18
Berggren WA, Kent DV, Swisher CC III, Aubry MP (1995) Arevised Cenozoic geochronology and chronostratigraphy. In:Berggren WA, Kent DV, Aubry MP, Hardenbol J (eds)Geochronology, time scales and global stratigraphic correla-tions. SEPM Soc Sedim Geol Spec Publ 54:129–212
Blumenthal NC, Betts F, Posner AS (1975) Effect of carbonate andbiological macromolecules on formation and properties of hy-droxyapatite. Calcified Tissue Res 18:81–90
Calzada S, Astibia H (1996) Sobre Plicatula pamplonensis Carez:revisi�n. Bull Soc Hist Nat Toulouse 132:65–69
Cendon DI, Ayora C, Pueyo JJ (1998) The origin of barren bodiesin the Subiza potash deposit, Navarra, Spain: implications forsylvite formation. J Sediment Res 68:43–52
Clarkson ENK (1992) Invertebrate palaeontology and evolution.Chapman and Hall, London, 434 pp
Elorza J, Astibia H, Murelaga X, Pereda Suberbiola X (1999)Francolite as a diagenetic mineral in dinosaurs and other UpperCretaceous reptile fossil bones (La�o, Iberian Peninsula): mi-crostructural, petrological and geochemical features. Cret Res20:169–187
Fern�ndez L�pez S (1990) El significado de la autocton�a/alocton�atafon�mica. In: Fernandez L�pez S (ed) Comunicaciones Re-uni�n Tafonom�a y Fosilizacion, Univ. Complutense Madrid,115–124
Fern�ndez L�pez S (2000) Temas de Tafonom�a. Dept. Paleon-tolog�a, Univ. Complutense Madrid, 167 pp. Internet site: http://www.ucm.es/info/paleo/personal/sixto.htm
Francillon-Vieillot H, Buffr�nil V de, Castanet J, G�raudie J, Me-unier FJ, Sire JY, Zylberger L, Ricqls A de (1990) Mi-crostructure and mineralization of vertebrate skeletal tissues.In: Carter JG (ed) Skeletal biomineralization: patterns, pro-cesses and evolutionary trends. 1:471–671, Van NostrandReinhold, New York
Galobart A, Maroto J, Ros J (1992) Troballa d’un sireni de l’eoc ala vall del Ll�mena. Quaderns C.E.C.B. Banyoles 1990–91:85–94
G�mez G (1998) Dos sirenios del Eoceno en la formaci�n Belsu�-Atar�s. Serrablo 110
Haines PW, Jago JB, Gum JC (2001) Turbidite deposition in theCambrian Kanmantoo Group, South Australia. Aust J Earth Sci48:465–478
Hubert JF, Panish PT, Chure DJ, Prostak KS (1996) Chemistry,microstructure, petrology, and diagenetic model of Jurassicdinosaur bones, Dinosaur National Monument, Utah. J Sedi-ment Res 66:531–547
Jarvis I (1992) Sedimentology, geochemistry and origin of phos-phatic chalks: the Upper Cretaceous deposits of NW Europe.Sedimentology 39:55–97
L�cuyer C, Bogey C, Garcia J-P, Grandjean P, Barrat J-A, FloquetM, Bardet N, Pereda Suberbiola X (2003) Stable isotopecomposition and rare earth element content of vertebrate re-mains from the Late Cretaceous of northern Spain (La�o): didthe environmental record survive? Palaeogeogr PalaeoclimatolPalaeoecol 193:457–471 [and Corrigendum, PalaeogeogrPalaeoclimatol Palaeoecol 196:427–428]
Mangin J (1959–60) Le nummulitique sud-pyr�n�en a l’ouest del’Aragon. Pirineos 51–58:1–656
Martill DM (1991) Bones as stones: the contribution of vertebrateremains to the lithologic record. In: Donovan SK (ed) Theprocess of fossilization. Belhaven Press, London, 270 pp
474

Martinsen OJ (1990) Fluvial, inertia-dominated deltaic depositionin the Namurian (Carboniferous) of Northern England. Sedi-mentology 37:1099–1114
McLeod P, Carey S, Sparks SJ (1999) Behaviour of particle-ladenflows into the ocean: experimental simulation and geologicalimplications. Sedimentology 46:523–536
Mellere D, Plink-Bj�rklund P, Steel R (2002) Anatomy of shelfdeltas at the edge of a prograding Eocene shelf margin, Spits-bergen. Sedimentology 49:1181–1206
Morellet L, Morellet J (1945) Faune des sables Nummulitesvariolarius de Barisseuse, prs de Saint-Vast-les-Mello (Oise),et remarques pal�ontologiques. Bull Soc G�ol Fr Ser 5, 15:337–356
Morton A (2003) A Collection of Eocene and Oligocene fossils.Internet site: http://www.dmap.co.uk/fossils/
Mulder T, Syvitski JPM (1995) Turbidity currents generated atriver mouths during exceptional discharges to the world oceans.J Geol 103:285–299
Mulder T, Syvitsky JPM, Migeon S, Faugres JC, Savoye B (2003)Marine hyperpycnal flows: initiation, behavior and related de-posits. A review. Mar Petrol Geol 20:861–882
Mutti E (1984) The Hecho Eocene submarine fan system, south-central Pyrenees, Spain. Geo-Mar Lett 3:199–202
Mutti E, Davoli G, Tinterri R, Zavala C (1996) The importance offluvio-deltaic systems dominated by catastrophic flooding intectonically active basins. Mem Sci Geol 48:233–291
Mutti E, Tinterri R, Benevelli G, Di Biase D, Cavanna G (2003)Deltaic, mixed and turbidite sedimentation of ancient forelandbasin. Mar Petrol Geol 20:733–755
Mutti E, Tinterri R, Di Biase D, Fava L, Mavilla N, Angella S,Calabrese L (2000) Delta-front facies associations of ancientflood-dominated fluvio-deltaic systems. Rev Soc Geol Espa�a13:165–190
Nemec W (1995) The dynamics of deltaic suspension plumes. In:Oti MN, Posta G (eds) Geology of deltas. AA Balkema, Rot-terdam, 31–93
Patzkowsky M, Bezusko-Layou K (1999) The paleobiology data-base (PBDB), collection 3494, Barisseuse, France. Internet site:http://www.paleodb.org
Payros A (1997) El Eoceno de la Cuenca de Pamplona: estrati-graf�a, facies y evoluci�n paleogeogr�fica. PhD Thesis, Uni-versidad del Pa�s Vasco/EHU, Bilbao (unpublished)
Payros A, Astibia H, Cearreta A, Pereda Suberbiola X, Murelaga X,Badiola A (2000) The Upper Eocene South Pyrenean coastaldeposits (Liedena Sandstone, Navarre): sedimentary facies,benthic foraminifera and avian ichnology. Facies 42:107–132
Payros A, Pujalte V, Orue-Etxebarria X, Baceta JI (1997) Un sis-tema turbid�tico de tipo “channel-levee” en la Cuenca dePamplona: implicaciones tect�nicas y paleogeogr�ficas. Ge-ogaceta 22:145–148
Pfretzschner H-U (2000) Pyrite formation in Pleistocene bones—acase of very early mineral formation during diagenesis. NeuesJahrb Geol Pal�ont Abh 217:143–160
Pfretzschner H-U (2001a) Pyrite in fossil bone. Neues Jahrb GeolPal�ont Abh 220:1–23
Pfretzschner H-U (2001b) Iron oxides in fossil bone. Neues JahrbGeol Pal�ont Abh 220:417–429
Pilleri G, Biosca J, Via L (1989) The Tertiary Sirenia of Catalonia.Brain Anatomy Institute, Univ Berne, Ostermundingen, 98 pp
Plink-Bj�rklund P, Steel RJ (2004) Initiation of turbidity currents:outcrop evidence for Eocene hyperpycnal flow turbidites.Sediment Geol 165:29–52
Puigdefabregas C (1975) La sedimentaci�n mol�sica en la Cuencade Jaca. Pirineos 140:1–188
Pujalte V, Baceta JI, Payros A (2002) Tertiary, Western Pyreneesand Basque-Cantabrian Region. In: Gibbons W, Moreno MT(eds) The geology of Spain. Geol Soc Lond, pp 293–301
Reynard B, L�cuyer C, Grandjean P (1999) Crystal-chemicalcontrols on rare-earth element concentrations in fossil biogenicapatites and implications for paleoenvironmental reconstruc-tions. Chem Geol 155:233–241
Ricqls A de, Buffr�nil V de (2001) Bone histology, heterochroniesand the return of Tetrapods to life in water: where are we? In:Mazin JM, Buffr�nil V de (eds) Secondary adaptations ofTetrapods to life in water. Verlag Dr. Friedrich Pfeil, Munich,pp 289–310
Roman P, Gutierrez-Zorrilla JM (1985) A quick method for de-termining the density of single crystals. J Chem Education63:167
Ruiz de Gaona M (1947) El Bartoniense de la Cuenca de Pamplona.Notas y Com Inst Geol Min Espa�a 17:155–165
Ruiz de Gaona M, Colom G (1950) Estudio de las sinecias de losforamin�feros eoc�nicos de la vertiente meridional de Pirineos(Catalu�a y Vizcaya). Estud Geol Inst Lucas Mallada 12:293–434
Sagne C (2001a) Halitherium taulannense, nouveau sir�nien(Sirenia, Mammalia) de l’�ocene sup�rieur provenant du do-maine Nord-T�thysien (Alpes-de-Haute-Provence, France). CRAcad Sci Paris, Sc Terre et plantes 333:471–476
Sagne C (2001b) La diversification des sir�niens l’Eocne(Sirenia, Mammalia): �tude morphologique et analyse phy-log�n�tique du sir�nien de Taulanne, Halitherium taulannense.PhD thesis, Mus�um National d’Histoire Naturelle, Paris, 292pp
Serra-Kiel J, Hottinger J, Caus, E, Drobne, K, Ferr ndez C., JauhriAK, Less G, Pavlovec R, Pignatti J, Sams� J.M, Schaub H,Sirel E, Strougo A, Tambareau Y, Tosquella J, Zakrevskaya E(1998) Larger foraminiferal biostratigraphy of the TethyanPaleocene and Eocene. Bull Soc G�ol Fr 169:281–299
Shipman P (1981) Life history of a fossil. An introduction totaphonomy. Harvard University Press, Cambridge, 222 pp
Sillen A (1989) Diagenesis of the inorganic phase of compact bone.In: Price TD (ed) The chemistry of prehistoric human bone.Cambridge Univ Press, Cambridge, pp 211–229
Sparks SJ, Bonnecaze RT, Huppert HE, Lister JR, Hallworth MA,Mader H, Phillips J (1993) Sediment-laden gravity currentswith reversing buoyancy. Earth Planet Sci Lett 114:243–257
Sprinkle J, Kier PM (1987) Phylum Echinodermata. In: BoardmanRS, Cheetham AH, Rowell AJ (eds) Fossil invertebrates.Blackwell, Palo Alto, pp 550–611
Taylor SR, McLennan SM (1985) The continental crust: its com-position and evolution. Blackwell, Oxford, 312 pp
Trueman CN (1999) Rare earth element geochemistry andtaphonomy of terrestrial vertebrate assemblages. Palaios14:555–568
Trueman CN, Benton MJ, Palmer MR (2003) Geochemicaltaphonomy of shallow marine vertebrate assemblages. Palaeo-geogr Palaeoclimatol Palaeoecol 197:151–169
Trueman CN, Martill DM (2002) The long-term survival of bone:the role of bioerosion. Archaeometry 44:371–3
West I (2002) Barton and Highcliffe, Eocene strata: geology of thesouth coast of England. Internet site: http://www.soton.ac.uk/~imw/barton.htm (School of Ocean and Earth Sciences,Southampton University, U.K. Version U.04.02.02)
Wright LD (1977) Sediment transport and deposition at rivermouths: a synthesis. Geol Soc Am Bull 88:857–868
475
Related Documents











