1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61... 1 of 26 12.04.13 12:36

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.sciencemag.org/content/340/6129/1234598/suppl/DC1
Supplementary Materials for
Mosaic Morphology in the Thorax of Australopithecus sediba
Peter Schmid,* Steven E. Churchill, Shahed Nalla, Eveline Weissen, Kristian J. Carlson,Darryl J. de Ruiter, Lee R. Berger
*Corresponding author. E-mail: [email protected]
Published 12 April 2013, Science 340, 1234598 (2013)DOI: 10.1126/science.1234598
This PDF file includes:
Methods and ResultsTables S1 to S8Figs. S1 to S11References (42–45)
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
1 of 26 12.04.13 12:36

Methods and Results
Comparative analysis of MH1 and MH2 ribs
Materials: Comparative data were collected on ribs 1-12 from samples of Homo sapiens (n=30), Pan troglodytes (n=16), Pongo pygmaeus/abelii (n=12) and Hylobates lar (N=20) from the A.H. Schultz Collection at the Anthropological Institute and Museum of the University of Zurich. Specimens were selected for completeness of the rib series and were primarily adult. To augment sample sizes, subadults (nearing maturity, and of a developmental age roughly comparable to that of Malapa Hominin 1) were included in the human (n = 1), chimpanzee (n = 7), and orangutan (n = 3) samples (table S3). Data were collected on the original specimens of Au. sediba MH1 and MH2. Methods: Seriation of the ribs was accomplished following (41), using the following criteria: (a) Distance between the costotransverse tubercles and the iliocostalis line: The distance from the costotransverse tubercle and the inferior-most aspect of the roughened oblique line on the external surface of the rib at the angle, which forms from the insertion of iliocostalis, increases sequentially in ribs 3-11. In lateral view, the inferior iliocostalis point can usually be seen as a clear inferior projection. (b) Differential shaft shape: the shafts of ribs 3-6 steadily increase in relative width mediolaterally (i.e. internal to external surfaces); beginning with rib 7 they then gradually become relatively narrower and caudally “bladelike” until rib 12. (c) Differential rib size: typical ribs sequentially increase in length and overall cross- sectional size (the latter is more important for fragmentary ribs) to a maximum usually at rib 7, after which they steadily decrease in both parameters. (d) Rib declination: in anatomical position, the angle of the lateral segment of the shaft (i.e. mid-shaft of the rib body) relative to the transverse plane sequentially increases in ribs 3-10 as each lateral shaft is slightly more inferiorly declined than its preceding counterpart. (e) Degree and form of shaft torsion: as rib declination sequentially increases, the torsion of the rib (i.e. spiraling of the shaft about its long axis) produces different orientations of the external border of the mid-shaft. In ribs 3-10, this results in a sequential gradation in which the superior part of the border is first medially-inclined, then vertically-aligned with the inferior border and finally laterally-inclined in the most caudal ribs. (f) Posterior angle: the posterior angle (the point of intersection of the iliocostalis line with the caudal margin of the shaft, generally coincident with the area of the greatest flexion of the shaft) becomes sequentially more obtuse in ribs 3-12. (g) Costal arc prominence: the costal arc is most prominent in ribs 5, 6 and 7. We collected six linear measurements and calculated two indices following (41). All data were collected by a single observer (EW). (1) Tubercle-iliocostal line distance (TID): straight-line distances from the center of the articular tubercle to the furthest extent of the iliocostal line (i.e. the inferior costal point). (2) Posterior angle chord (PAC): straight-line distance measured from the center of the articular tubercle to a point on the inferior aspect of the shaft, whose ventral linear distance from the iliocostal line is equidistant to the TID. Taken with calipers.
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
2 of 26 12.04.13 12:36

(3) Posterior angle subtense (PAS): the subtense from the inferior iliocostal point to the chord PAC. The value for the subtense is geometrically derived from a right triangle whose base is 1/2 PAC and whose hypotenuse is TID. (4) Posterior angle index (PA-INDEX): (PAS/PAC)*100. Relatively larger values for this index reflect a more curved posterior angle. (5) Tuberculo-ventral chord (TVC): straight-line distance between the ventral-most margin of the articular tubercle to the ventral-most point of the sternal end of the rib (taken to the nearest millimeter). (6) Tuberculo-ventral subtense (TVS): perpendicular distance from TVC to the lateral-most extent of the shaft of the rib. (7) Total rib curvature index (TRC-INDEX): (TVS/TVC)*100. Larger values for this index reflect a less “open” curvature, with less mediolateral expansion. (8) Humerus length (HL): Maximum length of the humerus was used to standardize certain rib measurements. The fragmentary nature of the ribs of MH1 and MH2 hampered the collection of osteometric data, and data could only be taken on ribs 1, 2, 4 and 6 (tables S4-S7). Serial position of the ribs of MH2 and MH1 Methods: UW 88-61 is a nearly complete right vertebrosternal rib that was recovered adjacent to and on top of the right upper limb remains of MH2. The size of the rib, along with its torsion, position of the iliocostal line, and shaft cross-sectional morphology all suggest that the rib derives from the upper-to-middle portion of the verterbrosternal series, most likely representing a 4th, 5th or 6th rib. Given the completeness of the rib, and its association with a complete 1st and partial 2nd and 3rd ribs from the same individual, accurate identification of the anatomical position of this rib is important in understanding thoracic shape in Au. sediba. Based on the inclination of the rib, its curvature, the degree of torsion evident in the body, and the rugosity of its iliocostal line (all relative to the preserved morphology evident in the right 3rd rib fragment UW88-166), we have provisionally identified this as a 4th rib. Using the same set of characters, the partial rib represented by the two fragments UW88-167 and -154 appears to be the rib subjacent to UW88-61, and thus the right 5th. We thus appear to have representation of the right side series R1-R5 for MH2, involving two complete ribs and three proximal fragments. The relative position of the iliocostalis line (that is, relative to humeral length) increases uniformly across R2-R5 in MH2 (fig. S3), supporting the serial attributions of each element. Based on size and morphology, the MH1-associated proximal rib fragments UW88-86/41 and UW88-15/131 appear to be right- and left-side (respectively) antimeres. Comparison of their proximal shaft curvatures and iliocostalis attachment area morphology with the proximal ribs of MH2 suggests that these ribs most likely represent the right and left sixth ribs (although they may possibly be seventh ribs). The relative iliocostalis attachment position (TID/HL) of UW88-86/41 (at 0.164) is large for a 6th rib compared to the means of the comparative groups (fig. S3), but within the range of variation observed in the human sample (mean 0.147 ± 0.015, n=29, range 0.121-0.177). The large TID/HL value of UW88-86/41 would also be consistent with the rib having
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
3 of 26 12.04.13 12:36

derived from more inferiorly in the costal series, but would appear to rule out the possibility that the rib comes from higher up in the series. Based on its size, iliocostalis attachment area morphology, and degree of curvature of the posterior angle, the left side shaft fragment UW88-155 appears to be the rib subjacent to UW88-15/131, and thus we have tentatively identified this as the left 7th (or possibly 8th) rib. Posterior angle index Methods: The morphology of the posterior angle of the rib reflects two functionally important aspects of thoracic shape: mediolateral expansion of the rib cage and degree of invagination of the vertebral column. We used the posterior angle index (PA-Index) to quantify the amount of rib curvature expressed at the posterior angle. The PA-Index is derived from the posterior angle subtense and chord, such that higher values indicate a greater amount of curvature at the posterior angle (fig. S4). As can be seen in fig. S4, mediolateral expansion of the thorax is associated with high index values in the ribs. The degree of invagination of the vertebral column is primarily determined by the angular relationship between the neck and the shaft, although curvature at the posterior angle is also likely to contribute to invagination (see figure 13.13 in 16). We examined the extent of curvature at the posterior angle across taxa by plotting the PA-Index by rib for our fossil specimens along with mean values of the comparative groups (fig. 2). We also examined between-group differences in median PA-Index values using the Kruskal-Wallis one-way analysis of variance (with df = 4 for all tests). This nonparametric test was chosen because small fossil sample sizes and moderate comparative sample sizes precluded the reliable assessment of distributional normality in the data and equal variance across samples. Post hoc testing was done using the Tukey-Kramer honestly significant difference (HSD) test. All statistical treatments were conducted in JMP 8.0.2 (42). Because the PA-Index could only be determined in ribs 2, 4, 5 and 6 in Au. sediba, analyses were limited to these ribs. Results: Homo sapiens have PA-Index values that are substantially higher across almost all of the ribs than those of apes or Au. sediba (fig. 2A). Human ribs increase steadily in curvature at the posterior angle from the second through the eighth rib, before becoming straighter in the lower ribs (because the iliocostalis attachment is difficult to identify consistently in human 11th and 12th ribs, we did not calculate the PA-Index for those ribs). Chimpanzee ribs, in contrast, are fairly straight in the upper part of the series, becoming increasingly curved posteriorly from about the sixth to ninth rib (fig. 2A). In the Asian apes (Pongo and Hylobates), the mean values are somewhat variable from rib to rib (perhaps the consequence of small sample sizes), but the overall curvature at the posterior angle does not change markedly from the superior to inferior ribs (fig. 2A). The observed index values for Au. sediba are also variable, but all fall among the mean values for the ape samples and well below the human means. For the second rib, Au sediba has an index value (17.5) that is intermediate between that of the Pan (17.0 ± 5.9, n = 16) and Pongo (18.6 ± 4.5, n = 12) samples, and below that of the Homo (20.5 ± 4.8, n = 27) sample. However, the Kruskal-Wallis test
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
4 of 26 12.04.13 12:36

failed to detect a significant difference in mean values between any of the groups (fig. S5). The Kruskal-Wallis test did, however, detect a significant between-group difference in posterior curvature in the fourth rib (chi-square = 38.42, p < 0.0001). The index value for the right fourth rib of MH2 (18.6) falls above the mean value of Pan (16.7 ± 3.0, n = 16) and very close to that of Pongo (18.7 ± 4.0, n = 12), and is roughly one standard deviation below the mean value for Homo (23.6 ± 4.6, n = 30). Post-hoc testing revealed that the human sample was significantly different from all of the ape samples, while none of the ape samples differed from one another. The single value for Au. sediba did not differ significantly from any of the samples (fig. S6). Similar results were obtained for the fifth and sixth ribs. For the fifth rib, differences between the ape samples and the human sample are even more pronounced (fig. 2 and S7), and this is again reflected in a significant Kruskal-Wallis test statistic (chi-square = 51.31, p < 0.0001). Post-hoc testing again revealed that the mean of the human sample was significantly different from those of all of the ape samples, while again none of the ape sample means differed from any other. Despite falling below the Pan (17.8 ± 2.5, n = 13) and Pongo (17.2 ± 4.2, n = 12) sample means, and more than 3.5 standard deviations below the Homo mean (24.9 ± 2.3, n = 30), the index value for the fifth rib of MH2 (16.4) was not significantly different from any of the comparative samples. The PA-Index for the right sixth rib of MH1 (20.5) is slightly greater than the mean values of the Pan (20.2 ± 1.9, n = 16) and Pongo (19.7 ± 6.2, n = 12) samples, yet is more than 2 standard deviations below the Homo sample mean (26.5 ± 2.7, n = 30). The Kruskal-Wallis test detected a significant difference between groups (chi-square = 50.02, p < 0.0001), and post-hoc testing showed that the human sample mean was significantly different from the means of all of the ape samples, and that the mean for Hylobates was also significantly different from all of the other sample means. Pan and Pongo did not differ from one another in mean index values, but were both significantly higher than the Hylobates mean and lower than the Homo mean (fig. S8). Without exception, the lowest mean PA-Index values for each rib were observed in Hylobates, which is interesting given that the genus is characterized by a less conical, more barrel-shaped rib cage than other apes (12, 13). Our data, along with casual observation of rib morphology in gibbons, suggest that hylobatids exhibit relatively slight invagination of the vertebral column, combined with very little curvature at the posterior angle, and attain a more barrel shape to the thorax through a greater amount of total rib curvature (see fig. 2B). In every rib measured, the PA-Index of Au. sediba falls with the mean values of Pan and Pongo (reflecting relatively straight posterior ribs, consistent with a mediolaterally-narrow upper rib cage) and well below the mean values observed in the Homo sample. Based on the differences in observed medians between samples, and the sample distributions of the PA-Index values in ribs four, five and six (fig. S6-S8), we attribute the lack of significant difference between Au. sediba and H. sapiens to be a problem of small sample sizes, and not a reflection of morphological parity between these species.
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
5 of 26 12.04.13 12:36

Total rib curvature and respiratory area Methods: The curvature of the costal arc was quantified by the total rib curvature index (TRC-Index), which is calculated using the tuberculo-ventral chord (TVC) and subtense (TVS) as 100*TVS/TVC. Larger values of this index indicate more curved ribs. We explored TRC-Index across ribs in the extant hominoid samples, and in the two ribs (1st and 4th) for which this value could be determined for Au. sediba (fig. 2B). We also tested for differences in curvature of the first and fourth ribs between samples using the Kruskal-Wallis one-way analysis of variance (with df = 4 for all tests). This nonparametric test was chosen because small fossil sample sizes and moderate comparative sample sizes precluded the reliable assessment of distributional normality in the data and equal variance across samples. Post hoc testing was done using the Tukey-Kramer honestly significant difference (HSD) test. All statistical treatments were conducted in JMP 8.0.2 (42). Upper thorax size at the level of the 4th rib was explored by reference to the respiratory area enclosed by the rib (41). This area is estimated by approximating half the area of an ellipse using the tuberculo-ventral chord (TVC) and subtense (TVS) as 0.5 (0.5TVC)(TVS). Gómez-Olivencia and colleagues (40) have noted problems with using tuberculo-ventral chords and subtenses with fossils that have, or potentially have, undergone taphonomic distortion. They note that the tuberculo-ventral arc length remains unchanged in plastically deformed ribs, and advocate its use as a general measure of rib size. While we see no evidence of plastic deformation in the 4th rib of MH2 (and thus we feel confident that the total curvature indices reported here accurately reflect the morphology of Au. sediba), we note that estimates of respiratory area should also be immune to problems of taphonomic distortion (since any gain in one dimension is offset by a proportional loss in the other dimension, which keeps the area under the curve constant). Since lung volume scales isometrically with body mass across mammals (37), we used the (square root of 4th rib respiratory area)/HL (= respiratory area index) as a size-standardized measure of upper thoracic size. Statistical testing was conducted as with TRC-Index (above). Results: Extant hominoids vary markedly in the pattern of rib curvature across the rib series (fig. 2B). All of the comparative samples show a large increase in total rib curvature from the first to the second rib. Hylobatids have the highest curvature indices of any of the comparative groups in the superior part of the series (ribs 2-7), peaking in the third rib and then becoming progressively straighter down the sequence (such that the lower ribs have mean values below chimps and humans). Humans are also characterized by high curvature indices in the upper thorax, although not to the extent seen in Hylobates. Humans differ from the gibbon pattern not only in having a somewhat more moderate curvature of the upper ribs, but also in maintaining relatively high curvature in the lower ribs. The ribs of Pan and Pongo are straighter (lower mean indices) than those of Homo (all ribs) and Hylobates (ribs 2-8). In the lower part of the series (ribs 9-11), the mean values for Pan are higher (more curved), and those of Pongo are lower (less curved), than those of Hylobates.
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
6 of 26 12.04.13 12:36

An insufficient number of ribs are represented to discern the overall pattern of curvature across the rib series in Au. sediba. First rib curvature indices for MH1 and MH2 fall well above the mean values of all of the comparative groups, suggesting that Au. sediba might have had greater mediolateral expansion of the cupola of the thorax, although the Kruskal-Wallis test failed to detect a significant difference between any of the groups. The low PA-Index value of the MH2 second rib (see above) suggests that mediolateral expansion of the cupola in Au. sediba, if any, was limited to the first rib. While the first ribs appear to be more curved in Au. sediba than in the comparative groups, the fourth rib appears to be less so. The TRC-Index value from MH2 (32.3) falls very close to (but slight below) the mean values for Pan and Pongo, and well below the means of Homo and Hylobates (fig. 2B). The Kruskal-Wallis test returned a significant chi-square value (50.18, p < 0.0001) and the HSD test revealed that Hylobates had a significantly higher mean than all other groups, that Homo differed significantly from Hylobates on one hand and Pan plus Pongo on the other, and that Pan and Pongo did not differ from one another (but had means significantly below Homo and Hylobates). The single index value for Au. sediba fell significantly below the mean of Hylobates, but was not significantly different from the mean of any other group (fig. S9). Significant differences were found between all of the extant comparative groups in humeral length standardized 4th rib respiratory area (chi-square = 63.58, p < 0.0001), with humans having the largest mean values (reflecting their enlarged upper rib cage and barrel-shaped chests) (table S8, fig. S10). The more conical shape of the chimpanzee thorax, with relatively less respiratory volume superiorly, is reflected in a mean respiratory area index below that of humans (table S8). The lowest mean values are observed in orangutans and gibbons (table S8), which no doubt reflects in part the elongation of the forelimb relative to body size in the Asian apes (43). The fourth rib of MH2 produces an index value 1.5 standard deviations above that of Homo, reflecting a human-like expansion of the volume of the upper thorax. Post-hoc HSD tests show Au. sediba to have an index value that is significantly higher than Pan, Pongo and Hylobates, but not significantly different than that of Homo. However, the low PA-Index values of the upper ribs of MH2 (see above) indicate that expansion of the volume of the upper rib cage was not gained via mediolateral expansion of the superior thorax. Consideration of 4th rib TVS and TVC values (fig. S11) supports this inference: MH2 can be seen to have a fourth rib that has a relatively low subtense value (reflecting a mediolaterally-narrow upper thorax) which falls among those of Pan and Pongo, but which has a relatively large chord (reflecting a large anteroposterior dimension to the upper rib cage). The TVC of MH2 is greater than the mean values of all of the comparative samples (table S6), despite Au. sediba being of a body size considerably smaller than average for Homo or Pongo.
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
7 of 26 12.04.13 12:36
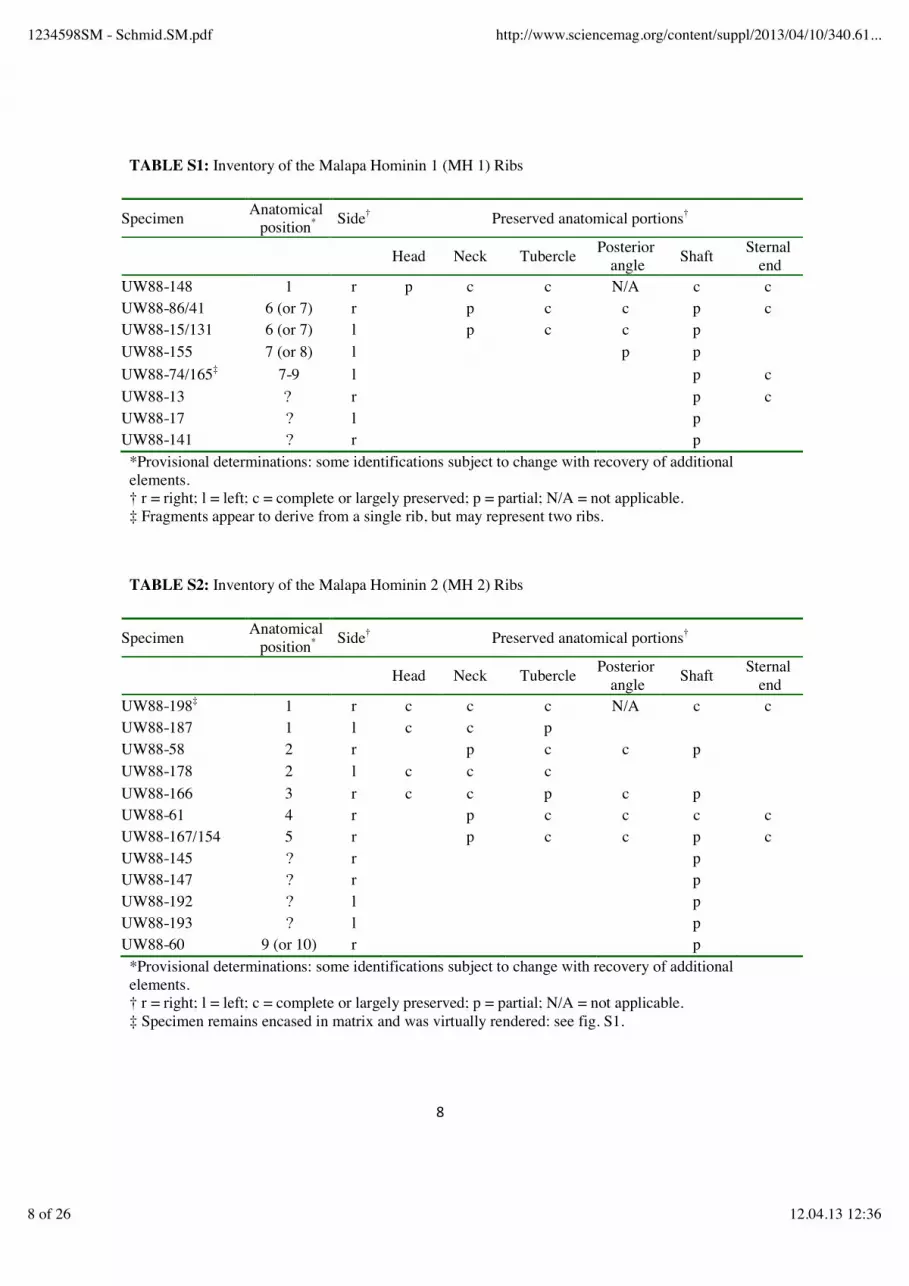
TABLE S1: Inventory of the Malapa Hominin 1 (MH 1) Ribs
Specimen Anatomical position* Side† Preserved anatomical portions†
Head Neck Tubercle Posterior angle Shaft Sternal
end UW88-148 1 r p c c N/A c c UW88-86/41 6 (or 7) r p c c p c UW88-15/131 6 (or 7) l p c c p UW88-155 7 (or 8) l p p UW88-74/165‡ 7-9 l p c UW88-13 ? r p c UW88-17 ? l p UW88-141 ? r p
*Provisional determinations: some identifications subject to change with recovery of additional elements. † r = right; l = left; c = complete or largely preserved; p = partial; N/A = not applicable. ‡ Fragments appear to derive from a single rib, but may represent two ribs.
TABLE S2: Inventory of the Malapa Hominin 2 (MH 2) Ribs
Specimen Anatomical position* Side† Preserved anatomical portions†
Head Neck Tubercle Posterior angle Shaft Sternal
end UW88-198‡ 1 r c c c N/A c c UW88-187 1 l c c p UW88-58 2 r p c c p UW88-178 2 l c c c UW88-166 3 r c c p c p UW88-61 4 r p c c c c UW88-167/154 5 r p c c p c UW88-145 ? r p UW88-147 ? r p UW88-192 ? l p UW88-193 ? l p UW88-60 9 (or 10) r p
*Provisional determinations: some identifications subject to change with recovery of additional elements. † r = right; l = left; c = complete or largely preserved; p = partial; N/A = not applicable. ‡ Specimen remains encased in matrix and was virtually rendered: see fig. S1.
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
8 of 26 12.04.13 12:36
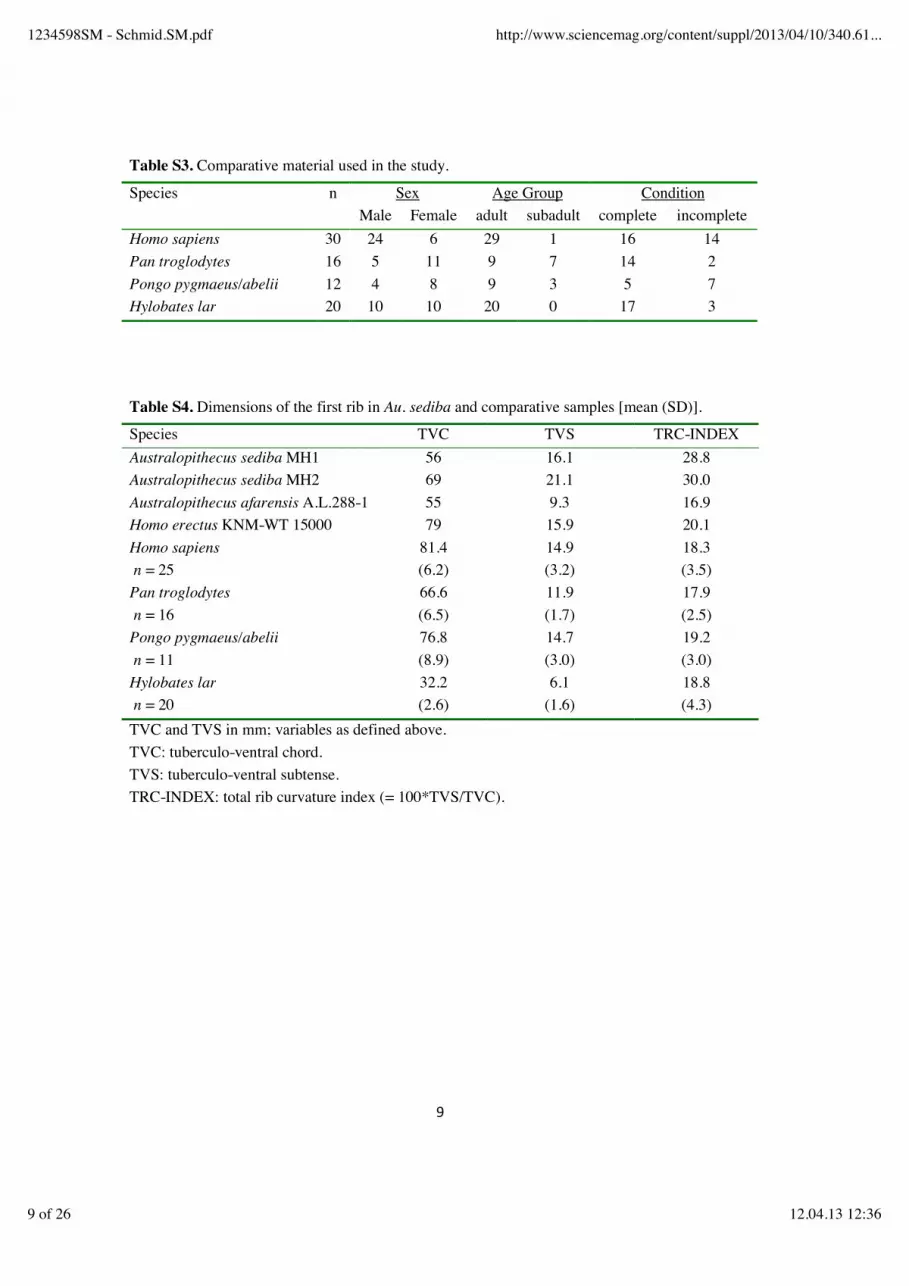
Table S3. Comparative material used in the study.
Species n Sex Age Group Condition Male Female adult subadult complete incomplete Homo sapiens 30 24 6 29 1 16 14 Pan troglodytes 16 5 11 9 7 14 2 Pongo pygmaeus/abelii 12 4 8 9 3 5 7 Hylobates lar 20 10 10 20 0 17 3 Table S4. Dimensions of the first rib in Au. sediba and comparative samples [mean (SD)].
Species TVC TVS TRC-INDEX Australopithecus sediba MH1 56 16.1 28.8 Australopithecus sediba MH2 69 21.1 30.0 Australopithecus afarensis A.L.288-1 55 9.3 16.9 Homo erectus KNM-WT 15000 79 15.9 20.1 Homo sapiens n = 25
81.4 (6.2)
14.9 (3.2)
18.3 (3.5)
Pan troglodytes n = 16
66.6 (6.5)
11.9 (1.7)
17.9 (2.5)
Pongo pygmaeus/abelii n = 11
76.8 (8.9)
14.7 (3.0)
19.2 (3.0)
Hylobates lar n = 20
32.2 (2.6)
6.1 (1.6)
18.8 (4.3)
TVC and TVS in mm; variables as defined above. TVC: tuberculo-ventral chord. TVS: tuberculo-ventral subtense. TRC-INDEX: total rib curvature index (= 100*TVS/TVC).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
9 of 26 12.04.13 12:36

Table S5. Dimensions of the second rib in Au. sediba and comparative samples [mean (SD)].
Species TID PAC PAS PA-INDEX Au. sediba MH2 17.8 35.2 7.7 21.9 Au. africanus Sts 14 14.2 28.4 2.5 8.8 Homo erectus KNM-WT 15000 37.8 68.1 11.4 16.7 Homo sapiens n = 27
31.1 (4.7)
57.3 (7.8)
10.5 (3.2)
18.5 (5.4)
Pan troglodytes n = 16
24.0 (4.5)
45.7 (7.8)
3.9 (1.0)
8.6 (1.6)
Pongo pygmaeus/abelii n = 12
23.4 (1.4)
43.7 (2.7)
3.6 (1.5)
8.2 (2.9)
Hylobates lar n = 20
12.5 (1.3)
23.8 (2.3)
6.1 (1.6)
18.8 (4.3)
TID, PAC and PAS in mm; variables as defined above. TID: tubercle-iliocostalis line distance. PAC: posterior angle chord. PAS: posterior angle subtense. PA-INDEX: posterior angle index (= 100*PAS/PAC).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
10 of 26 12.04.13 12:36

Table S6. Dimensions of the fourth rib in Au. sediba and comparative samples [mean (SD)].
Species TID PAC PAS PA-INDEX Au. sediba MH2 24.7 46.8 3.5 7.5 Au. africanus Sts 14 10.6 21.0 2.1 10.0 Homo sapiens n = 30
34.6 (4.3)
62.4 (7.2)
10.8 (2.4)
17.1 (2.8)
Pan troglodytes n = 16
28.7 (4.4)
54.4 (8.2)
5.4 (1.6)
9.8 (2.3)
Pongo pygmaeus/abelii n = 12
30.2 (2.4)
56.4 (3.8)
6.0 (1.6)
10.6 (2.6)
Hylobates lar n = 20
17.8 (2.0)
33.8 (3.7)
4.5 (1.1)
13.2 (2.1)
Table S6, continued Species TVC TVS TRC-INDEX Au. sediba MH2 176 56.8 32.3 Homo sapiens n = 29
174.1 (9.7)
64.5 (4.5)
37.2 (3.1)
Pan troglodytes n = 16
153.1 (9.6)
50.5 (3.2)
33.0 (2.1)
Pongo pygmaeus/abelii n = 12
162.0 17.2
52.7 (4.3)
32.8 (4.1)
Hylobates lar n = 20
87.6 (7.8)
38.2 (3.2)
43.8 (3.5)
TID, PAC, PAS, TVC and TVS in mm; variables as defined above. TID: tubercle-iliocostalis line distance. PAC: posterior angle chord. PAS: posterior angle subtense. PA-INDEX: posterior angle index (= 100*PAS/PAC). TVC: tuberculo-ventral chord. TVS: tuberculo-ventral subtense. TRC-INDEX: total rib curvature index (= 100*TVS/TVC).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
11 of 26 12.04.13 12:36
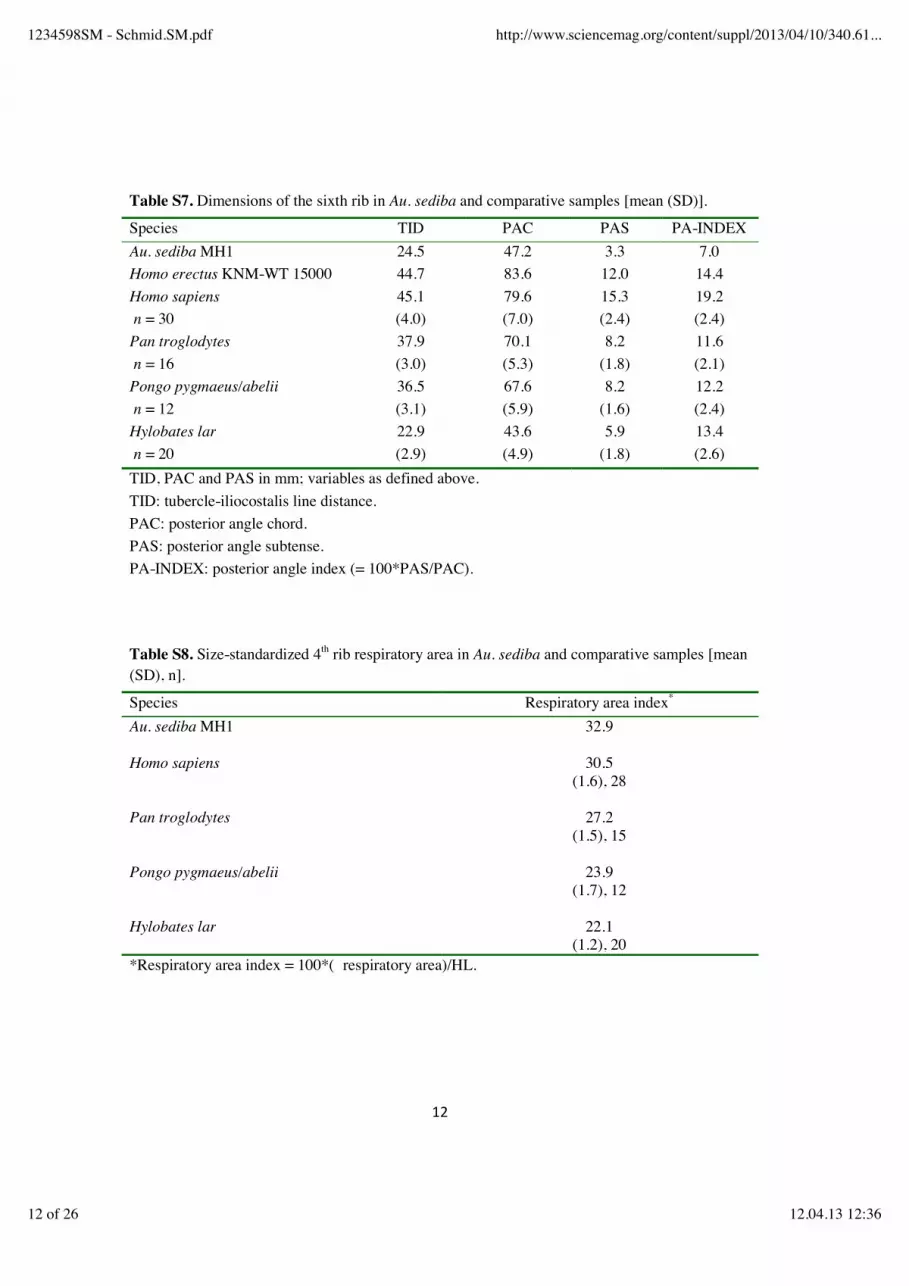
Table S7. Dimensions of the sixth rib in Au. sediba and comparative samples [mean (SD)].
Species TID PAC PAS PA-INDEX Au. sediba MH1 24.5 47.2 3.3 7.0 Homo erectus KNM-WT 15000 44.7 83.6 12.0 14.4 Homo sapiens n = 30
45.1 (4.0)
79.6 (7.0)
15.3 (2.4)
19.2 (2.4)
Pan troglodytes n = 16
37.9 (3.0)
70.1 (5.3)
8.2 (1.8)
11.6 (2.1)
Pongo pygmaeus/abelii n = 12
36.5 (3.1)
67.6 (5.9)
8.2 (1.6)
12.2 (2.4)
Hylobates lar n = 20
22.9 (2.9)
43.6 (4.9)
5.9 (1.8)
13.4 (2.6)
TID, PAC and PAS in mm; variables as defined above. TID: tubercle-iliocostalis line distance. PAC: posterior angle chord. PAS: posterior angle subtense. PA-INDEX: posterior angle index (= 100*PAS/PAC). Table S8. Size-standardized 4th rib respiratory area in Au. sediba and comparative samples [mean (SD), n].
Species Respiratory area index*
Au. sediba MH1
32.9
Homo sapiens
30.5 (1.6), 28
Pan troglodytes
27.2 (1.5), 15
Pongo pygmaeus/abelii
23.9 (1.7), 12
Hylobates lar
22.1 (1.2), 20
*Respiratory area index = 100*( respiratory area)/HL.
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
12 of 26 12.04.13 12:36

Fig. S1: 3D virtual surface reconstruction of the MH2 right first rib (UW88-198). The specimen is embedded in matrix and in contact with the dorsal surface of the manubrium, making its physical removal from the matrix impossible at this time. From top to bottom: lateral, dorsal, cranial (left), caudal (right), ventral, and medial perspectives. Note that the rib has a single facet on the head for articulation with the first thoracic but not the seventh cervical vertebra (the “univertebral pattern”) as found in Au. afarensis and the genus Homo (2, 44) and some other primates (45).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
13 of 26 12.04.13 12:36

Fig. S2: 3D virtual surface reconstruction of the MH2 manubrium. The specimen is embedded in matrix and in contact with the right first rib, making its physical removal from the matrix impossible at this time. Left: dorsal view; right: ventral view.
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
14 of 26 12.04.13 12:36
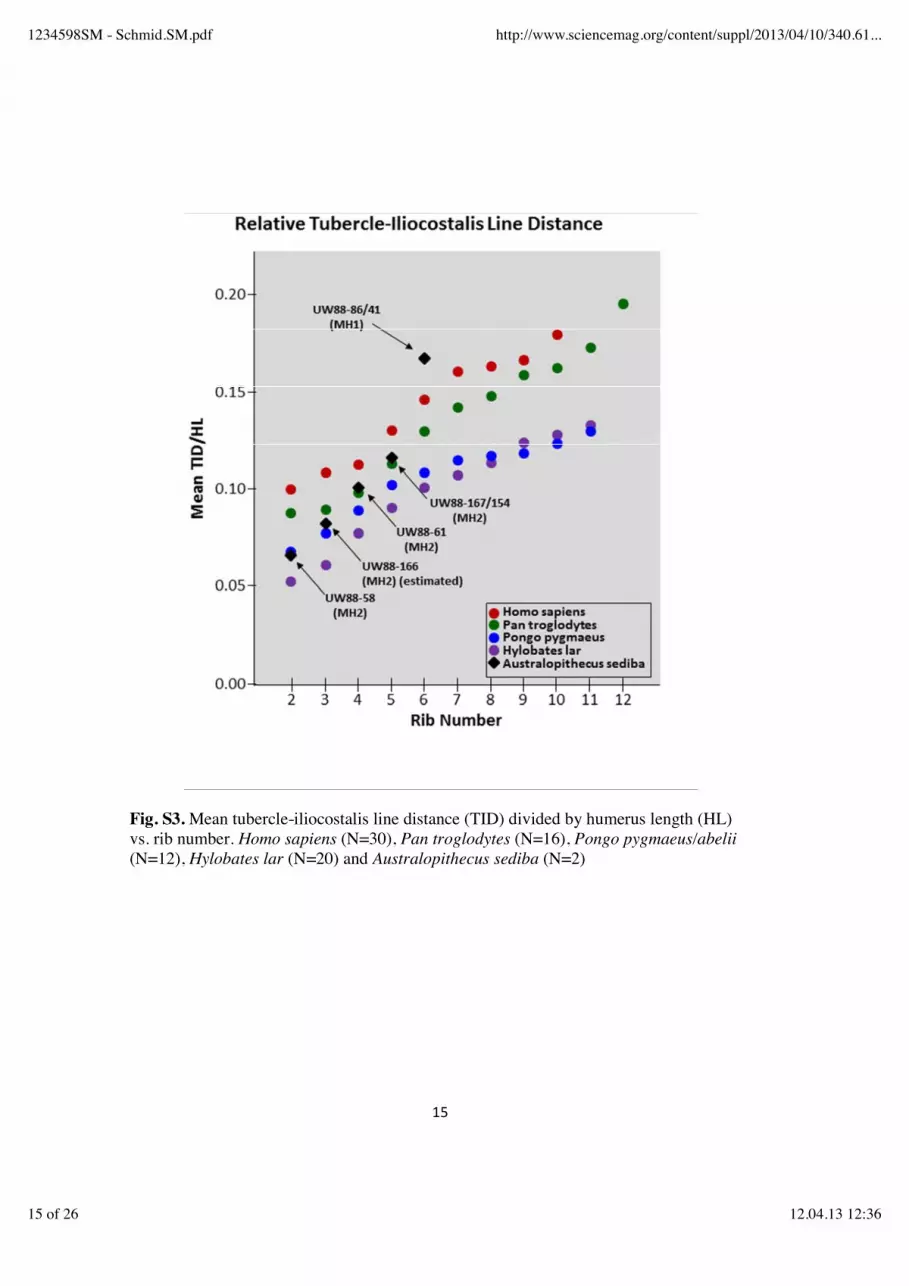
Fig. S3. Mean tubercle-iliocostalis line distance (TID) divided by humerus length (HL) vs. rib number. Homo sapiens (N=30), Pan troglodytes (N=16), Pongo pygmaeus/abelii (N=12), Hylobates lar (N=20) and Australopithecus sediba (N=2)
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
15 of 26 12.04.13 12:36

Fig. S4. Inferior view of ribs 1-6 in a chimpanzee (right) and a human (left), illustrating differences in posterior angle curvature, as reflected in the posterior angle index (PA-Index). The upper ribs of chimpanzees are relatively straight and thus their upper thoraces are not mediolaterally flared. This results in a relatively large posterior angle chord (line marked A in the offset ribs to the right and left) and a short posterior angle subtense (line marked B in the offset ribs), and thus a low value for the PA-Index. Differences between chimpanzees and humans in curvature of the posterior rib can be appreciated by reference to the area subtended by the posterior angle (shaded yellow).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
16 of 26 12.04.13 12:36

Fig. S5. Box and whiskers plot of the posterior angle index for the 2nd rib in Au. sediba and recent hominoids. Plots show the median, upper and lower quartiles (box) and range (whiskers).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
17 of 26 12.04.13 12:36

Fig. S6. Box and whiskers plot of the posterior angle index for the 4th rib in Au. sediba and recent hominoids. Plots show the median, upper and lower quartiles (box) and range (whiskers).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
18 of 26 12.04.13 12:36

Fig. S7. Box and whiskers plot of the posterior angle index for the 5th rib in Au. sediba and recent hominoids. Plots show the median, upper and lower quartiles (box) and range (whiskers).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
19 of 26 12.04.13 12:36

Fig. S8. Box and whiskers plot of the posterior angle index for the 6th rib in Au. sediba and recent hominoids. Plots show the median, upper and lower quartiles (box) and range (whiskers).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
20 of 26 12.04.13 12:36
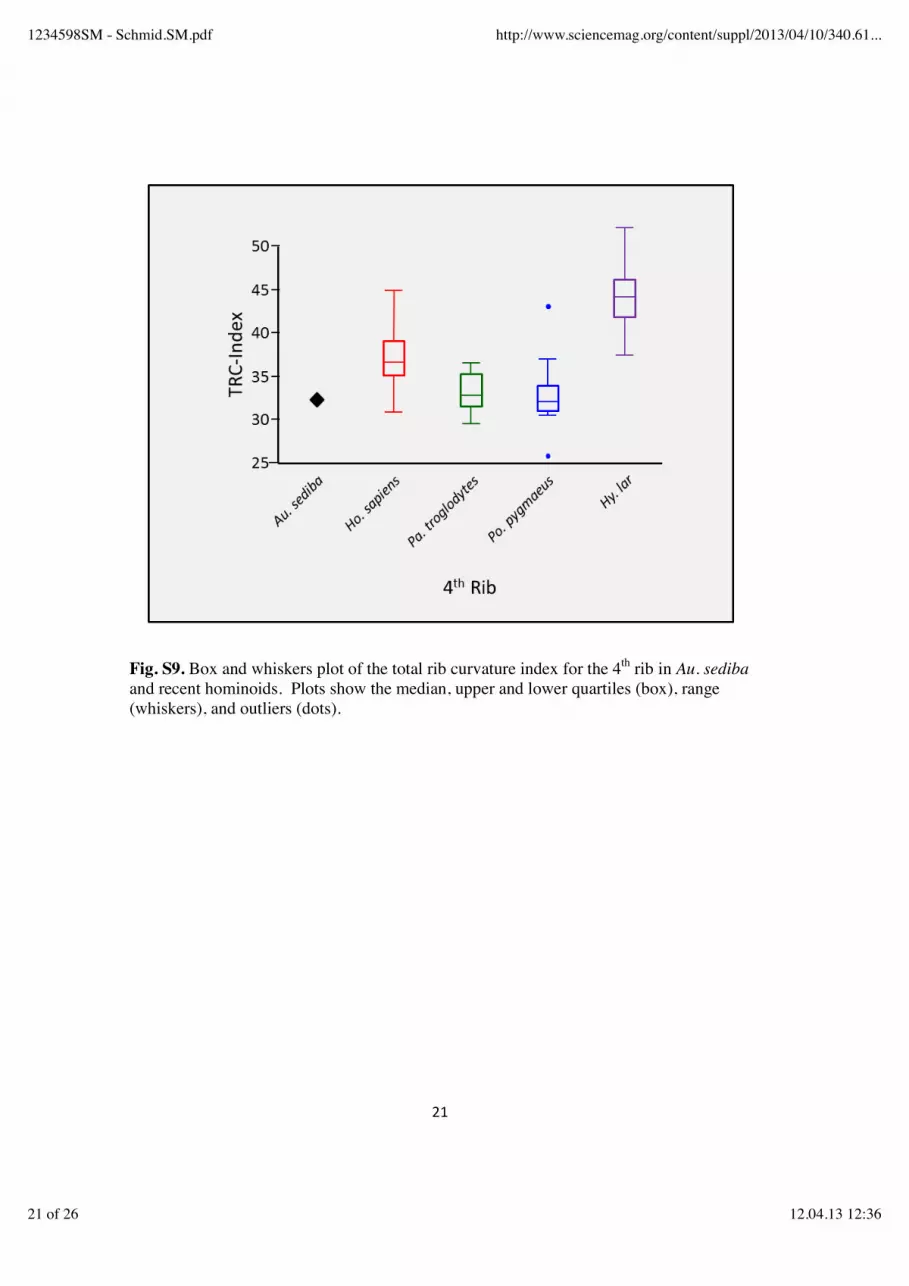
Fig. S9. Box and whiskers plot of the total rib curvature index for the 4th rib in Au. sediba and recent hominoids. Plots show the median, upper and lower quartiles (box), range (whiskers), and outliers (dots).
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
21 of 26 12.04.13 12:36
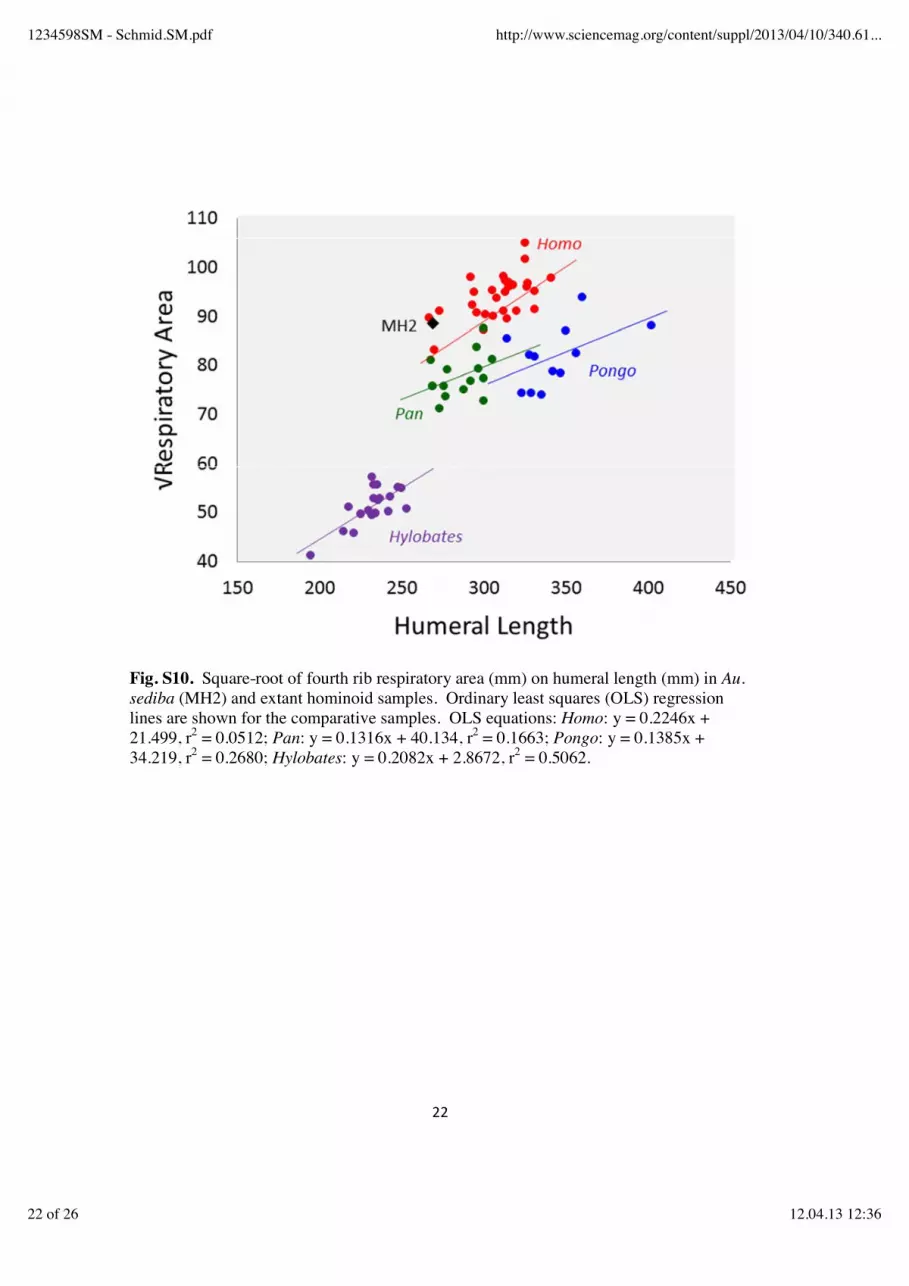
Fig. S10. Square-root of fourth rib respiratory area (mm) on humeral length (mm) in Au. sediba (MH2) and extant hominoid samples. Ordinary least squares (OLS) regression lines are shown for the comparative samples. OLS equations: Homo: y = 0.2246x + 21.499, r2 = 0.0512; Pan: y = 0.1316x + 40.134, r2 = 0.1663; Pongo: y = 0.1385x + 34.219, r2 = 0.2680; Hylobates: y = 0.2082x + 2.8672, r2 = 0.5062.
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
22 of 26 12.04.13 12:36

Fig. S11. Fourth rib tuberculo-ventral subtense (mm) on tuberculo-ventral chord (mm) in Au. sediba (MH2) and extant hominoid samples. Ordinary least squares (OLS) regression lines are shown for the comparative samples. OLS equations: Homo: y = 0.0717x + 52.059, r2 = 0.0239; Pan: y = 0.1575x + 26.374, r2 = 0.2303; Pongo: y = 0.0632x + 42.428, r2 = 0.0629; Hylobates: y = 0.2261x + 18.424, r2 = 0.3070.
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
23 of 26 12.04.13 12:36

24
References and Notes1. P. Schmid, Eine Rekonstruktion des Skelettes von A.L. 288-1 (Hadar) und deren
Konsequenzen [A reconstruction of the skeleton of A.L. 288-1 (Hadar) and itsconsequences]. Folia Primatol. (Basel) 40, 283 (1983). doi:10.1159/000156111Medline
2. J. C. Ohman, The first rib of hominoids. Am. J. Phys. Anthropol. 70, 209 (1986).doi:10.1002/ajpa.1330700208 Medline
3. P. Schmid, The trunk of the australopithecines, in Origine(s) de la Bipédie chez lesHominidés. Cahier de Paléoanthropologie, Y. Coppens, B. Senut, Eds. (Editionsdu CNRS, Paris, 1991), pp. 225–234.
4. C. O. Lovejoy, G. Suwa, S. W. Simpson, J. H. Matternes, T. D. White, The greatdivides: Ardipithecus ramidus reveals the postcrania of our last common ancestorswith African apes. Science 326, 100 (2009). doi:10.1126/science.1175833Medline
5. C. O. Lovejoy, G. Suwa, L. Spurlock, B. Asfaw, T. D. White, The pelvis and femur ofArdipithecus ramidus: The emergence of upright walking. Science 326, 71 (2009).doi:10.1126/science.1175831
6. Y. Haile-Selassie et al., An early Australopithecus afarensis postcranium fromWoranso-Mille, Ethiopia. Proc. Natl. Acad. Sci. U.S.A. 107, 12121 (2010).doi:10.1073/pnas.1004527107 Medline
7. C. V. Ward, Torso morphology and locomotion in Proconsul nyanzae. Am. J. Phys.Anthropol. 92, 291 (1993). doi:10.1002/ajpa.1330920306 Medline
8. L. K. Chan, Scapular position in primates. Folia Primatol. (Basel) 78, 19 (2007).doi:10.1159/000095683 Medline
9. C. V. Ward, Interpreting the posture and locomotion of Australopithecus afarensis:Where do we stand? Am. J. Phys. Anthropol. 119 (suppl. 35), 185 (2002).doi:10.1002/ajpa.10185 Medline
10. D. C. Johanson et al., Morphology of the Pliocene partial hominid skeleton (A.L.288-1) from the Hadar Formation, Ethiopia. Am. J. Phys. Anthropol. 57, 403(1982). doi:10.1002/ajpa.1330570403
11. J. T. Robinson, Early Hominid Posture and Locomotion (Univ. of Chicago Press,Chicago, 1972).
12. A. H. Schultz, Vertebral column and thorax, in Primatolgia, H. Hofer, A. H. Schultz,D. Starck, Eds. (S. Karger, Basel, 1961), vol. 4, pp. 1–66.
13. M. Kagaya, N. Ogihara, M. Nakatsukasa, Morphological study of the anthropoidthoracic cage: Scaling of thoracic width and an analysis of rib curvature. Primates49, 89 (2008). doi:10.1007/s10329-007-0064-z Medline
14. H. Preuschoft, Mechanisms for the acquisition of habitual bipedality: Are therebiomechanical reasons for the acquisition of upright bipedal posture? J. Anat. 204,363 (2004). doi:10.1111/j.0021-8782.2004.00303.x Medline
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
24 of 26 12.04.13 12:36

25
15. K. D. Hunt, Mechanical implications of chimpanzee positional behavior. Am. J. Phys.Anthropol. 86, 521 (1991). doi:10.1002/ajpa.1330860408 Medline
16. L. M. Jellema, B. Latimer, A. Walker, The rib cage, in The Nariokotome Homoerectus Skeleton, A. Walker, R. Leakey, Eds. (Hardvard Univ. Press, Cambridge,MA, 1993), pp. 294-325.
17. D. M. Bramble, D. E. Lieberman, Endurance running and the evolution of Homo.Nature 432, 345 (2004). doi:10.1038/nature03052 Medline
18. C. O. Lovejoy, Evolution of human walking. Sci. Am. 118 (November 1988).19. L. R. Berger et al., Australopithecus sediba: A new species of Homo-like australopith
from South Africa. Science 328, 195 (2010). doi:10.1126/science.1184944Medline
20. T. L. Kivell, J. M. Kibii, S. E. Churchill, P. Schmid, L. R. Berger, Australopithecussediba hand demonstrates mosaic evolution of locomotor and manipulativeabilities. Science 333, 1411 (2011). doi:10.1126/science.1202625 Medline
21. J. M. Kibii et al., A partial pelvis of Australopithecus sediba. Science 333, 1407(2011). doi:10.1126/science.1202521 Medline
22. B. Zipfel et al., The foot and ankle of Australopithecus sediba. Science 333, 1417(2011). doi:10.1126/science.1202703 Medline
23. Methods and background are available as supplementary materials on Science Online.24. C. O. Lovejoy, The natural history of human gait and posture. Part 1. Spine and
pelvis. Gait Posture 21, 95 (2005). Medline25. S. A. Williams et al., The vertebral column of Australopithecus sediba. Science
10.1126/science.1232996 (2013).26. W. C. O. Hill, Observations on a giant Sumatran orang. Am. J. Phys. Anthropol. 24,
449 (1939). doi:10.1002/ajpa.133024032427. S. E. Churchill et al., The upper limb of Australopithecus sediba. Science
10.1126/science.1233477 (2013).28. J. M. DeSilva et al., The lower limb and mechanics of walking in Australopithecus
sediba. Science 10.1126/science.1232999 (2013).29. M. Haeusler, R. Schiess, T. Boeni, New vertebral and rib material point to modern
bauplan of the Nariokotome Homo erectus skeleton. J. Hum. Evol. 61, 575(2011). doi:10.1016/j.jhevol.2011.07.004 Medline
30. L. C. Aiello, P. Wheeler, The expensive-tissue hypothesis. Curr. Anthropol. 36, 199(1995). doi:10.1086/204350
31. C. Ruff, A. C. Walker, Body size and shape, in The Nariokotome Skeleton, A. C.Walker, R. E. Leakey, Eds. (Harvard Univ. Press, Cambridge, 1993), pp. 234-265.
32. P. Schmid, Functional interpretation of the Laetoli footprints, in From Biped toStrider: The Emergence of Modern Human Walking, Running, and Resource
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
25 of 26 12.04.13 12:36

26
Transport., D. J. Meldrum, C. E. Hilton, Eds. (Kluwer Academic/Plenum, NewYork, 2004), pp. 49–62.
33. P. A. Kramer, A. D. Sylvester, Bipedal form and locomotor function: Understandingthe effects of size and shape on velocity and energetics. PaleoAnthropology 2009,238 (2009). doi:10.4207/PA.2009.ART32
34. C. N. Shaw, J. T. Stock, The influence of body proportions on femoral and tibialmidshaft shape in hunter-gatherers. Am. J. Phys. Anthropol. 144, 22 (2011).doi:10.1002/ajpa.21363 Medline
35. S. Cachel, J. W. K. Harris, The paleobiology of Homo erectus: Implications forunderstanding the adaptive zone of this species, in Aspects of AfricanArchaeology, G. Pwiti, R. Soper, Eds. (Univ. of Zimbabwe Publications, Harare,1996), pp. 3-9.
36. H. Pontzer et al., Locomotor anatomy and biomechanics of the Dmanisi hominins. J.Hum. Evol. 58, 492 (2010). doi:10.1016/j.jhevol.2010.03.006 Medline
37. P. Gehr et al., Design of the mammalian respiratory system. V. Scaling morphometricpulmonary diffusing capacity to body mass: Wild and domestic mammals. Respir.Physiol. 44, 61 (1981). doi:10.1016/0034-5687(81)90077-3 Medline
38. L. J. Shapiro, W. L. Jungers, Back muscle function during bipedal walking inchimpanzee and gibbon: Implications for the evolution of human locomotion. Am.J. Phys. Anthropol. 77, 201 (1988). doi:10.1002/ajpa.1330770208 Medline
39. A. Gómez-Olivencia et al., The costal skeleton of Homo antecessor: Preliminaryresults. J. Hum. Evol. 59, 620 (2010). doi:10.1016/j.jhevol.2010.07.023 Medline
40. A. Gómez-Olivencia, K. L. Eaves-Johnson, R. G. Franciscus, J. M. Carretero, J. L.Arsuaga, Kebara 2: New insights regarding the most complete Neandertal thorax.J. Hum. Evol. 57, 75 (2009). doi:10.1016/j.jhevol.2009.02.009 Medline
41. R. G. Franciscus, S. E. Churchill, The costal skeleton of Shanidar 3 and a reappraisalof Neandertal thoracic morphology. J. Hum. Evol. 42, 303 (2002).doi:10.1006/jhev.2001.0528 Medline
42. JMP (SAS Institute Inc., Cary, NC, 2009).43. M. S. M. Drapeau, C. V. Ward, Forelimb segment length proportions in extant
hominoids and Australopithecus afarensis. Am. J. Phys. Anthropol. 132, 327(2007). doi:10.1002/ajpa.20533 Medline
44. J. C. Ohman, A change in morphology in the cervicothoracic region of the vertebralcolumn in hominid evolution. Am. J. Phys. Anthropol. 60, 233 (1983).
45. J. T. Stern Jr., W. L. Jungers, The capitular joint of the first rib in primates: A re-evaluation of the proposed link to locomotion. Am. J. Phys. Anthropol. 82, 431(1990). doi:10.1002/ajpa.1330820405 Medline
1234598SM - Schmid.SM.pdf http://www.sciencemag.org/content/suppl/2013/04/10/340.61...
26 of 26 12.04.13 12:36
Related Documents











