REVIEW Salt stress inhibits photosystems II and I in cyanobacteria Suleyman I. Allakhverdiev Norio Murata Received: 12 May 2008 / Accepted: 12 July 2008 / Published online: 1 August 2008 Ó Springer Science+Business Media B.V. 2008 Abstract Recent studies of responses of cyanobacterial cells to salt stress have revealed that the NaCl-induced decline in the photosynthetic activities of photosystems II and I involves rapid and slow changes. The rapid decreases in the activities of both photosystems, which occur within a few minutes, are reversible and are associated with osmotic effects, which induce the efflux of water from the cytosol through water channels and rapidly increase intracellular concentrations of salts. Slower decreases in activity, which occur within hours, are irreversible and are associated with ionic effects that are due to the influx of Na + and Cl - ions through K + (Na + ) channels and, probably, Cl - channels, with resultant dissociation of extrinsic proteins from photosystems. In combination with light stress, salt stress significantly stimulates photoinhibition by inhibiting repair of photodamaged photosystem II. Tolerance of photosys- tems to salt stress can be enhanced by genetically engineered increases in the unsaturation of fatty acids in membrane lipids and by intracellular synthesis of compatible solutes, such as glucosylglycerol and glycinebetaine. In this review, we summarize recent progress in research on the effects of salt stress on photosynthesis in cyanobacteria. Keywords Cyanobacteria Membrane lipids Photosystem II Photosystem I Salt stress Tolerance Abbreviations BQ 1,4-Benzoquinone CCCP Carbonyl cyanide m-chlorophenylhydrazone Chl Chlorophyll CSA p-Chloromercuriphenyl-sulfonic acid DCIP 2,6-Dichlorophenolindophenol DCMU 3-(3 0 ,4 0 -Dichlorophenyl)-1,1-dimethylurea DPC Diphenylcarbazide FCCP Carbonyl cyanide p-trifluoro-methoxyphenyl- hydrazone GG Glucosylglycerol MV Methylviologen PSII Photosystem II PSI Photosystem I ROS Reactive oxygen species Introduction Photosynthesis converts light energy into chemical energy via electron transport through two photosystems, namely, photosystem II (PSII) and photosystem I (PSI). PSII and, sometimes, PSI are particularly sensitive to environmental stress (Powles 1984; Aro et al. 1993; Hagemann and Erd- mann 1997; Adir et al. 2003; Allakhverdiev and Murata 2004; Nishiyama et al. 2005, 2006; Murata et al. 2007; Mohanty et al. 2007; Takahashi and Murata 2008). Since the efficiency and productivity of photosynthesis are clo- sely related to the activities of the two photosystems, considerable attention has been paid to the effects of environmental stress on these photosystems. Cyanobacteria provide suitable model systems for studies of the effects of environmental stress on photosynthesis since these prokaryotes perform oxygenic photosynthesis using a photosynthetic apparatus similar to that found in chloroplasts S. I. Allakhverdiev Institute of Basic Biological Problems, Russian Academy of Sciences, Pushchino, Moscow Region 142290, Russia N. Murata (&) National Institute for Basic Biology, Okazaki 444-8585, Japan e-mail: [email protected] 123 Photosynth Res (2008) 98:529–539 DOI 10.1007/s11120-008-9334-x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REVIEW
Salt stress inhibits photosystems II and I in cyanobacteria
Suleyman I. Allakhverdiev Æ Norio Murata
Received: 12 May 2008 / Accepted: 12 July 2008 / Published online: 1 August 2008
� Springer Science+Business Media B.V. 2008
Abstract Recent studies of responses of cyanobacterial
cells to salt stress have revealed that the NaCl-induced
decline in the photosynthetic activities of photosystems II
and I involves rapid and slow changes. The rapid decreases in
the activities of both photosystems, which occur within a few
minutes, are reversible and are associated with osmotic
effects, which induce the efflux of water from the cytosol
through water channels and rapidly increase intracellular
concentrations of salts. Slower decreases in activity, which
occur within hours, are irreversible and are associated with
ionic effects that are due to the influx of Na+ and Cl- ions
through K+(Na+) channels and, probably, Cl- channels,
with resultant dissociation of extrinsic proteins from
photosystems. In combination with light stress, salt stress
significantly stimulates photoinhibition by inhibiting repair
of photodamaged photosystem II. Tolerance of photosys-
tems to salt stress can be enhanced by genetically engineered
increases in the unsaturation of fatty acids in membrane
lipids and by intracellular synthesis of compatible solutes,
such as glucosylglycerol and glycinebetaine. In this review,
we summarize recent progress in research on the effects of
salt stress on photosynthesis in cyanobacteria.
Keywords Cyanobacteria � Membrane lipids �Photosystem II � Photosystem I � Salt stress � Tolerance
Abbreviations
BQ 1,4-Benzoquinone
CCCP Carbonyl cyanide m-chlorophenylhydrazone
Chl Chlorophyll
CSA p-Chloromercuriphenyl-sulfonic acid
DCIP 2,6-Dichlorophenolindophenol
DCMU 3-(30,40-Dichlorophenyl)-1,1-dimethylurea
DPC Diphenylcarbazide
FCCP Carbonyl cyanide p-trifluoro-methoxyphenyl-
hydrazone
GG Glucosylglycerol
MV Methylviologen
PSII Photosystem II
PSI Photosystem I
ROS Reactive oxygen species
Introduction
Photosynthesis converts light energy into chemical energy
via electron transport through two photosystems, namely,
photosystem II (PSII) and photosystem I (PSI). PSII and,
sometimes, PSI are particularly sensitive to environmental
stress (Powles 1984; Aro et al. 1993; Hagemann and Erd-
mann 1997; Adir et al. 2003; Allakhverdiev and Murata
2004; Nishiyama et al. 2005, 2006; Murata et al. 2007;
Mohanty et al. 2007; Takahashi and Murata 2008). Since
the efficiency and productivity of photosynthesis are clo-
sely related to the activities of the two photosystems,
considerable attention has been paid to the effects of
environmental stress on these photosystems.
Cyanobacteria provide suitable model systems for studies
of the effects of environmental stress on photosynthesis since
these prokaryotes perform oxygenic photosynthesis using a
photosynthetic apparatus similar to that found in chloroplasts
S. I. Allakhverdiev
Institute of Basic Biological Problems, Russian Academy of
Sciences, Pushchino, Moscow Region 142290, Russia
N. Murata (&)
National Institute for Basic Biology, Okazaki 444-8585, Japan
e-mail: [email protected]
123
Photosynth Res (2008) 98:529–539
DOI 10.1007/s11120-008-9334-x

of higher plants and algae (Pfenning 1978; Oquist et al.
1995). Moreover, cyanobacterial cells can easily be exposed
directly to defined stress conditions in culture (Joset et al.
1996; Hagemann and Erdmann 1997) and they are able to
acclimate to a wide range of environmental stresses (Nishida
and Murata 1996; Hagemann and Erdmann 1997). The
cyanobacterial genes whose expression is enhanced by salt
stress have been identified. Levels of expression of 28 genes
increase more than threefold in response to salt stress spe-
cifically, while those of 11 genes increase specifically in
response to osmotic stress, and those of 34 genes are
increased both by salt stress and by osmotic stress (Kanesaki
et al. 2002). The products of half of these genes are hypo-
thetical proteins, whose functions have not been
characterized to date. Homologs of the salt-induced genes
found in Synechocystis sp. PCC 6803 (hereafter referred to as
Synechocystis) are similarly regulated by salt stress in higher
plants, confirming that this cyanobacterium provides a good
model for investigations of the responses of photosynthetic
organisms to salt stress (Bohnert et al. 2001). However, the
anti-oxidant systems in cyanobacteria differ from those in
higher plants (Demmig-Adams and Adams 1992, 2002;
Asada 2006) and we need to take this observation into
account, especially under conditions of strong light and salt
stress, when anti-oxidant systems are very important.
Nonetheless, using cyanobacterial cells, we can study the
direct effects of salt stress on the photosynthetic machinery,
and are confident that we should be able to apply the results
obtained in cyanobacteria to higher plants.
The basic physiological responses of cyanobacterial cells
to salt stress have been investigated in great detail (for review
of early works, see Hagemann and Erdmann 1997). The
responses occur in three phases. The first phase is observed
after a sudden increase in the ambient concentration of NaCl,
and it includes the influx of Na+ and Cl- ions, within sec-
onds, into the cytoplasm. The second phase, which occurs
within an hour, is characterized by replacement of Na+ ions
by K+ ions, leading to a decline in the toxic effects of high
concentrations of Na+ ions. The third and longest phase lasts
for several hours. This phase is important for the acclimation
of cells to elevated concentrations of ions. During this phase,
the synthesis or uptake of compatible solutes occurs to mit-
igate the toxic effects of salts and to conserve the structures
of complex proteins and cell membranes (Reed et al. 1985),
and also the cyclic electron transport via PSI is activated
(Joset et al. 1996; Hibino et al. 2000. Over a longer period of
time, namely, several days, salt stress inhibits cell division
(Ferjani et al. 2003).
Cyanobacteria have evolved a variety of protective
mechanisms that allow them to acclimate to unfavorable
salt-stress conditions, to survive, and to proliferate. Mech-
anisms of tolerance to salt stress have also been investigated
in detail, with emphasis on the role of Na+ /H+ antiporters
(Allakhverdiev et al. 1999, 2000b; Inaba et al. 2001), water
and ionic channels (Allakhverdiev et al. 2000a, b), the
synthesis of compatible solutes (Hagemann and Erdmann
1997; Hayashi and Murata 1998; Chen and Murata 2002)
and of salt stress-induced proteins (Bhagwat and Apte 1989;
Hagemann et al. 1990, 1991), and membrane-lipid com-
position (Allakhverdiev et al. 1999, 2001).
In this review, we focus on the effects of salt (NaCl)
stress on photosystems in cyanobacteria, the mechanisms
of tolerance to salt stress, and in particular, the inhibition of
the repair of PSII by salt stress. We also discuss the role of
membrane lipids and compatible solutes in the tolerance of
cyanobacterial cells to salt stress.
Osmotic stress decreases the cytoplasmic volume
and inhibits PSI and PSII reversibly
The effects of osmotic stress on the cytoplasmic volume
and the photosynthetic machinery have been studied
extensively in Synechococcus sp. PCC 7942 (hereafter
referred to as Synechococcus) and in Synechocystis. When
Synechocystis cells are incubated in the presence of 0.5 M
sorbitol, the cytoplasmic volume decreases to 30–50% of
the original volume in \5 min (Kanesaki et al. 2002;
Shapiguzov et al. 2005), indicating that shrinkage of the
cytoplasm occurs instantaneously. The Synechocystis gen-
ome contains the apqZ gene for a water channel (Kaneko
et al. 1996). In a Synechocystis cells with a mutant aqpZ
gene, osmotic stress had no effect at all on cytoplasmic
volume (Shapiguzov et al. 2005).
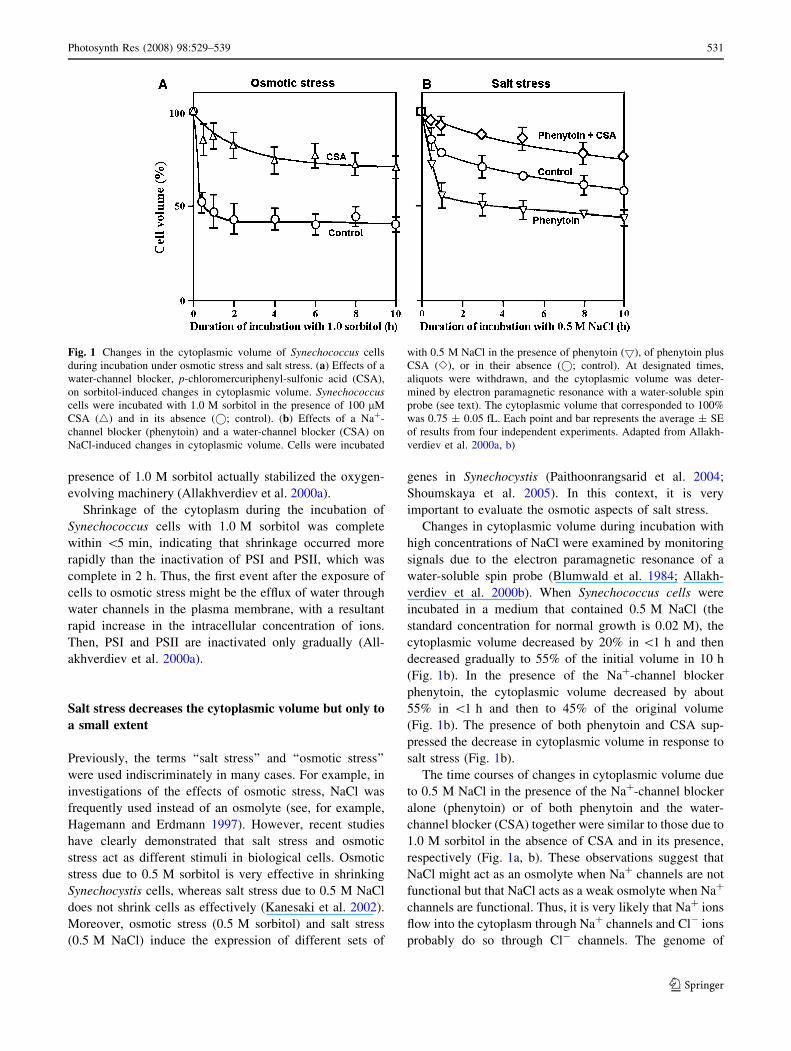
In Synechococcus, osmotic stress due to 1.0 M sorbitol
decreased the cytoplasmic volume to 50% of the original
value in\10 min (Fig. 1a). The presence of a water-channel
blocker, p-chloromercuriphenyl-sulfonic acid (CSA), sig-
nificantly limited the decrease in cytoplasmic volume in
terms of both rate and extent (Fig. 1a). Under normal con-
ditions, the cytoplasmic volume returned to the original
volume after removal of the osmotic stress, but CSA mark-
edly inhibited such recovery (Allakhverdiev et al. 2000a).
These observations indicate that water channels participate
in the efflux and influx of water across the plasma membrane
and in the shrinkage and swelling of the cytoplasm.
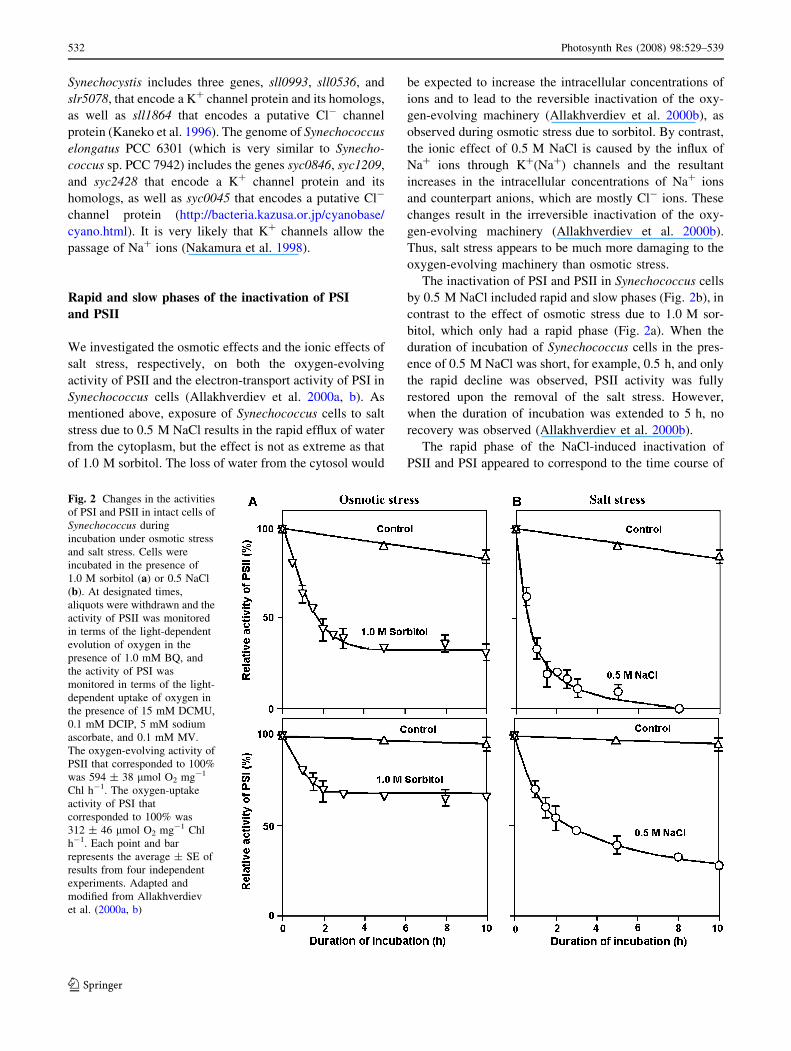
The effects of osmotic stress on the activities of PSII and
PSI in intact cells were examined by monitoring the light-
dependent evolution and uptake of oxygen, respectively, in
Synechococcus (Fig. 2a). Within 2 h, osmotic stress due to
1.0 M sorbitol depressed the oxygen-evolving activity of
PSII to 40% of the original value and decreased the electron-
transport activity of PSI to 70% of the original value
(Fig. 2a). By contrast, an examination of the effects of sor-
bitol on the PSII- and PSI-mediated transport of electrons in
vitro revealed that, in isolated thylakoid membranes, the
530 Photosynth Res (2008) 98:529–539
123
presence of 1.0 M sorbitol actually stabilized the oxygen-
evolving machinery (Allakhverdiev et al. 2000a).
Shrinkage of the cytoplasm during the incubation of
Synechococcus cells with 1.0 M sorbitol was complete
within \5 min, indicating that shrinkage occurred more
rapidly than the inactivation of PSI and PSII, which was
complete in 2 h. Thus, the first event after the exposure of
cells to osmotic stress might be the efflux of water through
water channels in the plasma membrane, with a resultant
rapid increase in the intracellular concentration of ions.
Then, PSI and PSII are inactivated only gradually (All-
akhverdiev et al. 2000a).
Salt stress decreases the cytoplasmic volume but only to
a small extent
Previously, the terms ‘‘salt stress’’ and ‘‘osmotic stress’’
were used indiscriminately in many cases. For example, in
investigations of the effects of osmotic stress, NaCl was
frequently used instead of an osmolyte (see, for example,
Hagemann and Erdmann 1997). However, recent studies
have clearly demonstrated that salt stress and osmotic
stress act as different stimuli in biological cells. Osmotic
stress due to 0.5 M sorbitol is very effective in shrinking
Synechocystis cells, whereas salt stress due to 0.5 M NaCl
does not shrink cells as effectively (Kanesaki et al. 2002).
Moreover, osmotic stress (0.5 M sorbitol) and salt stress
(0.5 M NaCl) induce the expression of different sets of
genes in Synechocystis (Paithoonrangsarid et al. 2004;
Shoumskaya et al. 2005). In this context, it is very
important to evaluate the osmotic aspects of salt stress.
Changes in cytoplasmic volume during incubation with
high concentrations of NaCl were examined by monitoring
signals due to the electron paramagnetic resonance of a
water-soluble spin probe (Blumwald et al. 1984; Allakh-
verdiev et al. 2000b). When Synechococcus cells were
incubated in a medium that contained 0.5 M NaCl (the
standard concentration for normal growth is 0.02 M), the
cytoplasmic volume decreased by 20% in \1 h and then
decreased gradually to 55% of the initial volume in 10 h
(Fig. 1b). In the presence of the Na+-channel blocker
phenytoin, the cytoplasmic volume decreased by about
55% in \1 h and then to 45% of the original volume
(Fig. 1b). The presence of both phenytoin and CSA sup-
pressed the decrease in cytoplasmic volume in response to
salt stress (Fig. 1b).
The time courses of changes in cytoplasmic volume due
to 0.5 M NaCl in the presence of the Na+-channel blocker
alone (phenytoin) or of both phenytoin and the water-
channel blocker (CSA) together were similar to those due to
1.0 M sorbitol in the absence of CSA and in its presence,
respectively (Fig. 1a, b). These observations suggest that
NaCl might act as an osmolyte when Na+ channels are not
functional but that NaCl acts as a weak osmolyte when Na+
channels are functional. Thus, it is very likely that Na+ ions
flow into the cytoplasm through Na+ channels and Cl- ions
probably do so through Cl- channels. The genome of
Fig. 1 Changes in the cytoplasmic volume of Synechococcus cells
during incubation under osmotic stress and salt stress. (a) Effects of a
water-channel blocker, p-chloromercuriphenyl-sulfonic acid (CSA),
on sorbitol-induced changes in cytoplasmic volume. Synechococcuscells were incubated with 1.0 M sorbitol in the presence of 100 lM
CSA (4) and in its absence (�; control). (b) Effects of a Na+-
channel blocker (phenytoin) and a water-channel blocker (CSA) on
NaCl-induced changes in cytoplasmic volume. Cells were incubated
with 0.5 M NaCl in the presence of phenytoin (5), of phenytoin plus
CSA (e), or in their absence (�; control). At designated times,
aliquots were withdrawn, and the cytoplasmic volume was deter-
mined by electron paramagnetic resonance with a water-soluble spin
probe (see text). The cytoplasmic volume that corresponded to 100%
was 0.75 ± 0.05 fL. Each point and bar represents the average ± SE
of results from four independent experiments. Adapted from Allakh-
verdiev et al. 2000a, b)
Photosynth Res (2008) 98:529–539 531
123
Synechocystis includes three genes, sll0993, sll0536, and
slr5078, that encode a K+ channel protein and its homologs,
as well as sll1864 that encodes a putative Cl- channel
protein (Kaneko et al. 1996). The genome of Synechococcus
elongatus PCC 6301 (which is very similar to Synecho-
coccus sp. PCC 7942) includes the genes syc0846, syc1209,
and syc2428 that encode a K+ channel protein and its
homologs, as well as syc0045 that encodes a putative Cl-
channel protein (http://bacteria.kazusa.or.jp/cyanobase/
cyano.html). It is very likely that K+ channels allow the
passage of Na+ ions (Nakamura et al. 1998).
Rapid and slow phases of the inactivation of PSI
and PSII
We investigated the osmotic effects and the ionic effects of
salt stress, respectively, on both the oxygen-evolving
activity of PSII and the electron-transport activity of PSI in
Synechococcus cells (Allakhverdiev et al. 2000a, b). As
mentioned above, exposure of Synechococcus cells to salt
stress due to 0.5 M NaCl results in the rapid efflux of water
from the cytoplasm, but the effect is not as extreme as that
of 1.0 M sorbitol. The loss of water from the cytosol would
be expected to increase the intracellular concentrations of
ions and to lead to the reversible inactivation of the oxy-
gen-evolving machinery (Allakhverdiev et al. 2000b), as
observed during osmotic stress due to sorbitol. By contrast,
the ionic effect of 0.5 M NaCl is caused by the influx of
Na+ ions through K+(Na+) channels and the resultant
increases in the intracellular concentrations of Na+ ions
and counterpart anions, which are mostly Cl- ions. These
changes result in the irreversible inactivation of the oxy-
gen-evolving machinery (Allakhverdiev et al. 2000b).
Thus, salt stress appears to be much more damaging to the
oxygen-evolving machinery than osmotic stress.
The inactivation of PSI and PSII in Synechococcus cells
by 0.5 M NaCl included rapid and slow phases (Fig. 2b), in
contrast to the effect of osmotic stress due to 1.0 M sor-
bitol, which only had a rapid phase (Fig. 2a). When the
duration of incubation of Synechococcus cells in the pres-
ence of 0.5 M NaCl was short, for example, 0.5 h, and only
the rapid decline was observed, PSII activity was fully
restored upon the removal of the salt stress. However,
when the duration of incubation was extended to 5 h, no
recovery was observed (Allakhverdiev et al. 2000b).
The rapid phase of the NaCl-induced inactivation of
PSII and PSI appeared to correspond to the time course of
Fig. 2 Changes in the activities
of PSI and PSII in intact cells of
Synechococcus during
incubation under osmotic stress
and salt stress. Cells were
incubated in the presence of
1.0 M sorbitol (a) or 0.5 NaCl
(b). At designated times,
aliquots were withdrawn and the
activity of PSII was monitored
in terms of the light-dependent
evolution of oxygen in the
presence of 1.0 mM BQ, and
the activity of PSI was
monitored in terms of the light-
dependent uptake of oxygen in
the presence of 15 mM DCMU,
0.1 mM DCIP, 5 mM sodium
ascorbate, and 0.1 mM MV.
The oxygen-evolving activity of
PSII that corresponded to 100%
was 594 ± 38 lmol O2 mg-1
Chl h-1. The oxygen-uptake
activity of PSI that
corresponded to 100% was
312 ± 46 lmol O2 mg-1 Chl
h-1. Each point and bar
represents the average ± SE of
results from four independent
experiments. Adapted and
modified from Allakhverdiev
et al. (2000a, b)
532 Photosynth Res (2008) 98:529–539
123

osmotic stress-induced inactivation (Fig. 2a), suggesting
that the rapid reversible decline in the activities of PSII and
PSI might have been caused by the osmotic effect of NaCl.
The slow and irreversible phase appeared to be specific to
ionic effects since the presence of a specific blocker of Na+
channels protected both PSII and PSI against the NaCl-
induced slow inactivation but not against the rapid inacti-
vation (Allakhverdiev et al. 2000b). In the presence of the
Na+ channel blocker, the NaCl-induced inactivation
resembled the sorbitol-induced inactivation. These obser-
vations suggest that Na+ and/or Cl- ions damage the
machinery that is necessary for the repair of PSII after
NaCl-induced inactivation.
Possible sites of inactivation of PSII and PSI by NaCl
Incubation of Synechococcus cells in the presence of 0.5 M
NaCl for 5 h inactivates both oxygen evolution in PSII and
electron transport in PSI (Allakhverdiev et al. 2000b).
Examination of the light-dependent quenching of chloro-
phyll fluorescence due to the reduction of pheophytin in
intact cells demonstrated that the site of inactivation is the
electron-donating side of PSII. Furthermore, incubation of
Synechococcus cells in the presence of NaCl did not
damage QA, pheophytin, and P680 but blocked the trans-
port of electrons from water to P680. These observations
indicate that the photochemical reaction center is not
involved in the NaCl-induced inactivation of PSII (All-
akhverdiev et al. 2000b).
In thylakoid membranes isolated from either Synecho-
cystis (Allakhverdiev et al. 1999) or Synechococcus
(Allakhverdiev et al. 2000b), the transport of electrons
from water to 2,6-dichlorophenolindophenol (DCIP), but
not from diphenylcarbazide (DPC) to DCIP, was sup-
pressed by the presence of 0.5 M NaCl. Since DPC donates
electrons to P680 (Yamashita and Butler 1969; Izawa
1980; Trebst 1980), it seems likely that salt stress inacti-
vates the oxygen-evolving machinery (Kuwabara and
Murata 1983; Miyao and Murata 1983; Murata and Miyao
1985).
We also examined the effects of salt stress on the
activity of PSI by monitoring the uptake of oxygen by
intact cells in the presence of DCIP, sodium ascorbate,
methyl viologen (MV), and 3-(30,40-dichlorophenyl)-1,1-
dimethylurea (DCMU) (Allakhverdiev et al. 2000b). In this
system, electrons are transported from the reduced form of
DCIP to MV through plastocyanin, P700, phylloquinone
(vitamin K1), and iron sulfur centers (Izawa 1980; Golbeck
1994). Incubation of Synechococcus cells with 0.5 M NaCl
suppressed the reduction of P700+ (Allakhverdiev et al.
2001). Since P700+ is reduced by plastocyanin, it seems
likely that the association of plastocyanin with the PSI
complex was disturbed by the presence of NaCl (Allakh-
verdiev et al. 2000b, 2001).
Salt-induced inactivation depends on the activity
and synthesis of Na+/H+ antiporters
The Synechocystis genome includes five genes for putative
Na+/H+ antiporters (Inaba et al. 2001). The Na+/H+
antiport system is responsible for maintaining the intra-
cellular concentration of Na+ ions at a certain low level.
The tolerance of Synechocystis cells to salt stress is related
to the activity of Na+/H+ antiporters, and moreover,
incubation of Synechocystis or Synechococcus cells with
NaCl inactivates the Na+/H+ antiport system in these
cyanobacterial cells (Allakhverdiev et al. 1999, 2000b).
Such inactivation might be due to inhibition of the syn-
thesis of proteins that are involved in the Na+/H+ antiport
system. Thus, the inactivation of Na+/H+ antiporters might
be related to the ionic effect of NaCl on the inactivation of
PSI and PSII (Allakhverdiev et al. 2000b). The inactivation
might be due to inhibition of the synthesis of the proteins
that are required for the function of Na+/H+ antiporters.
By contrast, water channels are predominantly respon-
sible for the hyperosmotic stress-induced inactivation of
PSII and PSI by sorbitol, as well as for some aspects of the
rapid phase of the NaCl-induced inactivation of PSII and
PSI (Allakhverdiev et al. 2000a, b). The rapid inactivation
is reversible, and protein synthesis is not required for the
recovery of the activities of PSI and PSII from the rapid
inactivation (Allakhverdiev et al. 2000b). It seems likely
that the Na+/H+ antiport system is involved in the rapid
and reversible inactivation of PSI and PSII and in the
recovery of their activities.
A hypothetical scheme for the salt-induced inactivation
of electron transport in PSI and PSII
Figure 3 shows a hypothetical scheme that might explain
the NaCl-induced inactivation of the photosynthetic
machinery. K+(Na+) channels, Cl- channels, water chan-
nels, and Na+/H+ antiporters are located on the plasma
membrane. The oxygen-evolving machinery of the PSII
complex is located on the luminal side of thylakoid
membranes. In cyanobacteria, this machinery is stabilized
by three extrinsic proteins, namely, PsbO (33-kD protein),
PsbV (cytochrome c550), and PsbU (Shen et al. 1998; Ni-
shiyama et al. 1999). Cyt c550 and PsbU are loosely bound
to the donor side of the core complex of PSII (Nishiyama
et al. 1997, 1999). These proteins are easily dissociated
from the cyanobacterial PSII complex in the presence of
elevated concentrations of NaCl (Shen et al. 1992, 1998).
Photosynth Res (2008) 98:529–539 533
123

When the extracellular concentrations of Na+ and Cl- ions
increase, water molecules in the intracellular space rapidly
leave the cell through water channels, whereas Na+ and
Cl- ions enter the cytoplasm through K+(Na+) channels
and Cl- channels, respectively, with resultant increases in
the cytosolic concentrations of Na+ and Cl- ions. The
Na+/H+ antiport system probably pumps Na+ ions out of
the cell. It seems likely that Na+ ions in the cytosol then
cross the thylakoid membrane to increase the concentration
of Na+ and Cl- ions in the intrathylakoid space (lumen),
although the mechanism of such movement of ions is
unknown. As a result, the extrinsic proteins of PSII dis-
sociate from the complex and the oxygen-evolving
machinery is partially inactivated. When the salt stress is
removed, water molecules enter the cytosol through the
water channels, and Na+ and Cl- ions leave the cytosol
through their respective channels, with resultant decreases
in the cytosolic concentrations of Na+ ions and Cl- ions.
The intrathylakoid concentrations of Na+ ions and Cl- ions
also decrease, and as a result, the integrity of PSII and PSI
is restored by the renewed binding of the extrinsic proteins
to these complexes.
The Na+/H+ antiport system is inactivated during long-
term incubation of cyanobacterial cells with 0.5 M NaCl,
and when it becomes inoperative, resultant increases occur
in the cytosolic concentrations and intrathylakoid concen-
trations of Na+ ions and Cl- ions. Extended incubation in
the presence of 0.5 M NaCl, during which the extrinsic
proteins remain dissociated from the complexes,
inactivates the Mn cluster, which is the catalytic center of
the oxygen-evolving complex.
A similar mechanism has been postulated for the NaCl-
induced inactivation of PSI. An increase in the intra-
thylakoid concentration of Na+ ions might lead to the
dissociation of plastocyanin or cytochrome c553 from the
PSI complex, causing a decrease in the rate of the PSI-
mediated transport of electrons (Allakhverdiev et al.
2000b, 2001).
Role of cellular energization in the recovery
of the activities of PSI and PSII
In most of the studies of the effects of salt stress on
cyanobacterial cells, the effects were examined in dark-
ness. However, cyanobacteria, such as Synechococcus and
Synechocystis, are photoautotrophic organisms, utilizing
light energy to drive energy production and the assimila-
tion of carbon via photosynthesis. It appears that light is an
important factor in the restoration of the activities of PSII
and PSI during incubation of cyanobacterial cells under
stress (Allakhverdiev et al. 2005). When light is applied to
Synechococcus cells that have lost the activities of PSI and
PSII as a result of salt stress in darkness, these activities are
fully restored. Light also restores the activity of the Na+/
H+ antiporters in Synechococcus. Weak light at 70 lE
m-2 s-1 is sufficient to generate ATP, which seems to
support recovery. An uncoupler, either carbonyl cyanide
Fig. 3 A hypothetical model of the NaCl-induced inactivation of PSI
and PSII in cyanobacterial cells. d, Extrinsic proteins of the oxygen-
evolving machinery of PSII, namely, the 33-kD protein, cytochrome
c550, and PsbU; m, a protein associated with the PSI complex,
namely, plastocyanin or cytochrome c553; red circle, water channel;
yellow circle, K+ (Na+) channel; pink circle, Cl- channel; white
circle, Na+/H+ antiporter. Thylakoid lumen and cytoplasm are
indicated by white and green color, respectively. Adapted and
modified from Allakhverdiev et al. (2000b)
534 Photosynth Res (2008) 98:529–539
123

m-chlorophenylhydrazone (CCCP) or carbonyl cyanide p-
trifluoro-methoxyphenylhydrazone (FCCP), each of which
induces the de-energization of cells, prevents the light-
dependent restoration of the activities of PSII, PSI, and
Na+/H+ antiporters (Allakhverdiev et al. 2005). These
observations confirm that the light-dependent energization
of cyanobacterial cells is important for the tolerance of
photosystems to salt stress. This effect of light is com-
pletely eliminated in the presence of lincomycin, an
inhibitor of protein synthesis (Allakhverdiev et al. 2005).
Thus, it seems likely that, when cyanobacterial cells are
exposed to salt stress, protein synthesis is important for the
recovery of the photosystems and the Na+/H+ antiporters.
The data obtained in experiments with Synechococcus
(Allakhverdiev et al. 2005) and Synechocystis (Allakhver-
diev et al. 1999) are in accord with the observations of
Hagemann et al. (1991), who demonstrated a close corre-
lation between the synthesis of proteins and the tolerance to
salt stress in Synechocystis cells. The involvement of pro-
tein synthesis in recovery was confirmed by protein-
labeling experiments. Furthermore, light not only stimu-
lated the overall synthesis of proteins but also induced the
synthesis of several specific proteins, which were absent
under dark conditions where no recovery occurred (All-
akhverdiev et al. 2005).
The presence of exogenous glucose in darkness also
resulted in regeneration of the activity of Na+/H+ anti-
porters. It is likely that exogenously added glucose
activated respiration since a high, but only transient, stim-
ulation of oxygen consumption was observed in darkness
after the addition of glucose to a culture of Synechococcus
cells (Allakhverdiev et al. 2005). Both photosynthesis and
respiration generate energy, in the form of ATP and of
proton gradients across cytoplasmic and thylakoid mem-
branes. The hypothesis that cellular energization in light or
in the presence of glucose in darkness accelerates the
recovery of the activities of PSII, PSI, and Na+/H+ anti-
porters is supported by the actions of ionophores, namely,
CCCP, FCCP, nigericin plus valinomycin, and N,N0-dicy-
clohexyl-carbodiimide (DCCD) each of which inhibits the
synthesis of ATP. The addition of these compounds pre-
vents the recovery of the activities of PSII and PSI in light
and in the presence of exogenous glucose in darkness
(Allakhverdiev et al. 2005), suggesting that the synthesis of
ATP is essential for the recovery.
Unsaturation of fatty acids in membrane lipids
increases the tolerance of PSII to salt stress
The state of thylakoid membranes plays a prominent role in
the tolerance of the photosynthetic machinery to environ-
mental stresses, and in particular, to cold stress (Wada et al.
1990; Murata et al. 1992) and salt stress (Allakhverdiev
et al. 1999, 2001). The ratio of saturated and unsaturated
fatty acids and the phospholipid composition of mem-
branes are critical factors in this regard (Nishida and
Murata 1996; Sakurai et al. 2003, 2007). When photosyn-
thetic organisms are exposed to salt stress, fatty acids of
membrane lipids are desaturated (Huflejt et al. 1990).
Therefore, it is possible that unsaturation of fatty acid in
membrane lipids might affect the tolerance of the photo-
synthetic machinery to salt stress.
Tasaka et al. (1996) isolated a desA-/desD- mutant
strain of Synechocystis, in which the desA and desD genes
for the D12 and D6 desaturases, respectively, had been
inactivated by targeted mutagenesis. The desA-/desD-
mutant cells contained mono-unsaturated, but not polyun-
saturated, fatty acids whereas wild-type cells contain
polyunsaturated fatty acids, such as di-, tri-, and tetra-
unsaturated fatty acids (Wada and Murata 1989). By
comparing these two strains, Tasaka et al. (1996) demon-
strated that polyunsaturated fatty acids are essential for
growth in Synechocystis at low temperatures. We examined
the contribution of the unsaturation of fatty acids in
membrane lipids to the tolerance of Synechocystis cells to
salt stress by comparing desA-/desD- mutant cells with
wild-type cells (Allakhverdiev et al. 1999). The loss of the
oxygen-evolving activity of PSII in the presence of 0.5 M
NaCl was much more rapid in desA-/desD- cells than in
wild-type cells. The oxygen-evolving activity that had been
lost during incubation with 0.5 M NaCl recovered when
cells were transferred to conditions that allowed photo-
synthesis in light or respiration as a result of the addition of
glucose to the medium. This recovery was much faster in
wild-type than in desA-/desD- cells, and in addition, it
was prevented by lincomycin (Allakhverdiev et al. 1999).
The relationship between the unsaturation of fatty acids
and the tolerance of PSII to salt stress was examined after a
genetically engineered increase in the unsaturation of fatty
acids in the membrane lipids of Synechococcus (Wada et al.
1990; Allakhverdiev et al. 2001). Wild-type cells of Syn-
echococcus synthesize saturated and mono-unsaturated
fatty acids, whereas desA+ cells, which had been trans-
formed with the desA gene for the D12 acyl-lipid desaturase
of Synechocystis, synthesized di-unsaturated fatty acids in
addition to mono-unsaturated fatty acids. Loss of the
activities of PSI and PSII and of the Na+/H+ antiport sys-
tem during incubation with 0.5 M NaCl occurred more
slowly in desA+ cells than in wild-type cells. In addition, we
observed that the PSII activity of isolated thylakoid mem-
branes from desA+ cells was more resistant to 0.5 M NaCl
than that of isolated membranes from wild-type cells (All-
akhverdiev et al. 2001). These findings indicate that the
unsaturation of fatty acids in membrane lipids enhances the
tolerance of PSI and PSII to salt stress.
Photosynth Res (2008) 98:529–539 535
123

There are at least five possible explanations for the
enhanced tolerance of PSII to salt stress upon the unsatu-
ration of membrane lipids, as follows: (i) Water channels,
the activity of which is responsible for the rapid phase of the
NaCl-induced inactivation, are located on the plasma
membrane. Therefore, it is quite plausible that their activity
might be affected by the unsaturation of membrane lipids
and by resultant changes in the fluidity of the membrane.
Such effects might explain why the unsaturation of fatty
acids minimized the rapid phase of the NaCl-induced
inactivation; (ii) K+(Na+) channels are located on the
plasma membrane, and their activities might be enhanced
by the unsaturation of fatty acids of membrane lipids. Such
effects might explain why the unsaturation of fatty acids
counteracted the slow phase of NaCl-induced inactivation;
(iii) The Na+/H+ antiport system, which consists of Na+/
H+ antiporter(s) and H+-ATPase(s), is located in the plasma
membrane. The unsaturation of fatty acids in membrane
lipids might activate the Na+/H+ antiport system via
enhanced fluidity of the membrane, resulting in a decrease
in concentrations of Na+ ions in the cytosol; (iv) The un-
saturation of fatty acids might stimulate the synthesis of the
Na+/H+ antiporter(s) and/or H+-ATPase(s). An increase in
the levels of these components of the antiport system might
result in a decrease in the concentration of Na+ ions in the
cytosol, which would tend to protect PSII and PSI against
NaCl-induced inactivation; (v) The repair of PSII includes a
step of insertion of the precursor to the D1 protein into the
membrane. The unsaturation of fatty acids might accelerate
the insertion, resulting in the enhanced activity of repair.
Synergistic actions of salt stress and light stress
Light stress and salt stress are major environmental factors
that limit the efficiency of photosynthesis. However, the
mechanisms of their effects on PSII are different. These
effects of salt stress and light stress on PSII can be sepa-
rately examined much more easily in cyanobacterial cells
than in leaves of higher plants. PSII of Synechocystis is
resistant to salt stress and the light stress, when each is
applied separately at certain moderate levels (Allakhver-
diev et al. 1999, 2002). However, when they are combined,
they inhibit PSII very efficiently. Moreover, this synergistic
effect is due to inhibition of the repair of PSII from light-
induced inactivation (Allakhverdiev et al. 2002). Labeling
of proteins in vivo and Western and Northern blotting
analyses indicated that salt stress inhibited the translation
and, to a much smaller extent, the transcription of the psbA
genes, which encode the D1 protein (Allakhverdiev et al.
2002). Pulse-chase experiments revealed that salt stress
inhibited the synthesis of D1 de novo in Synechococcus
(Ohnishi and Murata 2006).
Recent studies of photoinhibition of PSII in cyanobac-
teria suggest that oxidative stress due to reactive oxygen
species (ROS) inhibits the repair of PSII but does not
stimulate the photodamage to PSII (Nishiyama et al. 2005,
2006; Murata et al. 2007). The site of action of ROS seems
to be the elongation factor G (EFG) of the translation of
gene expression (Kojima et al. 2007). A further study has
suggested that it is likely that various kinds of stress, such
as salt and moderate heat stress, suppress the fixation of
CO2 and, thus, decrease utilization of NADPH that would
stimulate the formation of ROS (Allakhverdiev et al. 2007;
Takahashi and Murata 2008). These ROS, in turn, inhibit
protein synthesis and, thus, the repair of PSII (see also
Murata et al. 2007; Takahashi and Murata 2008). Another
possibility is that salt stress directly inactivates the trans-
lation machinery by dissociation of its subunits. Further
studies are necessary for a full understanding of the
mechanisms responsible for the effects of salt stress on
photoinhibition of photosynthesis.
In tobacco leaves, Al-Taweel et al. (2007) observed that
salt stress had no effect on photodamage to PSII but did
inhibit the repair of photodamaged PSII via inhibition of
protein synthesis in chloroplasts. In higher plants also, salt
stress seemed to inhibit the fixation of CO2 and decreased
utilization of NADPH that would stimulate the formation
of ROS in PSI (see also Takahashi and Murata 2008).
Compatible solutes increase the tolerance
of the photosynthetic machinery to salt stress
Many plants accumulate compatible solutes in response to
salt stress (Chen and Murata 2002). These compatible sol-
utes include proline, betaine, polyols, sugar alcohols, and
soluble sugars (Fulda et al. 1999). The expression of genes
that are involved in the biosynthesis of compatible solutes is
enhanced under salt stress, but not by osmotic stress (Marin
et al. 2006), and concentrations of accumulated compatible
solutes are correlated with the extent of tolerance to salt
stress (Zhu 2002). Glycinebetaine acts as a compatible
solute by stabilizing the quaternary structures of proteins,
cell membranes and, in particular, the oxygen-evolving
complex of PSII (Papageorgiou and Murata 1995).
Synechocystis cells accumulate glucosylglycerol (GG) as
a major compatible solute and they also accumulate traces of
sucrose transiently in response to salt stress. These com-
patible solutes allow cells to tolerate as much as 1.2 M NaCl
(Reed and Stewart 1988). Synechocystis cells synthesize
GG-phosphate from ADP-glucose and glycerol 3-phosphate
in a reaction catalyzed by GG-phosphate synthase (GgpS),
and they dephosphorylate the intermediate GG-phosphate
to yield GG in a reaction catalyzed by GG-phosphate
phosphatase (Hagemann and Erdmann 1994). GgpS and
536 Photosynth Res (2008) 98:529–539
123

GG-phosphate phosphatase are encoded by the ggpS and
stpA genes, respectively (Hagemann et al. 1997). Prior to salt
stress, Synechocystis cells do not synthesize GG. After
incubation of cells in darkness for 20 to 30 h in the presence
of 0.5 or 1.0 M NaCl, Synechocystis cells accumulate GG at
a level as high as 180–200 lg per mg of Chl, suggesting a
protective role for this compound at high concentrations of
salt in light (Hagemann and Erdmann 1994).
Ferjani et al. (2003) demonstrated that the presence of
GG is important in the tolerance of the cell-division
machinery to salt stress. In DggpS mutant cells of Syn-
echocystis, NaCl at 0.45 M inhibited cell division and
significantly increased cell size, whereas in wild-type cells,
cell division and increases in cell size were inhibited at
high concentrations of NaCl, such as 0.8 M. Electron
microscopy revealed that, in DggpS cells, separation of
daughter cells was incomplete, and aborted division could
be recognized by the presence of a structure that resembled
a division ring. The addition of GG to the culture medium
protected the mutant cells from salt stress and reversed the
adverse effects of NaCl on cell division and cell size. These
observations suggest that GG is important for salt tolerance
and, thus, for the proper division of cells under salt stress.
Our group demonstrated that salt stress due to 0.22 M
NaCl represses the repair of photodamaged PSII by
inhibiting the degradation and synthesis of D1 protein in
salt-stressed Synechococcus cells (Ohnishi and Murata
2006). We had previously transformed Synechococcus cells
with the codA gene for choline oxidase, which catalyzes
the synthesis of glycinebetaine from choline (Deshnium
et al. 1995). The transformed cells accumulated glycineb-
etaine at a concentration between 60 and 80 mM. Salt
stress might reduce the integrity and activity of the trans-
lational machinery in Synechococcus cells, and the
synthesis of glycinebetaine in the codA-transformed cells
might counteract the effects of salt stress. In addition, salt
stress might destabilize and glycinebetaine might stabilize
proteases that are required for the degradation of D1 and
other proteins in PSII (Ohnishi and Murata 2006). In this
respect, it is noted that FtsH, a bacterial homolog of such
proteases, is active when it is in a hexameric structure
(Suno et al. 2006), which might be destabilized by salt and
stabilized by the presence of glycinebetaine.
Conclusion
Changes in cytoplasmic volume and in the activities of PSI
and PSII suggest that NaCl has both osmotic and ionic
effects on cyanobacterial cells. The osmotic effect elicits
the efflux of water from the cytoplasm through water
channels and, thus, most probably increases cytoplasmic
concentrations of cations and anions, such as K+ ions.
These increases in concentrations of ions in the cytosol, in
turn, increase the concentrations of ions in the lumen (in-
trathylakoid space) with resultant dissociation of the
extrinsic proteins, which can lead to the reversible inacti-
vation of both PSI and PSII.
The ionic effect is caused by an influx of Na+ and Cl-
ions across the plasma membrane through K+(Na+) and
Cl- channels, respectively, and it increases the concen-
tration of NaCl in the cytosol. NaCl penetrates the
thylakoid membranes by an unknown mechanism to
increase the concentration of NaCl in the lumen. As a
result, the extrinsic proteins dissociate from both PSII and
PSI, with the resultant irreversible inactivation of PSI and
PSII after long-term incubation. The restoration and
recovery of the photosystems and the Na+/H+ antiporters
in salt-stressed cells might be regulated by the synthesis of
ATP and the subsequent synthesis of proteins.
Light and salt stress have a strong synergistic effect on
photoinhibition of PSII. Salt stress inhibits the repair of PSII
from light-induced inactivation. Synthesis of compatible
solutes, such as glucosylglycerol and glycinebetaine, and
the unsaturation of fatty acids in membrane lipids are key
factors in the repair of and the tolerance to salt stress of
PSII. Hence, a full understanding of the molecular basis of
the mechanisms of tolerance of photosystems to salt stress is
essential for the introduction, by plant breeding and genetic
engineering, of salt tolerance in crop plants. A particularly
promising approach in efforts to increase the ability of
plants to tolerate high concentrations of salt is genetic
engineering that increases levels of compatible solutes and
levels of unsaturation of membrane lipids.
Acknowledgments This work was supported, in part, by the
Cooperative Research Program on Stress-Tolerant Plants of the
National Institute for Basic Biology, Japan, and by grants from
the Russian Foundation for Basic Research (Nos. 08-04-00241 and
08-04-91300) and from the Molecular and Cell Biology Programs of
the Russian Academy of Sciences (to S.I.A.).
References
Adir N, Zer H, Shochat S, Ohad I (2003) Photoinhibition a historical
perspective. Photosynth Res 76:343–370. doi:10.1023/A:1024969
518145
Allakhverdiev SI, Murata N (2004) Environmental stress inhibits the
synthesis de novo of proteins involved in the photodamage-
repair cycle of photosystem II in Synechocystis sp. PCC 6803.
Biochim Biophys Acta 1657:23–32. doi:10.1016/j.bbabio.2004.
03.003
Allakhverdiev SI, Nishiyama Y, Suzuki I, Tasaka Y, Murata N (1999)
Genetic engineering of the unsaturation of fatty acids in
membrane lipids alters the tolerance of Synechocystis to salt
stress. Proc Natl Acad Sci USA 96:5862–5867. doi:
10.1073/pnas.96.10.5862
Allakhverdiev SI, Sakamoto A, Nishiyama Y, Murata N (2000a)
Inactivation of photosystems I and II in response to osmotic
Photosynth Res (2008) 98:529–539 537
123

stress in Synechococcus: Contribution of water channels. Plant
Physiol 122:1201–1208. doi:10.1104/pp.122.4.1201
Allakhverdiev SI, Sakamoto A, Nishiyama Y, Inaba M, Murata N
(2000b) Ionic and osmotic effects of NaCl-induced inactivation
of photosystems I and II in Synechococcus sp. Plant Physiol
123:1047–1056. doi:10.1104/pp.123.3.1047
Allakhverdiev SI, Kinoshita M, Inaba M, Suzuki I, Murata N (2001)
Unsaturated fatty acids in membrane lipids protect the photosyn-
thetic machinery against salt-induced damage in Synechococcus.
Plant Physiol 125:1842–1853. doi:10.1104/pp.125.4.1842
Allakhverdiev SI, Nishiyama Y, Miyairi S, Yamamoto H, Inagaki N,
Kanesaki Y et al (2002) Salt stress inhibits the repair of
photodamaged photosystem II by suppressing the transcription
and translation of psbA genes in Synechocystis. Plant Physiol
130:1443–1453. doi:10.1104/pp.011114
Allakhverdiev SI, Klimov VV, Hagemann M (2005) Cellular
energization protects the photosynthetic machinery against salt-
induced inactivation in Synechococcus. Biochim Biophys Acta
1708:201–208. doi:10.1016/j.bbabio.2005.01.002
Allakhverdiev SI, Deshnium P, Mohanty P, Murata N (2007)
Glycinebetaine alleviates the inhibitory effect of moderate heat
stress on the repair of photosystem II during photoinhibition.
Biochim Biophys Acta-Bioenergetics 1767:1363–1371
Al-Taweel K, Iwaki T, Yabuta Y, Shigeoka S, Murata N, Wadano A
(2007) A bacterial transgene for catalase protects translation of D1
protein during exposure of salt-stressed tobacco leaves to strong
light. Plant Physiol 145:258–265. doi:10.1104/pp.107.101733
Aro E-M, Virgin I, Andersson B (1993) Photoinhibition of photosystem
II: inactivation, protein damage and turnover. Biochim Biophys
Acta 1143:113–134. doi:10.1016/0005-2728(93)90134-2
Asada K (2006) Production and scavenging of reactive oxygen
species in chloroplasts and their functions. Plant Physiol
141:391–396. doi:10.1104/pp.106.082040
Bhagwat AA, Apte SK (1989) Comparative analysis of proteins
induced by heat shock, salinity, and osmotic stress in the
nitrogen-fixing cyanobacterium Anabaena sp. strain L-31. J
Bacteriol 171:5187–5189
Blumwald E, Wolosin JM, Packer L (1984) Na+/H+ exchange in the
cyanobacterium Synechococcus 6311. Biochem Biophys Res
Commun 22:452–459. doi:10.1016/0006-291X(84)90497-2
Bohnert HJ, Ayoubi P, Borchert C, Bressan RA, Burnap RL,
Cushman JC et al (2001) A genomics approach towards salt
stress tolerance. Plant Physiol Biochem 39:295–311. doi:
10.1016/S0981-9428(00)01237-7
Chen THH, Murata N (2002) Enhancement of tolerance of abiotic
stress by metabolic engineering of betaines and other compatible
solutes. Curr Opin Plant Biol 5:250–257. doi:10.1016/S1369-
5266(02)00255-8
Demmig-Adams B, Adams WWIII (1992) Photoprotection and other
responses of plants to high light stress. Annu Rev Plant Physiol
Plant Mol Biol 43:599–626. doi:10.1146/annurev.pp.43.060192.
003123
Demmig-Adams B, Adams WWIII (2002) Antioxidants in photosyn-
thesis and human nutrition. Science 298:2149–2153. doi:
10.1126/science.1078002
Deshnium P, Los DA, Hayashi H, Mustardy L, Murata N (1995)
Transformation of Synechococcus with a gene for choline
oxidase enhances tolerance to salt stress. Plant Mol Biol
29:897–907. doi:10.1007/BF00014964
Ferjani A, Mustardy I, Sulpice R, Marin K, Suzuki I, Hagemann M,
Murata N (2003) Glucosylglycerol, a compatible solute, sustains
cell division under salt stress. Plant Physiol 131:1628–1637. doi:
10.1104/pp.102.017277
Fulda S, Huckauf J, Schoor A, Hagemann M (1999) Analysis of stress
responses in the cyanobacterial strains Synechococcus sp. PCC
7942, Synechocystis sp. PCC 6803, Synechococcus sp. PCC:7418
Osmolyte accumulation and stress protein synthesis. J Plant
Physiol 154:240–249
Golbeck JH (1994) Photosystem I in cyanobacteria. In: Bryant DA
(ed) The molecular biology of cyanobacteria. Kluwer Academic
Publishers, Dordrecht, The Netherlands, pp 319–360
Hagemann M, Erdmann N (1994) Activation and pathway of
glucosylglycerol synthesis in the cyanobacterium Synechocystissp. PCC 6803. Microbiology 140:1427–1431
Hagemann M, Erdmann N (1997) Environmental stresses. In: Rai AK
(ed) Cyanobacterial Nitrogen Metabolism and Environmental
Biotechnology. Springer-Verlag, Heidelberg, pp 156–221
Hagemann M, Wolfel L, Kruger B (1990) Alterations of protein
synthesis in the cyanobacterium Synechocystis sp. PCC 6803
after a salt shock. J Gen Microbiol 136:1393–1399
Hagemann M, Techel D, Rensing L (1991) Comparison of salt- and
heat-induced alterations of protein synthesis in the cyanobacte-
rium Synechocystis sp. PCC 6803. Arch Microbiol 155:587–592.
doi:10.1007/BF00245354
Hagemann M, Schoor A, Jeanjean R, Zuther E, Joset F (1997) The
stpA gene from Synechocystis sp. strain PCC 6803 encodes the
glucosylglycerolphosphate phosphatase involved in cyanobacte-
rial osmotic response to salt shock. J Bacteriol 179:1727–1733
Hayashi H, Murata N (1998) Genetically engineered enhancement of
salt tolerance in higher plants. In: Sato K, Murata N (eds) Stress
responses of photosynthetic organisms. Molecular mechanisms
and molecular regulation. Elsevier, Amsterdam, pp 133–148
Hibino T, Lee BH, Rai AK, Ishikawa H, Kojima H, Tawada H et al
(2000) Salt enhances photosystem I content and cyclic electron
flow via NAD(P)H dehydrogenase in the halotolerant cyanobac-
terium Aphanothece halophytica. Aust J Plant Physiol 23:321–330
Huflejt ME, Tremolieres A, Pineau B, Lang JK, Hatheway J, Packer L
(1990) Changes in membrane lipid composition during saline
growth of the fresh water cyanobacterium Synechococcus 6311.
Plant Physiol 94:1512–1521
Inaba M, Sakamoto A, Murata N (2001) Functional expression in
Escherichia coli of low-affinity and high-affinity Na+(Li+)/H+
antiporters of Synechocystis. J Bacteriol 183:1376–1384. doi:
10.1128/JB.183.4.1376-1384.2001
Izawa S (1980) Acceptors and donors for chloroplast electron
transport. Methods Enzymol 69:413–435. doi:10.1016/S0076-
6879(80)69041-7
Joset F, Jeanjean R, Hagemann M (1996) Dynamics of the response of
cyanobacteria to salt stress: deciphering the molecular events.
PhysiolPlant 96:738–744. doi:10.1111/j.1399-3054.1996.tb00251.x
Kaneko T, Sato S, Kotani H, Tanaka A, Asamizu E, Nakamura Y et al
(1996) Sequence analysis of the genome of the unicellular
cyanobacterium Synechocystis sp. PCC 6803. II. Sequence deter-
mination of the entire genome and assignment of potential protein-
coding regions. DNA Res 3:109–136. doi:10.1093/dnares/3.3.109
Kanesaki Y, Suzuki I, Allakhverdiev SI, Mikami K, Murata N (2002)
Salt stress and hyperosmotic stress regulate the expression of
different sets of genes in Synechocystis sp. PCC 6803. Biochem
Biophys Res Commun 290:339–348. doi:10.1006/bbrc.2001.6201
Kojima K, Oshita M, Nanjo Y, Kasai K, Tozawa Y, Hayashi H et al
(2007) Oxidation of elongation factor G inhibits the synthesis of
the D1 protein of photosystem II. Mol Microbiol 65:936–947.
doi:10.1111/j.1365-2958.2007.05836.x
Kuwabara T, Murata N (1983) Quantitative analysis of the inactiva-
tion of photosynthetic oxygen evolution and the release of
polypeptides and manganese in the photosystem II particles of
spinach chloroplasts. Plant Cell Physiol 24:741–747
Marin K, Stirnberg M, Eisenhut M, Kramer R, Hagemann M (2006)
Osmotic stress in Synechocystis sp. PCC 6803: low tolerance
towards nonionic osmotic stress results from lacking activation
of glucosylglycerol accumulation. Microbiology 152:2023–
2030. doi:10.1099/mic.0.28771-0
538 Photosynth Res (2008) 98:529–539
123

Miyao M, Murata N (1983) Partial disintegration and reconstitution of
the photosynthetic oxygen-evolution system: binding of 24 kD
and 18 kD polypeptides. Biochim Biophys Acta 725:87–93. doi:
10.1016/0005-2728(83)90227-X
Mohanty P, Allakhverdiev SI, Murata N (2007) Application of low
temperatures during photoinhibition allows characterization of
individual steps in photodamage and the repair of photosystem
II. Photosynth Res 94:217–224. doi:10.1007/s11120-007-9184-y
Murata N, Miyao M (1985) Extrinsic membrane proteins in the
photosynthetic oxygen-evolving complex. Trends Biochem Sci
10:122–124. doi:10.1016/0968-0004(85)90272-5
Murata N, Ishizaki-Nishizawa O, Higashi S, Hayashi H, Tasaka Y,
Nishida I (1992) Genetically engineered alteration in the
chilling sensitivity of plants. Nature 356:710–713. doi:10.1038/
356710a0
Murata N, Takahashi S, Nishiyama Y, Allakhverdiev SI (2007)
Photoinhibition of photosystem II under environmental stress.
Biochim Biophys Acta 1767:414–421. doi:10.1016/j.bbabio.
2006.11.019
Nakamura T, Yuda R, Unemoto T, Bakker EP (1998) KtrAB, a new
type of bacterial K+ -uptake system from Vibrio alginolyticus. J
Bacteriol 180:3491–3494
Nishida I, Murata N (1996) Chilling sensitivity in plants and
cyanobacteria: the crucial contribution of membrane lipids.
Annu Rev Plant Physiol Plant Mol Biol 47:541–568. doi:
10.1146/annurev.arplant.47.1.541
Nishiyama Y, Los DA, Hayashi H, Murata N (1997) Thermal
protection of the oxygen-evolving machinery by PsbU, an
extrinsic protein of photosystem II in Synechococcus sp PCC
7002. Plant Physiol 115:1473–1480. doi:10.1104/pp.115.4.1473
Nishiyama Y, Los DA, Murata N (1999) PsbU, a protein associated
with photosystem II, is required for the acquisition of cellular
thermotolerance in Synechococcus sp PCC 7002. Plant Physiol
120:301–308. doi:10.1104/pp.120.1.301
Nishiyama Y, Allakhverdiev SI, Murata N (2005) Inhibition of the
repair of photosystem II by oxidative stress in cyanobacteria.
Photosynth Res 84:1–7. doi:10.1007/s11120-004-6434-0
Nishiyama Y, Allakhverdiev SI, Murata N (2006) A new paradigm
for the action of reactive oxygen species in the photoinhibition of
photosystem II. Biochim Biophys Acta 1757:742–749. doi:
10.1016/j.bbabio.2006.05.013
Ohnishi N, Murata N (2006) Glycinebetaine counteracts the inhib-
itory effects of salt stress on the degradation and synthesis of the
D1 protein during photoinhibition in Synechococcus sp. PCC
7942. Plant Physiol 141:758–765. doi:10.1104/pp.106.076976
Oquist G, Campbell D, Clarke A, Gustafsson P (1995) The
cyanobacterium Synechococcus modulates photosystem II func-
tion in response to excitation stress through D1 exchange.
Photosynth Res 46:151–158. doi:10.1007/BF00020425
Paithoonrangsarid K, Shoumskaya MA, Kanesaki Y, Satoh S, Tabata
S, Los DA et al (2004) Five histidine kinases perceive osmotic
stress and regulate distinct sets of genes in Synechocystis. J Biol
Chem 279:53078–53086. doi:10.1074/jbc.M410162200
Papageorgiou GC, Murata N (1995) The unusually strong stabilizing
effects of glycine betaine on the structure and function of the
oxygen-evolving photosystem II complex. Photosynth Res
44:243–252. doi:10.1007/BF00048597
Pfenning N (1978) General physiology and ecology of photosynthetic
bacteria. In: Clayton RK, Sistrom WR (eds) The photosynthetic
bacteria. Plenum Press, New York, pp 3–18
Powles SB (1984) Photoinhibition of photosynthesis induced by
visible light. Annu Rev Plant Physiol 35:15–44. doi:
10.1146/annurev.pp.35.060184.000311
Reed RH, Stewart WDP (1988) The responses of cyanobacteria to salt
stress. In: Rogers LJ, Gallan LJ (eds) Biochemistry of the algae
and cyanobacteria, vol 12. Clarendon Press, Oxford, pp 217–231
Reed RH, Warr SRC, Richardson DL, Moore DJ, Stewart WDP
(1985) Multiphasic osmotic adjustment in a euryhaline cyano-
bacterium. FEMS Microbiol Lett 28:225–229. doi:10.1111/j.
1574-6968.1985.tb00796.x
Sakurai I, Hagio M, Gombos Z, Tyystjarvi T, Paakkarinen V, Aro
E-M et al (2003) Requirement of phosphatidylglycerol for
maintenance of photosynthetic machinery. Plant Physiol
133:376–1384. doi:10.1104/pp.103.026955
Sakurai I, Mizusawa N, Ohashi S, Kobayashi M, Wada H (2007)
Effects of the lack of phosphatidylglycerol on the donor side of
photosystem II. Plant Physiol 144:1336–1346. doi:10.1104/pp.
107.098731
Shapiguzov A, Lyukevich AA, Allakhverdiev SI, Sergeyenko TV,
Suzuki I, Murata N et al (2005) Osmotic shrinkage of cells of
Synechocystis sp. PCC 6803 by water efflux via aquaporins
regulates the osmostress-inducible gene expression. Microbiol-
ogy 151:447–455. doi:10.1099/mic.0.27530-0
Shen J-R, Ikeuchi M, Inoue Y (1992) Stoichiometric association of
extrinsic cytochrome c550 and 12 kDa protein with a highly
purified oxygen-evolving photosystem II core complex from
Synechococcus vulcanus. FEBS Lett 301:145–149. doi:10.1016/
0014-5793(92)81235-E
Shen J-R, Qian M, Inoue Y, Burnap RL (1998) Functional charac-
terization of Synechocystis sp. PCC 6803 DpsbU and DpsbVmutants reveals important roles of cytochrome c-550 in cyano-
bacterial oxygen evolution. Biochemistry 37:1551–1558. doi:
10.1021/bi971676i
Shoumskaya MA, Paithoonrangsarid K, Kanesaki Y, Los DA,
Zinchenko VV, Tanticharoen M et al (2005) Identical Hik-Rre
systems are involved in perception and transduction of salt
signals and hyperosmotic signals but regulate the expression of
individual genes to different extents in Synechocystis. J Biol
Chem 280:21531–21538. doi:10.1074/jbc.M412174200
Suno R, Niwa H, Tsuchiya D, Zhang X, Yoshida M, Morikawa K
(2006) Structure of the whole cytosolic region of ATP-dependent
protease FtsH. Mol Cell 22:575–585. doi:10.1016/j.molcel.
2006.04.020
Takahashi S, Murata N (2008) How do environmental stresses
accelerate photoinhibition? Trends Plant Sci 13:178–182. doi:
10.1016/j.tplants.2008.01.005
Tasaka Y, Gombos Z, Nishiyama Y, Mohanty P, Ohba T, Ohki K et al
(1996) Targeted mutagenesis of acyl-lipid desaturases in
Synechocystis: evidence for the important roles of polyunsatu-
rated membrane lipids in growth, respiration and photosynthesis.
EMBO J 15:6416–6425
Trebst A (1980) Inhibitors in electron flow: tools for the functional
and structural localization of carriers and energy conservation
sites. Methods Enzymol 69:675–715. doi:10.1016/S0076-6879
(80)69067-3
Wada H, Murata N (1989) Synechocystis PCC 6803 mutants defective
in desaturation of fatty acids. Plant Cell Physiol 30:971–978
Wada H, Gombos Z, Murata N (1990) Enhancement of chilling
tolerance of a cyanobacterium by genetic manipulation of fatty
acid desaturation. Nature 347:200–203. doi:10.1038/347200a0
Yamashita T, Butler WL (1969) Inhibition of the Hill reaction by Tris
and restoration by electron donation to photosystem II. Plant
Physiol 44:435–438
Zhu J-K (2002) Salt and drought stress signal transduction in plants.
Annu Rev Plant Biol 53:247–273. doi:10.1146/annurev.arplant.
53.091401.143329
Photosynth Res (2008) 98:529–539 539
123
Related Documents



![STRUCTURE AND FUNCTION OF PHOTOSYSTEMS I …...by two photosystems [photosystem I (PSI) and photosystem II (PSII)], an ATP synthase (F-ATPase) that produces ATP at the expense of the](https://static.cupdf.com/doc/110x72/5e6a9bf3b881810a8b6cdf92/structure-and-function-of-photosystems-i-by-two-photosystems-photosystem-i.jpg)







