Seasonal cambial activity and tree-ring formation of Pinus merkusii and Pinus kesiya in Northern Thailand in dependence on climate Nathsuda Pumijumnong * , Toonsak Wanyaphet Faculty of Environment and Resource Studies, Mahidol University, Nakhon Pathom 73170, Thailand Received 28 January 2005; received in revised form 3 January 2006; accepted 30 January 2006 Abstract This study is aimed at characterizing the cambial dynamics and its dependence on climate of two pine species native to Thailand, Pinus merkusii and Pinus kesiya and at describing their climatic response over 148 years. The samples for cambial activity analysis were taken monthly from March 2000 to February 2001, and the cambial activity was determined by counting the number of undifferentiated cell layers between mature xylem and phloem in transverse sections. Statistical analysis was done using Pearson’s correlation. For the dendroclimatological analysis, samples were collected from the same sites in March 2004, and dendroclimatological standard techniques were applied. The results indicated that soil moisture influenced the cambial activity of P. merkusii and P. kesiya, however that rainfall and temperature had no significant effect on the cambial activity of both species. The response function described the relationship between tree-ring widths indices and monthly rainfall and temperature and revealed that the growth of P. merkusii at Hung Boung depended positively from rainfall in May. P. merkusii at Bao Kaew, however, had a positive correlation with rainfall from previous November to current July, whereas temperature in the preceding autumn and winter should be above-average and in the current spring and summer should be below-average. P. kesiya at Nong Kra Ting showed a slightly positive correlation with rainfall from February to June, but a strong dependence on rainfall in September. It can be concluded that the study of cambial activity could support our understanding of intra-annual variations and duration of tree-ring development, whereas the response function could help explain the average response of tree-growth to climate. # 2006 Elsevier B.V. All rights reserved. Keywords: Dendrochronology; Tropical pines; Thailand; Cambial activity; Tree-ring formation 1. Introduction Few subtropical tree species in Southeast Asia have been successfully used for dendrochronological research, in particular teak (Tectona grandis) and pine (Pinus spp.). Various approaches have been tried, e.g. dendrochronological networking and dendroclimatology with teak by Pumijumnong et al. (1995a). Moreover, Buckley et al. (1995) established a pine chronology in Northeast Thailand. Later, D’Arrigo et al. (1997) presented progress in dendroclimatic studies of mountain pine in Northern Thailand. All these studies provided information on the inter-annual but not on the intra-annual variation of growth and on the duration of wood formation. Likewise, whereas research has been undertaken to understand the reaction of teak and pine to the monsoon, the basic mechanisms of tree physiology and cell development recorded in the sequence of ring widths are not well understood and need further investigation. In Thailand, few studies have been published on the seasonal formation of xylem, e.g. by Pumijumnong et al. (1995b) for teak. But there is still a lack of information on cambial activity, on its increase at the beginning of the growth season and its decrease towards the dormant season, along with temporal variations of these processes. None of these studies compared the average climatic response of tree-ring width series with the seasonal cambial activity, especially in subtropical pine species. The present study was done for two pine species native in northern Thailand, Pinus merkusii (two needle pine) and Pinus kesiya (three needle pine), which are considered the most important ones with respect to reforestation of watershed areas and forest conservation as well as to their potential for dendroclimatology. It is focused on their intra-annual variation www.elsevier.com/locate/foreco Forest Ecology and Management 226 (2006) 279–289 * Corresponding author. Tel.: +66 2 4415000x228; fax: +66 2 4419509/4419510. E-mail address: [email protected] (N. Pumijumnong). 0378-1127/$ – see front matter # 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.foreco.2006.01.040

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Seasonal cambial activity and tree-ring formation of Pinus merkusii
and Pinus kesiya in Northern Thailand in dependence on climate
Nathsuda Pumijumnong *, Toonsak Wanyaphet
Faculty of Environment and Resource Studies, Mahidol University, Nakhon Pathom 73170, Thailand
Received 28 January 2005; received in revised form 3 January 2006; accepted 30 January 2006
Abstract
This study is aimed at characterizing the cambial dynamics and its dependence on climate of two pine species native to Thailand, Pinus merkusii
and Pinus kesiya and at describing their climatic response over 148 years. The samples for cambial activity analysis were taken monthly from
March 2000 to February 2001, and the cambial activity was determined by counting the number of undifferentiated cell layers between mature
xylem and phloem in transverse sections. Statistical analysis was done using Pearson’s correlation. For the dendroclimatological analysis, samples
were collected from the same sites in March 2004, and dendroclimatological standard techniques were applied.
The results indicated that soil moisture influenced the cambial activity of P. merkusii and P. kesiya, however that rainfall and temperature had no
significant effect on the cambial activity of both species.
The response function described the relationship between tree-ring widths indices and monthly rainfall and temperature and revealed that the
growth of P. merkusii at Hung Boung depended positively from rainfall in May. P. merkusii at Bao Kaew, however, had a positive correlation with
rainfall from previous November to current July, whereas temperature in the preceding autumn and winter should be above-average and in the
current spring and summer should be below-average. P. kesiya at Nong Kra Ting showed a slightly positive correlation with rainfall from February
to June, but a strong dependence on rainfall in September.
It can be concluded that the study of cambial activity could support our understanding of intra-annual variations and duration of tree-ring
development, whereas the response function could help explain the average response of tree-growth to climate.
# 2006 Elsevier B.V. All rights reserved.
Keywords: Dendrochronology; Tropical pines; Thailand; Cambial activity; Tree-ring formation
www.elsevier.com/locate/foreco
Forest Ecology and Management 226 (2006) 279–289
1. Introduction
Few subtropical tree species in Southeast Asia have been
successfully used for dendrochronological research, in
particular teak (Tectona grandis) and pine (Pinus spp.).
Various approaches have been tried, e.g. dendrochronological
networking and dendroclimatology with teak by Pumijumnong
et al. (1995a). Moreover, Buckley et al. (1995) established a
pine chronology in Northeast Thailand. Later, D’Arrigo et al.
(1997) presented progress in dendroclimatic studies of
mountain pine in Northern Thailand. All these studies
provided information on the inter-annual but not on the
intra-annual variation of growth and on the duration of wood
formation. Likewise, whereas research has been undertaken
* Corresponding author. Tel.: +66 2 4415000x228;
fax: +66 2 4419509/4419510.
E-mail address: [email protected] (N. Pumijumnong).
0378-1127/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.foreco.2006.01.040
to understand the reaction of teak and pine to the monsoon,
the basic mechanisms of tree physiology and cell development
recorded in the sequence of ring widths are not well understood
and need further investigation.
In Thailand, few studies have been published on the
seasonal formation of xylem, e.g. by Pumijumnong et al.
(1995b) for teak. But there is still a lack of information on
cambial activity, on its increase at the beginning of the growth
season and its decrease towards the dormant season, along
with temporal variations of these processes. None of these
studies compared the average climatic response of tree-ring
width series with the seasonal cambial activity, especially in
subtropical pine species.
The present study was done for two pine species native in
northern Thailand, Pinus merkusii (two needle pine) and Pinus
kesiya (three needle pine), which are considered the most
important ones with respect to reforestation of watershed areas
and forest conservation as well as to their potential for
dendroclimatology. It is focused on their intra-annual variation

N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289280
in monthly growth during one growing season as well as on
their long-term average climate/growth response. We had two
main objectives, first, to compare the variation of cambial
activity between the two pine species with trees of the same
age, and among trees of the same species with different ages,
and second, to describe the climatic response of the pine by
means of dendrochronology.
2. Materials and methods
2.1. Study area
In order to obtain meaningful climate-related results, the
sites should be as devoid of human activities as possible. Two
study sites were selected for cambial analysis. The Bao Kaew
(BK) Silviculture Research Station is located at Bao Loung,
Hod District, Chiang Mai Province, at latitude 18.16N,
longitude 98.38E and 1036 m above sea level. Here, P.
merkusii is growing.
The Nong Kra Ting (NT) Pine Production Area is located
in the Om-Koi District, Chiang Mai Province, at latitude
17.98N, longitude 98.38E and 1070 m above sea level. Both
sites are about 36 km apart from each other. Here, P. kesiya
is growing.
The original forest at both sites is a mixed dry dipterocarp
forest where pine is the dominant species, commonly in
association with native hardwoods, such as Shorea obtusa,
Dipterocarpus tuberculatus, D. obtusifolius, Phallanthus
emblica, etc. The undergrowth consists of Imperata cylindrical
and Cycas siamensis.
The site for dendrochronological analysis was Hung Boung
(HB), which is located at Bao Loung, Hod District, Chiang Mai
Province, at latitude 18.16N, longitude 98.42E and 1025 m
above sea level. Here, P. merkusii is growing. Hung Boung is
very near the Bao Kaew site, about 4 km down the road, and
about 40 km from the Nong Kra Ting site (Fig. 1).
The soil at HB and BK is red-yellow podzolic and gray
podzolic of medium depth with a pH between 4.5 and 5.5 and a
good drainage. As for the NT site, the main soil is a reddish-
brown laterite with a depth between 100 and 150 cm; its texture
is sandy loam or loam over clay and loam to clay with a pH
between 6.0 and 7.0.
In northern Thailand the southwest monsoon brings
rain with a monthly average of 167 mm from April/May to
October; from November to March it is dry with only 23 mm of
rainfall per month. The average annual rainfall at the three
study sites is 1000–1400 mm. The annual average minimum
and maximum temperatures are 4 and 40 8C, respectively
(Fig. 2).
3. Methods
3.1. Cambial activity
The age of each tree was determined by tree-ring counting,
and the trees were divided into three age classes: 30–60
(young), 60–90 (middle age) and >90 (old) years old.
3.2. Field sample collection
Samples were taken monthly from the same trees (three
middle aged and old aged, each, in BK and four young aged and
middle aged, each, in NT) from March 2000 to February 2001.
The specimens include bark, cambial zone and some part of
wood tissue of the main trunk at breast height (1.3 m). Two
specimens were taken from the north- and south-facing sides of
each tree. The sample size was about 2 cm � 2 cm with a depth
of 2 cm using a chisel. They were fixed with 3% glutaraldehyde
directly at the study site.
In the lab the specimens were thoroughly washed with water
to remove any remaining chemicals and then infiltrated and
embedded with PEG 4000. Cross sections of 20–30 mm
thickness were cut using a sliding microtome. Safranine and
fast green were used for staining.
The cambial activity of the pine trees was determined
by counting the number of undifferentiated cambial layers
having narrow rectangular cells between the mature xylem
and phloem under the light microscope. Earlywood and
latewood formation was observed by anatomical character-
istics.
The soil moisture content was one of the parameters used in
this study. Soil samples were taken from each site at a depth of
30 cm. Their moisture content was determined by weighing the
wet soil samples, then drying them at 100–110 8C and
reweighing. The soil moisture content was measured season-
ally. The results were expressed as percentage of moisture
based on the dry weight of the soil.
3.3. Sample collection for tree-ring analysis
In March 2004, altogether 37 pine trees from Hung Boung
(7 trees, 28 cores), Bao Kaew (10 trees, 30 cores) and Nong
Kra Ting (20 trees, 80 cores) were cored at breast height
(1.3 m) for age determination and tree-ring width measure-
ment. The Bao Kaew and Nong Kra Ting sites were the same
as those for cambial activity analysis. All cores were glued
to wooden core mounts and surfaced with up to 400 grit
sand paper, in order to render the tree-ring boundaries more
clearly visible.
3.4. Meteorological data
Average monthly temperature and total monthly rainfall data
were available from both sites, Hung Bong and Nong Kra Ting.
Since there is no meteorological station at Bao Kaew, we used
the meteorological data from Hung Boung.
Monthly total rainfall from Om-Koi meteorological station
(about 40 km from Houng Boung and Bo Kaew and 30 km from
Nong Kra Ting) for the period 1977–2003 was used for the
dendroclimatological study. This period of 26 years, while
rather short for statistical comparison, was all that was available
for this study. Since the Om-Koi meteorological station does
not record temperatures, we used temperature data from Chiang
Mai, about 100 km east of the study area, available for the
period 1951–2003 (52 years).
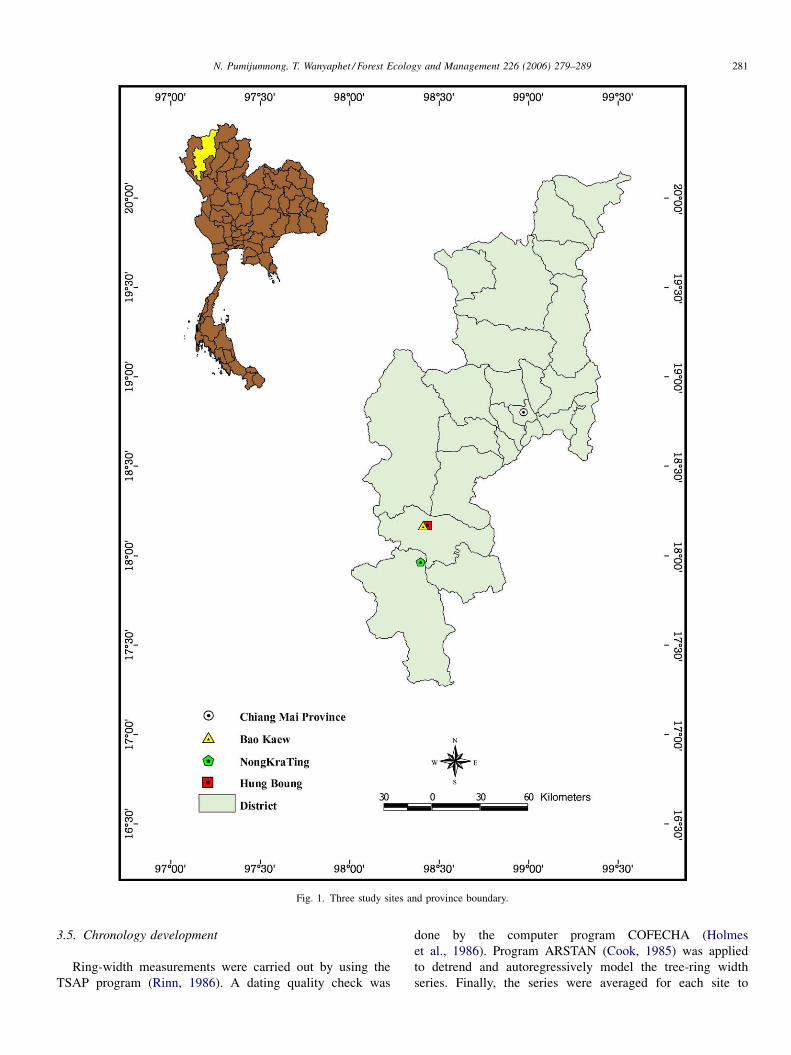
N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289 281
Fig. 1. Three study sites and province boundary.
3.5. Chronology development
Ring-width measurements were carried out by using the
TSAP program (Rinn, 1986). A dating quality check was
done by the computer program COFECHA (Holmes
et al., 1986). Program ARSTAN (Cook, 1985) was applied
to detrend and autoregressively model the tree-ring width
series. Finally, the series were averaged for each site to
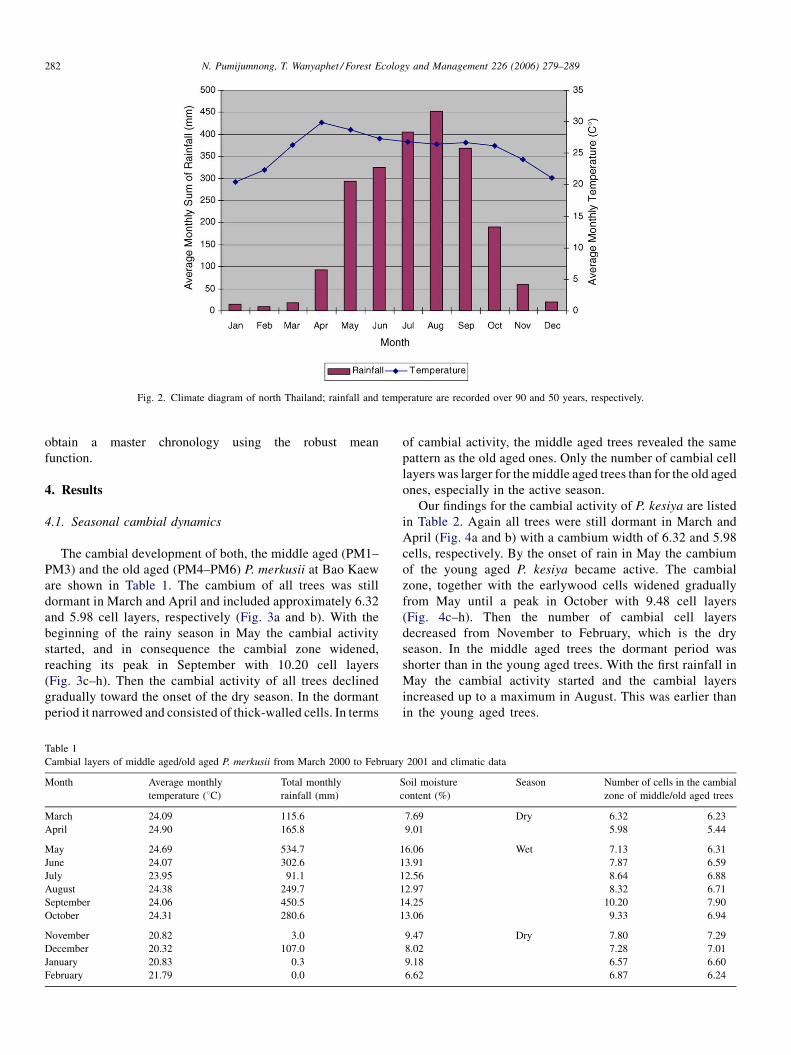
N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289282
Fig. 2. Climate diagram of north Thailand; rainfall and temperature are recorded over 90 and 50 years, respectively.
obtain a master chronology using the robust mean
function.
4. Results
4.1. Seasonal cambial dynamics
The cambial development of both, the middle aged (PM1–
PM3) and the old aged (PM4–PM6) P. merkusii at Bao Kaew
are shown in Table 1. The cambium of all trees was still
dormant in March and April and included approximately 6.32
and 5.98 cell layers, respectively (Fig. 3a and b). With the
beginning of the rainy season in May the cambial activity
started, and in consequence the cambial zone widened,
reaching its peak in September with 10.20 cell layers
(Fig. 3c–h). Then the cambial activity of all trees declined
gradually toward the onset of the dry season. In the dormant
period it narrowed and consisted of thick-walled cells. In terms
Table 1
Cambial layers of middle aged/old aged P. merkusii from March 2000 to February
Month Average monthly
temperature (8C)
Total monthly
rainfall (mm)
March 24.09 115.6
April 24.90 165.8
May 24.69 534.7
June 24.07 302.6
July 23.95 91.1
August 24.38 249.7
September 24.06 450.5
October 24.31 280.6
November 20.82 3.0
December 20.32 107.0
January 20.83 0.3
February 21.79 0.0
of cambial activity, the middle aged trees revealed the same
pattern as the old aged ones. Only the number of cambial cell
layers was larger for the middle aged trees than for the old aged
ones, especially in the active season.
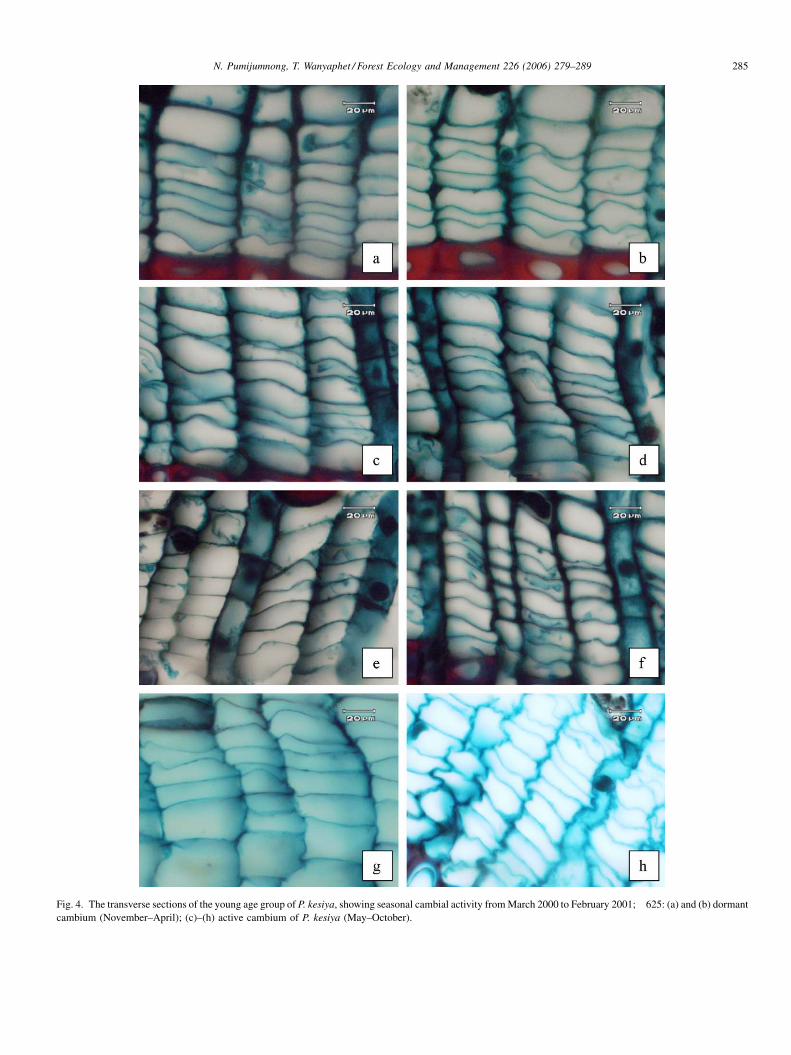
Our findings for the cambial activity of P. kesiya are listed
in Table 2. Again all trees were still dormant in March and
April (Fig. 4a and b) with a cambium width of 6.32 and 5.98
cells, respectively. By the onset of rain in May the cambium
of the young aged P. kesiya became active. The cambial
zone, together with the earlywood cells widened gradually
from May until a peak in October with 9.48 cell layers
(Fig. 4c–h). Then the number of cambial cell layers
decreased from November to February, which is the dry
season. In the middle aged trees the dormant period was
shorter than in the young aged trees. With the first rainfall in
May the cambial activity started and the cambial layers
increased up to a maximum in August. This was earlier than
in the young aged trees.
2001 and climatic data
Soil moisture
content (%)
Season Number of cells in the cambial
zone of middle/old aged trees
7.69 Dry 6.32 6.23
9.01 5.98 5.44
16.06 Wet 7.13 6.31
13.91 7.87 6.59
12.56 8.64 6.88
12.97 8.32 6.71
14.25 10.20 7.90
13.06 9.33 6.94
9.47 Dry 7.80 7.29
8.02 7.28 7.01
9.18 6.57 6.60
6.62 6.87 6.24
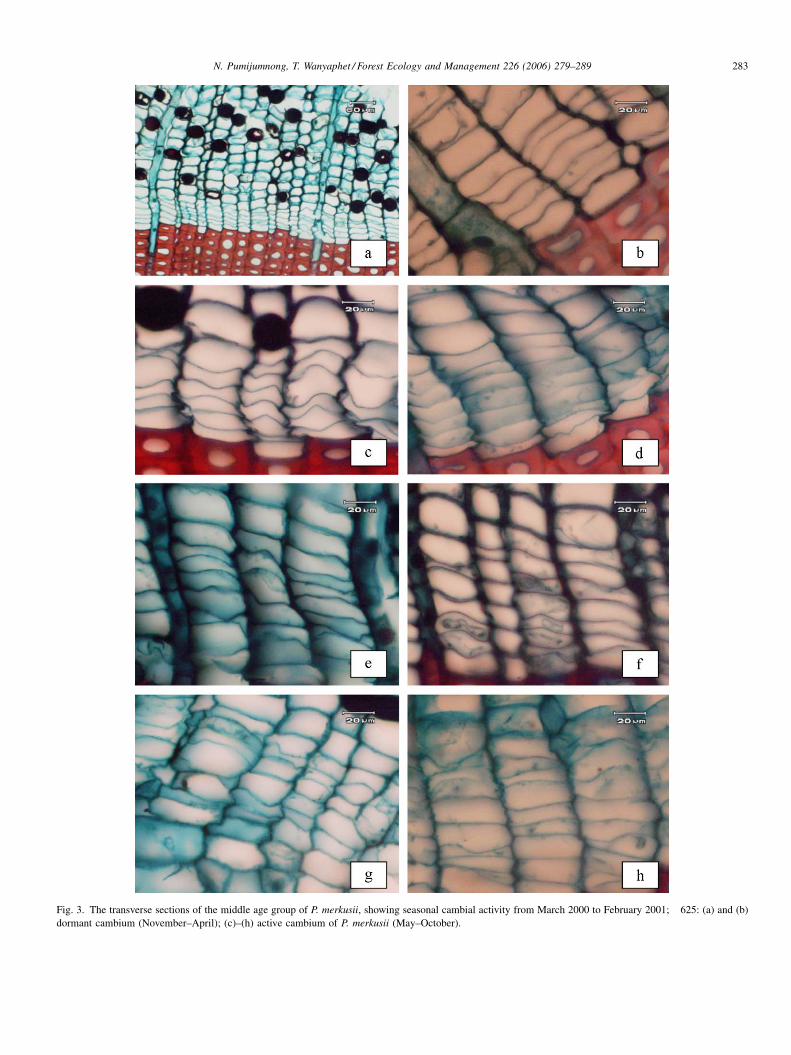
N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289 283
Fig. 3. The transverse sections of the middle age group of P. merkusii, showing seasonal cambial activity from March 2000 to February 2001; �625: (a) and (b)
dormant cambium (November–April); (c)–(h) active cambium of P. merkusii (May–October).
N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289284
Table 2
Cambial layers of young aged/middle aged P. kesiya from March 2000 to February 2001 and climatic data
Month Average monthly
temperature (8C)
Total monthly
rainfall (mm)
Soil moisture
content (%)
Season Number of cells in the cambial
zone of young/old aged trees
March 21.90 95.88 9.88 Dry 6.32 5.40
April 24.60 110.84 12.44 5.98 5.38
May 24.50 287.8 20.75 Wet 7.65 6.28
June 24.70 141.44 19.45 7.88 6.56
July 24.60 178.84 18.58 8.51 7.26
August 24.90 120.36 16.69 8.45 7.82
September 25.10 148.24 19.28 8.79 7.76
October 24.15 217.60 19.98 9.48 7.23
November 19.00 0.0 12.64 Dry 7.51 7.09
December 18.55 31.96 9.26 6.81 6.25
January 18.45 0.0 7.25 6.54 5.75
February 19.55 0.0 9.05 7.16 5.51
To summarize, the trees of both pine species and of all age
classes were dormant until March and April. In all cases, it was
May rainfall that reactivated cambial divisions.
4.2. The relationship between cambial activity and climatic
variables
Table 3 illustrates the Pearson correlation between the
average number of cambial layers of the trees of both age
groups and the climatic variables as well as the intercorrela-
tion between the climatic variables. The average temperature,
e.g. has a significant positive relationship with rainfall and
tends to have a positive relationship with soil moisture.
Rainfall is significantly positively correlated with soil
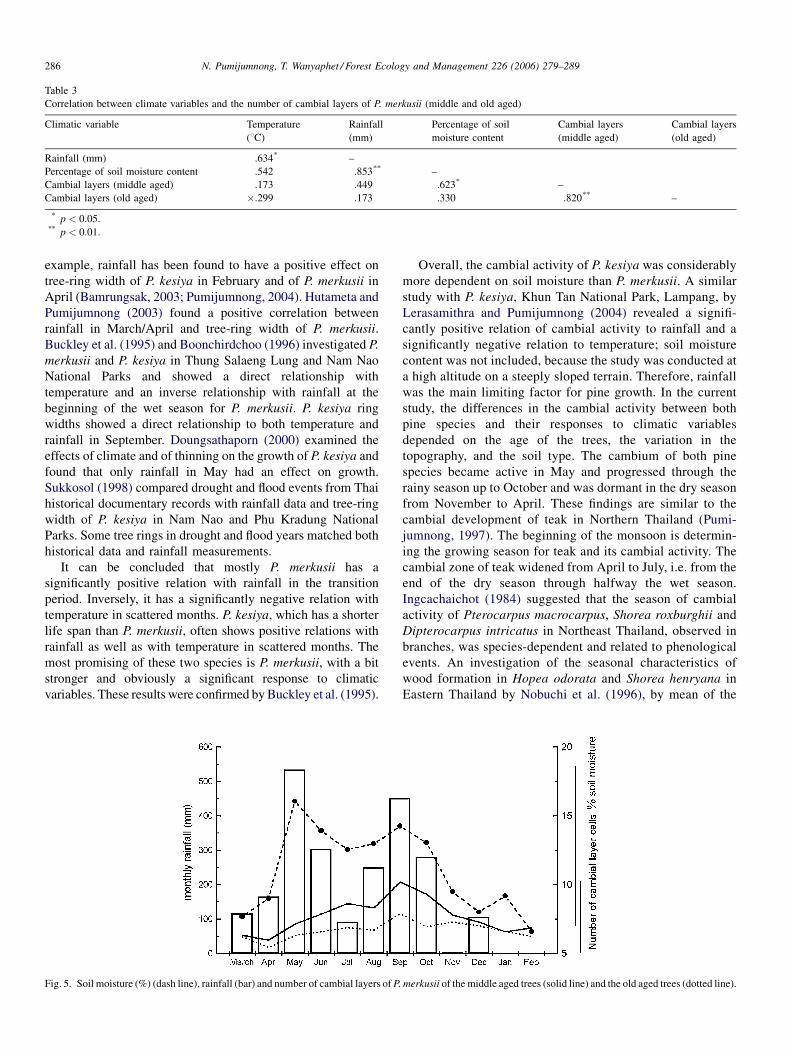
moisture. Fig. 5 displays the connection between soil moisture
content, rainfall and cambial layers of the middle aged and the
old aged trees of P. merkusii over 12 months. The number of
cambial layers of the middle aged trees is significantly
positively correlated with soil moisture but not with to rainfall.
Although, the number of cambial layers of the old aged trees
showed a similar pattern, the correlation with soil moisture
and rainfall is insignificant.
During dormancy (November to April) rainfall at the Hung
Boung site in 2000–2001 was nearly zero in November (3 mm
of rain) and in January (0.3 mm); in February there was no rain
at all. However, rainfall was steady from March to June, then
dropped in July and increased again from August to the end of
October. The soil moisture content generally coincided with
the amount of rainfall, although it was never zero during the
observation year (see Table 1). Even if there is no rain,
moisture can still accumulate in the soil. Another considera-
tion supporting this conclusion is that the site of Hung Boung
is only slightly sloped (about 5–7%), at a high altitude
(1025 m above sea level) and its soil drains easily. In such
conditions, there are no climatic factors limiting pine growth.
It would be logical that growth is enhanced by increased
moisture available over a long portion of the year, particularly
during the dry months.
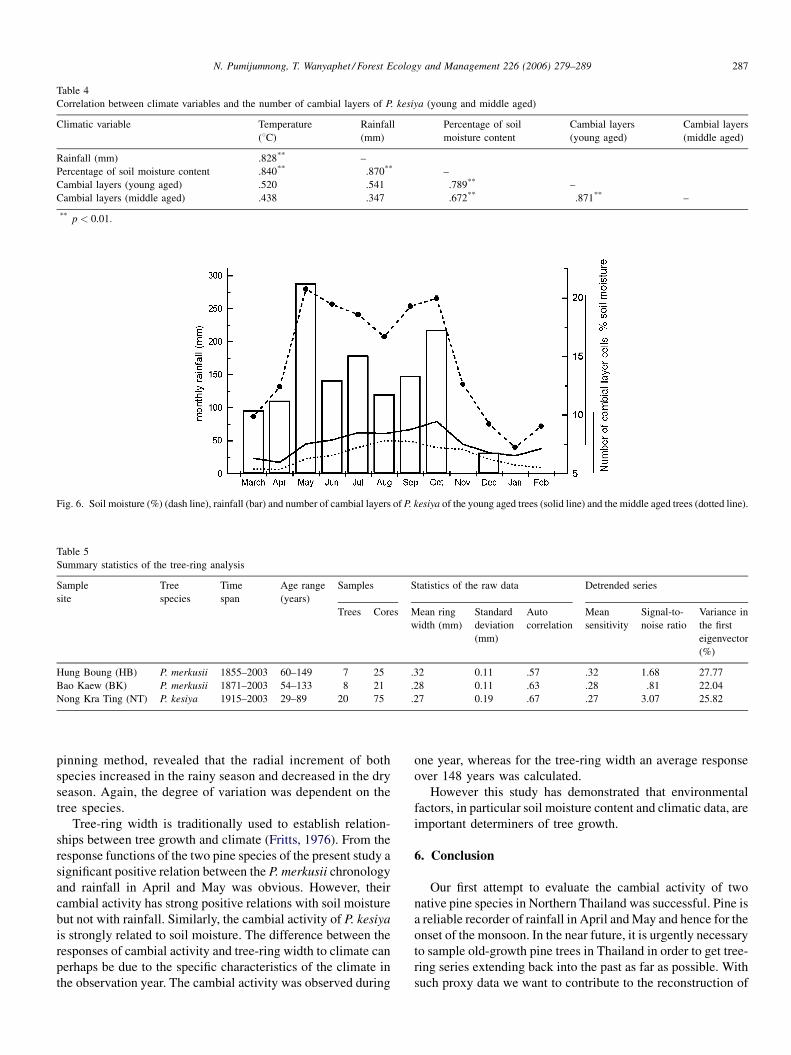
Regarding P. kesiya, high intercorrelations between rainfall,
temperature and soil moisture were found. The number of
cambial layers of the young aged and the middle aged trees
showed a highly positive relationship with soil moisture and in
tendency also with rainfall but with insignificant correlation
(Table 4 and Fig. 6).
4.3. Tree ring analysis
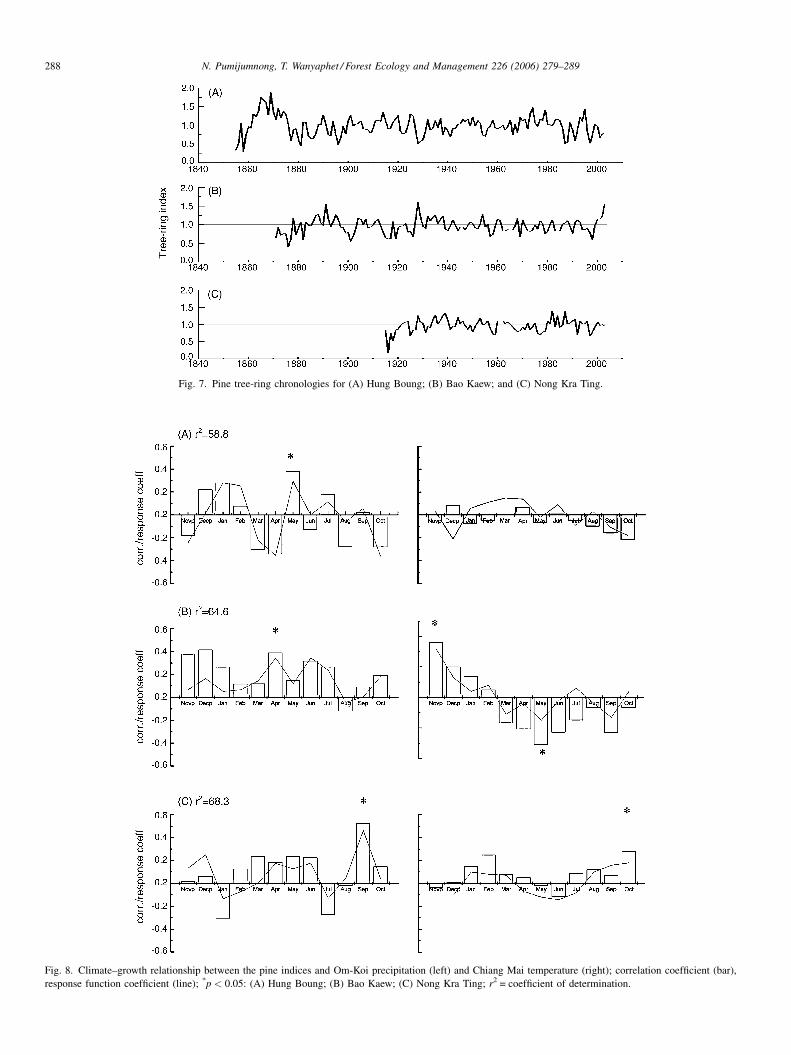
Tree-ring series are widely considered to be appropriate for
dendroclimatological studies if they have high values of mean
sensitivity and common variance in the first eigenvector, a high-
signal-to-noise ratio, and a low value of lag-1 autocorrelation
(Fritts, 1976). In Table 5 the statistics of the chronologies of the
three study sites are summarized; the chronologies are shown as
time series in Fig. 7.
In order to describe the climate–growth relationship, the
monthly rainfall and temperature from November prior to the
growing season to current October were correlated with the
so-called residual tree-ring chronology—that is the chronol-
ogy in which the autocorrelation has been eliminated by
autoregressive modeling (Fig. 8). At the HB site, the highest
positive correlation was with rainfall in May, which is the
beginning of the rainy season in Thailand, whereas tempera-
ture was not limiting growth (=no significant correlation); the
ring-width variation is climatically determined by 58.8%. At
the BK site, the highest positive correlation was with rainfall in
April, which is the transition from the dry to the wet season but
the trees responded positively to above-average rainfall nearly
throughout the entire period from November to July. At the
same time, the correlation with temperature is positive in from
November to July and negative during the whole period from
March to October; the ring-width variation is climatically
determined by 64.6%. At the NT site the highest positive
correlation was unexpectedly with rainfall in September and
with temperature in October, the ring-width variation is
climatically determined by 68.3%.
5. Discussion
Previous studies on the climate–growth response of pine
trees at various sites in Thailand had varying results. For
N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289 285
Fig. 4. The transverse sections of the young age group of P. kesiya, showing seasonal cambial activity from March 2000 to February 2001;�625: (a) and (b) dormant
cambium (November–April); (c)–(h) active cambium of P. kesiya (May–October).
N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289286
Table 3
Correlation between climate variables and the number of cambial layers of P. merkusii (middle and old aged)
Climatic variable Temperature
(8C)
Rainfall
(mm)
Percentage of soil
moisture content
Cambial layers
(middle aged)
Cambial layers
(old aged)
Rainfall (mm) .634* –
Percentage of soil moisture content .542 .853** –
Cambial layers (middle aged) .173 .449 .623* –
Cambial layers (old aged) �.299 .173 .330 .820** –
* p < 0.05.** p < 0.01.
example, rainfall has been found to have a positive effect on
tree-ring width of P. kesiya in February and of P. merkusii in
April (Bamrungsak, 2003; Pumijumnong, 2004). Hutameta and
Pumijumnong (2003) found a positive correlation between
rainfall in March/April and tree-ring width of P. merkusii.
Buckley et al. (1995) and Boonchirdchoo (1996) investigated P.
merkusii and P. kesiya in Thung Salaeng Lung and Nam Nao
National Parks and showed a direct relationship with
temperature and an inverse relationship with rainfall at the
beginning of the wet season for P. merkusii. P. kesiya ring
widths showed a direct relationship to both temperature and
rainfall in September. Doungsathaporn (2000) examined the
effects of climate and of thinning on the growth of P. kesiya and
found that only rainfall in May had an effect on growth.
Sukkosol (1998) compared drought and flood events from Thai
historical documentary records with rainfall data and tree-ring
width of P. kesiya in Nam Nao and Phu Kradung National
Parks. Some tree rings in drought and flood years matched both
historical data and rainfall measurements.
It can be concluded that mostly P. merkusii has a
significantly positive relation with rainfall in the transition
period. Inversely, it has a significantly negative relation with
temperature in scattered months. P. kesiya, which has a shorter
life span than P. merkusii, often shows positive relations with
rainfall as well as with temperature in scattered months. The
most promising of these two species is P. merkusii, with a bit
stronger and obviously a significant response to climatic
variables. These results were confirmed by Buckley et al. (1995).
Fig. 5. Soil moisture (%) (dash line), rainfall (bar) and number of cambial layers of P.
Overall, the cambial activity of P. kesiya was considerably
more dependent on soil moisture than P. merkusii. A similar
study with P. kesiya, Khun Tan National Park, Lampang, by
Lerasamithra and Pumijumnong (2004) revealed a signifi-
cantly positive relation of cambial activity to rainfall and a
significantly negative relation to temperature; soil moisture
content was not included, because the study was conducted at
a high altitude on a steeply sloped terrain. Therefore, rainfall
was the main limiting factor for pine growth. In the current
study, the differences in the cambial activity between both
pine species and their responses to climatic variables
depended on the age of the trees, the variation in the
topography, and the soil type. The cambium of both pine
species became active in May and progressed through the
rainy season up to October and was dormant in the dry season
from November to April. These findings are similar to the
cambial development of teak in Northern Thailand (Pumi-
jumnong, 1997). The beginning of the monsoon is determin-
ing the growing season for teak and its cambial activity. The
cambial zone of teak widened from April to July, i.e. from the
end of the dry season through halfway the wet season.
Ingcachaichot (1984) suggested that the season of cambial
activity of Pterocarpus macrocarpus, Shorea roxburghii and
Dipterocarpus intricatus in Northeast Thailand, observed in
branches, was species-dependent and related to phenological
events. An investigation of the seasonal characteristics of
wood formation in Hopea odorata and Shorea henryana in
Eastern Thailand by Nobuchi et al. (1996), by mean of the
merkusii of the middle aged trees (solid line) and the old aged trees (dotted line).
N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289 287
Fig. 6. Soil moisture (%) (dash line), rainfall (bar) and number of cambial layers of P. kesiya of the young aged trees (solid line) and the middle aged trees (dotted line).
Table 5
Summary statistics of the tree-ring analysis
Sample
site
Tree
species
Time
span
Age range
(years)
Samples Statistics of the raw data Detrended series
Trees Cores Mean ring
width (mm)
Standard
deviation
(mm)
Auto
correlation
Mean
sensitivity
Signal-to-
noise ratio
Variance in
the first
eigenvector
(%)
Hung Boung (HB) P. merkusii 1855–2003 60–149 7 25 .32 0.11 .57 .32 1.68 27.77
Bao Kaew (BK) P. merkusii 1871–2003 54–133 8 21 .28 0.11 .63 .28 .81 22.04
Nong Kra Ting (NT) P. kesiya 1915–2003 29–89 20 75 .27 0.19 .67 .27 3.07 25.82
Table 4
Correlation between climate variables and the number of cambial layers of P. kesiya (young and middle aged)
Climatic variable Temperature
(8C)
Rainfall
(mm)
Percentage of soil
moisture content
Cambial layers
(young aged)
Cambial layers
(middle aged)
Rainfall (mm) .828** –
Percentage of soil moisture content .840** .870** –
Cambial layers (young aged) .520 .541 .789** –
Cambial layers (middle aged) .438 .347 .672** .871** –
** p < 0.01.
pinning method, revealed that the radial increment of both
species increased in the rainy season and decreased in the dry
season. Again, the degree of variation was dependent on the
tree species.
Tree-ring width is traditionally used to establish relation-
ships between tree growth and climate (Fritts, 1976). From the
response functions of the two pine species of the present study a
significant positive relation between the P. merkusii chronology
and rainfall in April and May was obvious. However, their
cambial activity has strong positive relations with soil moisture
but not with rainfall. Similarly, the cambial activity of P. kesiya
is strongly related to soil moisture. The difference between the
responses of cambial activity and tree-ring width to climate can
perhaps be due to the specific characteristics of the climate in
the observation year. The cambial activity was observed during
one year, whereas for the tree-ring width an average response
over 148 years was calculated.
However this study has demonstrated that environmental
factors, in particular soil moisture content and climatic data, are
important determiners of tree growth.
6. Conclusion
Our first attempt to evaluate the cambial activity of two
native pine species in Northern Thailand was successful. Pine is
a reliable recorder of rainfall in April and May and hence for the
onset of the monsoon. In the near future, it is urgently necessary
to sample old-growth pine trees in Thailand in order to get tree-
ring series extending back into the past as far as possible. With
such proxy data we want to contribute to the reconstruction of
N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289288
Fig. 7. Pine tree-ring chronologies for (A) Hung Boung; (B) Bao Kaew; and (C) Nong Kra Ting.
Fig. 8. Climate–growth relationship between the pine indices and Om-Koi precipitation (left) and Chiang Mai temperature (right); correlation coefficient (bar),
response function coefficient (line); *p < 0.05: (A) Hung Boung; (B) Bao Kaew; (C) Nong Kra Ting; r2 = coefficient of determination.

N. Pumijumnong, T. Wanyaphet / Forest Ecology and Management 226 (2006) 279–289 289
the variability of the monsoon climate and thus to its better
understanding.
Acknowledgements
The research was funded by Thailand Research Fund grant
RSA4580035. We thank all members of forest stations for their
gracious help in collecting the data. We would also like to thank
Mr. Harry Toigo and Ms. Patcharin Pumchumnong for their
kind reading of the manuscript. Special thanks are owed to Prof.
Dr. Dieter Eckstein of the University of Hamburg for his strong
comments and suggestions. Finally, we thank anonymous
reviewers for reviewing of the manuscript.
References
Bamrungsak, J., 2003. Response of the tree rings of P. merkusii to climate: a
case study in Mae Hong Son Province, Supan Buri Province and Phechaburi
Province. M.Sc. thesis in Technology of Environmental Planning for Rural
Development, Faculty of Graduate Studies, Mahidol University (in Thai
with English abstract).
Boonchirdchoo, S., 1996. Correlative study of Pinus tree-ring width and
climatic variables. Case Study: Nam Nao and Phu Kradung National Parks,
Thailand. M.Sc. thesis in Technology of Environmental Management,
Faculty of Graduate Studies, Mahidol University.
Buckley, B., Barbetti, M., Watanasak, M., D’Arrigo, R., Boonchirdchoo, S.,
Suratanoon, S., 1995. Dendrochronological investigation in Thailand.
IAWA J. 16 (4), 393–409.
Cook, E.R., 1985. A time-series analysis approach to tree-ring standardization,
Ph.D. dissertation, University of Arizona, Tucson, AZ, USA.
D’Arrigo, R., Barbetti, M., Watanasak, M., Buckley, B., Krusic, P., Boonch-
irdchoo, S., Sarutanon, S., 1997. Progress in dendroclimatic studies of
Mountain pine in Northern Thailand. IAWA J. 18 (4), 433–444.
Doungsathaporn, K., 2000. Effect of climatic factors and thinning on growth of
Pinus kesiya Royle ex Gordon, at Bo Luang Plantation, Amphoe Hod,
Changwat Chiang Mai. M.Sc. thesis in Forestry, Faculty of Graduate
Studies, Kasetsart University (in Thai with English abstract).
Fritts, H.C., 1976. Tree Rings and Climate, second ed. Academic Press,
London.
Holmes, R.L., Adams, R.K., Fritts, H.C., 1986. Tree-ring chronologies of
Western North America: California, Eastern Oregon and Northern Great
Basin with procedures used in chronology development work including
users manual for computer program COFECHA and Arstan chronology
series. Laboratory of tree-ring research. University of Arizona, Tucson, AZ,
USA.
Hutameta, A., Pumijumnong, N., 2003. Pine tree-ring response to climate and
ENSO at Ban Wat Chan, Chiang Mai. Environ. Nat. Resour. J. 1 (2), 69–76.
Ingcachaichot, C., 1984. Seasonal variation of cambial activity of branch of
Pterocarpus macrocarpus, Shorea siamensis, Dipterocarpus intricatus.
M.Sc. thesis in Forestry, Faculty of Graduate Studies, Forestry: Kasetsart
University.
Lerasamithra, V., Pumijumnong, N., 2004. Seasonal variation of cambial
activity of Pinus kesiya in Doi Khuntan National Park, Lampang Province.
Environ. Nat. Resour. J. 2 (1), 1–10.
Nobuchi, T., Janmahasatien, S., Sakai, M., 1996. Seasonal changes of wood
formation and some characteristics of heartwood formation in Teak (Tec-
tona grandis L.) plantation. Kasetsart J. Nat. Sci. 30 (2), 254–263.
Pumijumnong, N., 1997. Cambium development of teak (Tectona grandis L.) in
Thailand and its relationship to climate.In: International Symposium on
Wood Science and Technology. Wood–Human–Environment, Seoul, Korea,
23–24 October 1997, pp. 61–72.
Pumijumnong, N., 2004. El Nino-Southern Oscillation (ENSO) in Thailand by
using tree-ring indices. National Research Council of Thailand, Bangkok,
Thailand (in Thai with English abstract).
Pumijumnong, N., Eckstein, D., Sass, U., 1995a. Tree-ring research on Tectona
grandis in Northern Thailand. IAWA J. 16 (4), 385–392.
Pumijumnong, N., Eckstein, D., Sass, U., 1995b. Reconstruction of rainfall in
northern Thailand from tree-ring series of teak. In: Proceedings of the
IGBP-PAGES/PEP II Symposium on Palaeoclimate and Environmental
Variability during the past 2000 Years in Austral-Asian Transect, Nagoya
University, Nagoya, Japan, 28 November–1 December 1995, pp. 186–191.
Rinn, F., 1986. TSAP. Time Series Analysis Program, Version 2.3. Reference
Manual. Heidelburg, Germany.
Sukkosol, V., 1998. A Study of drought and flood events in Thai historical data
of the 20th century and comparison with meteorological data and tree-ring
indices. M.Sc. thesis in Technology of Environmental Management, Faculty
of Graduate Studies, Mahidol University.
Related Documents











