Santander et al., Infection and Immunity 1 Title: 1 Inflammatory Effects of Edwardsiella ictaluri Lypopolysaccharide Modifications in 2 Catfish Gut 3 4 Authors: 5 Javier Santander 1,2* , Jacquelyn Kilbourne 1 , Jie-Yeun Park 1 , Taylor Martin 1,3 , 6 Amanda Loh 1,3 , Ignacia Diaz 1,4 , Robert Rojas 2 , Cristopher Segovia 2 , Dale DeNardo 3 7 and Roy Curtiss 3rd 1,3 . 8 9 1 Center for Infectious Diseases and Vaccinology, The Biodesign Institute, Arizona State 10 University, Tempe, AZ 85287; 2 Microbiology and Immunity Laboratory, Faculty of 11 Sciences, Universidad Mayor, Huechuraba, Chile 8580745; 3 School of Life and Sciences, 12 Arizona State University, Tempe, Arizona 85287; 4 Department of Physics, Master 13 Program in Nanoscience, Arizona State University, Tempe, Arizona 85287. 14 15 Keyword: Edwardsiella, Lipopolysaccharide, catfish, intestinal loops 16 17 Running Title: 18 LPS and Fish Intestinal Inflammation 19 *Corresponding Author: Javier Santander. University Mayor, Faculty of Sciences, Center 20 for Genomic and Bioinformatics, Microbiology and Immunity Laboratory, Camino la 21 Piramide 5750, Huechuraba, Santiago 8580745, Chile. Phone: 56-2-2518-92-05; e-mail: 22 [email protected] 23 24 IAI Accepts, published online ahead of print on 27 May 2014 Infect. Immun. doi:10.1128/IAI.01697-14 Copyright © 2014, American Society for Microbiology. All Rights Reserved. on March 12, 2018 by guest http://iai.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Santander et al., Infection and Immunity
1
Title: 1
Inflammatory Effects of Edwardsiella ictaluri Lypopolysaccharide Modifications in 2
Catfish Gut 3
4
Authors: 5
Javier Santander1,2*
, Jacquelyn Kilbourne1, Jie-Yeun Park
1, Taylor Martin
1,3, 6
Amanda Loh1,3
, Ignacia Diaz1,4
, Robert Rojas2, Cristopher Segovia
2, Dale DeNardo
3 7
and Roy Curtiss 3rd1,3
. 8
9
1Center for Infectious Diseases and Vaccinology, The Biodesign Institute, Arizona State 10
University, Tempe, AZ 85287; 2Microbiology and Immunity Laboratory, Faculty of 11
Sciences, Universidad Mayor, Huechuraba, Chile 8580745; 3School of Life and Sciences, 12
Arizona State University, Tempe, Arizona 85287; 4Department of Physics, Master 13
Program in Nanoscience, Arizona State University, Tempe, Arizona 85287. 14
15
Keyword: Edwardsiella, Lipopolysaccharide, catfish, intestinal loops 16
17
Running Title: 18
LPS and Fish Intestinal Inflammation 19
*Corresponding Author: Javier Santander. University Mayor, Faculty of Sciences, Center 20
for Genomic and Bioinformatics, Microbiology and Immunity Laboratory, Camino la 21
Piramide 5750, Huechuraba, Santiago 8580745, Chile. Phone: 56-2-2518-92-05; e-mail: 22
24
IAI Accepts, published online ahead of print on 27 May 2014Infect. Immun. doi:10.1128/IAI.01697-14Copyright © 2014, American Society for Microbiology. All Rights Reserved.
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
2
ABSTRACT 25
Bacterial lipopolysaccharides (LPS) are structural components of the outer membranes of 26
Gram-negative bacteria and also are potent inducers of inflammation in mammals. Higher 27
vertebrates are extremely sensitive to LPS but lower vertebrates, like fish, are resistant to 28
their systemic toxic effects. However, LPS effects on the fish intestinal mucosa remain 29
unknown. Edwardsiella ictaluri is a primitive member of the Enterobacteriaceae family 30
that causes enteric septicemia in channel catfish (Ictalurus punctatus). E. ictaluri infects 31
and colonizes deep lymphoid tissues upon oral or immersion infection. Both gut and 32
olfactory organs are the primary sites of invasion. At the systemic level E. ictaluri 33
pathogenesis is relatively well characterized, but our knowledge about E. ictaluri 34
intestinal interaction is limited. Recently, we observed that E. ictaluri oligo-35
polysaccharide (O-PS) LPS mutants have differential effects on the intestinal epithelia of 36
orally inoculated catfish. Here we evaluate the effects of E. ictaluri O-PS LPS mutants 37
using a novel catfish intestinal loop model and compared it to the rabbit ileal loop model 38
inoculated with Salmonella Typhimurium LPS. We found evident differences in rabbit 39
ileal loop and catfish ileal loop responses to E. ictaluri and S. Typhimurim LPS. We 40
determined that catfish respond to E. ictaluri LPS, but not to S. Typhimurium LPS. We 41
also determined that E. ictaluri inhibits cytokine production and induces disruption of the 42
intestinal fish epithelia in an O-PS dependent fashion. E. ictaluri wild type and wibT 43
LPS mutant caused intestinal tissue damage and inhibited pro-inflammatory cytokine 44
synthesis in contrast to E. ictaluri gne and ugd LPS mutants. We concluded that the E. 45
ictaluri O-PS subunits play a major role during pathogenesis, since that they influence the 46
recognition of the LPS by the intestinal mucosal immune system of the catfish. The LPS 47
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
3
structure of E. ictaluri mutants is need to understand the mechanism of interaction. 48
INTRODUCTION 49
The genus Edwardsiella, which consists of four species E. tarda, E. hoshinae, E. 50
piscida and E. ictaluri, is one of the most primitive members of the Enterobacteriaceae 51
family (1). E. ictaluri is one of the most important pathogens of commercially raised 52
channel catfish (I. punctatus) (2), which account for more than 80% of U.S. aquaculture 53
production, in spite of the recent production decrease (3,4). E. ictaluri infects and 54
colonizes catfish internal lymphoid tissues upon oral or bath infection making it a 55
promising strain to develop effective live attenuated recombinant vaccines for the catfish 56
industry (5). Both gut and olfactory organs are the primary sites of invasion of E. ictaluri 57
in natural outbreaks (6). E. ictaluri crosses the intestinal mucosa of channel catfish in 15 58
min after oral inoculation with 109 CFU (7). Although there are substantial descriptive 59
data relative to the invasion, spread, and persistence of E. ictaluri in channel catfish 60
(7,8,9), little is known about the molecular mechanisms of E. ictaluri fish intestinal 61
pathogenicity and pathogen associate molecular pattern (PAMPs) recognized by fish. 62
One of the most studied PAMPs is the lipopolysaccharide (LPS) that in mammals 63
is recognized by the Toll-like receptor 4 (TLR-4) (10,11,12). LPS is the major component 64
of the external layer of the outer membrane of Gram-negative bacteria. LPS is composed 65
of three distinct parts: carbohydrate subunits or oligo-polysaccharides (O-PS), the 66
oligosaccharide core region and the lipid A that is responsible for the activation of the 67
innate immune response in mammals and confers the endotoxic properties of the LPS 68
(13). On the other hand, fish, in contrast to mammals, are remarkable resistant to the 69
toxic effects of the LPS (14,15,16). 70
LPS is an important virulence factor for E. ictaluri (17,18,19). The E. ictaluri LPS 71
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
4
gene cluster has been identified by transposon mutagenesis (17,18) and recently fully 72
described (19). E. ictaluri LPS O-PS mutants exhibited different levels of virulence, 73
tissue colonization, and intestinal gut inflammation in orally inoculated catfish (19). In 74
deed, E. ictaluri wild type causes diarrhea-like symptoms in orally infected fish in 75
contrast to E. ictaluri LPS mutant strains (19). This observation correlates with the 76
current idea that fish recognize the O-PS of the LPS instead of the lipid A (20) and that 77
fish recognize LPS at the intestinal level. 78
Ligated ileal loops have been used to evaluate the contribution of LPS to 79
intestinal bacterial colitis in rabbit, mice and calves (21,22,23,24). Initially ligated loops 80
of rabbit small intestine have been used as a model to assess the contribution of putative 81
virulence factors to bacterial pathogen-induced diarrhea. This model was used for Vibrio 82
cholerae, where the injection of whole cultures (25,26), culture supernatants (27) and cell 83
extracts (28) caused dilation of the loop due to fluid accumulation. The rabbit ileal loop 84
model also has been used to study pathogenesis of Escherichia coli (29,30,31), 85
Salmonella (21,32,33), Shigella (34), Pseudomonas aeruginosa (35), Clostridium 86
perfringens (36), V. parahemolyticus, V. alcaligenes (37) and Bacillus cereus (38). In the 87
search for a model that mimics the human intestinal bacterial infection and inflammatory 88
responses, murine and bovine ligated ileal loops also have been used (39,40,41). 89
The complete LPS structure of E. ictaluri has not been elucidated. Nevertheless, 90
the composition and structure of the E. ictaluri O-PS has been reported (42). The E. 91
ictaluri typical O-chain was found to be an unbranched linear polymer of a repeating 92
tetrasaccharide unit composed of D-glucose, 2-acetamido-2-deoxy-D-galactose, and D-93
galactose in a 1:2:1 ratio having the structure: [4)--D-Glcp-(14)--D-GalpNAc-94
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
5
(13)--D-GalpNAc-(14)--D-Galp-(1]n (42). The E. ictaluri O-PS biosynthesis 95
enzymes are encoded by four genes, wibT, galF, gne and ugd, located in the O-PS gene 96
cluster (18,19). As mentioned previously, we determined that orally inoculated catfish 97
with E. ictaluri wild type developed diarrhea-like symptoms in contrast to fish inoculated 98
with LPS defective mutants (wibT, gne and ugd) (19). Intestinal diseases often lead 99
to disruption of the intestinal epithelial barrier either trough attachment and 100
internalization mediated effector molecule release, or through stimulation of host 101
inflammatory responses which ultimately compromise junctional integrity (43). Several 102
studies have begun to explore the cellular and molecular composition of mucosal surfaces 103
in salmonids (44,45), carp (46), cod (47), flounder (48) and catfish (49,50,51). Recently, 104
it has been suggested that E. ictaluri survive in intestinal macrophages (19) and caused 105
intestinal barrier disruption and immune suppression (49). Using a novel the catfish 106
intestinal loop model we corroborated that E. ictaluri caused intestinal barrier disruption 107
and immune suppression in a LPS O-PS dependent fashion. Furthermore, we determined 108
that E. ictaluri LPS O-PS plays a mayor role during catfish intestinal infection and 109
immune protective stimulation by a live attenuated E. ictaluri vaccine. 110
111
MATERIALS AND METHODS 112
113
Ethics statement. All research involving fish was conducted as per Protocol #09-1042R, 114
approved by the Arizona State University Institutional Animal Care and Use Committee. 115
116
Bacterial strains, media, and regents. The bacterial strains and plasmids are listed in 117
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
6
Table 1. Bacteriological media and components are from Difco (Franklin Lakes, NJ). 118
Antibiotics and reagents are from Sigma (St. Louis, MO). LB broth (tryptone, 10 g; yeast 119
extract 5 g; NaCl, 10 g; dextrose 1 g; ddH2O, 1L) (56) and Bacto-Brain Heart Infusion 120
(BHI) were used routinely. When required, the media were supplemented with 1.5% agar 121
or colistin sulphate (Col; 12.5 μg/ml). Bacterial growth was monitored 122
spectrophotometrically and/or by plating. 123
124
Bacteria inoculate preparation. Bacterial strains were grown overnight standing and 125
then the cultures were diluted 1:20 in prewarmed BHI broth and grown with mild 126
aeration (180 rpm) at 28C to an OD600 of 0.8 to 0.9 (~108 CFU/ml). Bacteria were 127
sedimented 10 min by centrifugation (7,000 rpm) at room temperature and resuspended 128
in saline (NaCl 0.85%) to appropriate densities for inoculation. 129
130
Bile Sensitivity. Sensitivity to bile was determined by the microplate serial dilution 131
assay. This assay was performed using flat bottom 96-well clear microtitre plates. Ox bile 132
and sodium deoxycholate were serially diluted in BHI broth and then inoculated with 133
mid-log-phase cultures of the E. ictaluri strains. The plates were incubated during 48 h 134
28C. 135
136
LPS purification and analysis. LPS extraction was performed by using TRI-regent 137
(Sigma) as described previously (57). LPS profile was evaluated by sodium dodecyl 138
sulfate-polyacrylamide gel electrophoresis and visualized by silver staining (58,59). 139
Protein contamination was evaluated by SYPROTM
Ruby (Invitrogene) staining and UV 140
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
7
scanning visualization (Typhoon Trio multi-mode imager, GE Healthcare) compared to 141
commercial LPS (Sigma). 142
143
Outer membrane protein (OMPs) preparation and purification. Sarkosyl-insoluble 144
outer membrane proteins (OMPs) were obtained as previously described (60). The outer 145
membrane proteins were LPS detoxified and normalized to 25 μg/μl by using the 146
nanodrop spectrophotometer (ND-1000, NanoDrop) and separated by 10% (wt/vol) 147
sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis for verification. 148
Coomassie blue staining was performed to visualize proteins. The residual LPS was 149
removed from the OMPs by Detoxi-GelTM Endotoxin columns (Thermo) and verified by 150
LPS profiles. 151
152
Rabbit surgery. The rabbits were fasted (but provided ad lib water) overnight 153
(maximum 16 hours) and then pre-medicated with 30 mg/kg ketamine and 6 mg/kg 154
xylazine intra muscular (i.m). Then, while masked with isoflurane in oxygen to a surgical 155
level of anesthesia (as indicated by lack of toe pinch, ear twitch reflexes and stable heart 156
rate via pulse oximetry), the ventral surface of the neck was shaved and disinfected by 157
sequential washes of chlorhexidine and alcohol. A cut down tracheotomy was performed 158
and an endotracheal tube placed to ensure a patent airway throughout the procedure. At 159
this point, mask administration of isoflurane was discontinued and further administration 160
was delivered via the endotracheal tube. Additionally, an intravenous catheter was placed 161
in an ear vessel to deliver lactated Ringers solution. The rabbits were monitored for depth 162
of anesthesia using pulse oximetry (heart rate and SpO2), ventilatory rate, and the lack of 163
a toe pinch reflex. Abdominal hair was removed along the mid-ventral body wall and the 164
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
8
skin disinfected using sequential washes with chlorhexidine and alcohol. An 165
approximately 10 cm midline incision was made through the skin and body wall. The 166
distal most region of the ileum was isolated, and a 4-6 cm section was double-ligated at 167
each end. Working proximally up the intestine, 6 more intestinal loops were created, each 168
1-2 cm apart. Throughout the procedure, sterile saline and moist gauze were used to keep 169
the viscera and body walls moist. After creation of all loops, 1.0 ml of LPS (100 g) or 170
bacterial culture (about 107 CFU) or sterilized saline (NaCl 0.85%) as a negative control 171
was injected into each loop. The body wall and skin were separately closed using a 172
simple continuous suture pattern. The rabbit was anesthetized for six hours, being 173
continuously observed. After 6 h, the rabbit was euthanized using an overdose of sodium 174
pentobarbital (150 mg/kg) injected intravenously. The intestinal loops were then 175
removed, the length of each segment measured, and the intestinal fluid extracted with the 176
volume-to-length ratio in milliliters per centimeter for each loop recorded. A piece of the 177
intestinal tissue was fixed in 10% formalin and subjected to histopathological study. Each 178
slide was graded on the basis of degree of mucosal disruption, cellularity, and vascular 179
congestion. 180
181
Channel catfish surgery. Thirty five outbreed channel catfish specific-pathogen-free 182
with a mean weight of 2 kg ± 10 g were used. The animals were acclimatized during 1 183
week prior to surgery in 100-liter tanks at 26 ± 1°C. Each tank is equipped with a re-184
circulating, biofiltered, mechanical filtered, and U.V. water treated system with 12 h light 185
cycle per day. The fish were fed daily with commercial Aquamax (Purina Mills Inc., St. 186
Louis, MO). The water quality was monitored for pH, NO2, and NO3 with standard kits. 187
Two days prior surgery the animals were fastened. The fish were anesthetized with 188
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
9
buffered tricaine methanesulfonate (pH 7.5)(61). Four anesthesia doses were used, fish 189
handling dose (15 mg/L; 25-26C), fish surgery dose (100 mg/L; 20-22C), recovery (30 190
mg/L; 20C-22C) and euthanasia (300 mg/L; 10-15C). From the acclimatization tanks, 191
the fish were moved to the handling anesthesia dose during at least 20 min. Then the fish 192
was moved to the surgery platform in a supine position (Fig. 1) and connected to the 193
surgical anesthesia dose. The fish platform developed here is a modified version of 194
previously described fish surgery platforms (62,63). The body is partially submerged in 195
water (the surgical site will remain above the water line) and the recirculating water flows 196
continuously through the mouth and over the gills (Figs. 1A and 1B). This permitted to 197
have the fish in position for the surgery while effectively ventilating the fish. Once the 198
fish was fully anesthetized, its body wall was cut and the coelomic cavity entered via 199
blunt dissection (Fig. 1C). The lower small intestine was isolated and up to 6 sections of 200
3 cm each were prepared by double-ligation (Fig. 1D). One hundred μg of purified LPS 201
in 500 l of phosphate saline buffer (PBS) or 500 l of 107 CFU/ml of E. ictaluri strains 202
were injected into each intestinal section (Fig. 1E). The control consisted in loops 203
inoculated with PBS, OMPs (100 g) or peptidoglycan from Staphylococcus aureus (100 204
g; Sigma). After injection, the body wall was sutured closed (Fig. 1F), and the fish was 205
moved to a bath containing a recovery anesthesia dose where it remained for up to 6 h to 206
allow time for the intestine to respond to the inoculate (Fig. 1G). After this period, the 207
fish was euthanized with a high concentration of buffered tricaine (300 mg/L) followed 208
by the harvesting of vital organs as a secondary method. The intestinal loops were then 209
removed, the length of each segment measured, and the intestinal fluid extracted with the 210
volume-to-length (V:L) ratio in milliliters per centimeter for each loop recorded. A piece 211
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
10
of the intestinal tissue was fixed in 10% formalin and subjected to histopathological 212
study. Each slide was graded on the basis of degree of mucosal disruption, cellularity, 213
and vascular congestion. 214
215
Quantitative real-time polymerase chain reaction (qRT- PCR). Gut samples were 216
frozen in liquid nitrogen prior to grinding. Total RNA was isolated using TRIzol®
LS 217
Reagent (Invitrogen) according to manufacturer’s instructions. Extracted RNA was 218
quantified by UV absorption using a NanoDrop ND-1000 spectrophotometer (NanoDrop 219
Technologies). The RNA was stored at -80℃ before use in complementary DNA (cDNA) 220
synthesis. Double-stranded cDNA was synthesized using Super Script®
III 1st Strand 221
Synthesis Kit (Invitrogen) in a final volume of 20 µl containing 2 µg of total RNA, 50 ng 222
of random hexamers, 1 µl dNTP (10 mM), 2 l DTT (0.1M), 2 μl 10 x RT buffer, 4 μl 223
MgCl2 (25 mM), 1 μl RNase-out (40U/μl), and 1 μl SuperScript III RT (200 U/μl) and 224
incubated at 50℃ for 50 min and at 85℃ for 5 min to terminate the reactions. 1 l of cDNA 225
was subsequently used as template in quantitative real time PCR using catfish specific 226
inflammatory cytokine primers listed in Table 2. RT-PCR were performed using the iQTM
227
SYBR®
Green Supermix (Bio-Rad) on the Multicolor Real-Time PCR Detection System 228
(Bio-Rad) with programmed thermal cycling conditions consisting of 40 cycles of 95°C 229
for 10 sec, 60 °C for 30 sec, and 72 °C for 30 sec. Each samples normalized to the 230
equivalent of the house-keeping gene, β-actin. The relative expression of the target gene 231
was estimated from the threshold cycles (Ct) according to the 2-ΔΔCt
methods (64). 232
Statistical analyses were performed using GraphPad Prism Version 6.00 for Windows 233
(Graph-Pad Software). Statistical comparison was performed using unpaired Student’s t-234
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
11
test. The significance level of the student t-test was set at P<0.05. 235
236
Statistics. An ANOVA (SPSS Software) analysis, followed by LSD (Least Significant 237
Difference) method, was used to evaluate differences in bacterial titers discerned to 95% 238
confidence intervals. P<0.05 was considered statistically significant. 239
240
RESULTS 241
242
E. ictaluri bile sensitivity. Bile is one of the main antibacterial components of the 243
intestinal fluids. Thus, we evaluate whether the E. ictaluri LPS (Fig. 2) mutant strains are 244
sensitive to Ox bile and sodium deoxycholate (Table 1). All the strains used in this study 245
were highly resistant to bile, suggesting that the LPS does not play role on bile resistance 246
in E. ictaluri. 247
248
Rabbit intestinal ileal loops inoculated with purified LPS. S. Typhimurium LPS 249
triggered significant fluid secretion with presence of blood (Fig. 3A), which correlated 250
with the intestinal inflammation and tissue damage (Fig. 4). In contrast, E. ictaluri wild 251
type LPS triggered low fluid secretion (Fig. 3A) and mild inflammation of the intestinal 252
epithelia without severe tissue damage (Fig. 4). E. ictaluri LPS mutants, including 253
wibT, gne and ugd, did not trigger fluid secretion (Fig. 3A). E. ictaluri wibT and 254
gne LPS triggered mild epithelial damage (Fig. 4). In contrast, E. ictaluri ugd LPS did 255
not cause tissue damage on the rabbit ligated loops (Fig. 4). Loops injected with OMPs or 256
peptidoglycan did not show any differences compared to the control. 257
258
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
12
Catfish intestinal loops inoculated with LPS. All purified LPS molecules from S. 259
Typhimurium or E. ictaluri triggered similar and significant levels of fluid secretion in 260
the catfish gut (Fig. 3B). However, S. Typhimurium LPS did not cause evident tissue 261
damage in contrast to E. ictaluri wild-type LPS that caused massive tissue damage and 262
gut inflammation (Fig. 4 and 5). Purified LPS from E. ictaluri wibT caused notorious 263
tissue damage in contrast to E. ictaluri gne LPS, which did not cause tissue damage or 264
significant inflammation (Figs. 4 and 5). E. ictaluri ugd LPS triggered a mild 265
inflammation with evident epithelial cell-cell junction disruption (Figs. 4 and 5). Loops 266
injected with OMPs or peptidoglycan did not show differences with respect to the 267
control. It has been established that fish intra-coelomic injected with LPS or fish 268
macrophages exposed to LPS do not generate an inflammatory immune response (20). 269
However, fish macrophages exposed to peptidoglycan mount an inflammatory immune 270
response (20). Catfish intestinal loops injected with purified peptidoglycan (100 g/dose) 271
did not show differences compared to the control (Fig. 6). 272
273
Catfish intestinal loops inoculated with E. ictaluri strains. E. ictaluri wild-type and S. 274
Typhimurium triggered significant fluid secretion in contrast with the mock negative 275
control (Figs. 3C and 4). However, E. ictaluri wild type caused extensive tissue damage 276
and inflammation in contrast with S. Typhimurium, which did not cause tissue damage 277
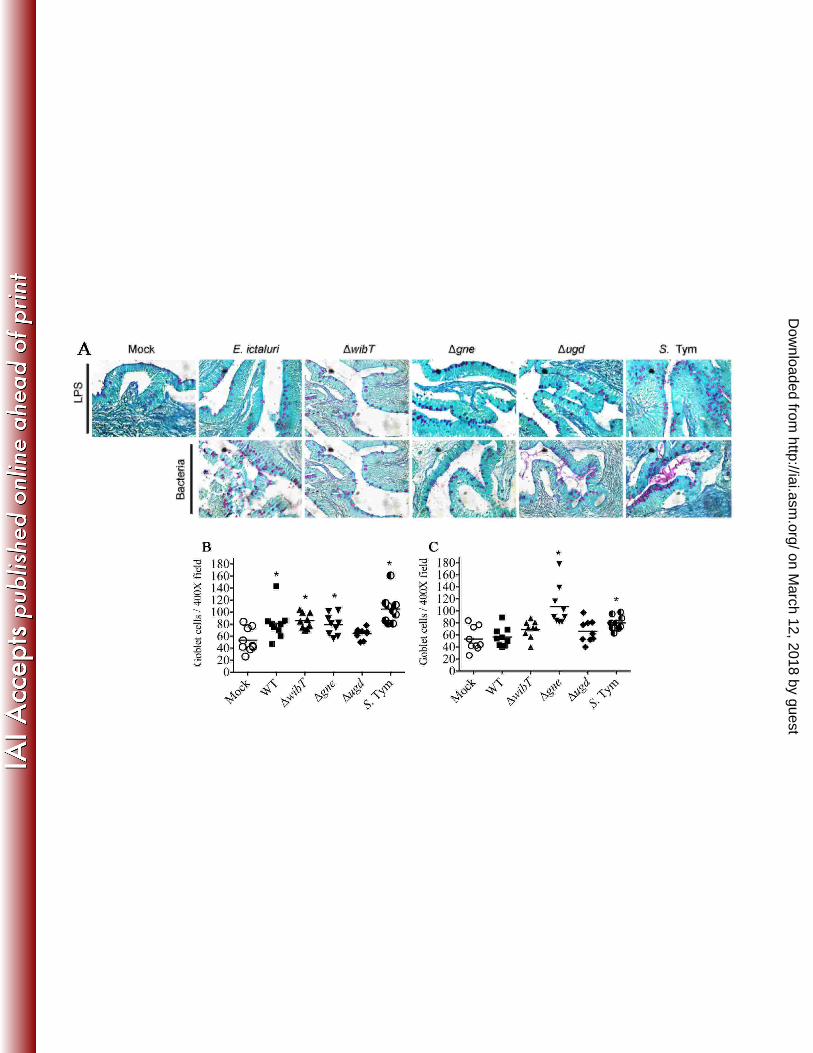
nor inflammation (Figs. 4 and 5). We noted that purified LPS form E. ictaluri gne, ugd 278
and S. Typhimurium stimulates goblet cells and mucus secretion (Fig. 7). Both E. ictaluri 279
and S. Typhimurium were recovered from the intestinal fluids and tissue in similar levels. 280
E. ictaluri wild type and S. Typhimurium were able to grow in the intestinal milieu (Fig. 281
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
13
8A) and colonized intestinal epithelial tissues (Fig. 8B). 282
E. ictaluri wibT caused significant levels of fluid secretion (Fig. 3C) and 283
significant tissue damage, similar to E. ictaluri wild-type (Figs. 4 and 5). We noted that 284
E. ictaluri gne, ugd and S. Typhimurium stimulate goblet cells and mucus secretion 285
without casing tissue damage (Figs. 4, 5 and 7). E. ictaluri wibT titers increased one log 286
fold in the intestinal milieu (Fig. 8A) and colonized epithelial tissues similar to the E. 287
ictaluri wild type (Fig. 8B). E. ictaluri gne generated significant low fluid secretion, 288
causing a mild inflammation with low tissue damage (Figs. 3C, 5 and 6). E. ictaluri gne 289
grow in the intestinal fluids in similar levels than the E. ictaluri wild type (Fig. 8A). 290
However, E. ictaluri gne was recovered in higher numbers from the intestinal 291
epithelium than E. ictaluri wild type (Fig. 8B). These results are in concordance with the 292
low tissue damage observed (Figs. 4 and 5) where epithelial cells containing E. ictaluri 293
are not in the intestinal fluids like the loops infected with E. ictaluri wild type and wibT. 294
Previous studies suggested that E. ictaluri gne is attenuated and immune protective 295
when orally administered to the fish, triggering a mild intestinal inflammation without 296
significant tissue damage (19). These results are coincident with our results, suggesting 297
that the gne mutant is a good candidate for oral live attenuated vaccine development. E. 298
ictaluri ugd titers recovered form the intestinal fluids and intestinal tissue were similar 299
to E. ictaluri wild type (Figs. 8A and 8B). 300
301
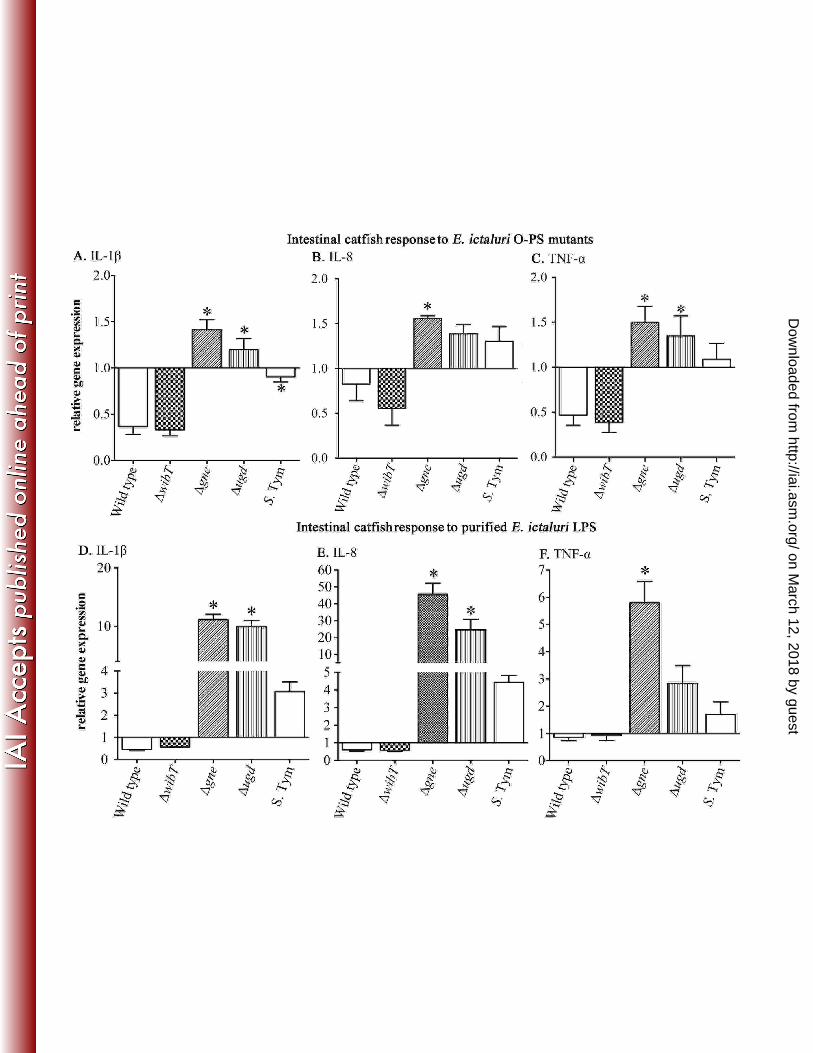
Cytokine expression. Cytokines play a major role in inflammatory responses. 302
Interleukins (IL) and tumor necrosis factors (TNFs) are a large group of cytokines 303
involved in innate immunity. In catfish only IL-1a, IL-1b, TNF- and IL-8 have been 304
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
14
identified (65,66,67). IL-1 plays a pivotal role in early pro-inflammatory cytokines that 305
enable the fish to respond to infection and enhance immune response induced by vaccines 306
(68). Also, it has been reported that in salmonid species IL-1, TNF- are induced in 307
presence of LPS and Gram-negative bacteria (69,70), triggering synthesis of IL-8 (71). 308
However, these studies focused on internal tissues like, spleen, liver and head kidney and 309
few studies have focused on the fish gut. Recently, we found that orally inoculated catfish 310
with E. ictaluri LPS O-PS mutants trigger lower inflammatory symptoms in contrast to 311
the wild type inoculated catfish (19), suggesting that LPS plays a role in inflammation at 312
the intestinal level in fish. Thus, here we evaluated expression of IL-1a, TNF- and IL-313
8 in the intestinal tissues 6 h post inoculation. During qRT-PCR settings, we detected that 314
the primers described for IL-1b generate two bands, one with the described molecular 315
weight and other just with the same sizes of -actin amplification fragment. Therefore, 316
here IL-1b was not measured. We found that E. ictaluri wild type and wibT down 317
regulated IL-1a, IL-8 and TNF- in contrast to gne and ugd which up regulated these 318
cytokines (Figs. 9A-C). S. Typhimurium slightly down regulated IL-a in a similar 319
fashion than E. ictaluri wild type, but with a significant difference between the fish and 320
non-fish bacterial pathogen (Fig. 9C). Although, TNF- and IL-8 levels were slightly up 321
regulated in S. Typhimurium inoculated loops, no significant differences were observed 322
in contrast to E. ictaluri WT (Figs. 9B-C). These results suggest that E. ictaluri wild type 323
and wibT have the ability to inhibit early innate immunity detection. Results from the 324
loops inoculated with gne and ugd indicate that this inhibitory ability might be LPS 325
mediated. Therefore, catfish intestinal loops were injected with purified LPS. Similar 326
results, but with ~10-fold increase in gne and ugd LPS samples, were found in loops 327
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
15
injected with purified LPS (Figs. 9D-F). This suggests that E. ictaluri LPS play an innate 328
immunity inhibitory role during fish intestinal colonization. Also, indicates that E. 329
ictaluri gne and ugd LPS molecules have intestinal immunestimulatory activity in the 330
fish gut, but the structure of these molecules is required to better understand these 331
interactions. Although the significant fold differences between the whole bacteria and the 332
purified LPS are evident, the results in both conditions have the same pattern. This might 333
be due to the fact that in the entire bacteria the LPS is in the outer membrane, not totally 334
accessible to interact with its putative receptor in contrast to the purified LPS. 335
336
DISCUSSION 337
338
In mammals, the lipid A portion of the LPS acts as a toxin by over-stimulating the 339
TLR-4 innate immune signaling, which induces pathogenic inflammatory responses. The 340
LPS is recognized by the serum circulating protein LBP (72) that facilitates the transfer 341
of LPS to the co-stimulatory molecule CD14 (73) and then to the myeloid differentiation 342
protein 2 (MD2; also called LY96) (74). MD2 is associated with the Toll-like receptor 4 343
(TLR4) and specifically recognized the endotoxic lipid A molecule (75), triggering a 344
downstream signaling that involves several intracellular TIR domain-containing adaptors, 345
like MDy88 and TICAM (76,77). 346
On the other hand, it is well established that fish and amphibians are very resistant 347
to the toxic effects of LPS (14). Several reports suggest that fish do not respond to LPS 348
because of the lack of PLB, CD14, LY96 and TCAM2, essential components for the 349
TLR4 function (20). Until today, functional LBP, CD14, MD2 and TCAM2 molecules 350
have not been described in fish. Indeed, more evolutionary advanced pufferfish lack 351
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
16
TLR4 ortholog (78,79). Also, it is suggested that LBP, CD14, MD2 and TCAM2 have 352
recently arisen during vertebrate evolution, being limited only to higher vertebrates 353
(20,80). This is supported by the extant literature indicating that fish are resistant to LPS 354
toxicity (14,19,20,81). It is evident that lower vertebrates interact with much higher 355
bacterial load in their living environment than land vertebrates, especially trough the 356
mucosal tissues. Perhaps due to the environmental selective pressure, imposed by the 357
aquatic “bacterial soup”, fish evolution did not favor high sensitivity to LPS. 358
However, several reports indicate that fish macrophages detect and respond to 359
high doses of LPS (15,16,19,20,80), but the mechanisms of detention and response are 360
unknown. 361
In the context of bacterial pathogenesis, recently we observed that catfish orally 362
infected with E. ictaluri wild type presented diarrhea-like symptoms, excreting mucoid 363
feces with high E. ictaluri titers (104-10
5 CFU/ml of feces) (19). In contrast, catfish orally 364
inoculated with E. ictaluri wibT, gne, or ugd (O-PS mutants) did not present 365
diarrhea-like symptoms (19). These results suggested that fish respond to LPS at the gut 366
level influencing E. ictaluri infection. These observations prompted us to investigate the 367
role of E. ictaluri LPS during intestinal inflammation by using the catfish intestinal loop 368
model and comparing it to the rabbit intestinal loop model and S. Typhimurium LPS. 369
Intestinal ligated loops have been used since 1950s to investigate intestinal interaction 370
with bacterial pathogens and their virulence factors. As mentioned previously all these 371
studies have been done in mammals, including mice, rats, rabbits, and calves 372
(22,23,24,27). In contrast to mammals, little is known about fish intestinal interaction 373
with bacterial pathogen LPS. 374
We observed that E. ictaluri LPS has effects on fish intestinal inflammation, 375
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
17
which is O-PS depending in fish but not in mammals (Figs. 3, 4 and 5). For instance, we 376
determined that purified E. ictaluri wild-type LPS triggers a mild inflammation in rabbit 377
ligated ileal loops in contrast to S. Typhimurium LPS that caused massive tissue damage 378
and inflammation (Figs. 4 and 5). The opposite results were observed in catfish intestinal 379
loops inoculated with purified E. ictaluri wild-type LPS. Catfish ligated loops inoculated 380
with S. Typhimurium LPS did not show significant tissue damage and inflammation in 381
contrast with E. ictaluri LPS that caused evident inflammation and tissue damage (Figs. 382
4, 5 and 6). E. ictaluri wibT LPS caused inflammation in both rabbits and fish ligated 383
loops, indicating that the E. ictaluri wibT LPS molecule is recognized by the innate 384
immune systems of fish and mammals. E. ictaluri gne LPS trigger a reduced epithelial 385
damage in rabbits, but no tissue damage was detected in intestinal fish loops. Although, 386
the LPS profile of E. ictaluri wibT and gne are similar, their glycosil compositions are 387
different (Fig. 2), suggesting that the structure of the O-PS is relevant to bacteria-fish 388
intestinal epithelia interaction and pathogenesis. E. ictaluri ugd LPS did not cause 389
effects on rabbit ligated loops, but in fish trigger a mild inflammation with an epithelial 390
cell-cell junction disruption (Figs. 4 and 5). 391
Immune stimulants represent a promising tool in aquaculture for enhancing 392
disease and stress resistance in cultured fish. It has been shown that oral administration of 393
LPS prevents disease in fish (82,83). However, the mechanisms of this oral immune 394
stimulation are unknown. Here we determined that purified LPS from E. ictaluri gne 395
and ugd mutants trigger synthesis of pro-inflammatory cytokines like IL-1, IL-8 and 396
TNF- in contrast to LPS from E. ictaluri wild type and wibT (Fig. 9). 397
IL-1 plays a pivotal role in early pro-inflammatory cytokines that enable 398
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
18
organism to respond to infection. Also, IL-1 has the potential to enhance immune 399
response induced by vaccines (68) and recombinant IL-1 has been use as an immune 400
stimulant for vaccines in sheep (84,85), pigs (86), cattle (87) and sea bass (88,89). In 401
teleost, the administration of immunostimulants, such as -1,3 glucan and peptidoglycan, 402
helps to prevent infections through activation of phagocytes, such as neutrophils, 403
monocytes, and macrophages, suggesting that the activation of innate immunity in teleost 404
fish by immune stimulants is a useful method of disease prevention that can replace the 405
use of antibiotics. Recent studies showed that orally administered LPS form Pantoea 406
agglomerans has a preventive effect against infection in fish such as yellowtail, carp, and 407
ayu (82). Among the possible receptors for LPS in fish it has been suggested that 2-408
integrin could play a role in LPS recognition (90,91). 2 integrins are one of the most 409
abundant receptor found in macrophages and they transmit intracellular activation signal 410
through MAP kinases and NF-B (90,91). 2 integrins recognize the hydrophilic 411
carbohydrate moiety that is buried in the outer bacterial membrane, but not the 412
hydrophobic lipid A (92). Also, the concentrations of LPS required to activate integrin 2 413
mediated activation of NF-B are high (93,94). We observed that the response to LPS at 414
the gut level is O-PS dependent (Fig. 9), suggesting an interaction with integrin 2 415
receptors or another carbohydrate receptor. This idea is supported by previous 416
observations, where fish intravenously injected with LPS and fish macrophages 417
inoculated with LPS respond in an O-PS dependent fashion (20). We observed that LPS 418
derived from gne and ugd E. ictaluri form supramolecules or aggregates, which seems 419
to increase interaction with fish macrophages (20). This observation correlates with the 420
increased stimulation of IL-1, IL-8 and TNF synthesis in the intestinal loops inoculated 421
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
19
with gne and ugd LPS, suggesting their potential utility as immune stimulants for fish. 422
Currently, there is a need for effective orally delivered vaccines (95). Thus studies 423
about fish gut bacterial interaction become important to develop effective oral vaccines 424
for aquaculture. Recently, we determined that an E. ictaluri gne strain conferred 425
immune protection to orally immunized fish (19). In contrast to the rest of E. ictaluri LPS 426
mutants study here, only gne confers immune protection to the orally immunized fish 427
(19). We observed that E. ictaluri gne do not cause tissue damage and fluid secretion, 428
increasing its colonization and interaction with lymphoid intestinal cells, influencing 429
positively the immune response to E. ictaluri. Perhaps, the immune stimulatory properties 430
of E. ictauri gne LPS, combines to its increased colonization of intestinal mucosa, lower 431
capability to survive in catfish macrohages, resistance to the antimicrobial peptides and 432
motility (19) make of E. ictauri gne a good candidate to be use as live attenuated 433
vaccine for the catfish aquaculture industry. 434
Here we have developed a catfish intestinal loop model to study the bacterial 435
interaction with the fish intestinal epithelia. By using this model, we determined that the 436
response to LPS at the intestinal level could differ depending on the LPS molecule and 437
the host. For instance, E. ictaluri LPS did not case inflammation in rabbit, perhaps its 438
lipid A has a different structure. The fish intestinal loop model is a useful methodology to 439
study pathogenesis and intestinal immunology, but also could be applied to evaluate 440
feeding diets, probiotics and therapeutic drugs. 441
442
443
444
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
20
445
446
ACKNOWLEDGMENTS 447
448
This work was supported by USDA grant CRIS-ARZR-2009-01801 and 449
Comisión Nacional de Investigación Científica y Tecnológica (CONICYT), Gestión 450
Propia Fellowship, Chile. We thank Tim Corsi and Joanne Tetens for their assistance at 451
The Biodesign Institute, Arizona State University, animal facility, and to Erika Arch, 452
Paul Hartig and Tina Hartig for their logistic support. 453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
Santander et al., Infection and Immunity
21
469
470
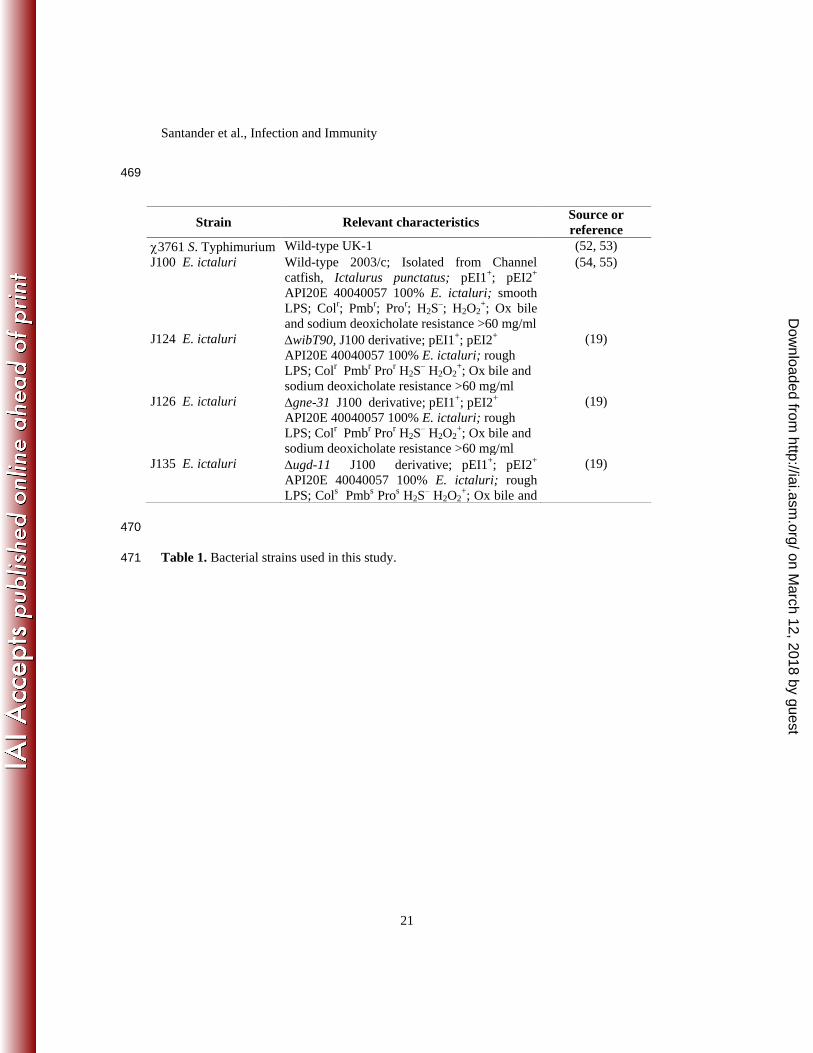
Table 1. Bacterial strains used in this study. 471
Strain Relevant characteristics Source or
reference
3761 S. Typhimurium Wild-type UK-1 (52, 53)
J100 E. ictaluri Wild-type 2003/c; Isolated from Channel
catfish, Ictalurus punctatus; pEI1+; pEI2
+
API20E 40040057 100% E. ictaluri; smooth
LPS; Colr; Pmb
r; Pro
r; H2S
–; H2O2
+; Ox bile
and sodium deoxicholate resistance >60 mg/ml
(54, 55)
J124 E. ictaluri wibT90, J100 derivative; pEI1+; pEI2
+
API20E 40040057 100% E. ictaluri; rough
LPS; Colr Pmb
r Pro
r H2S
– H2O2
+; Ox bile and
sodium deoxicholate resistance >60 mg/ml
(19)
J126 E. ictaluri gne-31 J100 derivative; pEI1+; pEI2
+
API20E 40040057 100% E. ictaluri; rough
LPS; Colr Pmb
r Pro
r H2S
– H2O2
+; Ox bile and
sodium deoxicholate resistance >60 mg/ml
(19)
J135 E. ictaluri ugd-11 J100 derivative; pEI1+; pEI2
+
API20E 40040057 100% E. ictaluri; rough
LPS; Cols Pmb
s Pro
s H2S
– H2O2
+; Ox bile and
(19)
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
22
472
473
474
475
476
477
478
479
480
481
482
483
Table 2. Primers used for Q Real time-PCR. 484
485
F: forward primer; R: reverse primer 486
487
sodium deoxicholate resistance >60 mg/ml
Gene Primer sequences Reference
β-actin
F AGAGAGAAATTGTCCGTGACATC
(67)
R CTCCGATCCAGACAGAGTATTTG
TNF-α F GGCCTCTACTTCGTCTAC
(66) R GCAGCAGCTTCTCGTCCAT
IL-1βa F CGGCAGATGTGACCTGCACA
(65) R CAGAGTAAAAGCCAGCAGAAG
IL-8 F CACCACGATGAAGGCTGCAACTC
(67) R TGTCCTTGGTTTCCTTCTGG
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
Santander et al., Infection and Immunity
23
488
489
490
491
492
493
494
495
496
497
498
499
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
24
REFERENCES 500
501
1. Abayneh, T., D. J. Colquhoun, and H. Sorum. 2013. Edwardsiella piscicida 502
sp. nov., a novel species pathogenic to fish. J Appl Microbiol 114:644-654. 503
2. Shoemaker, C. A., P. H. Klesius, C. R. Arias, and J. J. Evans. 2009. Uses of 504
modified live vaccines in the aquaculture. J World Aqua Soc 5:573e85. 505
3. Hanson, T. and D. Sites. 2012. Catfish database 2011. 506
http://www.aces.edu/anr/fish/aquaculture/files/2012/03/2011-catfish-report-507
whole1.pdf 508
4. Harvey, D. 2006. Aquaculture Outlook. USDA, ERS, LDP – AQS-23. 509
5. Santander, J. and R. Curtiss III. 2012. Presented at the AFRI/NRI Animal 510
Health and Walfare Annual Project Director Meeting, Chicago IL. 511
6. Shotts, E. B., V. S. Blazer, and W. D. Waltman. 1986. Pathogenesis of 512
experimental Edwardsiella ictaluri infections in channel catfish (Ictalurus 513
punctatus). Can J Fish Aquat Sci 43:36-42. 514
7. Baldwin, T. J., and J. C. Newton 1993. Pathogenesis of enteric septicemia of 515
channel catfish, caused by Edwardsiella ictaluri: bacteriologic and light and 516
electron microscopic findings. J Aquat Anim Health 5:189–198. 517
8. Thune, R. L., D. H. Fernandez, J. L. Benoit, M. Kelly-Smith, M. L. Rogge, N. 518
J. Booth, C. A. Landry, and R. A. Bologna. 2007. Signature-tagged 519
mutagenesis of Edwardsiella ictaluri identifies virulence-related genes, including 520
a salmonella pathogenicity island 2 class of type III secretion systems. Appl 521
Environ Microbiol 73:7934-7946. 522
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
25
9. Morrison, R. N., G. A. Cooper, B. F. Koop, M. L. Rise, A. R. Bridle, M. B. 523
Adams, and B. F. Nowak. 2006. Transcriptome profiling the gills of amoebic gill 524
disease (AGD)-affected Atlantic salmon (Salmo salar L.): a role for tumor 525
suppressor p53 in AGD pathogenesis? Physiol Genomics 26:15-34. 526
10. Poltorak, A., K. Peppel, and B. Beutler. 1994. Receptor-mediated label-transfer 527
assay (RELAY): a novel method for the detection of plasma tumor necrosis factor 528
at attomolar concentrations. J Immunol Methods 169:93-99. 529
11. Hoshino, K., O. Takeuchi, T. Kawai, H. Sanjo, T. Ogawa, Y. Takeda, K. 530
Takeda, and S. Akira. 1999. Cutting edge: Toll-like receptor 4 (TLR4)-deficient 531
mice are hyporesponsive to lipopolysaccharide: evidence for TLR4 as the LPS 532
gene product. J Immunol 162:3749-3752. 533
12. Qureshi, S. T., L. Lariviere, G. Leveque, S. Clermont, K. J. Moore, P. Gros, 534
and D. Malo. 1999. Endotoxin-tolerant mice have mutations in Toll-like receptor 535
4 (Tlr4). J Exp Med 189:615-625. 536
13. Raetz, C. R., and C. Whitfield. 2002. Lipopolysaccharide endotoxins. Annu Rev 537
Biochem 71:635-700. 538
14. Berczi, I., L. Bertok, and T. Bereznai. 1966. Comparative studies on the toxicity 539
of Escherichia coli lipopolysaccharide endotoxin in various animal species. Can J 540
Microbiol 12:1070-1071. 541
15. MacKenzie, S., J. V. Planas, and F. W. Goetz. 2003. LPS-stimulated expression 542
of a tumor necrosis factor-alpha mRNA in primary trout monocytes and in vitro 543
differentiated macrophages. Dev Comp Immunol 27:393-400. 544
16. Pelegrin, P., J. Garcia-Castillo, V. Mulero, and J. Meseguer. 2001. 545
Interleukin-1beta isolated from a marine fish reveals up-regulated expression in 546
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
26
macrophages following activation with lipopolysaccharide and lymphokines. 547
Cytokine 16:67-72. 548
17. Lawrence, M., M. Banes, and M. Williams. 2001. Phenotype and virulence of a 549
transposon- derived lipopolysaccharide O side-chain mutant of Edwardsiella 550
ictaluri. J Aquat Anim Health 13:291-299. 551
18. Lawrence, M. L., M. M. Banes, P. Azadi, and B. Y. Reeks. 2003. The 552
Edwardsiella ictaluri O polysaccharide biosynthesis gene cluster and the role of 553
O polysaccharide in resistance to normal catfish serum and catfish neutrophils. 554
Microbiol 149:1409-1421. 555
19. Santander, J., T. Martin, A. Loh, C. Pohlenz, D. M. Gatlin, and R. Curtiss 556
III. 2013. Mechanisms of intrinsic resistance to antimicrobial peptides of 557
Edwardsiella ictaluri and its influence on fish gut inflammation and virulence. 558
Microbiol 159:1471-1486. 559
20. Iliev, D. B., J. C. Roach, S. Mackenzie, J. V. Planas, and F. W. Goetz. 2005. 560
Endotoxin recognition: in fish or not in fish? FEBS Lett 579:6519-6528. 561
21. Everest, P., J. Ketley, S. Hardy, G. Douce, S. Khan, J. Shea, D. Holden, D. 562
Maskell, and G. Dougan. 1999. Evaluation of Salmonella typhimurium mutants 563
in a model of experimental gastroenteritis. Infect Immun 67:2815-2821. 564
22. Kong, Q., D. A. Six, Q. Liu, L. Gu, S. Wang, P. Alamuri, C. R. Raetz, and R. 565
Curtiss, 3rd. 2012. Phosphate groups of lipid A are essential for Salmonella 566
enterica serovar Typhimurium virulence and affect innate and adaptive immunity. 567
Infect Immun 80:3215-3224. 568
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
27
23. Menge, C., M. Blessenohl, T. Eisenberg, I. Stamm, and G. Baljer. 2004. 569
Bovine ileal intraepithelial lymphocytes represent target cells for Shiga toxin 1 570
from Escherichia coli. Infect Immun 72:1896-1905. 571
24. Menge, C., I. Stamm, P. M. Van Diemen, P. Sopp, G. Baljer, T. S. Wallis, and 572
M. P. Stevens. 2004. Phenotypic and functional characterization of intraepithelial 573
lymphocytes in a bovine ligated intestinal loop model of enterohaemorrhagic 574
Escherichia coli infection. J Med Microbiol 53:573-579. 575
25. De, S. N., M. L. Ghose, and A. Sen. 1960. Activities of bacteria-free 576
preparations from Vibrio cholerae. J Pathol Bacteriol 79:373-380. 577
26. Syngkon, A., S. Elluri, H. Koley, P. K. Rompikuntal, D. R. Saha, M. K. 578
Chakrabarti, R. K. Bhadra, S. N. Wai, and A. Pal. 2010. Studies on a novel 579
serine protease of a hapA prtV Vibrio cholerae O1 strain and its role in 580
hemorrhagic response in the rabbit ileal loop model. PLoS One 5. 581
27. De, S. N., and D. N. Chatterje. 1953. An experimental study of the mechanism 582
of action of Vibriod cholerae on the intestinal mucous membrane. J Pathol 583
Bacteriol 66:559-562. 584
28. Burrows, W., and G. M. Musteikis. 1966. Cholera infection and toxin in the 585
rabbit ileal loop. J Infect Dis 116:183-190. 586
29. Kohler, E. M. 1971. Enterotoxic activity of whole cell lysates of Escherichia coli 587
in young pigs. Am J Vet Res 32:731-737. 588
30. Moon, H. W., S. C. Whipp, G. W. Engstrom, and A. L. Baetz. 1970. Response 589
of the rabbit ileal loop to cell-free products from Escherichia coli 590
enteropathogenic for swine. J Infect Dis 121:182-187. 591
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
28
31. Smith, H. W., and S. Halls. 1967. Observations by the ligated intestinal segment 592
and oral inoculation methods on Escherichia coli infections in pigs, calves, lambs 593
and rabbits. J Pathol Bacteriol 93:499-529. 594
32. Fromme, I., and S. Schlecht. 1973. [Influence of aeration conditions in 595
fermentation cultures on the chemical composition of Salmonella 596
lipopolysaccharides (author's transl)]. Zentralbl Bakteriol Orig A 224:331-344 597
33. Wallis, T. S., S. M. Paulin, J. S. Plested, P. R. Watson, and P. W. Jones. 1995. 598
The Salmonella dublin virulence plasmid mediates systemic but not enteric phases 599
of salmonellosis in cattle. Infect Immun 63:2755-2761. 600
34. Taylor, J., M. P. Wilkins, and J. M. Payne. 1961. Relation of rabbit gut reaction 601
to enteropathogenic Escherichia coli. Br J Exp Pathol 42:43-52. 602
35. Kubota, Y., and P. V. Liu. 1971. An enterotoxin of Pseudomonas aeruginosa. J 603
Infect Dis 123:97-98. 604
36. Duncan, C. L., H. Sugiyama, and D. H. Strong. 1968. Rabbit ileal loop 605
response to strains of Clostridium perfringens. J Bacteriol 95:1560-1566. 606
37. Bhattacharya, S., A. K. Bose, and A. K. Ghosh. 1971. Permeability and 607
enterotoxic factors of nonagglutinable vibrios Vibrio alcaligenes and Vibrio 608
parahaemolyticus. Appl Microbiol 22:1159-1161. 609
38. Spira, W. M., and J. M. Goepfert. 1972. Bacillus cereus-induced fluid 610
accumulation in rabbit ileal loops. Appl Microbiol 24:341-348. 611
39. Frost, A. J., A. P. Bland, and T. S. Wallis. 1997. The early dynamic response of 612
the calf ileal epithelium to Salmonella typhimurium. Vet Pathol 34:369-386. 613
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
29
40. Santos, R. L., S. Zhang, R. M. Tsolis, A. J. Baumler, and L. G. Adams. 2002. 614
Morphologic and molecular characterization of Salmonella typhimurium infection 615
in neonatal calves. Vet Pathol 39:200-215. 616
41. Wallis, T. S., R. J. Hawker, D. C. Candy, G. M. Qi, G. J. Clarke, K. J. 617
Worton, M. P. Osborne, and J. Stephen. 1989. Quantification of the leucocyte 618
influx into rabbit ileal loops induced by strains of Salmonella typhimurium of 619
different virulence. J Med Microbiol 30:149-156. 620
42. Vinogradov, E., L. Nossova, M. B. Perry, and W. W. Kay. 2005. The structure 621
of the antigenic O-polysaccharide of the lipopolysaccharide of Edwardsiella 622
ictaluri strain MT104. Carbohydr Res 340:1509-1513. 623
43. Guttman, J. A., and B. B. Finlay. 2009. Tight junctions as targets of infectious 624
agents. Biochim Biophys Acta 1788:832-841. 625
44. Niklasson, L., H. Sundh, F. Fridell, G. L. Taranger, and K. Sundell. 2011. 626
Disturbance of the intestinal mucosal immune system of farmed Atlantic salmon 627
(Salmo salar), in response to long-term hypoxic conditions. Fish Shellfish 628
Immunol 31:1072-1080. 629
45. Komatsu, K., S. Tsutsui, K. Hino, K. Araki, Y. Yoshiura, A. Yamamoto, O. 630
Nakamura, and T. Watanabe. 2009. Expression profiles of cytokines released in 631
intestinal epithelial cells of the rainbow trout, Oncorhynchus mykiss, in response 632
to bacterial infection. Dev Comp Immunol 33:499-506. 633
46. Rombout, J. H., S. J. van der Tuin, G. Yang, N. Schopman, A. Mroczek, T. 634
Hermsen, and J. J. Taverne-Thiele. 2008. Expression of the polymeric 635
Immunoglobulin Receptor (pIgR) in mucosal tissues of common carp (Cyprinus 636
carpio L.). Fish Shellfish Immunol 24:620-628. 637
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
30
47. Rajan, B., J. M. Fernandes, C. M. Caipang, V. Kiron, J. H. Rombout, and M. 638
F. Brinchmann. 2011. Proteome reference map of the skin mucus of Atlantic cod 639
(Gadus morhua) revealing immune competent molecules. Fish Shellfish Immunol 640
31:224-231. 641
48. Palaksha, K. J., G. W. Shin, Y. R. Kim, and T. S. Jung. 2008. Evaluation of 642
non-specific immune components from the skin mucus of olive flounder 643
(Paralichthys olivaceus). Fish Shellfish Immunol 24:479-488. 644
49. Li, C., Y. Zhang, R. Wang, J. Lu, S. Nandi, S. Mohanty, J. Terhune, Z. Liu, 645
and E. Peatman. 2012. RNA-seq analysis of mucosal immune responses reveals 646
signatures of intestinal barrier disruption and pathogen entry following 647
Edwardsiella ictaluri infection in channel catfish, Ictalurus punctatus. Fish 648
Shellfish Immunol 32:816-827. 649
50. Hebert, P., A. J. Ainsworth, and B. Boyd. 2002. Histological enzyme and flow 650
cytometric analysis of channel catfish intestinal tract immune cells. Dev Comp 651
Immunol 26:53-62. 652
51. Skirpstunas, R. T., and T. J. Baldwin. 2002. Edwardsiella ictaluri invasion of 653
IEC-6, Henle 407, fathead minnow and channel catfish enteric epithelial cells. Dis 654
Aquat Organ 51:161-167. 655
52. Hassan, J. O., and R. Curtiss, 3rd. 1990. Control of colonization by virulent 656
Salmonella typhimurium by oral immunization of chickens with avirulent delta 657
cya delta crp S. typhimurium. Res Microbiol 141:839-850. 658
53. Luo, Y., Q. Kong, J. Yang, G. Golden, S. Y. Wanda, R. V. Jensen, P. B. 659
Ernst, and R. Curtiss, 3rd. 2012. Complete genome sequence of the universal 660
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
31
killer Salmonella enterica Serovar Typhimurium UK-1 (ATCC 68169). J 661
Bacteriol 193:4035-4036. 662
54. Santander, J., Xin, W., Yang, Z. and Curtiss, R., III. 2010. The aspartate-663
semialdehyde dehydrogenase of Edwardsiella ictaluri and its use as balanced-664
lethal system in fish vaccinology. Plos One 5: e15944. 665
55. Petrie-Hanson, L., C. L. Romano, R. B. Mackey, P. Khosravi, C. M. Hohn, 666
and C. R. Boyle. 2007. Evaluation of zebrafish Danio rerio as a model for enteric 667
septicemia of catfish (ESC). J Aquat Anim Health. 19:151-158. 668
56. Bertani, G. 1951. Studies on lysogenesis. I. The mode of phage liberation by 669
lysogenic Escherichia coli. J Bacteriol 62:293-300. 670
57. Yi, E. C., and M. Hackett. 2000. Rapid isolation method for lipopolysaccharide 671
and lipid A from gram-negative bacteria. Analyst 125:651-656. 672
58. Hitchcock, P. J., and T. M. Brown. 1983. Morphological heterogeneity among 673
Salmonella lipopolysaccharide chemotypes in silver-stained polyacrylamide gels. 674
J Bacteriol 154:269-277. 675
59. Tsai, C. M., and C. E. Frasch. 1982. A sensitive silver stain for detecting 676
lipopolysaccharides in polyacrylamide gels. Anal Biochem 119:115-119. 677
60. Santander, J., G. Golden, S. Y. Wanda, and R. Curtiss III. 2012. The Fur 678
Regulated Iron Uptake System of Edwardsiella ictaluri and its Influence on 679
Pathogenesis and Immunogenicity in the Catfish Host. Infect Immun 80:2689-680
2703. 681
61. Popovic, N. T., I. Strunjak-Perovic, R. Coz-Rakovac, J. Barisic, M. Jadan, A. 682
Persin, Berakovic, R. Sauerborn Klobucar. 2012. Tricaine methane-sulfonate 683
(MS-222) application in fish anaesthesia. J Appl Ichthyol 28:553-564. 684
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
32
62. Brown, L. A. 1987. Recirculation anaesthesia for laboratory fish. Lab Anim 685
21:210-215. 686
63. Harms, C. A. 2005. Surgery in fish research: common procedures and 687
postoperative care. Lab Anim (NY) 34:28-34. 688
64. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expression 689
data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. 690
Methods 25:402-408. 691
65. Wang, Y., Q. Wang, P. Baoprasertkul, E. Peatman, and Z. Liu. 2006. 692
Genomic organization, gene duplication, and expression analysis of interleukin-693
1beta in channel catfish (Ictalurus punctatus). Mol Immunol 43:1653-1664. 694
66. Zou, J., C. J. Secombes, S. Long, N. Miller, L. W. Clem, and V. G. Chinchar. 695
2003. Molecular identification and expression analysis of tumor necrosis factor in 696
channel catfish (Ictalurus punctatus). Dev Comp Immunol 27:845-858. 697
67. Chen, L., C. He, P. Baoprasertkul, P. Xu, P. Li, J. Serapion, G. Waldbieser, 698
W. Wolters, and Z. Liu. 2005. Analysis of a catfish gene resembling interleukin-699
8: cDNA cloning, gene structure, and expression after infection with Edwardsiella 700
ictaluri. Dev Comp Immunol 29:135-142. 701
68. Nash, P. V., and A. M. Mastro. 1993. Activation of primary lymphocytes 702
requires prolonged lectin stimulation. J Leukoc Biol 53:73-78. 703
69. Peddie, S., J. Zou, and C. J. Secombes. 2002. Immunostimulation in the 704
rainbow trout (Oncorhynchus mykiss) following intraperitoneal administration of 705
Ergosan. Vet Immunol Immunopathol 86:101-113. 706
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
33
70. Secombes, C. J., T. Wang, S. Hong, S. Peddie, M. Crampe, K. J. Laing, C. 707
Cunningham, and J. Zou. 2001. Cytokines and innate immunity of fish. Dev 708
Comp Immunol 25:713-723. 709
71. Wang, Z., X. Liu, A. Dacanay, B. A. Harrison, M. Fast, D. J. Colquhoun, V. 710
Lund, L. L. Brown, J. J. Li, and E. Altman. 2007. Carbohydrate analysis and 711
serological classification of typical and atypical isolates of Aeromonas 712
salmonicida: A rationale for the lipopolysaccharide-based classification of A-713
salmonicida. Fish & Shellfish Immunology 23:1095-1106. 714
72. Mathison, J. C., P. S. Tobias, E. Wolfson, and R. J. Ulevitch. 1992. Plasma 715
lipopolysaccharide (LPS)-binding protein. A key component in macrophage 716
recognition of gram-negative LPS. J Immunol 149:200-206. 717
73. Tobias, P. S., K. Soldau, J. A. Gegner, D. Mintz, and R. J. Ulevitch. 1995. 718
Lipopolysaccharide binding protein-mediated complexation of lipopolysaccharide 719
with soluble CD14. J Biol Chem 270:10482-10488. 720
74. Gioannini, T. L., A. Teghanemt, D. Zhang, E. N. Levis, and J. P. Weiss. 2005. 721
Monomeric endotoxin:protein complexes are essential for TLR4-dependent cell 722
activation. J Endotoxin Res 11:117-123. 723
75. Akashi, S., S. Saitoh, Y. Wakabayashi, T. Kikuchi, N. Takamura, Y. Nagai, 724
Y. Kusumoto, K. Fukase, S. Kusumoto, Y. Adachi, A. Kosugi, and K. 725
Miyake. 2003. Lipopolysaccharide interaction with cell surface Toll-like receptor 726
4-MD-2: higher affinity than that with MD-2 or CD14. J Exp Med 198:1035-727
1042. 728
76. O'Neill, L. A., and A. G. Bowie. 2007. The family of five: TIR-domain-729
containing adaptors in Toll-like receptor signalling. Nat Rev Immunol 7:353-364. 730
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
34
77. Vogel, S. N., and M. Fenton. 2003. Toll-like receptor 4 signalling: new 731
perspectives on a complex signal-transduction problem. Biochem Soc Trans 732
31:664-668. 733
78. Oshiumi, H., T. Tsujita, K. Shida, M. Matsumoto, K. Ikeo, and T. Seya. 2003. 734
Prediction of the prototype of the human Toll-like receptor gene family from the 735
pufferfish, Fugu rubripes, genome. Immunogenetics 54:791-800. 736
79. Jault, C., L. Pichon, and J. Chluba. 2004. Toll-like receptor gene family and 737
TIR-domain adapters in Danio rerio. Mol Immunol 40:759-771. 738
80. Sepulcre, M. P., F. Alcaraz-Perez, A. Lopez-Munoz, F. J. Roca, J. Meseguer, 739
M. L. Cayuela, and V. Mulero. 2009. Evolution of lipopolysaccharide (LPS) 740
recognition and signaling: fish TLR4 does not recognize LPS and negatively 741
regulates NF-kappaB activation. J Immunol 182:1836-1845. 742
81. Swain, P., S. K. Nayak, P. K. Nanda, and S. Dash. 2008. Biological effects of 743
bacterial lipopolysaccharide (endotoxin) in fish: a review. Fish Shellfish Immunol 744
25:191-201 745
82. Kohchi, C., H. Inagawa, T. Nishizawa, T. Yamaguchi, S. Nagai, and G. Soma. 746
2006. Applications of lipopolysaccharide derived from Pantoea agglomerans (IP-747
PA1) for health care based on macrophage network theory. J Biosci Bioeng 748
102:485-496. 749
83. Kadowaki, T., Y. Yasui, O. Nishimiya, Y. Takahashi, C. Kohchi, G. Soma, 750
and H. Inagawa. 2013. Orally administered LPS enhances head kidney 751
macrophage activation with down-regulation of IL-6 in common carp (Cyprinus 752
carpio). Fish Shellfish Immunol 34:1569-1575. 753
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
35
84. Elhay, M. J., and P. Andersen. 1997. Immunological requirements for a subunit 754
vaccine against tuberculosis. Immunol Cell Biol 75:595-603. 755
85. Lofthouse, S. A., A. E. Andrews, G. J. Barcham, and A. D. Nash. 1995. 756
Parameters related to the application of recombinant ovine interleukin-1 beta as 757
an adjuvant. Vaccine 13:1277-1287. 758
86. Blecha, F., D. N. Reddy, C. G. Chitko-McKown, D. S. McVey, M. M. 759
Chengappa, R. D. Goodband, and J. L. Nelssen. 1995. Influence of 760
recombinant bovine interleukin-1 beta and interleukin-2 in pigs vaccinated and 761
challenged with Streptococcus suis. Vet Immunol Immunopathol 44:329-346. 762
87. Reddy, D. N., P. G. Reddy, H. C. Minocha, B. W. Fenwick, P. E. Baker, W. 763
C. Davis, and F. Blecha. 1990. Adjuvanticity of recombinant bovine interleukin-764
1 beta: influence on immunity, infection, and latency in a bovine herpesvirus-1 765
infection. Lymphokine Res 9:295-307. 766
88. Buonocore, F., M. Mazzini, M. Forlenza, E. Randelli, C. J. Secombes, J. Zou, 767
and G. Scapigliati. 2004. Expression in Escherchia coli and purification of sea 768
bass (Dicentrarchus labrax) interleukin 1beta, a possible immunoadjuvant in 769
aquaculture. Mar Biotechnol (NY) 6:53-59. 770
89. Buonocore, F., M. Forlenza, E. Randelli, S. Benedetti, P. Bossu, S. Meloni, C. 771
J. Secombes, M. Mazzini, and G. Scapigliati. 2005. Biological activity of sea 772
bass (Dicentrarchus labrax L.) recombinant interleukin-1beta. Mar Biotechnol 773
(NY) 7:609-617. 774
90. Ingalls, R. R., and D. T. Golenbock. 1995. CD11c/CD18, a transmembrane 775
signaling receptor for lipopolysaccharide. J Exp Med 181:1473-1479. 776
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
36
91. Ingalls, R. R., M. A. Arnaout, R. L. Delude, S. Flaherty, R. Savedra, Jr., and 777
D. T. Golenbock. 1998. The CD11/CD18 integrins: characterization of three 778
novel LPS signaling receptors. Prog Clin Biol Res 397:107-117. 779
92. Chateau, M. T., and R. Caravano. 1997. The oxidative burst triggered by 780
Salmonella typhimurium in differentiated U937 cells requires complement and a 781
complete bacterial lipopolysaccharide. FEMS Immunol Med Microbiol 17:57-66. 782
93. Santos, D. R., J. B. Calixto, and G. E. Souza. 2003. Effect of a kinin B2 783
receptor antagonist on LPS- and cytokine-induced neutrophil migration in rats. Br 784
J Pharmacol 139:271-278. 785
94. Aurell, C. A., and A. O. Wistrom. 1998. Critical aggregation concentrations of 786
gram-negative bacterial lipopolysaccharides (LPS). Biochem Biophys Res 787
Commun 253:119-123. 788
95. Plant, K. P., and S. E. Lapatra. 2013. Advances in fish vaccine delivery. Dev 789
Comp Immunol 35:1256-1262. 790
791
792
793
794
795
796
797
798
799
800
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
37
FIGURE LEGENDS 801
802
Figure 1. Fish intestinal loops surgery process. A. Fish surgery platform. The arrows 803
indicate the water flow; B. Fish anesthesia application; C. Incision in the catfish to enter 804
the coelomic cavity; D. Catfish intestinal loops; E. Injection of intestinal loop; F. Suture 805
of the incision; G. Recovery bath (MS222 30 mg/l). 806
807
Figure 2. Phenotype of E. ictaluri O-PS mutants. A. Map of E. ictaluri oligo-808
polysaccharide genes; B. LPS profile of E. ictaluri O-PS mutants; C-E. Complementation 809
of O-PS mutants; F. Table LPS glycosyl composition analysis. N.D.: not detected; 810
*Values are expressed as mole per cent of total carbohydrate; Gal: galactose; Glu: 811
glucose; GalNAc: N-acetyl galactosamine; GlcNAc: N-acetyl glucosamine. Adapted 812
from Santander et al., 2013 (21). 813
814
Figure 3. Catfish intestine loops and fluid secretion. A. Rabbit loops 6 h post-815
inoculation with 100 g of LPS; B. Catfish loops 6 h post-inoculation with 100 g of 816
LPS; C. Catfish fluid secretion from the intestinal loops 6 h post-inoculation with 107 817
CFU/ ml of E. ictaluri; S. Typhimurium was used as control 108 CFU/ml. The number of 818
animals per group is 3. The experiment was repeated 3 times independently. The total of 819
animals used per group was 9. The error bars indicate the standard deviation. *: 820
significant difference versus Mock control (P<0.05). 821
822
Figure 4. Comparative intestinal loop histology 6 h post inoculation with 100 μg of 823
LPS or 107 CFU/dose of bacteria. Mock intestinal loops were injected with saline (NaCl 824
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
38
0.85%). The ileal loops were injected with 1 ml of the respective sample. These 825
experiments were repeated 2 to 3 times independently. 826
827
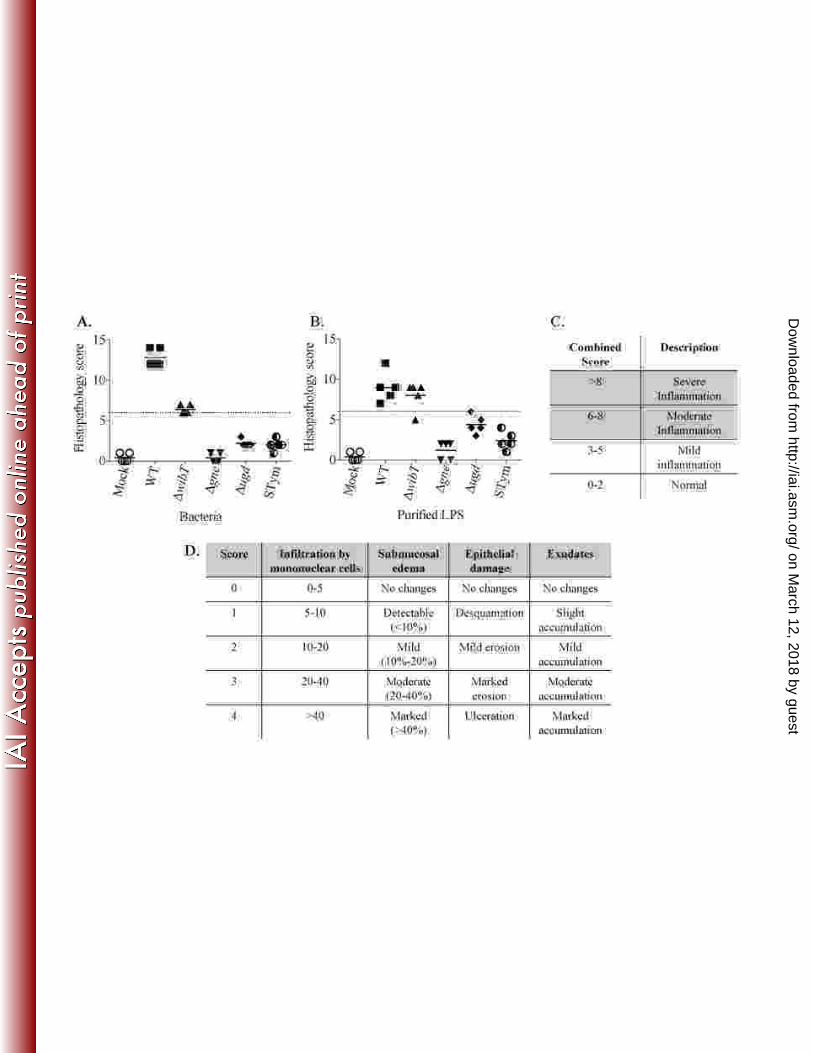
Figure 5. Histopathology score. A. Whole bacteria; B. Purified LPS; C. Combined 828
score; D. Scoring table. 829
830
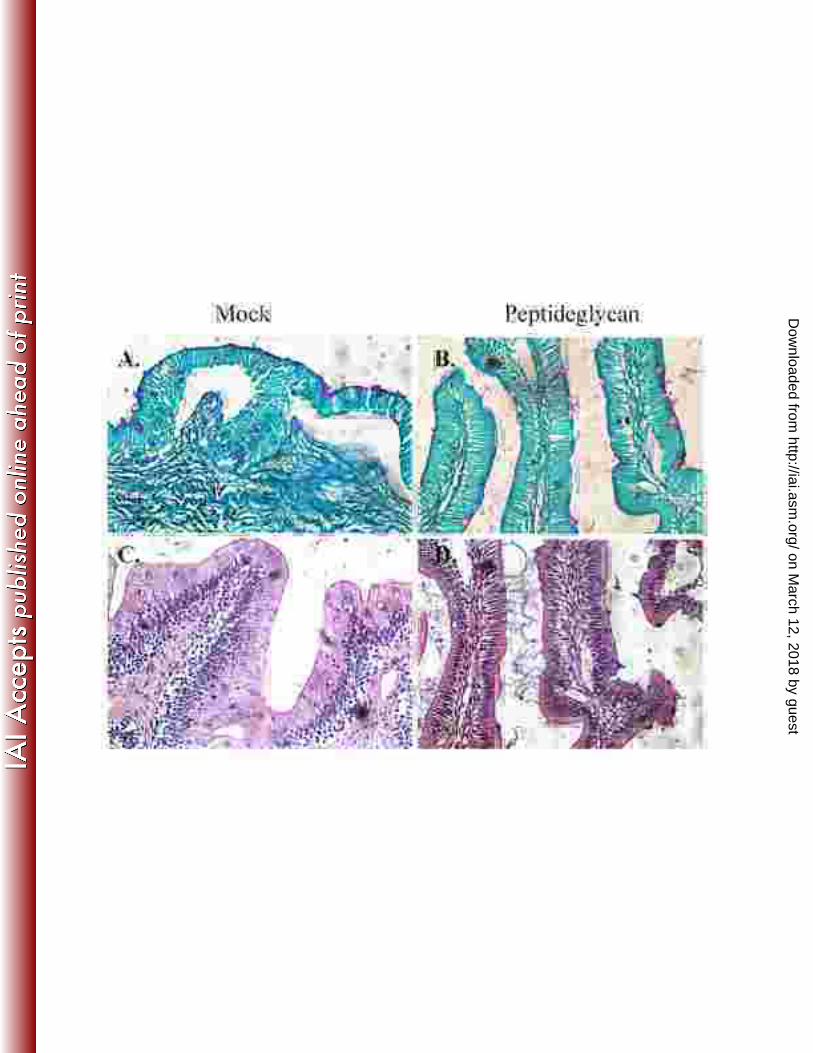
Figure 6. Catfish intestinal loop histology 6 h post inoculation with peptidoglycane. 831
A. Mock, PAS stained; B. Pepetidoglycan, PAS stained; C. Mock, HE stained; D. 832
Peptidoglycan, HE stained. A and C. Mock intestinal loops were injected with 1 ml of 833
saline (NaCl 0.85%). B and D. Intestinal loops injected with 1 ml of peptidoglycan (100 834
g). These experiments were repeated 2 to 3 times independently. 835
836
Figure 7. Comparative intestinal loop histology 6 h post inoculation with 100 μg of 837
LPS or 107 CFU/dose of bacteria. A. Catfish loop, PAS stained; B. Number of goblet 838
cells in a bacterial injected loop; C. Number of goblet cells in a LPS injected loop. Mock 839
intestinal loops were injected with saline (NaCl 0.85%). Catfish intestinal loops were 840
injected with 1 ml of the respective LPS sample (100 ug). These experiments were 841
repeated 3-4 times independently. Each dot represents a field; *: significant difference 842
(P<0.05). 843
844
Figure 8. E. ictaluri colonization ratio in intestinal loop histology 6 h post-845
inoculation with 107
CFU/dose of bacteria. A. Fluid colonization; B. Tissue 846
colonization; *: significant difference versus wild type (P<0.05). Each dot represents an 847
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Santander et al., Infection and Immunity
39
independent experiment. Total of 9 animals were used. The error bars indicate the 848
standard deviation. 849
850
Figure 9. qRT-PCT of intestinal catfish cytokines induced by E. ictaluri LPS. A-C. 851
Intestinal response to E. ictaluri O-PS mutants. The intestinal loops were inoculated with 852
107 CFU of each mutant; D-F. Intestinal response to purified E. ictaluri LPS. The 853
intestinal loops were inoculated with 100 g of LPS from each mutant; The experiment 854
was repeated 2 times independently. The total of animals used per group was 6. The error 855
bars indicate the standard deviation; *:significant difference versus wild type. 856
on March 12, 2018 by guest
http://iai.asm.org/
Dow
nloaded from





Related Documents











