1 23 Gesunde Pflanzen Pflanzenschutz - Verbraucherschutz - Umweltschutz ISSN 0367-4223 Volume 67 Number 1 Gesunde Pflanzen (2015) 67:21-31 DOI 10.1007/s10343-014-0333-x Alleviation of Salt Stress Adverse Effect and Enhancing Phenolic Anti-oxidant Content of Eggplant by Seaweed Extract Amira M. Hegazi, Amal M. El-Shraiy & A. A. Ghoname

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 23
Gesunde PflanzenPflanzenschutz - Verbraucherschutz -Umweltschutz ISSN 0367-4223Volume 67Number 1 Gesunde Pflanzen (2015) 67:21-31DOI 10.1007/s10343-014-0333-x
Alleviation of Salt Stress Adverse Effect andEnhancing Phenolic Anti-oxidant Contentof Eggplant by Seaweed Extract
Amira M. Hegazi, Amal M. El-Shraiy &A. A. Ghoname

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

1 3
Original article
Received: 1 September 2014 / Accepted: 27 October 2014 / Published online: 19 November 2014© Springer-Verlag Berlin Heidelberg 2014
Alleviation of Salt Stress Adverse Effect and Enhancing Phenolic Anti-oxidant Content of Eggplant by Seaweed Extract
Amira M. Hegazi · Amal M. El-Shraiy · A. A. Ghoname
Gesunde Pflanzen (2015) 67:21–31DOI 10.1007/s10343-014-0333-x
ferent salinity stress levels, the activity of SOD and APX, significantly increased K content. These results provide support for the field application of bio-stimulants and an-tioxidant compounds to alleviate the symptoms and effects of salty soils.
Keywords Salinity · Salt stress · Eggplant
Minderung des unerwünschten Salzstresseffekts und Erhöhung des Gehalts an phenolischen Antioxidantien in Auberginen durch Meeresalgenextrakt
Zusammenfassung Der Salzgehalt entweder im Boden oder im Gießwasser mindert das Wachstum, den Ertrag sowie die Qualität verschiedener Gemüsesorten, wie z. B. der Aubergine. Um die unerwünschten Auswirkungen von Salzstress auf das Wachstum und den Ertrag von Auber-ginen zu reduzieren, wurden an der Experimental Station Farm, Faculty of Agriculture, Ain Shams University, in zwei aufeinanderfolgenden Sommern (2011 und 2012) Topfexperimente durchgeführt. Zum Experiment gehörten neun Behandlungen mit einer Kombination aus drei Salz-gehalten im Gießwasser, nämlich 320, 3200 und 4800 ppm Natriumchlorid und drei Meeresalgenextraktkonzentra-tionen (Ascophyllum nodosum) von Acadian Sea Plants Limited, Dartmouth, Nova Scotia, Kanada) (0, 2,5 und 5,0 cm3 L−1).
Der Salzstress (320, 3200 und 4800 ppm) minderte si-gnifikant das vegetative Wachstum und reduzierte den Ge-samtertrag sowie die Fruchtqualität. Die Anwendung von Biostimulanzien minderte diese negativen Auswirkungen mäßiger Salzstresswerte (3200 ppm) und wirkten teilwei-se den schädlichen Effekten des Salzgehalts bei höchstem Salzstress (4800 ppm) entgegen. Meeresalgenextrakt in
Abstract Salinity either of soil or of irrigation water causes restriction in growth, yield, and quality of sever-al vegetable crops such as eggplant. To alleviate the ad-verse effect of salt stress on the growth and fruit yield of eggplant, a pot experiment was conducted at the Experi-mental Station Farm, Faculty of Agriculture, Ain Shams University, during two successive summer seasons (2011 and 2012). The experiment included nine treatments which were the combination of three salinity levels of irrigation solutions (320, 3200, and 4800 ppm sodium chloride), and three levels of seaweed (Ascophyllum nodosum) extract levels from Acadian Sea plants Limited, Dartmouth, Nova Scotia, Canada(0, 2.5, and 5.0 cm3 L−1).
Salt stress (320, 3200, and 4800 ppm) significantly decreased vegetative growth and reduced total yield and fruit quality. Applied bio-stimulants positively allevi-ated the negative effects of moderate salinity stress lev-els (3200 ppm) and partially counteracted the harmful effects of salinity under the highest salinity stress level (4800 ppm). Sea weed extract (SWE) at highest concen-tration (5.0 cm3 L−1) was the most effective anti-salinity stress agents. Salinity stress levels increased super oxide dismutase (SOD), ascorbic peroxidase (APX) activity, and also Na content but decreased K content in the shoot and fruit of eggplants. In general, all of the applied seaweed slightly increased, either by itself or combined with dif-
A. A. Ghoname ()Vegetable Research Department, National Research Center,Dokki, Cairo, Egypte-mail: [email protected]
A. M. Hegazi · A. M. El-ShraiyAgric. Botany Department, Ain Shams Univ.,Cairo, Egypt
Author's personal copy

22
1 3
A. M. Hegazi et al.
der höchsten Konzentration (5,0 cm3 L−1) war als Anti-Stress-Wirkstoff gegen Salzstress am wirksamsten. Salz-stress erhöhte die Aktivität von Superoxiddismutase (SOD) und Ascorbatperoxidase (APX) sowie den Na-Gehalt und minderte den K-Gehalt in Sprossen und Früchten der Au-berginenpflanze. Allgemein erhöhte die Anwendung von Meeresalgen allein oder in Kombination mit verschiedenen Salzstresswerten leicht die Aktivität von SOD, APX sowie signifikant den K-Gehalt. Diese Ergebnisse unterstützen die Feldanwendung von Biostimulanzien und antioxidati-ven Verbindungen zur Minderung der Symptome und Aus-wirkungen salziger Böden.
Schlüsselwörter Salzgehalt · Salzstress · Auberginen
Introduction
Water scarcity and limitation of good quality water has become one of the major constraints for agricultural devel-opment in Egypt, especially after the commencement of con-structing the Grand Ethiopian Renaissance Dam. Hence, the increasing demands for irrigation water force the growers to utilize semi-saline underground water. The use of poor-quality ground water or brackish water has become inevi-table for irrigation to compensate rapidly increasing water requirements for irrigation and the competition between human and industrial water use, especially in arid and semi-arid regions (Katerji et al. 2000). Therefore, the use of saline or brackish water requires more knowledge and new inno-vative technologies and techniques with further research studies concerning the alleviation of salt stress, enhancing salt tolerance of cultivars, as well as water and soil manage-ment under saline conditions. However, the low quality of irrigation water (mainly saline) in combination with over-fertilization often causes problems and sharply reduces the productivity of eggplant (Bybordi et al. 2010).
Eggplant (Solanum melongena, L.) is a crop moderately sensitive to salt with a threshold of 1.1 dS/m and about 6.9 % of yield decrease when the electrical conductivity of irriga-tion water is increased by 1 dS/m (Yilmaz 1997). Eggplant is an important vegetable cash crop offering advantages to new Egyptian farmers in the new reclaimed lands where cli-matic conditions are rarely limiting. The importance of egg-plant is due to its higher yield and low productivity cost, as well as its higher antioxidant and nutritional value (Noreen and Ashraf 2009).
Several investigations have shown that cytotoxicity effects induced by salt stress can be alleviated by the exog-enous application of antioxidants (Sakhabutdinova et al. 2003) or by compounds that are able to enhance the natural defense systems of the plant (Demir et al. 1999; Saleh et al. 2010; El-Tohamy et al. 2012; El-Bassiony et al. 2012).
One of the most effective anti-stress compounds is sea-weed extract which is a bio-stimulant that has been used as soil conditioner for improving plant growth (Hurtado et al. 2009). Several studies have revealed the benefits of seaweed extracts on plants such as improved crop performance and yield, and better resistance to biotic and abiotic stress (Eyras et al. 2008; Norrie and Keathley 2006). The increased plant growth, yield, and quality due to seaweed extracts applica-tion, result from its influence on cell metabolism and induc-tion of synthesis of antioxidant molecules which could improve plant growth and stress resistance (Cardozo et al. 2007). Seaweed extracts also could improve nutrient avail-ability and productivity (Aziz et al. 2011). Seaweed extracts have also the ability to enhance stress tolerance in many plant species by increasing the concentration of bioactive molecules including antioxidants in treated plants (Fan et al. 2011; Rayirath et al. 2009).
The objective of the current study is to investigate the possibility of alleviating the negative effect of salt stress and improving growth and biochemical attributes of eggplant using seaweed extract.
Materials and Methods
Location and Growth Conditions
Two pot experiments were conducted at the Experimental Station Farm, Faculty of Agriculture, Ain Shams University, Cairo, Egypt, during two successive summer seasons (2011 and 2012). 45-day-old eggplant seedlings (cv. Black King) were obtained from a local commercial nursery where healthy seedlings of uniform size were selected and trans-planted on the 15th February, 2011, and 18th February, 2012, respectively, into pots (50 cm diameter) containing 10 kg of air-dried clay soil with two plants/pot. Before planting, each pot was supplied with nitrogen (N) as ammonium sulphate (20.5 % N) at 3 g per pot, phosphorous (P) as calcium super-phosphate (15.5 % P2O5) at 2 g per pot, and potassium (K) as potassium sulphate (K2O) at 1.5 g per pot. Likewise, further N (ammonium sulphate 20.5 % N) was added at 30, 60, and 120 days after transplanting at 2.5 g per pot.
Plant Material and Treatments
The experiment included three salinity levels of irrigation solutions (320, 3200, and 4800 ppm sodium chloride) and three levels of seaweed (Ascophyllum nodosum) extract levels (0, 2.5 and 5.0 cm3 L−1) from Acadian Sea plants Limited, Dartmouth, Nova Scotia, Canada. Irrigation solu-tions were supplied daily according to the plant’s need. The experimental soil had a sandy texture with pH of 7.6, and EC of 0.18 (Ds/m in soil paste), and the organic matter was
Author's personal copy

23
1 3
Alleviation of Salt Stress Adverse Effect and Enhancing Phenolic
(F.Wt.). In order to determine turgid weight (T.Wt.), leaves were floated in distilled water inside a closed petri dish. Leaf samples were weighed periodically, after gently wiping the water from the leaf surface with tissue paper until a steady state was achieved. At the end of the imbibition period, leaf samples were placed in a pre-heated oven at 80°C for 48 h, in order to determine dry weight (D.Wt.). Values of F.Wt., T.Wt., and D.Wt. were used to calculate LRWC using the equation below (Kaya et al. 2003):
LRWC (%) = [(F.Wt.–D.Wt.)/(T.Wt.–D.Wt.)] × 100
Fruit Yield and Yield Component
The fruits were harvested five times when having attained full size but still shining purple for fresh use. Fruit physical characteristics (fruit length and diameter), as well as fresh and dry weight were recorded. Total number of fruits per plant, early yield (the collection of the first two harvests as g/plant), and total fruit yield per plant were also recorded.
Bio-chemical Composition
The following chemical determinations were investigated in fruits, total soluble solids (TSS), total soluble sugar content, total phenols, tannins, super oxide dismutase (SOD) activ-ity, ascorbate peroxidase activity (APX), as well as potas-sium and sodium content.
Total soluble solids were determined by hand-held refrac-tometer AOAC (1975). Reducing and non-reducing sugars were extracted from 5 g crude dried material of the third ter-minal leaf using 70 % ethanol and kept overnight at room temperature according to Kayani et al. (1990), and then were filtered and recorded as total soluble sugar content (TSS).
Super oxide dismutase (SOD) activity was measured in fresh leaf samples collected in an ice bucket and brought to the laboratory. Leaves were washed with distilled water and then blotted dry. Leaf samples (0.5 g) were homogenized in an ice cold previously prepared 0.1 M pH 7.5 phosphate buffer containing 0.5 mM EDTA in a pre-chilled pestle and mortar. The homogenate was centrifuged at 15,000 x g for 15 min at 4 ºC. The supernatant was transferred to 30 mL tubes, and SOD activity was estimated by recording the decrease in absorbance of superoxide-nitro blue tetrazolium complex by the enzyme. About 3 mL of reaction mixture containing 0.1 mL of 1.5 M sodium carbonate, 0.2 mL of 200 mM methionine, 0.1 mL of 2.25 mM nitro-blue tetra-zolium, 0.1 mL of 3 mM EDTA, 1.5 mL of 100 mM potas-sium phosphate buffer, 1 mL distilled water, and 0.05 mL of enzyme were taken in test tubes in duplicate from each enzyme sample. Two tubes without enzyme extract were taken as control. The reaction was started by adding 0.1 mL riboflavin (60 mM) and placing the tubes below a light
0.19 % with 15, 9.40, and 16 mg/100 g soil of N, P, and K, respectively.
Plants were foliar-sprayed to runoff with seaweed extract at 30, 60, and 90 days after transplanting. Each experiment included three salinity levels and three exogenous spray treatments (nine treatments), replicated three times in a completely randomized design.
Sampling and Data recording
Vegetative Growth Parameters
In both growing seasons, five sample pots were taken at 75 days after transplanting, and the following vegetative growth characters were recorded: plant height (cm); num-ber of branches/plant; number of leaves/plant; shoot fresh weight (g); shoot dry weight (g); leaf area (cm2/plant).
Determination of Chlorophylls
Leaf discs (0.1 g) were taken from the interveinal areas of the fully opened sixth leaf from the top of three plants per replicate. Chlorophyll was extracted by grinding the tissue with a mortar and pestle using ammoniacal acetone (10 mL). The resulting extracts were centrifuged at 3000 × g for 3 min. The total chlorophyll concentrations were deter-mined by UV-vis spectrophotometry (CT 200 spectropho-tometer). The absorbance of the solution was measured at 470, 647, and 664 nm. Formulae and extinction coefficients used for the determination of chlorophyll were described by Lichtenthaler and Wellburn (1983).
Determination of Membrane Permeability
Twenty leaf discs (10 mm diameter) obtained from two plants per replicate from the young fully expanded leaves were placed in 50-ml glass vials and rinsed with distilled water to remove electrolytes released during leaf disc exci-sion. Vials were then filled with 30 mL of distilled water and allowed to stand in the dark for 24 h at room temperature. Electrical conductivity (EC1) of the bathing solution was determined at the end of the incubation period using an elec-trical conductivity meter (HANNA H199301). Vials were heated in a temperature-controlled water bath at 95°C for 20 min and then cooled to room temperature, and the electri-cal conductivity (EC2) was measured. Electrolyte leakage was calculated as percentage of EC1/EC2 (Shi et al. 2006).
Determination of Leaf Relative Water Content (LRWC)
Samples were taken from two plants per replicate (the sixth leaf from the top). Individual leaves were first detached from the stem and then weighed to determine fresh weight
Author's personal copy
24
1 3
A. M. Hegazi et al.
Determination of Tannins
Tannins of the dried fruits were determined using the modi-fied vanillin-hydrochloric acid (MV-HCl) as reported by Maxson and Rooney (1972). Samples of 1 g dried fruits were extracted with 1 % concentrated hydrochloric acid in methanol, and the samples were shaken on a reciprocating shaker for 24 h (the same length of time was used for all samples). 5 mL of vanillin-HCl reagent (50:50 mixtures of 4 % vanillin/8 % HCl in methanol) was quickly added to 1 mL fruit sample extract. The optical density of the devel-oped color was measured at 500 nm using the Shimadzu spectrophotometer model UV 1201. By plotting the optical densities against catechin concentrations, the standard curve was obtained and tannins % were calculated.
Potassium (K) and sodium (Na) contents were estimated by flame photometry (Peterburgski 1968).
The determination of membrane permeability (MP), leaf relative water content (LRWC) and enzymatic antioxidants, as well as bio-chemical compounds and nutrient elements was only carried out in the first experimental season.
Experimental Design and Statistical Analysis
The experimental design used in the two successive sea-sons was the complete randomized block design with four replicates. The analysis of variance was calculated using ANOVA, and the mean separation at 5 % level was done using the LSD test according to the method described by Gomez and Gomez (1984).
Results
Vegetative growth characters of eggplants (expressed as plant height, number of branches, number of leaves, and leaf area) were significantly decreased with the increase of salinity stress levels, with the greatest reduction observed at
source of two 15 w florescent lamps for 15 min. The reac-tion was stopped by switching off the light and covering the tube with a black cloth. Tubes without enzyme developed maximal color. A non-irradiated complete reaction mixture which did not develop color served as blank. Absorbance was recorded at 560 nm, and one unit of enzyme activity was taken as the quantity of enzyme which reduced the absorbance reading of samples 50 % in comparison with types lacking enzymes (Dhindsa et al. 1981).
Ascorbate peroxidase (APX) was assayed spectropho-tochemically according to Fielding (1978). The assay was carried out at 25 °C in a 1.0 cm light path cuvette, and the reaction mixture consisted of 1500 µL phosphate buf-fer, 20 µL EDTA, 1000 µL sodium ascorbate, and enzyme extract (20 µL). After mixing the reaction was initiated by adding the 480 µL H2O2, and absorbance was measured by spectrophotometer (CT 200 spectrophotometer) at 290 nm.
Determination of Total Polyphenols
Half gram of dried fruits was blended with 40 mL of 70 % aqueous acetone, boiled for 2−5 h under a condenser to extract the total polyphenols, and left to get cool. The mix-ture was then filtered using Wattman No. 1 filter paper, the residue was mixed with another 40 mL of 70 % aque-ous acetone, and the extraction process was repeated twice. The collected methanolic extracts were combined and com-pleted to a final volume of 100 mL. Ten drops of concen-trated hydrochloric acid were added to 1 mL of the extract, and then heated rapidly to boiling in a water bath for about 10 min. After cooling, 1 mL of the reagent and 1.5 mL of sodium carbonate solution (14 %) were added. The mixture was diluted to 10 mL with distilled water and then thor-oughly mixed. The reaction mixture was heated in boiling water for 5 min and then cooled, and the developing color was spectrophotometrically measured at 520 nm using the Folin-Ciocalteu reagent (Aldrich Chemical Co. St. Louis, MO, USA) as described by Wettasinghe and Shahidi (1999).
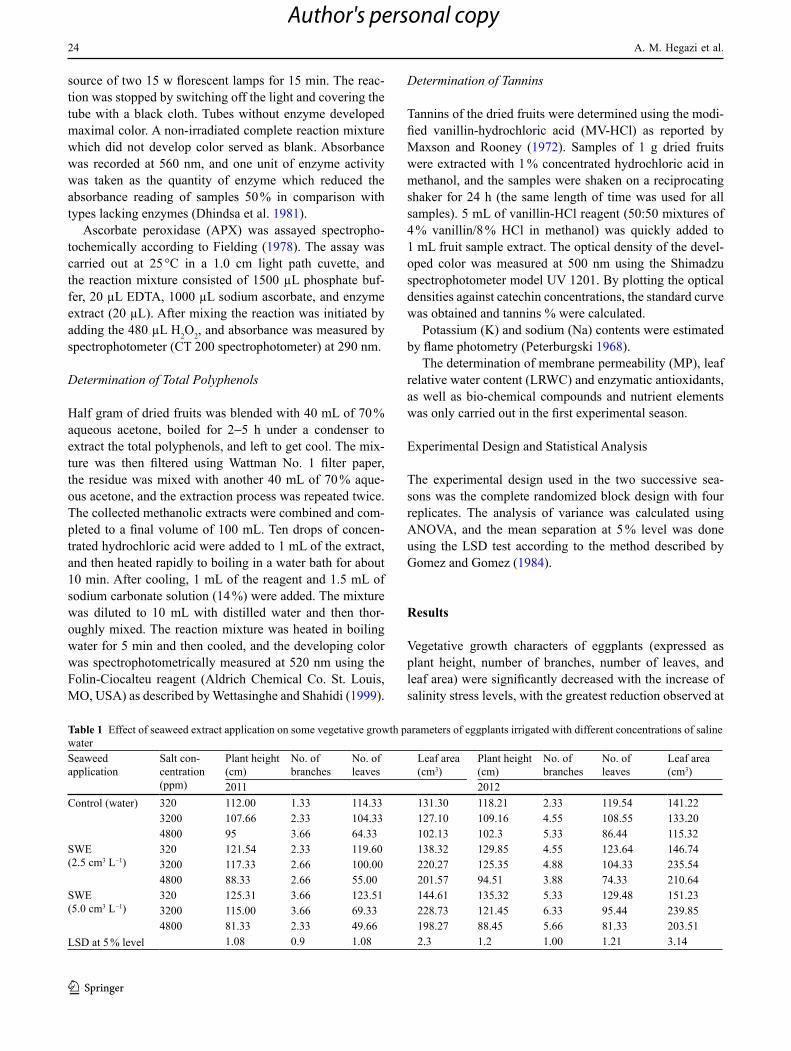
Table 1 Effect of seaweed extract application on some vegetative growth parameters of eggplants irrigated with different concentrations of saline waterSeaweed application
Salt con-centration (ppm)
Plant height (cm)
No. of branches
No. of leaves
Leaf area (cm3)
Plant height (cm)
No. of branches
No. of leaves
Leaf area (cm3)
2011 2012Control (water) 320 112.00 1.33 114.33 131.30 118.21 2.33 119.54 141.22
3200 107.66 2.33 104.33 127.10 109.16 4.55 108.55 133.204800 95 3.66 64.33 102.13 102.3 5.33 86.44 115.32
SWE (2.5 cm3 L−1)
320 121.54 2.33 119.60 138.32 129.85 4.55 123.64 146.743200 117.33 2.66 100.00 220.27 125.35 4.88 104.33 235.544800 88.33 2.66 55.00 201.57 94.51 3.88 74.33 210.64
SWE (5.0 cm3 L−1)
320 125.31 3.66 123.51 144.61 135.32 5.33 129.48 151.233200 115.00 3.66 69.33 228.73 121.45 6.33 95.44 239.854800 81.33 2.33 49.66 198.27 88.45 5.66 81.33 203.51
LSD at 5 % level 1.08 0.9 1.08 2.3 1.2 1.00 1.21 3.14
Author's personal copy
25
1 3
Alleviation of Salt Stress Adverse Effect and Enhancing Phenolic
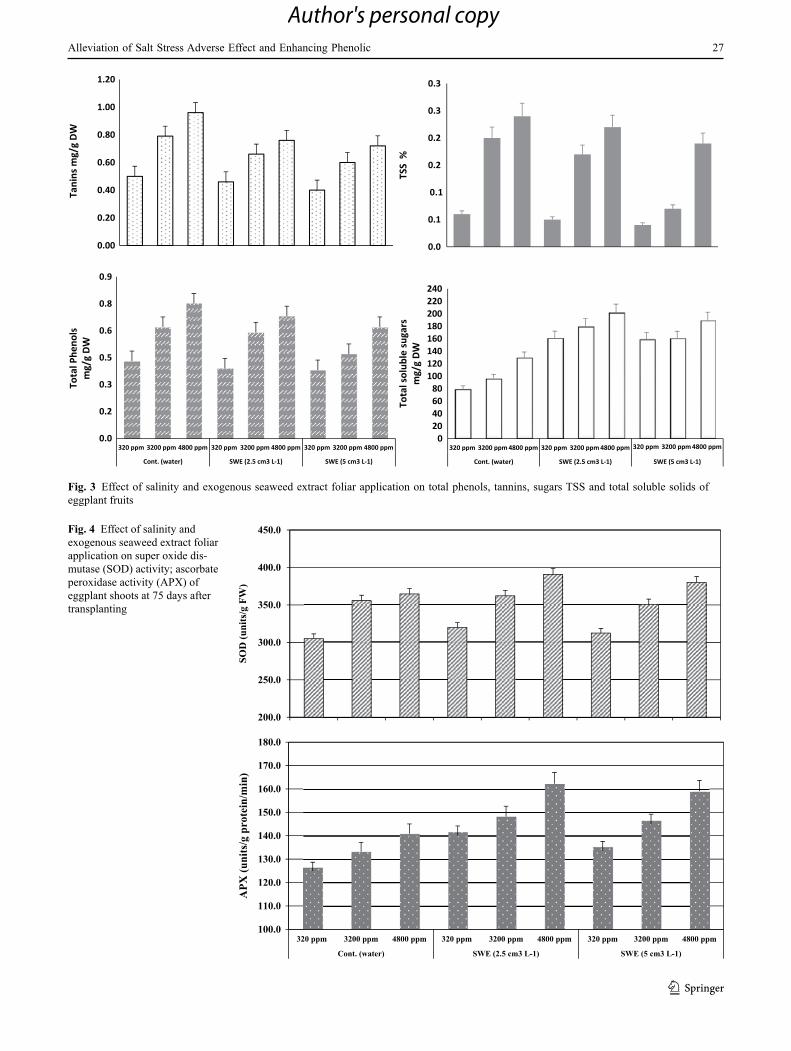
Data illustrated in Fig. 2 shows clearly that membrane permeability was significantly increased as salinity level increased, while seaweed application had a positive effect on enhancing control of membrane permeability. As for leaf relative water content, it was significantly decreased as salinity stress level increased. Seaweed application at both used concentrations counteracted the negative effect on (LRWC %).
Data illustrated in Figs. 3, 4, and 5 shows that all salin-ity stress levels (320, 3200, and 4800 ppm) significantly increased the antioxidant content expressed as total phenols and tannins, as well as the T.S.S. content, total soluble sug-ars, and Na content. Furthermore, salinity stress increased the enzymatic activity of super oxide dismutase (SOD) and ascorbate peroxidase (APX).
It was observed that the applied seaweed extract signifi-cantly increased total phenols, tannins, and T.S.S. contents, and increased the enzymatic activity of SOD and APX, as
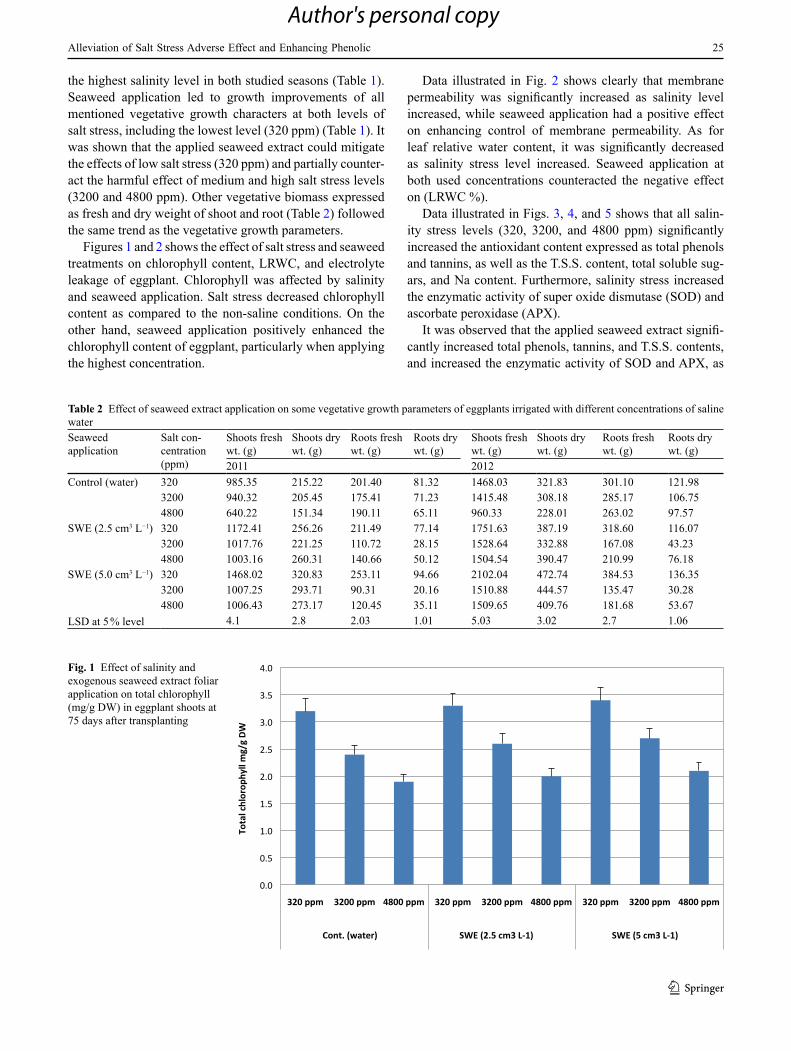
the highest salinity level in both studied seasons (Table 1). Seaweed application led to growth improvements of all mentioned vegetative growth characters at both levels of salt stress, including the lowest level (320 ppm) (Table 1). It was shown that the applied seaweed extract could mitigate the effects of low salt stress (320 ppm) and partially counter-act the harmful effect of medium and high salt stress levels (3200 and 4800 ppm). Other vegetative biomass expressed as fresh and dry weight of shoot and root (Table 2) followed the same trend as the vegetative growth parameters.
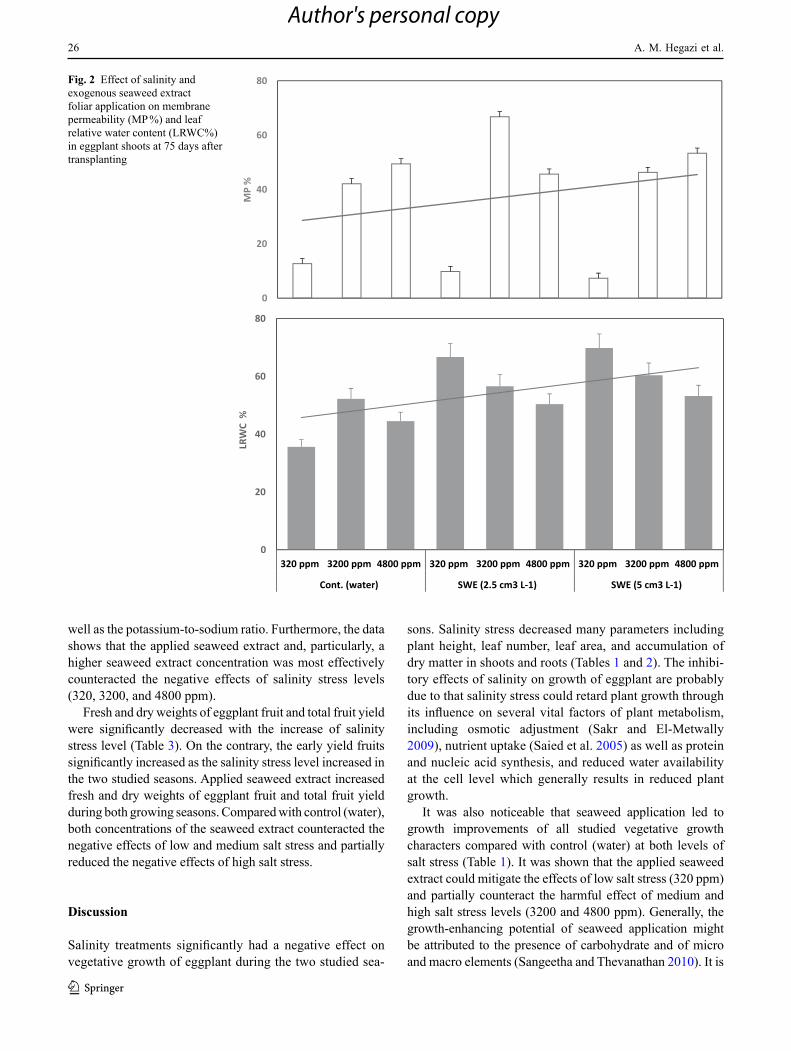
Figures 1 and 2 shows the effect of salt stress and seaweed treatments on chlorophyll content, LRWC, and electrolyte leakage of eggplant. Chlorophyll was affected by salinity and seaweed application. Salt stress decreased chlorophyll content as compared to the non-saline conditions. On the other hand, seaweed application positively enhanced the chlorophyll content of eggplant, particularly when applying the highest concentration.
Table 2 Effect of seaweed extract application on some vegetative growth parameters of eggplants irrigated with different concentrations of saline waterSeaweed application
Salt con-centration (ppm)
Shoots fresh wt. (g)
Shoots dry wt. (g)
Roots fresh wt. (g)
Roots dry wt. (g)
Shoots fresh wt. (g)
Shoots dry wt. (g)
Roots fresh wt. (g)
Roots dry wt. (g)
2011 2012Control (water) 320 985.35 215.22 201.40 81.32 1468.03 321.83 301.10 121.98
3200 940.32 205.45 175.41 71.23 1415.48 308.18 285.17 106.754800 640.22 151.34 190.11 65.11 960.33 228.01 263.02 97.57
SWE (2.5 cm3 L−1) 320 1172.41 256.26 211.49 77.14 1751.63 387.19 318.60 116.073200 1017.76 221.25 110.72 28.15 1528.64 332.88 167.08 43.234800 1003.16 260.31 140.66 50.12 1504.54 390.47 210.99 76.18
SWE (5.0 cm3 L−1) 320 1468.02 320.83 253.11 94.66 2102.04 472.74 384.53 136.353200 1007.25 293.71 90.31 20.16 1510.88 444.57 135.47 30.284800 1006.43 273.17 120.45 35.11 1509.65 409.76 181.68 53.67
LSD at 5 % level 4.1 2.8 2.03 1.01 5.03 3.02 2.7 1.06
Fig. 1 Effect of salinity and exogenous seaweed extract foliar application on total chlorophyll (mg/g DW) in eggplant shoots at 75 days after transplanting
Author's personal copy
26
1 3
A. M. Hegazi et al.
sons. Salinity stress decreased many parameters including plant height, leaf number, leaf area, and accumulation of dry matter in shoots and roots (Tables 1 and 2). The inhibi-tory effects of salinity on growth of eggplant are probably due to that salinity stress could retard plant growth through its influence on several vital factors of plant metabolism, including osmotic adjustment (Sakr and El-Metwally 2009), nutrient uptake (Saied et al. 2005) as well as protein and nucleic acid synthesis, and reduced water availability at the cell level which generally results in reduced plant growth.
It was also noticeable that seaweed application led to growth improvements of all studied vegetative growth characters compared with control (water) at both levels of salt stress (Table 1). It was shown that the applied seaweed extract could mitigate the effects of low salt stress (320 ppm) and partially counteract the harmful effect of medium and high salt stress levels (3200 and 4800 ppm). Generally, the growth-enhancing potential of seaweed application might be attributed to the presence of carbohydrate and of micro and macro elements (Sangeetha and Thevanathan 2010). It is
well as the potassium-to-sodium ratio. Furthermore, the data shows that the applied seaweed extract and, particularly, a higher seaweed extract concentration was most effectively counteracted the negative effects of salinity stress levels (320, 3200, and 4800 ppm).
Fresh and dry weights of eggplant fruit and total fruit yield were significantly decreased with the increase of salinity stress level (Table 3). On the contrary, the early yield fruits significantly increased as the salinity stress level increased in the two studied seasons. Applied seaweed extract increased fresh and dry weights of eggplant fruit and total fruit yield during both growing seasons. Compared with control (water), both concentrations of the seaweed extract counteracted the negative effects of low and medium salt stress and partially reduced the negative effects of high salt stress.
Discussion
Salinity treatments significantly had a negative effect on vegetative growth of eggplant during the two studied sea-
Fig. 2 Effect of salinity and exogenous seaweed extract foliar application on membrane permeability (MP %) and leaf relative water content (LRWC%) in eggplant shoots at 75 days after transplanting
Author's personal copy
27
1 3
Alleviation of Salt Stress Adverse Effect and Enhancing Phenolic
Fig. 3 Effect of salinity and exogenous seaweed extract foliar application on total phenols, tannins, sugars TSS and total soluble solids of eggplant fruits
Fig. 4 Effect of salinity and exogenous seaweed extract foliar application on super oxide dis-mutase (SOD) activity; ascorbate peroxidase activity (APX) of eggplant shoots at 75 days after transplanting
Author's personal copy
28
1 3
A. M. Hegazi et al.
For the present study, a tendency towards elevated elec-trolyte leakage in leaves of salinized eggplant plants was detected (Fig. 2). Similar reports were presented by Parida and Das (2005) and Yildirim et al. (2008) for several crops. Electrolyte leakage causes cell membrane injury when plants are grown under salinity stress. Maintaining integrity of the cellular membranes under salt stress is considered an integral part of the salinity tolerance mechanism (Stevens et al. 2006). Our results showed that seaweed application has positive effects on membrane permeability and can protect the cell membrane from the adverse effects of NaCl salinity in eggplants.
Increased salinity lowered LRWC of eggplants (Fig. 2). Parida and Das (2005) reported that the relative water con-tent, water potential, and osmotic potential of plants become more negative with an increase in salinity. Our present study showed that seaweed application treatments induced an increase in LRWC of the salt-stressed plants as compared to the control plants (Fig. 1). Increases in LRWC of plants treated with seaweed foliar application were also reported on salvia officinalis (El Kaoua et al. 2013).
also reported that seaweed extract contains growth-promot-ing hormones such as auxins, gibberellins, cytokinins, and betaines (Sridhar and Rengasamy 2010).
As for chlorophyll content, salt stress significantly decreased chlorophyll content as compared with control. The adverse effect of salt stress on chlorophyll content of eggplant has been shown by Kaya et al. (2002). Further-more, salt stress inhibits the chlorophyll content in leaves of many crops (Parida and Das 2005). However, seaweed application significantly increased chlorophyll content under both levels of salt stress. The increase in chlorophyll content with seaweed confirmed the reports of Bozorgi (2012) and Ramarajan et al. (2013) who mentioned that seaweed application of bean plant applied as foliar spray or incorporated into medium culture increases leaf pigment (particularly chl A) for bean plants. The increase in chloro-phyll content of plants treated with seaweed might depend on betaines present in the extract and could be a result of reduction in chlorophyll degradation (Wapham et al. 1994). Seaweed extracts are rich in glycine betaine content which delays the loss of photosynthetic activity by inhibiting chlo-rophyll degradation (Genard et al. 1991).
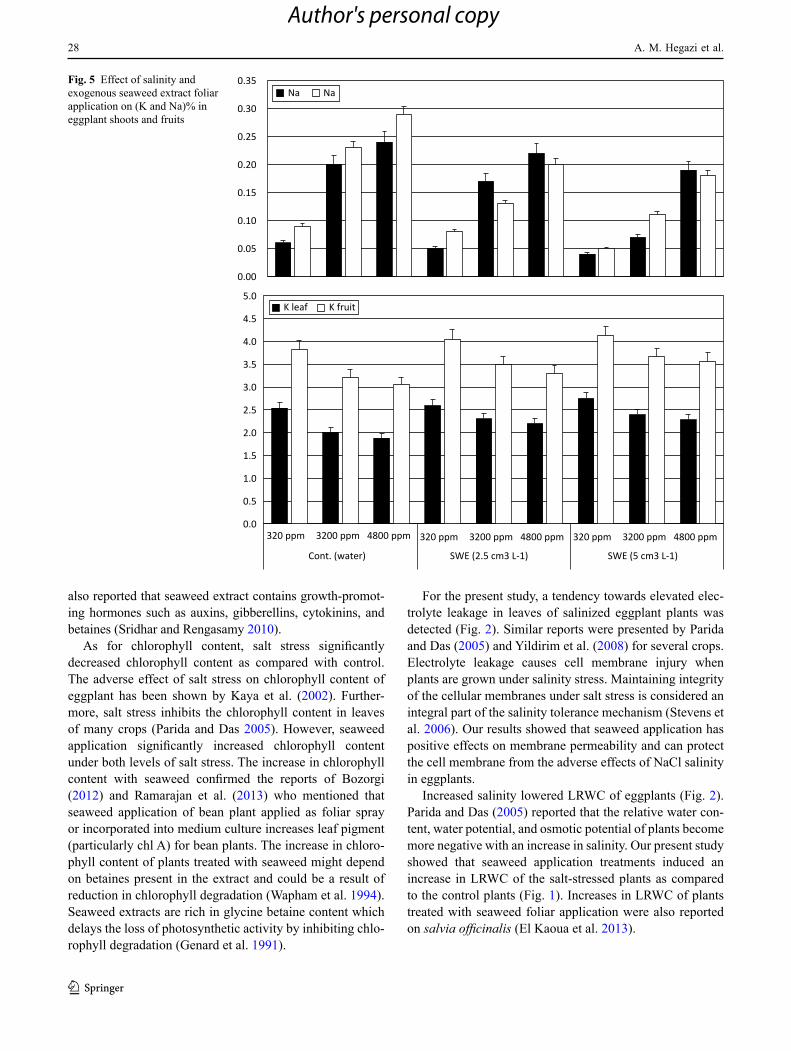
Fig. 5 Effect of salinity and exogenous seaweed extract foliar application on (K and Na)% in eggplant shoots and fruits
Author's personal copy
29
1 3
Alleviation of Salt Stress Adverse Effect and Enhancing Phenolic
ment of antioxidant enzymes (SOD and ASP) for protection against adverse environmental conditions (Schmidt 2005).
As for fruit yield, saline irrigation water significantly decreased fruit fresh and dry weight as well as total fruit yield. However, application of seaweed extract counteracted the negative effects of low and medium salt stress and par-tially reduced the negative effects of high salt stress. Reduc-tions in fruit yield could be due to decreases in the viability of pollen or the receptivity of the stigmatic surface (Sakr et al. 2004), and substantially increased abscission of flowers or young fruits due to ethylene induction by salinity (Sakr et al. 2012). Moreover, a reduction in the leaf area caused by salinity stress led to a reduction in the supply of assimilates due to a decrease in the net photosynthetic rate and biomass accumulation, and consequently decreased total fruit yield.
The enhanced vegetative growth by seaweed extract was reflected by fruit growth and fruit yield. Such results could be due to the fact that more assimilates went to the fruits and due to the high sink strength of the fruits which attracted more assimilates and water to the fruits (Yazied et al. 2012).
The results presented here clearly indicate that there is a possibility to ameliorate the adverse effects of salin-ity by the exogenous application either of anti-oxidants or natural anti-stress compounds known to enhance the natural defenses of plants against salt stress. It was interesting to note that the seaweed extract is relatively inexpensive and commercially available.
The antioxidant content (expressed as total phenols and tannins, and TSS), as well as the content of total soluble sugars and Na were significantly increased as salinity level increased while the K content was decreased (Figs. 3, 4, and 5). The data show that salinity stress levels increased sodium and decreased potassium contents in the shoot of eggplants which is a typical response of plants in saline environments arising from the inability of plants to distinguish between sodium and potassium ions (Storey et al. 1983). Likewise, salt stress significantly increased the enzymatic activity of super oxide dismutase (SOD) and ascorbate peroxidase (APX) in eggplants. The increase in enzymatic activity may be due to that plants grown under salt stress have devel-oped a complex antioxidant system to repair the damage initiated by salt stress. The primary components of this sys-tem include enzymes such as superoxide dismutase (SOD) and ascorbate peroxidase (APX). Many components of this antioxidant defense system can be found in different sub-cellular compartments (Vaidyanathan et al. 2003). Antioxi-dant enzymes and metabolites are reported to increase under various environmental stresses (Yu and Rengel 1999), and this increase was also evident in the current work.
The enhancing effect of seaweed application may be due to that it led to stimulation of the uptake of K and also can alleviate the inhibitory effect of Na toxicity and restore growth. In addition, seaweed application led to an enhance-
Table 3 Effect of seaweed extract application on fruit yield and yield components of eggplants irrigated with different concentrations of saline waterSeaweed application Salt concentration
(ppm)Fruit freshwt. (g)
Fruit drywt. (g)
No. of fruits/plant Early yield(g/plant)
Total yield(g/plant)
2011Control (water) 320 127.2 24.4 17.2 428.2 2189.1
3200 163.3 31.7 10.2 726.8 1661.04800 182.7 37.5 8.2 634.9 1489.0
SWE (2.5 cm3 L−1) 320 157.8 24.0 18.7 838.1 2328.63200 124.9 30.6 26.2 868.5 4131.24800 113.4 21.7 31.3 950.6 3543.8
SWE (5.0 cm3 L−1) 320 179.5 24.3 21.8 589.7 2758.43200 126.4 34.9 23.3 965.6 4173.84800 151.1 28.2 29.1 1123.0 4404.2
LSD at 5 % level 19.4 2.1 1.8 23.2 42.12012
Control (water) 320 156.4 21.8 14.7 366.5 1873.93200 139.8 28.0 8.7 622.2 1421.84800 108.9 31.3 7.0 543.4 1274.6
SWE (2.5 cm3 L−1) 320 97.1 21.4 16.0 717.4 3033.53200 135.1 27.0 22.4 743.5 3536.34800 106.9 19.4 26.8 813.7 1993.3
SWE (5.0 cm3 L−1) 320 129.4 21.6 18.7 504.8 3770.03200 153.7 30.7 19.9 826.6 3572.84800 108.2 25.9 24.9 961.3 2361.2
LSD at 5 % level 18.6 2.75 2.54 19.80 16.09
Author's personal copy

30
1 3
A. M. Hegazi et al.
Kaya C, Bekir Erol AK, Higgs D (2003) Response of salt-stressed strawberry plants to supplementary calcium nitrate and/or potas-sium nitrate. J Plant Nutr 26:543–560
Kayani SA, Naqvi HH, Ting IP (1990) Salinity effects on germi-nation and mobilization of reserves in Jojoba seed. Crop Sci 30(3):704−708
Khan W, Rayirath U, Subramanian S, Jithesh M, Rayorath P, Hodges DM, Critchley AT, Craigie JS, Norrie J, Prithiviraj B (2009) Sea-weed extracts as biostimulants of plant growth and development. J Plant Growth Reg 28:386–399
Lichtenthaler HK, Wellburn AR (1983) Determinations of total carot-enoids and chlorophylls a and b of leaf extracts in different sol-vents. Biochem SocTransact 11:591–592
Maxson ED, Rooney LW (1972) Evaluation of methods for tannin analysis in sorghum grain. Cereal Chem 49:719–729
Noreen Z, Ashraf M (2009) Assessment of variation in antioxida-tive defense system in salt treated pea (Pisum sativum) cultivars and its putative use as salinity tolerance markers. J Plant Physiol 166:1764–1774
Norrie J, Keathley J (2006) Benefits of ascophyllum nodosum marine-plant extract applications to ‘thompson seedless’ grape production. Acta Horticul 185:191–198
Parida AK, Das AB (2005) Salt tolerance and salinity effects on plants: a review. Ecotox Environ Safety 60(3):324–349
Peterburgski AV (1968) Handbook of agronomic chemistry. Klop Pub-lishing House, Moscow, Russia
Ramarajan S, Beschi ARS, Saravana GA (2013) Effect of seaweed ex-tracts mediated changes in leaf area and pigment concentration in soybean under salt stress condition. Research and reviews. J Life Sci 3(1):17–21
Rayirath P, Benkel B, Hodges D, Allan-Wojtas P, MacKinnon S, Critchley AT, Prithiviraj B (2009) Lipophilic components of the brown seaweed, Ascophyllum nodosum, enhance freezing toler-ance in Arabidopsis thaliana. Planta 230:135–147
Saleh SA, Glala AA, Ezzo MI, Ghoname AA (2010) An attempt for reducing mineral fertilization in lettuce production by using bio-organic farming system. Acta Hort 852:311–318
Saied AS, Keutgen AJ, Noga G (2005) The influence of NaCl salinity on growth, yield and fruit quality of strawberry Cvs. ‘Elsanta’ and ‘Korona’. Sci Hortic 103:289–303
Sakhabutdinova AR, Fatkhutdinova DR, Bezrukova MV, Shakirova FM (2003) Salicylic acid prevents the damaging action of stress factors on wheat plants. Bulg J Plant Physiol 1:314–319
Sakr MT, El-Metwally MA (2009) Alleviation of the harmful effects of soil salt stress on growth, wield and endogenous antioxidant content of wheat plant by application of antioxidants. Pakistan J Biol Sci 12(8):624–630
Sakr MT, El-Hadidy M, Abo El-Kheer AM, Farouk S (2004) Physi-ological studies of some osmo-regulator on canola. International Conf. on Microbiol. And Biotechnol. In Favour of Man and En-vironment in Africa and Arab Region. 27th–29th of April 2004, Cairo, Egypt. pp 295–321
Sakr TMM, El-Sarkassy NM, Fuller P (2012) Exogenously applied antioxidants and biostimulants alleviate salt stress in sweet pepper. J Agric Res
Sangeetha V, Thevanathan R (2010) Biofertilizer potential of tradi-tional and panchagavya amended with seaweed extract. J Amer Sci 6:61–67
Schmidt RE (2005) Biostimulants function in turfgrass nutrition. Ph.D. Emeritus Virginia Tech
Shi Q, Bao Z, Zhu Z, Ying Q, Qian Q (2006) Effects of different treat-ments of salicylic acid on heat tolerance, chlorophyll fluorescence, and antioxidant enzyme activity in seedlings of Cucumis sativa L. Plant Growth Regul 48:127–135
References
AOAC (1975) Official methods of analysis, 13th edn, Association of Official Agricultural Chemists, Washington
Aziz N, Mahgoub M, Siam Z (2011) Growth, flowering and chemi-cal constituents performance of amaranthus tricolor plants as in-fluenced by seaweed (Ascophyllum nodosum) extract application under salt stress conditions. J Appl Sci Res 7:1472–1484
Bozorgi H (2012) Effects of foliar spraying with marine plant Asco-phyllum nodosum extract and nano iron chelate fertilizer on fruit yield and several attributes of eggplant (solanum melongena.). ARPNJ Agricul Biol Sci 7:357–362
Bybordi A, Tabatabaei SJ, Ahmadev A (2010) Effect of salinity on the growth and peroxidase and IAA oxidase activities in canola. J Food Agric Environ 8(1):109–112
Cardozo KHM, Guaratini T, Barros MP, FalcaoVR, Tonon AP, Lopes NP, CamposS, Torres MA, Souza AO, Colepicolo P, Pinto E (2007) Metabolites from algae with economic impact. Com Bio-chem Physiol 146:60–78
Demir K, Gunes A, Inal A, Alpaslan M (1999) Effects of humic acids on the yield and mineral nutrition of cucumber (Cucumis sativus L.) grown with different salinity levels. Acta Hort 492:95–103
Dhindsa RA, Plumb-Dhindsa P, Thorpe TA (1981) Leaf senescence correlated with increased permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J Exp Bot 126:93–101
El-Bassiony AM, Ghoname AA, El-Awadi ME, Fawzy ZF, Gruda N (2012) Ameliorative effects of brassinosteroids on growth and productivity of snap beans grown under high temperature. Ges Pfl 64:175–182
El Kaoua M, Chernane H, Benaliat A, Neamallah L (2013) Seaweed liquid extracts effect on Salvia officinalis growth, biochemi-cal compounds and water deficit tolerance. African J Biotech 12(28):4481−4489
El-Tohamy WA, El-Abagy HM, Badr MA, Ghoname AA, Abou-Hus-sein SD (2012) Improvement of productivity and quality of Cape gooseberry (Physalis peruviana L.) by foliar application of some chemical substances. J Appl Sci Res 8(4):2366−2370
Eyras M, Defosse GE, Dellatorre F (2008) Seaweed compost as an amendment for horticultural soils in Patagonia, Argentina. Com-pos Sci Utilization 16:119−124
Fan D, Hodges DM, Zhang J, Kirby CW, Ji X, Locke SJ, Critchley AT, Prithiviraj B (2011) Commercial extract of the brown seaweed Ascophyllum nodosum enhances phenolic antioxidant content of spinach (Spinacia oleracea l.) which protects caenorhab-ditis elegans against oxidative and thermal stress. Food Chem 124:195–202.
Fielding J (1978) A biochemical and cytochemical study of peroxidase activity in root of Pisum sativum. J Exp Bot 29:969−981
Gomez KA, Gomez AA (1984) Statistical procedures for agricultural research, 2nd edn, Wiley and Sons, New York
Genard H, Le-Saos J, Billard JP, Tremolieres A, Boucaud J (1991) Ef-fect of salinity on lipid composition, glycine betaine content and photosynthetic activity in chloroplasts of Suaeda maritime. Plant Physiol Biochem 29:421–427
Hurtado AQ, Yunque DA, Tibubos K, Critchley AT (2009) Use of Aca-dian marine plant extract powder from Ascophyllum nodosum in tissue culture of Kappaphycus varieties. J Appl Phycol 21:633–639
Katerji N, Van Hoorn JW, Hamdy A, Mastrorilli M (2000). Salt toler-ance classification of crops according to soil salinity and to water stress day index. Agric Water Manag 43:99−109
Kaya C, Higgs D, Saltali K, Gezeral O (2002) Response of strawberry grown at high salinity and alkalinity to supplementary potassium. J Plant Nutr 25(7):1415−1427
Author's personal copy

31
1 3
Alleviation of Salt Stress Adverse Effect and Enhancing Phenolic
Yazied A, Gizawy AM, Ragab MI, Hamed ES (2012) Effect of sea-weed extract and compost treatments on growth, yield and quality of snap bean. J Am Sci 8:1–20
Yildirim E, Turan M, Guvenc I (2008) Effect of foliar salicylic acid applications on growth, chlorophyll and mineral content of cu-cumber (Cucumis sativus L.) grown under salt stress. J Plant Nutr 31:593–612
Yilmaz Z (1997) Adaptation and plant growth mechanisms for salt stress. M.Sc. Seminar, Agriculture Faculty, Ankara University
Yu Q, Rengel Z (1999) Drought and salinity differentially influence activities of superoxide dismutases in narrow-leafed lupins. Plant Sci 142:1–11
Amira Mahmoud Hegazi, born on the 6th of January 1972. Associ-ated Professor of Plant Physiology, Agricultural Botany Dept, Fac. of Agriculture, Ain Shams Univ., Cairo, Egypt. Fields of interest: Plant Physiology, Plant Abiotic Stress, Plant Nutrition, Eco-Physiology and Plant Phytoremediation.
Sridhar S, Rengasamy R (2010) Effect of seaweed liquid fertilizer on the growth, biochemical constituents and yield of Tagetes erecta, under field trial. J Phytol 2(6):61–68
Stevens J, Senaratna T, Sivasithamparam K (2006) Salicylic acid in-duces salinity tolerance in tomato (Lycopersicon esculentum cv. Roma): associated changes in gas exchange, water relations and membrane stabilization. Plant Growth Regul 49:77–83
Storey R, Pitman MG, Stelzer R, Carter C (1983) X-ray microanalysis of cells and cell compartments of Atriplex spongiosa. I. Leaves. J Exp Bot 34:778–794
Vaidyanathan H, Sivakumar P, Chakrabarsty R, Thomas G (2003) Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.) differential response in salt-tolerant and sensi-tive varieties. Plant Sci 165:1411–1418
Wapham CA, Jenkıns T, Blunden G, Hankins SD (1994) The role of seaweed extracts, Ascophyllum nodosum in the reduction in fecun-dity of Meloidogyne javanica. Fund Appl Nematol 17(2):181–183
Wettasinghe M, Shahidi F (1999) Evening primrose meal: a source of natural antioxidants and scavenger of hydrogen peroxide and oxy-gen-derived free radicals. J Agricul Food Chem 47(5):1801–1812
Author's personal copy
Related Documents



![Ascorbic Acid and It S Effects on Alleviation of Salt Stress in ... - … · 2016-07-07 · limit the growth and productivity of plants [2]. The inhibitory effect of salinity stress](https://static.cupdf.com/doc/110x72/5e281a91321842186a55c140/ascorbic-acid-and-it-s-effects-on-alleviation-of-salt-stress-in-2016-07-07.jpg)







