Mid-Holocene vegetation history of the central Mediterranean Laura Sadori, 1 S. Jahns 2 and O. Peyron 3 Abstract This paper provides a synthesis of available palynological data from central Mediterranean lacustrine records, studying the causes of the important vegetation changes which occurred in the mid Holocene. They are illustrated by seven well-dated records, lakes Accesa, Mezzano, Vico, and Pergusa in Italy, Maliq, Voulkaria and Malo J. in the Balkans. A good tool for disentangling climate change and human impact is the combined interpretation of pollen percentage and concentration/influx data. Pollen concentration/influx drops are an indication of increased erosion in the catchment, mainly ascribable to forest reduction. There is no doubt that major synchronous changes could not have been caused by humans alone. Many southern European records show substantial differences between early- and late-Holocene vegetation, suggesting a general evolution from wetter to drier climatic conditions, in agreement with arid phases recognised by other, independent palaeoclimatic methods. Other pollen records show a different trend. The role of changing seasonality seems important for this region of the Mediterranean. Precipitation seasonality increased during the early to mid Holocene with winter precipitation attaining a maximum, and summer precipitation a minimum. At least three rapid climate events with changes in plant biomass are in evidence: an abrupt and short change around 8200 yr BP, another one centred around 6000 yr BP, and one soon after 3000 yr BP. From the beginning of the Bronze Age (c. 4400 yr BP in this region) human impact overlapped with a climate change, probably bipartite, towards dryness. Our results show that this aridification trend began around 8000 yr BP, and culminated around 4000 yr BP. Keywords central Mediterranean, climate change, Holocene, human impact, mid-Holocene changes, pollen The Holocene 21(1) 117–129 © The Author(s) 2011 Reprints and permission: sagepub.co.uk/journalsPermissions.nav DOI: 10.1177/0959683610377530 http://hol.sagepub.com Introduction The Mediterranean region possesses a large number of Holocene lacustrine records. These terrestrial archives are widespread across a range of environments in a region that is located at the transition between high- and low-latitude influences. The occur- rence of a pronounced seasonal precipitation rhythm is controlled by the northerly migration of the ITCZ in summer, bringing the Mediterranean under the influence of the subtropical anticyclone belt (‘Azores High’) and widespread dryness, and its southerly retreat in winter, allowing the influence of mid-latitude westerlies and cyclogenesis to be established (Tzedakis, 2007). These west- erly depressions originate mainly from the North Atlantic and the Mediterranean basin and therefore are subject to influence by the NAO. The region is also affected by the Siberian anticyclone presence in winter, and by ENSO and the south Asian monsoon (Lionello et al., 2006; Raicich et al., 2003). Thus, complicated present-day climate processes can explain some of the differences observed in palaeovegetational records. The massive boost given to pollen research in the last decades has allowed reconstructions of both vegetational dynamics and climatic trends during the Holocene, which also show that the Mediterranean is not a homo- geneous region. Close examination of pollen records reveals that specific local and regional patterns characterize the postglacial vegetation succession, reflecting climatic differences and intrin- sic geological and geomorphological features. Additionally, anthropogenic effects are often hard to discern from climate causes, as both can produce similar alterations of natural vegeta- tion successions (Sadori et al., 2010). A clear and simple pattern over the Mediterranean region is therefore often difficult to find. Two reviews on Mediterranean vegetation history, published in the last few years (de Beaulieu et al., 2005; Tzedakis, 2007), both show the complexity of the area, not only during the Holocene, but also during the last million years. De Beaulieu et al. (2005) indicated that some common trends appeared in the northwestern Mediterranean (Spain, France and Italy), but also strong diver- gences, especially during the Holocene. They may be due to real regional differences in the climate, but also to problems in chron- ological assessments of individual records. In Italy, besides a probable general trend toward aridity during the second half of the Holocene, short simultaneous climatic crises are well docu- mented. Another problem discussed by de Beaulieu et al. (2005) is the role of human populations in shaping the present Mediter- ranean environment, given that synanthropic taxa, the ‘anthropo- genic indicators’ defined in northern Europe, are natural elements of open Mediterranean ecosystems. A key question in Tzedakis’ (2007) review is how different lines of often-conflicting evidence 1 Università ‘La Sapienza’, Italy 2 Brandenburgisches Landesamt für Denkmalpflege und Archäologisches Landesmuseum, Germany 3 Université de Franche-Comté, France Received 16 June 2009; revised manuscript accepted 19 May 2010 Corresponding author: Laura Sadori. Dipartimento di Biologia Ambientale, Università ‘La Sapienza’, Piazzale Aldo Moro 5, 00185 Rome, Italy Email: [email protected] Holocene Special Issue

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mid-Holocene vegetation history of the central Mediterranean
Laura Sadori,1 S. Jahns2 and O. Peyron3
AbstractThis paper provides a synthesis of available palynological data from central Mediterranean lacustrine records, studying the causes of the important vegetation changes which occurred in the mid Holocene. They are illustrated by seven well-dated records, lakes Accesa, Mezzano, Vico, and Pergusa in Italy, Maliq, Voulkaria and Malo J. in the Balkans. A good tool for disentangling climate change and human impact is the combined interpretation of pollen percentage and concentration/influx data. Pollen concentration/influx drops are an indication of increased erosion in the catchment, mainly ascribable to forest reduction. There is no doubt that major synchronous changes could not have been caused by humans alone. Many southern European records show substantial differences between early- and late-Holocene vegetation, suggesting a general evolution from wetter to drier climatic conditions, in agreement with arid phases recognised by other, independent palaeoclimatic methods. Other pollen records show a different trend. The role of changing seasonality seems important for this region of the Mediterranean. Precipitation seasonality increased during the early to mid Holocene with winter precipitation attaining a maximum, and summer precipitation a minimum. At least three rapid climate events with changes in plant biomass are in evidence: an abrupt and short change around 8200 yr BP, another one centred around 6000 yr BP, and one soon after 3000 yr BP. From the beginning of the Bronze Age (c. 4400 yr BP in this region) human impact overlapped with a climate change, probably bipartite, towards dryness. Our results show that this aridification trend began around 8000 yr BP, and culminated around 4000 yr BP.
Keywordscentral Mediterranean, climate change, Holocene, human impact, mid-Holocene changes, pollen
The Holocene21(1) 117 –129© The Author(s) 2011Reprints and permission:sagepub.co.uk/journalsPermissions.navDOI: 10.1177/0959683610377530http://hol.sagepub.com
Introduction
The Mediterranean region possesses a large number of Holocene lacustrine records. These terrestrial archives are widespread across a range of environments in a region that is located at the transition between high- and low-latitude influences. The occur-rence of a pronounced seasonal precipitation rhythm is controlled by the northerly migration of the ITCZ in summer, bringing the Mediterranean under the influence of the subtropical anticyclone belt (‘Azores High’) and widespread dryness, and its southerly retreat in winter, allowing the influence of mid-latitude westerlies and cyclogenesis to be established (Tzedakis, 2007). These west-erly depressions originate mainly from the North Atlantic and the Mediterranean basin and therefore are subject to influence by the NAO. The region is also affected by the Siberian anticyclone presence in winter, and by ENSO and the south Asian monsoon (Lionello et al., 2006; Raicich et al., 2003). Thus, complicated present-day climate processes can explain some of the differences observed in palaeovegetational records. The massive boost given to pollen research in the last decades has allowed reconstructions of both vegetational dynamics and climatic trends during the Holocene, which also show that the Mediterranean is not a homo-geneous region. Close examination of pollen records reveals that specific local and regional patterns characterize the postglacial vegetation succession, reflecting climatic differences and intrin-sic geological and geomorphological features. Additionally, anthropogenic effects are often hard to discern from climate causes, as both can produce similar alterations of natural vegeta-tion successions (Sadori et al., 2010). A clear and simple pattern over the Mediterranean region is therefore often difficult to find.
Two reviews on Mediterranean vegetation history, published in the last few years (de Beaulieu et al., 2005; Tzedakis, 2007), both show the complexity of the area, not only during the Holocene, but also during the last million years. De Beaulieu et al. (2005) indicated that some common trends appeared in the northwestern Mediterranean (Spain, France and Italy), but also strong diver-gences, especially during the Holocene. They may be due to real regional differences in the climate, but also to problems in chron-ological assessments of individual records. In Italy, besides a probable general trend toward aridity during the second half of the Holocene, short simultaneous climatic crises are well docu-mented. Another problem discussed by de Beaulieu et al. (2005) is the role of human populations in shaping the present Mediter-ranean environment, given that synanthropic taxa, the ‘anthropo-genic indicators’ defined in northern Europe, are natural elements of open Mediterranean ecosystems. A key question in Tzedakis’ (2007) review is how different lines of often-conflicting evidence
1Università ‘La Sapienza’, Italy2 Brandenburgisches Landesamt für Denkmalpflege und Archäologisches Landesmuseum, Germany
3Université de Franche-Comté, France
Received 16 June 2009; revised manuscript accepted 19 May 2010
Corresponding author:Laura Sadori. Dipartimento di Biologia Ambientale, Università ‘La Sapienza’, Piazzale Aldo Moro 5, 00185 Rome, ItalyEmail: [email protected]
Holocene Special Issue

118 The Holocene 21(1)
from Mediterranean palaeoenvironmental studies might be recon-ciled. Interesting comparisons are made between the present and previous interglacials, for example the presence of sapropel layers in the central and eastern Mediterranean Seas, whose causes can be better understood using a wider temporal and geographical approach.
The central Mediterranean region The central Mediterranean regions show important latitudinal and altitudinal variety, which are accompanied by important differ-ences in sea-surface temperatures. Italy, with its southeastward-protruding peninsula appears to be a natural division between western and eastern Mediterranean and a key region to understand Holocene climatic changes. In this way, the Italian peninsula, the western Balkan coast and associated islands, supply important data for understanding mid-Holocene vegetation trends.
The onset of reforestation in the central Mediterranean region is dated to c. 12 000 yr BP with minor differences from site to site (Table 1), showing different vegetation dynamics and woodland densities. All the tree taxa present today, with the exception of the introduced exotic ones, were present in the region at least since the end of the last glacial period. They spread from multiple refugia (mainly from southern Mediter-ranean ones) located at different elevations. A complex pattern of vegetation response to climatic change prevailed across the region through the Holocene, favouring the spread of particular vegetation types (Sadori, 2007; e.g. Abies in Tuscany and Liguria, Bellini et al., 2009; Drescher-Schneider et al., 2007; Pistacia at coastal Sicily, Tinner et al., 2009): a similar vegetation behav-iour had occurred in previous interglacial periods (de Beaulieu et al., 2005).
Many southern Mediterranean pollen records show substantial differences between early- and late-Holocene vegetation develop-ment, suggesting a general evolution from wetter to drier climatic conditions (e.g. Ben Tiba and Reille, 1982; Carrión et al., 2001;
Pantaleon-Cano et al., 2003; Pérez-Obiol and Sadori, 2007; Sadori and Narcisi, 2001). These vegetation changes have often been interpreted as reflecting aridification phases leading to the estab-lishment of the present-day Mediterranean climate (Follieri et al., 2000; Jalut et al., 2000, 2009). According to an alternative interpre-tation, the expansion of sclerophyllous Mediterranean woodland since the mid Holocene was linked to increasing human impact from the early Neolithic onwards (Pons and Quézel, 1998; Quézel, 1999; Reille and Pons, 1992) and associated in some cases with forest clearance by fires which may have favoured Mediterranean taxa. This hypothesis, however, is not consistent with recent studies of fire history (e.g. Colombaroli et al., 2007, 2009; Sadori and Giardini, 2007, 2008; Sadori et al., 2008; Vannière et al., 2008, 2011, this issue). Nonetheless, changes in biomass burning would have contributed to this landscape transformation.
In recent years it became clear that the chronological frame-work of pollen records needs to be improved if we are to find out whether natural events were really synchronous and to establish their precise timing and duration. Chronology is therefore still a limiting factor when comparing pollen records and correlation of both single events and phase durations. If we admit that every attempt to define the chronology of each ter-restrial sediment record has limits (Zanchetta et al., 2011, this issue), we have to try to define similarities and differences, syn-chronous and asynchronous vegetation patterns using the avail-able data, always keeping in mind that over-precise correlations may turn out to be erroneous. The dates mentioned in the text and illustrations are all expressed in calendar age BP. At most Italian sites the so-called climate optimum was generally char-acterised by maximum percent pollen of deciduous oak. Within this phase, the c. 8200 cal. yr BP cool event recognised in the North Atlantic (Alley et al., 1997) is not easily detected in the central Mediterranean, even if a signal is present in some sites (Pross et al., 2009). In Mediterranean records, a similar behav-iour is found also for other centennial-scale climatic events
Table 1. Synoptic scheme for major Holocene vegetation and climate events of central Mediterranean (from Sadori, 2007, modified)
Date (yr BP) Vegetation change based on pollen data Inferred cause of change
a) Italian peninsula and SicilyAfter 12 000 BP Spread of thermophilous forests (deciduous Quercus dominant) in
central and southernItaly, with Abies in northern Italy
Woodland migration linked to postglacial climatic amelioration
After 9500 BP Beginning of continuous Quercus ilex curve in central Italy (after 9500 BP) and increase of Olea in Sicily (from 8000 BP)
Start of summer-dry climatic conditions; increasing after 6000 BP. Occasional evidence of human activity (from c. 8000 BP)
After 4400–4200 BP Forest loss in Sicily, Apulia and Latium mostly without fires. Decrease of Abies in northern Appennines
Arid climatic phase at most sites
After 4000 BP Forest clearance, cultivation of cereals, legumes and olives (chestnut), presence of secondary anthropogenic indicators and use of fire
Human presence clearly detectable at most sites
After 2800 BP Extensive cultivation, Juglans in Sicily, Castanea widespread near Rome Large-scale human land-cover change
b) Southwestern BalkansAfter 12 000 BP Spread of thermophilous forests (deciduous Quercus dominant) and
conifer woods at some upland sitesWoodland migration linked to postglacial climatic amelioration
10 000–7000 BP Maximum of Pistacia pollen in many sites, associated with open woodland environment in lowlands
Climatic requirements vary according to Pistacia species
9200–7000 BP Reduction of deciduous Quercus and increase of Corylus, Carpinus orientalis/Ostrya, Ulmus and/or Tilia
Period of wetter climate
c. 8000 BP Increase of Phillyrea and Juniperus detected on Dalmatian coast at c. 8000 BP
Short-lived arid event
After 5800 BP Probable human forest clearance, increasing pasture and secondary anthropic indicators
Human presence indicated
After 4500 BP Extensive cultivation of olives since the Bronze Age, Juglans after c. 3300 BP
Clear human impact

Sadori et al. 119
(Bond et al., 1997, 2001) recorded in central and western Medi-terranean (Magny et al., 2009). An aridification trend in vegeta-tion of the central Mediterranean began after 8000–7500 yr BP as represented by either a weak forest reduction or a slight increase of sclerophyllous taxa (in central Italy associated with the spread of beech, in the Balkans with hornbeam) and became more marked around 5500–5000 yr BP. After this time strong changes in woodland extent, density, and composition occurred, and mediterranean-type vegetation spread all over the whole basin. While this may have been caused by a further increase in aridity, it is often masked by human impact which by then becomes unquestionable.
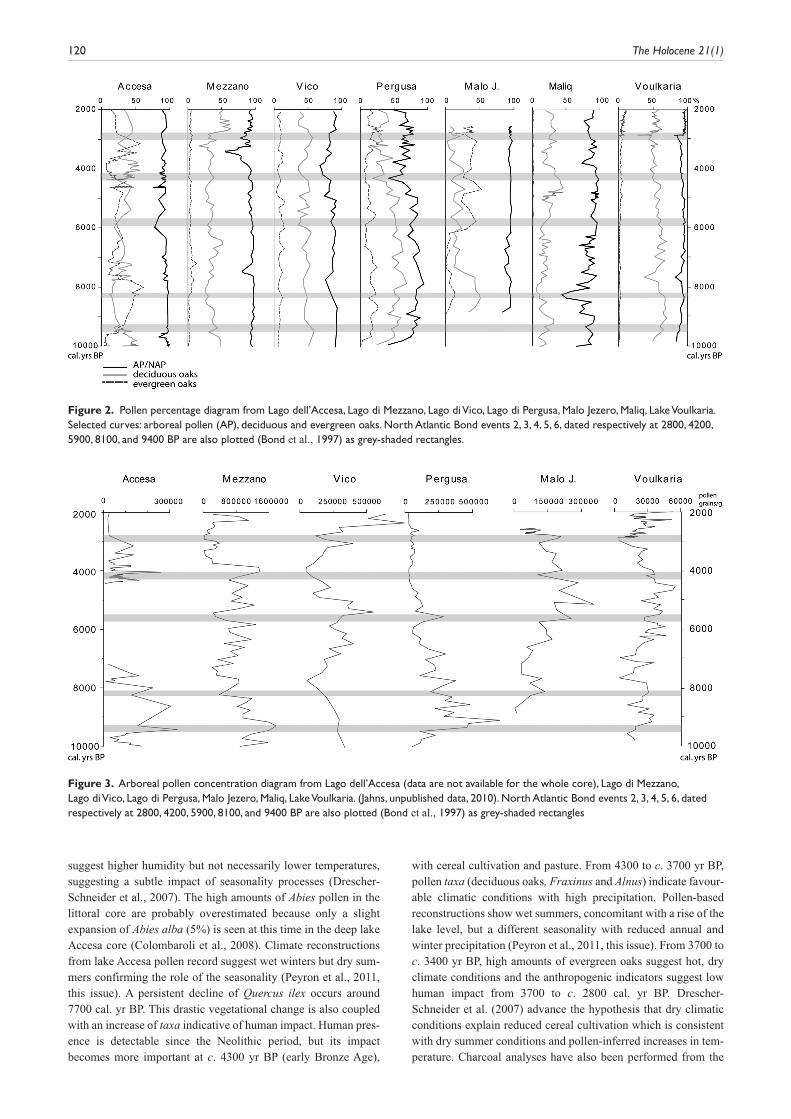
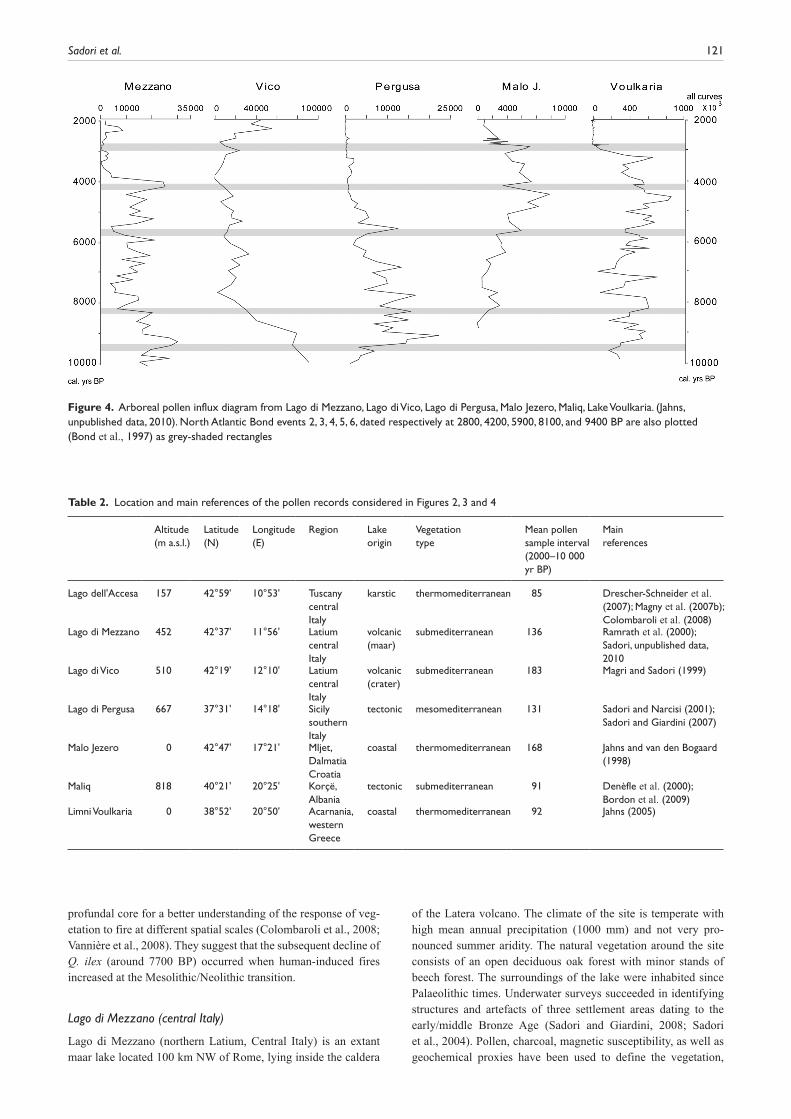
Central Mediterranean key sites Seven sites have been selected for this synthesis (Figure 1, Table 2), three extant lakes from peninsular Italy (Lago dell’Accesa, Lago di Mezzano, Lago di Vico), one from Sicily (Lago di Pergusa), two from the southwestern side of the Balkan peninsula (ancient lake Maliq in Albania and extant lakes Limni Voulkaria in Greece) and Malo Jezero from the Croatian island of Mljet. Most but not all sites experience truly Mediterranean (i.e. summer-dry) cli-matic regimes and they lie from coastal through to mountain envi-ronments. They all have well-established chronologies, based principally on AMS radiocarbon dates and tephra layers (Tables 3 and 4, supplementary information, available online). The main limit for comparison is the fact that dating, in cases in which ter-restrial plant macroremains were not found, was carried out on bulk organic matter, except for a few cases in which pollen was concentrated and dated (Jahns, 2002, 2005). Linear interpolation between two radiocarbon dates (2s interval) was used to assess individual chronologies. Arboreal pollen (AP) percentages, decid-uous oak curves (Figure 2) and arboreal pollen concentration/ influx curves (Figures 3 and 4) have been used to indicate mid-Holocene vegetation changes.
Lago dell’Accesa (central Italy)
Lago dell’Accesa is a small lake (16 ha, 39 m max. depth with a 5 km2 catchment basin) located in Tuscany, central Italy, 10 km from the Tyrrhenian Sea. The climate shows typical Mediterranean fea-tures with annual precipitation of c. 780 mm, highest in autumn with a marked summer drought. Relic areas of natural vegetation around the lake consist of maquis, and evergreen woods. Mean annual tem-perature is c. 13°C, ranging from 4.5°C in the coldest month to 22°C in the warmest. A multiproxy approach based on lake levels, pollen, charcoal and fires, chironomids, magnetic susceptibility and pollen-based transfer functions allowed the main vegetation and climate fluctuations to be defined over the Lateglacial and the Holocene (Drescher-Schneider et al., 2007; Magny et al., 2007a, b , Millet et al., 2007, Peyron et al., 2011, this issue; Vannière et al., 2008). The chronology, spanning the last 16 000 years, is based on 21 14C dates and 8 tephra layers (Magny et al., 2007a, b).
Two pollen records obtained from different localities within the lake describe the vegetation history of Lago dell’Accesa. The pollen diagram from Lake Accesa littoral core AC3/4 (Drescher-Schneider et al., 2007) provides a record of vegetation and cli-matic change spanning 15 000 years, while the deep core AC05 focuses only on the interval from 11 000 until 6500 yr BP (Colom-baroli et al., 2008). Results are roughly in agreement and show that the Holocene vegetation was dominated by Quercus (proba-bly Q. pubescens at the beginning of the Holocene) and character-ised by alternating dominance of deciduous and evergreen oaks (Figure 2). Pollen concentration was not calculated for the whole time window (see Figure 3) and for this reason the influx curve is not provided. Three zones characterised by evergreen oak pollen are accompanied by peat layers associated with low lake levels (Magny et al., 2007b) at c. 8600–7900, 4600–4300 and 3700–2800 yr BP. From 9500 to 7700 yr BP, high percentages of ever-green oaks and low lake levels appear to indicate warm, dry conditions, while high values of Abies (20%) and other taxa
Figure 1. Location of the selected Mediterranean palynological records and of other sites quoted in text (1, Gorgo Basso; 2, Grotta di Carburangeli; 3, Corchia cave; 4, Lago Frassino in northern Italy; 5, Lago Grande di Monticchio; 6, Tenaghi Philippon; 7, Ohrid; 8, Lago di Massacciuccoli; 9, Rascafría; 10, Villaverde; 11, Padul; 12, Olot; 13, Almería; 14, Biot; 15, Palavas; 16, Delphinos; 17, Cres; 18, Minorca
120 The Holocene 21(1)
suggest higher humidity but not necessarily lower temperatures, suggesting a subtle impact of seasonality processes (Drescher-Schneider et al., 2007). The high amounts of Abies pollen in the littoral core are probably overestimated because only a slight expansion of Abies alba (5%) is seen at this time in the deep lake Accesa core (Colombaroli et al., 2008). Climate reconstructions from lake Accesa pollen record suggest wet winters but dry sum-mers confirming the role of the seasonality (Peyron et al., 2011, this issue). A persistent decline of Quercus ilex occurs around 7700 cal. yr BP. This drastic vegetational change is also coupled with an increase of taxa indicative of human impact. Human pres-ence is detectable since the Neolithic period, but its impact becomes more important at c. 4300 yr BP (early Bronze Age),
with cereal cultivation and pasture. From 4300 to c. 3700 yr BP, pollen taxa (deciduous oaks, Fraxinus and Alnus) indicate favour-able climatic conditions with high precipitation. Pollen-based reconstructions show wet summers, concomitant with a rise of the lake level, but a different seasonality with reduced annual and winter precipitation (Peyron et al., 2011, this issue). From 3700 to c. 3400 yr BP, high amounts of evergreen oaks suggest hot, dry climate conditions and the anthropogenic indicators suggest low human impact from 3700 to c. 2800 cal. yr BP. Drescher-Schneider et al. (2007) advance the hypothesis that dry climatic conditions explain reduced cereal cultivation which is consistent with dry summer conditions and pollen-inferred increases in tem-perature. Charcoal analyses have also been performed from the
Figure 2. Pollen percentage diagram from Lago dell’Accesa, Lago di Mezzano, Lago di Vico, Lago di Pergusa, Malo Jezero, Maliq, Lake Voulkaria. Selected curves: arboreal pollen (AP), deciduous and evergreen oaks. North Atlantic Bond events 2, 3, 4, 5, 6, dated respectively at 2800, 4200, 5900, 8100, and 9400 BP are also plotted (Bond et al., 1997) as grey-shaded rectangles.
Figure 3. Arboreal pollen concentration diagram from Lago dell’Accesa (data are not available for the whole core), Lago di Mezzano,Lago di Vico, Lago di Pergusa, Malo Jezero, Maliq, Lake Voulkaria. (Jahns, unpublished data, 2010). North Atlantic Bond events 2, 3, 4, 5, 6, dated respectively at 2800, 4200, 5900, 8100, and 9400 BP are also plotted (Bond et al., 1997) as grey-shaded rectangles
Sadori et al. 121
profundal core for a better understanding of the response of veg-etation to fire at different spatial scales (Colombaroli et al., 2008; Vannière et al., 2008). They suggest that the subsequent decline of Q. ilex (around 7700 BP) occurred when human-induced fires increased at the Mesolithic/Neolithic transition.
Lago di Mezzano (central Italy)
Lago di Mezzano (northern Latium, Central Italy) is an extant maar lake located 100 km NW of Rome, lying inside the caldera
of the Latera volcano. The climate of the site is temperate with high mean annual precipitation (1000 mm) and not very pro-nounced summer aridity. The natural vegetation around the site consists of an open deciduous oak forest with minor stands of beech forest. The surroundings of the lake were inhabited since Palaeolithic times. Underwater surveys succeeded in identifying structures and artefacts of three settlement areas dating to the early/middle Bronze Age (Sadori and Giardini, 2008; Sadori et al., 2004). Pollen, charcoal, magnetic susceptibility, as well as geochemical proxies have been used to define the vegetation,
Figure 4. Arboreal pollen influx diagram from Lago di Mezzano, Lago di Vico, Lago di Pergusa, Malo Jezero, Maliq, Lake Voulkaria. (Jahns, unpublished data, 2010). North Atlantic Bond events 2, 3, 4, 5, 6, dated respectively at 2800, 4200, 5900, 8100, and 9400 BP are also plotted (Bond et al., 1997) as grey-shaded rectangles
Table 2. Location and main references of the pollen records considered in Figures 2, 3 and 4
Altitude(m a.s.l.)
Latitude (N)
Longitude (E)
Region Lake origin
Vegetation type
Mean pollen sample interval (2000–10 000 yr BP)
Main references
Lago dell'Accesa 157 42°59' 10°53' Tuscanycentral Italy
karstic thermomediterranean 85 Drescher-Schneider et al. (2007); Magny et al. (2007b); Colombaroli et al. (2008)
Lago di Mezzano 452 42°37' 11°56' Latium central Italy
volcanic (maar)
submediterranean 136 Ramrath et al. (2000); Sadori, unpublished data, 2010
Lago di Vico 510 42°19' 12°10' Latiumcentral Italy
volcanic (crater)
submediterranean 183 Magri and Sadori (1999)
Lago di Pergusa 667 37°31' 14°18' Sicilysouthern Italy
tectonic mesomediterranean 131 Sadori and Narcisi (2001);Sadori and Giardini (2007)
Malo Jezero 0 42°47' 17°21' Mljet, DalmatiaCroatia
coastal thermomediterranean 168 Jahns and van den Bogaard (1998)
Maliq 818 40°21' 20°25' Korçë,Albania
tectonic submediterranean 91 Denèfle et al. (2000); Bordon et al. (2009)
Limni Voulkaria 0 38°52' 20°50' Acarnania, western Greece
coastal thermomediterranean 92 Jahns (2005)

122 The Holocene 21(1)
climate, and human-induced changes that occurred at Lago di Mezzano in the last 15 000 years (Ramrath et al., 2000; Sadori, unpublished data, 2010). A chronology of the whole sediment record spanning the last 30 000 years was proposed on the basis of varve thickness measurements, AMS radiocarbon dates, tephra layers study, and interpolated sedimentation rates.
Deciduous oaks are the most important trees up until the present day (Figure 2), even if important expansions of hazel and beech are found (Sadori, unpublished data, 2010). The Holocene vegetation at Lago di Mezzano, besides important changes of composition, shows a number of abrupt and dramatic drops of pollen concentration/ influx indicating considerable variations of the forest density, with minima at c. 8200, 7600–7300 (matching the maximum percentage of evergreen oaks), 5400, 3700–3400, and 2900 yr BP (Figures 3 and 4). Evidence of environmental change is found for several peri-ods and in some cases it is possible to disentangle different causes of vegetation change. Human impact on the environment is evident (cultivation, grazing and forest clearance) from 3600 to 3400 yr BP during the Bronze Age, following evidence of an abrupt climatic change occurring after 3900 BP (Sadori, 2006; Sadori et al., 2004). Earlier changes in vegetation are generally interpreted as climate-induced although it is not possible to exclude human land use. Clear effects of human impact are also found in Etruscan/Roman times and during the Middle Ages. Important natural fires (Sadori, unpublished data, 2010) occurred at the beginning of the Holocene up until 10 000 years BP, in a period in which the climate was rather unstable and human presence is hardly detected. In the long period between 10 000 to 3800 yr BP (Sadori and Giardini, 2008; Sadori et al., 2004), only a number of sporadic and isolated fires occurred. The interaction of climate and human impact on the environment is particularly clear during the Bronze age period, from about 4100 to about 2900 yr BP. Fires played an important role in modifying the landscape also during the Roman and the Middle Ages, as well as in the last centuries, when a strong human pressure on the environ-ment is evident.
Lago di Vico (central Italy)
The Vico volcano has a central collapse caldera, at present occu-pied by the homonymous lake. Lago di Vico (maximum depth of c. 50 m, maximum diameter of c. 5 km, surface area of c. 12 km2) is located in Latium, central Italy, 50 km NW of Rome. Rainfall for the highest peaks in the catchment area is estimated at about 2000 mm. No summer aridity, typical of the Mediterranean cli-mate, is found. Mainly deciduous oaks and beech forests, chestnut coppices, together with sparse holm oaks and hazel woods form the caldera vegetation. Human activity in the caldera is docu-mented from the beginning of the Neolithic to the end of the Eneolithic. During the Bronze Age there is evidence of occupa-tion which increased during Etruscan and Roman times. The sedi-ment record of Holocene vegetation history shows rather stable environmental conditions (Magri and Sadori, 1999). A sharp increase in tree percentages is recorded at c. 12 000 yr BP, which remain at high values (70–90%) up to the present time. The most characteristic feature of the Holocene appears to be the over-whelming abundance of deciduous oaks (Figure 2), never lower than 35%, while none of the other arboreal taxa reaches 20%. The mean sample interval, due to the low sedimentation rate (1.25 m for the whole Holocene) is only 183 years. The landscape does not undergo important floristic changes and the vegetation does not show a typical interglacial succession; there are only few AP
reductions, emphasized by dramatic drops of pollen concentration/ influx indicating considerable variations of the forest density, with minima recorded at c. 7800, 5900, 4800, 4000–3900, and 2800 yr BP, highlighting the main vegetational changes of the Holocene (Figure 3).
Lago di Pergusa (Sicily, Italy)
Lago di Pergusa (37°31′N; 14°18′E, 667 m a.s.l.) is the main inland natural lake of Sicily, located in the centre of the island, a few kilometres from the chief town of Enna; it occupies an endo-reic basin of subelliptical shape. The mean annual precipitation in the area is between 500 and 700 mm, and the mean annual tem-perature is 13.4°C. The potential vegetation is supposed to be formed by mesophilous Mediterranean evergreen woods with prevailing Quercus ilex L., but the surroundings of Lago di Per-gusa are among the most open landscapes of Sicily and are widely cultivated, the only traces of natural woody vegetation being sparse stands of deciduous and evergreen oaks. In central Sicily there is evidence of human activity since the Palaeolithic. Copper, Bronze and Iron Ages occupation are documented in several sites close to the lake (Sadori and Giardini, 2007, 2008). Since the first millennium bc the area was inhabited first by Sicans and Siculis, then by Greeks, Syracusans, Carthagenians, and Romans.
The chronological framework of the postglacial part of the sedi-ment record has been provided by seven AMS radiocarbon dates and one tephra layer. The early-Holocene tree increase appears to have been rather slow and a couple of millennia was necessary for the transition from an open vegetation formation with sparse trees to real forest conditions, which was completed at c. 9900 yr BP.A forest dominated by deciduous and evergreen oaks was established in the estimated time interval 10 000–5000 yr BP (Figure 2), with the maximum arboreal biomass expansion recorded in the period between soon after 9400 and around 8200 yr BP. This expansion is however bipartite, showing a decrease between c. 8900 and 8700 yr BP. An important change is found soon after 8000 yr BP, indicating the beginning of the continuous curve of Olea and of a climatic trend towards more arid conditions. The arboreal pollen concentration/ influx curves (Figures 3 and 4) show minima at c. 8700, 8100, 7600, 6100–5900 yr BP. Excepting a temporary recovery of the for-est at c. 5500 yr BP, the environment appears to be mostly open in the last millennia, with deciduous and evergreen oaks and olive as the main taxa. The AP percentage curve however shows further important forest clearance at 4400–4300 and at 2000 yr BP. Impor-tant fires were recorded corresponding to the main forest expansion of deciduous oaks, in the period 9400–8200 yr BP and from 4500 yr BP onwards (Sadori and Giardini, 2007).
Malo Jezero (Mljet island, Croatia)
Mljet is the southernmost of the big islands along the Dalmatian coast. The island is largely wooded, especially in the western part, the site of a National Park where Malo Jezero is located. At higher elevation dense Quercus ilex forests grow, in other parts Pinus halepensis occurs frequently. Where the forests are degraded to maquis, evergreen bushes such as Pistacia lentiscus, Myrtus com-munis and Arbutus unedo grow. Today, Mljet is characterized by a climate with annual mean precipitation of 973 mm and annual mean temperature of 16.2°C. Even if moister conditions can be supposed for the early Holocene, the water supply on the islands must have been less stable than on the mainland, where the clouds

Sadori et al. 123
accumulate around the mountains. Therefore the vegetation on the islands can be assumed to have reacted more sensitively to climatic change than in places with more precipitation. Archaeo-logical findings date back to the Iron Age. At latest since Roman times, Mljet was permanently populated to a larger extent.
Pollen diagrams from Malo Jezero and Veliko Jezero (two communicating salty lagoons connected with the sea) show that in the early Holocene the island of Mljet was covered by dense decid-uous oak forest until c. 7700 BP (Beug, 1961, Jahns and van den Bogaard, 1998). Only in the oldest part Pinus and Pistacia show higher values, which might indicate drier conditions before 8900 BP. Diatom studies on the lake sediments, which were carried out by Wunsam et al. (1999) point to a relatively moist climate for the period which started around 8400 BP. Around 8200 BP an expan-sion of Carpinus orientalis/Ostrya type can be observed. The same phenomenon is visible in diagrams from northern Greece (Lawson et al., 2004; Willis, 1992). A change in climate as a trigger for this vegetation development is suggested by most authors, although its meaning is not specified. Beug (1961) proposed plant migration to explain this phenomenon. Around 7600 BP the diagrams from Mljet show a distinct change to an evergreen vegetation, mainly composed of Juniperus and Phillyrea, which replaced the decidu-ous oak woodland. At c. 6100 BP evergreen oaks spread. As the present knowledge of the settlement history of the island does not indicate strong deforestation in the Neolithic period, it is more likely that this change in vegetation is the result of a climatic change to drier conditions. This interpretation corresponds to the results of the investigations of Colombaroli et al. (2009) on Malo Jezero, which linked the increase of evergreen oaks to decreasing frequencies of regional fire. This vegetational change shows a good correspondence with the results from a palynological inves-tigation of a core from Lake Vrana on the island of Cres (Schmidt et al., 2000), where evergreen woody taxa increase after c. 6000 BP. There, a distinct increase of evergreen oaks is dated at 4200 BP, much later than on the island of Mljet. Arboreal pollen concen-tration minima at Malo J. are found at c. 5800, 4100, 3000 yr BP (Figure 3; Jahns, this paper).
From the nearby mainland coast of Dalmatia the studies of Brande (1973) from the Neretva lowlands are available for com-parison. These pollen diagrams show an expansion of Phillyrea, which could be dated to c. 7500 BP at Vid; however, the signal is less prominent than that of Mljet. Subsequently, also an increase of evergreen oaks can be observed.
Maliq (Albania)
In Albania, the pollen record from past lake Maliq documents the vegetation dynamics of the Balkans since 12 000 yr BP (Denèfle et al., 2000). Owing to the proximity of the Adriatic Sea and the surrounding mountains, the climate in the area of Maliq has both Mediterranean and continental influences. The regional climate is characterized by low temperatures in winter (around 0°C) and mild temperatures in summer (from 16 to 20°C). Annual rainfall is around 790 mm (40 mm/month in summer, around 70 mm/month in winter).
The present-day vegetation at Maliq is sub-Mediterranean, and follows an altitudinal distribution with mixed deciduous woods dominating the lower mountain slopes, oak forests covering the upper slopes, and above, beech woods and sub-alpine moors and grasslands (Bordon et al., 2009). This site is one of the rare ones which give vegetation information concerning middle-high altitudes
in the Balkan peninsula, and is well dated with 14 radiocarbon dates (Bordon et al., 2009; Denèfle et al., 2000). At the beginning of the Holocene, a mixed forest characterized by deciduous Quer-cus associated with Corylus, Tilia, and Ulmus developed after 10 200 yr BP (Denèfle et al., 2000). The development of lake-shore trees, followed by the progressive increase of Abies, could indicate high precipitation and low temperatures in summer (Denèfle et al., 2000). This interpretation is confirmed by the study of Bordon et al. (2009) which reconstructed warm and wet condi-tions for the beginning of the Holocene, as well as remarkably stable climatic conditions from 11 300 to 2000 BP, with the excep-tion of a short-lived cooling event, dated at 8300 to 8100 BP. After 5000 yr BP, a modest expansion of the Mediterranean evergreen oaks and Olea indicates different environmental conditions, char-acterized by lower available summer moisture and warmer win-ters, in agreement with the ‘drier trend’ reconstructed by Bordon et al. (2009) from 5000 BP to today. At the same time, the increased importance of Fagus, the Abies decrease, and the appear-ance of cultivated plants has been attributed to the development of human activity in the Maliq region (Denèfle et al., 2000).
Limni Voulkaria (Greece)
Lake Voulkaria is situated along the coast of western Greece, south of the Gulf of Ambrakia, in very close vicinity of the Ionian Sea, just in front of Lefkada island. Although the climate is of Mediterranean character with winter rain and summer drought (at the nearby Korfu island, located c. 80 km NW of the lake, the mean annual precipitation is 1094 mm and the mean annual tem-perature is 17.5°C) the clouds accumulating around the inland mountains provide a better water supply than on the Island of Mljet. Even if the area was already inhabited in Palaeolithic times, known prehistoric sites are rare. The most famous sites date back to the Classic Hellenistic period, the ancient city of Palairos lies very close to Lake Voulkaria. In Roman times the population declined after the Battle of Actium in 31 bc. The pres-ent-day vegetation mainly consists of partly high-growing maquis and partly shrub phrygana (for example Quercus coccifera, Phillyrea latifolia, Pistacia lentiscus). Trees are scarce, (for example Quercus macrolepis and other deciduous Quercus,Carpinus orientalis, Fraxinus ornus, Celtis australis).
Lake Voulkaria shows a pollen record for the last 9800 years. In contrast to the pollen diagram from Mljet, deciduous oaks dominated throughout the Holocene, similarly to the development shown in most Italian lacustrine records. Until c. 9000 BP, Pista-cia occurred frequently (see also Lawson et al., 2004 for Ioannina basin). Afterwards the area was covered by deciduous oak wood-land. The prevalence of deciduous oaks can be attributed to the better water supply to the western Greece coast, implying that deciduous forest was widespread in this lowland area. The increase of Carpinus orientalis/Ostrya at c. 8200 BP, shown in the pollen diagram from Mljet, can also be found at Lake Voulkaria. In the mid Holocene no distinct change to evergreen vegetation as on Mljet can be observed al Lake Voulkaria, only a less prominent spreading of Erica is visible around 7100 BP.
An unpublished pollen concentration estimate is available for the record, with a trend very similar to that of Mljet island. The curve indicates that some forest biomass crises happened, with harshest conditions recorded at c. 8700, 8300, 7600 and 2900 yr BP and minor negative peaks matching minima of other central Mediterranean curves (Figures 3 and 4).

124 The Holocene 21(1)
Principal vegetation changes in the central Mediterranean
In Figures 2, 3 and 4 the AP/NAP ratio, the main taxa pollen curves and concentrations/influx (when available) are drawn for each site, as the basis for the interpretation and discussion of main landscape changes. The chronology of the pollen records used in Figures 2 and 3 will not be discussed here, as it was established in original papers through radiocarbon and tephra analyses. Avail-able radiocarbon dates and tephras are reported in Tables 3 and 4 (supplementary information, available online). All the records show the existence of a dense forest cover around the lakes included in this study for most of the early–mid Holocene (Table 1; Figure 2), independent of their geographical location and avail-ability of atmospheric moisture. In the early millennia of the pres-ent interglacial (from c. 11 000 to c. 8000 yr BP), deciduous oaks were widespread all over the northern shores of the central Medi-terranean basin, weakening first at true Mediterranean sites like Mljet island and Accesa lake. The first signal of forest reduction is found at Italian sites at c. 9000 BP, deduced by absolute pollen values and also by AP percentages in Sicily (Figures 2, 3 and 4). This does not appear to match the interruption of S1 deposition in the Mediterranean Sea, which seems to have occurred later (Ariztegui et al., 2000). The vegetational change may correspond to the 9400 yr BP event found by Bond et al. (1997) in the North Atlantic and also identified in the pollen-based climate reconstructions from Accesa (Peyron et al., 2011, this issue) and the Aegean sea (Dor-moy et al., 2009). This forest opening happened during the so-called ‘climate optimum’ of the early Holocene. Palynologists (e.g. Rossignol-Strick, 1999) have indicated that this was the period in which deciduous forests achieved their maximum expansion in the Holocene, probably the period with highest win-ter precipitation. This climate favoured deciduous oaks, but we should at least wonder if this is really an ‘optimum’ climate for the region, as drought-tolerant Mediterranean-type taxa, for example, did not profit from it.
The 8200 yr BP event (Alley et al., 1997, Bond et al., 1997), thought to be the strongest short-term climate anomaly within the Holocene, is represented by sharp changes in some of the records considered here (Figures 2, 3 and 4). At the Italian lakes of Pergusa and Mezzano both forest reduction and a change in composition are found, suggesting an arid event; in contrast to this evidence, a lake-level rise is found at Accesa (Magny et al., 2007a, b) synchronous with a short-lived increase in reconstructed summer precipitation (Peyron et al., 2011, this issue). The biome recon-struction inferred from the Lago Grande di Monticchio pollen sequence indicates a shift from a temperate deciduous biome to a warm mixed forest one at the same time (Allen et al., 2002). The new pollen data from Tenaghi Philippon, NE Greece, also provide evidence for a major climate-induced ecosystem turnover in the northern Aegean region associated with the 8200 yr BP event (Peyron et al., 2011, this issue; Pross et al., 2009). At Maliq a rapid event is clearly visible in the pollen diagram and well recon-structed as a cooling by Bordon et al. (2009). A small change is found at lake Ohrid (Wagner et al., 2009), while it seems clearer at Voulkaria and Mljet lakes, where deciduous oaks show a decrease and Carpinus orientalis/Ostrya an increase. At Mljet there was also an important coeval increase of juniper. The increase of these two pollen taxa appears to be contradictory, because the first may indicate wetter conditions, and the second one drier ones. A long discussion has taken place on the hornbeam
spread in the Balkans (e.g. Beug, 1961; Tonkov, 2003; Willis, 1994). Probably the most convincing hypothesis is that it is linked to the Holocene re-migration of this taxon from glacial refugia to east. An argument in favour of this hypothesis could be the later spread of Carpinus orientalis/Ostrya into Italy and its absence in the western Mediterranean.
Subsequent vegetation change was longer-lasting and is recorded in all sites soon after 8000 BP. It coincides with a decrease of AP percentages (and concentration at some lakes) and with the start of the expansion of Mediterranean-type vegetation at most sites. In the European central Mediterranean region the Neolithic period started at c. 8100 yr BP, so a climatic link to changes in human society cannot be excluded. At around 6000 yr BP another change in forest density occurred; Accesa appears to be a particularly sensitive record as not only forest reduction, but also low lake level is found (Magny et al., 2007b). A consistent reduction of deciduous oaks is also visible at Vico and Mezzano lakes and a change at Mljet, where a sharp increase of evergreen oaks is found.
A further increase in mediterranean-type vegetation seems to be achieved in the following centuries, while a rapid, strong change occurred in the period 4400–4100 yr BP in all the inves-tigated sites except Accesa, where a different trend (Giraudi et al., 2011, this issue; Magny et al., 2007b) seems to be visible. At Lago di Vico, where the low sedimentation rate made it diffi-cult to obtain a high-resolution record for the Bronze Age, the arboreal biomass drop beginning around 4300 BP seems to be accompanied by an important human exploitation of the sur-rounding territory, lasting without interruption for more than four centuries. At Lago di Mezzano, the event appears bipartite, with a first minor pollen concentration drop at around 4400 BP, before the occurrence of the Avellino tephra, associated with a slight decrease of AP percentages. A second, more distinct change lasted from c. 3800 to 3400 BP. At the time of the second forest reduction a strong human impact is detected, with clear traces of cultivation, forest exploitation and fire (Sadori et al., 2004). At Lago dell’Accesa a consistent human impact is found from 4300 to c. 3700 BP, but during the middle and late Bronze Age (3700 to c. 2800 BP) human impact seems low. Little is known about the human population in the Accesa area during the Bronze Age; however a large number of graves around the lake provide evi-dence of the start of intensive human colonisation (Drescher-Schneider et al., 2007). At Lago di Pergusa the human role is not easily disentangled from a climatic one; at 4300 BP and at 3800 BP AP percentages drop to c. 40%, indicating a forest decline characterized by slight decreases in tree pollen concentration (AP concentration is already very much reduced in the second half of the Holocene) and an increase of herbaceous pollen, as well as evidence of fires. At Maliq a strong decrease in deciduous oaks is indicated around 3800 BP. At Malo Jezero a decrease in arboreal pollen concentration is found slightly before 4100 BP, accompanied only after 3900 BP by a decrease of AP percent-ages, with a pattern similar to that of Lago di Mezzano. At lake Voulkaria, a first clearance is documented by a minimum of AP pollen concentration at 4200 BP and by a second, more important one, from 3800 to 3400 BP. The environmental change of the early and middle Bronze Age seems therefore to be bipartite in the central Mediterranean (Magny et al., 2009). The date of the first signal deserves a closer examination, and both improved chronologies (e.g. the detection of Avellino tephra, Sulpizio et al., 2008; Zanchetta et al., 2011, this issue) and sampling of individual

Sadori et al. 125
records will reveal if it is synchronous or if it reflects a shift in the location of anticyclones in the Mediterranean basin (Di Rita and Magri, 2009). A first signal therefore occurs between 4400 and 4100 yr BP, and is sudden and short. It seems to affect first arboreal pollen concentration and then percentages, and is often accompa-nied by human presence/impact indicators. The second one, occur-ring between 3900 and 3400 yr BP, appears to be more prolonged and seems to generally involve intensive land exploitation. These changes have been coupled to the climate event detected by Magny et al. (2009) in the period from 4300 to 3800 yr BP, at a global scale centred on 4200 yr BP (e.g. Booth et al., 2005).
Wider comparisons and causes Correlations with pollen records from southwestern Europe (Jalut et al., 2000) and from Mediterranean islands (Pérez-Obiol and Sadori, 2007) have been attempted. Pollen diagrams from the Balearic islands (Yll et al., 1997) and central Sicily (Sadori and Narcisi, 2001) show remarkable synchronisms prior to 8000 yr BP, when the densest forest canopies are found in both areas. The data from the northwestern Iberian regions under Atlantic influ-ence (Muñoz-Sobrino et al., 2004) confirm this trend. But the sig-nal is in disagreement with the results from investigations of mountainous sites from central and southeastern Spain (Rascafría: Franco-Múgica et al., 1998; Villaverde: Carrión et al., 2001; Padul: Pons and Reille, 1988), in northern Catalonia (Olot: Pérez- Obiol, 1988) and in Almería (Pantaleon-Cano et al., 2003), where a probable increase of humidity is found in the period correspond-ing to the 8200 ka BP event (Alley et al., 1997). In the coastal records of southern France, Abies occurred more frequently after 9500 yr BP (Biot: Nicol-Pichard and Dubard, 1998; Palavas: Planchais, 1987), suggesting an increase in humidity, although a delay in the migration of this tree cannot be excluded. Mediter-ranean plants began to expand in the region after 8000 BP, with a pattern similar to that pointed out in the central Mediterranean.
At the beginning of the Holocene, a remarkable expansion of Pistacia (probably P. terebinthus in most Mediterranean sites,P. atlantica in Anatolian ones) is ubiquitous in the eastern Medi-terranean (e.g. Lawson et al., 2004; Willis, 1994; see also Roberts et al., 2011, this issue). Its peaks, mainly occurring between 10 000 and 8000 yr BP have sometimes been inferred to indicate an arid phase, i.e. climatically optimal for drought-adapted Medi-terranean vegetation, coeval with the period of deciduous oak expansion found in central Mediterranean sites. At the coastal lake of Voulkaria, in northwestern Greece (Jahns, 2005) Pistacia is present from the beginning of the postglacial reafforestation. A Pistacia expansion is documented (with maximum values at 8000 BP) in the open forest with deciduous and evergreen oaks found at Crete (Delphinos: Bottema and Sarpaki, 2003), resembling the expansion of Olea in Sicily (Lago di Pergusa: Sadori and Narcisi, 2001) and of Juniperus at Mljet (Malo Jezero: Jahns and van den Bogaard, 1998) while this spread of Mediterranean taxa is not documented in Bulgaria and in Croatia, at Cres (Schmidt et al., 2000) or in the northern Adriatic (Cheddadi et al., 1991). Marine records indicate that the Pistacia maxima corresponds to sapropel 1 deposition, while in coastal Sicily (Tinner et al., 2009) the stron-gest Pistacia expansion of the early–mid Holocene (up to 40%) took place slightly earlier, between 10 000 and 7500 yr BP. Tinner et al. (2009) interpreted this phase, with a dominance of grasses and Pistacia, as dry, even if humidity was increasing with respect to the start of the Holocene. This interpretation is in evident
contrast with the evidence found in pollen and isotope curves at Lago di Pergusa (central Sicily) (Sadori and Narcisi, 2001; Zanch-etta et al., 2007a) and at Grotta di Carburangeli (northern Sicily) in a speleothem record (Frisia et al., 2006).
A problem with discussions on the distribution of Pistacia in the Mediterranean during the early Holocene remains the fact that there are three main species of this low pollen producer genus (with apparently indistinguishable pollen grains), with different spatial occurrences and ecological requirements, one being a deciduous species (Pistacia terebinthus L.), one deciduous/semi-deciduous (P. atlantica L.) and the other evergreen (P. lentiscus L.). Different vegetation associations and climatic interpretations would result depending on which of these species was dominant during the early Holocene. The deciduous species could be better adapted to a subtropical savannah (Rossignol-Strick, 1999), while the evergreen one could find better adaptation in a relatively dry grassland. So, in the absence of unequivocal species identifica-tion, every climatic interpretation is a matter of opinion.
Even if in some sites evergreen oaks showed an earlier expan-sion (e.g. at Lago dell’Accesa, Drescher-Schneider et al., 2007) it was after around 8000–7500 yr BP that most pollen diagrams show an expansion of arboreal mediterranean-type taxa from Spain (e.g. Majorca: Burjachs et al., 1994; Salinas: Burjachs and Riera, 1995; Minorca: Yll et al., 1997; Navarres: Carrión and Van Geel, 1999) and France (Jalut et al., 2000; Capestang: Jalut, 1995; Jalut et al., 2009) to the eastern Mediterranean (Mljet: Jahns and van den Bogaard, 1998; Crete: Bottema and Sarpaki, 2003), pass-ing through central Italy (Lago dell’Accesa: Drescher-Schneider et al., 2007; Lago di Vico: Magri and Sadori, 1999; Lago Albano and Lago di Nemi: Lowe et al., 1996) and Sicily (Lago di Per-gusa: Sadori and Narcisi, 2001; Gorgo Basso: Tinner et al., 2009). In the same period, in almost all the sites situated at higher alti-tudes in Greece, Bulgaria and Albania and in most central Italian sites, a change in woodland composition is recorded with decreas-ing amounts of deciduous oaks and in an increase of one or more other tree taxa (e.g. Abies, Corylus, Fagus, Ostrya/Carpinus ori-entalis) demanding more humidity. Among the possible explana-tions of this contrasting evidence, often in the same site, a delay in the migration of mesophilous taxa seems likely, also consider-ing that water-demanding taxa need well-developed soils, which may not yet have been formed in previous millennia, whereas deciduous oaks forest demands a less organic soil.
To explain such a different behaviour in vegetation distribu-tion in the Mediterranean basin, we have to consider that at the start of interglacials seasonality was probably more extreme, with increased aridity in summer and higher precipitation in autumn/winter (Peyron et al., 2011, this issue). This rain appears to have been more in the form of intense events (Tzedakis, 2007). If a soil is saturated and not able to retain most of the excess water, this would have escaped as runoff on a slope or down karst systems in limestone geological situations. This can explain why at some sites we get sclerophylls during this period rather than deciduous trees. Only if the soils retained the moisture could the deciduous trees outcompete the sclerophylls despite the summer aridity. This intense autumn/winter precipitation and strong summer aridity might explain, for example, why the highest peaks of fires are found at Lago di Pergusa between c. 10 000 and 8000 yr BP, in correspondence with the highest concentration of deciduous oak pollen (Sadori and Giardini, 2007) and why, in a period of enhanced rainfall (Frisia et al., 2006) glaciers were reduced with respect to the second half of the Holocene (Giraudi et al., 2011,

126 The Holocene 21(1)
this issue). Moving northwards, at Corchia cave, a period with enhanced rainfall was found between 8900 and 7300 yr BP (Zanchetta et al., 2007b), but at Lago Frassino in northern Italy, an opposite trend was found, with isotopes suggesting a dry early Holocene from 9100 to 7000 yr BP (Baroni et al., 2006).
At 6000 yr BP a clear change is visible at Accesa, also present but less evident at Lago di Vico and Lago di Mezzano and Mljet lakes. In addition, at Lago di Massaciuccoli (central Italy, Colom-baroli et al., 2007; Marchetto et al., 2008) at Cres (Schmidt et al., 2000) and at Minorca (Yll et al., 1997) an increase of xeric condi-tions is found at about 6000 yr BP. A change after c. 6000 yr BP is also recorded in speleothems from Israel (Bar-Matthews and Ayalon, 2011, this issue; Bar-Matthews et al., 1997).
Changes in the Bronze Age in the northern central Mediterra-nean area (Figures 2, 3 and 4), occurring from around 4400 yr BP, are detected not only in many other central Mediterranean records (e.g. Caroli and Caldara, 2007; Drescher-Schneider et al., 2007; Lowe and Watson, 1994; Magri and Sadori, 1999; Noti et al., 2009; Oldfield et al., 2003; Reille, 1992; Sadori and Narcisi, 2001) but also in western (e.g. Carrión et al., 2003, 2004; Carrión García et al., 2000; Pérez-Obiol et al., 2011, this issue; Yll et al., 1997), eastern (e.g. Bottema and Sarpaki, 2003; Roberts et al., 2001, 2004; Wick et al., 2003) and southern Mediterranean coun-tries (Ben Tiba and Reille, 1982; Ritchie, 1984; Salamani, 1993). From central Mediterranean North African countries the past veg-etation record is so poor that a direct comparison with cultural changes (Mercuri et al., 2011, this issue) was not undertaken. The signal towards drier conditions in the period that corresponds to the changes in isotopic composition of speleothems (Drysdale et al., 2006) and other geological proxies (Giraudi et al., 2011, this issue) from central Italy and of plant macrofossils from Anatolia (Riehl et al., 2008) for the mid Holocene is also clear. The recent climate reconstructions inferred from Lake Accesa and Tenaghi Philippon (Greece) pollen sequences confirms this understanding of an evolution from wetter (early Holocene) to drier climatic condition but brings new insight on the seasonality process (Peyron et al., 2011, this issue). The early-Holocene period is reconstructed as a moist period from 9500 to 7800 cal. BP with wet winters, but dry summers, resulting in a strongly seasonal hydrological contrast (stronger than today). A later aridification phase, with a smaller seasonal contrast and lower overall precipi-tation lasts from 7800 to 5000 cal. BP.
Conclusion A well-known ‘mediterraneanization’ of lands surrounding the Mediterranean Sea took place during the mid and late Holocene (e.g. Pérez-Obiol et al., 2011, this issue). Even if evergreen spe-cies at present adapted to summer aridity were already present at some sites (e.g. Lago dell’Accesa: Drescher-Schneider et al., 2007) the expansion of drought-tolerant taxa all over the Mediter-ranean basin occurred principally after 8000 yr BP, associated with a re-organisation of regional climate. In the pollen records discussed here from the central Mediterranean this landscape transformation is clearest at Pergusa in Sicily (the driest and most southerly site) and at Malo Jezero in Dalmatia which saw a switch from deciduous to evergreen oak dominance. Voulkaria in west-ern Greece also showed some decline in deciduous oak percent-ages, but here the changes before c. 4000 yr BP were quite modest, due to the existence of a deciduous lowland forest still found in preserved coastal areas of the central Mediterranean. On the other
hand, the trend towards drought-tolerant plant taxa is much less evident in the other pollen records shown in Figure 2 (Vico, Maliq, Mezzano). In most of these other sequences, percentage values of AP and deciduous oak were as high at c. 3000 BP as at c. 9000 BP. This might imply that the landscape ecology of cen-tral and northern Italy and interior parts of the Balkans was less affected by mid-Holocene ‘mediterraneanization’ than other regions such as Spain, Sicily, Croatia or southern Greece. Even today, central and northern Italy is one of the wettest and greenest regions around the Mediterranean Sea. Moreover the land sur-rounding Maliq and Vico show rather high annual precipitation, placing the sites at the limit between mediterranean and humid temperate climate. The maintenance of deciduous forest cover in much of the Italian and Balkan peninsulas is also evident in pollen-based biomisation reconstructions comparing 6000 yr BP and the present day across the wider Mediterranean region (e.g. Roberts et al., 2004: plate 7). So, even though there is clear inde-pendent evidence of a change in climate between c. 8000 and c. 4000 yr BP (e.g. Giraudi et al., 2011, this issue), overall woodland cover and composition was not greatly changed everywhere on the northern shores of the central Mediterranean, presumably because many of these areas (with higher rainfall, well-developed soils, etc.) were less sensitive to ‘mediterraneanisation’ than other parts of the Mediterranean basins. Pollen concentration and influx data (Figures 3 and 4) imply that forest density fluctuated through time, but overall vegetation formations appear to have been less affected. Overall, we can conclude that before c. 4000 yr BP the main cause of vegetation change in the central Mediterranean was climatic variations, even if locally human impact can be detected. From c. 4000 to c. 2000 yr BP (i.e. from the Bronze Age to the Roman period), and also after this date, on the other hand, the main change in vegetation seems to have been human-induced, notably caused by land-cover change, even if significant fluctua-tions in climate also occurred.
Acknowledgements
Thanks are due to Gianni Zanchetta for advice in chronological assessment and critical reading. Fruitful discussions on vegeta-tion and climate dynamics with Chronis Tzedakis, Anna Maria Mercuri and Carlo Giraudi are gratefully acknowledged. We sin-cerely thank Neil Roberts for the fruitful advice and the language editing of the last version of this article. This study is a part of two projects financially supported by the French CNRS (National Centre for Scientific Research) within the framework of the ECLIPSE program (Past Environments and Climates), and the LAMA ANR project (MSHE Ledoux, USR 3124, CNRS) coordi-nated by M. Magny and N. Combourieu-Nebout. The authors sin-cerely thank Jacques Louis de Beaulieu, Ruth Drescher-Schneider, Anne-Marie Lézine, Michele Denefle, Elena Ortu and Amandine Bordon for their help and for providing access to the original pol-len data of Maliq and Accesa lakes.
References
Allen JRM, Watts WA, McGee E and Huntley B (2002) Holocene environ-
mental variability – The record from Lago Grande di Monticchio, Italy.
Quaternary International 88: 69–80.
Alley RB, Mayewski PA, Sowers T, Stuiver M, Taylor KC and Clark PU
(1997) Holocene climatic instability; A prominent, widespread event 8200
yr ago. Geology 25: 483–486.
Ariztegui D, Asioli A, Lowe JJ, Trincardi F, Vigliotti L, Tamburini F et al.
(2000) Palaeoclimatic reconstruction and formation of sapropel S1:

Sadori et al. 127
Inferences from Late Quaternary lacustrine and marine sequences in the
Central Mediterranean region. Palaeogeography, Palaeoclimatology,
Palaeoecology 158: 215–240.
Bar-Matthews M, Ayalon A and Kaufman A (1997) Late Quaternary paleo-
climate in the Eastern Mediterranean region from stable isotope analysis
of speleothems at Soreq Cave, Israel. Quaternary Research 47: 155–168.
Baroni C, Zanchetta G, Fallick AE and Longinelli A (2006) Mollusca stable
isotope record of a core from Lake Frassino, northern Italy: Hydrological
and climatic changes during the last 14 ka. The Holocene 16: 827–837.
Bellini C, Mariotti-Lippi M and Montanari C (2009) The Holocene landscape
history of the NW Italian coasts. The Holocene 19: 1161–1172.
Ben Tiba B and Reille M (1982) Recherches pollenanalytiques dans les mon-
tagnes de Kroumirie (Tunisie septentrionale), premiers résultats. Ecologia
Mediterranea 7: 75–86.
Beug H-J (1961) Beiträge zur postglazialen Floren- und Vegetationsgeschichte
Süddalmatiens. Der See ‘Malo Jezero’ auf Mljet, Teil 1 Vegetationsent-
wicklung. Flora 150: 600–630.
Bond G, Showers W, Cheseby M, Lotti R, Almasi P, deMenocal P et al. (1997)
A pervasive millennial-scale cycle in North Atlantic Holocene and glacial
climates. Science 278: 1257–1266.
Bond G, Kromer B, Beer J, Muscheler R, Evans MN, Showers W et al. (2001) Persistent solar influence on North Atlantic climate during the Holocene. Science 294: 2130–2136.
Booth RK, Jackson ST, Forman SL, Kutzbach JE, Bettis EA, Kreigs J et al.
(2005) A severe centennial-scale drought in midcontinental North America
4200 years ago and apparent global linkages. The Holocene 15: 321–328.
Bordon A, Peyron O, Lézine AM, Brewer S and Fouache E (2009) Pollen-
inferred Late-Glacial and Holocene climate in southern Balkans (lake
Maliq). Quaternary International 200: 19–30.
Bottema S and Sarpaki A (2003) Environmental change in Crete: A 9000-year
record of Holocene vegetation and the effect of the Santorini eruption. The
Holocene 13: 733–749.
Brande A (1973) Untersuchungen zur postglazialen Vegetationsgeschichte im
Gebiet der neretva-Niederungen (Dalmatien, Herzegowina). Flora 162:
1–44.
Burjachs F and Riera S (1995) Canvis vegetals i climatics durant el Neolitic
a la fac¸ana mediterrania iberica: 1 Congrés del Neolitic a la Peninsula
Iberica, Cava-Bellaterra. Rubricatum 1: 21–27.
Burjachs F, Pérez-Obiol R, Roure JM and Julia R (1994) Dinámica de la vege-
tación durante el Holoceno en la isla de Mallorca. In: Dupré Ollivier M,
Burgaz Moreno ME, Mateu Andrés I and Heras Güemes J (eds) Abstracts
del X Simposio de Palinología. Valencia: A.P.L.E., 199–210.
Calanchi N, Cattaneo A, Dinelli E, Gasparotto G and Lucchini F (1998) Tephra
layers in Late Quaternary sediments of the central Adriatic Sea. Marine
Geology 149: 191–209.
Caroli I and Caldara M (2007) Vegetation history of Lago Battaglia (Eastern
Gargano coast, Apulia Italy) during the middle–late Holocene. Vegetation
History and Archaeobotany 16: 317–327.
Carrión JS and van Geel B (1999) Fine resolution Upper Weichselian and
Holocene Palynological record from Navarrés (Valencia, Spain) and a dis-
cussion about factors of Mediterranean forest succession. Review of Pal-
aeobotany and Palynology 106: 209–236.
Carrión JS, Munuera M, Dupré M and Andrade A (2001) Abrupt vegetation
changes in the Segura Mountains of southern Spain throughout the Holo-
cene. Journal of Ecology 89: 783–797.
Carrión JS, Sánchez-Gómez P, Mota JF, Yll R and Chaìn C (2003) Holocene
vegetation dynamics, fire and grazing in the Sierra de Gádor, southern
Spain. The Holocene 13: 839–849.
Carrión JS, Yll EI, Willis KJ and Sánchez P (2004) Holocene forest history
of the eastern plateaux in the Segura Mountains (Murcia, Southeastern
Spain). Review of Palaeobotany and Palynology 132: 219–236.
Carrión García JS, Munuera Giner M, Navarro Camacho C and Sáez Soto F
(2000) Paleoclimas e historia de la vegetación quaternaria en España a
travéò del análisis polínico. Viejas falacias y nuevo paradigmas. Complu-
tum 11: 115–142.
Cheddadi R, Rossignol-Strick M and Fontugne M (1991) Eastern Mediterra-
nean paleoclimates from 26 to 5 Kyr BP documented by pollen and isotopic
analysis of a core in the anoxic Bannock basin. Marine Geology 100: 53–66.
Colombaroli D, Marchetto A and Tinner W (2007) Long-term interactions
between Mediterranean climate, vegetation and fire regime at Lago di
Massaciuccoli (Tuscany, Italy). Journal of Ecology 95: 755–770.
Colombaroli D, Vannière B, Chapron E, Magny M and Tinner W (2008) Fire–
vegetation interactions during the Mesolithic–Neolithic transition at Lago
dell’Accesa, Tuscany, Italy. The Holocene 18: 679–692.
Colombaroli D, Tinner W, van Leeuwen J, Noti R, Vescovi E, Vannière B et al.
(2009) Response of broad-leaved evergreen Mediterranean forest veg-
etation to fire disturbance during the Holocene: Insights from the peri-
Adriatic region. Journal of Biogeography 36: 314–326.
Coltelli M, Del Carlo P and Vezzoli L (2000) Stratigraphic constraints for
explosive activity in the past 100 ka at Etna Volcano, Italy. International
Journal of Earth Sciences 89: 665–677.
de Beaulieu J-L, Miras Y, Andrieu-Ponel V and Guiter F (2005) Vegetation
dynamics in north-western Mediterranean regions: Instability of the Medi-
terranean bioclimate. Plant Biosystems 139: 114–126.
Denèfle M, Lezine AM, Fouache E and Dufaure JJ (2000) A 12,000-year pol-
len record from Lake Maliq, Albania. Quaternary Research 54: 423–432.
Di Rita F and Magri D (2009) Holocene drought, deforestation and evergreen veg-
etation development in the central Mediterranean: A 5500 year record from
Lago Alimini Piccolo, Apulia, southeast Italy. The Holocene 19: 295–306.
Di Vito MA, Isaia R, Orsi G, Southon J, Di Vito S, D’Antonio M et al. (1999)
Volcanism and deformation since 12 000 years at Campi Flegrei caldera
(Italy). Journal of Volcanology and Geothermal Research 91: 221–246.
Dormoy I, Peyron O, Combourieu-Nebout N, Goring S, Kotthoff U, Magny M
et al. (2009) Terrestrial climate variability and seasonality changes in the
Mediterranean region between 15,000 and 4,000 years B.P. deduced from
marine pollen records. Climate of the Past 5: 615–632.
Drescher-Schneider R, de Beaulieu J-L, Magny M, Walter-Simonnet AV,
Bossuet G, Millet L et al. (2007) Vegetation history, climate and human
impact over the last 15 000 years at Lago dell’Accesa (Tuscany, Central
Italy). Vegetation History and Archaeobotany 16: 279–299.
Drysdale RN, Zanchetta G, Hellstrom J, Maas R, Fallick AE, Pickett M et al.
(2006) Late Holocene drought responsible for the collapse of Old World
civilizations is recorded in an Italian cave flowstone. Geology 34: 101–104.
Follieri M, Roure JM, Giardini M, Magri D, Narcisi B, Pantaléon-Cano J
et al. (2000) Desertification trends in Spain and Italy based on pollen anal-
ysis. In: Balabanis P, Peter D, Ghazi A and Tsogas M (eds) Mediterranean
Desertification. Research Results and Policy Implications. Proceedings
of the International Conference, 29 October to 1 November 1996, Crete,
Greece. European Commission, 2: 33–44.
Franco-Múgica F, García Antón M and Sainz-Ollero H (1998) Vegetation
dynamics and human impact in the Sierra de Guadarrama, Central System,
Spain. The Holocene 8: 69–82.
Frisia S, Borsato A, Mangini A, Spötl Ch, Madonia G and Sauro U (2006)
Holocene climate variability in Sicily from a discontinuous stalagmite
record and the Mesolithic to Neolithic transition. Quaternary Research 66:
388–400.
Giraudi C, Magny M, Zanchetta G and Drysdale R (2011) The Holocene cli-
matic evolution of the Mediterranean Italy: A review of the continental
geological data. The Holocene 21(1): 105–115 (this issue).
Jahns S (2002) An improved time scale for the Holocene history of vegetation
and environment on the South Dalmatian Island of Mljet. Vegetation His-
tory and Archaeobotany 11: 315–316.

128 The Holocene 21(1)
Jahns S (2005) The Holocene history of vegetation and settlement at the coastal
site of Lake Voulkaria in Acarnania, western Greece. Vegetation History
and Archaobotany 14: 55–66.
Jahns S and van den Bogaard C (1998) New palynological and tephrostrati-
graphical investigations of two salt lagoons on the island of Mljet, south
Dalmatia, Croatia. Vegetation History and Archaeobotany 7: 219–234.
Jalut G (1995) Analyse pollinique de sédiments holocènes de l’étang de Capes-
tang (Hérault). In: Guilaine J (ed.) Temps et Espace dans le Bassin de
l’Aude du Néolithique à l’Age du Fer. Resp. A.T.P. ‘Grands Projets d’Ar-
chéologie Métropolitaine’, CNRS, 293–302.
Jalut G, Esteban Amat A, Bonnet L, Gauquelin T and Fontugne M (2000)
Holocene climatic changes in the Western Mediterranean, from south-east
France to south-east Spain. Palaeogeography, Palaeoclimatology, Palaeo-
ecology 160: 255–290.
Jalut G, Deboutat JJ, Fontugne M and Otto T (2009) Holocene circum Medi-
terranean vegetation changes: Climate forcing and human impact. Quater-
nary International 200: 4–18.
Lawson I, Frogley M, Bryant C, Preece R and Tzedakis P (2004) The Lategla-
cial and Holocene environmental history of the Ioannina basin, northwest
Greece. Quaternary Science Reviews 23: 1599–1625.
Lionello P, Malanotte-Rizzoli P and Boscolo R (eds) (2006) Mediterranean
Climate Variability. Elsevier.
Lowe JJ and Watson C (1994) Lateglacial and early Holocene pollen stratig-
raphy of the northern Apennines, Italy. Quaternary Science Reviews 12:
727–738.
Lowe JJ, Accorsi CA, Bandini Mazzanti M, Bishop A, van der Kaars S, Forlani
L et al. (1996) Pollen stratigraphy of sediment sequences from lakes Albano
and Nemi (near Rome) and from the central Adriatic, spanning the interval
from oxygen isotope Stage 2 to the present day. In: Guilizzoni P and Old-
field F (eds) Palaeoenvironmental analysis of Italian crater lakes and Adri-
atic sediments. Memorie dell’Istituto Italiano di Idrobiologia 55: 71–98.
Magny M, de Beaulieu J-L, Drescher-Schneider R, Vannière B, Walter-Simonnet
A-V, Miras Y et al. (2007a) Early-Holocene climatic oscillations recorded
by lake-level fluctuations in west-central Europe and in central Italy. Quater-
nary Science Reviews 26: 1736–1758.
Magny M, Vannière B, de Beaulieu J-L, Bégeot C, Heiri O, Millet O et al.
(2007b) Holocene climate changes in the central Mediterranean as
recorded by lake-level fluctuations at Lake Accesa (Tuscany, Italy). Qua-
ternary Science Reviews 26: 1951–1964.
Magny M, Vannière B, Zanchetta G, Fouache E, Touchais G, Petrika L et al.
(2009) Possible complexity of the climatic event around 4300–3800 cal.
BP in the central and western Mediterranean. The Holocene 19: 1–11.
Magri D and Sadori L (1999) Late Pleistocene and Holocene pollen stratigra-
phy at Lago di Vico, central Italy. Vegetation History and Archaeobotany
8: 247–260.
Marchetto A, Colombaroli D and Tinner W (2008) Diatom response to mid-
Holocene climate change in Lago di Massaciuccoli (Tuscany, Italy). Jour-
nal of Paleolimnology 40: 235–245.
Mercuri AM, Sadori L and Uzquiano Ollero P (2011) Mediterranean and
north-African cultural adaptations to mid-Holocene environmental and cli-
matic changes. The Holocene 21(1): 189–206 (this issue).
Millet L, Vannière B, Verneaux V, Magny M, Disnar JR, Laggoun-Defarge F
et al. (2007) Response of littoral chironomid communities and organic
matter to late glacial lake-level, vegetation and climate changes at Lago
dell’Accesa (Tuscany, Italy). Journal of Paleolimnology 38: 525–539.
Muñoz-Sobrino C, Ramil-Rego P and Gómez-Orellana L (2004) Vegetation
of the Lago de Sanabria area (NW Iberia) since the end of the Pleistocene:
A palaeoecological reconstruction on the basis of two new pollen sequences.
Vegetation History and Archaeobotany 13: 1–22.
Nicol-Pichard S and Dubard M (1998) Reconstruction of Late-glacial and
Holocene environments in south-east France based on the study of a 66-m
long core from Biot, Alpes Maritimes. Vegetation History and Archaeo-
botany 7: 11–15.
Noti R, van Leeuwen JFN, Colombaroli D, Vescovi E, Pasta S, La Mantia T
et al. (2009) Mid- and late-Holocene vegetation and fire history at Biviere
di Gela, a coastal lake in southern Sicily, Italy. Vegetation History and
Archaeobotany 18: 371–387.
Oldfield F, Asioli A, Accorsi CA, Mercuri AM, Juggins S, Langone L et al.
(2003) A high resolution late Holocene palaeoenvironmental record from
the central Adriatic Sea. Quaternary Science Reviews 22: 319–342.
Pantaleon-Cano J, Yll EI, Perez-Obiol R and Roure JM (2003) Palynological
evidence for vegetational history in semi-arid areas of the western Mediter-
ranean (Almeria, Spain). The Holocene 13: 109–119.
Paterne M, Guichard F and Labeyrie J (1988) Explosive activity of the south Italian
volcanoes during the past 80 000 years as determinded by marine tephrochro-
nology. Journal of Volcanology and Geothermal Research 34: 153–172.
Pérez-Obiol R (1988) Histoire Tardiglaciaire et Holocène de la vegetation de
la region volcanique d’Olot (N.E. Péninsule Ibérique). Pollen et spores
30: 189–202.
Pérez-Obiol R and Sadori L (2007) Similarities and dissimilarities, synchro-
nisms and diachronisms in the Holocene vegetation history of the Balearic
Islands and Sicily. Vegetation History and Archaeobotany 16: 259–265.
Pérez-Obiol R, Jalut G, Julià R, Pèlachs A, Iriarte MJ, Otto T et al. (2011)
Mid-Holocene vegetation and climatic history of the Iberian Peninsula.
The Holocene 21(1): 75–93 (this issue).
Peyron O, Goring S, Dormoy I, Kotthoff U, Pross J, de Beaulieu J-L et al.
(2011) Holocene seasonality changes in the central Mediterranean region
reconstructed from the pollen sequence of Lake Accesa (Italy) and Tenaghi
Philippon (Greece). The Holocene 21(1): 131–146 (this issue).
Planchais N (1987) Impact de l’homme lors du remplissage de l’estuaire du
Lez (Palavas, Hérault) mis en évidence par l’analyse pollinique. Pollen et
spores 29: 73–88.
Pons A and Quézel P (1998) A propos de la mise en place du climat méditerra-
néen. Comptes Rendus de l’Académie des Sciences de Paris 327: 755–760.
Pons A and Reille M (1988) The Holocene and upper Pleistocene pollen record
from Padul (Granada, Spain): A new study. Palaeogeography, Palaeocli-
matology, Palaeoecology 66: 243–263.
Pross J, Kotthoff U, Müller UC, Peyron O, Dormoy I, Schmiedl G et al. (2009)
Massive perturbation in terrestrial ecosystems of the Eastern Mediter-
ranean region associated with the 8.2 kyr climatic event. Geology 37:
887–890.
Quézel P (1999) Les grandes structures de la végétation en région méditerra-
néen: Facteurs déterminants dans leur mise en place postglaciaire. Geobios
32: 19–32.
Raicich F, Pinardi N and Navarra A (2003) Teleconnections between Indian
monsoon and Sahel rainfall and the Mediterranean. International Journal
of Climatology 23: 173–186.
Ramrath A, Sadori L and Negendank JFW (2000) Sediments from Lago di
Mezzano, central Italy: A record of Lateglacial/Holocene climatic varia-
tions and anthropogenic impact. The Holocene 10: 87–95.
Reille M (1992) New pollen-analytical researches in Corsica: The problem of
Quercus ilex L. and Erica arborea L., the origin of Pinus halepensis Miller
forests. New Phytologist 122: 359–378.
Reille M and Pons A (1992) The ecological significance of sclerophyllous oak
forests in the west part of Mediterranean Basin: A note on pollen analytical
data. Vegetatio 99/100: 13–17.
Riehl S, Bryson RA and Pustovoytov K (2008) Changing growing condi-
tions for crops during the Near Eastern Bronze Age (3000–1200 B.C.):
The stable carbon isotope evidence. Journal of Archaeological Science 35:
1011–1022.
Ritchie JC (1984) Analyse pollinique des sédiments Holocène supérieur des
hauts plateaux du Maghreb Oriental. Pollen et spores 26: 489–496.

Sadori et al. 129
Roberts CN, Eastwood W, Kuzucuoğlu C, Fiorentino G and Caracuta V (2011)
Climatic, vegetation and cultural change in the eastern Mediterranean
during the mid-Holocene environmental transition. The Holocene 21(1):
147–162 (this issue).
Roberts N, Reed JM, Leng MJ, Kuzucuoglu C, Fontugne M, Bertaux J et al.
(2001) The tempo of Holocene climatic change in the eastern Mediter-
ranean region: New high-resolution crater-lake sediment data from central
Turkey. The Holocene 11: 721–736.
Roberts N, Stevenson T, Davis B, Cheddadi R, Brewster S and Rosen A
(2004) Holocene climate, environment and cultural change in the circum-
mediterranean region. In: Battarbee RW, Gasse F and Stickley CE (eds)
Past Climate Variability Through Europe and Africa. Dordrecht: Springer,
343–362, 627–637.
Rossignol-Strick M (1999) The Holocene climatic optimum and pollen records
of sapropel I in the eastern Mediterranean, 9000–6000 years BP. Quater-
nary Science Reviews 18: 515–530.
Sadori L (2006) Variazioni climatiche ed impatto umano: la storia della vege-
tazione e degli incendi durante l’età del Bronzo al Lago di Mezzano (VT).
Informatore Botanico Italiano 38: 117–122.
Sadori L (2007) Pollen records, postglacial. Southern Europe. Encyclopaedia
of Quaternary Sciences. Elsevier, 2763–2773.
Sadori L and Giardini M (2007) Charcoal analysis, a method to study veg-
etation and climate of the Holocene: The case of Lago di Pergusa, Sicily
(Italy). Geobios 40: 173–180.
Sadori L and Giardini M (2008) Environmental history in the Mediterranean
basin: Microcharcoal as a tool to disentangle human impact and climate
change. In: Fiorentino G and Magri D (eds) Charcoals from the Past:
Cultural and Palaeoenvironmental Implications. Proceedings of the Third
International Meeting of Anthracology, Cavallino – Lecce (Italy), 28
June–1 July 2004. BAR International Series, 1807: 229–236.
Sadori L and Narcisi B (2001) The postglacial record of environmental history
from Lago di Pergusa (Sicily). The Holocene 11: 655–671.
Sadori L, Giraudi C, Petitti P and Ramrath A (2004) Human impact at Lago
di Mezzano (central Italy) during the Bronze Age: A multidisciplinary
approach. Quaternary International 113: 5–17.
Sadori L, Zanchetta G and Giardini M (2008) Last Glacial to Holocene pal-
aeoenvironmental evolution at Lago di Pergusa (Sicily, Southern Italy) as
inferred by pollen, microcharcoal, and stable isotopes. Quaternary Inter-
national 181: 4–14.
Sadori L, Mercuri AM and Mariotti Lippi M (2010) Reconstructing past cul-
tural landscape and human impact using pollen and plant macroremains.
Plant Biosystems 144: DOI: 10.1080/11263504.2010.491982, in press.
Salamani M (1993) Prémières données paléophytogéographiques du Cèdre de
l’Atlas (Cedrus atlantica) dans la région de Grand Kabylie (NE Algérie).
Palynosciences 2: 147–155.
Santacroce R, Cioni R, Marianelli P, Sbrana A, Sulpizio R, Zanchetta G et al.
(2008) Age and whole rock–glass compositions of proximal pyroclastics
from the major explosive eruptions of Somma-Vesuvius: A review as a
tool for distal tephrostratigraphy. Journal of Volcanology and Geothermal
Research 177: 1–18.
Schmidt R, Müller J, Drescher-Schneider R, Krisai R, Szeroczyńska K and
Barić A (2000) Changes in lake level and trophy at Lake Vrana, a large
karstic lake on the Island of Cres (Croatia), with respect to palaeoclimate
and anthropogenic impacts during the last approx. 16,000 years. Jounal of
Limnology 59: 113–130.
Sulpizio R, Bonasia R, Dellino P, Di Vito MA, La Volpe L, Mele D et al. (2008)
Discriminating the long distance dispersal of fine ash from sustained col-
umns or near ground ash clouds: The example of the Pomici di Avellino
eruption (Somma-Vesuvius, Italy). Journal of Volcanology and Geother-
mal Research 17: 263–276.
Tinner W, van Leeuwen J, Colombaroli D, Vescovi E, van der Knaap WO,
Henne PD et al. (2009) Holocene environmental and climatic changes at
Gorgo Basso, a coastal lake in southern Sicily, Italy. Quaternary Science
Reviews 28: 1498–1510.
Tonkov S (2003) Holocene palaeovegetatìon of the northwestern Pirin moun-
rains (Bulgaria) as reconstructed from pollen analysis. Review of Palaeo-
botany and Palynology 124: 5l–61.
Tzedakis PC (2007) Seven ambiguities in the Mediterranean palaeoenviron-
mental narrative. Quaternary Science Reviews 26: 2042–2066.
Vannière B, Colombaroli D, Chapron E, Leroux A, Tinner W and Magny M
(2008) Climate versus human-driven fire regimes in Mediterranean land-
scapes: The Holocene record of Lago dell’Accesa (Tuscany, Italy). Quater-
nary Science Reviews 27: 1181–1196.
Vannière B, Power MJ, Roberts N, Tinner W, Carrión J, Magny M et al. (2011)
Circum-Mediterranean fire activity and climate changes during the mid-
Holocene environmental transition (8500–2500 cal. BP). The Holocene
21(1): 53–73 (this issue).
Wagner B, Lotter AF, Nowaczyk N, Reed JM, Schwalb A, Sulpizio R
et al. (2009) A 40,000-year record of environmental change from ancient
Lake Ohrid (Albania and Macedonia). Journal of Paleolimnology 41:
407–430.
Wick L, Lemcke G and Sturm M (2003) Evidence of Lateglacial and
Holocene climatic change and human impact in eastern Anatolia:
High-resolution pollen, charcoal, isotopic and geochemical records
from the laminated sediments of Lake Van, Turkey. The Holocene 13:
665–675.
Willis KJ (1992) The late Quaternary vegetation history of northwestern
Greece. 1. Lake Gramousti, 2. Rezina marsh, 3. comparative study of two
contrasting sites. New Phytologist 121: 101–155.
Willis KJ (1994) The vegetational history of the Balkans. Quaternary Science
Reviews 13: 769–788.
Wunsam S, Schmidt R and Müller J (1999) Holocene lake development of
two Dalmatian lagoons (Malo Jezero and Veliko Jezero, Isle of Mljet) in
respect to changes in Adriatic sea level and climate. Palaeogeography,
Palaeoclimatology, Palaeoecology 146: 251–281.
Yll EL, Pérez-Obiol R, Pantaleon-Cano J and Roure JM (1997) Palynological
evidence for climatic change and human activity during the Holocene on
Minorca (Balearic Islands). Quaternary Research 48: 339–347.
Zanchetta G, Borghini A, Fallick AE, Bonadonna FP and Leone G (2007a)
Late Quaternary palaeohydrology of Lake Pergusa (Sicily, southern Italy)
as inferred by stable isotopes of lacustrine carbonates. Journal of Paleo-
limnology 38: 227–239.
Zanchetta G, Drysdale RN, Hellstrom JC, Fallick AE, Isola I, Gagan MK
et al. (2007b) Enhanced rainfall in the Western Mediterranean during depo-
sition of sapropel S1: Stalagmite evidence from Corchia cave (Central
Italy). Quaternary Science Reviews 26: 279–286.
Zanchetta G, Sulpizio R, Roberts CN, Cioni R, Eastwood WJ, Siani G et al.
(2011) Tephrostratigraphy, chronology and climatic events of the Medi-
terranean basin during the Holocene: An overview. The Holocene 21(1):
33–52.
Related Documents











