Research Plan On Halophilic Enzymes and Their Biotechnological Applications Submitted by Under the Supervision of Sumit Kumar Dr. S. K. Khare (2007CYZ8227) DEPARTMENT OF CHEMISTRY INDIAN INSTITUTE OF TECHNOLOGY, DELHI

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 1/27
Research Plan
On
Halophilic Enzymes
and
Their Biotechnological Applications
Submitted by Under the Supervisionof
Sumit Kumar Dr. S. K. Khare
(2007CYZ8227)
DEPARTMENT OF CHEMISTRY
INDIAN INSTITUTE OF TECHNOLOGY, DELHI

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 2/27
Research Plan
Submitted by: Sumit Kumar
(2007CYZ8227)
Registered Topic: Halophilic Enzyme and
Their Biotechnological
Applications
Ph.D. Supervisor: Dr. S. K. Khare
Course Work: Completed 12 Credits
CGPA: 9.5/10

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 3/27
Introduction
Hydrolases, especially proteases, lipases and amylases are the largest sellingindustrial enzymes. These are widely used in detergent formulations and
peptide/esters/oligosaccharide synthesis. Their demand is predicted to increase by
4-5 folds in coming years. In question, however, is the stability of these enzymesin extreme conditions, such as high salt, surfactants and organic solvents.
Enzymes in halophiles and haloalkaliphiles have evolved to function in highly
saline and alkaline conditions. These are best suited to function under suchconditions.
A systematic investigation to screen the halophilic diversity that exists in the vast
saline habitats of our country is needed for obtaining novel enzyme preparations.
It is proposed to work on above lines with following overall objectives:
1. Studies on microbial diversity of different saline habitat of India with respect to
their morphological, biochemical and enzymatic profile.
2. Screening of lipolytic activity in selected halophilic isolates and selection of
potential lipase producers.
3. Production, purification and characterization of potential lipases especially with
respect to their catalysis in high salt/ solvent medium.
4. Identification of halophilic lipase genes, cloning, overexpression, structure andfunction analysis.
5. Lipase application in high salt/ solvent system.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 4/27
Review of literature
Halophiles
Halophiles have been studied from saline soils, mud and salt piles in hypersalineenvironment of California, Nevada, Great salt lake and Utah, USA; Soda lake in
Kenya; Japanese sea; inland and marine saltern of France, Canada, Germany, UK
and many other countries. Among these Halococcus and Halobacterium are prominently found halophiles. These have been extensively studied for structural
and genetic diversity and some of the species, such as, Halobacterium halobium,
Halomonas halophiles and Halocella cellulytica are well characterized (Anton et
al., 2000, Vreeland et al., 2000)
Little is known about the enzymes from these halophiles. Amongst, few reports
that have been published in this area, a cellulase complex have been purified from Halocella cellulytica which grows at 2.6 M NaCl. The half life of enzyme was
found to be 68 minutes at 50°C. A serine protease from Archbacterium strain 172
P1 is also purified and characterized. The purification to homogeneity in this case
was achieved by ammonium sulfate precipitation followed by chromatography on butyl Toyopearl 650-C. The enzyme had a Molecular weight of 44,000-46,000. It
had pH optima at 10.7, temperature optima 75-80°C and reported to be stable at
high NaCl concentration.
The biotechnological applications of halophiles and their enzymes are lessexplored. The understanding of enzymes from halophilic and haloalkaliphilic
organisms is still hazy. Their uses in hypersaline waste treatment, enhanced oil
recovery and peptide synthesis are predicted to be promising. A cellulase from a
halophilic bacterium has been successfully used for degradation of cellulose inhypersaline waste at US Waste Isolation Pilot Plant (Rodriguez-Valera, 1992). In
another interesting application, an extracellular protease from Halobacterium
halobium was used for peptide synthesis in aqueous organic system (Kim andDordick, 1997). The esterification activity of enzyme is found to be 80 fold higher
in DMSO medium. This unique behavior of halophilic enzymes in organic media
may have advantage in organic synthesis.
Structure and function analysis of halophilic enzyme gives insight in proteinfolding under a combination of extreme conditions (Favilla et al., 1997). Some of
the enzymes from halophilic and haloalkaliphilic organisms are extremely
resistant to chemical denaturation, a feature which would attract several novelapplications besides providing clues on the protein stabilities. Investigations on
these aspects would be quite interesting to pursue.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 5/27
Lipases
Lipases (EC 3.1.1.3) catalyse both the hydrolysis and esterification reactions (Fig.1). These reactions usually proceed with high regio- and/or enantioselectivity. Thereasons for the enormous biotechnological potential of lipases include the facts
that they are (1) stable in organic solvents, (2) do not require cofactors, (3)
possess broad substrate specificity and (4) exhibit a high enantioselectivity(Jaeger and Reetz, 1998)
Figure 1
Their ability to accept wide range of substrate (lipids, sugar, alcohol and esters)
and to maintain activity and selectivity in organic solvents have enabled their
wide use as biocatalysts in food, detergent, pharmaceutical, leather, textile,cosmetic and paper industries (Houde et al ., 2004; Salameh and Wiegel, 2007).
Though a large number of microbes secrete lipase, only few are of industrialimportance. The production of lipases is greatly influenced by nutritional and
physico-chemical factors (Jaeger et al ., 1994; Kim et al ., 1996). For enhancement
of yields, process optimization is carried out by “one-at-a-time approach” or byResponse Surface Methodology (RSM).
Lipases have been purified and characterized from diverse microbial sources.However, no single technique or generic protocol can be defined to be the best.
The purification strategy varies from source to source.
Cell free supernatant obtained by filtration or centrifugation of fermentation brothcan be concentrated by means of ultrafiltration (Castro-Ochoa et al., 2005) or
precipitated using ammonium sulphate, acetone or ethanol (Ogino et al., 2000;Karadzic et al., 2006) or extracted with organic solvents (Hiol et al., 1999). The
precipitation step concentrates the enzyme and often purifies to about two to three
fold (Aires-Barros et al ., 1994). Such partially purified lipase preparations are aptfor use in detergent formulations.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 6/27
In general, single chromatographic step does not purify lipase to required level.
Combination of chromatographic steps works better. Ion-exchange
chromatography is the most common chromatographic method. The mostfrequently employed ion-exchangers are the diethylaminoethyl (DEAE) and
carboxymethyl (CM) cellulose (Gupta et al ., 2004).
Anion-exchange chromatography on Amberlite IRA 410 for B. licheniformis
MTCC 6824 resulted in 33.5 fold purification (Chakraborty and Raj, 2008). C.
antarctica lipase B has been purified by cation-exchange chromatography to 2.4fold (Trodler et al., 2008). P. aeruginosa lipase has been purified using anion
exchanger Q-Sepharose to 12.7 fold (Singh and Banerjee, 2007). DEAE-
Sepharose ion exchange chromatography was successful in purification of B.
cereus C71 lipase to 65 fold purification (Shaoxin et al ., 2007).
Lipases having large hydrophobic surfaces may be better purified by hydrophobic
interaction chromatography (Queiroz et al ., 2001). The use of hydrophobic
interaction chromatography has increased in the past few years. Various HICmatrices have been employed for the purification of microbial lipases e.g. octyl-
sepharose CL-4B for Pseudomonas sp. strain ATCC 21808 (Kordel et al ., 1991)and Bacillus sp. H257 (Imamura and Kitaura, 2000); Fractogel TSK-isobutyl 650
for P. aeruginosa YS-7 (Shabtai and Daya-Mishne, 1992); Butyl-Sepharose 4
Fast Flow for Rhizopus chinensis (Sun and Xu, 2008) and Butyl-toyopearl for P.
aeruginosa (Karadzic et al ., 2006).
Different affinity matrices/ supports have been used to purify lipases from various
microbial sources. Some of them are ConA-Sepharose for Pseudomonas sp. strainS5 lipase (Rahman et al ., 2005); oleic acid affinity column for Rhizopus delemar
lipase (Haas et al ., 1992) and hydroxyapitite for P. simplicissimum lipase (Sztajer
et al ., 1992) leading to 30, 10.3 and 56 fold purification respectively.
Cloning of lipase gene from Galactomyces geotrichum Y05 into pPIC9K and
overexpression in Pichia pastoris GS115 has been reported by Yan et al . (2007).Overexpression led to 10.4-fold higher production over the wild type strain. The
genes of organic solvent-tolerant LST-03 lipase (Lip9) and lipase-specific foldase
(Lif9) have been cloned and expressed in E. coli (pET system). The
overexpression of the lipase gene (lip9) was achieved upon using T7 promoter and deleting the signal peptide of the lipase. Under these conditions, the
overexpressed lipase accumulated in the form of inclusion bodies (Ogino et al .,
2007). Yet another lipase gene from Pseudomonas fluorescens JCM5963 has beencloned, sequenced and overexpressed as an N-terminus His-tag fusion protein in
E. coli. The recombinant lipase (rPFL) was purified to homogeneity by Ni-NTA
affinity chromatography and Sephacryl S-200 gel filtration chromatography(Zhang et al., 2009).
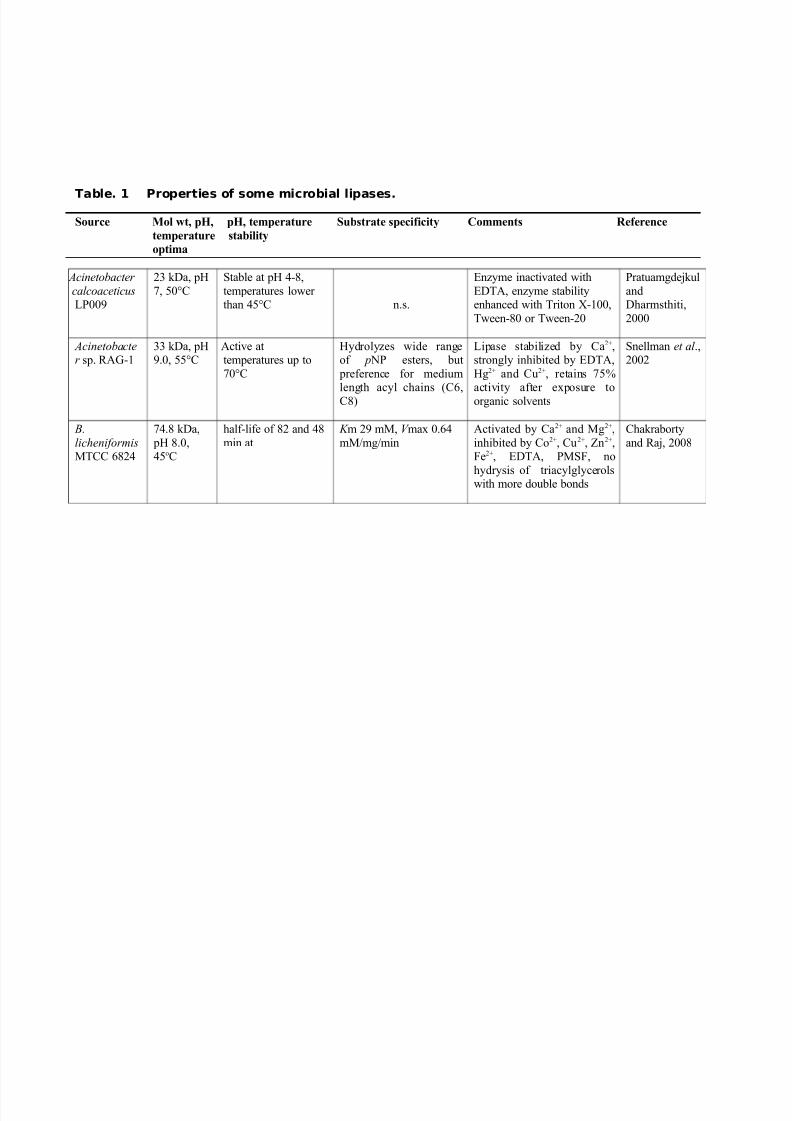
Properties of Lipases: Properties of various microbial lipases are summarized in
Table 1.
7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 7/27
Table. 1 Properties of some microbial lipases.
Source Mol wt, pH, pH, temperature Substrate specificity Comments Reference
temperature stability
optima
Acinetobacter
calcoaceticus LP009
23 kDa, pH
7, 50°C
Stable at pH 4-8,
temperatures lower than 45°C
n.s.
Enzyme inactivated with
EDTA, enzyme stabilityenhanced with Triton X-100,
Tween-80 or Tween-20
Pratuamgdejkul
andDharmsthiti,
2000
Acinetobacter sp. RAG-1
33 kDa, pH9.0, 55°C
Active attemperatures up to
70°C
Hydrolyzes wide rangeof p NP esters, but
preference for medium
length acyl chains (C6,C8)
Lipase stabilized by Ca2+,strongly inhibited by EDTA,
Hg2+ and Cu2+, retains 75%
activity after exposure toorganic solvents
Snellman et al .,2002
B.
licheniformisMTCC 6824
74.8 kDa,
pH 8.0,45oC
half-life of 82 and 48
min at
K m 29 mM, V max 0.64
mM/mg/min
Activated by Ca2+ and Mg2+,
inhibited by Co2+, Cu2+, Zn2+,Fe2+, EDTA, PMSF, no
hydrysis of triacylglycerolswith more double bonds
Chakraborty
and Raj, 2008
7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 8/27
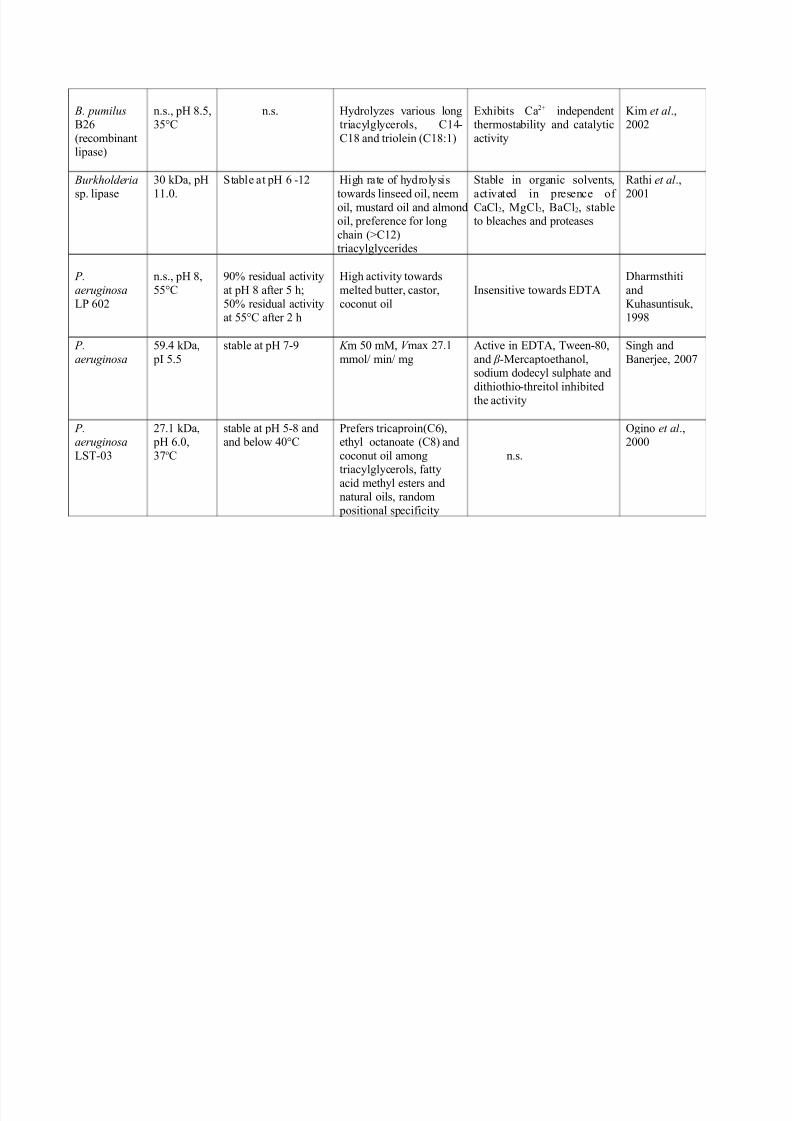
B. pumilus B26
(recombinantlipase)
n.s., pH 8.5,35°C
n.s. Hydrolyzes various longtriacylglycerols, C14-
C18 and triolein (C18:1)
Exhibits Ca2+ independentthermostability and catalytic
activity
Kim et al .,2002
Burkholderia
sp. lipase
30 kDa, pH
11.0.
Stable at pH 6 -12 High rate of hydrolysis
towards linseed oil, neemoil, mustard oil and almondoil, preference for long
chain (>C12)triacylglycerides
Stable in organic solvents,
activated in presence of CaCl2, MgCl2, BaCl2, stableto bleaches and proteases
Rathi et al .,
2001
P.
aeruginosa
LP 602
n.s., pH 8,
55°C
90% residual activity
at pH 8 after 5 h;
50% residual activityat 55°C after 2 h
High activity towards
melted butter, castor,
coconut oil
Insensitive towards EDTA
Dharmsthiti
and
Kuhasuntisuk,1998
P.
aeruginosa
59.4 kDa,
pI 5.5
stable at pH 7-9 K m 50 mM, V max 27.1
mmol/ min/ mg
Active in EDTA, Tween-80,
and β -Mercaptoethanol,sodium dodecyl sulphate and
dithiothio-threitol inhibitedthe activity
Singh and
Banerjee, 2007
P.aeruginosa
LST-03
27.1 kDa, pH 6.0,
37oC
stable at pH 5-8 andand below 40°C
Prefers tricaproin(C6),ethyl octanoate (C8) and
coconut oil amongtriacylglycerols, fatty
acid methyl esters andnatural oils, random
positional specificity
n.s.
Ogino et al .,2000

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 9/27
against triolein, high
solvent stability
P.
fluorescens
AK 102
33 kDa,pH 8.0-10.0,55°C, pI4.0
pH 4-10, stablebelow 50 ºC for 1h.
Broad specificity Enzyme stable in anionicsurfactants
Kojima et al.,1994
P.
fluorescens
NS2W
n.s., pH9.0, 55°C
Stable over pH 3-
11; stable at 0°C
with more than70% residualactivity for at least2 h
n.s. n.s.
Kulkarni andGadre, 2002
P. luteola n.s., 55°C Half-life of 116 minat 65°C
Preference formedium chainsaturated andunsaturated fattyacids
Inhibited by Sn and Zn Litthauer et
al., 2002
P.
mendocina
3121-1
62 kDa,pH 7.2-9.5, 50-65°C
Different fordifferentsubstrates
Hydrolyzes pNPbutyrate, Tween-80,olive oil
pH and temperaturekinetics, effect of variousmetal ions and EDTAdepended on the natureof the substrate.
Surinenaiteet al.,2002
Pseudomon
as sp. strain
S5
60 kDa, pH9.0, 45°C
stable at 45°C and pH6-9
highest activity in the presence of palm oil andtriolein among natural oilsand synthetic triglyceride,
Ca2+ and Mg2+ stimulated activ-ity, EDTA had no effect, highsolvent stability
Rahman et al .,2005

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 10/27
random positional specifi-city
Serratia
marcescens
52 kDa,pH 8.0-9.0, 37°C
n.s.Michelis-Mentenconstant 1.35 mM ontributyrin
n.s.
Abdou, 2003

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 11/27
Halophilic Lipase
Isolation and characterization of salt stable lipase from halophilic sources haveattracted considerable interest in recent years. Availability of such enzymes would
facilitate industrial processes that require activity at high salt concentrations.
Moderately halophilic bacteria adapted to live in a wide range of saltconcentrations (3–15% NaCl) constitute an interesting group of micro-organisms
that could be used as a source of such salt-adapted enzymes (Ventosa et al.,
1998).
It is quite possible that the structural features of halophilic enzymes that impart
stability at high salt concentrations will also confer stability in organic solvents
and at high temperatures (Adams et al., 1995).
Therefore, it seems promising to screen halophiles for lipases having novel/ new
biochemical properties.
In a recent study, Sanchez- Porro et al. (2003) isolated hydrolase-producing
moderately halophilic and halotolerant eubacteria from Spanish salterns. Only23% of the 892 strains produced extracellular lipolytic activity.
In the course of screening programme a novel, moderately halophilic bacterium(strain SM19T) that displays novel lipolytic activity has been isolated and
characterized. Strain SM19T is a Gram-negative rod that grows optimally in
culture media containing 7.5% NaCl. This is classified in the genus Marinobacter
with proposed name Marinobacter lipolyticus sp. nov.(Martı´n et al., 2003).
An extremely halophilic isolate, Salicola strain IC10, showing lipase and proteaseactivities has been marked for potential biotechnological applications. The
optimum growth conditions for this strain were 15-20% (w/v) NaCl, pH 8.0, and
370C. Its lipase showed highest activity against p-nitrophenyl-butyrate (deLourdes Moreno et al., 2009).
In the study related to archaea, total 118 halophilic strains were screened for
lipolytic activity. Eighteen were found positive on rhodamine agar plates. Highestlipase activities were found at pH 8, temperature 45-650C and NaCl 3.5-4 M.
These results indicate the presence of salt-dependent and thermostable lipases in
halophilic archaeal groups (Ozcan et al., 2009).
Fifty strains of moderately halophilic bacteria were isolated from various saltyenvironments in Iran. A strain designated SA-2 was shown to be the best
producer of extracellular lipase. Biochemical and physiological characterization
along with 16S rDNA sequence analysis placed SA-2 in the genus Salinivibrio
(Amoozegar et al., 2008).

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 12/27
Lipolytic activity has also been reported and characterized from an extremelyhalophilic archaeon, Natronococcus sp. (Boutaiba et al., 2006). Recently,
lipolytic enzymes have also been reported in B. halodurans strain originating
from a Kenyan alkaline soda lake (Vargas et al., 2004).
Thus variety of potential lipase producers has been studied in different part of
world. In Indian context the proposed study is likely to be a useful initiative.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 13/27
Methods
● Sample collection and purification of halophiles
Samples were collected from different saline habitats along the western coast of
India in sterile plastic containers. pH and other physical characteristics of thesample were recorded at the time of sample collection. Halophilic bacteria were
isolated by salt enrichment♦. Pure colonies were obtained by repeated streaking
on nutrient agar plates containing 10%/ 20% NaCl (w/v).
● Screening of enzyme activity
Lipase producers among the isolated halophilic strains were screened ontributyrin/ Rhodamine B agar plates. Potential isolates showing good zone of
hydrolysis were cultured in broth and enzyme activity was reconfirmed. Potential
enzyme producers have been selected for further studies.
• Lipase assay
Lipase activity was determined by the method of Kilcawley et al . [2002] using p-nitrophenyl palmitate as substrate. The amount of liberated p-nitrophenol ( p NP)
will be recorded at 400 nm.
Maintenance and preservation
The pure cultures were preserved on the CMB (Complete Medium Broth)* agar
media containing (10% w/v NaCl and adjusted to pH 8-10) and stored at 4°C.Subculturing was done at monthly interval.
♦ (10-20% NaCl in EM1 media containing (%): olive oil 2; MgSO4.7H2O 0.04.; MgCl2.6H2O
0.07; CaCl2.2H2O 0.05; KH2PO4 0.03; K 2HPO4 0.03; (NH4)2SO4 0.05; 0.01% of trace elements
solution containing 0.026 B, 0.05 Cu, 0.05 Mn, 0.006 Mo and 0.07 Zn).
* Complex Medium Broth (CMB) (g/l): glucose, 10; peptone, 5; Yeast Extract, 5; KH2PO4, 5.
Identification of Isolates by 16S rRNA gene Sequencing

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 14/27
Genomic DNA was isolated by PrepManTM Ultra sample preparation kit (Applied
Biosystems Inc., CA, and USA). The purity of DNA was checked by comparing
260/280 nm, 260/230 nm absorbance ratios and electrophorectic mobility onagarose gels. The 16S rDNA sequence was obtained by using MicroSeq ® fullgene
16S rDNA sequencing kit (Applied Biosystems Inc., CA, USA) and bacterial
identification was done by using MicroSeq®
3130 microbial identification system.
Identification of Isolates by 16S rRNA gene Bioinformatic analysis
These sequences were compared with gene sequences available at “ribosomalDNA database” (http://rdp.cme.msu.edu/) and the identity of the isolates was
established.
● Transmission Electron Microscopy (TEM)
The cells of log phase, grown in presence of salt (NaCl) were harvested bycentrifugation at 10,000 x g for 10 min at 4oC. The pellet was washed with 0.1 M
phosphate buffer (pH 7.4). The washed cells were fixed in modified Karnovsky’sfluid and processed as per the procedure of David et al . (1973). Micrographs wererecorded at TEM facility of IIT Delhi.
Optimization of culture conditions for growth and enzyme production
Nitrogen and carbon source/ concentrations, salt, dO2, pH requirements and other
relevant parameters will be optimized using response surface methodology. Scale-
up will be carried out in 3.5 L bioreactor.
● Purification of halophilic lipase
Halophilic isolates rich in lipase activity will be harvested at active growth stage
and crude extract will be prepared in appropriate buffer. Firstly ammonium sulfate
precipitation will be carried out. The ammonium sulfate fraction containing
desired enzyme activity would be subjected to combination of ion exchange/molecular sieving chromatography for further purification. Recent techniques of
affinity precipitation/ HIC will also be attempted to develop a single step
purification procedure. The homogeneity of enzyme preparation will bedetermined by SDS-PAGE. Protein content at each stage will be estimated by
Bradford method (Bradford, 1976).
Characterization of Halophilic lipase

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 15/27
Km of lipase will be determined using p-nitrophenyl palmitate as substrate.
Temperature and pH optima will also be determined towards same substrate.
Substrate specificity of enzyme will be determined using range of substrates andestimating corresponding activity. Enzyme kinetics and stability will also be
monitored in presence of different salt concentrations.
Molecular weight of the enzyme will be determined by SDS-PAGE and gel
exclusion chromatography.
Behavior of lipase in aqueous-organic solvent/ salt concentrations/ surfactants anddetergent systems with respect to activity and stability will be studied by
following standard protocols (Gupta et al., 2005).
Application of halophilic lipase
Lipase will be used in organic solvent/ low water system for fatty acid ester
synthesis. The reaction conditions will be optimized for maximum product yields.Enzymes stable in presence of surfactant and detergents will be studied for cleansing efficiency in detergent formulations.
Gene Cloning and Overexpression
The identity of the lipase encoding genes would be determined. N-terminal
sequencing of the purified protein will be obtained and matched with homologous
sequences in database. The gene sequence of matching protein will be used for designing appropriate primers. PCR amplified product of the test DNA (based on
above primers) will be cloned and sequenced.The lipase genes would also be cloned in inducible expression vectors (e.g. pET
system from clontech) with ‘N’ or ‘C’ terminal tags for easy purification of therecombinant lipase, which will be characterized subsequently.
● Structural and functional analysis
Bioinformatic analysis will be done to predict the three-dimensional structure of
halophilic lipase and to study the possible reasons for salt, temperature and
organic solvent stability of the protein.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 16/27
Work done so far
Sample Collection
Various saline and hyper saline samples (water and soil) were collected from sea of
Calicut (Kerala), saltern near old Goa (Goa), Nagoa beach (Diu, Gujarat), Somanath
(Gujarat) and Triveni sangam (Gujarat).
Isolation of Halophilic bacteria by salt enrichment
Halophilic bacteria were isolated by salt enrichment (10-20% NaCl). The isolates were
subjected to repeat streaking on nutrient agar plates containing NaCl (10-20% W/V) (Fig.
2a.). The pure halophilic colonies were plated on tributyrin agar to isolate lipase
producers based on zone of hydrolysis (Fig.2b).
Fig. 2a. Isolated halophiles streaked on nutrient agar plate
Fig. 2b. Lipase activity of Halophiles on tributyrin agar plate

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 17/27
Cell Morphology and Gram Staining
The isolates exhibited variation in their cell morphology, cell arrangement and Gram’s
reaction. The microscopic observations revealed that majority of the isolates were Gram
negative. Only few were gram positive, overall results are summarized in Table 2.
Table 2
A typical Gram negative reaction of isolate K 1 (from saline samples of Kerala coast) is
shown in Fig. 3.
Fig 3. Gram’s staining of isolate K 1
Lipase Activity in broth
Site of sample collection Gram positive isolates Gram negative isolates
Kozhikhode beach, Kerala 3 9
Nagoa beach, Diu, Gujarat 4 8
Somanath, Gujarat 4 11
Triveni sangam, Gujarat 6 12saltern near old Goa (Goa) 1 7

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 18/27
To monitor lipase production in liquid broth, media with following composition was
used:
Peptone- 1.0%
Yeast extract- 0.5%
MgSo4,7H2O- 0.02%.
CaCl2, 2H2O- 0.01%
Olive oil- 1%
Salt (NaCl) – 10%.
Medium was inoculated with 4%, v/v of 24 h grown mother culture (OD~1.3) and
incubated at 30oC in orbital shaker maintained at 120 rpm. Samples were withdrawn at
various time intervals and centrifuged at 10,000 x g for 10 min at 4 oC. Lipase activity
was determined in the cell-free supernatant as per following procedure:
Lipase assay
p-nitrophenyl palmitate was used as substrate. Briefly, 1.8 ml of solution containing 0.15
M NaCl and 0.5% Triton X-100 in 0.05 M Tris-HCl buffer (pH 8.0) was preincubated at
40°C. To this solution, 200 µl of suitable dilution of culture supernatant and 20 µl of
substrate (50 mM p NPP in acetonitrile) were added followed by incubation at 40°C for
30 min. The amount of liberated p-nitrophenol ( p NP) was recorded at 400 nm. One unit
of lipase activity is defined as the amount of enzyme liberating 1 nmol of p NP under
standard assay conditions (Kilcawley et al ., 2002).
Level of lipase production in different isolates is summarized in Table 3. On the basis of
level of production isolate K 1 was selected for further study. Halophilic isolate S-15-9
was a generous gift, from Prof. S.P Singh (Saurashtra University) that was compared with
other isolates.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 19/27
Table 3. Lipase production by halophilic isolates
Isolates Lipase production (U/ml)
K 1 129K 2 35
K 3 88
K 4 33
DL-20-1 51
DL-20-2 43
TL-20-2 65
SL-20-2 42
S-15-9 57
G4 32
G5 36
K- Isolate from sea water and sand (clear water) sample, Kozhikhode beach, Kerala
D- Isolate from sea water and sand. (Turbid water with soil particles) sample, Nagoa beach, Diu, Gujarat
T- Isolate from sea water, soil and sand. (Turbid water with soil particles) sample, Triveni sangam, Gujarat
S- Isolate from sea water, pebble and sand (clear water) sample, Somanath, Gujarat
G- Isolate from soil sample, saltern near old Goa (Goa)
Biochemical characterization of isolate K 1 was done by Hi25TM (HiMedia) kit. Results
are shown in Table. 4.
Table 4. Biochemical characteristics of isolate K 1

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 20/27
Parameters Observation Parameters Observation
Gram’s staining Negative Oxidase Positive
Cell shape Rod Methyl Red Negative
Pigmentation White Indole production Negative
Catalase Positive Xylose utilization Negative
ONPG Negative Serine utilization Positive
Lysine decarboxylase Positive Cellobiose utilization Positive
Ornithine decarboxylase Positive Melibiose utilization Negative
Urease Positive Fructose utilization Positive
Nitrate production Positive Raffinose utilization Negative
H2S production Negative Glucose utilization Positive
Citrate utilization Positive Lactose utilization Negative
Voges Proskauer’s Negative Glycerol utilization Positive
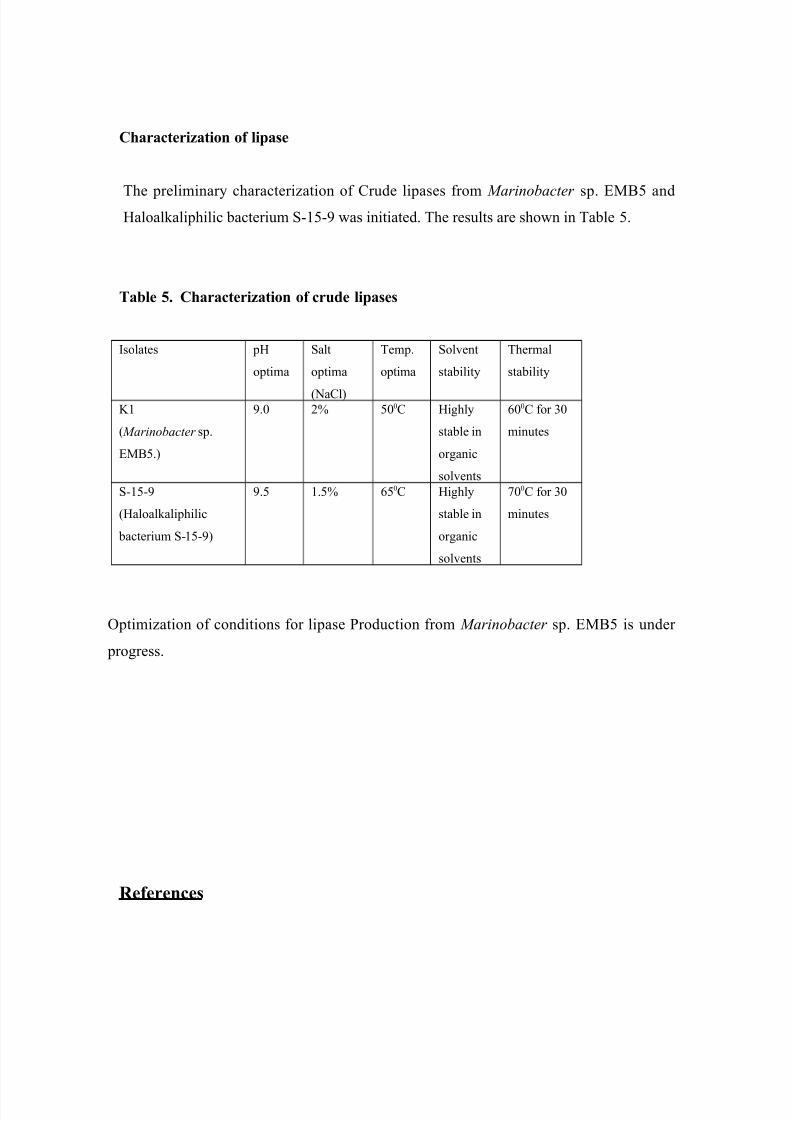
Identification of K 1 and S-15-9 by 16S rRNA gene sequencing
The full length gene of isolate K 1 showed ~99 % relatedness with Marinobacter sp.
Therefore, the new isolate is named as Marinobacter sp. EMB5. Isolate S-15-9 showed
~99 % relatedness with a Haloalkaliphilic bacterium. Therefore, this is named as
Haloalkaliphilic bacterium S-15-9. Phylogenetic tree of isolates are shown in Fig. 4.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 21/27
7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 22/27
Characterization of lipase
The preliminary characterization of Crude lipases from Marinobacter sp. EMB5 andHaloalkaliphilic bacterium S-15-9 was initiated. The results are shown in Table 5.
Table 5. Characterization of crude lipases
Isolates pH
optima
Salt
optima
(NaCl)
Temp.
optima
Solvent
stability
Thermal
stability
K1
( Marinobacter sp.
EMB5.)
9.0 2% 500C Highly
stable in
organic
solvents
600C for 30
minutes
S-15-9
(Haloalkaliphilic
bacterium S-15-9)
9.5 1.5% 650C Highly
stable in
organic
solvents
700C for 30
minutes
Optimization of conditions for lipase Production from Marinobacter sp. EMB5 is under
progress.
References

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 23/27
Abdou, A. M. (2003). Purification and partial characterization of psychrotrophic
Serratia marcescens lipase. J. Dairy Sci., 86: 127-132.
Adams, M.W.W., Perler, F.B. and Kelly, R.M. (1995). Extremozymes: expanding
the limits of biocatalysis. Biotechnology, 13: 662–668.
Aires-Barros, M. R., Taipa, M. A., and Cabral, J. M. S. (1994). Isolation and
purification of lipases, in Lipases: Their structure, biochemistry and application
(Woolley, P., and Petersen, S. B. Eds.), Cambridge University Press, Cambridge,UK, pp. 243-270.
Amoozegar, M. A., Salehghamari, E., Khajeh, K., Kabiri, M. and Naddaf, S.
(2008). Production of an extracellular thermohalophilic lipase from a moderatelyhalophilic bacterium, Salinivibrio sp. strain SA-2. J. Basic Microbiol., 48(3): 160-
167.
Anton, J., Rossello-Mora, R., Rodríguez-Valera, F. and Amann, R. (2000).Extremely halophilic bacteria in crystallizer ponds from solar salterns. Appl.
Environ. Microbiol., 66: 3052-3057.
Boutaiba, S., Bhatnagar, T., Hacene, H. , Mitchell D.A. and Baratti, J.C. (2006).Preliminary characterisation of a lipolytic activity from an extremely halophilic
archaeon, Natronococcus sp. J. Mol. Cat. B: Enzym., 41: 21–26.
.
Bradford, M.M. (1976). A rapid and sensitive method for the quantification of
microgram quantities of proteins utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248–254.
Castro-Ochoa, L. D., Rodr ıguez-G´omez, C., Valerio-Alfaro, and Ros, R. O.(2005). Screening, purification and characterization of the thermoalkalophilic lipase
produced by Bacillus thermoleovorans CCR11. Enzyme Microb. Technol., 37: 648-
654.
Chakraborty, K., and Raj, R. P. (2008). An extra-cellular alkaline metallo-lipase
from Bacillus licheniformis MTCC 6824: Purification and biochemical
characterization. Food Chem., 109: 727-736.
David, G. F., Herbert, J., and Wright, G. D. (1973). The ultrastructure of pineal
ganglion in the ferret. J. Anat., 115: 79-89.
de Lourdes Moreno, M., Garcia, M. T., Ventosa, A. and Mellado, E.(2009).
Characterization of Salicola sp. IC10, a lipase- and protease-producing extreme
halophile. FEMS Microbiol. Ecol., 68(1): 59-71.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 24/27
Dharmsthiti, S., and Kuhasuntisuk, B. (1998). Lipase from Pseudomonas
aeruginosa LP602: Biochemical properties and application for wastewater
treatment. J. Ind. Microbiol. Biotechnol., 21 : 75-80.
Favilla, R., Parisoli, A. and Mazzini, A. (1997). Alkaline denaturation and partial
refolding of pepsin investigated with DAPI as an extrinsic probe. Biophys. Chem.,67: 75-83.
Gupta, A., Patel, R. K., Roy, I., Singh, S. P., Khare, S. K. and Gupta, M. N.
(2005). One step purification and characterization of an alkaline protease from
haloalkaliphilic Bacillus sp. J. Chromatogr. A., 1075: 103-108.
Gupta, R., Gupta, N., and Rathi, P. (2004). Bacterial lipases: An overview of
production, purification and biochemical properties. Appl. Microbiol. Biotechnol.64: 763-781.
Haas, M. J., Cichowicz, D. J., and Bailey, D. G. (1992). Purification andcharacterization of an extracellular lipase from the fungus Rhizopus delemar .
Lipids, 27: 571-576.
Hiol, A., Jonzo, M. D., Druet, D., and Comeau, L. (1999). Production, purificationand characterization of an extracellular lipase from Mucor hiemalis f. hiemalis.
Enzyme Microb. Technol., 25: 80-87.
Houde, A., Kademi, A., and Leblanc, D. (2004). Lipases and their industrial
applications. Appl. Biochem. Biotechnol., 118: 155-170.
Imamura, S., and Kitaura, S. (2000). Purification and characterization of a
monoacylglycerol lipase from the moderately thermophilic Bacillus sp. H-257. J.
Biochem., 127: 419-425.
Jaeger K. E. and Reetz M. T. (1998). Microbial lipases form versatile tools for
biotechnology. Trends Biotechnol., 16: 396–403.
Jaeger, K. E., Ransac, S., Dijkstra, B. W., Colson, C., Heuvel, M. v., and Misset, O.
(1994). Bacterial lipases. FEMS Microbiol. Rev., 15: 29-63.
Karadzic, I., Masui, A., Zivkovic, L. I., and Fujiwara, N. (2006). Purification and
characterization of an alkaline lipase from Pseudomonas aeruginosa isolated from
putrid mineral cutting oil as component of metalworking fluid. J. Biosci. Bioeng.,102: 82-89.
Kilcawley, K.N., Wilkinson, M.G. and Fox, P.F. (2002). Determination of key
enzyme activities in commercial peptidase and lipase preparations from microbialor animal sources. Enzyme Microbial. Technol., 31: 310-320.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 25/27
Kim, H. K., Choi, H. J., Kim, M. H., Sohn, C. B., and Oh, T. K. (2002). Expression
and characterization of Ca2+-independent lipase from Bacillus pumilus B26.
Biochem. Biophys. Acta, 1583: 205-212.
Kim, J. and Dordick J. S. (1997). Unusual salt and solvent dependence of a proteasefrom an extreme halophile. Biotech. Bioeng., 55: 471-479.
Kim, S. S., Kim, E. K., and Rhee, J. S. (1996). Effects of growth rate on the
production of Pseudomonas fluorescens lipase during the fed batch cultivation of Escherichia col . Biotechnol. Prog., 12: 718-722.
Kojima, Y., Yokoe, M., and Mase, T. (1994). Purification and characterization of analkaline lipase from Pseudomonas fluorescens AK 102. Biosci. Biotechnol.
Biochem., 58: 1564-1568.
Kordel, M., Hofmann, B., Schaumburg, D., and Schmid, R. D. (1991). Extracellular lipase of Pseudomonas sp. strain ATCC 21808: Purification, characterization,
crystallization and preliminary X ray diffraction data. J. Bacteriol., 173: 4836-4841.
Kulkarni, N., and Gadre, R. V. (2002). Production and properties of an alkaline,
thermophilic lipase from Pseudomonas fluorescens NS2W. J. Ind. Food Microbiol.,
28: 344-348.
Litthauer, D., Ginster, A. and Skein, E. V. E. (2002). Pseudomonas luteola lipase:
A new member of the 320-residue Pseudomonas lipase family. Enzyme Microb.
Technol., 30: 209-215.
Martı´n, S., Ma´ rquez, M. C. , Sa´ nchez-Porro, C., Mellado, E., Arahal D. R. and
Ventosa, A. (2003). Marinobacter lipolyticus sp. nov., a novel moderate halophilewith lipolytic activity. Int. J. of Syst. Evol. Microbiol., 53: 1383–1387
Ogino, H., Katou, Y., Akagi, R., Mimitsuka, T., Hiroshima, S., Gemba, Y.,Doukyu, N., Yasuda, M., Ishimi, K., and Ishikawa, H. (2007). Cloning and
expression of gene, and activation of an organic solvent-stable lipase from
Pseudomonas aeruginosa LST-03. Extremophiles, 11: 809-817.
Ogino, H., Nakagawa, S., Shinya, K., Muto, T., Fujimura, N., Yasuda, M., and
Ishikawa, H. (2000). Purification and characterization of organic solvent-stablelipase from organic solvent-tolerant Pseudomonas aeruginosa LST-03. J. Biosci. Bioeng., 89: 451-457.
Ozcan, B., Ozyilmaz, G., Cokmus, C. and Caliskan, M. (2009). Characterization of
extracellular esterase and lipase activities from five halophilic archaeal strains. J.
Ind. Microbiol. Biotechnol., 36(1): 105-110.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 26/27
Pratuamgdejkul, J., and Dharmsthiti, S. (2000). Purification and characterization of
lipase form psychrophilic Acinetobacter calcoaceticus LP009. Microbiol. Res., 155:95-100.
Queiroz, J. A., Tomaz, C. T., and Cabral, J. M. S. (2001). Hydrophobic interactionchromatography of proteins. J. Biotechnol., 87: 143-159.
Rahman, R. N. Z. R. A., Baharum, S. N., Basri, M., and Salleh, A. B. (2005) High-yield purification of an organic solvent-tolerant lipase from Pseudomonas sp. strain
S5. Anal. Biochem., 341: 267-274.
Rathi, P., Saxena, R. K., and Gupta, R. A. (2001) Novel lipase from Burkholderiacepacia for detergent formulation. Process Biochem., 37: 187-192.
Rodriguez-Valera, F. (1992). Biotechnological potential of halobacteria.In: Danson,
M.J., Hough, D.W. and Lunt, G.G. (eds) The Archaebacteria: Biochemistry and Biotechnology, pp. 135–147.
Salameh, M., and Wiegel, J. (2007). Lipases from extremophiles and potential for
industrial applications, Adv. Appl. Microbiol., 61: 253-283.
Sanchez-Porro, C., Martin, S., Mellado, E. and Ventosa, A. (2003) Diversity of
moderately halophilic bacteria producing extracellular hydrolytic enzymes. J. Appl. Microbiol., 94: 295–300.
Shabtai, Y., and Daya-Mishne, N. (1992). Production, purification and properties of a lipase from a bacterium ( Pseudomonas aeruginosa YS-7) capable of growing in
water-restricted environments. Appl. Environ. Microbiol., 58: 174-180.
Shaoxin, C., Lili, Q., and Bingzhao, S. (2007). Purification and properties of
enantioselective lipase from a newly isolated Bacillus cereus C71. Process
Biochem., 42: 988-994.
Singh, S. and Banerjee, U. C. (2007). Purification and characterization of trans-3-
(4-methoxyphenyl) glycidic acid methyl ester hydrolyzing lipase from
Pseudomonas aeruginosa. Process Biochem., 42: 1063-1068.
Snellman, E. A., Sullivan, E. R., and Colwell, R. R. (2002). Purification and
properties of the extracellular lipase, Lip A, of Acinetobacter sp. RAG-1. Eur. J. Biochem., 269: 5771-5779.
Sun, S. Y., and Xu, Y. (2008). Solid-state fermentation for ‘whole-cell syntheticlipase’ production from Rhizopus chinensis and identification of the functional
enzyme. Process Biochem., 43: 219-224.

7/28/2019 sample of a research plan
http://slidepdf.com/reader/full/sample-of-a-research-plan 27/27
Surinenaite, B., Bendikiene, V., Juodka, B., Bachmatova, I. and Marcinkevichiene,
L. (2002). Characterization and physicochemical properties of a lipase from Pseudomonas mendocina 3121-1. Biotechnol. Appl. Biochem., 36: 47-55.
Sztajer, H., Liinsdorf, H., Erdmann, H., Menge, U. and Schmid, R. (1992).Purification and properties of lipase from Penicillium simplicissimum. Biochim.
Biophys. Acta, 1124: 253-261.
Trodler, P., Nieveler, J., Rusnak, M., Schmid, R. D. and Pleiss, J. (2008). Rational
design of a new one-step purification strategy for Candida antarctica lipase B by
ion-exchange chromatography. J. Chromatogr. A, 1179: 161-167.
Vargas A.V., Delgado O.D., Hatti-Kaul R. and Mattiasson B. (2004). Lipase
producing microorganisms from a Kenyan alkaline soda lake. Biotechnol. Lett., 26:
81–86.
Ventosa, A., Nieto, J. J. and Oren, A. (1998). Biology of moderately halophilic
aerobic bacteria. Microbiol. Mo.l Bio.l Rev., 62: 504–544.
Vreeland, R.H., Rosenzweig, W.D. and Powers, D.W. (2000). Isolation of a 250
million-year-old halotolerant bacterium from a primary salt crystal. Nature., 407:897 - 900.
Yan, J., Yang, J., Xu, L. and Yan, Y. (2007). Gene cloning, overexpression and
characterization of a novel organic solvent tolerant and thermostable lipase from
Galactomyces geotrichum Y05. J. Mol. Cat. B: Enzym., 49: 28-35.
Zhang, A., Renjun, G., Diao, N., Xie, G., Gao, G. and Cao, S. (2009). Cloning,expression and characterization of an organic solvent tolerant lipase from
Pseudomonas fluorescens JCM5963. J. Mol. Cat. B: Enzym., 56: 78–84.
Related Documents











